Submitted:
07 July 2026
Posted:
08 July 2026
You are already at the latest version
Abstract
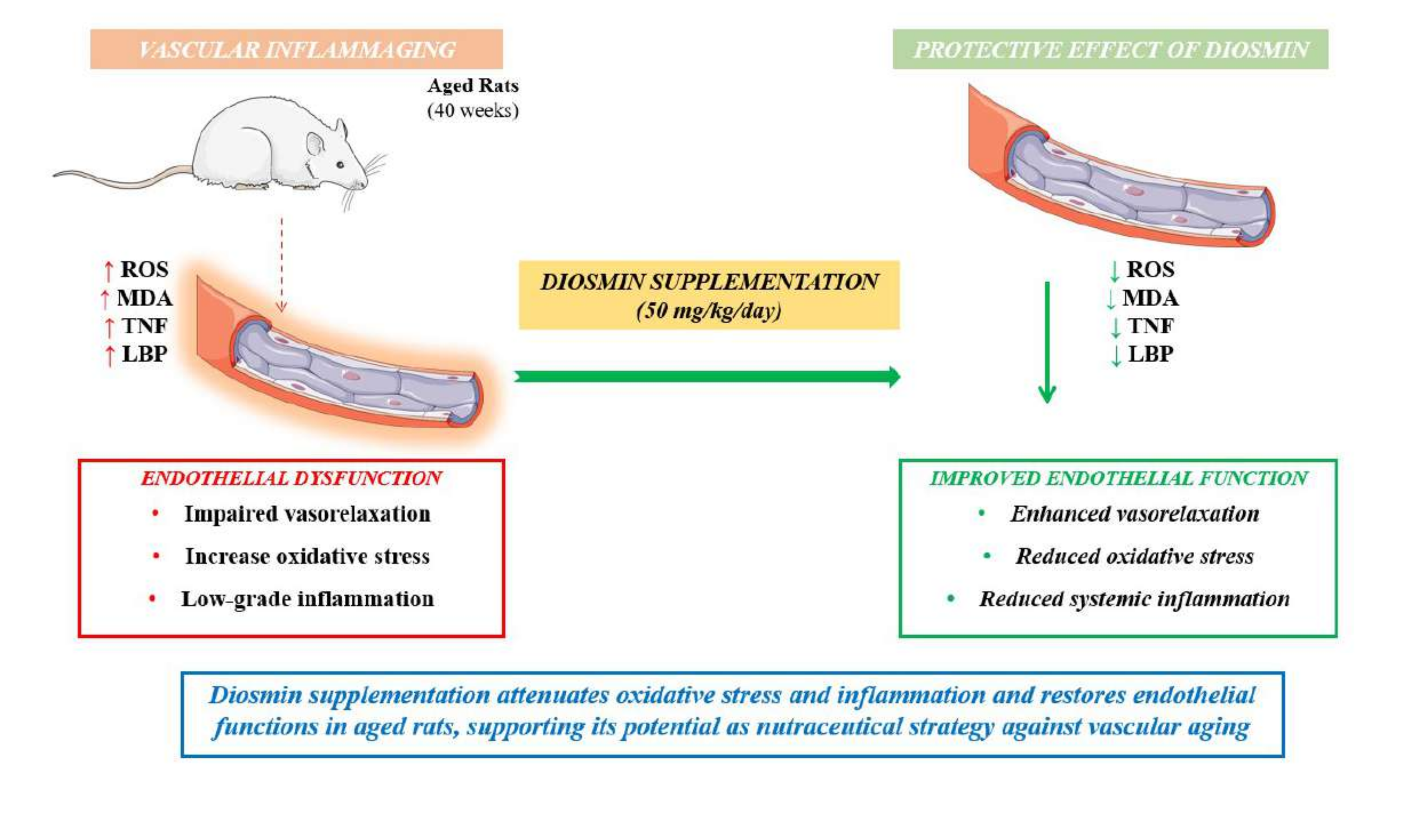
Vascular aging is driven by a self-sustaining interplay between oxidative stress and chronic low-grade inflammation, collectively referred to as inflammaging. Diosmin, a cit-rus-derived flavonoid with antioxidant and anti-inflammatory properties, has not previ-ously been investigated in this context. Young (10-week-old) and aged (40-week-old) male Sprague–Dawley rats received diosmin (50 mg/kg/day) or vehicle for 3 months. Vascular function was assessed in mesenteric resistance arteries using pressure myography, while oxidative stress was evaluated by dihydroethidium and mitoSOX staining. Circulating malondialdehyde (MDA), tumor necrosis factor (TNF) and lipopolysaccharide-binding protein (LBP) were measured and vascular expression of Enos, Gpx, Sod2 and Xdh was an-alyzed. Aging was associated with impaired endothelium-dependent vasorelaxation, increased vascular ROS production and elevated circulating MDA and LBP levels. Chronic diosmin supplementation partially restored endothelial function and significantly reduced both NADPH oxidase–derived and mitochondrial ROS in aged vessels. These effects were ac-companied by decreased plasma MDA and LBP levels, while TNF remained unchanged. Diosmin did not significantly affect Enos, Gpx, or Sod2 gene’s expression, but attenuated the age-related upregulation of Xdh. Overall, diosmin mitigates vascular inflammaging by reducing ROS burden and improving nitric oxide bioavailability, highlighting its potential to counteract age-related vascular dysfunction through modulation of redox–inflammatory crosstalk.
Keywords:
diosmin
; inflammaging
; vascular aging
; oxidative stress
1. Introduction
The term inflammaging—a fusion of “inflammation” and “aging”- refers to findings from various preclinical and clinical studies that highlight a close relationship between the processes that lead to aging and the presence of a persistent and systemic mild inflammatory response, without visible clinical symptoms (latent), but which can be the basis for several chronic degenerative diseases, commonly observed in the elderly [1]. The condition of inflammaging seems to result from a progressive reduction of the biological reserve mechanisms that allow the body cells to counteract changes and challenges to the tissue homeostasis. These changes depend on the environmental exposure and chronic accumulation of inflammatory damages over the course of an individual’s existence, inducing an impairment of the biological reserve greater than that associated solely with aging [1].
