Submitted:
27 May 2026
Posted:
28 May 2026
You are already at the latest version
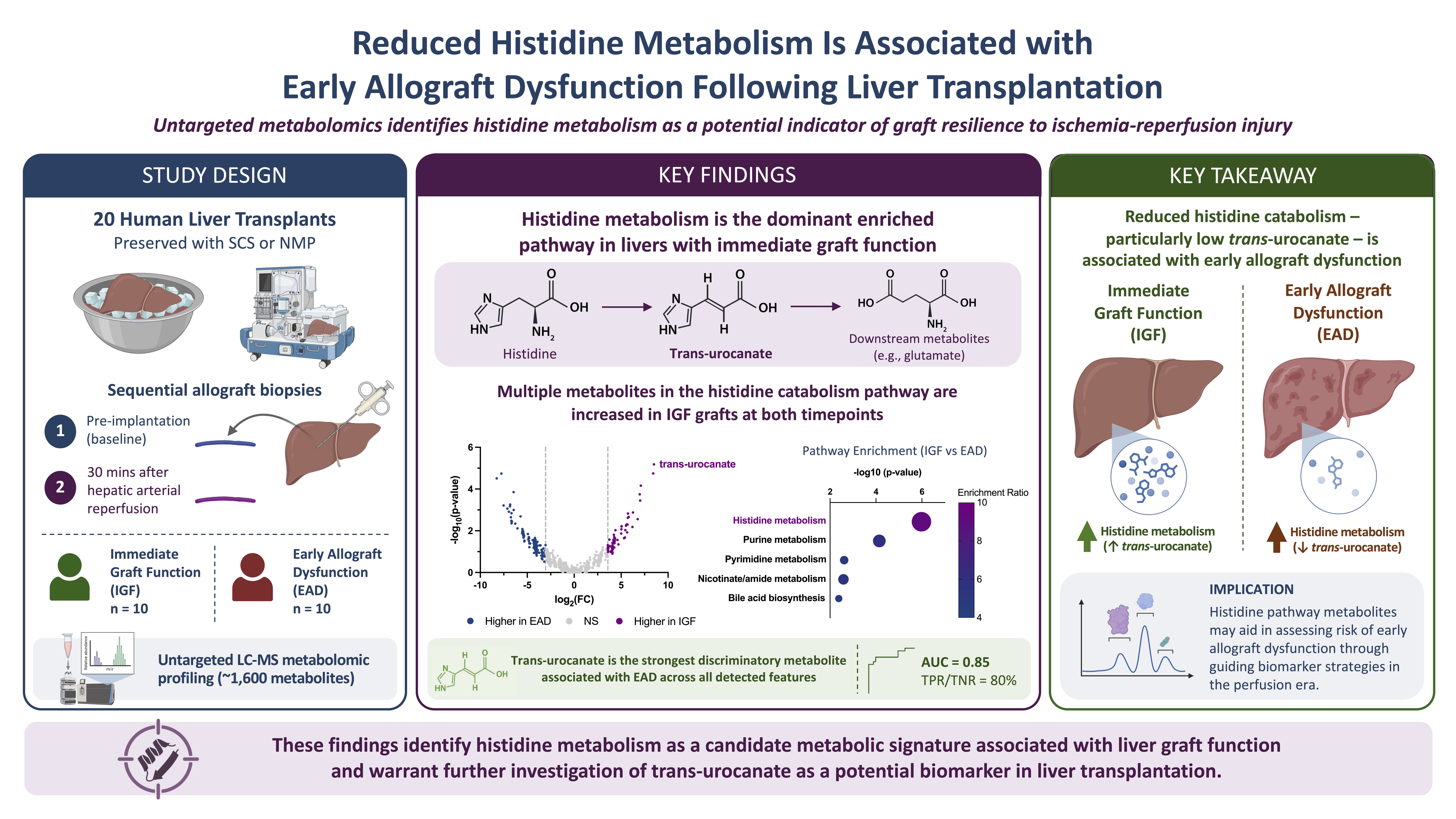
Abstract
Background/Objectives: Early allograft dysfunction (EAD) is a common complication after liver transplantation and is associated with inferior graft survival While normothermic machine perfusion (NMP) has reduced EAD incidence, prediction of early graft performance prior to implantation remains elusive. We aimed to correlate peri-transplant energetic and metabolic profile of liver grafts with post-transplant outcome in a cohort that included grafts preserved with NMP. Methods: Sequential biopsies were taken from 20 transplanted livers (10 immediate graft function [IGF] and 10 EAD), preserved by either static cold storage or NMP. Samples were collected immediately prior to implantation and 30 minutes after hepatic arterial reperfusion. Untargeted liquid chromatography-mass spectrometry was performed, and energy charge was calculated as (ATP + ½ ADP)/(ATP + ADP + AMP). Univariate and receiver operating characteristic analysis identified metabolites correlated with EAD and assessed predictive accuracy. Results: Hepatic concentrations of adenine nucleotides and calculated energy charge did not differ between outcome groups either before implantation or after reperfusion. In contrast, trans-urocanate was significantly enriched in IGF livers across both time points, and additional histidine catabolism pathway metabolites were preferentially increased in IGF grafts. Trans-urocanate demonstrated discriminatory performance for EAD with 80% sensitivity and 80% specificity, confirmed as the single strongest predictive feature among >1,600 detected metabolites. Conclusions: These data identify histidine catabolism as a novel metabolic pathway associated with early graft function and a potential indicator of allograft resilience to ischemia-reperfusion injury. Integration of histidine-pathway metabolites into perfusion-era viability assessment may serve as a discriminative biomarker of EAD and support future metabolite-guided graft optimization strategies.
