Submitted:
17 June 2026
Posted:
18 June 2026
You are already at the latest version
Abstract
Background: Due to the difficulty of identifying the window of receptivity, a molecular understanding of endometrial changes after fertilization is essential for improving the effectiveness of in vitro fertilization (IVF). This multi-level study examines the compo-sition of cytokines and extracellular vesicles (EVs) in uterine lavage samples obtainable via a noninvasive procedure in mice. The goal is to offer a new perspective on the tim-ing of embryo transfer. Methods: We performed flow cytometric analyses of inflamma-tory cytokines and EVs in uterine lavage samples collected from CD1 mice at the pre-receptive stage (2.5 days post coitum [dpc]) and the receptive stage (4.5 dpc). We also assessed endometrial progesterone receptor expression via immunohistochemistry and evaluated the expression of adhesion molecules related to implantation by RT-PCR. Re-sults: Significant differences were found in the concentrations of inflammatory cyto-kines (IL-6, IFN-γ, and MCP-1), the EV pattern in uterine lavage fluid, and the expres-sion of some genes related to implantation (CD44, CD81, and CD29). These results re-veal uterine lavage fluid markers associated with endometrial receptivity. Using these noninvasive markers can improve the effectiveness of IVF by enabling more precise timing of embryo transfer.
Keywords:
uterine lavage
; adhesion
; extracellular vesicles
; receptivity markers
1. Introduction
Pregnancy loss is a significant event in human reproduction and is also notable among mammals. Estimates suggest that it occurs in approximately 70% of all pregnancies, with a substantial proportion—around 60%—happening within the first 6 weeks [1]. Moreover, more than 50% of these losses manifest as biochemical miscarriages [2]. This profound impact also persists in IVF-ICSI procedures, even when the transferred embryos are of good quality and undergo genetic testing in assisted reproductive procedures. This situation is concerning because, while the global fertility rate is declining, more and more children are being born worldwide through assisted reproductive technologies (ART). This proportion usually falls between 2% and 3%, regarding developed countries [3].
A pivotal factor in ART is the implantation of a competent embryo at the time when the endometrium is receptive [4]. Although both maternal and embryonic factors influence uterine receptivity, the embryonic side is now better characterized [5]. Poor-quality embryos send inhibitory messages that prevent the development of the essential endometrial microenvironment, including the transformation of endometrial stromal cells into decidual stromal cells [6]. Despite the transfer of high-quality, genetically examined embryos in assisted reproductive procedures, the critical event remains the implantation of a competent embryo within the narrow window of the decidualized endometrium, known as WI. However, information about the implantation window is limited. The implantation window, essential for proper embryo implantation, spans a mere 12-24 hours in mice and 36 hours in humans [7,8]. Deviations from this optimal timeframe by 12 hours in humans reduce the clinical pregnancy rate from 44% to 23% in IVF cycles, significantly contributing to miscarriages, especially during biochemical pregnancies [9]. Determining the ideal time for embryo transfer presents challenges, as ultrasonographic measurements have not yielded the expected results. Commercial tests utilizing gene examinations and endometrial assays during transfer cycles lack general acceptance. Molecular examinations of uterine fluid offer a promising method for investigation [10,11,12,13,14].
Although progesterone (P), estradiol (E2), and their receptors [15,16,17,18,19] are the main contributors to implantation, embryo-endometrial crosstalk is more complex. It involves adhesion molecules, extracellular matrix remodeling, soluble mediators, and extracellular vesicles.
EVs are cell-derived particles surrounded by a double phospholipid membrane. EV production is evolutionarily conserved and characteristic of prokaryotic, eukaryotic, and plant cells, regardless of their life cycle stage. Due to their molecular composition of proteins, lipids, carbohydrates, and nucleic acids on both exterior and interior surfaces, EVs participate in complex physiological and pathological processes. EV-mediated cellular communication can occur locally (the paracrine effect) or systemically at distant sites in the body. Because the molecular composition of EVs is unique to the cells that secrete them, they can serve as potential biomarkers for diagnostics and prognostics. EVs can be detected in various biological fluids, including blood and urine. This makes collecting EV-containing samples relatively simple with minimally invasive procedures (’liquid biopsy’), making EVs a promising alternative to solid tissue biopsies. Additionally, more invasive tissue biopsies only reflect the biopsy site, not the entire endometrium [14,20]. Several studies have demonstrated the role of extracellular vesicles in human reproduction, including follicle development (folliculogenesis), fertilization, implantation, and maintenance of pregnancy [21,22]. From a diagnostic perspective, EVs can be used to monitor embryo quality during in vitro fertilization (IVF) treatment [23,24].
In summary, EVs carry complex messages that influence the function of target cells. Therefore, the cargo of embryo-derived EVs can affect the activity of endometrial epithelial and immune cells, thus modulating the biosensor function of the endometrium.