The condition of inflammaging is characterized by high circulating levels of pro-inflammatory markers, including interleukin (IL)-1β, IL-6, IL-8, IL-13, IL-18, C-reactive protein, interferon (IFN)-α and IFN-β, transforming growth factor (TGF)-β, tumor necrosis factor (TNF), and serum amyloid type A [1,2]. Although initially these inflammatory alterations were considered an inevitable consequence of aging, an increasing body of experimental and human data suggests that they may represent also the primary causes of aging, thus configuring the occurrence of a vicious circle of events where aging represents the source of inflammation and, at the same time, the pathological target of its consequences [3].
Despite the mechanisms underlying the above vicious circle remain largely to be defined, the increase in oxidative stress with advancing age, resulting from an impairment of antioxidant defenses and an increase in the production of oxygen free radicals, seems to play a central role as mediator of the relationship between inflammation and cellular aging. Indeed, an increase in the production of oxygen free radicals, especially of mitochondrial origin, represents an important stimulus to the activation of the inflammatory response [4] and, simultaneously, it can damage also the major cellular macro-elements (proteins, lipids, nucleic acids). In the early stages, this damage can be repaired by cellular defense mechanisms. However, its chronicization makes such intracellular defenses insufficient, leading to malfunctioning of enzymes and cellular barrier structures as well as to altered gene expression that are believed to be at the basis of the evolution of organic aging [5]. At the same time, the inflammatory response, sustained by the increase in oxidative stress, is almost invariably associated with an increase in the production of oxygen free radicals, thus favoring a pro-oxidant environment that could support the onset and persistence of a chronic inflammatory process [1,2].
The tight relationship between oxidative stress, inflammation and aging is confirmed by the scientific literature that has clearly identified inflammation and oxidative stress as the main actors underlying the development of chronic degenerative diseases commonly occurring in the elderly, such as Alzheimer’s, type II diabetes, and cardiovascular diseases [1,2].
Recently, diosmin, a citrus-derived bioflavonoid used clinically to treat chronic venous disorders, has emerged as a promising agent for addressing several conditions, including those related to aging. [6,7]. It exerts anti-inflammatory effects by inhibiting the expression of pro-inflammatory cytokines through the NF-κB pathway and reducing T cell receptor activity. Diosmin also has antioxidant properties, scavenging oxygen free radicals and lowering oxidative stress biomarkers [6,7]. However, to date, no studies have assessed the potential of diosmin to counteract vascular inflammaging. Based on this background, the present study was designed to evaluate the potential beneficial effects of diosmin in a rat model of vascular inflammaging.
2. Materials and Methods
2.1. Experimental Design and Administration
Aged (40-weeks-old, 500-600 g of body weight) and young (10-weeks-old, 200-225 g of body weight) Albino male Sprague–Dawley rats were purchased from ENVIGO S.r.l (San Pietro al Natisone, UD, Italy) and employed throughout the study. Animals had free access to standard laboratory chow and tap water and were given at least 1 week to acclimatize before the beginning of experimental procedures.
They have been housed, three in a cage, in temperature-controlled rooms at 22–24 °C and 50–60% humidity with a 12-h light cycle. Animal care and handling were in accordance with the provisions of the European Community Council Directive 210/63/UE, implemented by the Italian Government. The experiments have been approved by the Ethical Committee for Animal Experimentation of the University of Pisa and by the Italian Ministry of Health (authorization n 1023/2023-PR). Animals were monitored daily to asses any sign of distress and all efforts were made to minimize animal suffering.
Aged rats were considered as a natural model of vascular inflammaging. 32 animals were randomly assigned to four groups, as follows: a) young rats + vehicle (n=8); b) young rats + diosmin 50 mg/kg [8] (n=8); c) aged rats + vehicle (n=8); d) aged rats+ diosmin 50 mg/kg (n=8). Animals were treated with diosmin or vehicle daily for three months by oral gavage. Diosmin was suspended in 0.2 ml of methocell 3%. After the treatment period, animals were anaesthetized and sacrificed to collect tissues.
The investigational product is a diosmin-based ingredient (μsmin® Plus) standardized to > 80% diosmin and 90% total flavonoids, supplied by Giellepi S.p.A. (Milan, Italy).
2.2. Preparation of Small Mesenteric Arteries for Functional Experiments
After dissection, the first branch of mesenteric artery was placed in cold (4 °C) physiological salt solution (PSS) containing (in mmol/L): NaCl 120, NaHCO3 25, KCl 4.7, KH2PO4 1.18, MgSO4 1.18, CaCl2 2.5, EDTA 0.026, and glucose 5.5, as previously published [9]. A second-order branch of the mesenteric arterial tree (≈2 mm in length) was dissected and mounted on 2-glass microcannulas in a pressurized myograph, as previously described [9]. Vessels were equilibrated for 60 min under constant intraluminal pressure (45 mmHg) in warmed (37 °C) and bubbled (95% air and 5% CO2) PSS, at pH 7.4. Vessels were considered viable and used if they constricted >70% of their resting lumen diameter in response to an extraluminal application of high-potassium solution (125 mmol/L of KCl) containing 100 μmol/L of noradrenaline (NA).
2.3. Detection of Vascular Superoxide Anion Generation
The in situ production of superoxide anion from 30 mm frozen mesenteric vessel sections was evaluated at the confocal microscope by means of the fluorescent dye dihydroethidium [(DHE), Sigma], as previously described [10]. Three slides per segment were analyzed simultaneously after incubation with Krebs solution at 37 °C for 30 min. Krebs-HEPES buffer containing 2 μM DHE was then applied to each section and evaluated under fluorescence microscopy. In the presence of superoxide, DHE undergoes oxidation and intercalates in cell DNA, thus staining the nucleus with red fluorescence (excitation at 488 nm, emission 610 nm). The percentage of arterial wall area stained with the red signal was normalized to the total area examined and quantified using an imaging analysis software (McBiophotonics Image J; National Institutes of Health, Bethesda, MD, United States).
