Submitted:
18 June 2026
Posted:
18 June 2026
You are already at the latest version
Abstract
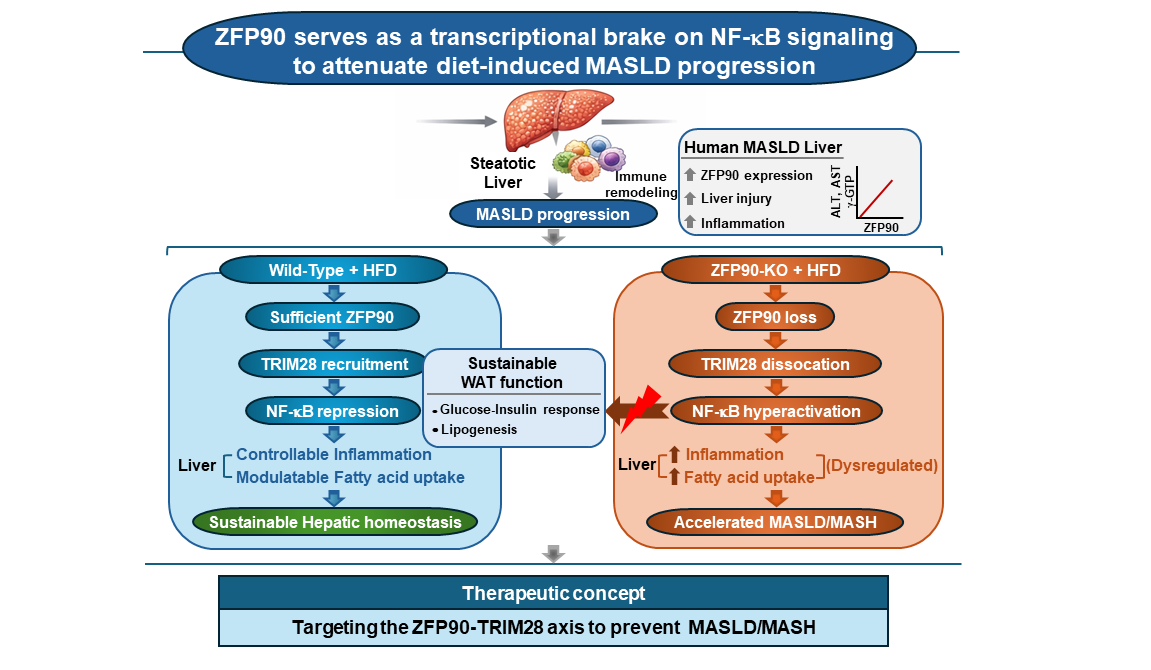
Background & Objectives: The increasing prevalence of metabolic dysfunction-associated steatotic liver disease (MASLD) is intrinsically linked to nutrient overload and obesity, and adipose dysfunction. While continuous dietary stress triggers adipose-tissue-derived lipotoxicity and disrupts hepatic metabolic homeostasis and provokes inflammation, the transcriptional scaffolds that mitigate this lipotoxicity remain incompletely understood. We investigated the role of zinc finger protein 90 (ZFP90) in defending against diet-induced metabolic stress and MASLD pathogenesis. Methods: Wild-type and ZFP90-knockout mice were subjected to a high-fat diet (HFD) to model nutrient-overload-induced MASLD. Hepatic phenotypes were characterized using metabolic profiling and RNA sequencing. Mechanistic dynamics were evaluated through protein interaction assays, and clinical relevance was validated using human MASLD liver biopsies. Results: ZFP90 deficiency significantly accelerated HFD-induced steatosis, systemic insulin resistance, and inflammatory infiltration. Crucially, ZFP90 depletion drove severe white adipose tissue (WAT) dysfunction, characterized by impaired lipogenic capacity, exacerbated lipolysis, and diminished local insulin signaling. This was accompanied by a pro-inflammatory secretory shift in WAT, evident from decreased Adipoq and increased Cd68/Ccl3 expression. In the liver, transcriptomic analysis revealed a profound induction of pathways related to fatty acid uptake and cytokine signaling. Mechanistically, ZFP90 forms a repressive complex with TRIM28, acting as a crucial molecular brake on NF-κB signaling. Loss of ZFP90 unleashes p65-mediated hyper-inflammation. Clinically, hepatic ZFP90 expression is significantly upregulated in patients with MASLD. Conclusions: ZFP90 is a novel regulator of immunometabolic homeostasis under dietary stress. By forming of complex with Trim28 to inhibit the nuclear translocation of NF-κB, ZFP90 suppresses pro-inflammatory responses and protects the liver from obesity-associated systemic lipotoxicity. These finding provide critical insights into the adipo-hepatic axis and highlight the ZFP90 as a promising therapeutic target to mitigate the progression to metabolic dysfunction-associated steatohepatitis (MASH).
Keywords:
MASLD
; MASH
; adipose tissue
; liver
; ZFP
1. Introduction
Metabolic dysfunction–associated steatotic liver disease (MASLD), formerly termed non-alcoholic fatty liver disease (NAFLD), has emerged as the most prevalent chronic liver disease worldwide, currently affecting over 30% of the global population [1]. Its prevalence is increasing in parallel with obesity and type 2 diabetes, reflecting the broader expansion of metabolic syndrome driven by chronic nutrient overload [2,3]. MASLD is characterized by hepatic steatosis in > 5% of hepatocytes, fundamentally linked to metabolic risk factors, namely obesity and insulin resistance, in the absence of excessive alcohol consumption or secondary causes of chronic liver diseases.
The pathological spectrum of MASLD is extensive, progressing from simple steatosis to metabolic dysfunction-associated steatohepatitis (MASH), which is defined by varying degrees of inflammation and fibrosis that can culminate in cirrhosis and hepatocellular carcinoma (HCC) [4,5,6]. Beyond hepatic outcomes, MASLD significantly elevates the risk of extrahepatic complications, including cardiovascular and chronic kidney disease. Despite this substantial clinical and socioeconomic burden, pharmacological interventions remain limited, emphasizing the urgent need to elucidate the molecular drivers governing its pathogenesis [7]. Current paradigms suggest that MASLD progression is orchestrated by a “multiple-hit” process. Central to this process is the impact of continuous dietary stress, which precipitates adipose tissue dysfunction, impairs intestinal barrier integrity, and ultimately leads to the chronic activation of hepatic immune cell populations [8].
In particular, the adipo-hepatic axis plays a pivotal role in determining the liver’s response to nutritional stress. As the largest immunological organ in the body, the liver hosts a diverse repertoire of innate and adaptive immune cells, including Kupffer cells, macrophages, and lymphocytes [9]. Substantial alterations in these immune populations have been observed in both murine and human MASLD, contributing to a dysregulated inflammatory milieu that exacerbates hepatocellular injury [10,11,12]. Recent evidence highlights that obesity-induced adipose tissue dysfunction—characterized by lipolysis and pro-inflammatory adipokine secretion—creates a state of systemic lipotoxicity. This systemic stress, combined with intrahepatic immune-parenchymal crosstalk, coordinates the delicate balance between lipid metabolism and inflammatory signaling [13,14,15,16]. While perturbations in these cellular interactions are recognized as the primary drivers of MASLD progression [16], the specific molecular mediators that integrate systemic metabolic cues with local immune regulation remain largely elusive.