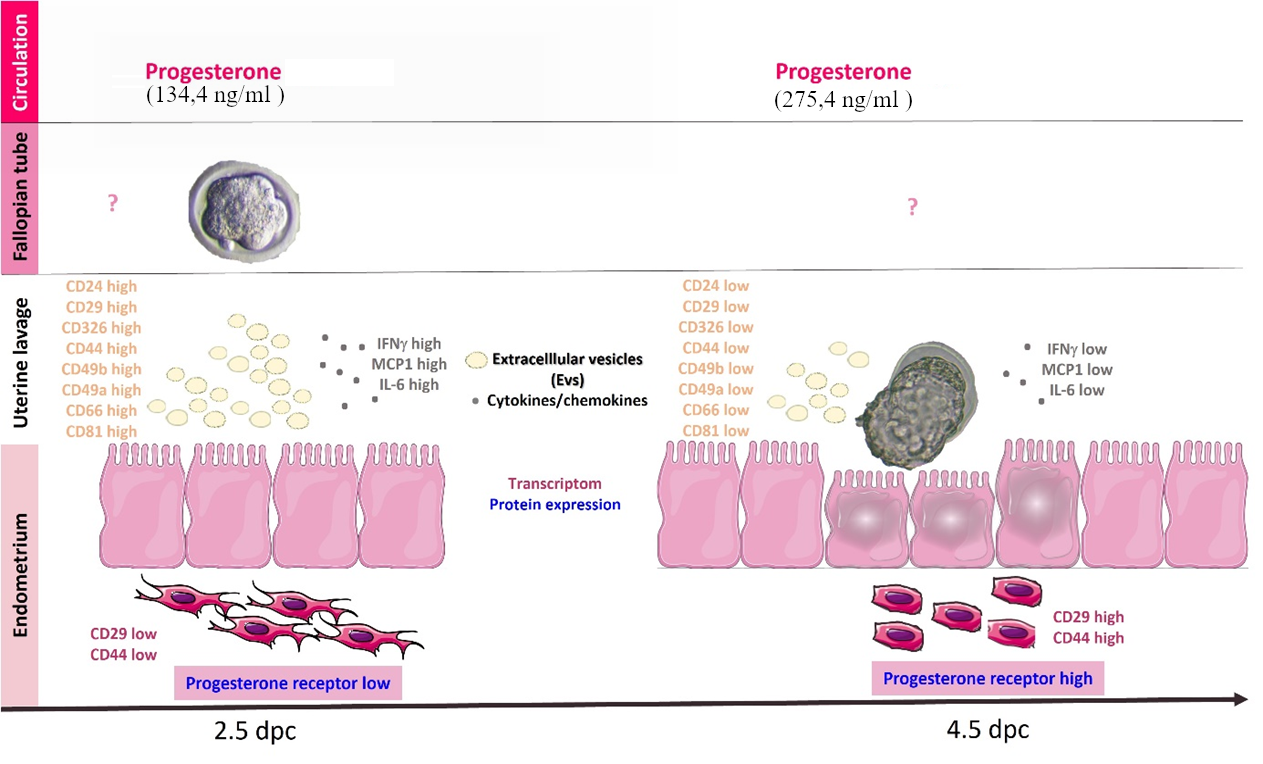
Our research aims to identify biomarkers that could improve the success rate of assisted reproductive procedures and deepen our understanding of the mechanisms of embryo implantation. The mouse model is appropriate for our questions because WI in mice occurs at 4.5–5 days post-coitum (dpc), enabling clear differentiation between the pre-receptive and receptive stages. These accurate spatiotemporal results in mice could help pinpoint truly relevant molecular indicators, including their levels and trends, which may also serve as valuable markers in humans. Therefore, we aimed to investigate serum progesterone levels, progesterone receptor expression in the endometrium, and soluble mediators, including cytokines and extracellular vesicles, in uterine lavage fluid. We also assessed the expression of adhesion molecule genes in the endometrium.
2. Materials and Methods
2.1. Animals and Ethics
Superovulation Treatment: Sixty-five eight- to twelve-week-old CD1 female mice (Charles River, Germany) received an injection of five international units (IU) of Meriofert (Kit, IBSA Institut Biochimique SA, Lugano, Switzerland). Forty-eight hours later, the mice received 5 IU of human chorionic gonadotropin (Chorulon; Ferring Pharmaceuticals A/S, Copenhagen, Denmark) and were paired with CD1 males. The presence of a vaginal plug was considered an indication of 0.5 days of pregnancy.
Sixty-one mice tested positive and were sacrificed at 2.5 or 4.5 days post coitum (dpc). Blood samples were collected after decapitation. At 2.5 dpc, embryos at the four- to six-cell stage were retrieved from 26 mice. At 4.5 dpc, blastocyst-stage embryos were flushed from the fallopian tubes and uterine cornua of 27 mice. The retrieved embryos were cultured for further experiments. The resident embryos in the oviducts and uterine horns were counted and recorded. Only the embryo-positive cases (59 mice) and their corresponding endometrial samples were included in subsequent analysis at both time points (26 mice at 2.5 dpc and 27 mice at 4.5 dpc). The uteri were then dissected and prepared for endometrial tissue collection. The remaining six animals were used for immunohistochemical and immunofluorescent staining. The uterine cornua were flushed separately. To confirm that implantation had already occurred by that time, histological examinations were also performed at 5.5 dpc.
All methods were carried out according to relevant guidelines and regulations. The Animal Health Committee of Baranya County approved the experimental protocols. No: BAI/35/859-6/2019).
2.2. Histopathology and Immunohistochemistry
Immunohistochemical and immunofluorescence staining were conducted on paraffin-embedded uterine sections at 2.5, 4.5, and 5.5 days post-coitum (dpc). (n=6/dpc) Antigen retrieval was achieved using a citrate buffer solution (Agilent Dako, Glostrup, Denmark) at pH 6.0 with microwave heating. Endogenous peroxidase activity was inhibited by incubating the samples in 3% hydrogen peroxide at room temperature for 10 minutes. The sections were labeled with a polyclonal anti-progesterone receptor antibody (Cell Signaling Technology, Danvers, MA, USA) to detect progesterone receptors. We visualized the binding of the primary antibodies using either horseradish peroxidase-conjugated goat anti-rabbit IgG (Abcam plc., Cambridge, UK) with 3,3’-diaminobenzidine (DAB) substrate (Agilent Dako, Glostrup, Denmark) or fluorescein isothiocyanate (FITC)-conjugated goat anti-rabbit IgG (Abcam plc., Cambridge, UK). We analyzed the histological sections using an Olympus FV1000 confocal microscope (Olympus Europe GmbH, Hamburg, Germany). We conducted analysis using a 20x dry objective (NA: 0.75) in photon counting mode. That allowed us to quantify fluorescence intensities post-analysis with ImageJ software (NIH, Bethesda, MD).
