Submitted:
16 June 2026
Posted:
17 June 2026
You are already at the latest version
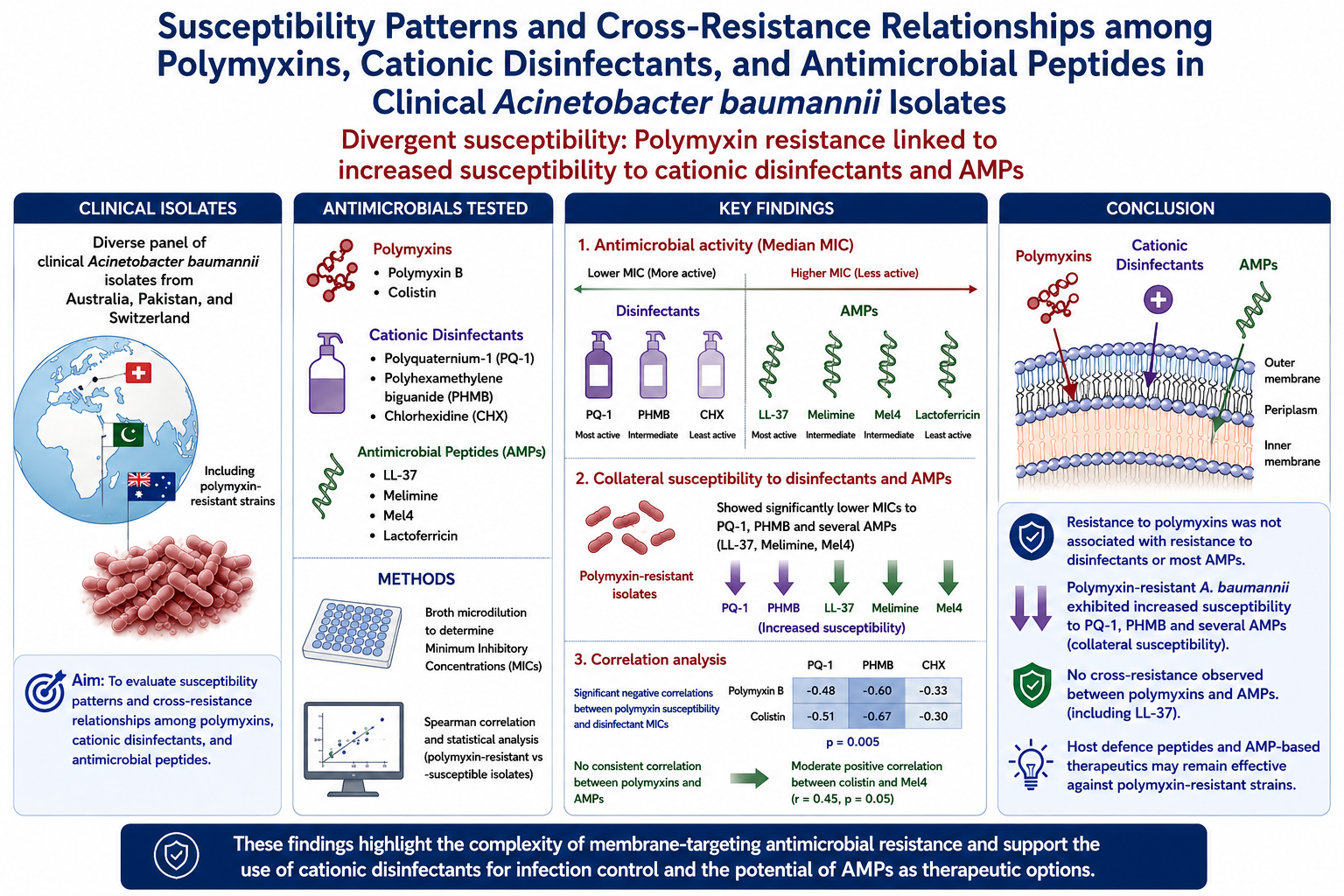
Abstract
Background: Acinetobacter baumannii is a critical priority pathogen characterized by extensive multidrug resistance, leading to increased reliance on last-line polymyxins and widespread use of cationic disinfectants in healthcare settings. However, shared membrane-targeting mechanisms raise concerns regarding potential cross-resistance between disinfectants, polymyxins, and cationic antimicrobial peptides. Methods: A diverse panel of clinical A. baumannii isolates from Australia, Pakistan, and Switzerland, including polymyxin-resistant strains, was used to evaluate their antimicrobial resistance profiles. Minimum inhibitory concentrations (MICs) of polymyxin B, colistin, three cationic disinfectants (polyquaternium-1; PQ-1), polyhexamethylene biguanide (PHMB), and chlorhexidine (CHX)), and four AMPs (LL-37, melimine, Mel4, and lactoferricin) were determined using broth microdilution. Correlations between MICs were assessed using Spearman analysis, and differences between polymyxin-resistant and susceptible isolates were analysed statistically. Results: Most isolates were susceptible to polymyxins, although selected strains (12.9%) displayed high-level resistance. PQ-1 exhibited the greatest antibacterial activity among disinfectants, while LL-37 showed the lowest and most consistent MICs among AMPs; lactoferricin was largely inactive. Polymyxin-resistant isolates demonstrated significantly lower MICs to PQ-1 and PHMB, indicating collateral susceptibility. Correlation analysis revealed significant negative correlations between polymyxin susceptibility and disinfectant MICs (r=-0.48 to -11 and p= 0.005), whereas no consistent correlation was observed between polymyxins and AMPs, including LL-37. A moderate positive correlation was identified between colistin and Mel4 activity (r =0.45 and p =0.05). Conclusions: These findings demonstrate divergent susceptibility patterns of A. baumannii to cationic antimicrobials. Resistance to polymyxins was not associated with resistance to disinfectants or most AMPs and instead was linked to increased susceptibility to certain disinfectants. Importantly, the absence of cross-resistance with LL-37 suggests that host defense peptides and AMP-based therapeutics may remain effective against polymyxin-resistant strains. This study highlights the complexity of membrane-targeting antimicrobial resistance and underscores the need for further mechanistic investigations into cross-resistance and collateral susceptibility.
Keywords:
cationic antimicrobials
; clinical isolates
; Acinetobacter baumannii
; polymyxin
; resistance
1. Introduction
The Acinetobacter genus is diverse, comprising oxidase-positive and -negative, nonpigmented, Gram-negative coccobacilli. With over 50 species identified [1] most are environmental and non-pathogenic. The species most frequently linked to infections are A. baumannii, followed by A. calcoaceticus and A. lwoffii [2], then other species including A. haemolyticus, A. johnsonii, A. junii, and A. ursingii [3,4,5,6,7,8,9]. A. baumannii is the most virulent species among them [10].
