
Submitted:
24 November 2025
Posted:
27 November 2025
You are already at the latest version
Abstract
What mechanisms drive and shape the stepwise evolution from simple carbon-based materials (CBMs) into complex organisms and societies? This fundamental question spans life's origin, biological diversification, and societal development, yet remains unresolved because it has often been studied in isolation. Through cross-disciplinary integrative reasoning, empirical validation, and mathematical modeling, here we extend Darwinian theory to formulate the Carbon-Based Evolutionary Theory (CBET) ¾ the first explicit framework that unifies chemical, biological, and social evolution. CBET highlights energy flow, functions of CBMs, environmental influence, and mathematical rules. It derives three axioms and two key mechanisms ¾ spriodynamic feedback and natural selection, which drive and shape the stepwise evolution from carbon atoms to societies, respectively. CBET schematically explains how CBMs diversify, elevate hierarchies, increase fitness, and enhance orderliness and complexity. Furthermore, CBET refines fitness calculation and clarifies five common misunderstandings about evolution. It elucidates the natural balance of competition versus inclusiveness, selfishness versus altruism, and individual versus collective interests, thereby offering a scientific foundation for fostering balanced, harmonious societal development.
Keywords:
carbon-based material
; mechanism
; chemical evolution
; biological evolution
; social evolution
; natural selection
; theory
Introduction
Why have simple carbon-based materials (CBMs) — such as carbon atoms, methane, and carbon dioxide — evolved stepwise into the complex, diverse, and ordered organisms and societies we observe today? This fundamental question has long fascinated scholars but remains unresolved [1,2,3,4,5,6,7,8]. It involves the chemical evolution of organic molecules [5], the biological evolution of organisms, and the social evolution of animal and human societies [7,8]. Traditionally, these three domains have been studied in isolation [1,2,3,4,5,6,7,8], and thus theories developed in one domain often fail to apply to others. For instance, while Darwinian theory provides a robust account for biological evolution, it does not address chemical or social evolution [1,2]. Consequently, it offers neither a foundational explanation for life’s origin nor reliable insights for understanding societal development [1,2,3,4]. Darwinian theory has even been improperly extended to societal development to rationalize selfishness, slavery, massacres, and colonialism [2,4]. Additionally, certain biological phenomena, such as epigenetic changes, neutral mutations, and non-random mutations, have not been seamlessly integrated into Darwinian theory [2]. To bridge these gaps, here we propose the Carbon-Based Evolutionary Theory (CBET), which builds upon and extends Darwinian theory to unify chemical, biological, and social evolution within a single coherent framework. CBET is derived through integrative reasoning from extensive established knowledge across diverse disciplines that researchers have accumulated since Darwin’s era.
Results
Hierarchies and Functions of CBMs
The central concept of ‘organisms’ in Darwinian theory is extended into the more inclusive ‘CBMs’ in CBET. As shown in Figure 1, CBMs are classified into nine hierarchies (H1 to H9) based on the criterion that higher hierarchies are constructed by lower ones through chemical, physical, or more complex integrative interactions. H1-CBMs refer to carbon atoms, which likely originated in stellar interiors [9]. H2-CBMs are small carbon-containing molecules (CCMs), like CH₄ and CO₂. H3-CBMs are medium CCMs, like glucose and lysine. Polymerization of some H3-CBMs forms H4-CBMs, which refer to large CCMs, like proteins and nucleic acids — some of which can assemble into H5-CBMs, namely multi-molecule aggregates (MMAs), like phospholipid bilayers. The transition to life occurs at H6 (cells) and culminates at H7 (multicellular organisms). Some H7 individuals form H8-CBMs, which represents single-level societies (e.g., ant colonies, small companies). Some H8-CBMs form H9-CBMs, which are multi-level societies (e.g., universities, armies, nations). Boundaries between hierarchies are not rigid, and transitional forms such as peptides (between H3 and H4) and viruses (between H4 and H5) can act as bridges facilitating hierarchical elevation of CBMs.
The following functions of CBMs are crucial to CBM evolution.
(1) Macromolecular backbone function. Carbon is unique in its ability to form long, stable chains and complex rings through relatively strong, versatile C-C bonds, which can link to various atoms (such as hydrogen, oxygen, nitrogen, sulfur, and halogens) and functional groups (such as hydroxyl, carbonyl, amino, and carboxyl groups), giving rise to immense molecular diversity. In contrast, other elements lack the ability to form stable extended chains through self-bonding, although silicon and boron can form weak and short covalent chains (e.g., Si-Si, B-B) that are prone to cleavage. Therefore, carbon atoms are uniquely capable of serving as the backbones of myriad stable macromolecules [10]. This fundamental disparity explains why CBMs, not silicon or boron-based ones, have naturally evolved into life.
(2) Hierarchy elevation function. CBMs across H1–H9 can form higher-hierarchy CBMs through physical, chemical, biological, and/or social interactions. This is evidenced by transitions from CO₂ to amino acids, from amino acids to proteins, from individual ants to colonies, from tribes to alliances, and from small companies to large corporations.
(3) Relative stability function. Various CBMs are relatively stable. For instance, diverse H3-CBMs could persist for billions of years in meteorites [11]; some proteins and nucleic acids were preserved in fossils for millions of years [12]; humans typically live for decades; and some companies have existed for years.
(4) Changeability function. The repeated formation and degradation of higher-hierarchy CBMs constitute Earth’s carbon cycles [13], and various CBMs (e.g., proteins and organisms) produced in these cycles exhibit variations relative to their progenitors.
(5) Information function. H6-CBMs and H7-CBMs can encode, read, translate, change, store, and transmit genetic information through the interactions of nucleic acids and proteins [2]. They also use diverse signaling pathways to respond to internal and external changes. Some H8-CBMs and H9-CBMs harness additional information forms — such as scents, sounds, behaviors, scripts, and electromagnetic waves.
(6) Other functions. The energy-storage and energy-release functions of various H3-CBMs (e.g., glucose) and H4-CBMs (e.g., lipids), the catalytic functions of various H4-CBMs (e.g., proteins), the reproduction of H6-CBMs, and the knowledge accumulation function of humans are all crucial to CBM evolution.