2.4. MitoSOX Red Staining
The level of mitochondrial ROS was determined by staining mesenteric arteries with MitoSOX Red dye (Life Technologies, USA). Briefly, mesenteric arteries have been washed 3 times with PBS buffer and then loaded with MitoSOX Red in the dark at a temperature of 37 °C for 30 min. A fluorescence microscope (Zeiss, Germany) was used to detect fluorescent signals with an excitation wavelength at 510 nm and emission wavelength at 580 nm.
2.5. Plasma Levels of TNF and MDA
Serum levels of TNF and MDA were analyzed by ELISA kit (Prodotti Gianni, Milan, Italy), as previously described [11]. Aliquots of 100 μL were used for the test. TNF levels were expressed as a percentage of Young+Vehicle (% vs Young+Vehicle) of serum.
For the evaluation of MDA plasma levels, a colorimetric assay kit (Nordic Biosite, Stockholm, Sweden) was employed and performed following manufacture instructions. MDA levels were indicated as ng/ml of serum.
2.6. Evaluation of lipopolysaccharide-binding protein (LBP) levels in plasma
Blood samples were centrifuged for 5 minutes at 4000g at 2–8 °C and, after the centrifugation, supernatants were collected. Aliquots (100 μl) were used for the assay. LBP levels were expressed as ρg/mL of plasma.
2.7. Real Time qPCR
Total RNA was extracted using TRIzol Reagent (Invitrogen, Carlsbad, CA) according to the manufacturer’s recommendations. Before extraction, vascular samples were manually lysed using Microtube Pestles (Starlab, Milan, Italy) and then sonicated via Ultrasonic Processor (Merck, Milan, Italy). Conversion of total cellular RNA to cDNA was carried out using High-capacity cDNA reverse transcription kit (Thermofisher, Milan, Italy) in a final volume of 20 μl, according to manufacturer instructions. Real-time PCR was performed using the SYBR Select Master Mix (Applied Biosystems, Thermo Fisher Scientific, Milan, Italy) on a CFX Duet Real-Time PCR System (Bio-Rad, Milan, Italy) according to the manufacturer’s instructions. Gapdh was used as endogenous control to normalise RNA concentration. The amplification program consisted of 1 cycle at 95 °C for 10 min, followed by 40 cycles with a denaturing phase at 95˚C for 30 s and an annealing and elongation phase of 1 min at 60˚C. A melting curve analysis was performed after amplification to verify the accuracy of the amplicon. Differences in Ct values between test genes and endogenous controls (ΔCt) and between different groups and control group (ΔΔCt) used as negative power of 2 (2-ΔΔCt) were calculated and used for statistical analysis.
2.8. Statistical Analysis
The experimental unit was defined as a single animal (rat). Data were expressed as mean ± SEM or violin plot and analyzed with GraphPad Prism 7.0 (GraphPad Software Inc., United States). Statistical significances were determined by unpaired t-test, one-way ANOVA (when appropriate) followed by Tukey’s post hoc test. A p value < 0.05 was considered statistically significant.
3. Results
3.1. Isolation of Mesenteric Arteries and in Vitro Assessment of Their Structure and Reactivity
No differences in vascular remodeling were observed in young rats, although an improvement in media/lumen ratio was observed in aged rats treated with diosmin (Figure 1A). In vascular relaxation assays, no differences between treatment groups were observed in young rats (Figure 1B). In aged rats, chronic supplementation with diosmin alone induced a partial restoration of vascular relaxation to ACh (Figure 1C,D) also in terms of L-NAME vasoinhibition (Figure 1E), a reliable marker of endothelial function. Taken together, our data indicate that chronic diosmin supplementation is able to partially rescue the vascular phenotype in aged rats.
3.2. NADPH-Oxidase and Mitochondrial-Derived Reactive Oxygen Species Production in the Vascular Wall of Second Order Mesenteric Arterioles
Fluorescence assays showed that in the vascular wall of young rats no differences were detected in terms of mtROS (assessed via mitoSOX staining) or NADPH-oxidase derived ROS (assessed via DHE staining) between untreated and diosmin-treated animals. In aged rats, levels of mtROS and DHE were significantly higher, as compared with young animals. In this setting, chronic supplementation with diosmin was able to mitigate the mtROS and NAPDH-oxidase derived ROS burden (Figure 2A,B). This further confirmed our functional data, showing that chronic diosmin supplementation is able to partially mitigate the reactive oxygen species levels in the vascular wall of aged rats.
3.3. Evaluation of Plasma Inflammatory Parameters TNF and Oxidative Stress Index (MDA)
MDA levels in plasma from aged animals administered with vehicle were significantly increased, as compared with young animals and young animals administered with diosmin (Figure 3A). Administration of aged animals with diosmin significantly decreased MDA. No significant changes were observed in TNF levels (Figure 3B) in young and aged rats treated with vehicle or diosmin.
3.4. Evaluation of Plasma LBP Levels
LBP levels in plasma from aged animals administered with vehicle were enhanced compared to young animals administered with vehicle (Figure 4). Administration of aged animals with diosmin significantly reduced such parameter.
3.5. Enos, Sod2, Gpx and Xdh Gene Expression in Vascular Tissues
No significant differences were observed in Enos expression among groups, with diosmin treatment in aged animals showing no detectable effect compared to untreated aged controls (Figure 5A). Gpx expression showed a tendency to increase in aged animals treated with diosmin compared to untreated aged rats; although, this difference did not reach statistical significance (Figure 5B). Similarly, Sod2 levels were not significantly altered by diosmin treatment in either young or aged groups (Figure 5C), indicating no major effect on this antioxidant enzyme.