Infections caused by Acinetobacter spp. became more widespread in the 1960s and 1970s, coinciding with the growing use of intensive care techniques [11,12], mechanical ventilation, central venous and urinary catheterization, and associated antibacterial therapies [13,14,15,16]. Analysis of data from 2018-2020 found Acinetobacter spp. accounted for 1.2% of all healthcare-associated infections in USA [17]. There is an estimated 1 million cases globally each year [18]. Similar infection rates are observed in ICUs across Europe and Latin America [19,20,21,22,23]. In some Asian and Latin American countries, Acinetobacter ranks among the top three causes of bacteraemia and nosocomial pneumonia [24,25,26,27,28].
Acinetobacter baumannii is ranked as a top priority pathogen of critical importance by the World Health Organisation for which new antimicrobials are urgently required [29]. It has an increasing resistance to all traditional antibiotics, with up to 90% of strains being resistant to cephalosporins, carbapenem, fluoroquinolones, and β-lactams [30]. Furthermore, multidrug resistance has been reported in up to 99.8% of carbapenem-resistant A. baumannii [31,32]. Infection with carbapenem-resistant strains results in higher in-hospital mortality [17]. Polymyxins are becoming more widely used to treat Acinetobacter infections due to this increase in resistance to traditional antibiotics. However, resistance to polymyxins is also increasing, [33,34] with 21% of strains being resistant to colistin in one study [30]. Acinetobacter baumannii can become resistant to polymyxins via a variety of mechanisms including addition of phosphoethanolamine to its outer membrane, loss of lipopolysaccharide from its outer membrane, loss of outer membrane proteins such as OmpA, and expression of efflux pumps [35,36].
Due to the potential for cross contamination, disinfection of hospital surfaces and appliances is common. Cationic disinfectants such as chlorhexidine (CHX), polyhexamethylene biguanide (PHMB) and polyquaternium-1 (PQ-1) are widely used for disinfection purposes in health care settings [37,38,39]. However, excessive or improper use of disinfectants can contribute to bacterial resistance and promote cross-resistance to antibiotics, thereby worsening the global antimicrobial resistance crisis [40]. Common disinfectants such as CHX may trigger adaptive bacterial mechanisms such as efflux pump activation, biofilm formation, membrane alterations, stress-response pathways, and horizontal gene transfer [41]. These adaptations can increase bacterial tolerance not only to disinfectants but also to multiple antibiotics, leading to multidrug-resistant pathogens [41]. There is the potential for resistance to cationic disinfectants to affect the susceptibility to polymyxins [42]. Cationic antimicrobial peptides are possible alternative new antibiotics or disinfecting agents that are being developed. They typically consist of short chains of amino acids and their net positive charge at physiological pH is usually due to the presence of lysine or arginine [43]. This positive charge is critical for their interaction with microbial cell membranes, which are often negatively charged [44]. The naturally occurring human cathelicidin LL-37 and the chimeric AMP melimine have been studied for their antimicrobial activity [45,46,47]. Use of LL-37 can improve polymicrobial wound infection in clinical trials [48,49], and a modified version of melimine, Mel4, successfully completed a Phase III clinical trial as an antimicrobial contact lens coating [50]. The changes that occur in membranes to develop polymyxins and cationic disinfectants, can also affect sensitivity to antimicrobial peptides [51], although this is not always the case [52,53].
This research was designed to determine the susceptibility pattern of clinical isolates of Acinetobacter baumannii against disinfectants, polymyxins and cationic antimicrobial peptides, and to evaluate whether resistance to disinfectants correlated to resistance to polymyxins, and whether resistance to polymyxins was correlated with resistance to antimicrobial peptides. The hypotheses being tested were (1) resistance to disinfectants would correlate with resistance to polymyxin antibiotics, and (2) resistance to polymyxin antibiotics would correlate with resistance to naturally occurring and synthetic antimicrobial peptides.
2. Results
2.1. Conformation of Strains Identity
All strains were confirmed as Acinetobacter baumannii (Table 1). Apart from strains ATCC 19606, ATCC 19606 (R), Ab08, Ab39 and Ab61, other strains were not selected on the basis of their resistance to polymyxins or any of the other antimicrobials being studied.
2.2. Minimum Inhibitory Concentration of Cationic Antimicrobials
The results demonstrate diversified patterns of susceptibility toward different tested cationic antimicrobials. Except four strains, (ATCC 19606R, Ab08, Ab39 and Ab 61) all strains were susceptible to polymyxin B (1-2 μg/ml) and colistin (0.25-2 μg/ml). The MICs to polymyxin B for the resistant strains ranged from 16 μg/ml for Ab08 to 1250 μg/ml for ATCC 19606 (R) (Table 2). For the disinfectants, PQ-1 was the most effective with a median MIC of 18.75 μg/ml, ranging from 6.3–50 μg/ml (Table 2). The median MICs for PHMB and CHX were the same, at 25 μg/ml, although there was a slightly greater range for PHMB of 3.13 to 50 μg/ml (Table 2).
For the antimicrobial peptides, LL-37 gave the most consistently low MICs, with a median of 62.5 μg/ml, ranging from 31.3 to 125 μg/ml (Table 2). This was followed by Mel4 and melimine, with median MICs of 125 μg/ml, but with a slightly greater range from 15.6 to 250 μg/ml for Mel4 (Table 2). Lactoferricin was virtually inactive against the strains tested, with MICs >500 μg/ml (Table 2).
Overall, Australian and Pakistani isolates had very similar median MICs to each antimicrobial, with the exception of Mel4, where the median MIC for Pakistani isolates was 250 μg/ml compared to 62.5 μg/ml for Australian isolates.
When the MICs to Mel4, Melimine, LL-37, PQ-1, PMHB, CHX of the polymyxin resistance isolates (ATCC 19606 (R), Ab08, Ab39, Ab61) were compared with the polymyxin susceptible strains, there were markedly lower MICs to PQ-1 (median 6.3 vs. 25; p = 0.01) and PHMB (median 3.1 vs. 25; p = 0.003), with a trend for the same difference occurring with CHX (15.6 vs. 25; p = 0.08) suggesting collateral sensitivity to membrane-active antiseptic agents. In contrast, susceptibility to peptide antimicrobials (Mel4, Melimine, LL-37) showed no consistent directional change.