Keywords:
liver transplantation
; early allograft dysfunction
; histidine metabolism
; ischemia-reperfusion injury
; graft resilience
; biomarkers
; machine perfusion
1. Introduction
Early allograft dysfunction (EAD) occurs in up to 20-40% of liver transplants despite advances in donor graft procurement, preservation, and peri-operative management [1,2]. EAD is associated with increased intensive care utilization, prolonged hospital length of stay, and inferior short and long-term outcomes including reduced graft survival and increased mortality [3,4]. Extended-criteria donors, including donation after circulatory death (DCD), steatotic and older grafts, are increasingly used in transplant programs despite their elevated risk for EAD [5,6], highlighting a critical unmet need for biomarkers that can predict graft performance [7,8].
Normothermic machine perfusion (NMP) has emerged as a transformative preservation technology that not only extends preservation time but also enables real-time functional assessment of donor livers prior to transplantation [9,10]. Current viability assessment during NMP relies mostly on lactate clearance, bile production, and vascular flows [11], all metrics based on the central role of adenosine triphosphate (ATP) depletion and mitochondrial failure during ischemia-reperfusion injury (IRI) [12,13]. Previous mechanistic studies have demonstrated that adenine nucleotide content and hepatic energy charge were relevant molecular indicators of organ viability [14,15]. Finding additional metabolic metrics that can be measured during NMP would allow us to discriminate between grafts that will develop EAD and those that will achieve immediate graft function (IGF) in modern perfusion era.
Prior metabolomic studies have demonstrated that amino acid metabolism, redox buffering pathways, and mitochondrial substrate utilization are dynamically altered during machine perfusion and transplantation [16,17]. In this study, we performed untargeted metabolomic profiling of human liver allograft biopsies obtained immediately prior to implantation and following hepatic arterial reperfusion in livers preserved with static cold storage (SCS) or having undergone NMP. Our goal was to identify molecular signatures associated with early graft outcome that could be targeted for intervention during machine perfusion. Specifically, we examined traditional energy metrics and alternative metabolic pathways to better discriminate between grafts that developed EAD and those that achieved IGF.
2. Materials and Methods
2.1. Study Population
Adult candidate recipients (aged 18-75 years) for deceased donor liver transplantation were included in this study following informed consent. To identify relevant metabolites, we selected 10 control patients with IGF and 10 patients that suffered from EAD from a larger cohort of patients that underwent transplant between February 2022 and November 2023. Selection was done by chronological order of transplantation and with the goal of donor-matching between groups for sex and DBD/DCD status (Table 1). In each group, half of the grafts were placed on NMP, while the other half was transported on SCS. Exclusion criteria included HIV-1 seropositivity.
Patients received perioperative standard of care for liver transplantation. Post-transplant recipient liver function tests, international normalized ratio, ICU length of stay (LOS), pressor use, HCV status, and recipient warm ischemic time (WIT) were determined by retrospective chart review. Donor age, gender, donor type (DCD vs. donation after brain death [DBD]) and cold ischemic time (CIT) were determined by retrospective review of the recipient medical chart, which had information transferred from the donor packet accompanying the donor organ. EAD was defined by the presence of at least one of the following criteria: bilirubin > 10 mg/dL on post-op day 7 (POD7), INR > 1.6 on POD7, or ALT or AST >2000 IU/L within the first 7 days post-op (1). The Mass General Brigham Investigational Review Board approved this clinical study (IRB protocol #2013P000872).
2.2. Surgical and Biopsy Procedure
All donor livers for transplantation were obtained following standard procurement procedure for DBD or DCD. None of the livers underwent normothermic regional perfusion (NRP). Livers were cold-flushed in situ with University of Wisconsin (UW) solution. Livers undergoing SCS were transported in UW preservation solution on ice. Livers undergoing NMP were perfused using the liver Organ Care System (TransMedics Inc., Andover, MA) according to the manufacturer instructions. Livers were flushed through the portal vein with lactated ringers-albumin solution immediately before implantation. Arterial anastomoses and reperfusion were performed separately and after portal reperfusion. A maximum of 2 needle biopsies were taken at sequential time points: immediately before implantation (out of ice [OOI]) and 30 minutes after hepatic artery reperfusion (HAR). Samples were immediately (<10 sec) flash frozen in the operating room using liquid nitrogen and stored at -80 °C until further analysis.