2.3. Isolation of Endometrial Samples
Endometrial samples were obtained by carefully scraping the longitudinal dissected uterine horns. The uterus was subsequently scraped in a sterile PBS solution. The collected material was transferred with minimal liquid volume. The tissue mass was determined gravimetrically, and the samples were stored at -80℃ in 200 µL of RNA Later (InvitrogenTM, a Thermo Fisher Scientific company, Waltham, MA, USA). (Supplementary Figure S1)
2.4. RNA Isolation from Endometrium and RT-PCR
Total RNA was extracted using the Quick-RNA™ MiniPrep Kit (Zymo Research, USA) according to the manufacturer’s instructions. The concentration and purity of the isolated RNA were quantified and assessed using a NanoDrop microvolume spectrophotometer (NanoDrop ND-1000, ThermoFisher Scientific, USA).
cDNA synthesis was performed using a SensiFast cDNA synthesis kit (Bioline, Taunton, MA, USA). All primers used in this study were synthesized by ThermoFisher Scientific (USA).
Relative expression levels of CD24, CD29, CD326, CD44, CD45, CD49b, CD49e, CD66a, CD9, CD81, β2-microglobulin, and β-actin were measured using the SensiFAST SYBR Hi-ROX Kit (Bioline, Taunton, MA). (Supplementary Table S1) Samples were analyzed using a Bio-Rad CFX96 Touch Real-Time PCR machine (Bio-Rad Laboratories, Inc., USA) with the following program settings: two minutes at 95 °C, followed by 40 cycles of five seconds at 95 °C and 30 seconds at 60 °C. The results are relative to the expression of GAPDH and HPRT as housekeeping genes. Each sample was tested in triplicate, and the comparative critical threshold (ΔΔCT) method was used to determine the relative expression levels of the genes.
2.5. Uterine Lavage Collection
Uterine cornua were flushed separately with 300 µl of PBS solution under sterile conditions. We excised both horns of the uterus just above the portio uteri. As we always conducted tube flushes, the end of the fallopian tube was left free. Our aim was to retain the longest possible portion of the horn for the flushing procedure. At the end of the fallopian tube, we inserted a strip connected to a 1 ml syringe. To prevent backflow, we occluded the lumen with a clamp. We flushed each horn once with 300 µl sterile PBS. After centrifugation at 3000 rpm, the supernatant was removed and supplemented with 2.5 µl of protease inhibitor (Complete™ Mini Protease Inhibitor Cocktail, Roche Diagnostics, GmbH, Mannheim, Germany). The samples were stored at -80°C to study soluble factors and extracellular vesicles. For flushing the uterine horn, we first isolated a short segment of the horn and its tube. The embryos were flushed from the fallopian tubes separately, and the number of embryos present in the tubes was counted. The number of blastocysts staying in the uterine horns at the 4.5 dpc was also counted in the uterine lavage. After the embryos were removed, the lavage was preserved for storage
2.6. Cytokine Measurements
Levels of inflammatory cytokines and chemokines, including IL-1β, IFN-γ, TNF-α, MCP-1, IL-10, IL-6, IL-17A, CX3CL1, IL-15, TGF-β, and LIF, were measured in uterine lavage using a LEGENDplex™ Mouse Mix-and-Match 11-plex panel (BioLegend, San Diego, CA) according to the manufacturer’s instructions. Legendplex is a highly sensitive, bead-based flow cytometry method that can detect several analytes simultaneously. The custom panel was designed to detect cytokines that are potentially associated with embryo implantation [25,26].
2.7. EV Isolation and Characterization
We characterized uterine lavage extracellular vesicles using the MACSPlex EV kit (Miltenyi Biotec, USA), which detects 37 different surface epitopes. Exosomes were pre-enriched by ultracentrifugation at 100,000 x g for one hour at 4 °C using an Optima Max XP benchtop ultracentrifuge with an MLS-50 swinging-bucket rotor (Becton-Dickinson, San Jose, CA, USA). The protein content was determined using the Pierce™ BCA Protein Assay Kit (Thermo Fisher Scientific, USA), following the manufacturer’s recommendations to ensure sample comparability. (Supplementary Figure S2)
2.8. Serum Separation and Hormone Measurement
Peripheral blood samples were collected by decapitating the animals immediately before flushing the fallopian tubes. After coagulation, the samples were centrifuged at 1,000 x g for 10 minutes in a refrigerated centrifuge, after which the serum was collected. Serum progesterone concentrations were determined using automated, competitive, chemiluminescent immunoassays (B84493, 33550), which utilized the Beckman Coulter UniCel DxL 800 Access Immunoassay System (Beckman Coulter, Inc., Brea, CA, USA).
2.9. Statistical Analyses
We used a chi-square test to analyze Pr expression in the endometrium and primary decidua. To prevent type I errors, we adjusted the p-values using the Bonferroni correction. We used a two-tailed unpaired Student’s t-test for normally distributed data.
We employed IBM SPSS Statistics (version 22.0, IBM Corp., USA) and GraphPad Prism (version 9.4, GraphPad Software, La Jolla, California, USA) for statistical analysis. We used Cytoscape 3.10.3 (https://cytoscape.org/) software to visualize and analyze the protein-protein interactions between uterine lavage soluble mediators and EVs. The significance level was set at p <0.05 for all tests.
We conducted a predictive analysis of endometrial receptivity using a combination of logistic regression and principal component analysis.