Zinc finger proteins (ZFPs), the most expansive family of transcriptional regulators, are indispensable for diverse biological processes, ranging from cellular differentiation to immune modulation [17,18]. Although select ZFP family members have been implicated in metabolic homeostasis [19,20,21], the functional role of zinc finger protein 90 (ZFP90) remains poorly understood. While ZFP90 has been previously associated with hematopoietic cell proliferation and cardiac development [22,23], its potential involvement in systemic adipo-hepatic and immune–metabolic regulation has yet to be explored.
In the present study, we identified ZFP90 as a critical regulator of systemic and hepatic immune-metabolic homeostasis under dietary stress. Utilizing ZFP90-knockout mice challenged with a high-fat diet, integrated transcriptomic profiling, and mechanistic investigations of the ZFP90-TRIM28 interaction, we demonstrate that ZFP90 deficiency drives profound white adipose tissue (WAT) dysfunction, exacerbates hepatic lipotoxicity, alters intrahepatic T-cell dynamics, and promotes NF-κB–dependent hyper-inflammation. Our findings characterize ZFP90 as a novel, previously unrecognized modulator of MASLD pathogenesis and highlight its potential as a therapeutic target for mitigating MASH progression driven by nutrient overload.
2. Materials and Methods
2.1. Animal Models and Diets
All animal procedures and experimental protocols were strictly performed in accordance with the guidelines approved by the Ethics Review Committee for Animal Experimentation at Nagasaki University. Zfp90-knockout (Zfp90−/−) mice on a C57BL/6 background were generated by deleting exon 3 of the mouse Zfp90 transcript. Animals were maintained in a specific pathogen-free facility under a controlled environment (temperature: 21–24 °C; 12 h light/dark cycle) with ad libitum access to water and a standard CRF-1 diet (Oriental Yeast Co. Ltd., Tokyo, Japan). At 12 weeks of age, male mice were randomly assigned to either a standard chow diet or a high-fat diet (HFD; 60% kcal from fat) for 20 weeks to model nutrient overload. Body weight was monitored regularly, and body composition (total, visceral, and subcutaneous adipose tissue) was evaluated at 6 months of age using 3D micro-computed tomography (Rigaku Co., Tokyo, Japan).
2.2. Metabolic and Biochemical Phenotyping
Whole-body energy consumption (VO2), carbon dioxide production (VCO2), and respiratory quotient (RQ) were monitored using an indirect calorimetry system (MK-5000RQ, Muromachi Kikai Co., Tokyo, Japan). Glucose homeostasis was evaluated via glucose tolerance tests (GTT); following a 4 h fast, mice received an intraperitoneal injection of glucose (1 g/kg body weight). Blood glucose levels were measured at 0, 15, 30, 60, and 120 min post-injection using an Accu-Chek Aviva Nano Meter (Roche Diagnostics, Tokyo, Japan). Serum concentrations of insulin, alanine aminotransferase (ALT), and aspartate aminotransferase (AST) were quantified using validated ELISA systems (Morinaga Institute of Biological Science, Yokohama, Japan; FUJIFILM Wako Pure Chemical Corp., Osaka, Japan) according to the manufacturers’ protocols.
2.3. Mass Spectrometry Analysis
Protein samples were fractionated on a 4–20% gradient SDS-PAGE (Cosmo Bio, Tokyo, Japan) and stained with GelCode Blue Reagent (Thermo Fisher Scientific, Waltham, MA, USA). Protein bands ranging from 13 to 120 kDa were excised and subjected to in-gel digestion according to the manufacturer’s instructions (Bruker Daltonics, Bremen, Germany). Briefly, the gel pieces were destained with 25 mM ammonium bicarbonate in 50% acetonitrile (ACN), reduced with 10 mM dithiothreitol at 56 °C for 45 min, and alkylated with 55 mM iodoacetamide for 30 min at room temperature. After dehydration with ACN, the gel was incubated with 20 ng/µL Trypsin Gold (Promega, Madison, WI, USA) at 37 °C overnight. Digested peptides were extracted using 50% ACN/5% trifluoroacetic acid (TFA), followed by sonication and vortexing. The extract was concentrated by evaporation and reconstituted in 2% ACN/0.1% TFA. Samples were analyzed using a custom liquid chromatography-electrospray ionization–tandem mass spectrometry (LC-ESI-MS/MS) system (LTQ-XL; Thermo Fisher Scientific). Raw MS/MS data were processed and searched against the UniProt Mus musculus database using Proteome Discoverer v.3.3 (Thermo Fisher Scientific) to identify high-confidence protein-protein interactions.
2.4. RNA Sequencing and Transcriptomic Analysis
Library preparation was performed using a TruSeq Stranded mRNA Sample Prep Kit (Illumina, San Diego, CA, USA) according to the manufacturer’s instructions. Sequencing was performed on an Illumina NovaSeq 6000 platform in 151-base single-end mode. The sequenced reads were trimmed using Trimmomatic version 0.39 and mapped to the mouse reference genome sequences (GRCm38.p4) using HISAT2 version 2.1.0. Fragments per kilobase of exon per million mapped fragments (FPKMs) were calculated using Cufflinks version 2.0.6. Differentially expressed genes (DEGs) were identified based on a fold-change threshold of > 2.0 or < 0.5, with a significance level of p < 0.05. Raw data were deposited in the NCBI Gene Expression Omnibus (GEO) under accession number GSE324098.
2.5. Quantitative Real-Time PCR
Total RNA was extracted from the liver and adipose tissues using the RNeasy Tissue Kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. cDNA was synthesized from the isolated RNA using the ReverTra Ace qPCR RT Kit (Toyobo, Osaka, Japan). The relative amount of mRNA expression was analyzed by quantitative PCR (qPCR) using the THUNDERBIRD™ SYBR and probe qPCR Mix (Toyobo, Japan) on a real-time PCR detection system. The reaction conditions followed the standardized thermal cycling protocol provided by the manufacturer. The expression levels of mouse Zfp90 and human ZFP90 were analyzed using TaqMan probes (Mm00496071_m1 and Hs01001573_g1, respectively). Relative mRNA expression levels were calculated and normalized to the endogenous controls, 18S ribosomal RNA (18S), ribosomal protein lateral stalk subunit P0 (Rplp0), and glyceraldehyde-3-phosphate dehydrogenase (Gapdh). All primer sequences used in this study are listed in Tables S1 and S2.