2.3. Correlation Analysis of MICs
Within the antimicrobial groups, polymyxin B and colistin showed strong positive correlation (r = 0.51; p = 0.0041) (Figure 1). Mel4 and colistin demonstrated the next strongest correlation (r = 0.45; p = 0.0127), followed by Mel4 and melimine (r = 0.41; p = 0.0258). On the other hand, colistin was significantly negatively correlated with PQ-1 (r = -0.67; 0.0001), and PHMB (r = -0.44; p = 0.0151). Similarly, susceptibility was significantly negatively correlated with PHMB susceptibility (r = -0.48; 0.0074). In contrast, LL-37 susceptibility was not significantly correlated with any of the other antimicrobials tested.
3. Discussion
The present study was designed to evaluate the pattern of susceptibility among Acinetobacter strains against different cationic antimicrobials and to evaluate whether high MICs to disinfectants correlated to resistance to polymyxins or MICs to cationic antimicrobial peptides. The impetus for this research was the use of disinfectants to control spread of Acinetobacter and the increasing isolation of polymyxin resistant strains. The hypothesis was that resistance to disinfectants might cross-react to resistance to polymyxins, as these can act in similar ways on the membrane of bacteria. Furthermore, mechanisms of resistance to disinfectants such as efflux pump activation and membrane alterations [41] are also used be Acinetobacter to become resistant to polymyxins [33].
The results of this study show that the hypothesis was not proven, and resistance to polymyxin in A. baumannii negatively correlated with resistance to disinfectants (especially PQ-1 and PHMB). Whilst the antibacterial mechanism of action of PHMB has been traditionally thought to be via membrane disruption, recent studies have shown that whilst PHMB can attach to Gram-negative bacterial membranes, this does not result in pore formation (often a key mechanism for antibacterial activity) but it then penetrates through the membranes and interacts with intracellular DNA, which is likely the antibacterial mode of action [55,56]. Similarly, the quaternary amine compound benzalkonium chloride can have two modes of action against A. baumannii [57]. At high concentrations benzalkonium chloride acts on the bacterial membrane, but at lower concentrations it can induce protein aggregation and this can lead to cell death [57]. This may also occur for polyquats such as PQ-1. The polymyxin antibiotics appear to have predominant modes of action on the outer membrane of A. baumannii and other gram-negative bacteria, although they may also act intercellularly by inhibiting respiratory enzymes and triggering production of reactive oxygen species [58,59]. These differences may at least partially account for the lack of correlation between the MICs for the polymyxins and disinfectants. Reinforcing this, the polymyxin resistant mutant A. baumannii ATCC (R)19606 with complete loss of outer membrane [54] with MIC 1250 μg/ml and >2500 μg/ml against polymyxin B and colistin, respectively, showed high sensitively to the disinfectants. A previous study found that loss of LPS caused a 4 fold reduction in MIC of CHX [60].
This study also evaluated whether high MICs to the disinfectants or resistance to polymyxins resulted in high MICs to antimicrobial peptides [61]. The AMPs chosen for study were two naturally occurring AMPs, LL-37 (the only human cathelicidin) and lactoferricin (a small peptide from lactoferrin), as well as the synthetic AMPs melimine and Mel4. Should there have been a correlation, it would mean that resistance to polymyxins might have large impacts on the pathogenesis of disease caused by A. baumannii. Epithelial cells upregulate AMPs during A. baumannii infection [62] and AMPs such as LL-37 are key components of the innate defence system [63]. Furthermore, the AMPs melimine and Mel4 are being examined as potential new therapeutic or prophylactic agents [64,65,66], and so cross-resistance might curtail this research and development. Previous studies have determined that polymyxin resistance in A. baumannii is mediated by several factors including phosphoethanolamine addition to lipopolysaccharides in the outer membrane, loss of lipopolysaccharide from the outer membrane, and overexpression of efflux pumps [33,35,54]. These same mechanisms can confer resistance to antimicrobial peptides such as LL-37 [45]. Indeed, clinical use of polymyxin can induce cross-resistance to host antimicrobial peptides in A. baumannii [42].
However, the current study found no statistically significance evidence for cross-resistance between the naturally occurring LL-37 and polymyxins. This implies a lack of effect of polymyxin resistance, especially loss of the outer membrane which is the known mechanism of resistance of strain ATCC 19600 (R), on susceptibility to LL-37. There was a significant correlation between colistin resistance and MIC to Mel4, implying the possibility of similar mechanisms being involved. Although, this correlation did not hold for polymyxin B, which is structurally similar to colistin. Further research is needed to identify if any known polymyxin resistance mechanisms can confer cross-resistance to Mel4, and why these do not correlate with resistance to polymyxin B or MICs to melimine (the parent peptide of Mel4). The correlation between MIC to Mel4 and PQ-1 is intriguing, suggesting some cross-resistance, and this should also be followed up in future experiments. These future experiments could mutate susceptible strains with genes known to be involved polymyxin, disinfectant and antimicrobial peptide and determine which ones confer cross resistance.
4. Materials and Methods
4.1. Collection and Preservation of Bacterial Strains
Clinical isolates of Acinetobacter were collected from Australia (Liping Li, Macquarie University, NSW, Australia) Pakistan (Bushra Jamil, Sir Syed CASE University, Islamabad, Pakistan) and Switzerland (Stefano Mancini, Institute of Medical Microbiology, University Zurich, Switzerland). This study included the polymyxin-resistant mutant A. baumannii ATCC 19606R, which has a complete loss of its outer membrane due to mutation in lpx genes [54]. All the strains were preserved in 25% glycerol and stored at −80 °C at the microbial culture collection of School of Optometry and Vision Science, Faculty of Medicine and Health, UNSW Sydney.
4.2. Confirmation of Strain Identities by MALDI-TOF MS
Identity of strains was confirmed by MALDI-TOF MS (Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry). Briefly, strains were cultured on blood agar overnight and protein extraction was done according to MALDI Biotyper® Protocol Guideline [67,68] with 300μl of sterile MilliQ water. After air-drying the pellet at room temperature, 25 μL of 70% aqueous formic acid was added and mixed, followed by addition of 25 μL 100% acetonitrile and mixing. The mixture was then centrifuged for 2 minutes at 13,000 rpm and 1 μL of the supernatant was deposited onto a MALDI target plate and allowed to dry at room temperature. A 1μl aliquot of Bruker Bacterial Test Standard (Bruker, Billerica, MA, USA) was transferred to a MALDI target plate and allowed to dry. After drying. 1 μl of HCCA matrix solution was added to each sample and standard. Identification was done by using Bruker ultrafleXtreme MALDI-TOF/TOF biotyper system.
4.3. Determination of Minimum Inhibitory Concentrations of the Antimicrobials
Two cationic polypeptide antibiotics, polymyxin B (PMB) (Fluka Chemie GmbH, Buchs, Switzerland) and colistin sulfate (Polymyxin E; Sigma-Aldrich, Massachusetts, United States), were used. Four cationic antimicrobial peptides (LL-37, Mel4, melimine, and lactoferricin) were synthesised by solid state synthesis and purchased from AusPep Peptide Company (Tullamarine, VIC, Australia) with a purity of ≥90%. The cationic disinfectants PQ-1 (Sigma Aldrich), PHMB (Biosynth, Gardner, MA USA) and CHX (Sigma Aldrich) were also used.