In contrast, Xdh expression was markedly elevated in aged animals compared to young controls, and this increase was significantly reduced by diosmin treatment in the aged group (Figure 5D), suggesting a potential modulatory effect of diosmin on age-associated oxidative stress pathways.
4. Discussion
The present study provides novel evidence that chronic diosmin supplementation exerts protective vascular effects in a rat model of vascular inflammaging. Specifically, diosmin partially restored endothelial function and improved vascular wall structure in aged animals, while attenuating oxidative stress–related alterations and inflammatory markers. These findings support the concept that diosmin effectively targets key pathophysiological mechanisms underlying vascular aging, including endothelial dysfunction, reactive oxygen species (ROS) accumulation, and chronic low-grade inflammation [1].
Vascular aging is a complex, multifactorial process characterized by structural remodeling, endothelial impairment, and increased arterial stiffness, all of which contribute to the heightened cardiovascular risk observed in elderly populations. A growing body of evidence identifies inflammaging, a persistent, low-grade inflammatory state, as a central driver of these alterations, promoting oxidative stress, impairing nitric oxide (NO) bioavailability, and fostering maladaptive vascular remodeling [1,12]. In this context, our findings suggest that diosmin interrupts, at least in part, the self-reinforcing interplay between oxidative stress and inflammation that sustains vascular aging.
Endothelial dysfunction represents one of the earliest and most clinically relevant manifestations of vascular aging. In the present study, aged animals exhibited impaired acetylcholine (ACh)-induced vasorelaxation, consistent with reduced endothelial NO bioavailability. Chronic diosmin supplementation significantly improved endothelium-dependent relaxation, as evidenced by partial restoration of ACh responses and normalization of L-NAME–mediated inhibition, indicating enhanced endothelial NO contribution. These findings suggest that diosmin improves endothelial function primarily through modulation of redox balance and inflammatory signaling pathways that converge on NO bioactivity [13].
Mechanistically, previous studies have demonstrated that diosmin inhibits nuclear factor-κB (NF-κB) signaling and reduces the expression of pro-inflammatory cytokines such as TNF, IL-1β, and IL-6 [6,7]. Given that NF-κB activation negatively regulates eNOS function and promotes endothelial activation, suppression of this pathway likely contributes to the improved vascular responses observed in the present study. In parallel, the well-documented antioxidant properties of diosmin, including free radical scavenging and inhibition of lipid peroxidation, may further enhance NO bioavailability by limiting the formation of peroxynitrite and other reactive nitrogen species [6,7].
A key finding of this study is the reduction of both NADPH oxidase–derived and mitochondrial ROS in the vascular wall of aged animals following diosmin treatment. Oxidative stress is a pivotal driver of vascular aging and is largely mediated by activation of NADPH oxidase isoforms (notably NOX2 and NOX4) and mitochondrial dysfunction [14,15]. The observed decrease in dihydroethidium (DHE) and mitoSOX fluorescence signals indicates that diosmin effectively suppresses ROS generation from both enzymatic and mitochondrial sources, suggesting a broad and integrated antioxidant mechanism of action.
The molecular pathways underlying these effects remain incompletely defined; however, flavonoids such as diosmin and its aglycone diosmetin are known to modulate redox-sensitive transcription factors, including NF-κB and Nrf2, thereby influencing the expression of endogenous antioxidant systems [7]. Consistent with this framework, diosmin-treated aged animals exhibited increased SOD expression, although not significant, indicative of activation of compensatory antioxidant defenses. By simultaneously reducing ROS production and enhancing antioxidant capacity, diosmin likely disrupts the feed-forward loop linking oxidative stress, mitochondrial dysfunction, and vascular inflammation [16].
In line with these observations, transcriptional profiling of oxidative stress–related genes revealed a selective and mechanistically informative pattern of modulation by diosmin. Notably, while Enos, Sod2 and Gpx expression remained largely unchanged across experimental groups, aging was associated with a pronounced upregulation of xanthine dehydrogenase (Xdh), which was significantly attenuated following diosmin treatment. Given the well-established role of Xdh/xanthine oxidase as a major enzymatic source of superoxide and hydrogen peroxide in the vasculature [17], its downregulation suggests that diosmin preferentially targets upstream ROS-generating systems rather than broadly inducing antioxidant gene expression. This interpretation is further supported by the absence of compensatory upregulation of canonical antioxidant enzymes, indicating that the observed redox improvement is unlikely to be driven by enhanced scavenging capacity. Instead, diosmin appears to mitigate oxidative stress at its source, thereby reducing the overall oxidative burden [18]. Importantly, the lack of transcriptional changes in Enos, despite clear functional improvement in endothelium-dependent vasorelaxation, strongly supports a model in which diosmin enhances nitric oxide bioavailability primarily by limiting ROS-mediated NO degradation, rather than by increasing Enos expression per se. Collectively, these findings provide mechanistic evidence that diosmin exerts vascular protection in aging through targeted suppression of pro-oxidant pathways, effectively uncoupling the feed-forward loop between ROS overproduction and endothelial dysfunction that characterizes vascular inflammaging.
Consistent with the systemic presence of inflammaging, aged animals displayed elevated plasma levels of malondialdehyde (MDA), a marker of lipid peroxidation, as well as increased lipopolysaccharide-binding protein (LBP), indicative of metabolic endotoxemia and impaired gut barrier function [19,20]. Diosmin treatment significantly reduced MDA and LBP levels supporting its combined antioxidant and anti-inflammatory effects at the systemic level.