2.6. Western Blotting
Proteins were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto nitrocellulose membranes. Membranes were blocked for 60 min in TBS-T buffer containing either 3% BSA or 5% non-fat dry milk, followed by overnight incubation at 4 °C with primary antibodies: anti-TRIM28, anti-p-AKT (Ser473) (Cell Signaling Technology, Danvers, MA, USA), anti-Flag (Sigma-Aldrich, St. Louis, MO, USA), or anti-β-actin (Abcam, Cambridge, UK). Following incubation with species-specific HRP-linked secondary antibodies (Amersham Pharmacia Biotech, Amersham, UK), protein bands were visualized using enhanced chemiluminescence (ECL) reagents. Signals were quantified via the FluorChem system (Alpha Innotech Corp., San Leandro, CA, USA) using AlphaEase FC software.
2.7. Histological Analyzes
Adipose and liver tissues were fixed in 4% paraformaldehyde (PFA), embedded in paraffin, and stained with hematoxylin and eosin (H&E). Images were acquired using a BZ-X700 microscope (Keyence, Osaka, Japan). Quantitative analysis of adipocyte size and brown adipocyte count was performed using the BZ-X700 application software and ImageJ (n = 4 per group).
2.8. Cell Culture
COS-7 cells were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM). MT-2 and SKW-3 cells were cultured in Roswell Park Memorial Institute medium (RPMI), supplemented with 10% fetal bovine serum, 100 U/mL penicillin, and 100 mg/mL streptomycin. Cells were incubated at 37 °C in humidified 5% CO2 and subcultured upon reaching confluence.
2.9. Liver Non-Parenchymal Cell (NPC) Isolation
To isolate non-parenchymal cells (NPCs), the liver was perfused with phosphate-buffered saline (PBS). The liver was then excised, minced, and filtered through a 70 µm nylon mesh to obtain a single-cell suspension. To separate NPCs from hepatocytes, the suspension was subjected to low-speed centrifugation at 50× g for 5 min at 4 °C. The supernatant containing the NPCs was collected and further centrifuged at 500× g for 10 min. The resulting NPC pellet was resuspended and further purified using a density gradient and red blood cell lysis buffer.
2.10. Clinical Study
Formalin-fixed paraffin-embedded (FFPE) human liver sections were obtained from patients with MASLD and healthy liver transplant donors at Nagasaki University Hospital (2010–2024). Total RNA was isolated using the RNeasy FFPE Kit (Qiagen, Germany) and cDNA was synthesized as described above. Clinical and biochemical characteristics of the study population were obtained from the Nagasaki University Hospital information system. All procedures were performed in accordance with protocols approved by the Clinical Research Ethics Committee of Nagasaki University Hospital (Permit No. 25080703).
2.11. Statistical Analysis
All data are presented as the mean ± SEM. Statistical significance was determined using an unpaired two-tailed Student’s t-test and two-way analysis of variance (ANOVA) with Bonferroni’s multiple comparisons test. Analysis was performed using GraphPad Prism 5, with p < 0.05 considered statistically significant.
3. Results
3.1. ZFP90 Deficiency Promotes Visceral Adiposity and Shifts Metabolic Substrate Utilization
We previously demonstrated that neuropeptide Y (NPY)-mediated lipid metabolic balance is essential for calorie restriction-induced lifespan extension, and that zinc finger protein 90 (ZFP90) serves as a key mediator in this NPY-related lipid metabolism [24]. We found that Zfp90 mRNA expression in white adipose tissue (WAT) was downregulated under a negative energy balance (30% CR; Figure 1A) but significantly upregulated under a positive energy balance (chronic nutrient overload via high-fat diet, HFD) or aging (Figure 1B,C). To investigate its functional role, 12-week-old ZFP90 wild-type (WT) and knockout (KO) mice were fed either a standard chow diet (CRF1) or an HFD for 20 weeks. While body weight and food consumption showed no significant differences between genotypes (Figure 2A and Figure S1), body fat mass was significantly higher in KO mice fed the CRF1 diet (Figure 1D). Histological analysis revealed larger adipocytes in the epididymal WAT (eWAT) of KO mice compared to WT mice fed the CRF1 diet (Figure 1E). These results indicate that ZFP90 deficiency promotes visceral adiposity.
3.2. Loss of ZFP90 Exacerbates HFD-Induced MASLD and Hepatic Transcriptomic Remodeling
On the HFD, both WT and KO mice gained significantly more weight than those on the chow diet, with no significant inter-genotype differences in weight gain (Figure 2A) or total WAT accumulation (Figure 2B and Figure S2A,B). However, KO mice exhibited exacerbated liver injury under HFD, as evidenced by significantly elevated serum alanine aminotransferase (ALT) and aspartate aminotransferase (AST) levels (Figure 2D). Regression analysis revealed a strong positive correlation between visceral fat weight and the liver weight-to-body weight (BW) ratio, underscoring that excessive adiposity and systemic lipid overflow directly drive hepatic steatosis (Figure 2C,E). Notably, Zfp90 mRNA expression in eWAT was positively correlated with the liver-to-BW ratio, suggesting its role as a reactive regulator against systemic lipid stress (Figure 2E).
Transcriptomic profiling (RNA-seq) identified 1,038 upregulated and 263 downregulated genes (fold-change > 2) in ZFP90-KO livers compared to WT livers (Figure S3A). Pathway enrichment analysis revealed that ZFP90 deficiency upregulated programs involved in immunity, inflammatory responses, and lipid metabolism—specifically cytokine-cytokine receptor interactions, NF-κB, and chemokine signaling (Figure S3B). To explore the mechanisms underlying ZFP90 deficiency-induced hepatic steatosis, we assessed specific hepatic gene expression based on the RNA-seq data. ZFP90 deficiency increased the mRNA levels of genes related to fatty acid uptake (Cd36, Lpl, Mogat1), inflammation (Tnfa, Cd68, Ccr2), and fibrosis (Col1a2) (Figure 2F,G). These findings indicate that the loss of ZFP90 accelerates diet-induced MASLD progression through the integrated dysregulation of inflammatory and metabolic pathways.
3.3. ZFP90-Deficiency Drives Adipose Tissue Dysfunction and Systemic Insulin Resistance
Adipose tissue dysfunction and systemic inflammation are known primary drivers of hepatic steatosis [25]. In HFD-fed mice, ZFP90 deficiency significantly decreased the expression of key lipogenic genes (Cebpa, Pparg2, Acc1, and Fasn) while increasing the expression of the lipolytic gene Lipa in WAT (Figure 3A). This imbalance suggests an impaired lipid storage capacity in WAT, which potentially exacerbates ectopic lipid deposition in the liver. Furthermore, ZFP90-KO mice exhibited a pronounced pro-inflammatory shift in WAT, indicated by the upregulation of macrophage and inflammatory markers (Cd68 and Ccl3) and the concurrent downregulation of the anti-inflammatory adipokine Adipoq (Figure 3B). While both genotypes developed HFD-induced hyperinsulinemia, ZFP90-KO mice exhibited markedly higher fasting blood glucose levels and diminished Akt phosphorylation (p-Akt(Ser473)) in WAT (Figure 3C–E). Collectively, these results demonstrate that ZFP90 deficiency exacerbates MASLD pathogenesis, at least in part, by driving adipose tissue dysfunction and impairing systemic insulin sensitivity.