Reasoning Steps of CBET
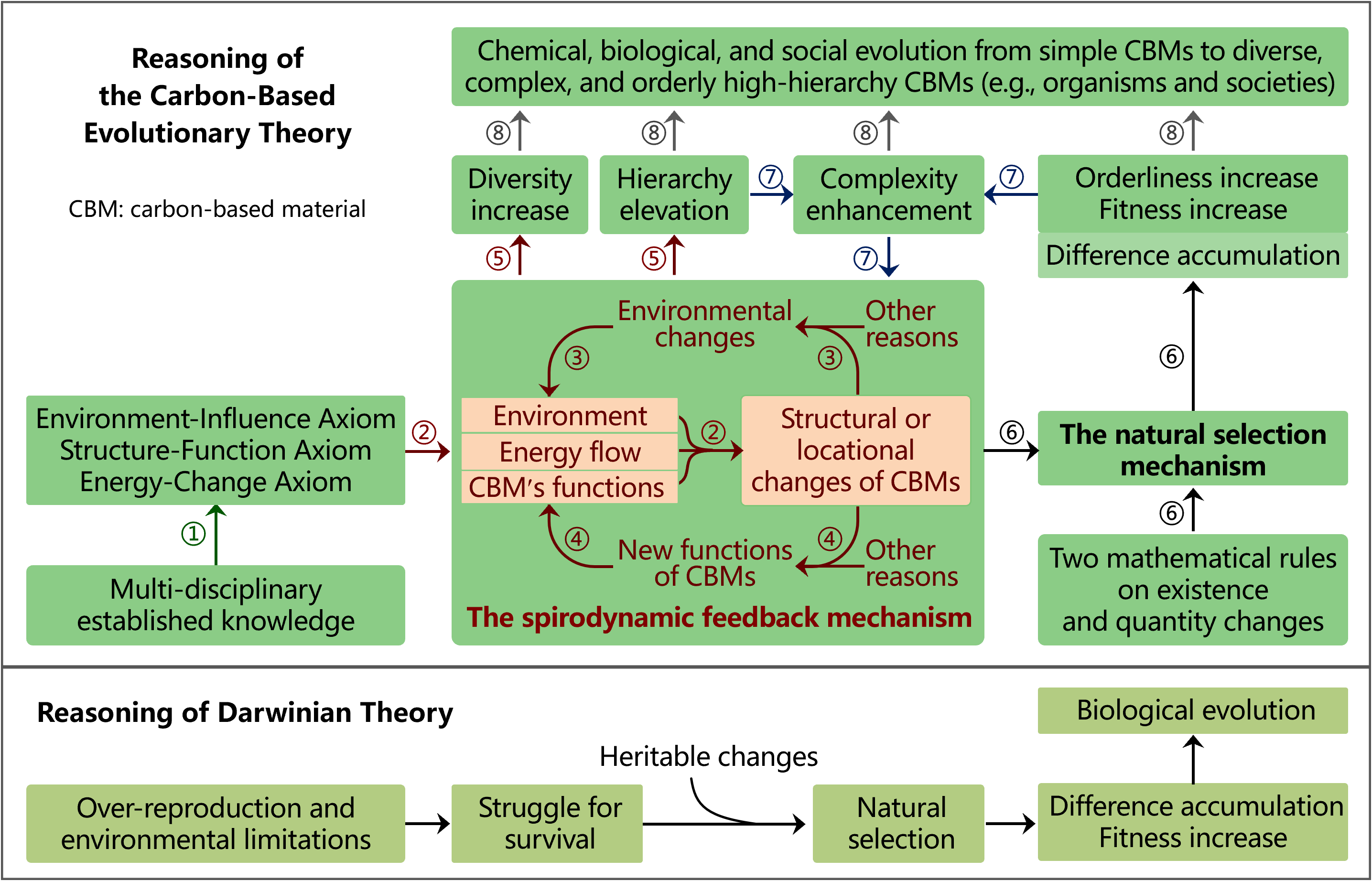
CBET was derived through eight steps and five tiers of integrative reasoning (Figure 2). Mathematical formulation of these steps is given in Supplementary Text 1.
Step 1. Reasoning of three axioms
The CBET framework begins with the derivation of three axioms from foundational principles across diverse disciplines, such as Newtonian mechanics from physics, the laws of thermodynamics and energy conservation, and reaction principles from chemistry. This is achieved by extracting the core qualitative relationships within these principles to establish their applicability to all molecules, organisms, and societies. This derivation represents Tier 1 integrative reasoning within CBET. These axioms are explicit and have long guided research in physics, chemistry, biology, and the social sciences.
(1) The Environment–Influence Axiom. Environment factors influence structural or locational changes in a material. These changes, in turn, can alter the environment of the material. For example, Earth’s habitable environment fosters CBM evolution [14,15], which in turn profoundly transforms Earth’s environment.
(2) The Structure–Function Axiom. A material’s structure partly determines its functions, and structural changes can generate new functions. Concurrently, functions are the internal causes for maintaining or modifying structures. For example, a cell’s structure dictates its metabolic and reproductive abilities, which in turn are essential for sustaining and replicating that structure. This axiom suggests that the execution of a complex function (e.g., a car’s operation and a cell’s reproduction) — defined as one requiring multiple coordinated steps — typically necessitates a complex structure of specialized, organized components, because each step requires a specialized structural component.
(3) The Energy-Change Axiom. Energy flow drives structural changes (e.g., protein synthesis) or locational changes (e.g., bird flying) in a material under the modulation of functions of the material (internal causes) and the environment (external causes). This axiom highlights energy flow as the universal driver of structural and locational changes across all CBMs, from carbon atoms to societies. It is derived from and supported by the two axioms above and principles from Newtonian mechanics and organic chemistry, as well as evidence from animal locomotion and social activities.
Steps 2–5. Reasoning of the spriodynamic feedback mechanism
Step 2. According to the Energy-Change Axiom, energy flow under the modulation of functions of CBMs and the environment drives structural or locational changes in CBMs.
Step 3. Structural changes in CBMs and other factors such as environmental shifts can generate novel functions. These new functions can feed back to Step 2 and thus initiate further structural changes in CBMs. An example is the formation of nucleotides, which enables their polymerization into nucleic acids — a novel function that emerges when nucleotides are formed.
Step 4. Structural or locational changes from Step 2 can cause environmental changes. Other factors like earthquakes can also cause environmental changes. Environmental changes can feed back to Step 2 and thus initiate further structural or locational changes in CBMs. For instance, the atmospheric oxygenation by photosynthetic bacteria ~2.5 billion years ago selected against anaerobes in favor of aerobes [2,16].
Step 5. The iterative feedback loops between changes in structures or locations (Step 2), functions (Step 3), and environments (Step 4) constitute the spriodynamic feedback mechanism. This represents Tier 2 integrative reasoning in CBET, which integrates the three axioms with the functions of CBMs and the foundational concept of feedback loops in systems theory. This mechanism serves as the primary engine for amplifying CBM diversity (partially due to the changeability function of CBMs) and elevating CBM hierarchies (partially due to the hierarchy elevation function of CBMs).
Step 6. Reasoning of the natural selection mechanism
The following two rules are self-evident in fundamental mathematics.
(1) The Quantity-Existence Rule. A material exists if its cumulative formation exceeds its cumulative degradation. This applies to the synthesis/breakdown of molecules, birth/death of organisms, and establishment/dissolution of social organizations.