Notably, the reduction in circulating LBP integrates with the observed suppression of Xdh expression and vascular ROS production, providing a coherent multi-level mechanism underlying diosmin protective effects. LBP is a surrogate marker of endotoxin exposure and gut-derived inflammation [20], which represent important upstream triggers of systemic oxidative stress and endothelial dysfunction. Thus, the decrease in LBP suggests that diosmin may attenuate circulating pro-inflammatory stimuli that contribute to the activation of vascular ROS-generating systems, including Xdh/xanthine oxidase pathways. In this framework, reduced metabolic endotoxemia may alleviate redox-sensitive signaling cascades, ultimately limiting ROS overproduction and preserving nitric oxide bioavailability [21]. These findings support an integrated model in which diosmin acts both locally, by suppressing vascular pro-oxidant enzymes, and systemically, by reducing gut-derived inflammatory inputs, thereby disrupting the bidirectional amplification between oxidative stress and inflammation that characterizes vascular inflammaging. Although direct assessment of intestinal permeability or microbiota composition was not performed, this interpretation is consistent with previous evidence indicating that flavonoids can modulate gut barrier function and host–microbiota interactions [22,23].
Although no significant changes were observed in circulating TNF levels, this does not exclude anti-inflammatory activity, as systemic cytokine measurements may not accurately reflect local vascular inflammation or subtle changes in low-grade inflammatory signaling. It is plausible that diosmin exerts its primary anti-inflammatory effects at the tissue level, where modulation of redox-sensitive pathways and endothelial activation occurs independently of overt systemic cytokine suppression.
The reduction in LBP is particularly noteworthy, as increased circulating LBP in aging has been associated with translocation of bacterial endotoxins from the gut, contributing to systemic inflammation and vascular dysfunction. Diosmin has been reported to improve intestinal barrier integrity and modulate gut microbiota composition [24], suggesting that its vascular protective effects may, in part, be mediated through modulation of the gut–vascular axis. This observation highlights a potential multi-organ mechanism of action, integrating metabolic, inflammatory, and vascular pathways.
Structural remodeling of the vasculature is another hallmark of aging. In the present study, aged animals exhibited an increased media-to-lumen ratio in mesenteric resistance arteries, indicative of hypertrophic remodeling. Chronic diosmin supplementation partially reversed these structural alterations, suggesting a protective effect on vascular architecture. These improvements may result from reduced oxidative injury to vascular smooth muscle cells and attenuation of pro-fibrotic signaling pathways, including those mediated by TGF-β and matrix metalloproteinases, both of which are regulated by redox-sensitive mechanisms [25].
Taken together, our findings support a mechanistic model in which diosmin disrupts the reciprocal amplification between oxidative stress and inflammation that drives vascular aging. Excess ROS production from mitochondrial dysfunction and NADPH oxidase activation promotes inflammatory signaling, endothelial dysfunction, and structural remodeling, establishing a self-perpetuating cycle of vascular damage [26]. Importantly, diosmin mechanism of action likely involves both direct radical scavenging and transcriptional regulation of redox-sensitive genes. Activation of the Nrf2 pathway represents a plausible unifying mechanism, potentially accounting for both enhanced antioxidant defenses and improved vascular phenotype [27], and warrants further investigation.
Diosmin is already widely used in clinical practice for chronic venous insufficiency and hemorrhoidal disease due to its venotonic and anti-inflammatory properties [7]. The present findings expand its therapeutic relevance, suggesting that diosmin may be repurposed for the prevention or treatment of vascular aging and related cardiovascular disorders. Its ability to simultaneously target oxidative stress, inflammation, and endothelial dysfunction positions it as a promising candidate among nutraceutical and pharmacological strategies aimed at modulating inflammaging.
Similar vascular protective effects have been reported for other flavonoids, including quercetin and hesperidin; however, diosmin may offer pharmacokinetic advantages, such as improved oral bioavailability and longer plasma half-life, potentially enhancing its efficacy in chronic settings [28,29]. In this regard, previous pharmacokinetic studies have shown that the diosmin-based formulation investigated in this study exhibits superior bioavailability compared with micronized diosmin in both rats and humans [30]. These considerations, together with the mechanistic insights provided here, support further translational investigation.
However, several limitations should be acknowledged. The present study was conducted exclusively in male animals, and sex-dependent differences in vascular aging and flavonoid responsiveness are well recognized. Additionally, although functional and biochemical improvements were clearly demonstrated, the specific molecular mediators of diosmin action, i.e., NF-κB inhibition, Nrf2 activation, and modulation of individual NOX isoforms, were not evaluated. Future studies incorporating gene and protein expression analyses will be essential to clarify these pathways. Moreover, the inflammatory profile was limited to a small set of circulating cytokines, and broader omics-based approaches may provide deeper insight into diosmin’s immunomodulatory effects. Finally, the translational relevance of these findings should be validated in long-term studies and in clinically relevant models of cardiovascular disease.
5. Conclusions
This study demonstrates that chronic diosmin supplementation improves vascular structure and function in aged animals by attenuating key features of vascular inflammaging. These effects are mediated by suppression of ROS generation from both NADPH oxidase and mitochondrial sources, restoration of endothelial function, and reduction of systemic oxidative and inflammatory markers. Collectively, these findings support a model in which diosmin interrupts the pathogenic cycle linking oxidative stress, inflammation, and vascular dysfunction, thereby slowing the progression of vascular aging. Given its established safety profile and clinical use, diosmin emerges as a promising candidate for targeting age-related vascular disorders. However, the extent to which these effects may be shared by other diosmin formulations remains to be established. Further studies are warranted to validate these findings in human populations and to elucidate the molecular pathways underlying its protective effects.
Author Contributions
Conceptualization, L.A; methodology, L.A.; validation, G.V., C.D.S., L.C., A.M., F.C. and E.D. ;formal analysis, G.V., C.D.S., L.C., A.M., F.C. and E.D.; investigation, L.A., G.V., C.D.S., L.C., A.M., F.C. and E.D; data curation, G.V., C.D.S., L.C., A.M., F.C. and E.D; Writing—original draft preparation, L.A., G.V.; Writing—review and editing, L.A., M.F., A.V., N.B., R.C., C.P. and A.M.; supervision, L.A., M.F., C.P. and A.V.; project administration, L.A.; funding acquisition, L.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Giellepi S.p.A.