2.3. Untargeted Metabolomic Analysis
Tissue samples were homogenized, centrifuged, and the supernatant was analyzed by Hydrophilic Interaction Liquid Chromatography coupled to tandem mass spectrometry (HILIC-MS/MS). Proteins were extracted in Tris-HCl buffer with protease/phosphatase inhibitors and quantified by BCA Protein Assay Kit (Thermo Scientific, Massachusetts, US). HILIC-MS/MS was performed in both positive and negative ionization modes using an Agilent 6495 QgQ system with 1290 UHPLC system. Data acquisition and metabolite quantification were performed using Agilent MassHunter software (B.07.00) based on Extracted Ion Chromatogram areas for multiple reaction monitoring (MRM) transitions. Peak areas were analyzed in R with drift correction and noise filtering (CV > 30%) performed using MRM PROBS software. Relative metabolite abundances were normalized prior to downstream analyses. Energy charge was calculated as [ATP + ADP * 0.5]/[ATP + ADP + AMP].
Multivariate analyses were performed using MetaboAnalyst. Partial least squares-discriminant analysis (PLS-DA) was used to assess metabolic separation between grafts with IGF and EAD, and PLS-DA model performance was assessed using repeated cross-validation. Variable importance in projection (VIP) scores were calculated to identify metabolites contributing most strongly to group discrimination. Metabolite set enrichment analysis was performed using pathway-associated metabolite libraries to identify pathways associated with graft outcome. Receiver operating characteristic (ROC) analyses were performed to evaluate discriminatory performance of candidate metabolites for prediction of EAD. Monte Carlo cross-validation was used for multivariate ROC modeling and feature ranking.
2.4. Statistical Analysis
Differences in donor characteristics between groups were tested using a Mann Whitney U or x2 test where applicable. A logistic regression was performed to test differences in cofactor ratios and DRI between groups. Analysis was performed using R version 4.4.2 (2024-10-31). Data are presented as median (Q1-Q3) unless otherwise noted.
3. Results
3.1. Donor and Recipient Characteristics
Donor and recipient characteristics are presented in Table 1. A total of 20 transplanted livers were analyzed, including 10 grafts that developed EAD and 10 with IGF. Donor characteristics were similar between groups including donor age, sex, donor type, and total WIT. CIT trended longer in the EAD group, but did not reach statistical significance. Recipient age and Model for End-stage Liver Disease (MELD) score were similar between groups. However, recipient body mass index (BMI) was significantly higher in the EAD group (median 31.6 vs. 25.6 kg/m2, p = 0.008).
3.2. Histidine Metabolism Is the Dominant Enriched Metabolic Pathway Associated with Immediate Graft Function
Direct quantification of adenine nucleotides demonstrated no significant differences in ATP, ADP, AMP concentrations or calculated energy charge between IGF and EAD grafts (Figure 1A). Calculated hepatic energy charge demonstrated substantial overlap between groups both before and after HAR, therefore failing to discriminate graft outcome in this cohort. Despite the absence of differences in conventional energetic metrics, untargeted metabolomic profiling revealed distinct metabolic separation between IGF and EAD grafts by PLS-DA (Figure 1B). Cross-validation analysis also demonstrated stable classifier performance across multiple feature sets, with optimal predictive accuracy achieved using approximately 15 metabolite features. VIP analysis identified lactobionate and trans-urocanate among the highest-ranking discriminatory metabolites contributing to separation between outcome groups (Figure 1C).
Univariate metabolomic analysis identified trans-urocanate as the most significantly enriched metabolite in IGF livers. This enrichment was observed across pre-reperfusion (Figure 2A), post-reperfusion (Figure 2B), and when analyzing the time points together (Figure 2C). Seven of the top 20 VIP-ranked metabolites in IGF livers belonged to the histidine degradation pathway, including trans-urocanate, imidazole propionate, formiminoglutamate, N-acetylhistidine, 4-imidazoleacetate, imidazole lactate, and cis-urocanate. Non-histidine metabolites such as lactobionate demonstrated weaker and less consistent discriminatory performance.
To determine whether coordinated metabolic pathways distinguished graft outcomes, metabolite set enrichment analysis was performed using MetaboAnalyst. Histidine metabolism emerged as the dominant enriched metabolic pathway associated with IGF across timepoints (Figure 3A-C). Additional enriched pathways demonstrated substantially lower enrichment ratios and statistical significance, supporting histidine metabolism as the principal metabolic signature associated with IGF.