3. Results
3.1. Endometrial Mass
Hormonal milieu-driven endometrial thickness was assessed by weighing isolated endometrial samples. The average weight of the samples was 22.7 (SD±9.3) mg on day 2.5, increasing to 57.6 (SD±13.4) mg by day 4.5. This increase in endometrial tissue mass was statistically significant (P = 0.0003). (Figure 1)
3.2. Serum Progesterone
No significant difference was observed in serum progesterone levels from 2.5 dpc (160.8 ng/ml ± 30.64) to 4.5 dpc (260.96 ng/ml ± 76). (Supplementary Figure S3)
3.3. Progesterone Receptor Expression in the Endometrium
To investigate endometrial progesterone sensitivity, we examined progesterone receptor (Pr) expression using immunofluorescence at 2.5, 4.5, and 5.5 days post-coitum (dpc). (Figure 1A,B) We conducted a comparative analysis of Pr expression using quantitative imaging across six animals, with triplicate measurements per sample. The expression of Pr was significantly higher in both the 4.5 dpc (P=0.002) and 5.5 dpc (P = 0.0002) samples than in the 2.5 dpc sample. (Figure 1C)
3.4. Inflammatory Mediators and EV Patterns in Uterine Lavage
A custom panel was designed based on scientific literature to characterize embryo implantation-related cytokines and chemokines in uterine lavage samples. The IL-1β, TNF-α, IL-17A, CX3CL1, IL-15, TGF-β, and LIF remained consistent, while the IL-10 level was below the detection limit. Three cytokines were significantly down-regulated at 4.5 dpc, including IL-6 (2.5 dpc: 2.045 ± 0.9 pg/mL; 4.5 dpc: 1.197 ± 0.6 pg/mL; p=0.04), MCP-1 (2.5 dpc: 3.091 ± 1.1 pg/mL; 4.5 dpc: 2.044 ± 0.7 pg/mL; p=0.004), and IFN-γ (2.5 dpc: 3.422 ± 1.5 pg/mL; 4.5 dpc: 1.726 ± 1.1 pg/mL; p<0.0001) (Figure 2A). We conducted a Pearson’s correlation test to assess the interactions among cytokines and chemokines in uterine lavage. The correlation analysis revealed positive correlations among the proinflammatory cytokines IL-1, IFN-γ, TNF-α, MCP-1, and IL-6 (Figure 2B).
We characterized extracellular vesicles (EVs) from uterine lavage using flow cytometry in accordance with the MIFlowCyt-EV guidelines. We enriched the EVs through ultracentrifugation and assessed their surface protein profile with a bead-based, multiplex assay. Cell adhesion-associated markers, including CD24, CD29, CD326, CD44, CD49b, CD49a, and CD66, were highly expressed on EVs in 2.5 dpc samples compared to 4.5 dpc samples (Figure 3). (Supplementary Figure S4)
Pathway analysis showed that parameters with significant variation in uterine lavage patterns are interrelated and part of a complex mechanism. Analyzing the network revealed a total of 14 nodes. Most of the highlighted Gene Ontology (GO) pathways are associated with cell adhesion: GO:0030155 (regulation of cell adhesion), GO:0045785 (positive regulation of cell adhesion), GO:0022407 (regulation of cell-cell adhesion), GO:0098609 (cell-cell adhesion), and GO:0007155 (cell adhesion). The regulation of the adaptive immune response (GO:0002819) was also displayed. (Figure 4)
3.5. Endometrial Adhesion Molecule Expression During the Preimplantation Period
The key characteristics of uterine lavage enhance the receptivity of the endometrium, indicating a series of adhesive interactions. In our experimental system, we examined 11 molecules associated with embryo attachment (CD24, CD29, CD326, CD44, CD45, CD49b, CD49e, CD66a, CD9, CD81, and β2-microglobulin) at the transcriptome level on days 2.5 and 4.5 dpc. CD29, CD44, and CD81 displayed significantly lower expression levels at 2.5 dpc [6]. (Figure 5)
4. Discussion
In this study, we developed a workflow to distinguish between pre-receptive and receptive endometria by analyzing extracellular vesicles (EVs) and soluble mediators in uterine lavage samples. Our work is significant because it uses an in vivo animal model in which the blastocyst awaiting implantation and the endometrium are in constant communication. Although our experimental system has limitations — namely, that no animal model can be directly applied to the human body — it is unique in enabling long-term, dynamic studies that are not permitted in humans. At the same time, our results can inform, supplement, and clarify human endpoint measurement systems.
The 2.5 and 4.5 dpc stages of mouse pregnancy were the focus of our study. 2.5 dpc has been proven to correspond to the pre-receptive phase, while 4.5 dpc is the most suitable time for embryo implantation. That is evident from the fact that blastocyst collection from the oviduct is easily performed in mice at 4.5 dpc. By 4.5 dpc, the implantation process is in the pre-adhesion stage; therefore, more embryos can be flushed from the uterus than from the tubes, but they are still flushable. By 5.5 dpc, the embryo has already implanted. Our experiment enabled us to examine embryos and the endometrium in parallel, spatio-temporally, with precise timing of the implantation window. Of course, this procedure cannot be performed on humans and requires extensive validation in humans, which is a limitation of our work.
It is well known that a balance of estrogen and progesterone is essential for the development of a receptive endometrium [27]. During the pre-ovulatory phase, estrogen stimulates endometrial proliferation and increases progesterone receptor expression, thereby enhancing endometrial sensitivity to progesterone. Following ovulation, progesterone triggers the transformation of the endometrium to prepare it for implantation. Additionally, progesterone helps induce maternal immunological tolerance in the early stages of pregnancy. In our study, serum progesterone levels showed only an insignificant increase (134.4 ng/ml ± 147.3 versus 275.4 ng/ml ± 288.3) between days 2.5 and 4.5 dpc. However, a significant increase in progesterone receptor (PR) expression was observed at day 4.5 dpc. That suggests that the endometrium had become more sensitive to progesterone.
In clinical practice, the ERA (endometrial receptivity assay) method is commonly used to assess gene expression and determine the optimal timing for embryo implantation during in vitro fertilization. This method requires an invasive tissue biopsy, which is not allowed during the embryo transfer cycle [28]. Additionally, in humans, the window of implantation (WI) date is based on the statistical average from earlier studies. It ranges from LH+6 to LH+10 days [29,30], which only partially reflects the actual WI timing in transfer cycles. It is well known that implantation timing can vary between cycles, even in fertile women [14].
In light of the ERA tests performed in clinical practice, we also conducted gene expression analyses of endometrial samples. As our work aimed to demonstrate the applicability of non-invasively collected uterine lavage fluid, we searched for genes whose expression differed between the 2.5 dpc and 4.5 dpc stages in uterine lavage samples [31].