Minimum inhibitory concentration (MIC) of the cationic antimicrobials was determined by broth micro-dilution assay [69]. Standard solutions of 128μg/mL for polymyxin B, colistin, PQ-1, PHMB and CHX were made sterile distilled water. For the AMPs, standard solutions of 5mg/mL were made in sterile distilled water. Cation adjusted Muller Hinton broth (Ca-MHB; Oxoid, Basingstoke, Hampshire, UK) was used as described previously [70]. Briefly, 100μl of each antimicrobial was diluted two fold in 100μl Ca-MHB, then serially diluted in Ca-MHB. A standard bacterial inoculum was prepared from overnight culture by adjusting to 0.5 McFarland standard (~1.5 × 108 colony forming units (CFU)/mL) followed by further dilution to achieve a final concentration of approximately 1.5 × 106 CFU/mL and 100μl of bacterial cells were added to each well and plates were incubated for 24 hours and MIC was determined as inhibition of visible growth.
4.4. Statistical Analysis
Correlation analysis among MICs of different cationic antimicrobials was determined by GraphPad Prism software (version 11.02.2. GraphPad Software, San Diego, CA, USA) by determining Spearman correlation coefficient from MIC data. Mann-Whitney U test was performed to determine whether differences in the median MICs of polymyxin resistant and susceptible strains to Mel4, Melimine, LL-37, PQ-1, PMHB, CHX were statistically different.
5. Conclusions
Acinetobacter baumannii strains demonstrate diverse patterns of susceptibility to cationic antimicrobial peptides and disinfectant. One hypothesis being tested was to determine whether high MICs to disinfectants that can be used to clean surfaces to reduce the spread of Acinetobacter spp. were correlated to resistance (high MICs) to polymyxin antibiotics which are antibiotics of last resort to treat Acinetobacter infections. The study found an inverse correlation, implying that disinfectant resistance does not result in resistance to polymyxins. The second hypothesis being tested was whether resistance to polymyxins was correlated to resistance to antimicrobial peptides involved in defence against Acinetobacter or being developed as new therapeutics or prophylactic agents. The lack of correlation between MIC for LL-37 and polymyxins implies no cross resistance.
This study has important translational implications for both infection control and therapeutic development. The finding that polymyxin-resistant A. baumannii remains susceptible, or even shows collateral sensitivity, to commonly used disinfectants suggests that current hospital disinfection strategies are unlikely to drive resistance to last-line antibiotics, and may in fact help control resistant strains. Furthermore, the absence of cross-resistance between polymyxins and key antimicrobial peptides such as LL-37 highlights the potential for host defence–based or AMP-derived therapies to remain effective against highly drug-resistant A. baumannii, supporting their ongoing clinical development as alternative treatments.
Author Contributions
Conceptualization, S.S. and M.D.P.W.; methodology, S.S., M.D.P.W.; formal analysis, S.S.; resources, M.D.P.W. and B.J.; writing - original draft preparation, S.S.; writing - review and editing, M.D.P.W. and B.R.; visualization, S.S. and M.D.P.W.; supervision, M.D.P.W. and M.Y.; project administration, M.D.P.W.; funding acquisition, M.D.P.W. All authors have read and agreed to the published version of this manuscript.
Funding
No additional funding was obtained for this work.
Institutional Review Board Statement
Not Applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The first author has received UIPA from UNSW, Sydney.
Competing Interests
The authors declare no conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| Lfcin | Lactoferricin |
| PQ-1 | polyquaternium-1 |
| PHMB | polyhexamethylene biguanide |
| CHX | chlorhexidine |
References
- Al Atrouni, A.; Joly-Guillou, M.L.; Hamze, M.; Kempf, M. Reservoirs of Non-baumannii Acinetobacter Species. Front. Microbiol. 2016, 7, 49. [Google Scholar] [CrossRef] [PubMed]
- Dijkshoorn, L.; van der Toorn, J. Acinetobacter species: which do we mean? Clin. Infect. Dis. 1992, 15, 748–749. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.N.; Lu, C.H.; Huang, C.R.; Chuang, Y.C. Community-acquired Acinetobacter meningitis in adults. Infection 2000, 28, 395–397. [Google Scholar] [CrossRef] [PubMed]
- Visca, P.; Seifert, H.; Towner, K.J. Acinetobacter infection--an emerging threat to human health. IUBMB Life 2011, 63, 1048–1054. [Google Scholar] [CrossRef] [PubMed]
- Dortet, L.; Legrand, P.; Soussy, C.J.; Cattoir, V. Bacterial identification, clinical significance, and antimicrobial susceptibilities of Acinetobacter ursingii and Acinetobacter schindleri, two frequently misidentified opportunistic pathogens. J. Clin. Microbiol. 2006, 44, 4471–4478. [Google Scholar] [CrossRef] [PubMed]
- Loubinoux, J.; Mihaila-Amrouche, L.; Le Fleche, A.; Pigne, E.; Huchon, G.; Grimont, P.A.; Bouvet, A. Bacteremia caused by Acinetobacter ursingii. J. Clin. Microbiol. 2003, 41, 1337–1338. [Google Scholar] [CrossRef] [PubMed]
- Endo, S.; Sasano, M.; Yano, H.; Inomata, S.; Ishibashi, N.; Aoyagi, T.; Hatta, M.; Gu, Y.; Yamada, M.; Tokuda, K.; et al. IMP-1-producing carbapenem-resistant Acinetobacter ursingii from Japan. J. Antimicrob. Chemother. 2012, 67, 2533–2534. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.H.; Lee, Y.T.; Wang, Y.C.; Yin, T.; Kuo, S.C.; Yang, Y.S.; Chen, T.L.; Lin, J.C.; Wang, F.D.; Fung, C.P. A retrospective study of the incidence, clinical characteristics, identification, and antimicrobial susceptibility of bacteremic isolates of Acinetobacter ursingii. BMC Infect. Dis. 2015, 15, 400. [Google Scholar] [CrossRef] [PubMed]