Institutional Review Board Statement
The animal study protocol was approved by the Ethics Committee for Animal Experimentation of the University of Pisa and by the Italian Ministry of Health (authorization n 1023/2023-PR).
Data Availability Statement
The data supporting the conclusions of this article will be made available by the authors on request.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ACh | Acetylcholine |
| ANOVA | Analysis Of Variance |
| cDNA | Complementary DNA |
| Ct | Cycle Threshold |
| DHE | Dihydroethidium |
| EDTA | Ethylenediaminetetraacetic Acid |
| ELISA | Enzyme-Linked Immunosorbent Assay |
| eNOS | Endothelial Nitric Oxide Synthase |
| GAPDH | Glyceraldehyde-3-Phosphate Dehydrogenase |
| GPx | Glutathione Peroxidase |
| IFN | Interferon |
| IL | Interleukin |
| LBP | Lipopolysaccharide-Binding Protein |
| L-NAME | N(Ω)-Nitro-L-Arginine Methyl Ester |
| MDA | Malondialdehyde |
| mtROS | Mitochondrial Reactive Oxygen Species |
| NA | Noradrenaline |
| NADPH | Nicotinamide Adenine Dinucleotide Phosphate |
| NF-κB | Nuclear Factor Kappa B |
| NO | Nitric Oxide |
| NOX | NADPH Oxidase |
| Nrf2 | Nuclear Factor Erythroid 2-Related Factor 2 |
| PBS | Phosphate-Buffered Saline |
| PCR | Polymerase Chain Reaction; |
| PSS | Physiological Salt Solution |
| RNA | Ribonucleic Acid |
| ROS | Reactive Oxygen Species |
| SEM | Standard Error Of The Mean |
| Sod2 | Superoxide Dismutase 2 |
| TGF-β | Transforming Growth Factor Beta |
| TNF | Tumor Necrosis Factor |
| Xdh | Xanthine Dehydrogenase |
References
- Ferrucci, L.; Fabbri, E. Inflammageing: Chronic inflammation in ageing, cardiovascular disease, and frailty. Nature reviews. Cardiology 2018, 15, 505-522.
- Franceschi, C.; Garagnani, P.; Parini, P.; Giuliani, C.; Santoro, A. Inflammaging: A new immune-metabolic viewpoint for age-related diseases. Nature reviews. Endocrinology 2018, 14, 576-590.
- Jurk, D.; Wilson, C.; Passos, J.F.; Oakley, F.; Correia-Melo, C.; Greaves, L.; Saretzki, G.; Fox, C.; Lawless, C.; Anderson, R., et al. Chronic inflammation induces telomere dysfunction and accelerates ageing in mice. Nature communications 2014, 2, 4172.
- Gurung, P.; Lukens, J.R.; Kanneganti, T.D. Mitochondria: Diversity in the regulation of the nlrp3 inflammasome. Trends in molecular medicine 2015, 21, 193-201.
- Hekimi, S.; Lapointe, J.; Wen, Y. Taking a “good” look at free radicals in the aging process. Trends in cell biology 2011, 21, 569-576.
- Mitra, R.; Nersesyan, A.; Pentland, K.; Melin, M.M.; Levy, R.M.; Ebong, E.E. Diosmin and its glycocalyx restorative and anti-inflammatory effects on injured blood vessels. FASEB journal : official publication of the Federation of American Societies for Experimental Biology 2022, 36, e22630.
- Huwait, E.; Mobashir, M. Potential and therapeutic roles of diosmin in human diseases. Biomedicines 2022, 10.
- El-Fawal, R.; El Fayoumi, H.M.; Mahmoud, M.F. Effects of diosmin and crocin on metabolic syndrome-associated cardio-vascular complications in rats. Naunyn-Schmiedeberg’s archives of pharmacology 2019, 392, 1523-1536.
- Neves, M.F.; Virdis, A.; Schiffrin, E.L. Resistance artery mechanics and composition in angiotensin ii-infused rats: Effects of aldosterone antagonism. Journal of hypertension 2003, 21, 189-198.
- Virdis, A.; Duranti, E.; Colucci, R.; Ippolito, C.; Tirotta, E.; Lorenzini, G.; Bernardini, N.; Blandizzi, C.; Taddei, S. Ghrelin restores nitric oxide availability in resistance circulation of essential hypertensive patients: Role of nad(p)h oxidase. European heart journal 2015, 36, 3023-3030.
- D’Antongiovanni, V.; Salvo, C.D.; Valdiserra, G.; Fornai, M.; Cappelli, F.; Mengozzi, A.; Virdis, A.; Masi, S.; Duranti, E.; Benvenuti, L., et al. Gut microbiota manipulation by probiotic lacticaseibacillus paracasei dg i1572 as new therapeutical strategy to counteract vascular inflammaging. Molecular nutrition & food research 2026, 70, e70399.
- Ajoolabady, A.; Pratico, D.; Tang, D.; Zhou, S.; Franceschi, C.; Ren, J. Immunosenescence and inflammaging: Mechanisms and role in diseases. Ageing research reviews 2024, 101, 102540.
- Virdis, A.; Duranti, E.; Rossi, C.; Dell’Agnello, U.; Santini, E.; Anselmino, M.; Chiarugi, M.; Taddei, S.; Solini, A. Tumour necrosis factor-alpha participates on the endothelin-1/nitric oxide imbalance in small arteries from obese patients: Role of perivascular adipose tissue. European heart journal 2015, 36, 784-794.
- Li, Y.J.; Jin, X.; Li, D.; Lu, J.; Zhang, X.N.; Yang, S.J.; Zhao, Y.X.; Wu, M. New insights into vascular aging: Emerging role of mitochondria function. Biomedicine & pharmacotherapy = Biomedecine & pharmacotherapie 2022, 156, 113954.