3.4. ZFP90 Interacts with TRIM28 to Suppress NF-κB Dependent Hepatic Inflammation
To identify ZFP90-binding partners and elucidate its molecular mechanisms, we performed immunoprecipitation and LC-MS/MS analysis in COS-7 cells expressing FLAG-tagged ZFP90 (Figure 4A). Ten protein bands containing putative ZFP90-binding candidates were used to identify nine potential interacting proteins (Figure 4A,B). Among them, tripartite motif-containing 28 (TRIM28) is of particular interest, as previous reports have suggested that ZFP90 forms a complex with TRIM28, a known regulator of NF-κB signaling [26,27]. To validate this interaction, western blot analysis was performed, confirming the physical association between ZFP90 and TRIM28 (Figure 4C). To test the functional impact of this interaction within the hepatic microenvironment, we stimulated liver non-parenchymal cells from LPS-injected mice. LPS-induced Tnfa and Ccl2 expression was significantly higher in ZFP90-KO cells, demonstrating that the ZFP90-TRIM28 complex serves as a critical suppressor of excessive NF-κB activation (Figure 4D).
3.5. ZFP90 Deficiency Alters T-Cell Dynamics
Data from the Human Protein Atlas indicate high expression of both ZFP90 and TRIM28 in liver immune cells, particularly T cells [28]. To further elucidate the hepatic immunomodulatory function of ZFP90, we measured the mRNA expression levels of T-cell markers (Cd4 and Cd8a), along with Tlr7, Trim28, and Tnfa. ZFP90 deficiency significantly altered the intrahepatic immune landscape, resulting in reduced Cd4 and increased Tlr7 and Tnfa expression (Figure 5A). Together, these findings suggest that ZFP90 is essential for maintaining hepatic T-cell homeostasis, and its absence shifts the immune microenvironment toward a pro-inflammatory state during metabolic stress.
To investigate the specific role of ZFP90 in T cells, we examined the mRNA expression profiles of T-cell-related genes in MT2, a human regulatory T (Treg)-like cell line [29], and SKW3, a T-cell leukemia cell line. The two cell lines exhibited distinct phenotypic characteristics; MT2 cells showed high expression of IL2RA and IL2RB, whereas SKW3 cells displayed elevated levels of TNFA and IL2 (Figure S4). Notably, ZFP90 expression was more than two-fold higher in MT2 cells compared to SKW3 cells (Figure 5B). Given that TRIM28 plays a crucial role in Treg cell function [30], we knocked down ZFP90 in MT2 cells to evaluate its impact. Interestingly, the knockdown of ZFP90 markedly increased the mRNA expression of TNFα (Figure 5C). Furthermore, nuclear p65 protein levels were significantly elevated in the livers of KO mice compared to those of WT mice (Figure 5D). These results suggest that ZFP90 may function as a transcriptional repressor of pro-inflammatory cytokines in Treg cells, potentially cooperating with TRIM28 to maintain their immunosuppressive identity.
3.6. Hepatic ZFP90 Expression Correlates with MASLD Severity in Humans
Finally, we assessed the clinical relevance of ZFP90 in humans. Table 1 summarizes the physical, anthropometric, and biochemical evaluations, including biomarkers of MASLD, for both patients and controls across all study phases. We first examined the hepatic mRNA expression of ZFP90 and found that it was significantly elevated in patients with MASLD compared to healthy controls (Figure 6A). Furthermore, we explored the association between ZFP90 expression and individual biomarkers in the MASLD cohort. Consistent with our murine data, ZFP90 expression showed a strong positive correlation with serum AST and ALT concentrations (Figure 6B,C). Additionally, ZFP90 levels were positively correlated with γ-GTP levels (Figure 6B). This clinical upregulation of ZFP90 expression likely reflects a compensatory, albeit insufficient, physiological response to chronic adipo-hepatic injury and severe metabolic stress.
4. Discussion
The present study identified ZFP90 as a novel transcriptional regulator that governs the intricate balance between hepatic lipid metabolism and inflammatory signaling under conditions of nutrient overload. Through ZFP90-deficient murine models and clinical validation, we demonstrated that the loss of this zinc finger protein significantly accelerates the transition from simple steatosis to an aggressive MASH-like phenotype. Our findings provide compelling evidence that ZFP90, via its physical and functional interaction with TRIM28, serves as a crucial molecular brake in the NF-κB signaling pathway, thereby protecting the liver from obesity-induced metabolic lipotoxicity and chronic inflammation.
Crucially, our study highlights the profound impact of ZFP90 deficiency on systemic metabolic homeostasis, primarily driven by adipose tissue dysfunction. Under chronic dietary stress, the observed imbalance between lipogenic and lipolytic processes, coupled with impaired insulin signaling (reduced p-Akt), suggests that ZFP90 is essential for maintaining healthy adipose expansion. Dysfunctional adipose tissue directly contributes to hepatic steatosis via the adipo-hepatic axis, where increased lipolysis and pro-inflammatory adipokine secretion drive systemic lipotoxicity and hepatic insulin resistance. These systemic alterations act in concert with intrahepatic mechanisms to accelerate MASLD progression.
Consequently, the influx of adipose-derived lipids and inflammatory signals exacerbates hepatic lipotoxicity. We found that ZFP90 deficiency promotes hepatic steatosis independently of caloric intake or total body weight (Figure 2 and Figure S1), suggesting a direct regulatory role in hepatic lipid handling in response to systemic overflow. Transcriptomic profiling revealed that ZFP90 deficiency leads to the robust induction of genes involved in fatty acid uptake (Cd36, Mogat1) and pro-inflammatory cascades (Ccl2, Tnfa). This dual dysregulation indicates that ZFP90 is not merely a metabolic regulator but a master scaffold that integrates extracellular nutrient sensing with local immune responses. Specifically, the induction of Ccr2 and Cd68 in Zfp90⁻/⁻ livers underscores the increased recruitment of monocyte-derived macrophages, which are fundamental drivers of the persistent inflammation characterizing MASLD progression.
The mechanistic nexus between ZFP90 and TRIM28 identified here adds a new layer of complexity to the known functions of Krüppel-associated box domain (KRAB)-ZFPs in the liver. While KRAB-ZFPs typically recruit TRIM28 for epigenetic silencing, their roles in hepatic metabolic diseases remain understudied. Our data suggest that the ZFP90-TRIM28 complex acts as a repressive unit that restricts the nuclear translocation and transcriptional activity of p65. This aligns with prior reports indicating that TRIM28 can sequester the NF-κB (p65) subunit to dampen inflammatory output [27]. Dysregulated NF-κB signaling is a well-established driver of MASLD, mediating hepatocyte injury and macrophage activation [31,32]. The loss of ZFP90 effectively “relieves the brake” in this system, priming the liver for a hyper-inflammatory response to nutrient-induced metabolic stress. This inflammatory priming is further corroborated by a significant shift in intrahepatic T-cell dynamics, specifically the reduced Cd4 and increased Tnfa [14,33,34,35,36,37]. We observed that ZFP90 deficiency drives this shift, likely through NF-κB-dependent chemokine signaling, which amplifies local inflammation and hepatocellular injury.