(2) The Quantity-Change Rule. The change in a material’s existing quantity over a time interval [a,b] is determined by its initial quantity and the ratio of its total formation () to its total degradation () within that period. This ratio reflects a material’s overall performance in formation and sustenance (OPFS). In the context of CBET, we define this OPFS ratio as the universal measure of fitness (Equation 1).
In modern biology, fitness is defined as the number of offspring an individual leaves in the next generation [2]. This measure fails to capture the fitness of molecules, social organizations, or worker bees, and it neglects generation time. The universal fitness calculation in Equation 1 overcomes these limitations.
Guided by the two mathematical rules, changes that increase fitness tend to accumulate, while those that decrease fitness tend to be eliminated. This process constitutes the natural selection mechanism and Tier 3 integrative reasoning within CBET. Natural selection filters changes resulting from the spriodynamic feedback mechanism and thus non-randomly shapes CBM evolution and enhances fitness across all hierarchies. As suggested by the Structure–Function Axiom, such fitness improvements typically entail greater structural orderliness and enhanced internal collaboration.
Step 7. Reasoning of complexity enhancement and its feedback
The hierarchical elevation in Step 5 and the enhanced internal collaboration in Step 6 can collectively increase the structural complexity of CBMs. This represents Tier 4 integrative reasoning in CBET, which integrates the effects of spriodynamic feedback and natural selection. The heightened structural complexity itself is a type of structural changes and can thus constitute an input into the spriodynamic feedback and natural selection mechanisms. This creates a new feedback loop that can, in some cases, intensify the pressure of natural selection — a phenomenon illustrated by coevolutionary arms races, such as those between predators and prey [17]. For instance, structural adaptations that grant gazelles greater speed exert stronger selection pressure on their predators (e.g., cheetahs) for corresponding speed enhancements. The success of these faster predators then exerts even stronger selective pressure for increased speed in the gazelle population.
Step 8. Synergy of the reasoning steps
The seven reasoning steps above demonstrate how CBET integrates three axioms, two mathematical rules, CBM functions, and environmental factors to formulate two key mechanisms that respectively drive and shape the stepwise evolution of CBMs from simple atoms into complex, diverse, and ordered organisms and social organizations. This synergetic step represents Tier 5 integrative reasoning in CBET. In this process, the vast diversity of CBMs at H4 and higher hierarchies—driven by the spriodynamic feedback mechanism—facilitates the origin of novel, higher-hierarchy CBMs, which will persist under natural selection provided their formation exceeds their degradation.
In comparison, Darwinian theory primarily aligns with Step 6 of CBET and derives natural selection from over-reproduction, environmental limitations, struggle for survival, and heritable changes (Figure 2). It clarifies neither the mathematical rules of natural selection nor the mechanism underlying the hierarchy elevation and diversification of CBMs.
Explanation of Life’s Origin
(1) Driving role of spriodynamic feedback. Life’s origin, viewed as the stepwise hierarchical elevation of CBMs from H1 to H6 on Earth, was driven by the spriodynamic feedback mechanism. This mechanism involves energy flow and feedback loops among changes in structures, functions, and environments of CBMs (Figure 2). In thermodynamics, certain H2-CBMs can spontaneously absorb energy from sunlight, radiation, geothermal heat, and other sources under specific environmental conditions, be thereby activated to form diverse H3-CBMs through chemical reactions [3,5,6,15]. Likewise, some of the resulting H3-CBMs can spontaneously absorb energy from energy flow sources and be thereby activated to form H4-CBMs through chemical reactions under specific environmental conditions (e.g., presence of catalysts) [5,6,15]. This stepwise process establishes a natural energy-driven trend toward the gradual accumulation and increasing complexity of organic molecules. In astronomy and geology, this process could have persisted on Earth for hundreds of millions of years before life’s origin [5,6,15], and thus would have led to the accumulation of myriad macromolecules (H4-CBMs). In physics, the energy flows — such as those from wind, water flow, and wet–dry cycles — can drive the assembly of these macromolecules into innumerable MMAs (H5-CBMs) and MMA clusters. Functions of relevant CBMs, such as their interaction through hydrogen bonds, also influence the formation of MMA clusters. Some MMA clusters, owing to their structural complexity, may have developed metabolic functions such as catalyzing the synthesis or degradation of organic molecules (e.g., amino acids, nucleotides). This capability could have accelerated the formation-degradation cycles of CBMs, including H3–H5, H4-CBMs, H5-CBMs, and MMA clusters.
(2) Shaping role of natural selection before life’s origin. Prior to life’s origin, natural selection preferentially accumulated those H3-CBMs, H4-CBMs, H5-CBMs, and MMA clusters with a higher formation-to-degradation ratio (i.e., higher fitness) during their formation-degradation cycles. Generally, fitness follows this ascending order: 1) an unstable nucleic acid; 2) a stable nucleic acid formed without catalysis; 3) a stable nucleic acid whose formation (but not replication) is catalyzed; and 4) a stable nucleic acid whose synthesis is catalyzed in a replicative mode (e.g., the RNA genomes of coronaviruses and other RNA viruses replicated by certain proteins). Likewise, stable proteins whose replication is catalyzed by certain MMA clusters — such as ribosomes [18] — also possess relatively high fitness and are thus favored by natural selection. In turn, due to their complex structures, some nucleic acids and proteins can act as catalysts or form part of MMA clusters that catalyze the replicative formation of nucleic acids and proteins. These scenarios demonstrate that natural selection can shape the evolution or the formation-degradation cycles of H4-CBMs, H5-CBMs, and MMA clusters in a direction conducive to life’s origin.
(3) Two mechanisms leading to life’s origin. Driven by the spriodynamic feedback mechanism, which amplified molecular diversity and hierarchical complexity, and shaped by natural selection, which preferentially accumulated stable macromolecules and catalytic MMA clusters, an exceedingly rare fraction of these MMA clusters eventually attained the emergent function of reproduction due to their extremely complex structures, becoming the first life structures (H6-CBMs).
(4) Shaping role of natural selection after life’s origin. The reproduction function provided a decisive advantage of life structures to increase their prevalence, as the function could facilitate the formation of life structures to consistently exceed their degradation. In contrast, no such sustaining mechanism exists for non-replicating MMA clusters to increase their quantities. Moreover, life structures have the changeability function, and consequently, myriad variants of life structures could have emerged on Earth over billions of years, and those variants with mutations that enhance their fitness are favored by natural selection, making life structures increasingly advantageous over non-reproductive MMA clusters. This could have led to the prosperity of life structures and the decline of non-reproductive MMA clusters on Earth, along with their constituent H2-CBMs, H3-CBMs, and H4-CBMs.
