Submitted:
25 May 2026
Posted:
26 May 2026
You are already at the latest version
Abstract
Prostate cancer remains one of the most biologically heterogeneous malignancies in men and continues to pose major clinical challenges despite substantial advances in diagnosis and treatment. Although androgen deprivation therapy and newer androgen receptor targeted agents have improved disease control, therapeutic resistance and progression to castration resistant prostate cancer remain common. Increasing evidence suggests that prostate cancer progression is influenced not only by tumor intrinsic genetic alterations, but also by complex interactions involving inflammatory signaling, metabolic reprogramming, oxidative stress, epigenetic regulation, immune dysfunction, and the gut microbiome. In parallel, interest has grown in the potential role of natural products as modulators of multiple interconnected biological pathways relevant to prostate cancer biology. This review examines the molecular mechanisms underlying prostate cancer progression with particular emphasis on androgen receptor signaling, PI3K/AKT and MAPK pathway activation, oxidative stress, lipid metabolism, inflammatory signaling, and epigenetic dysregulation. The emerging role of the gut microbiome in systemic androgen metabolism, immune regulation, microbial metabolite production, and inflammatory signaling is also discussed. In this context, the review evaluates current evidence describing the biological effects of selected natural products including curcumin, epigallocatechin gallate, resveratrol, quercetin, sulforaphane, genistein, and related phytochemicals that may influence tumor growth through modulation of microbiome composition, androgen receptor activity, inflammatory pathways, metabolic adaptation, and epigenetic regulation. Available translational and clinical evidence is critically examined together with important limitations related to bioavailability, pharmacokinetics, interindividual variability, microbiome heterogeneity, and incomplete mechanistic understanding. Rather than considering natural products as isolated therapeutic alternatives, this review adopts a systems level perspective in which dietary bioactive compounds may function as modulators of interconnected regulatory networks linking microbiome dynamics, metabolism, endocrine signaling, inflammation, and tumor biology. A more integrated understanding of these interactions may help inform future translational and precision based strategies in prostate cancer research.
Keywords:
prostate cancer
; natural products
; tumor microenvironment
; gut microbiome
; androgen receptor signaling
; epigenetics
; microRNA
; castration-resistant prostate cancer
; PI3K/AKT
; oxidative stress
1. Introduction
Prostate cancer remains one of the most frequently diagnosed malignancies among men worldwide and continues to impose a substantial burden on health systems, patients, and families [1,2]. Although advances in imaging, molecular stratification, surgery, radiotherapy, androgen receptor targeted therapy, chemotherapy, and radioligand approaches have improved clinical management, prostate cancer remains biologically heterogeneous, with outcomes ranging from indolent localized disease to metastatic and castration resistant phenotypes [3,4]. This variability reflects the interaction of tumor intrinsic alterations with endocrine signaling, inflammatory pathways, metabolic adaptation, epigenetic remodeling, immune regulation, and the tumor microenvironment [5,6].
Androgen receptor signaling remains central to prostate cancer biology [7]. Under physiological conditions, androgen signaling contributes to normal prostate development and epithelial function; however, in malignant disease, dysregulated androgen receptor activity supports proliferation, survival, metabolic reprogramming, and disease progression [8,9]. Androgen deprivation therapy therefore remains a cornerstone of treatment for advanced prostate cancer, but many tumors eventually develop mechanisms that sustain androgen receptor signaling under low androgen conditions [10,11]. These mechanisms include androgen receptor amplification, ligand binding domain mutations, intratumoral androgen synthesis, constitutively active splice variants such as AR-V7, and activation of bypass survival pathways [12,13,14].
Increasing evidence indicates that prostate cancer progression cannot be explained by androgen receptor signaling alone. The PI3K/AKT/mTOR and MAPK pathways interact with androgen receptor signaling through reciprocal feedback loops and contribute to therapeutic adaptation [15,16]. Chronic inflammation, oxidative stress, immune suppression, and altered lipid metabolism further support tumor evolution and resistance [17,18,19]. Epigenetic changes, including DNA methylation, histone modification, chromatin remodeling, and microRNA-mediated regulation, add another level of transcriptional plasticity that allows tumor cells to adapt under therapeutic pressure [20,21].