- Salzer, H.J.; Rolling, T.; Schmiedel, S.; Klupp, E.M.; Lange, C.; Seifert, H. Severe Community-Acquired Bloodstream Infection with Acinetobacter ursingii in Person who Injects Drugs. Emerg. Infect. Dis. 2016, 22, 134–137. [Google Scholar] [CrossRef] [PubMed]
- Chusri, S.; Chongsuvivatwong, V.; Rivera, J.I.; Silpapojakul, K.; Singkhamanan, K.; McNeil, E.; Doi, Y. Clinical outcomes of hospital-acquired infection with Acinetobacter nosocomialis and Acinetobacter pittii. Antimicrob. Agents Chemother. 2014, 58, 4172–4179. [Google Scholar] [CrossRef] [PubMed]
- Glew, R.H.; Moellering, R.C., Jr.; Kunz, L.J. Infections with Acinetobacter calcoaceticus (Herellea vaginicola): clinical and laboratory studies. Medicine (Balt.) . 1977, 56, 79–97. [Google Scholar] [CrossRef] [PubMed]
- Daly, A.K.; Postic, B.; Kass, E.H. Infections due to organisms of the genus Herellea. B5W and B. anitratum. Arch. Intern. Med. 1962, 110, 580–591. [Google Scholar] [CrossRef] [PubMed]
- Spellberg, B.; Bonomo, R.A. The deadly impact of extreme drug resistance in Acinetobacter baumannii. Crit. Care Med. 2014, 42, 1289–1291. [Google Scholar] [CrossRef] [PubMed]
- Maragakis, L.L.; Perl, T.M. Acinetobacter baumannii: epidemiology, antimicrobial resistance, and treatment options. Clin. Infect. Dis. 2008, 46, 1254–1263. [Google Scholar] [CrossRef] [PubMed]
- Joly-Guillou, M.L. Clinical impact and pathogenicity of Acinetobacter. Clin. Microbiol. Infect. 2005, 11, 868–873. [Google Scholar] [CrossRef] [PubMed]
- Sievert, D.M.; Ricks, P.; Edwards, J.R.; Schneider, A.; Patel, J.; Srinivasan, A.; Kallen, A.; Limbago, B.; Fridkin, S.; National Healthcare Safety Network; T.; et al. Antimicrobial-resistant pathogens associated with healthcare-associated infections: summary of data reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2009-2010. Infect. Control. Hosp. Epidemiol. 2013, 34, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lodise, T.P.; Nguyen, S.T.; Margiotta, C.; Cai, B. Clinical burden of Acinetobacter baumannii, including carbapenem-resistant A. baumannii, in hospitalized adult patients in the USA between 2018 and 2022. BMC Infect. Dis. 2025, 25, 549. [Google Scholar] [CrossRef] [PubMed]
- Spellberg, B.; Rex, J.H. The value of single-pathogen antibacterial agents. Nat. Rev. Drug Discov. 2013, 12, 963. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Bano, J.; Cisneros, J.M.; Fernandez-Cuenca, F.; Ribera, A.; Vila, J.; Pascual, A.; Martinez-Martinez, L.; Bou, G.; Pachon, J.; Grupo de Estudio de Infeccion, H. Clinical features and epidemiology of Acinetobacter baumannii colonization and infection in Spanish hospitals. Infect. Control Hosp. Epidemiol. 2004, 25, 819–824. [Google Scholar] [CrossRef] [PubMed]
- Agodi, A.; Auxilia, F.; Barchitta, M.; Brusaferro, S.; D'Alessandro, D.; Montagna, M.T.; Orsi, G.B.; Pasquarella, C.; Torregrossa, V.; Suetens, C.; et al. Building a benchmark through active surveillance of intensive care unit-acquired infections: the Italian network SPIN-UTI. J. Hosp. Infect. 2010, 74, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.F.; Kuchenbecker, R.S.; Pilger, K.O.; Pagano, M.; Barth, A.L.; Force, C.-P.S.T. High endemic levels of multidrug-resistant Acinetobacter baumannii among hospitals in southern Brazil. Am. J. Infect. Control. 2012, 40, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Labarca, J.A.; Salles, M.J.; Seas, C.; Guzman-Blanco, M. Carbapenem resistance in Pseudomonas aeruginosa and Acinetobacter baumannii in the nosocomial setting in Latin America. Crit. Rev. Microbiol. 2016, 42, 276–292. [Google Scholar] [CrossRef] [PubMed]
- Villar, M.; Cano, M.E.; Gato, E.; Garnacho-Montero, J.; Miguel Cisneros, J.; Ruiz de Alegria, C.; Fernandez-Cuenca, F.; Martinez-Martinez, L.; Vila, J.; Pascual, A.; et al. Epidemiologic and clinical impact of Acinetobacter baumannii colonization and infection: a reappraisal. Medicine (Balt.) . 2014, 93, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Chung, D.R.; Song, J.H.; Kim, S.H.; Thamlikitkul, V.; Huang, S.G.; Wang, H.; So, T.M.; Yasin, R.M.; Hsueh, P.R.; Carlos, C.C.; et al. High prevalence of multidrug-resistant nonfermenters in hospital-acquired pneumonia in Asia. Am. J. Respir. Crit. Care. Med. 2011, 184, 1409–1417. [Google Scholar] [CrossRef] [PubMed]
- Kuo, S.C.; Chang, S.C.; Wang, H.Y.; Lai, J.F.; Chen, P.C.; Shiau, Y.R.; Huang, I.W.; Lauderdale, T.L.; Hospitals, T. Emergence of extensively drug-resistant Acinetobacter baumannii complex over 10 years: nationwide data from the Taiwan Surveillance of Antimicrobial Resistance (TSAR) program. BMC Infect. Dis. 2012, 12, 200. [Google Scholar] [CrossRef] [PubMed]
- S., D.; S., T.; P., S.; N., A.; K, M. Antimicrobial resistance of Acinetobacter baumannii: six years of National Antimicrobial Resistance Surveillance Thailand (NARST) surveillance. J. Med. Assoc. Thai. 2011, 92, 34. [Google Scholar]
- Madani, N.; Rosenthal, V.D.; Dendane, T.; Abidi, K.; Zeggwagh, A.A.; Abouqal, R. Health-care associated infections rates, length of stay, and bacterial resistance in an intensive care unit of Morocco: findings of the International Nosocomial Infection Control Consortium (INICC). Int. Arch. Med. 2009, 2, 29. [Google Scholar] [CrossRef] [PubMed]
- Luna, C.M.; Rodriguez-Noriega, E.; Bavestrello, L.; Guzman-Blanco, M. Gram-negative infections in adult intensive care units of latin america and the Caribbean. Crit. Care. Res. Pract. 2014, 2014, 480463. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Prioritization of pathogens to guide discovery, research and development of new antibiotics for drug-resistant bacterial infections, including tuberculosis. 2017.