(5) Unique capacity of CBMs and Earth’s habitability. While the three axioms and two mechanisms in CBET could be extended from CBMs to other materials and allow their hierarchy elevation and fitness enhancement, only CBMs possess the function to reach and surpass the hierarchy of diverse and stable macromolecules (H4), as explained above. Moreover, the formation of the complex life structures requires a suitable environment like Earth, which provides abundant energy, ample CBMs, plentiful liquid water, and other conditions [14,15]. While various environments on early Earth, such as hydrothermal vents or volcanic lakes, have been proposed for life’s origin [2,4], our analysis, detailed in Supplementary Text 1, indicates that the early atmosphere could be much more conducive to life’s origin [19,20,21,22,23,24].
(6) Probabilistic nature and intermediate steps. Due to the extraordinary complexity of life structures — particularly exemplified by systems like ribosomes, which produces proteins according to the sequences of nucleic acids [18] — CBET posits that the probability of an inanimate MMA cluster transitioning into a life structure, while greater than zero, is exceedingly low. Consequently, some intermediate steps were likely essential for life’s origin. For instance, given that life structures require diverse metabolic functions, it is more plausible that they evolved from metabolic MMAs or MMA clusters, which already possessed some foundational catalytic capabilities, rather than directly from individual macromolecules.
Clarification of Misunderstandings
The CBET framework can be employed to clarify the following five common misunderstandings about natural selection. These misunderstandings are not flaws in Darwinian theory but rather in the common interpretations.
(1) Misunderstanding about ‘Survival of the fittest’. Darwin adopted this phrase as shorthand for natural selection, but it is often interpreted literally to mean that only the fittest can survive [1,4]. According to Equation 1 in CBET, a bacterial mutant with a detrimental mutation can increase in population size, even if its growth rate (e.g., 0.05% per day) is lower than that of its wild-type counterpart (e.g., 0.30% per day). This illustrates that ‘survival of the fit enough’ is more literally accurate than ‘survival of the fittest.’ Likewise, major evolutionary transitions — such as multicellularity, endothermy, and eusociality — could have been initiated by pioneers that were not fitter than their ancestors but were fit enough to establish new lineages. Additionally, a CBM’s fitness reflects the combined contribution of all its traits, while specific key traits (e.g., antelope speed) can have an outsized effect on fitness. Therefore, natural selection shapes both all-around and specialized adaptation.
(2) Misunderstanding that all traits are advantageous. The example above shows that neutral or even detrimental traits can persist, provided the relevant CBM is fit enough. This is also evidenced by the vulnerability of human infants and the prevalence of neutral mutations in genomes [2]. Furthermore, traits may be advantageous in some contexts but detrimental in others. For instance, giraffes’ height aids browsing and combat but hinders rising from ground [25].
(3) Misunderstanding that natural selection is purely competitive. As elucidated above, natural selection is also inclusive because it can tolerate advantageous, neutral, and even detrimental traits, provided the relevant CBM is fit enough, particularly in conductive environments or at the early stage of evolution when only a few competitors are available. This inclusiveness is also evidenced by the long-term increase in biodiversity throughout Earth’s history [2], though catastrophes such as asteroid impacts occasionally trigger sharp declines in biodiversity [2].
(4) Misunderstanding that natural selection equals selfishness. Selfish behavior is critical for acquiring energy, resources, favorable environments, and mating opportunities [1,26]. Yet CBET shows that selfishness and altruism coexist and balance throughout evolution. Diverse molecules often serve as catalysts, energy sources, or building blocks for the replication of other molecules [4]; diverse macromolecules in cells enable genetic transmission of genomic DNA; somatic cells in multicellular organisms support the passage of reproductive cells; ant workers sacrifice for colony reproduction [7]; and humans in armies or police may die for societies. According to the Energy-Change Axiom, these altruistic behaviors above are driven by energy flow that is modulated by the functions of the relevant CBMs and the environment.
(5) Misunderstanding that only heritable changes matter. It is frequently assumed that natural selection acts solely on heritable genetic changes, and that only these can lead to evolutionary change across generations [1,2]. In effect, non-heritable or acquired factors (e.g., vaccination, education), along with epigenetic changes (which may or may not be inherited), influence survival and reproduction [27]. Therefore, these changes also affect fitness and are under natural selection in shaping evolutionary outcomes. This suggests that both acquired and genetic factors matter in shaping evolutionary trajectories.
Validation of CBET
The validity of CBET is rooted in explicit and rigorous reasoning, extensive empirical evidence, its ability to explain cross-disciplinary phenomena, and robustness against iterative falsification, as detailed in Methods.
Discussion
Significance in natural sciences. CBET potentially stands as the first framework that seamlessly bridges chemical, biological, and social evolution — a goal long desired in research [2,4,8,28]. It inherits the natural selection mechanism — which shapes evolution — from Darwinian theory. CBET innovatively derives the spriodynamic feedback mechanism — which drives the stepwise evolution of CBMs from carbon atoms to life and societies. Furthermore, CBET reveals the mathematical rules governing natural selection, refines fitness calculation, and integrates multiple facts (e.g., the prevalence of neutral mutations) that have not been integrated in a single evolutionary theory. CBET clarifies the intricate relationships among energy, structures, functions, information, orderliness, and complexity (Figure 2), which have been debated for decades [29]. CBET suggests that complex functions of certain CBMs — such as reproduction, nonrandom mutagenesis, and epigenetic changes — stems from their complex structures [27,30]. Comparison of CBET to other theories is detailed in Supplementary Text 1, and certain differences between Darwinian theory and CBET were shown in Table 1.
Significance in social sciences. CBET offers an innovative explanation for the origin of societies. It reveals that changes in CBMs — from organic molecules to societies — are governed by the same axioms, mathematical rules, and mechanisms of spriodynamic feedback and natural selection. It elucidates the prevalence of altruistic behavior and the importance of acquired traits in social evolution. Furthermore, CBET illuminates the persistent natural balance between seemingly opposing forces, such as self-functioning versus environmental influence, competition versus inclusiveness, individual versus collective interests, and social development versus environmental protection. Thereby, CBET sheds new light from a panoramic perspective on sociology, ethics, and political philosophy, offering a scientific basis for balanced and harmonious societal development. Additionally, the spriodynamic feedback mechanism underscores the systemic and long-term effects of policies. Together, CBET establishes an innovative, natural-science-grounded ontological foundation for social sciences.
Boundaries of CBET. Both Darwinian theory and CBET only provide a schematic and qualitative explanation for the plausibility of evolution, not a detailed, quantitative account of the inevitability of evolution. For example, although CBET innovatively derives two mechanisms to drive and shape the evolution from MMA clusters to life’s origin, it clarifies few details about for life’s origin. To unveil the details about life’s origin, more studies in astronomy, chemistry, biology, and computer science under the guide of the CBET framework are needed. Additionally, both Darwinian theory and CBET do not present new principles in mathematics, physics, or chemistry, and their core viewpoints can be traced back to certain previous theories. For example, the spriodynamic feedback mechanism in CBET can be traced back to the feedback concept in systems theory. The above boundaries of Darwinian theory and CBET have lowered the threshold for establishing the two theories and enhanced their credibility. For example, CBET would be uncredible if it described protein synthesis and the emergence dynamics of companies using the same quantitative equation.