The gut microbiome has emerged as a potentially important regulator of systemic metabolism, immune homeostasis, inflammatory signaling, and endocrine function in cancer biology [22,23]. In prostate cancer, microbiome-associated pathways may influence androgen metabolism, inflammatory tone, short-chain fatty acid production, epithelial barrier function, and tumor-associated immune responses [24,25]. Experimental evidence suggesting that commensal bacteria may contribute to endocrine resistance through androgen biosynthesis has strengthened interest in the gut–prostate axis as part of the broader systems biology of prostate cancer [26].
Within this evolving framework, natural products and dietary bioactive compounds have gained renewed attention as modulators of interconnected biological pathways rather than as simple cytotoxic agents [27,28]. Phytochemicals such as curcumin, epigallocatechin-3-gallate (EGCG), resveratrol, sulforaphane, quercetin, genistein, and lycopene have been reported to influence androgen receptor signaling, inflammatory pathways, oxidative stress responses, mitochondrial function, epigenetic regulation, and microbiome composition in experimental models [28,29,30,31]. However, translation remains limited by poor bioavailability, pharmacokinetic variability, formulation differences, microbiome heterogeneity, and insufficient high-quality clinical evidence [32,33].
This review examines the molecular basis of prostate cancer progression with particular emphasis on interactions among androgen receptor signaling, microbiome-associated regulation, inflammation, metabolism, oxidative stress, immune modulation, and epigenetic adaptation. It further evaluates the emerging role of natural products as biologically active modulators of these interconnected pathways. Rather than presenting natural products as alternatives to established treatment, this review adopts a cautious systems-level perspective in which dietary bioactive compounds are considered within broader biological networks relevant to prostate cancer progression and translational research.
2. Molecular Basis of Prostate Cancer Progression
Prostate cancer progression is driven by an adaptive biological network involving androgen receptor signaling, compensatory survival pathways, metabolic rewiring, oxidative stress, inflammatory signaling, and epigenetic plasticity [6,7,12,16,18,19]. Although early disease often remains dependent on androgen receptor activation, tumor evolution under therapeutic pressure frequently selects for cellular states that sustain growth despite androgen deprivation [18,19]. This transition from androgen-sensitive disease to castration-resistant prostate cancer is therefore best understood as a dynamic process rather than a single molecular event.
2.1. Androgen Receptor Signaling in Prostate Cancer
The androgen receptor is a ligand-activated transcription factor that plays a central role in both normal prostate biology and prostate cancer progression [7,9]. Testosterone is converted within prostate tissue to dihydrotestosterone by 5α-reductase enzymes; dihydrotestosterone binds androgen receptor with high affinity and promotes receptor activation [7,8]. Ligand binding induces dissociation of heat shock proteins, receptor dimerization, nuclear translocation, and binding to androgen response elements within chromatin [7,9]. Recruitment of transcriptional co-regulators such as SRC-1, p300, and related chromatin-associated proteins enables expression of genes involved in proliferation, survival, differentiation, lipid metabolism, and prostate-specific antigen production [8,9].
Because androgen receptor signaling is central to tumor growth, androgen deprivation therapy and androgen receptor pathway inhibitors remain foundational treatments for advanced disease [10,11]. However, suppression of circulating androgens does not completely extinguish androgen receptor activity, particularly in advanced tumors that acquire adaptive resistance mechanisms [12,13,14].
2.2. Mechanisms of Castration Resistant Prostate Cancer
Castration-resistant prostate cancer develops through multiple mechanisms that preserve androgen receptor signaling or substitute alternative survival pathways under androgen-depleted conditions [7,8,34]. Androgen receptor amplification or overexpression increases receptor sensitivity to low androgen concentrations, while ligand binding domain mutations can broaden receptor responsiveness to alternative ligands or reduce antagonist efficacy [10,11,12]. Intratumoral steroidogenesis may also sustain local androgen availability, further supporting receptor activity [11,35].
A clinically important mechanism of resistance is the emergence of androgen receptor splice variants, particularly AR-V7 [27,35]. AR-V7 lacks the ligand binding domain but retains transcriptional activity, allowing ligand-independent activation of androgen receptor target genes [27]. Detection of AR-V7 in circulating tumor cells has been associated with resistance to enzalutamide and abiraterone in patients with castration-resistant prostate cancer [35]. These findings illustrate why receptor-targeted approaches alone may be insufficient in biologically advanced disease.