Furthermore, our clinical data provide a critical translational perspective. The significant upregulation of hepatic ZFP90 in patients with MASLD, which correlates with serum AST/ALT levels, likely reflects a compensatory but ultimately insufficient defense mechanism. We propose that the liver attempts to upregulate ZFP90 to mitigate inflammatory damage under chronic systemic metabolic stress; however, this regulatory capacity is eventually overwhelmed as lipotoxicity persists.
Despite these insights, this study has several limitations that warrant consideration. The precise downstream effectors of the ZFP90-TRIM28 complex in human T cells require further characterization, and the tissue-specific contributions of ZFP90 (such as in adipocytes versus hepatocytes) should be rigorously investigated using conditional knockout models. Future studies should focus on the pharmacological modulation of the ZFP90 pathway to evaluate its clinical utility in treating obesity-associated metabolic complications.
5. Conclusions
This study establishes ZFP90 as a critical gatekeeper of systemic and hepatic immunometabolic integrity under dietary stress. By elucidating the adipo-hepatic interplay and the role of ZFP90 in inflammatory signaling, we uncovered a potential therapeutic target that could be exploited to arrest MASLD progression. Strategies aimed at stabilizing ZFP90 activity or enhancing its interaction with co-repressors, such as TRIM28, may offer a novel avenue for mitigating systemic lipotoxicity and preventing the transition to advanced MASH.
Supplementary Materials
The following supporting information can be downloaded at website of this paper posted on Preprints.org, Figure S1: ZFP90 deficiency does not affect food intake; Figure S2: ZFP90 deficiency does not affect adipocyte size in white adipose tissue; Figure S3: ZFP90 deficiency alters hepatic gene expression profiles related to immunity and lipid metabolism; Figure S4: mRNA expression levels in MT-2 and SKW-3 cells; Table S1: Gene name, symbols, and primer sequences (Mus musculus) for quantitative real-time PCR; Table S2: Gene name, symbols, and primer sequences (Homo sapiens) for quantitative real-time PCR.
Author Contributions
S.P. designed the study and wrote the paper. S.P., T.K., K.M., and D.O. performed the experiments. I.S. helped with the data analysis and interpretation. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Grants-in-Aid for Scientific Research from the Japan Society for the Promotion of Science (No. 23K28021 and 24K14652).
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Clinical Research Ethics Committee of Nagasaki University Hospital (Permit No. 25080703) for studies involving humans. The animal study protocol was approved by the Animal Care and Use Committee of Nagasaki University (Protocol Nos. 1909171569 and 2404301945).
Informed Consent Statement
Patient consent was waived due to the retrospective nature of the study utilizing archived formalin-fixed paraffin-embedded (FFPE) tissue samples.
Data Availability Statement
The transcriptomic (RNA-seq) data presented in this study are openly available in the NCBI Gene Expression Omnibus (GEO) under accession number GSE324098.
Acknowledgments
We are grateful to the staff at the Nagasaki University Hospital Clinical Research Center, Research Center for Biomedical Models and Animal Welfare, and Biomedical Research Support Center for their technical assistance. We also thank Yutaka Araki, Yuko Moriyama, Hiromi Noda, and Ryota Noda for excellent technical assistance.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have influenced the work reported in this paper. Notably, Isao Shimokawa is now employed by SAGL Limited Liability Company (SAGL, LLC). SAGL LLC, in affiliation, had no role in the study’s design, data collection, analysis, interpretation, manuscript writing, or decision to publish the results.
References
- Younossi, Z.M.; Golabi, P.; Paik, J.M.; Henry, A.; Van Dongen, C.; Henry, L. The global epidemiology of nonalcoholic fatty liver disease (NAFLD) and nonalcoholic steatohepatitis (NASH): a systematic review. Hepatology 2023, 77, 1335-1347. [CrossRef]
- Younossi, Z.M.; Golabi, P.; de Avila, L.; Paik, J.M.; Srishord, M.; Fukui, N.; et al. The global epidemiology of NAFLD and NASH in patients with type 2 diabetes: a systematic review and meta-analysis. J. Hepatol. 2019, 71, 793-801. [CrossRef]
- Polyzos, S.A.; Kountouras, J.; Mantzoros, C.S. Obesity and nonalcoholic fatty liver disease: from pathophysiology to therapeutics. Metabolism 2019, 92, 82-97. [CrossRef]
- Kaya, E.; Yilmaz, Y. Metabolic-associated fatty liver disease (MAFLD): a multi-systemic disease beyond the liver. J. Clin. Transl. Hepatol. 2022, 10, 329-338.
- Zhou, X.D.; Targher, G.; Byrne, C.D.; Somers, V.; Kim, S.U.; Chahal, C.A.A.; et al. An international multidisciplinary consensus statement on MAFLD and the risk of CVD. Hepatol. Int. 2023, 17, 773-791. [CrossRef]
- Powell, E.E.; Wong, V.W.; Rinella, M. Non-alcoholic fatty liver disease. Lancet 2021, 397, 2212-2224. [CrossRef]
- Miao, L.; Targher, G.; Byrne, C.D.; Cao, Y.Y.; Zheng, M.H. Current status and future trends of the global burden of MASLD. Trends Endocrinol. Metab. 2024, 35, 697-707. [CrossRef]
- Yan, M.; Man, S.; Ma, L.; Guo, L.; Huang, L.; Gao, W. Immunological mechanisms in steatotic liver diseases: An overview and clinical perspectives. Clin. Mol. Hepatol. 2024, 30, 620-648. [CrossRef]
- Racanelli, V.; Rehermann, B. The liver as an immunological organ. Hepatology 2006, 43, S54-S62. [CrossRef]
- Zheng, X.; Zhang, H.; Yin, L.; Wang, C.R.; Liu, Y.; Zheng, P. Modulation of NKT cell development by B7-CD28 interaction: an expanding horizon for costimulation. PLoS One 2008, 3, e2703. [CrossRef]
- Xu, G.X.; Wei, S.; Yu, C.; Zhao, S.Q.; Yang, W.J.; Feng, Y.H.; et al. Activation of Kupffer cells in NAFLD and NASH: mechanisms and therapeutic interventions. Front. Cell Dev. Biol. 2023, 11, 1199519. [CrossRef]
- Barreby, E.; Chen, P.; Aouadi, M. Macrophage functional diversity in NAFLD - more than inflammation. Nat. Rev. Endocrinol. 2022, 18, 461-472. [CrossRef]
- Steixner-Kumar, A.A.; Santacruz, D.; Geiger, T.; Rust, W.; Böttner, D.; Krenkel, O.; et al. Single-cell landscape of peripheral immune cells in MASLD/MASH. Hepatol. Commun. 2025, 9, e0643. [CrossRef]
- Sawada, K.; Chung, H.; Softic, S.; Moreno-Fernandez, M.E.; Divanovic, S. The bidirectional immune crosstalk in metabolic dysfunction-associated steatotic liver disease. Cell Metab. 2023, 35, 1852-1871. [CrossRef]
- L’homme, L.; Sermikli, B.P.; Haas, J.T.; Fleury, S.; Quemener, S.; Guinot, V.; et al. Adipose tissue macrophage infiltration and hepatocyte stress increase GDF-15 throughout development of obesity to MASH. Nat. Commun. 2024, 15, 7173. [CrossRef]
- Kostallari, E.; Schwabe, R.F.; Guillot, A. Inflammation and immunity in liver homeostasis and disease: a nexus of hepatocytes, nonparenchymal cells and immune cells. Cell Mol. Immunol. 2025, 22, 1205-1225. [CrossRef]
- Ecco, G.; Cassano, M.; Kauzlaric, A.; Duc, J.; Coluccio, A.; Offner, S.; et al. Transposable elements and their KRAB-ZFP controllers regulate gene expression in adult tissues. Dev. Cell 2016, 36, 611-623. [CrossRef]
- Urrutia, R. KRAB-containing zinc-finger repressor proteins. Genome Biol. 2003, 4, 231.