3.3. Low Trans-Urocanate Associated with Early Allograft Dysfunction
ROC analysis demonstrated that low trans-urocanate was able to predict EAD with 80% sensitivity and 80% specificity (Figure 4A). Absolute tissue concentrations of trans-urocanate were significantly reduced in EAD grafts compared with IGF grafts (Figure 4B), consistent with metabolomic enrichment analyses observed across reperfusion states.
To further evaluate the discriminatory performance of metabolomic features associated with graft outcome, Monte Carlo cross-validated multivariate ROC modeling was performed. Composite metabolite classifiers demonstrated robust discrimination between IGF and EAD grafts (Figure 5A). Across all multivariate models, trans-urocanate consistently emerged as the most dominant predictive feature (Figure 5B). Notably, inclusion of additional metabolites provided only modest improvement in discriminatory performance, relative to trans-urocanate alone, as demonstrated by the top-performing two-metabolite classifiers (Figure 5C).
4. Discussion
This study identifies histidine metabolism, specifically trans-urocanate, as a previously unrecognized metabolic discriminator of early liver allograft function. Across unbiased metabolic profiling, multiple histidine-derived intermediates were enriched in grafts that achieve immediate function, with low trans-urocanate emerging as the single strongest predictive feature of EAD. These findings identify histidine catabolism as a previously underappreciated pathway that may contribute to hepatocellular resilience during IRI. Among all detected metabolites, trans-urocanate demonstrated the most dominant and most consistent association with IGF, predicting EAD with 80% sensitivity and 80% specificity. Notably, its elevation was observed both before implantation and after reperfusion, suggesting that it reflects an intrinsic metabolic state of the graft rather than a secondary response to reperfusion injury.
Importantly, the metabolite signature associated with graft function was not restricted to a single metabolite. Of the top 20 VIP-ranked features, seven belonged to the histidine degradation pathway, including multiple other imidazole derivatives. This coordinated enrichment strongly supports a pathway-level biological effect, rather than stochastic statistical association. The presence of both upstream and downstream histidine metabolites suggests that overall pathway flux, rather than isolated accumulation, is what differentiates protected grafts from those that fail early. This raises the question whether impaired activation of histidine catabolism is a metabolic vulnerability state that predisposes grafts to reperfusion injury.
Histidase (histidine ammonia-lyase, encoded by HAL), the rate-limiting enzyme in histidine catabolism, catalyzes the conversion of histidine to urocanate and is predominantly expressed in the liver, making hepatic tissue uniquely positioned for histidine degradation and urocanate production [18,19]. Histidine catabolism produces imidazole-containing intermediates, including trans-urocanate, that function as antioxidants, proton-buffers, and redox-modulators [20,21]. These properties are particularly relevant during the reperfusion phase of transplantation, when oxygen reintroduction drives mitochondrial ROS generation and oxidative damage [22]. Imidazole-containing intermediates modulate oxidative stress and inflammatory responses in experimental hepatic IRI models [23,24], supporting a potential link between histidine pathways and IRI/EAD pathogenesis. Elevated histidine flux in the IGF livers may enhance intracellular hepatic buffering capacity, stabilizing cytosolic and mitochondrial pH during abrupt metabolic transitions, while its derivatives scavenge ROS and limit oxidative damage to mitochondrial membranes and respiratory chain proteins. Preserving mitochondrial integrity with histidine-derived compounds could limit necrotic and apoptotic cascades underlying EAD. Consistent with this hypothesis, modified organ preservation solutions such as modified Histidine-Tryptophan-Ketoglutarate (HTK-N) solution incorporate N-acetylhistidine alongside iron chelators to enhance hepatoprotection during cold storage [25], providing proof-of-concept that histidine-based strategies can improve organ preservation outcomes.
Recipient BMI was significantly higher in the EAD group, consistent with known associations between obesity, mitochondrial stress, impaired antioxidant capacity, and heightened susceptibility to IRI and poor transplant outcome [26,27]. Obesity is also characterized by altered amino acid handling and diminished redox flexibility [28]. It is therefore plausible that grafts transplanted into recipients with higher BMI face a greater burden of oxidative and inflammatory injury, which may overwhelm any histidine-linked protective effect. However, the reproducibility of the histidine signal across donors, preservation conditions, and time points indicates that histidine metabolism more likely reflects a graft-intrinsic metabolic program associated with resilience to injury, rather than a surrogate of recipient metabolic risk.