Significantly higher levels of CD44, CD29, and CD81 were detected in the endometrium at 4.5 days compared to 2.5 days. CD44 plays a pivotal role in interacting with various surface ligands, such as fibronectins, collagens, and laminins, which adorn the endometrial landscape. The CD29 integrin promotes embryo attachment to the endometrium and enhances trophoblast cell invasiveness during implantation, playing an essential role in establishing pregnancy [32]. CD81 is a widely expressed tetraspanin that initiates signaling cascades by orchestrating interactions between membrane-associated proteins, thereby regulating cell adhesion, migration, and invasion [33]. CD81 forms a complex with integrins on the cell membrane and interacts with several transmembrane proteins, including CD9 and CD29 [34]. It also highlights their role in establishing direct cell-to-cell connections. Tetraspanins are involved in remodeling the affected cell membranes, which is essential during embryo implantation [35]. Changes in the expression levels of CD29 and CD81 may indicate a ’higher state of readiness’ in the endometrium, preparing it for embryo implantation between days 4.5 and 5.5 post coitum.
Uterine fluid collection is noninvasive and, based on the data, reflects the state of the endometrium. These tests involve washing the uterine cavity with sterile physiological solutions to collect endometrium-derived mediators [11,20]. Uterine lavage is of great interest, and several groups have demonstrated that its molecular composition — including proteins, microRNAs (miRNAs), and extracellular vesicles — can be associated with the physiological and pathological conditions of the endometrium [14,36]. That is why we used uterine fluid examinations in mice, despite knowing that there are critical interspecies differences, which is a limitation of our work.
Using a multiplex bead-based method, we characterized mouse uterine lavage EVs at 2.5 dpc and 4.5 dpc. Based on membrane protein patterns, we identified heterogeneous EV populations in uterine lavage. At 2.5 days post-coitum, which corresponds to the pre-receptive stage, we detected a significantly higher number of extracellular vesicles (EVs) expressing the tetraspanins CD81 and CD9, as well as a significantly higher number of EVs expressing molecules involved in cell adhesion, such as CD24, CD29, CD326, CD44, CD49b, CD49e, and CD66a. The question arises as to whether the observed differences reflect only higher total EV counts or represent actual biological variance.
Nevertheless, this limitation remains in our study. The content of EVs in biological fluids is determined by the balance between EV secretion and uptake by target cells. The increased number of EVs expressing adhesion molecules in the 2.5-day samples may indicate that the endometrium is in a pre-receptive state, which does not yet require these molecules. However, by day 4.5, the mouse endometrium may begin to retain these molecules in preparation for implantation.
In placental animals, implantation describes the process by which the blastocyst (conceptus) attaches to, and invades, the endometrium of the uterus. The first phase of the implantation process is apposition. During this phase, the newly hatched blastocyst loosely adheres to the endometrial epithelium and often ’rolls’ to the eventual site of implantation, where it becomes firmly attached. Successful implantation is associated with several regulatory factors, including local concentrations of chemokines and cytokines. It is widely accepted that proinflammatory cytokines, such as IL-6, are among the ’pro-invasive’ factors. However, our results contradict this, as the process is much more nuanced due to cytokines’ pleiotropy. On the other hand, it is well known that the glycocalyx composition of endometrial epithelial cells influences blastocyst adhesion. Decreased MUC1 expression is associated with receptivity. The presence of MUC1 prevents blastocyst attachment. Numerous studies confirm that proinflammatory cytokines increase MUC1 expression. Therefore, if the concentration of proinflammatory cytokines is lower, this will also affect MUC1 expression in the endometrium, potentially promoting implantation. During the later phase of implantation, an inflammatory immune environment is required, so a proinflammatory shift in the cytokine pattern can be expected. In our experimental system, we selected the 4.5 dpc time point as this represents the time of blastocyst attachment (’opportunity for attachment’) [37,38].
All of these factors highlight the importance of our experimental model, as postcoital time can be accurately tracked in mice and is demonstrably correlated with endometrial condition. In human practice, where WI is available over a wider time range, it is not possible to standardise the sampling times required for accurate kinetic analysis.
During data processing, we created a logistic regression model for predictive analysis, incorporating the 61 examined parameters. We designated ’receptivity’ as the target variable, based on the 4.5 dpc results. On the other hand, we performed a principal component analysis of the dataset to identify the main trends (or principal components). By comparing these models, we identified markers that can predict endometrial receptivity, including CD326, CD44, CD81, IL-1β, IL-10, IL-17A, and MCP1. Consistent with the literature, our results suggest that uterine lavage fluid may be a valuable indicator of endometrial receptivity.
5. Conclusion
There are significant differences in the possibilities for studying WI between human and mouse models. In humans, the date of WI is based on the statistical average from earlier studies. It only partially reflects the actual WI timing in transfer cycles. It is well known that implantation time can vary between cycles, even in fertile women. In our research, the data represent successful cycles in mice. In the cases examined, embryo-maternal crosstalk occurred from mating through sample collection. Differences at the two specific time points allow not only the measurement of their levels but also the examination of different aspects of their changes across successful cycles. This work lays a foundation for identifying shared, translatable biomarkers within the valid timing of WI.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
The experiments were conducted by ZB and TCS. ZB, BG, and TCS were involved in measuring and analyzing confocal microscopy data. EP and NF were involved in flow cytometric analysis. NB was involved in RT-PCR measurements. BZ and ÉP were involved in reviewing and editing the manuscript. ZB and ÉP were responsible for the conceptualization, data analysis, and writing and editing of the manuscript.
Funding
This work was supported by the National Laboratory on Human Reproduction. Project no. RRF-2.3.1-21-2022-00012, titled "National Laboratory on Human Reproduction," was conducted with backing from the European Union’s Recovery and Resilience Facility under the Széchenyi Plan Plus Program. The project received funding from the HUN-REN Hungarian Research Network and STIA_2024 (19279/PMKP/2025). Project no. TKP-2021-EGA-10 was executed with support from the National Research, Development, and Innovation Fund of Hungary, which was funded under the TKP-2021-EGA program. The PTE Medical School (Szolcsányi Foundation) supported the work.
Acknowledgments
We are grateful for the technical assistance of Judit Pejtsik.
Conflicts of Interest
The authors declare that the research was conducted without commercial or financial relationships that could be perceived as a potential conflict of interest.
Generative AI Statement: The author(s) declare that no Generative AI was used in the creation of this manuscript.