- Gharaibeh, M.H.; Abandeh, Y.M.; Elnasser, Z.A.; Lafi, S.Q.; Obeidat, H.M.; Khanfar, M.A. Multi-drug Resistant Acinetobacter baumannii: Phenotypic and Genotypic Resistance Profiles and the Associated Risk Factors in Teaching Hospital in Jordan. J. Infect. Public. Heal. 2024, 17, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Guo, Y.; Yang, Y.; Zheng, Y.; Wu, S.; Jiang, X.; Zhu, D.; Wang, F.; China Antimicrobial Surveillance Network Study, G. Resistance reported from China antimicrobial surveillance network (CHINET) in 2018. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 2275–2281. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.K.; Yasir, M.; Kuppusamy, R.; Wong, E.H.H.; Hui, A.; Sorensen, K.; Lin, J.S.; Jenssen, H.; Barron, A.E.; Willcox, M. Antimicrobial activity of peptoids against Metallo-beta-lactamase-producing Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and other WHO priority pathogens, including Candida auris. J. Appl. Microbiol. 2025, 136, lxaf031. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, S.; Willcox, M.D.P.; Rayamajhee, B. A Review of Resistance to Polymyxins and Evolving Mobile Colistin Resistance Gene (mcr) among Pathogens of Clinical Significance. Antibiotics 2023, 12, 1597. [Google Scholar] [CrossRef] [PubMed]
- Qin, P.; Ding, W.; Zhang, W. High-level polymyxin B resistance and underlying mechanism in a multidrug-resistant Acinetobacter strain isolated from the marine plastisphere. J. Antibiot. 2026, 79, 122–132. [Google Scholar] [CrossRef]
- Lima, W.G.; Alves, M.C.; Cruz, W.S.; Paiva, M.C. Chromosomally encoded and plasmid-mediated polymyxins resistance in Acinetobacter baumannii: a huge public health threat. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 1009–1019. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.I.; Kim, S.; Oh, M.H.; Na, S.H.; Kim, Y.J.; Jeon, Y.H.; Lee, J.C. Outer membrane protein A contributes to antimicrobial resistance of Acinetobacter baumannii through the OmpA-like domain. J. Antimicrob. Chemother. 2017, 72, 3012–3015. [Google Scholar] [CrossRef] [PubMed]
- Van den Poel, B.; Saegeman, V.; Schuermans, A. Increasing usage of chlorhexidine in health care settings: blessing or curse? A narrative review of the risk of chlorhexidine resistance and the implications for infection prevention and control. Eur. J. Clin. Microbiol. Infect. Dis. 2022, 41, 349–362. [Google Scholar] [CrossRef] [PubMed]
- Hadaway, L. Polyhexamethylene Biguanide Dressing – Another Promising Tool to Reduce Catheter-related Bloodstream Infection. J. Assoc. Vasc. Access. 2011, 15, 203–205. [Google Scholar] [CrossRef]
- de Azevedo Magalhaes, O.; Ribeiro Dos Santos, D.; Coch Broetto, B.G.; Corcao, G. Polyhexamethylene Biguanide Multipurpose Solutions on Bacterial Disinfection: A Comparison Study of Effectiveness in a Developing Country. Eye Contact Lens 2023, 49, 139–142. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, M.; Gholipour, S.; Ghodsi, S.; Nikaeen, M. Co-selection of antibiotic and disinfectant resistance in environmental bacteria: Health implications and mitigation strategies. Environ. Res. 2025, 267, 120708. [Google Scholar] [CrossRef] [PubMed]
- Walczak, Ł.J.; Kwiatkowska, M.; Twarowski, B.; Kubacka, M.; Paluch, J.; Herbet, M. Disinfectant-induced bacterial resistance and antibiotic cross-resistance—mechanisms and clinical relevance. Clin. Exp. Med. 2026, 26, 26. [Google Scholar]
- Napier, B.A.; Burd, E.M.; Satola, S.W.; Cagle, S.M.; Ray, S.M.; McGann, P.; Pohl, J.; Lesho, E.P.; Weiss, D.S. Clinical use of colistin induces cross-resistance to host antimicrobials in Acinetobacter baumannii. mBio 2013, 4, e00021-00013. [Google Scholar] [CrossRef] [PubMed]
- Bulet, P.; Stocklin, R.; Menin, L. Anti-microbial peptides: from invertebrates to vertebrates. Immunol. Rev. 2004, 198, 169–184. [Google Scholar] [CrossRef] [PubMed]
- Brogden, K.A. Antimicrobial peptides: pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Ridyard, K.E.; Overhage, J. The Potential of Human Peptide LL-37 as an Antimicrobial and Anti-Biofilm Agent. Antibiotics 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Sambanthamoorthy, K.; Palys, T.; Paranavitana, C. The human antimicrobial peptide LL-37 and its fragments possess both antimicrobial and antibiofilm activities against multidrug-resistant Acinetobacter baumannii. Peptides 2013, 49, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Rasul, R.; Cole, N.; Balasubramanian, D.; Chen, R.; Kumar, N.; Willcox, M.D. Interaction of the antimicrobial peptide melimine with bacterial membranes. Int. J. Antimicrob. Agents. 2010, 35, 566–572. [Google Scholar] [CrossRef] [PubMed]
- Duplantier, A.J.; van Hoek, M.L. The Human Cathelicidin Antimicrobial Peptide LL-37 as a Potential Treatment for Polymicrobial Infected Wounds. Front. Immunol. 2013, 4, 143. [Google Scholar] [CrossRef] [PubMed]
- Svensson, D.; Nilsson, B.O. Human antimicrobial/host defense peptide LL-37 may prevent the spread of a local infection through multiple mechanisms: an update. Inflamm. Res. 2025, 74, 36. [Google Scholar] [CrossRef] [PubMed]
- Kalaiselvan, P.; Konda, N.; Pampi, N.; Vaddavalli, P.K.; Sharma, S.; Stapleton, F.; Kumar, N.; Willcox, M.D.P.; Dutta, D. Effect of Antimicrobial Contact Lenses on Corneal Infiltrative Events: A Randomized Clinical Trial. Transl. Vis. Sci. Technol. 2021, 10, 32. [Google Scholar] [CrossRef] [PubMed]
- Band, V.I.; Weiss, D.S. Mechanisms of Antimicrobial Peptide Resistance in Gram-Negative Bacteria. Antibiotics 2015, 4, 18–41. [Google Scholar] [CrossRef] [PubMed]