2.3. PI3K/AKT/mTOR and MAPK Pathway Cross-Talk
The PI3K/AKT/mTOR pathway is one of the most important non-androgen receptor survival pathways in prostate cancer [6,37]. Loss of PTEN, a negative regulator of PI3K signaling, is common in advanced prostate cancer and contributes to persistent AKT activation, cell survival, metabolic adaptation, and resistance to apoptosis [37]. Importantly, reciprocal feedback exists between androgen receptor signaling and PI3K/AKT signaling, such that inhibition of one pathway may enhance activity of the other [37]. This compensatory biology helps explain why single-pathway inhibition may have limited durability in advanced disease.
MAPK signaling also contributes to proliferation, stress adaptation, inflammatory cross-talk, and androgen receptor phosphorylation [37,38]. Activation of ERK, JNK, and p38 MAPK pathways can influence receptor stability, transcriptional activity, and cellular survival under therapeutic pressure [37,38]. Together, PI3K/AKT/mTOR and MAPK pathways form adaptive signaling networks that intersect with androgen receptor biology and contribute to castration resistance [34,37].
2.4. Metabolic Reprogramming and Lipid Metabolism
Metabolic reprogramming is a major feature of prostate cancer progression [17,19]. Unlike many malignancies that rely heavily on glycolysis, prostate cancer demonstrates strong dependence on lipid synthesis, cholesterol metabolism, and fatty acid oxidation [17]. Androgen receptor signaling directly regulates genes involved in lipid metabolism, including fatty acid synthase and sterol regulatory element-binding proteins [17,18,19]. Increased lipid synthesis provides membrane components, energy substrates, and signaling intermediates necessary for tumor growth and adaptation [17].
Altered lipid metabolism may also influence oxidative stress responses, inflammatory signaling, mitochondrial function, and susceptibility to ferroptosis [17]. These metabolic changes are particularly relevant in advanced disease, where tumor cells must adapt to therapeutic pressure, nutrient stress, and changing microenvironmental conditions [17,18,19].
2.5. Oxidative Stress, Inflammation, and Genomic Instability
Oxidative stress and chronic inflammation contribute to prostate carcinogenesis, progression, and therapeutic resistance [18,19]. Reactive oxygen species generated through mitochondrial dysfunction, inflammatory signaling, and metabolic stress can promote DNA damage, genomic instability, and activation of survival pathways [18]. Inflammatory cytokines such as interleukin-6 and tumor necrosis factor-α activate STAT3 and NF-κB signaling, which support proliferation, angiogenesis, immune suppression, and resistance to apoptosis [18]. These inflammatory pathways also interact with androgen receptor signaling and may sustain receptor activity under low androgen conditions [18,19].
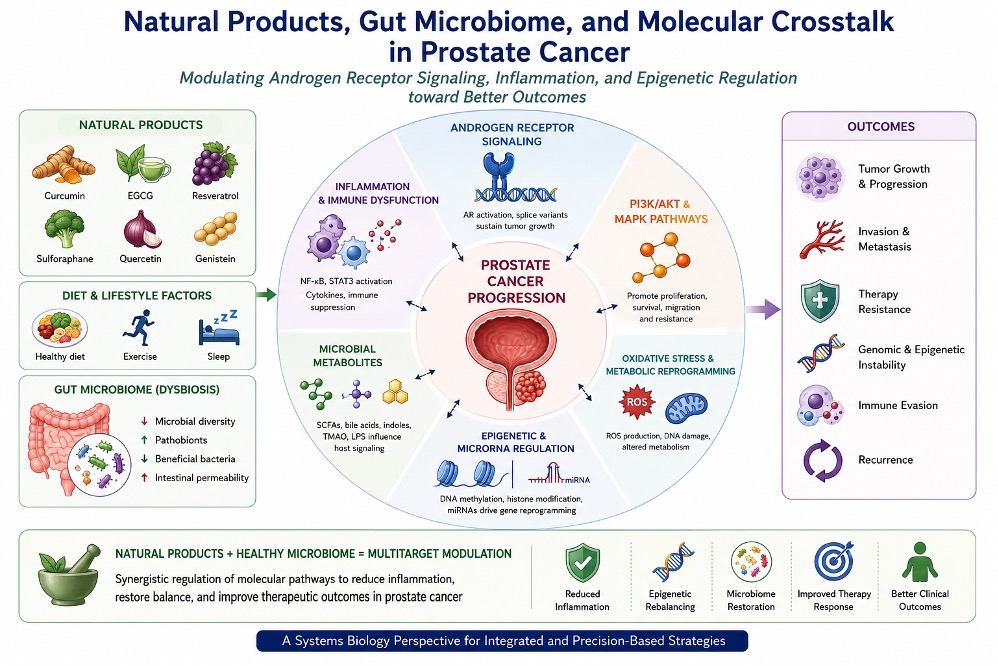
Collectively, the molecular basis of prostate cancer progression reflects a dynamic network of endocrine signaling, survival pathway cross-talk, metabolic adaptation, oxidative stress, inflammation, and epigenetic regulation. This complexity provides a rationale for evaluating natural products not as single-target agents, but as potential modulators of interconnected biological systems relevant to prostate cancer progression. The molecular progression of prostate cancer involves complex interactions among androgen receptor signaling inflammatory pathways, oxidative stress, metabolic reprogramming, and adaptive resistance mechanisms that collectively promote tumor survival and transition toward castration-resistant disease (Figure 1).