References
- Macklon, N.S.; Geraedts, J.P.; Fauser, B.C. Conception to ongoing pregnancy: the ‘black box’of early pregnancy loss. Hum. Reprod. Update 2002, 8, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Craciunas, L.; Gallos, I.; Chu, J.; Bourne, T.; Quenby, S.; Brosens, J.J.; Coomarasamy, A. Conventional and modern markers of endometrial receptivity: a systematic review and meta-analysis. Hum. Reprod. Update 2019, 25, 202–223. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, N.V.; Schumacher, A.E.; Aali, A.; Abate, Y.H.; Abbasgholizadeh, R.; Abbasian, M.; Abbasi-Kangevari, M.; Abbastabar, H.; Abd ElHafeez, S.; Abd-Elsalam, S. Global fertility in 204 countries and territories, 1950–2021, with forecasts to 2100: a comprehensive demographic analysis for the Global Burden of Disease Study 2021. The lancet 2024, 403, 2057–2099. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.; Sun, X.; Dey, S.K. Mechanisms of implantation: strategies for successful pregnancy. Nat. Med. 2012, 18, 1754–1767. [Google Scholar] [CrossRef] [PubMed]
- Paria, B.; Huet-Hudson, Y.; Dey, S. Blastocyst’s state of activity determines the" window" of implantation in the receptive mouse uterus. Proc. Natl. Acad. Sci. 1993, 90, 10159–10162. [Google Scholar] [CrossRef] [PubMed]
- Brosens, J.J.; Bennett, P.R.; Abrahams, V.M.; Ramhorst, R.; Coomarasamy, A.; Quenby, S.; Lucas, E.S.; McCoy, R.C. Maternal selection of human embryos in early gestation: insights from recurrent miscarriage. Proceedings of the Seminars in cell & developmental biology 2022, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Kalakota, N.R.; George, L.C.; Morelli, S.S.; Douglas, N.C.; Babwah, A.V. Towards an improved understanding of the effects of elevated progesterone levels on human endometrial receptivity and oocyte/embryo quality during assisted reproductive technologies. Cells 2022, 11, 1405. [Google Scholar] [CrossRef] [PubMed]
- Aplin, J.D. Adhesion molecules in implantation. Implant. Early Dev. 1997, 49–60. [Google Scholar]
- Enciso, M.; Aizpurua, J.; Rodríguez-Estrada, B.; Jurado, I.; Ferrández-Rives, M.; Rodríguez, E.; Pérez-Larrea, E.; Climent, A.; Marron, K.; Sarasa, J. The precise determination of the window of implantation significantly improves ART outcomes. Sci. Rep. 2021, 11, 13420. [Google Scholar] [CrossRef] [PubMed]
- Van der Gaast, M.; Macklon, N.; Beier-Hellwig, K.; Krusche, C.; Fauser, B.; Beier, H.; Classen-Linke, I. The feasibility of a less invasive method to assess endometrial maturation—comparison of simultaneously obtained uterine secretion and tissue biopsy. BJOG An. Int. J. Obstet. Gynaecol. 2009, 116, 304–312. [Google Scholar]
- Boomsma, C.; Kavelaars, A.; Eijkemans, M.; Lentjes, E.; Fauser, B.; Heijnen, C.; Macklon, N. Endometrial secretion analysis identifies a cytokine profile predictive of pregnancy in IVF. Hum. Reprod. 2009, 24, 1427–1435. [Google Scholar] [CrossRef] [PubMed]
- Hannan, N.; Paiva, P.; Meehan, K.; Rombauts, L.; Gardner, D.; Salamonsen, L. Analysis of fertility-related soluble mediators in human uterine fluid identifies VEGF as a key regulator of embryo implantation. Endocrinology 2011, 152, 4948–4956. [Google Scholar] [CrossRef] [PubMed]
- Edgell, T.A.; Evans, J.; Lazzaro, L.; Boyes, K.; Sridhar, M.; Catt, S.; Rombauts, L.J.; Vollenhoven, B.J.; Salamonsen, L.A. Assessment of potential biomarkers of pre-receptive and receptive endometrium in uterine fluid and a functional evaluation of the potential role of CSF3 in fertility. Cytokine 2018, 111, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Giacomini, E.; Scotti, G.; Vanni, V.; Lazarevic, D.; Makieva, S.; Privitera, L.; Signorelli, S.; Cantone, L.; Bollati, V.; Murdica, V. Global transcriptomic changes occur in uterine fluid-derived extracellular vesicles during the endometrial window for embryo implantation. Hum. Reprod. 2021, 36, 2249–2274. [Google Scholar] [CrossRef] [PubMed]
- Szekeres-Bartho, J.; Wegmann, T. A progesterone-dependent immunomodulatory protein alters the Th1Th2 balance. J. Reprod. Immunol. 1996, 31, 81–95. [Google Scholar] [CrossRef] [PubMed]
- Polgár, B.; Nagy, E.; Mikó, É.; Varga, P.; Szekeres-Barthó, J. Urinary progesterone-induced blocking factor concentration is related to pregnancy outcome. Biol. Reprod. 2004, 71, 1699–1705. [Google Scholar] [CrossRef] [PubMed]
- Szekeres-Bartho, J.; Polgar, B. PIBF: the double edged sword. Pregnancy and tumor. Am. J. Reprod. Immunol. 2010, 64, 77–86. [Google Scholar] [CrossRef] [PubMed]
- DeMayo, F.J.; Lydon, J.P. 90 YEARS OF PROGESTERONE: New insights into progesterone receptor signaling in the endometrium required for embryo implantation. J. Mol. Endocrinol. 2020, 65, T1–T14. [Google Scholar] [CrossRef] [PubMed]
- Young, S.L. Oestrogen and progesterone action on endometrium: a translational approach to understanding endometrial receptivity. Reprod. Biomed. Online 2013, 27, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Apostolov, A.; Mladenović, D.; Tilk, K.; Lõhmus, A.; Baev, V.; Yahubyan, G.; Sola-Leyva, A.; Bergamelli, M.; Görgens, A.; Zhao, C. Multi-omics analysis of uterine fluid extracellular vesicles reveals a resemblance with endometrial tissue across the menstrual cycle: biological and translational insights. Hum. Reprod. Open 2025, 2025, hoaf010. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez Fernandez, J.; Moncayo Arlandi, J.; Ochando, A.; Simon, C.; Vilella, F. The role of extracellular vesicles in intercellular communication in human reproduction. Clin. Sci. 2023, 137, 281–301. [Google Scholar] [CrossRef] [PubMed]