- Kelemen, J.; Sztermen, M.; Dakos, E.K.; Budai, J.; Katona, J.; Szekeressy, Z.; Sipos, L.; Papp, Z.; Stercz, B.; Dunai, Z.A.; et al. Complex Infection-Control Measures with Disinfectant Switch Help the Successful Early Control of Carbapenem-Resistant Acinetobacter baumannii Outbreak in Intensive Care Unit. Antibiotics 2024, 13. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Quintanilla, M.; Pulido, M.R.; Moreno-Martinez, P.; Martin-Pena, R.; Lopez-Rojas, R.; Pachon, J.; McConnell, M.J. Activity of host antimicrobials against multidrug-resistant Acinetobacter baumannii acquiring colistin resistance through loss of lipopolysaccharide. Antimicrob. Agents Chemother. 2014, 58, 2972–2975. [Google Scholar] [CrossRef] [PubMed]
- Moffatt, J.H.; Harper, M.; Harrison, P.; Hale, J.D.; Vinogradov, E.; Seemann, T.; Henry, R.; Crane, B.; St Michael, F.; Cox, A.D.; et al. Colistin resistance in Acinetobacter baumannii is mediated by complete loss of lipopolysaccharide production. Antimicrob. Agents Chemother. 2010, 54, 4971–4977. [Google Scholar] [CrossRef] [PubMed]
- Sowlati-Hashjin, S.; Carbone, P.; Karttunen, M. Insights into the Polyhexamethylene Biguanide (PHMB) Mechanism of Action on Bacterial Membrane and DNA: A Molecular Dynamics Study. J. Phys. Chem. B. 2020, 124, 4487–4497. [Google Scholar] [CrossRef] [PubMed]
- Chindera, K.; Mahato, M.; Sharma, A.K.; Horsley, H.; Kloc-Muniak, K.; Kamaruzzaman, N.F.; Kumar, S.; McFarlane, A.; Stach, J.; Bentin, T.; et al. The antimicrobial polymer PHMB enters cells and selectively condenses bacterial chromosomes. Sci. Rep. 2016, 6, 23121. [Google Scholar] [CrossRef] [PubMed]
- Knauf, G.A.; Cunningham, A.L.; Kazi, M.I.; Riddington, I.M.; Crofts, A.A.; Cattoir, V.; Trent, M.S.; Davies, B.W. Exploring the Antimicrobial Action of Quaternary Amines against Acinetobacter baumannii. mBio 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Wang, H.; Zhao, D.; Zhang, S.; Hu, C. Polymyxins: recent advances and challenges. Front. Pharmacol. 2024, 15, 1424765. [Google Scholar] [CrossRef] [PubMed]
- Diani, E.; Bianco, G.; Gatti, M.; Gibellini, D.; Gaibani, P. Colistin: Lights and Shadows of an Older Antibiotic. Molecules 2024, 29. [Google Scholar] [CrossRef] [PubMed]
- Carretero-Ledesma, M.; Garcia-Quintanilla, M.; Martin-Pena, R.; Pulido, M.R.; Pachon, J.; McConnell, M.J. Phenotypic changes associated with Colistin resistance due to Lipopolysaccharide loss in Acinetobacter baumannii. Virulence 2018, 9, 930–942. [Google Scholar] [CrossRef] [PubMed]
- Sampson, T.R.; Liu, X.; Schroeder, M.R.; Kraft, C.S.; Burd, E.M.; Weiss, D.S. Rapid killing of Acinetobacter baumannii by polymyxins is mediated by a hydroxyl radical death pathway. Antimicrob. Agents Chemother. 2012, 56, 5642–5649. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Jia, X.; Adams, M.D.; Ghosh, S.K.; Bonomo, R.A.; Weinberg, A. Epithelial innate immune response to Acinetobacter baumannii challenge. Infect. Immun. 2014, 82, 4458–4465. [Google Scholar] [CrossRef] [PubMed]
- Voronko, O.E.; Khotina, V.A.; Kashirskikh, D.A.; Lee, A.A.; Gasanov, V.A.O. Antimicrobial Peptides of the Cathelicidin Family: Focus on LL-37 and Its Modifications. Int. J. Mol. Sci. 2025, 26. [Google Scholar] [CrossRef] [PubMed]
- Dutta, D.; Cole, N.; Kumar, N.; Willcox, M.D. Broad spectrum antimicrobial activity of melimine covalently bound to contact lenses. Invest. Ophthalmol. Vis. Sci. 2013, 54, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Yasir, M.; Dutta, D.; Willcox, M.D.P. Comparative mode of action of the antimicrobial peptide melimine and its derivative Mel4 against Pseudomonas aeruginosa. Sci. Rep. 2019, 9, 7063. [Google Scholar] [CrossRef] [PubMed]
- Yasir, M.; Dutta, D.; Willcox, M.D.P. Mode of action of the antimicrobial peptide Mel4 is independent of Staphylococcus aureus cell membrane permeability. PLoS ONE 2019, 14, e0215703. [Google Scholar] [CrossRef] [PubMed]
- Schulthess, B.; Brodner, K.; Bloemberg, G.V.; Zbinden, R.; Bottger, E.C.; Hombach, M. Identification of Gram-positive cocci by use of matrix-assisted laser desorption ionization-time of flight mass spectrometry: comparison of different preparation methods and implementation of a practical algorithm for routine diagnostics. J. Clin. Microbiol. 2013, 51, 1834–1840. [Google Scholar] [CrossRef] [PubMed]
- Yasir, M.; Willcox, M.D.P. The use of ultraviolet light generated from light-emitting diodes for the disinfection of transvaginal ultrasound probes. PLoS. One. 2024, 19, e0298449. [Google Scholar] [CrossRef] [PubMed]
- Damtie, M.A.; Vijay, A.K.; Willcox, M.D.P. Detection of Genes Associated with Polymyxin and Antimicrobial Peptide Resistance in Isolates of Pseudomonas aeruginosa. Int. J. Mol. Sci. 2025, 26, 10499. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, I.; Hilpert, K.; Hancock, R.E. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Heatmap of antimicrobial agents showing the relationships between their activity profiles. Each cell represents the Spearman correlation coefficient (r) between a pair of compounds, with values ranging from -1.00 (strong negative correlation) to +1.00 (strong positive correlation). Dark red indicates strong positive correlations, white represents no correlation, and blue reflects strong negative correlations. ***, p <0.0001; **, p < 0.005; *, p <0.05.
Figure 1.
Heatmap of antimicrobial agents showing the relationships between their activity profiles. Each cell represents the Spearman correlation coefficient (r) between a pair of compounds, with values ranging from -1.00 (strong negative correlation) to +1.00 (strong positive correlation). Dark red indicates strong positive correlations, white represents no correlation, and blue reflects strong negative correlations. ***, p <0.0001; **, p < 0.005; *, p <0.05.


Table 1.