Of note, a previous study found increased histidine metabolites in pre-transplant biopsies of EAD livers [29]. However, these were pre-preservation biopsies taken immediately after procurement, with only 30-40 minutes of CIT, so that very little ischemic and no reperfusion injury would have occurred. The profile of histidine metabolism observed in their samples, therefore, would not reflect changes during cold storage, NMP, or reperfusion. In contrast, our biopsies were obtained after preservation and at reperfusion, capturing the metabolic state of the graft at the time most relevant to IRI-mediated injury. The reversed directionality of histidine metabolite enrichment between the two studies may reflect the dynamic, time-dependent nature of histidine catabolism during the transplant process.
NMP has shifted the paradigm of graft evaluation from static risk estimation to dynamic functional interrogation. Current assessment tools emphasize global physiologic performance, with emerging molecular markers such as flavin mononucleotide (FMN) being validated for mitochondrial viability assessment during hypothermic machine perfusion [30]. Our findings suggest that histidine-pathway metabolites may add a molecular layer of stress-resilience profiling, identifying grafts that are biochemically equipped to withstand reperfusion. From a translational perspective, histidine metabolites are attractive biomarker candidates because they are potentially modifiable during machine perfusion; histidine supplementation or pathway priming could be used to actively enhance graft resilience prior to implantation.
However, this study has several limitations. The small cohort size, along with multiple preservation modalities, limited statistical power for subgroup analyses and external validation. Given the exploratory nature of this untargeted metabolomic study, these findings should be considered hypothesis-generating and will require validation in larger independent cohorts. In addition, while coordinated pathway enrichment supports biologic plausibility, direct causal testing will require experimental manipulation of histidine flux during machine perfusion and transplantation. In addition, longitudinal outcome correlations beyond EAD remain to be investigated.
In summary, this work uncovers histidine metabolism, with trans-urocanate as a significant player, to be a previously unrecognized signal of early liver allograft resilience. Trans-urocanate and related imidazole metabolites may support a metabolic program associated with mitochondrial recovery during IRI. These findings help to move graft assessment beyond static measures of injury toward a systems-level view of metabolic stress tolerance, with implications for future perfusion-based metabolic reprogramming strategies.
Author Contributions
Conceptualization and methodology, A.L., K.U. and H.Y.; investigation, T.A. and S.B.; data curation, T.A. and E.B.; formal analysis, T.A. and A.C.; writing—original draft preparation, A.C.; writing—review and editing, A.C., S.B., A.L. and H.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by US National Institutes of Health (R01DK131543 to HY, R01DK096075 to HY and KU, R01DK114506 to KU, R21AI196804 to AL), and National Science Foundation EEC 1941543 to KU.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Institutional Review Board of Mass General Brigham (protocol code 2013P000872; approval date June 17, 2013).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
Application is extended to New England Donor Services and their generous support that enables research with human donor organs, and special appreciation to the donors of organs and their families whose gift of life enables us to perform research with the goal of reducing number of deaths due to organ failure. Appreciation is also extended to the Mass Spectroscopy Core Facility at Shriners Children’s – Boston for processing our samples. The views, opinion and/or findings contained in this manuscript are those of the authors and should not be construed as an official position, policy or decision of any of the institutions that supported the research, unless designated by other documentation.
Conflicts of Interest
The authors declare competing interests. Drs. Uygun and Yeh have provisional patent applications relevant to the study. Dr. Uygun serves on the Scientific Advisory Board for Sylvatica Biotech Inc., a company focused on developing high subzero organ preservation technology. All competing interests are managed by the Mass General Brigham in accordance with their conflict-of-interest policies.
Abbreviations