- Machtinger, R.; Baccarelli, A.A.; Wu, H. Extracellular vesicles and female reproduction. J. Assist. Reprod. Genet. 2021, 38, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Zheng, H.; Xiong, Y.; Li, K. Extracellular vesicles affecting embryo development in vitro: a potential culture medium supplement. Front. Pharmacol. 2024, 15, 1366992. [Google Scholar] [CrossRef] [PubMed]
- Pallinger, E.; Bognar, Z.; Bodis, J.; Csabai, T.; Farkas, N.; Godony, K.; Varnagy, A.; Buzas, E.; Szekeres-Bartho, J. A simple and rapid flow cytometry-based assay to identify a competent embryo prior to embryo transfer. Sci. Rep. 2017, 7, 39927. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Guo, A.; Yang, F.; Li, L.; Yan, J.; Deng, X.; Dai, C.; Li, Y. Alterations of Cytokine Profiles in Patients With Recurrent Implantation Failure. Front. Endocrinol. 2022, 13–2022. [Google Scholar] [CrossRef]
- Pantos, K.; Grigoriadis, S.; Maziotis, E.; Pistola, K.; Xystra, P.; Pantou, A.; Kokkali, G.; Pappas, A.; Lambropoulou, M.; Sfakianoudis, K.; et al. The Role of Interleukins in Recurrent Implantation Failure: A Comprehensive Review of the Literature. Int. J. Mol. Sci. 2022, 23, 2198. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Ashary, N.; Sharma, R.; Modi, D. Extracellular vesicles in embryo implantation and disorders of the endometrium. Am. J. Reprod. Immunol. 2021, 85, e13360. [Google Scholar] [CrossRef] [PubMed]
- Aghajanova, L.; Hamilton, A.; Giudice, L. Uterine receptivity to human embryonic implantation: histology, biomarkers, and transcriptomics. Proceedings of the Seminars in cell & developmental biology 2008, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Mackens, S.; Santos-Ribeiro, S.; van de Vijver, A.; Racca, A.; Van Landuyt, L.; Tournaye, H.; Blockeel, C. Frozen embryo transfer: a review on the optimal endometrial preparation and timing. Hum. Reprod. 2017, 32, 2234–2242. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.; Rai, A.; Nguyen, H.P.T.; Poh, Q.H.; Elglass, K.; Simpson, R.J.; Salamonsen, L.A.; Greening, D.W. Human Endometrial Extracellular Vesicles Functionally Prepare Human Trophectoderm Model for Implantation: Understanding Bidirectional Maternal-Embryo Communication. PROTEOMICS 2019, 19, 1800423. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Dubey, P.; Sunda, U.; Hard, T. A Comprehensive Review of the Endometrial Receptivity Array in Euploid Embryo Transfer Cycles. Cureus 2024, 16. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Cao, Y.; Zhou, M.; Han, M.; Liu, M.; Hu, Y.; Xu, B.; Zhang, A. Decreased CD44v3 expression impairs endometrial stromal cell proliferation and decidualization in women with recurrent implantation failure. Reprod. Biol. Endocrinol. 2022, 20, 170. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Diao, Z.; Sun, H.-X.; Yan, G.-J.; Wang, Z.; Li, R.-T.; Dai, Y.; Wang, J.; Li, J.; Ding, H.; et al. Up-regulation of CD81 inhibits cytotrophoblast invasion and mediates maternal endothelial cell dysfunction in preeclampsia. Proc. Natl. Acad. Sci. 2017, 114, 1940–1945. [Google Scholar] [CrossRef] [PubMed]
- SERRU, V.; François, L.N.; BILLARD, M.; AZORSA, D.O.; LANZA, F.; BOUCHEIX, C.; RUBINSTEIN, E. Selective tetraspan–integrin complexes (CD81/α4β1, CD151/α3β1, CD151/α6β1) under conditions disrupting tetraspan interactions. Biochem. J. 1999, 340, 103–111. [Google Scholar] [CrossRef]
- Jankovičová, J.; Sečová, P.; Michalková, K.; Antalíková, J. Tetraspanins, More than Markers of Extracellular Vesicles in Reproduction. Int. J. Mol. Sci. 2020, 21, 7568. [Google Scholar] [CrossRef] [PubMed]
- von Grothusen, C.; Frisendahl, C.; Modhukur, V.; Lalitkumar, P.G.; Peters, M.; Faridani, O.R.; Salumets, A.; Boggavarapu, N.R.; Gemzell-Danielsson, K. Uterine fluid microRNAs are dysregulated in women with recurrent implantation failure. Hum. Reprod. 2022, 37, 734–746. [Google Scholar] [CrossRef] [PubMed]
- Meseguer, M.; Pellicer, A.; Simón, C. MUC1 and endometrial receptivity. Mol. Hum. Reprod. 1998, 4, 1089–1098. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Chen, X.; Liu, Y.; Liang, B.; Xu, H.; Li, T.C.; Wang, C.C. Decreased MUC1 in endometrium is an independent receptivity marker in recurrent implantation failure during implantation window. Reprod. Biol. Endocrinol. 2018, 16, 60. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Progesterone receptor expression in decidua. (A) Representative confocal images (200x magnification) of progesterone receptor (Pr) expression levels at 2.5, 4.5, and 5.5 days post coitum in mouse uterine sections. Progesterone receptor expression is shown using immunofluorescence (green), and cell nuclei are stained with Hoechst 33342 (cyan). White arrows indicate the luminal epithelium, and red arrows indicate the endometrial stroma. The UC labels the uterine cavity. After implantation at 5.5 days post coitum (dpc), the yellow arrow marks the implanted embryo and the blue arrow designates the primary decidual zone. (B) Mosaic confocal images of mouse uterine cross-sections at 2.5 and 4.5 days post coitum. We used the “mosaic function” to combine the individual images into one large composite image. The images were acquired at 40× magnification and stitched together to reconstruct the complete uterine cross sections. (C) The difference in Pr levels of the 2.5 and 4.5 dpc aged endometrium stroma, as well as the prime decidual zone of the 5.5 day old pregnant uterine horn. The Pr expression was determined by comparing pixel intensities. The asterisk indicates statistical significance. (p=0.0003, Student’s T-test).