Outlook of CBET. Looking ahead, the CBET framework will guide studies that use relevant investigations, experiments, and computational modeling: (i) to establish a new theory of chemical evolution, detailing how the spriodynamic feedback and natural selection mechanisms drove and shaped the hierarchical ascent of macromolecules and life’s origin under early Earth conditions; (ii) to establish a new theory of biological evolution, which integrates all major findings in biology and details how the two mechanisms facilitated major transitions; (iii) to establish a new theory of social evolution, which integrates the entirety of human societal history and enables the forecasting of long-term evolutionary trends, including in the post-AI era. These cross-disciplinary applications will not only enrich respective fields but also serve to rigorously test, validate, and refine the CBET framework itself.
Methods
Methodology of CBET
Darwinian theory was initially developed by Darwin and Wallace through the integration of the insights from diverse fields, such as Malthusian demography, Lyellian geology, artificial selection, and biogeography [2]. Later, this theory has been empirically validated by more observations and scientific advancements and refined thereby (Figure 3). CBET extends this methodology through the integration of established knowledge from modern physics, chemistry, biology, and social sciences to derive its three axioms and two core mechanisms (Figure 3). This is followed by empirical validation and iterative refinements.
Integrative Reasoning of CBET
Selection of CBMs as CBET’s core concept. The core concept of ‘organism’ is to biological evolution what ‘carbon-based material (CBM)’ is to CBET. Unlike more restricted or abstract concepts like ‘molecule’ ‘gene’ ‘system’ or ‘entropy’, ‘CBM’ is an explicit and precise concept that seamlessly encompasses entities from carbon atoms and organic molecules to organisms and societies.
Integration of various key factors. CBET integrates key factors that are consistently crucial to CBM evolution across hierarchies, including energy, structure, function, environment, feedback, natural selection, and specific enabling factors (e.g., water and diverse hierarchies of CBMs). The deliberate selection of these concrete key factors allows CBET to provide a direct and explicit explanatory framework.
Eight steps and five tiers of integrative reasoning. CBET was derived through eight steps and five tiers of integrative reasoning, as outlined in Figure 2.
Circumvention of circular reasoning. CBET derives the spriodynamic feedback mechanism — grounded in three universal axioms drawn from extensive empirical facts — to address the origin and availability of new structural variations upon which natural selection acts. CBET has also clarified the mathematical rules governing natural selection. By clearly separating the generation of variation (via spriodynamic feedback) from the selection of variation (via natural selection), CBET circumvents circular reasoning (i.e., tautology) [32].
Empirical Validation of Spriodynamic Feedback
To empirically validate the spriodynamic feedback mechanism, we subjected it to a stringent test against 27 representative facts, randomly selected across a spectrum from molecular to social phenomena (Table 2). All these 27 facts align with the core characteristics of the spriodynamic feedback mechanism, as detailed below.
- ✧
- They require or rely on energy flow.
- ✧
- They are influenced by environmental factors.
- ✧
- They rely on functions of the relevant material, which are partially determined by the structure of the relevant material.
- ✧
- They demonstrate that structural changes can generate new, emergent functions, which can initiate further structural changes, establishing a feedback loop that can lead to hierarchical elevation, as illustrated by Facts 1, 4, 7, 10, 13, 16, 19, 22, and 25.
- ✧
- They show that structural or locational changes can alter the environment, which can in turn feedback to initiate further structural changes, as evidenced by Facts 1, 3, 5, 6, 8, 18, 20, and 27.
Empirical Validation of Natural Selection
In biological evolution, extensive facts have been identified for the effect of natural selection across multiple taxa and ecosystems [1,2]. In chemical evolution, molecular populations undergo natural selection in those organic chemical reactions with multiple possible products, where the products that are easily formed and difficult to degrade tend to accumulate in greater quantities than those that are difficult to form and easily degraded. In social evolution, companies or schools undergo competition and selection, and those with the formation-to-degradation ratio >1 demonstrate progressive accumulation.
Evaluation of Conceptual Clarity and Logical Coherence
Following the empirical support outlined above, we further evaluated the CBET framework by assessing the conceptual clarity and logical coherence of its core components. This evaluation was conducted through a combination of AI-assisted analysis and structured feedback from human evaluators, including five university professors and 36 students randomly selected across diverse scientific disciplines. The primary aim was to identify potential ambiguities, internal contradictions, or robust counterevidence that might falsify the theory. A consensus emerged across all evaluators that CBET’s core elements — including its hierarchical classification of CBMs, its three axioms, and its two mechanisms — are conceptually clear, rigorous in reasoning with the support of extensive empirical evidence. No robust counterevidence was identified by these evaluators to falsify these core viewpoints.
Refinement of CBET
The CBET framework was iteratively refined based on empirical validation and critical feedback, to enhance its coherence, rigor, clarity, and explanatory power. Key refinements included redefining the hierarchies of CBMs, reformulating the three axioms and two mechanisms, streamlining the reasoning steps, updating the mathematical formulations, and redesigning the relevant figures. All elusive concepts (e.g., entropy) have been excluded from the final version. CBET is open to be further refined, if needed.
Other Issues
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Comparison of CBET to other theories, mathematical formulation of CBET, mathematical modeling of CBET, certain questions and answers about CBET, and the glossary of CBET.
Author Contributions
J.M.C. conceived, designed, performed, and financially supported this study; J.W.C. performed and financially supported this study; J.M.C. wrote the manuscript; all authors revised and approved the manuscript.
Acknowledgments
This study was supported by the High-Level Talent Fund of Foshan University (No. 20210036). The authors thank Meng Yang, Feng Tang, Zhen Liu, Hai Xiang, Ying Li, and many other people for their comments on this study. The authors thank Yiqing Chen who revised figures and the mathematical parts of this work.
Competing Interests
The authors declare no competing interests.
References
- Darwin, C. On the Origin of Species 1st edn (John Murray, 1859).
- Futuyma, D. J. & Kirkpatrick, M. Evolution 4th edn (Sinauer Associates, 2017).
- Chen, J. M. A new evolutionary theory deduced mathematically from entropy amplification. Chin. Sci. Bull. 45, 91–96 (2000). [CrossRef]
- Xie, P. The Aufhebung and Breakthrough of the Theories on the Origin and Evolution of Life (Science Press, 2014).
- Oparin, A. I. Chemistry and the origin of life. R. Inst. Chem. Rev. 2, 1–12 (1969).