The following abbreviations are used in this manuscript:
| ADP | adenosine diphosphate |
| AMP | adenosine monophosphate |
| ATP | adenosine triphosphate |
| DBD | donor/donation after brainstem death |
| DCD | donor/donation after circulatory death |
| EAD | early allograft dysfunction |
| FMN | flavin mononucleotide |
| HAR | hepatic arterial reperfusion |
| HMP | hypothermic machine perfusion |
| HTK-N | histidine-tryptophan-ketoglutarate n-acetyl-histidine |
| IGF | immediate graft function |
| IRI | ischemia-reperfusion injury |
| NADPH | nicotinamide adenine dinucleotide phosphate |
| NMP | normothermic machine perfusion |
| NRP | normothermic regional perfusion |
| PLS-DA | partial least squares discrimination analysis |
| ROC | receiver operating characteristic |
| ROS | reactive oxygen species |
| SCS | static cold storage |
| UW | University of Wisconsin |
| VIP | variable importance in projection |
References
- Olthoff, K.M.; Kulik, L.; Samstein, B.; et al. Validation of a current definition of early allograft dysfunction in liver transplant recipients and analysis of risk factors. Liver Transpl. 2010, 16(8), 943–949. [Google Scholar] [CrossRef]
- Masior, Ł.; Grąt, M. Primary Nonfunction and Early Allograft Dysfunction after Liver Transplantation. Dig. Dis. 2022, 40(6), 766–776. [Google Scholar] [CrossRef]
- Shin, J.; Suh, S.W. Early Allograft Dysfunction After Liver Transplantation: Impact on Clinical Outcomes and Associated Risk Factors. Medicina 2025, 61(9), 1710. [Google Scholar] [CrossRef]
- Moosburner, S.; Sauer, I.M.; Förster, F.; et al. Early Allograft Dysfunction Increases Hospital Associated Costs After Liver Transplantation-A Propensity Score-Matched Analysis. Hepatol. Commun. 2020, 5(3), 526–537. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, T.; Chen, C.; et al. Transplantation of Extended Criteria Donor Livers Following Continuous Normothermic Machine Perfusion Without Recooling. Transplantation. 2022, 106(6), 1193–1200. [Google Scholar] [CrossRef] [PubMed]
- Croome, K.P.; Mathur, A.K.; Mao, S.; et al. Perioperative and long-term outcomes of utilizing donation after circulatory death liver grafts with macrosteatosis: A multicenter analysis. Am. J. Transplant. 2020, 20(9), 2449–2456. [Google Scholar] [CrossRef] [PubMed]
- Meszaros, A.T.; Weissenbacher, A.; Schartner, M.; et al. The Predictive Value of Graft Viability and Bioenergetics Testing Towards the Outcome in Liver Transplantation. Transpl. Int. 2024, 37, 12380. [Google Scholar] [CrossRef]
- Lin, Y.; Huang, H.; Chen, L.; Chen, R.; Liu, J.; Zheng, S.; Ling, Q. Assessing Donor Liver Quality and Restoring Graft Function in the Era of Extended Criteria Donors. J. Clin. Transl. Hepatol. 2023, 11(1), 219–230. [Google Scholar] [CrossRef] [PubMed]
- Mergental, H.; Perera, M.T.; Laing, R.W.; et al. Transplantation of Declined Liver Allografts Following Normothermic Ex-Situ Evaluation. Am. J. Transplant. 2016, 16(11), 3235–3245. [Google Scholar] [CrossRef]
- Okumura, K.; Dhand, A.; Misawa, R.; Sogawa, H.; Veillette, G.; Nishida, S. Normothermic Machine Perfusion Is Associated With Improvement in Mortality and Graft Failure in Donation After Cardiac Death Liver Transplant Recipients in the United States. Transpl. Direct 2024, 10(8), e1679. [Google Scholar] [CrossRef]
- Mergental, H.; Laing, R.W.; Kirkham, A.J.; et al. Transplantation of discarded livers following viability testing with normothermic machine perfusion. Nat. Commun. 2020, 11(1), 2939. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.A.; Murphy, N.; Richards, D.A.; et al. Interstitial Lactic Acidosis in the Graft During Organ Harvest, Cold Storage, and Reperfusion of Human Liver Allografts Predicts Subsequent Ischemia Reperfusion Injury. Transplantation. 2006, 82(2), 227–233. [Google Scholar] [CrossRef] [PubMed]
- Go, K.L.; Lee, S.; Zendejas, I.; Behrns, K.E.; Kim, J.S. Mitochondrial Dysfunction and Autophagy in Hepatic Ischemia/Reperfusion Injury. BioMed Res. Int. 2015, 2015, 183469. [Google Scholar] [CrossRef] [PubMed]
- Kamiike, W.; Burdelski, M.; Steinhoff, G.; Ringe, B.; Lauchart, W.; Pichlmayr, R. Adenine nucleotide metabolism and its relation to organ viability in human liver transplantation. Transplantation 1988, 45(1), 138–143. [Google Scholar] [CrossRef]
- Sumimoto, K.; Inagaki, K.; Yamada, K.; Kawasaki, T.; Dohi, K. Reliable indices for the determination of viability of grafted liver immediately after orthotopic transplantation. Bile flow rate and cellular adenosine triphosphate level. Transplantation 1988, 46(4), 506–509. [Google Scholar] [CrossRef]
- Bruinsma, B.G.; Sridharan, G.V.; Weeder, P.D.; et al. Metabolic profiling during ex vivo machine perfusion of the human liver. Sci. Rep. 2016, 6, 22415. [Google Scholar] [CrossRef]