The strains used in the current study.
| Strain code | Origin (disease, body site, country, year of isolation) |
|---|---|
| ATCC 17978 | Meningitis, ATCC, USA |
| ATCC 19606 | Urine, ATCC, USA |
| ATCC 19606 (R) | LPS mutant [54] |
| Pk.A1 | Sepsis, Blood, Pakistan, 2023 |
| Pk.A2 | Sepsis, Blood, Pakistan, 2023 |
| Pk.A3 | Sepsis, Blood, Pakistan, 2023 |
| Pk.A5 | Sepsis, Blood, Pakistan, 2023 |
| Pk.A6 | Sepsis, Blood, Pakistan, 2023 |
| Pk.A8 | Sepsis, Blood, Pakistan, 2023 |
| Pk.M1 | Sepsis, Blood, Pakistan, 2023 |
| Pk.M10 | Sepsis, Blood, Pakistan, 2023 |
| Pk.M11 | Sepsis, Blood, Pakistan, 2023 |
| Pk.M2 | Sepsis, Blood, Pakistan, 2023 |
| Pk.M3 | Sepsis, Blood, Pakistan, 2023 |
| Pk.M4 | Sepsis, Blood, Pakistan, 2023 |
| Pk.M5 | Sepsis, Blood, Pakistan, 2023 |
| Pk.M9 | Sepsis, Blood, Pakistan, 2023 |
| PW01a | Unknown origin, Australia, 2012 |
| PW01b | Unknown origin, Australia, 2012 |
| PW01c | Unknown origin, Australia, 2012 |
| RB01 | Sepsis, Blood, Australia, 2012 |
| RB01AA7 | Bronchiectasis, Sputum, Australia, 2012 |
| RB02d1 | Tracheal aspirate, Australia, 2012 |
| WM00 | Bronchiectasis, Sputum, Australia, 2012 |
| WM97a | Bronchiectasis, Sputum, Australia, 2012 |
| WM97b | Unknown origin, Australia, 2012 |
| WM98 | Wound, Australia, 2012 |
| WM99c | Sputum, Australia, 2012 |
| Ab08 | Inguinal swab, Switzerland, 2014 |
| Ab39 | Rectal swab, Switzerland, 2020 |
| Ab61 | Pharyngeal swab, Switzerland, 2016 |
Table 2.
Minimum inhibitory concentration (MIC) of different cationic antimicrobials against Acinetobacter baumannii isolates.
Table 2.
Minimum inhibitory concentration (MIC) of different cationic antimicrobials against Acinetobacter baumannii isolates.
| Strain | MIC (μg/ml) | MIC (μg/ml) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| S≤2, I=4, R≥8* | |||||||||
| Polymyxin B | Colistin | Mel4 | Melimine | LL-37 | LFcin | PQ-1 | PHMB | CHX | |
| ATCC 17978 | 2 | 0.5 | 15.6 | 62.5 | 62.5 | >500 | 25 | 12.5 | 25 |
| ATCC 19606 | 1 | 0.5 | 62.5 | 62.5 | 62.5 | >500 | 25 | 12.5 | 25 |
| ATCC 19606 (R) | 1250 | >2500 | 62.5 | 62.5 | 62.5 | >500 | 12.5 | 6.3 | 6.3 |
| Pk.A1 | 1 | 1 | 250 | 250 | 62.5 | ND | 12.5 | 25 | 12.5 |
| Pk.A2 | 2 | 2 | 250 | 250 | 31.3 | ND | 6.3 | 25 | 50 |
| Pk.A3 | 2 | 1 | 250 | 250 | 62.5 | ND | 12.5 | 50 | 12.5 |
| Pk.A5 | 1 | 1 | 15.6 | 250 | 62.5 | ND | 25 | 25 | 50 |
| Pk.A6 | 1 | 0.5 | 250 | 250 | 62.5 | ND | 25 | 25 | 50 |
| Pk.A8 | 2 | 1 | 250 | 250 | 62.5 | ND | 50 | 12.5 | 25 |
| Pk.M1 | 2 | 1 | 250 | 125 | 62.5 | ND | 12.5 | 25 | 25 |
| Pk.M2 | 1 | 1 | 250 | 125 | 31.3 | ND | 25 | 12.5 | 6.3 |
| Pk.M3 | 2 | 2 | 250 | 125 | 62.5 | ND | 25 | 6.3 | 25 |
| Pk.M4 | 1 | 2 | 250 | 125 | 62.5 | ND | 12.5 | 12.5 | 25 |
| Pk.M5 | 1 | 2 | 250 | 125 | 62.5 | ND | 12.5 | 12.5 | 12.5 |
| Pk.M8 | 1 | 2 | 250 | 125 | 31.3 | ND | 12.5 | 25 | 12.5 |
| Pk.M9 | 1 | 1 | 250 | 125 | 62.5 | ND | 25 | 12.5 | 12.5 |
| Pk.M10 | 1 | 1 | 62.5 | 62.5 | 62.5 | ND | 6.3 | 25 | 6.3 |
| Pk.M11 | 1 | 2 | 125 | 250 | 62.5 | ND | 6.3 | 25 | 25 |
| PW01a-AA1 | 1 | 1 | 125 | 62.5 | 62.5 | >500 | 50 | 25 | 25 |
| PW01b-AA2 | 1 | 1 | 125 | 62.5 | 62.5 | >500 | 12.5 | 50 | 25 |
| WM00-AA3 | 2 | 1 | 125 | 125 | 62.5 | >500 | 25 | 25 | 25 |
| WM98-AA4 | 1 | 0.5 | 62.5 | 62.5 | 62.5 | >500 | 25 | 25 | 25 |
| WM99c-AA5 | 2 | 1 | 15.6 | 125 | 125 | >500 | 25 | 25 | 25 |
| PW01c-AA6 | 1 | 0.5 | 125 | 125 | 125 | >500 | 12.5 | 25 | 25 |
| RB01-AA7 | 1 | 0.5 | 31.3 | 250 | 62.5 | >500 | 50 | 50 | 50 |
| RB02d1-AA8 | 1 | 0.25 | 15.6 | 125 | 125 | >500 | 50 | 12.5 | 25 |
| WM97a-AA9 | 2 | 1 | 15.6 | 62.5 | 125 | >500 | 25 | 6.25 | 25 |
| Ab08 | 16 | 8 | 62.5 | 62.5 | 62.5 | ND | 6.3 | 3.1 | 25 |
| Ab39 | 32 | 16 | 250 | 125 | 125 | ND | 6.3 | 3.1 | 50 |
| Ab61 | 64 | 64 | 500 | 125 | 125 | ND | 6.3 | 3.1 | 6.3 |
*R = resistant, I = intermediate and S = Sensitive, cut offs from CLSI for A. baumannii; Lfcin = lactoferricin; PQ-1 = polyquaternium-1; PHMB = polyhexamethylene biguanide; CHX = chlorhexidine. ND = not done.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