- Yan, Q.; Liu, S.; Sun, Y.; Chen, C.; Yang, S.; Lin, M.; Long, J.; Yao, J.; Lin, Y.; Yi, F., et al. Targeting oxidative stress as a preventive and therapeutic approach for cardiovascular disease. Journal of translational medicine 2023, 21, 519.
- Galla, R.; Mule, S.; Ferrari, S.; Molinari, C.; Uberti, F. Combined effects of diosmin, hesperidin, ruscus aculeatus, ananas comosus, and bromelain on endothelial function and gut barrier integrity in vitro. International journal of molecular sciences 2025, 26.
- Nelson, K.L.; Voruganti, V.S. Implication of xanthine oxidoreductase in oxidative stress-related chronic diseases. Frontiers in endocrinology 2025, 16, 1662037.
- Wojciak, M.; Feldo, M.; Borowski, G.; Kubrak, T.; Plachno, B.J.; Sowa, I. Antioxidant potential of diosmin and diosmetin against oxidative stress in endothelial cells. Molecules 2022, 27.
- Brandt, A.; Baumann, A.; Hernandez-Arriaga, A.; Jung, F.; Nier, A.; Staltner, R.; Rajcic, D.; Schmeer, C.; Witte, O.W.; Wessner, B., et al. Impairments of intestinal arginine and no metabolisms trigger aging-associated intestinal barrier dysfunction and ‘inflammaging’. Redox biology 2022, 58, 102528.
- Benvenuti, L.; D’Antongiovanni, V.; Pellegrini, C.; Fornai, M.; Bernardini, N.; Ippolito, C.; Segnani, C.; Di Salvo, C.; Colucci, R.; Martelli, A., et al. Dietary supplementation with the probiotic sf68 reinforces intestinal epithelial barrier in obese mice by improving butyrate bioavailability. Molecular nutrition & food research 2023, 67, e2200442.
- Xiong, Z.; Liao, Y.; Zhang, Z.; Wan, Z.; Liang, S.; Guo, J. Molecular insights into oxidative-stress-mediated cardiomyopathy and potential therapeutic strategies. Biomolecules 2025, 15.
- Xiong, H.H.; Lin, S.Y.; Chen, L.L.; Ouyang, K.H.; Wang, W.J. The interaction between flavonoids and intestinal microbes: A review. Foods 2023, 12.
- Wang, L.; Li, M.; Gu, Y.; Shi, J.; Yan, J.; Wang, X.; Li, B.; Wang, B.; Zhong, W.; Cao, H. Dietary flavonoids-microbiota crosstalk in intestinal inflammation and carcinogenesis. The Journal of nutritional biochemistry 2024, 125, 109494.
- Salem, M.B.; El-Lakkany, N.M.; Seif El-Din, S.H.; Hammam, O.A.; Samir, S. Diosmin alleviates ulcerative colitis in mice by increasing akkermansia muciniphila abundance, improving intestinal barrier function, and modulating the nf-kappab and nrf2 pathways. Heliyon 2024, 10, e27527.
- Liu, R.M.; Desai, L.P. Reciprocal regulation of tgf-beta and reactive oxygen species: A perverse cycle for fibrosis. Redox biology 2015, 6, 565-577.
- Zinkevich, N.S.; Gutterman, D.D. Ros-induced ros release in vascular biology: Redox-redox signaling. American journal of physiology. Heart and circulatory physiology 2011, 301, H647-653.
- Meephat, S.; Prasatthong, P.; Potue, P.; Bunbupha, S.; Pakdeechote, P.; Maneesai, P. Diosmetin ameliorates vascular dysfunction and remodeling by modulation of nrf2/ho-1 and p-jnk/p-nf-kappab expression in hypertensive rats. Antioxidants 2021, 10.
- Mas-Capdevila, A.; Teichenne, J.; Domenech-Coca, C.; Caimari, A.; Del Bas, J.M.; Escote, X.; Crescenti, A. Effect of hesperidin on cardiovascular disease risk factors: The role of intestinal microbiota on hesperidin bioavailability. Nutrients 2020, 12.
- Rahman, L.; Talha Khalil, A.; Ahsan Shahid, S.; Shinwari, Z.K.; Almarhoon, Z.M.; Alalmaie, A.; Sharifi-Rad, J.; Calina, D. Diosmin: A promising phytochemical for functional foods, nutraceuticals and cancer therapy. Food science & nutrition 2024, 12, 6070-6092.
- Russo, R.; Chandradhara, D.; De Tommasi, N. Comparative bioavailability of two diosmin formulations after oral administration to healthy volunteers. Molecules 2018, 23.
Figure 1.
Microvascular structure and function in the four study arms. (A) Media-to-lumen ratio, measure of vascular remodeling. Difference between aged + vehicle and aged + diosmin group were evaluated by unpaired t-test. Young + vehicle and young + diosmin groups were shown as reference control. Each column shows the mean ± SEM (n = 4-5, rats per group). Results are as follows: a—p < 0.05 for significant difference. (B-C) Vasorelaxation curves to acetylcholine in young (B) and aged (C) rats; (D) maximal vasorelaxation to acetylcholine in aged rats; (E) L-NAME vasoinhibition to maximal vasorelaxation to acetylcholine, measure of endothelial function, in aged rats. Maximal vasorelaxation with ACh alone and ACh+L-NAME were independently compared by unpaired t-test. (B-C) Each point shows the mean ± SEM (n = 4-5, rats per group). (D-E) Data are presented as violin plots showing the distribution of individual values (n = 4-5, rats per group). Results are as follows: a—p < 0.05; aa—p < 0.01 for significant difference. Abbreviations: L-NAME: N (ω)-nitro-L-arginine methyl ester.
Figure 1.