- Wang, X.; Jiao, A.; Sun, L.; Li, W.; Yang, B.; Su, Y.; et al. Zinc finger protein Zfp335 controls early T-cell development and survival through beta-selection-dependent and -independent mechanisms. eLife 2022, 11, e75508. [CrossRef]
- Charrier, A.; Wang, L.; Stephenson, E.J.; Ghanta, S.V.; Ko, C.W.; Croniger, C.M.; et al. Zinc finger protein 407 overexpression upregulates PPAR target gene expression and improves glucose homeostasis in mice. Am. J. Physiol. Endocrinol. Metab. 2016, 311, E869-E880. [CrossRef]
- Shao, M.; Ishibashi, J.; Kusminski, C.M.; Wang, Q.A.; Hepler, C.; Vishvanath, L.; et al. Zfp423 maintains white adipocyte identity through suppression of the beige cell thermogenic gene program. Cell Metab. 2016, 23, 1167-1184. [CrossRef]
- Hata, L.; Murakami, M.; Kuwahara, K.; Nakagawa, Y.; Kinoshita, H.; Usami, S.; et al. Zinc-finger protein 90 negatively regulates neuron-restrictive silencer factor-mediated transcriptional repression of fetal cardiac genes. J. Mol. Cell. Cardiol. 2011, 50, 972-981. [CrossRef]
- Liu, T.; Kong, W.X.; Tang, X.Y.; Xu, M.; Wang, Q.H.; Zhang, B.; et al. The transcription factor Zfp90 regulates the self-renewal and differentiation of hematopoietic stem cells. Cell Death Dis. 2018, 9, 677. [CrossRef]
- Park, S.; Komatsu, T.; Kim, S.E.; Tanaka, K.; Hayashi, H.; Mori, R.; et al. Neuropeptide Y resists excess loss of fat by lipolysis in calorie-restricted mice: A trait potential for the life-extending effect of calorie restriction. Aging Cell 2017, 16, 339-348. [CrossRef]
- Azzu, V.; Vacca, M.; Virtue, S.; Allison, M.; Vidal-Puig, A. Adipose tissue-liver cross talk in the control of whole-body metabolism: Implications in nonalcoholic fatty liver disease. Gastroenterology 2020, 158, 1899-1912. [CrossRef]
- Huang, C.; Martin, S.; Pfleger, C.; Du, J.; Buckner, J.H.; Bluestone, J.A.; et al. Cutting edge: a novel, human-specific interacting protein couples FOXP3 to a chromatin-remodeling complex that contains KAP1/TRIM28. J. Immunol. 2013, 190, 4470-4473. [CrossRef]
- Kamitani, S.; Togi, S.; Ikeda, O.; Nakasuji, M.; Sakauchi, A.; Sekine, Y.; et al. Kruppel-associated box-associated protein 1 negatively regulates TNF-alpha-induced NF-kappaB transcriptional activity by influencing the interactions among STAT3, p300, and NF-kappaB/p65. J. Immunol. 2011, 187, 2476-2483. [CrossRef]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [CrossRef]
- Hamano, R.; Wu, X.; Wang, Y.; Oppenheim, J.J.; Chen, X. Characterization of MT-2 cells as a human regulatory T cell-like cell line. Cell Mol. Immunol. 2015, 12, 780-782.
- Tanaka, S.; Pfleger, C.; Lai, J.F.; Roan, F.; Sun, S.C.; Ziegler, S.F. KAP1 regulates regulatory T cell function and proliferation in both Foxp3-dependent and -independent manners. Cell Rep. 2018, 23, 796-807. [CrossRef]
- Kohler, U.A.; Bohm, F.; Rolfs, F.; Egger, M.; Hornemann, T.; Pasparakis, M.; et al. NF-kappaB/RelA and Nrf2 cooperate to maintain hepatocyte integrity and to prevent development of hepatocellular adenoma. J. Hepatol. 2016, 64, 94-102.
- Oakley, F.; Teoh, V.; Ching, A.S.G.; Bataller, R.; Colmenero, J.; Jonsson, J.R.; et al. Angiotensin II activates I kappaB kinase phosphorylation of RelA at Ser 536 to promote myofibroblast survival and liver fibrosis. Gastroenterology 2009, 136, 2334-2344. [CrossRef]
- Van Herck, M.A.; Weyler, J.; Kwanten, W.J.; Dirinck, E.L.; De Winter, B.Y.; Francque, S.M.; et al. The differential roles of T cells in nonalcoholic fatty liver disease and obesity. Front. Immunol. 2019, 10, 82. [CrossRef]
- Moreno-Fernandez, M.E.; Giles, D.A.; Oates, J.R.; Chan, C.C.; Damen, M.S.M.A.; Doll, J.R.; et al. PKM2-dependent metabolic skewing of hepatic Th17 cells regulates pathogenesis of nonalcoholic fatty liver disease. Cell Metab. 2021, 33, 1187-1204. [CrossRef]
- Campbell, C.; Rudensky, A. Roles of regulatory T cells in tissue pathophysiology and metabolism. Cell Metab. 2020, 31, 18-25. [CrossRef]
- Ma, C.; Kesarwala, A.H.; Eggert, T.; Medina-Echeverz, J.; Kleiner, D.E.; Jin, P.; et al. NAFLD causes selective CD4(+) T lymphocyte loss and promotes hepatocarcinogenesis. Nature 2016, 531, 253-257. [CrossRef]
- Wolf, M.J.; Adili, A.; Piotrowitz, K.; Abdullah, Z.; Boege, Y.; Stemmer, K.; et al. Metabolic activation of intrahepatic CD8+ T cells and NKT cells causes nonalcoholic steatohepatitis and liver cancer via cross-talk with hepatocytes. Cancer Cell 2014, 26, 549-564. [CrossRef]
Figure 1.