- de Vries, R.J.; Cronin, S.E.J.; Romfh, P.; Pendexter, C.A.; Jain, R.; et al. Non-invasive quantification of the mitochondrial redox state in livers during machine perfusion. PLoS ONE 2021, 16(10), e0258833. [Google Scholar] [CrossRef]
- Torres, N.; Martínez, L.; Alemán, G.; Bourges, H.; Tovar, A.R. Histidase expression is regulated by dietary protein at the pretranslational level in rat liver. J. Nutr. 1998, 128(5), 818–824. [Google Scholar] [CrossRef]
- Fiadjoe, H.K.; Hoteit, T.; Jones, H.P.; Park, I.W.; Chaudhary, P. Histidine ammonia lyase expression characteristics in primary liver cancer: hepatocellular carcinoma and cholangiocarcinoma. Discov. Oncol. 2025, 16(1), 2179. [Google Scholar] [CrossRef]
- Saadati, S.; Kabthymer, R.H.; Aldini, G.; Mousa, A.; Feehan, J.; de Courten, B. Effects of carnosine and histidine-containing dipeptides on biomarkers of inflammation and oxidative stress: a systematic review and meta-analysis. Nutr. Rev. 2024, 82(12), 1696–1709. [Google Scholar] [CrossRef]
- Brosnan, M.E.; Brosnan, J.T. Histidine Metabolism and Function. J. Nutr. 2020, 150 (Suppl 1), 2570S–2575S. [Google Scholar] [CrossRef]
- Dar, W.A.; Sullivan, E.; Bynon, J.S.; Eltzschig, H.; Ju, C. Ischaemia reperfusion injury in liver transplantation: Cellular and molecular mechanisms. Liver Int. 2019, 39(5), 788–801. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Akahoshi, T.; Mizuta, Y.; et al. Histidine-Rich Glycoprotein Alleviates Liver Ischemia/Reperfusion Injury in Mice With Nonalcoholic Steatohepatitis. Liver Transpl. 2021, 27(6), 840–853. [Google Scholar] [CrossRef] [PubMed]
- Caruso, G.; Di Pietro, L.; Cardaci, V.; Maugeri, S.; Caraci, F. The therapeutic potential of carnosine: Focus on cellular and molecular mechanisms. Curr. Res. Pharmacol. Drug Discov. 2023, 4, 100153. [Google Scholar] [CrossRef] [PubMed]
- Mohr, A.; Brockmann, J.G.; Becker, F. HTK-N: Modified Histidine-Tryptophan-Ketoglutarate Solution-A Promising New Tool in Solid Organ Preservation. Int. J. Mol. Sci. 2020, 21(18), 6468. [Google Scholar] [CrossRef]
- Nair, S.; Verma, S.; Thuluvath, P.J. Obesity and its effect on survival in patients undergoing orthotopic liver transplantation in the United States. Hepatology 2002, 35(1), 105–109. [Google Scholar] [CrossRef]
- Tashiro, H.; Kuroda, S.; Mikuriya, Y.; Ohdan, H. Ischemia–reperfusion injury in patients with fatty liver and the clinical impact of steatotic liver on hepatic surgery. Surg. Today 2014, 44(9), 1611–1625. [Google Scholar] [CrossRef]
- Gaggini, M.; Carli, F.; Rosso, C.; et al. Altered amino acid concentrations in NAFLD: Impact of obesity and insulin resistance. Hepatology 2018, 67(1), 145–158. [Google Scholar] [CrossRef]
- Cortes, M.; Pareja, E.; García-Cañaveras, J.C.; et al. Metabolomics discloses donor liver biomarkers associated with early allograft dysfunction. J. Hepatol. 2014, 61(3), 564–574. [Google Scholar] [CrossRef]
- Eden, J.; Thorne, A.M.; Bodewes, S.B.; et al. Assessment of liver graft quality during hypothermic oxygenated perfusion: The first international validation study. J. Hepatol. 2025, 82(3), 523–534. [Google Scholar] [CrossRef]
Figure 1.
Untargeted metabolomic profiling identifies global metabolic separation between IGF and EAD grafts despite similar hepatic energy charge. (A) Hepatic ATP, ADP, AMP concentrations before and after reperfusion for IGF (in red) and EAD (in green) grafts. (B) PLS-DA of untargeted metabolomic profiles demonstrating metabolic separation between IGF (red) and EAD (green) grafts. (C) VIP scores identifying metabolites contributing most strongly to group discrimination in the PLS-DA, including trans-urocanate and lactobionate. Biopsies were obtained from 10 grafts per group.
Figure 1.
Untargeted metabolomic profiling identifies global metabolic separation between IGF and EAD grafts despite similar hepatic energy charge. (A) Hepatic ATP, ADP, AMP concentrations before and after reperfusion for IGF (in red) and EAD (in green) grafts. (B) PLS-DA of untargeted metabolomic profiles demonstrating metabolic separation between IGF (red) and EAD (green) grafts. (C) VIP scores identifying metabolites contributing most strongly to group discrimination in the PLS-DA, including trans-urocanate and lactobionate. Biopsies were obtained from 10 grafts per group.

Figure 2.
Trans-urocanate is enriched in IGF grafts across reperfusion states. Differential metabolite abundance in biopsies pre-implantation (A), post-reperfusion (B), and combined timepoint analyses (C), showing significant enrichment of trans-urocanate in IGF grafts.
Figure 2.