- Fiore, M. Prebiotic Chemistry and Life’s Origin (Royal Society of Chemistry, 2022).
- Nowak, M. A., Tarnita, C. E. & Wilson, E. O. The evolution of eusociality. Nature 466, 1057–1062 (2010). [CrossRef]
- Birch, J. The Philosophy of Social Evolution (Oxford Univ. Press, 2017).
- Cowan, J. J. One of the first of the second stars. Nature 488, 288–289 (2012). [CrossRef]
- Razeghi, M. The Mystery of Carbon: An Introduction to Carbon Materials (IOP Publishing, 2019).
- Heck, P. R. et al. Lifetimes of interstellar dust from cosmic ray exposure ages of presolar silicon carbide. Proc. Natl Acad. Sci. USA 117, 1884–1889 (2020). [CrossRef]
- Li, Z. H. et al. Exceptional preservation of an extinct ostrich from the Late Miocene Linxia Basin of China. Vertebrata PalAsiatica 59, 229–248 (2021). [CrossRef]
- Reichle, D. E. The Global Carbon Cycle and Climate Change 2nd edn (Elsevier, 2023).
- Seager, S. Exoplanet habitability. Science 340, 577–581 (2013). [CrossRef]
- Rodriguez, L. E. et al. A geological and chemical context for the origins of life on early Earth. Astrobiology 24 (S1), S76–S106 (2024). [CrossRef]
- Olejarz, J., Iwasa, Y., Knoll, A. H. & Nowak, M. A. The Great Oxygenation Event as a consequence of ecological dynamics modulated by planetary change. Nat. Commun. 12, 3985 (2021). [CrossRef]
- Benton, M. J. The Red Queen and the Court Jester: species diversity and the role of biotic and abiotic factors through time. Science 323, 728–732 (2009). [CrossRef]
- De Capitani, J., Mutschler, H. The long road to a synthetic self-replicating central dogma. Biochemistry 62(7), 1221–1232 (2023). [CrossRef]
- Marchi, S. & Korenaga, J. The shaping of terrestrial planets by late accretions. Nature 641, 1111–1120 (2025). [CrossRef]
- Guo, X., Fu, S., Ying, J. & Zhao, Y. Prebiotic chemistry: a review of nucleoside phosphorylation and polymerization. Open Biol. 13, 220234 (2023). [CrossRef]
- Nam, I., Lee, J. K., Nam, H. G. & Zare, R. N. Abiotic production of sugar phosphates and uridine ribonucleoside in aqueous microdroplets. Proc. Natl Acad. Sci. USA 114, 12396–12400 (2017). [CrossRef]
- Dobson, C. M., Ellison, G. B., Tuck, A. F. & Vaida, V. Atmospheric aerosols as prebiotic chemical reactors. Proc. Natl Acad. Sci. USA 97, 11864–11868 (2000). [CrossRef]
- Griffith, E. C. & Vaida, V. In situ observation of peptide bond formation at the water-air interface. Proc. Natl Acad. Sci. USA 109, 15697–15701 (2012). [CrossRef]
- Trainer, M. G. Atmospheric prebiotic chemistry and organic hazes. Curr. Org. Chem. 17, 1710–1723 (2013). [CrossRef]
- Wang, S. Q. et al. Sexual selection promotes giraffoid head-neck evolution and ecological adaptation. Science 376, eabl8316 (2022). [CrossRef]
- Dawkins, R. The Selfish Gene (Oxford Univ. Press, 1976).
- Low, D. A. Advantage of an epigenetic switch in response to alternate environments. Proc. Natl Acad. Sci. USA 121, e2416356121 (2024). [CrossRef]
- Veigl, S. J., Vasilyeva, Z. & Müller, R. Scientific-intellectual movements in the post-truth age: The case of the Extended Evolutionary Synthesis. Soc. Stud. Sci. 2025. [CrossRef]
- Chen, J. M., Chen, J. W. & Zivieri, R. Systematically challenging three prevailing notions about entropy and life. Qeios 2024. [CrossRef]
- Fitzgerald, D. M. & Rosenberg, S. M. What is mutation? A chapter in the series: How microbes “jeopardize” the modern synthesis. PLoS Genet. 15, e1007995 (2019). [CrossRef]
- Fine, J. L. & Moses, A. M. An RNA condensate model for the origin of life. J. Mol. Biol. 437, 169124 (2025). [CrossRef]
- Hunt, T. Reconsidering the logical structure of the theory of natural selection. Commun. Integr. Biol. 7, e972848 (2014). [CrossRef]
- Zhu, S. C. et al. Key for hexagonal diamond formation: theoretical and experimental study. J. Am. Chem. Soc. 147, 2158–2167 (2025). [CrossRef]
- Stobart, C. C. & Moore, M. L. RNA virus reverse genetics and vaccine design. Viruses 6, 2531–2550 (2014). [CrossRef]
- Hutchison, C. A. et al. Design and synthesis of a minimal bacterial genome. Science 351, aad6253 (2016). [CrossRef]
- Prigogine, I. Time, structure and fluctuation. Science 201, 777–785 (1978). [CrossRef]
- Ramstead, M. J. D., Badcock, P. B. & Friston, K. J. Answering Schrödinger’s question: A free-energy formulation. Phys. Life Rev. 24, 1–16 (2018). [CrossRef]
- Friston, K. The free-energy principle: a unified brain theory? Nat. Rev. Neurosci. 11, 127–138 (2010). [CrossRef]
- Rudman, L. A. & Saud, L. H. Justifying social inequalities: The role of social Darwinism. Pers. Soc. Psychol. Bull. 46, 1139–1155 (2020). [CrossRef]
- Sober, E. & Wilson, D. S. Unto Others: The Evolution and Psychology of Unselfish Behavior (Harvard Univ. Press, 1998).
- Eldredge, N., Pievani, T., Serrelli, E. & Tëmkin, I. Evolutionary Theory: A Hierarchical Perspective (Univ. Chicago Press, 2016).
- Miller, J. H. & Page, S. E. Complex Adaptive Systems: An Introduction to Computational Models of Social Life (Princeton Univ. Press, 2007).
- Sabater, B. Entropy Perspectives of Molecular and Evolutionary Biology. Int. J. Mol. Sci. 23, 4098 (2022). [CrossRef]
- Schrödinger, E. What is Life (Cambridge Univ. Press, 2012).
- Bejan, A. The principle underlying all evolution, biological, geophysical, social and technological. Phil. Trans. R. Soc. A 381, 20220288 (2023). [CrossRef]
- Dewar, R. C. Maximum entropy production and plant optimization theories. Phil. Trans. R. Soc. B 365, 1429–1435 (2010). [CrossRef]
- Borgnakke, C. & Sonntag, R. E. Fundamentals of Thermodynamics (Wiley, 2022).