ZFP90 deficiency increases adiposity in mice. (A-C) mRNA expression of Zfp90 in (A) caloric restricted (CR), (B) aged, and (C) HFD-fed mice. (D) Representative 3D-Micro CT images of subcutaneous and visceral adipose tissue (left) and quantification of total, visceral (vis), and subcutaneous (sub) adipose tissue percentages (right) (n = 7 per group). (E) Representative hematoxylin and eosin (H&E) staining of epididymal (eWAT) and inguinal (iWAT) white adipose tissue (upper). Scale bar, 100 μm. Mean adipocyte sizes from eWAT and iWAT are shown in the bottom panel (n = 4 per group). (F) Oxygen consumption (VO2) and (G) respiratory quotient (RQ) in WT and KO mice fed a HFD (n = 4 per group). All data are presented as mean ± SEM. Statistical significance was determined by two-way ANOVA with Bonferroni’s multiple-comparisons test or Student’s two-tailed t-test; *p < 0.05, **p < 0.01, and ***p < 0.001. Abbreviations: AL, ad libitum; mo, month; sAT, subcutaneous adipose tissue; vAT, visceral adipose tissue.
Figure 1.
ZFP90 deficiency increases adiposity in mice. (A-C) mRNA expression of Zfp90 in (A) caloric restricted (CR), (B) aged, and (C) HFD-fed mice. (D) Representative 3D-Micro CT images of subcutaneous and visceral adipose tissue (left) and quantification of total, visceral (vis), and subcutaneous (sub) adipose tissue percentages (right) (n = 7 per group). (E) Representative hematoxylin and eosin (H&E) staining of epididymal (eWAT) and inguinal (iWAT) white adipose tissue (upper). Scale bar, 100 μm. Mean adipocyte sizes from eWAT and iWAT are shown in the bottom panel (n = 4 per group). (F) Oxygen consumption (VO2) and (G) respiratory quotient (RQ) in WT and KO mice fed a HFD (n = 4 per group). All data are presented as mean ± SEM. Statistical significance was determined by two-way ANOVA with Bonferroni’s multiple-comparisons test or Student’s two-tailed t-test; *p < 0.05, **p < 0.01, and ***p < 0.001. Abbreviations: AL, ad libitum; mo, month; sAT, subcutaneous adipose tissue; vAT, visceral adipose tissue.

Figure 2.
ZFP90 deletion exacerbates HFD-induced MASLD. (A) Body weight and (B) tissue weight changes during 16 weeks of CRF1 (control) or HFD feeding in WT and KO mice. (C) Representative H&E staining of liver sections (left; scale bar, 100 μm) and quantification of lipid accumulation in WT and KO mice (right) (n = 4 per group). (D) Serum ALT and AST levels. (E) Correlation analysis: epididymal fat mass/BW ratio vs. liver weight/BW ratio (left); ZFP90 mRNA levels in the liver or eWAT vs. liver weight/BW ratio in HFD-fed mice (right). (F) Heatmaps showing differentially expressed genes (DEGs) from the transcriptomic analysis of livers from HFD-fed mice (n = 3 per group). (G) mRNA expression of genes related to fatty acid uptake (Cd36, Lpl, Mogat1), inflammation (Tnfa, Cd68, Ccr2), and fibrosis (Col1a2) in the liver. Data are presented as mean ± SEM. Statistical significance was determined by Student’s two-tailed t-test; *p < 0.05, **p < 0.01, and ***p < 0.001. Abbreviations: BAT, brown adipose tissue; Br, brain; eWAT, epididymal white adipose tissue; I, inguinal; pWAT, perirenal, inguinal; Sp, spleen.
Figure 2.
ZFP90 deletion exacerbates HFD-induced MASLD. (A) Body weight and (B) tissue weight changes during 16 weeks of CRF1 (control) or HFD feeding in WT and KO mice. (C) Representative H&E staining of liver sections (left; scale bar, 100 μm) and quantification of lipid accumulation in WT and KO mice (right) (n = 4 per group). (D) Serum ALT and AST levels. (E) Correlation analysis: epididymal fat mass/BW ratio vs. liver weight/BW ratio (left); ZFP90 mRNA levels in the liver or eWAT vs. liver weight/BW ratio in HFD-fed mice (right). (F) Heatmaps showing differentially expressed genes (DEGs) from the transcriptomic analysis of livers from HFD-fed mice (n = 3 per group). (G) mRNA expression of genes related to fatty acid uptake (Cd36, Lpl, Mogat1), inflammation (Tnfa, Cd68, Ccr2), and fibrosis (Col1a2) in the liver. Data are presented as mean ± SEM. Statistical significance was determined by Student’s two-tailed t-test; *p < 0.05, **p < 0.01, and ***p < 0.001. Abbreviations: BAT, brown adipose tissue; Br, brain; eWAT, epididymal white adipose tissue; I, inguinal; pWAT, perirenal, inguinal; Sp, spleen.

Figure 3.
ZFP90-deficiency induces adipose tissue dysfunction and insulin resistance. (A) mRNA expression of genes related to lipogenesis (Cebpa, Pparg2, Acc1, Fasn), lipolysis (Lipa), pro-inflammatory markers (Cd68, Ccl3), and Adipoq in eWAT. (B-D) Glucose tolerance test (GTT) showing (B) blood glucose curves, (C) fasting blood glucose, and (D) insulin levels in WT and KO mice (n = 6-7 per group). (E) Protein levels of p-Akt (ser473) and Akt (upper) measured by western blotting in eWAT, with quantification (bottom). Data are presented as mean ± SEM. Statistical significance was determined by Student’s two-tailed t-test; *p < 0.05, **p < 0.01, and ***p < 0.001.
Figure 3.
ZFP90-deficiency induces adipose tissue dysfunction and insulin resistance. (A) mRNA expression of genes related to lipogenesis (Cebpa, Pparg2, Acc1, Fasn), lipolysis (Lipa), pro-inflammatory markers (Cd68, Ccl3), and Adipoq in eWAT. (B-D) Glucose tolerance test (GTT) showing (B) blood glucose curves, (C) fasting blood glucose, and (D) insulin levels in WT and KO mice (n = 6-7 per group). (E) Protein levels of p-Akt (ser473) and Akt (upper) measured by western blotting in eWAT, with quantification (bottom). Data are presented as mean ± SEM. Statistical significance was determined by Student’s two-tailed t-test; *p < 0.05, **p < 0.01, and ***p < 0.001.

Figure 4.