3. Gut Microbiome and Prostate Cancer Biology
The human gut microbiome is increasingly recognized as an important regulator of systemic metabolism, immune homeostasis, endocrine signaling, and inflammatory responses [22,23,25]. Through the production of bioactive metabolites and modulation of host immunity, intestinal microbial communities may influence the development and progression of several malignancies, including prostate cancer [22,23,25]. Emerging evidence suggests that microbiome-associated signaling may contribute to androgen metabolism, chronic inflammation, oxidative stress, epigenetic remodeling, and therapeutic responsiveness in prostate cancer [22,23,24,25].
Unlike classical oncogenic pathways that originate primarily within tumor cells, the microbiome represents a dynamic and environmentally responsive biological system influenced by diet, obesity, aging, medications, and host genetics [22,23]. This complexity has generated increasing interest in understanding how microbial dysbiosis may interact with endocrine and immune pathways relevant to prostate cancer progression [22,23,24,25].
3.1. Gut Dysbiosis and Systemic Inflammation
Disruption of microbial homeostasis may promote chronic low-grade systemic inflammation through increased intestinal permeability, altered microbial metabolites, and activation of inflammatory signaling pathways [22,23,39]. Translocation of bacterial products such as lipopolysaccharides can activate Toll-like receptor signaling and stimulate NF-κB, interleukin-6, and tumor necrosis factor-α pathways [40]. Persistent inflammatory signaling contributes to oxidative stress, genomic instability, epithelial–mesenchymal transition, and resistance to apoptosis in prostate cancer [18,40].
Several studies have reported altered microbial composition in patients with prostate cancer compared with healthy individuals, including increased abundance of inflammatory taxa and reductions in bacteria associated with short-chain fatty acid production [22,23]. Although findings remain heterogeneous across populations and methodologies, these observations support a potential relationship between dysbiosis and prostate cancer-associated inflammation [22,23,39]. Emerging evidence suggests that gut microbial dysbiosis influences prostate cancer progression through complex interactions involving inflammatory signaling, microbial metabolites, androgen metabolism, oxidative stress, immune modulation, and epigenetic regulation as illustrated in Figure 2.
3.2. Microbial Regulation of Androgen Metabolism
One of the most important developments in microbiome-associated prostate cancer research involves the observation that intestinal bacteria may influence systemic androgen metabolism [25,39]. Certain microbial populations possess steroid-metabolizing enzymes capable of converting androgen precursors into biologically active metabolites [25]. Experimental evidence suggests that commensal bacteria may partially restore androgen receptor signaling under androgen-deprived conditions and thereby contribute to endocrine resistance [25,26].
The gut-prostate endocrine axis is likely influenced by diet, obesity, antibiotic exposure, hormonal therapy, and host metabolism [22,25]. Bidirectional interactions may also occur because androgen deprivation therapy itself appears capable of altering microbial diversity and metabolic activity [22,25]. Although mechanistic understanding remains incomplete, the microbiome-endocrine relationship represents an important emerging component of prostate cancer systems biology.