Trans-urocanate is enriched in IGF grafts across reperfusion states. Differential metabolite abundance in biopsies pre-implantation (A), post-reperfusion (B), and combined timepoint analyses (C), showing significant enrichment of trans-urocanate in IGF grafts.

Figure 3.
Metabolite set enrichment analysis identifies coordinated activation of histidine metabolism in grafts with immediate graft function. Pathway enrichment analysis performed in MetaboAnalyst using untargeted metabolomic profiles from liver graft biopsies. Histidine metabolism emerged as the most significantly enriched metabolite set in IGF vs. EAD grafts both before reperfusion (A), after reperfusion (B), and in combined analyses (C). Bubble size indicates enrichment ratio and bubble color corresponds to pathway significance (-log10 p-value).
Figure 3.
Metabolite set enrichment analysis identifies coordinated activation of histidine metabolism in grafts with immediate graft function. Pathway enrichment analysis performed in MetaboAnalyst using untargeted metabolomic profiles from liver graft biopsies. Histidine metabolism emerged as the most significantly enriched metabolite set in IGF vs. EAD grafts both before reperfusion (A), after reperfusion (B), and in combined analyses (C). Bubble size indicates enrichment ratio and bubble color corresponds to pathway significance (-log10 p-value).

Figure 4.
Trans-urocanate demonstrates strong discriminatory performance for EAD. (A) ROC curve demonstrating the ability of hepatic trans-urocanate to discriminate EAD from IGF grafts with 80% sensitivity and 80% specificity. (B) Absolute tissue concentrations of trans-urocanate in IGF vs. EAD grafts, with significantly reduced levels in grafts developing EAD.
Figure 4.
Trans-urocanate demonstrates strong discriminatory performance for EAD. (A) ROC curve demonstrating the ability of hepatic trans-urocanate to discriminate EAD from IGF grafts with 80% sensitivity and 80% specificity. (B) Absolute tissue concentrations of trans-urocanate in IGF vs. EAD grafts, with significantly reduced levels in grafts developing EAD.

Figure 5.
Multivariate metabolomic classifiers identify trans-urocanate as the dominant predictive feature associated with graft outcome. (A) Monte Carlo cross-validation analysis ROC curves demonstrating predictive accuracy of multivariate metabolites classifiers across increasing feature-set sizes. (B) Feature importance ranking showing trans-urocanate as the highest-ranking contributor to classifier performance across multivariate models. (C) ROC performance of the three top-performing two-metabolite classifiers, demonstrating only modest improvement in discriminatory performance with additional metabolites beyond trans-urocanate.
Figure 5.
Multivariate metabolomic classifiers identify trans-urocanate as the dominant predictive feature associated with graft outcome. (A) Monte Carlo cross-validation analysis ROC curves demonstrating predictive accuracy of multivariate metabolites classifiers across increasing feature-set sizes. (B) Feature importance ranking showing trans-urocanate as the highest-ranking contributor to classifier performance across multivariate models. (C) ROC performance of the three top-performing two-metabolite classifiers, demonstrating only modest improvement in discriminatory performance with additional metabolites beyond trans-urocanate.


Table 1.
Donor and Recipient Demographics.
| No EAD | EAD | p-value | |
| DONOR | |||
| Age | |||
| Median [IQR] | 41.5 [24.8, 53.0] | 51.0 [41.8, 59.3] | 0.265 |
| Sex | |||
| Female | 5 (50.0%) | 3 (30.0%) | 0.65 |
| Male | 5 (50.0%) | 7 (70.0%) | |
| Pump | |||
| No | 5 (50.0%) | 5 (50.0%) | 1 |
| Yes | 5 (50.0%) | 5 (50.0%) | |
| Donor Type | |||
| DBD | 6 (60.0%) | 6 (60.0%) | 1 |
| DCD | 4 (40.0%) | 4 (40.0%) | |
| Warm Ischemic Time | |||
| Median [IQR] | 23.0 [22.5, 24.0] | 28.0 [19.0, 30.0] | 0.408 |
| Cold Ischemic Time | |||
| Median [IQR] | 265.0 [150.0, 343.0] | 422.0 [385.0, 454.0] | 0.068 |
| RECIPIENT | |||
| Age | |||
| Median [IQR] | 59.5 [51.0, 63.5] | 56.0 [48.0, 62.3] | 0.692 |
| Sex | |||
| Female | 5 (50.0%) | 4 (40.0%) | 1 |
| Male | 5 (50.0%) | 6 (60.0%) | |
| BMI | |||
| Median [IQR] | 25.6 [24.7, 28.3] | 31.6 [29.7, 36.8] | 0.008 |
| MELD score | |||
| Median [IQR] | 27.0 [22.8, 28.0] | 27.5 [19.5, 30.5] | 0.572 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