ZFP90 interacts with TRIM28 to modulate NF-κB activation. (A-D) Co-immunoprecipitation (Co-IP) analysis in COS-7 cells. Cells were transfected with FLAG-tagged ZFP90 and lysed 48 h post-transfection. Cell lysates were subjected to immunoprecipitation with an anti-Flag antibody, followed by (A) Coomassie Blue-stained SDS-PAGE to visualize ZFP90-binding proteins. (B) Arrows and letters indicate bands excised for identification by LC-MS/MS. Molecular mass standards (kDa) are indicated on the left. (C) Representative immunoblots of FLAG-immunoprecipitated proteins and input lysates using anti-FLAG and anti-Trim28 antibodies. (D) mRNA expression of Tnfα and Ccl2 in isolated non-parenchymal cells from WT and KO mice treated with LPS (0.1 mg/kg) or saline for 2 h. (F) Schematic model: the ZFP90-Trim28 complex acts to suppress excessive NF-κB activation. Data are presented as mean ± SEM. Statistical significance was determined by Student’s two-tailed t-test; **p < 0.01 and ***p < 0.001.
Figure 4.
ZFP90 interacts with TRIM28 to modulate NF-κB activation. (A-D) Co-immunoprecipitation (Co-IP) analysis in COS-7 cells. Cells were transfected with FLAG-tagged ZFP90 and lysed 48 h post-transfection. Cell lysates were subjected to immunoprecipitation with an anti-Flag antibody, followed by (A) Coomassie Blue-stained SDS-PAGE to visualize ZFP90-binding proteins. (B) Arrows and letters indicate bands excised for identification by LC-MS/MS. Molecular mass standards (kDa) are indicated on the left. (C) Representative immunoblots of FLAG-immunoprecipitated proteins and input lysates using anti-FLAG and anti-Trim28 antibodies. (D) mRNA expression of Tnfα and Ccl2 in isolated non-parenchymal cells from WT and KO mice treated with LPS (0.1 mg/kg) or saline for 2 h. (F) Schematic model: the ZFP90-Trim28 complex acts to suppress excessive NF-κB activation. Data are presented as mean ± SEM. Statistical significance was determined by Student’s two-tailed t-test; **p < 0.01 and ***p < 0.001.

Figure 5.
ZFP90 deficiency alters intrahepatic T-cell dynamics. (A) mRNA expression of genes related to T-cell dynamics (Cd4, Cd8a, Tlr7), Zfp90, and Trim28 in eWAT and livers from WT and KO mice. (B) mRNA expression of ZFP90 in MT-2 and SKW-3 cells. (C) mRNA expression of ZFP90, TNFα, IL2TA, and IL2RB in MT-2 cells transfected with control or ZFP90-siRNA. (D) Representative immunoblot analysis of p65 and Lamin B1 protein levels in the livers from WT and KO mice. Data are presented as mean ± SEM. Statistical significance was determined by Student’s two-tailed t-test; *p < 0.05, **p < 0.01, and ***p < 0.001.
Figure 5.
ZFP90 deficiency alters intrahepatic T-cell dynamics. (A) mRNA expression of genes related to T-cell dynamics (Cd4, Cd8a, Tlr7), Zfp90, and Trim28 in eWAT and livers from WT and KO mice. (B) mRNA expression of ZFP90 in MT-2 and SKW-3 cells. (C) mRNA expression of ZFP90, TNFα, IL2TA, and IL2RB in MT-2 cells transfected with control or ZFP90-siRNA. (D) Representative immunoblot analysis of p65 and Lamin B1 protein levels in the livers from WT and KO mice. Data are presented as mean ± SEM. Statistical significance was determined by Student’s two-tailed t-test; *p < 0.05, **p < 0.01, and ***p < 0.001.

Figure 6.
Clinical correlation of ZFP90 with MASLD severity. (A) mRNA expression of ZFP90 in the livers of healthy controls and patients with MASLD. (B, C) Correlation between Zfp90 mRNA levels and serum ALT, AST, and GGT levels in (B) human MASLD patients and (C) HFD-fed mice. (D) Schematic summary: ZFP90 functions as a pivotal regulator of hepatic immunometabolic homeostasis. ZFP90 deficiency exacerbates MASLD/MASH through a multi-hit process involving lipid metabolism disorders, adipose tissue dysfunction, hyper-activation of NF-κB–dependent inflammatory signaling, and altered T-cell dynamics. Data are presented as mean ± SEM. Statistical significance was determined by Student’s two-tailed t-test; **p < 0.01.
Figure 6.
Clinical correlation of ZFP90 with MASLD severity. (A) mRNA expression of ZFP90 in the livers of healthy controls and patients with MASLD. (B, C) Correlation between Zfp90 mRNA levels and serum ALT, AST, and GGT levels in (B) human MASLD patients and (C) HFD-fed mice. (D) Schematic summary: ZFP90 functions as a pivotal regulator of hepatic immunometabolic homeostasis. ZFP90 deficiency exacerbates MASLD/MASH through a multi-hit process involving lipid metabolism disorders, adipose tissue dysfunction, hyper-activation of NF-κB–dependent inflammatory signaling, and altered T-cell dynamics. Data are presented as mean ± SEM. Statistical significance was determined by Student’s two-tailed t-test; **p < 0.01.


Table 1.
Clinical and biochemical characteristics of the whole population.
| Variable (mean±SD) | Non-MASLD control subjects | MASLD subjects | p Value |
|---|---|---|---|
| Number of subjects | 12 | 11 | |
| Female/male, % | 50/50 | 64/36 | |
| Age, years | 50.89±9.36 | 56.73±10.57 | NS |
| BMI, kg/m2 | 20.57±0.82 | 29.34±5.30 | <0.00002 |
| Fasting plasma glucose, mg/dL | 93.25±3.96 | 115.50±28.49 | <0.02 |
| SABP, mm Hg | 108.42±13.27 | 123.82±15.91 | <0.02 |
| DABP, mm Hg | 73.67±15.42 | 71.73±8.06 | NS |
| C-reactive protein, mg/dL | 0.03±0.03 | 0.16±0.08 | <0.00004 |
| HDL-cholesterol, mg/dL | 80.33±28.07 | 51.55±18.01 | <0.009 |
| LDL-cholesterol, mg/dL | 134.42±25.2 | 109.27±31.98 | <0.05 |
| Triglyceride, mg/dL | 76.50±23.00 | 170.09±69.40 | <0.0003 |
| ALT, U/L | 15.33±7.89 | 79.55±60.49 | <0.002 |
| AST, U/L | 17.25±3.02 | 62.36±59.17 | <0.02 |
| GGT, U/L | 15.75±6.55 | 66.18±35.33 | <0.00009 |
Results are expressed as mean ± SD. ALT and AST, serum alanine and aspartate aminotransferase; BMI, body mass index; DABP, diastolic arterial blood pressure; GGT, γ-glutamyl-transferase; HDL, high-density lipoprotein; LDL, low-density lipoprotein; NS, non-significant; SABP, systolic arterial blood pressure.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