- Schmitt-Kopplin, P. et al. High molecular diversity of extraterrestrial organic matter in Murchison meteorite revealed 40 years after its fall. Proc. Natl Acad. Sci. USA 107, 2763–2768 (2010). [CrossRef]
- Yang, Y., Wang, Z., Bai, J. & Qiao, H. Prebiotic peptide synthesis: How did longest peptide appear? J. Mol. Evol. 93, 193–211 (2025). [CrossRef]
- Yang, B., Liu, A., Li, X. & Miao, Z. Research progress on mRNA drug delivery strategies in the nervous system. Nanomedicine 20, 1955–1970 (2025). [CrossRef]
Figure 1.
| Nine hierarchies (H1−H9) of carbon-based materials (CBMs) from carbon atoms and carbon-containing molecules (CCMs) to social organizations. Arrows indicate the stepwise or cross-hierarchy formation or degradation of higher-hierarchy CBMs.
Figure 1.
| Nine hierarchies (H1−H9) of carbon-based materials (CBMs) from carbon atoms and carbon-containing molecules (CCMs) to social organizations. Arrows indicate the stepwise or cross-hierarchy formation or degradation of higher-hierarchy CBMs.
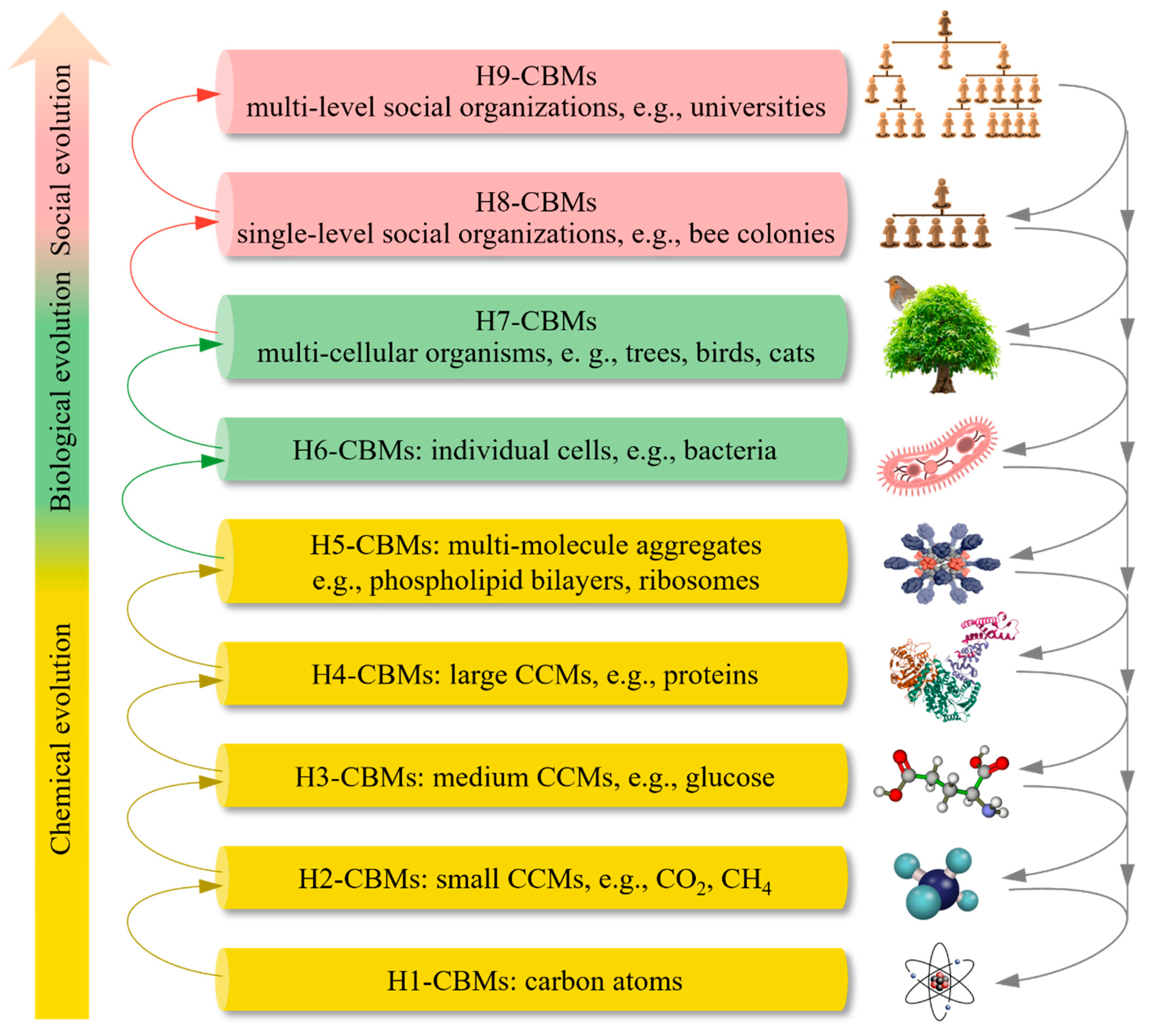
Figure 2.
| The reasoning structure of the Carbon-Based Evolutionary Theory (CBET) and its relationship to Darwinian theory. The derivation of CBET comprises eight reasoning steps (Steps 1–8), which are organized into five integrative tiers indicated by arrows with different colors. The first four tiers derive the three axioms, the spriodynamic feedback mechanism, the natural selection mechanism, and the reasons for complexity enhancement, respectively; the fifth tier represents their synergy. For comparison, Darwinian theory is shown to align primarily with Step 6 of this reasoning structure.
Figure 2.
| The reasoning structure of the Carbon-Based Evolutionary Theory (CBET) and its relationship to Darwinian theory. The derivation of CBET comprises eight reasoning steps (Steps 1–8), which are organized into five integrative tiers indicated by arrows with different colors. The first four tiers derive the three axioms, the spriodynamic feedback mechanism, the natural selection mechanism, and the reasons for complexity enhancement, respectively; the fifth tier represents their synergy. For comparison, Darwinian theory is shown to align primarily with Step 6 of this reasoning structure.