Figure 1.
Progesterone receptor expression in decidua. (A) Representative confocal images (200x magnification) of progesterone receptor (Pr) expression levels at 2.5, 4.5, and 5.5 days post coitum in mouse uterine sections. Progesterone receptor expression is shown using immunofluorescence (green), and cell nuclei are stained with Hoechst 33342 (cyan). White arrows indicate the luminal epithelium, and red arrows indicate the endometrial stroma. The UC labels the uterine cavity. After implantation at 5.5 days post coitum (dpc), the yellow arrow marks the implanted embryo and the blue arrow designates the primary decidual zone. (B) Mosaic confocal images of mouse uterine cross-sections at 2.5 and 4.5 days post coitum. We used the “mosaic function” to combine the individual images into one large composite image. The images were acquired at 40× magnification and stitched together to reconstruct the complete uterine cross sections. (C) The difference in Pr levels of the 2.5 and 4.5 dpc aged endometrium stroma, as well as the prime decidual zone of the 5.5 day old pregnant uterine horn. The Pr expression was determined by comparing pixel intensities. The asterisk indicates statistical significance. (p=0.0003, Student’s T-test).

Figure 2.
Inflammatory mediators in the uterine lavage. (A) Cytokine and chemokine levels at 2.5 and 4.5 dpc. Significant differences were detected in the concentrations of monocyte chemoattractant protein-1 (MCP-1/CCL2) (** p= 0.004), interferon gamma (IFN-γ) (*** p<0.0001), and interleukin 6 (IL-6) (* p=0.04). The concentration of IL-10 was below the detection threshold in both groups (n=20). (B) A correlation matrix heatmap of the Pearson correlation coefficients for all soluble mediators studied in uterine lavage. Positive values are shown in blue and negative values in red (n=20).
Figure 2.
Inflammatory mediators in the uterine lavage. (A) Cytokine and chemokine levels at 2.5 and 4.5 dpc. Significant differences were detected in the concentrations of monocyte chemoattractant protein-1 (MCP-1/CCL2) (** p= 0.004), interferon gamma (IFN-γ) (*** p<0.0001), and interleukin 6 (IL-6) (* p=0.04). The concentration of IL-10 was below the detection threshold in both groups (n=20). (B) A correlation matrix heatmap of the Pearson correlation coefficients for all soluble mediators studied in uterine lavage. Positive values are shown in blue and negative values in red (n=20).

Figure 3.
Characterization of the surface epitopes of uterine lavage EVs by using the MACSPlex kit. The violin plots show the normalized mean fluorescence intensity (MFI) for the 11 markersthat showed significant changes at 2.5 and 4.5 dpc (mean ± SD) (* p<0,05; ** p=0,007,Student’s T-test) (n=12).
Figure 3.
Characterization of the surface epitopes of uterine lavage EVs by using the MACSPlex kit. The violin plots show the normalized mean fluorescence intensity (MFI) for the 11 markersthat showed significant changes at 2.5 and 4.5 dpc (mean ± SD) (* p<0,05; ** p=0,007,Student’s T-test) (n=12).

Figure 4.
A comprehensive analysis of soluble cytokines, chemokines, and extracellular vesicles in uterine lavage fluid. We used Cytoscape (version 3.7.2), an open-source bioinformatics software platform, to visualize protein interaction networks. The hubs represent proteins that changed significantly in uterine lavage. The colored circular arcs around the hubs represent Gene Ontology (GO) pathways. Light blue: GO:0007155 (cell adhesion); dark blue: GO:0098609 (cell-cell adhesion); light green: GO:0030155 (regulation of cell adhesion); dark green: GO:0022407 (regulation of cell-cell adhesion); pink: GO:0022407 (regulation of cell-to-cell adhesion); pink: GO:0045785 (positive regulation of cell adhesion). The gray lines in the figure illustrate the interconnection between the analytes, indicating this process. The thickness of the lines indicates the confidence of a real biological interaction in the Protein-Protein Interaction (PPI) network.
Figure 4.
A comprehensive analysis of soluble cytokines, chemokines, and extracellular vesicles in uterine lavage fluid. We used Cytoscape (version 3.7.2), an open-source bioinformatics software platform, to visualize protein interaction networks. The hubs represent proteins that changed significantly in uterine lavage. The colored circular arcs around the hubs represent Gene Ontology (GO) pathways. Light blue: GO:0007155 (cell adhesion); dark blue: GO:0098609 (cell-cell adhesion); light green: GO:0030155 (regulation of cell adhesion); dark green: GO:0022407 (regulation of cell-cell adhesion); pink: GO:0022407 (regulation of cell-to-cell adhesion); pink: GO:0045785 (positive regulation of cell adhesion). The gray lines in the figure illustrate the interconnection between the analytes, indicating this process. The thickness of the lines indicates the confidence of a real biological interaction in the Protein-Protein Interaction (PPI) network.

Figure 5.
Relative gene expression in the endometrium. Results of qRT-PCR analysis of 11 differentially expressed genes in the mouse endometrium. The violin plots illustrate the relative expression levels at 2.5 and 4.5 dpc (n=12; *p<0,05; **p=0.001, Student’s T-test).
Figure 5.
Relative gene expression in the endometrium. Results of qRT-PCR analysis of 11 differentially expressed genes in the mouse endometrium. The violin plots illustrate the relative expression levels at 2.5 and 4.5 dpc (n=12; *p<0,05; **p=0.001, Student’s T-test).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