Microvascular structure and function in the four study arms. (A) Media-to-lumen ratio, measure of vascular remodeling. Difference between aged + vehicle and aged + diosmin group were evaluated by unpaired t-test. Young + vehicle and young + diosmin groups were shown as reference control. Each column shows the mean ± SEM (n = 4-5, rats per group). Results are as follows: a—p < 0.05 for significant difference. (B-C) Vasorelaxation curves to acetylcholine in young (B) and aged (C) rats; (D) maximal vasorelaxation to acetylcholine in aged rats; (E) L-NAME vasoinhibition to maximal vasorelaxation to acetylcholine, measure of endothelial function, in aged rats. Maximal vasorelaxation with ACh alone and ACh+L-NAME were independently compared by unpaired t-test. (B-C) Each point shows the mean ± SEM (n = 4-5, rats per group). (D-E) Data are presented as violin plots showing the distribution of individual values (n = 4-5, rats per group). Results are as follows: a—p < 0.05; aa—p < 0.01 for significant difference. Abbreviations: L-NAME: N (ω)-nitro-L-arginine methyl ester.

Figure 2.
Oxidative stress at the vascular wall level. Vascular wall levels of mitochondrial derives reactive oxygen species (A) and NADPH-oxidase derived reactive oxygen species (B). Difference between aged + vehicle and aged + diosmin group were evaluated by unpaired t-test. Young + vehicle and young + diosmin groups were shown as reference control. Each column shows the mean ± SEM (n = 3, rats per group). Results are as follows: a—p < 0.05 significant difference of diosmin-treated groups and respective controls. Abbreviations: MitoSOX: Mitochondrial Superoxide Indicator; DHE: Dihydroethidium.
Figure 2.
Oxidative stress at the vascular wall level. Vascular wall levels of mitochondrial derives reactive oxygen species (A) and NADPH-oxidase derived reactive oxygen species (B). Difference between aged + vehicle and aged + diosmin group were evaluated by unpaired t-test. Young + vehicle and young + diosmin groups were shown as reference control. Each column shows the mean ± SEM (n = 3, rats per group). Results are as follows: a—p < 0.05 significant difference of diosmin-treated groups and respective controls. Abbreviations: MitoSOX: Mitochondrial Superoxide Indicator; DHE: Dihydroethidium.

Figure 3.
Inflammatory parameters and oxidative stress index. (A) MDA and (B) TNF levels in plasma obtained young and aged rats treated with vehicle or diosmin for 3 months. Each column shows the mean ± SEM (n = 6-8, rats per group). One-way ANOVA and Tukey post hoc test results are as follows: ***—p < 0.001 for significant difference vs. young rats and aged rats; a—p < 0.05 for significant difference vs. diosmin-treated groups and respective controls. Abbreviations: TNF: tumor necrosis factor; MDA: malondialdehyde.
Figure 3.
Inflammatory parameters and oxidative stress index. (A) MDA and (B) TNF levels in plasma obtained young and aged rats treated with vehicle or diosmin for 3 months. Each column shows the mean ± SEM (n = 6-8, rats per group). One-way ANOVA and Tukey post hoc test results are as follows: ***—p < 0.001 for significant difference vs. young rats and aged rats; a—p < 0.05 for significant difference vs. diosmin-treated groups and respective controls. Abbreviations: TNF: tumor necrosis factor; MDA: malondialdehyde.

Figure 4.
Effects of aging and diosmin on circulating LBP levels. Plasma levels of lipopolysaccharide-binding protein (LBP) in young and aged animals treated with vehicle or diosmin. Data are presented as mean ± SEM (n = 5-8, rats per group); one-way ANOVA and Tukey post hoc test results are as follows: **—p < 0.01 for significant difference vs. young rats and aged rats; a—p < 0.05 for significant difference vs. diosmin-treated groups and respective controls. Abbreviations: LBP: Lipopolysaccharide Binding Protein.
Figure 4.
Effects of aging and diosmin on circulating LBP levels. Plasma levels of lipopolysaccharide-binding protein (LBP) in young and aged animals treated with vehicle or diosmin. Data are presented as mean ± SEM (n = 5-8, rats per group); one-way ANOVA and Tukey post hoc test results are as follows: **—p < 0.01 for significant difference vs. young rats and aged rats; a—p < 0.05 for significant difference vs. diosmin-treated groups and respective controls. Abbreviations: LBP: Lipopolysaccharide Binding Protein.

Figure 5.
Effects of aging and diosmin on oxidative stress–related gene expression. Relative mRNA levels of Enos (A), Gpx (B), Sod2 (C) and Xdh (D) in vascular tissues from young and aged animals with or without diosmin. Aging significantly increased Xdh expression, which was reduced by diosmin treatment, while Enos, Gpx and Sod2 remained unchanged. Difference between aged + vehicle and aged + diosmin group were evaluated by unpaired t-test. Young + vehicle and young + diosmin groups were shown as reference control. Data are shown as mean ± SEM (n = 4-7, per group). Results are as follows: a—p < 0.05 for significant difference vs. diosmin-treated groups and respective controls. Abbreviations: Enos: endothelial Nitric Oxide Synthase; Gpx: Glutathione Peroxidase; Sod2: Superoxide Dismutase 2; Xdh: Xanthine Dehydrogenase.
Figure 5.
Effects of aging and diosmin on oxidative stress–related gene expression. Relative mRNA levels of Enos (A), Gpx (B), Sod2 (C) and Xdh (D) in vascular tissues from young and aged animals with or without diosmin. Aging significantly increased Xdh expression, which was reduced by diosmin treatment, while Enos, Gpx and Sod2 remained unchanged. Difference between aged + vehicle and aged + diosmin group were evaluated by unpaired t-test. Young + vehicle and young + diosmin groups were shown as reference control. Data are shown as mean ± SEM (n = 4-7, per group). Results are as follows: a—p < 0.05 for significant difference vs. diosmin-treated groups and respective controls. Abbreviations: Enos: endothelial Nitric Oxide Synthase; Gpx: Glutathione Peroxidase; Sod2: Superoxide Dismutase 2; Xdh: Xanthine Dehydrogenase.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