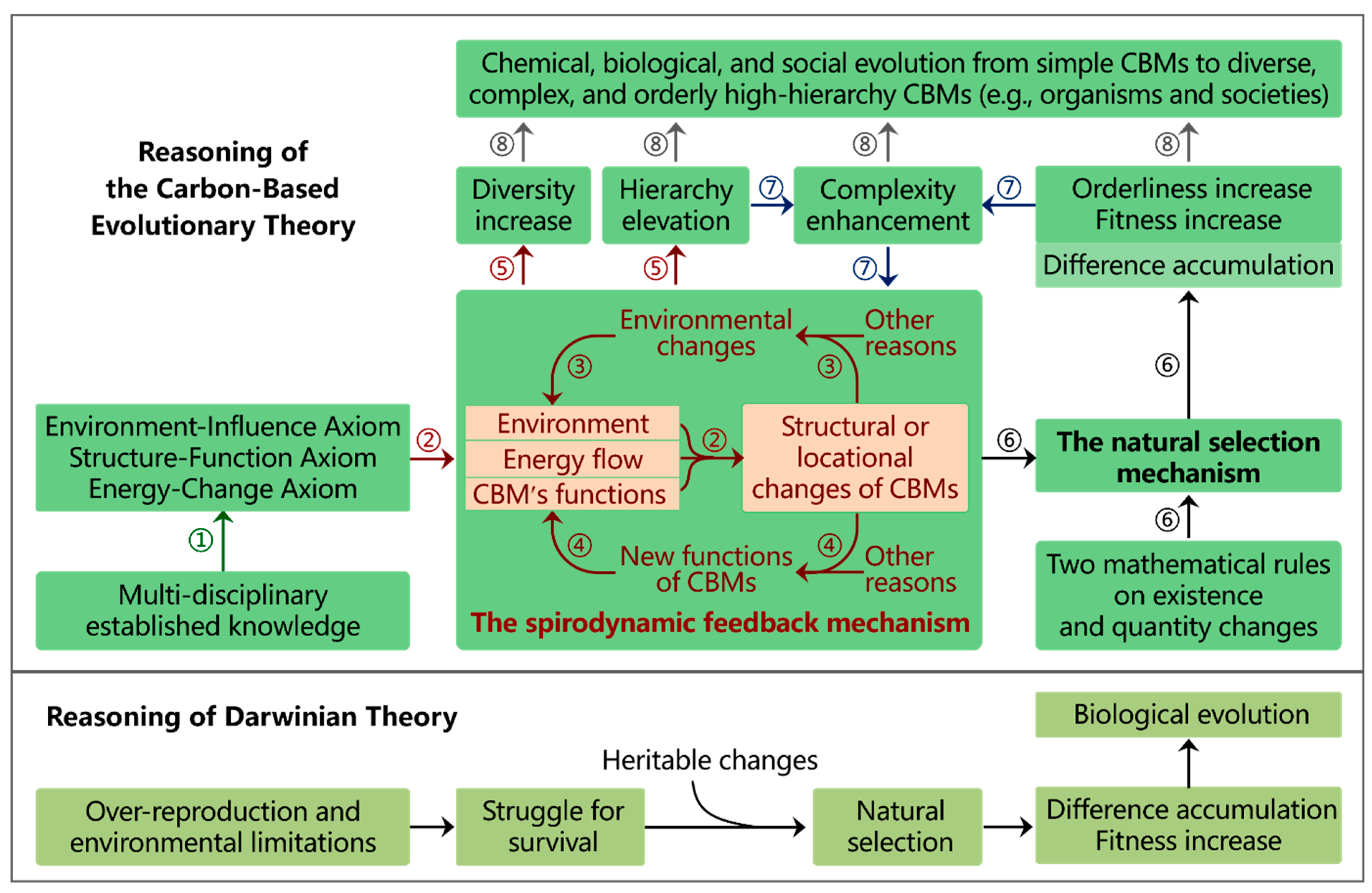
Figure 3.
The methodology of Darwinian theory and the Carbon-Based Evolutionary Theory (CBET) starting from integrative reasoning. CBMs: carbon-based materials.
Figure 3.
The methodology of Darwinian theory and the Carbon-Based Evolutionary Theory (CBET) starting from integrative reasoning. CBMs: carbon-based materials.

Table 1.
Selected differences between two theories of evolution in biology.
| Darwinian theory | The Carbon-Based Evolutionary Theory | |
|---|---|---|
| Scope | Biological evolution | Chemical, biological, and social evolution |
| Core concept | Organisms | Carbon-based materials (CBMs) |
| Mechanisms | Natural selection that shapes evolution | Spriodynamic feedback (drives evolution) and natural selection (shapes evolution) |
| Fitness calculation | The number of an individual’s offspring in the next generation | The formation-to-degradation ratio of a CBM over a time interval |
| Life’s origin | Did not explain life’s origin | Schematically accounts for life’s origin |
| Inclusiveness | Could not fully integrate neutral or detrimental mutations with natural selection | Integrates the prevalence of neutral or detrimental mutations with natural selection |
| Changes under selection | Focused on heritable genetic changes | Highlights the roles of genetic, epigenetic, and non-heritable changes |
| Primary significance | Unveiled the natural selection mechanism; challenged creationism and inheritance of acquired traits | Establishes a coherent framework theory that explicitly and schematically explains chemical, biological, and social evolution |
| Social significance | Could not establish natural-science-grounded foundations for social sciences or for harmonious societal development | Establishes natural-science-grounded foundations for social sciences and for harmonious societal development |
Table 2.
The 27 representative facts used to validate the spriodynamic feedback mechanism.
| Order | Description of the fact |
|---|---|
| 1 | Charcoal combustion which consumes oxygen and forms carbon dioxide |
| 2 | Diamond synthesis from graphite under certain conditions [33] |
| 3 | Water adsorption of activated carbon |
| 4 | CO2, NH3, and other molecules form amino acids via chemical reactions [2,5] |
| 5 | Methane explosion in coal mines |
| 6 | Evaporation of ethanol from an open bottle |
| 7 | The dehydration condensation of amino acids into peptides in tubes |
| 8 | The binding of gentamicin to the 30S ribosomal subunit of some bacteria |
| 9 | The degradation of glucose into carbon dioxide and water in cells |
| 10 | Some recombinant proteins self-assembling into virus-like particles |
| 11 | DNA synthesis catalyzed by a protein |
| 12 | Starch degradation in the stomach |
| 13 | Inputting certain DNA molecules into certain cells can create new viruses [34] |
| 14 | Inputting certain DNA molecules into certain cells can create new cells [35] |
| 15 | The formation of bilayer vesicles by amphiphilic lipid molecules |
| 16 | The development of a fertilized egg into a tadpole |
| 17 | Escherichia coli absorbs glucose via its membrane carrier proteins |
| 18 | The reproduction of bacteria within chicken intestines |
| 19 | A group of ant individuals form a eusocial ant colony |
| 20 | Some trees undergo spring budding and autumn leaf shedding |
| 21 | A car collision causes human casualties |
| 22 | The formation of a basketball team by a group of young people |
| 23 | Leafcutter ants harvest foliage to cultivate symbiotic fungi in their nests |
| 24 | The destruction of a beehive by a forest fire |
| 25 | Several institutions merged into Paris-Saclay University in 2018 |
| 26 | University of the Arts Philadelphia was closed in 2024 |
| 27 | The ongoing war between Russia and Ukraine |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