3.3. Short-Chain Fatty Acids and Epigenetic Regulation
Short-chain fatty acids generated through microbial fermentation of dietary fiber participate in immune regulation, epithelial barrier maintenance, and metabolic homeostasis [18,23,25,42,43]. Butyrate is particularly important because it functions as a histone deacetylase inhibitor and may therefore influence chromatin accessibility and gene expression [23,25,42]. Through epigenetic regulation, short-chain fatty acids may affect inflammatory signaling, apoptosis, differentiation, and immune responses relevant to tumor biology [20,21,43].
Short-chain fatty acids additionally influence regulatory T-cell differentiation, macrophage activation, and cytokine production [42,43]. However, the biological effects of these metabolites are context dependent and may vary according to concentration, microbial composition, metabolic state, and tumor stage [24,43]. Consequently, simplistic interpretations regarding uniformly protective microbiome metabolites should be avoided.
3.4. Microbiome-Immune Interactions
The gut microbiome exerts substantial influence on both innate and adaptive immunity [25,44]. Microbial metabolites and bacterial antigens shape immune cell differentiation, cytokine production, and systemic inflammatory tone [25]. Dysregulated microbiome-associated signaling may therefore contribute to immune suppression within the prostate tumor microenvironment [45].
Several immune cell populations implicated in prostate cancer progression, including tumor-associated macrophages, myeloid-derived suppressor cells, and regulatory T cells, may be influenced by microbiome-derived signaling molecules [25,45]. Chronic inflammatory activation may promote immunosuppressive cytokine profiles that facilitate tumor escape from immune surveillance [18,43]. Emerging evidence also suggests that microbiome diversity may influence therapeutic responsiveness, including responses to immunotherapy and hormonal therapy [24,25].
3.5. Translational Implications
Despite increasing mechanistic interest, important limitations continue to affect microbiome research in prostate cancer [22,23,24,25]. Many studies involve small cohorts, heterogeneous sequencing methodologies, and cross-sectional designs. Establishing causal relationships remains difficult because microbial alterations may represent either contributors to disease progression or secondary consequences of cancer biology and treatment exposure [24,25]. Nevertheless, current evidence supports the concept that the gut microbiome participates in interconnected regulatory networks involving inflammation, endocrine signaling, metabolism, and immune modulation in prostate cancer.
4. Natural Products as Microbiome Modulators
Natural products and dietary phytochemicals have attracted increasing attention because of their ability to influence multiple biological systems simultaneously, including inflammatory signaling, oxidative stress responses, metabolic regulation, and microbial homeostasis [31,32,33]. Unlike conventional single target pharmacological agents, many plant derived compounds exhibit pleiotropic biological activities that may affect both host cellular pathways and intestinal microbial ecology [46,47]. This multidimensional interaction is particularly relevant in prostate cancer, where chronic inflammation, endocrine signaling, metabolic adaptation, and microbiome associated immune regulation appear to interact dynamically during disease progression [46,47].
The relationship between natural products and the gut microbiome is bidirectional [46,47,48]. On one hand, dietary phytochemicals may alter microbial diversity, intestinal permeability, metabolite production, and inflammatory signaling [47,48]. On the other hand, microbial enzymes metabolize many natural compounds into biologically active derivatives that may possess altered bioavailability and pharmacological activity [48]. These reciprocal interactions may partially explain the variability observed in experimental and clinical responses to natural products across different individuals and populations [48].
Increasing evidence suggests that natural products may exert biologically relevant effects not only through direct tumor associated signaling pathways, but also indirectly through modulation of microbiome associated regulatory networks [49].
4.1. Curcumin and Microbial Homeostasis
Curcumin, a polyphenolic compound derived from Curcuma longa, is among the most extensively studied natural products in cancer biology [50,51]. Curcumin exhibits anti-inflammatory, antioxidant, epigenetic, and immunomodulatory properties in several experimental models [51]. In prostate cancer, curcumin has been reported to influence androgen receptor signaling, nuclear factor kappa B activation, STAT3 signaling, and oxidative stress pathways [51,52].
Beyond its direct cellular effects, curcumin may also alter gut microbial composition and intestinal inflammatory responses [53]. Experimental studies suggest that curcumin supplementation may increase microbial diversity while promoting the growth of beneficial bacterial taxa associated with short chain fatty acid production and mucosal immune regulation [54]. Curcumin has additionally been associated with reductions in pro-inflammatory microbial populations linked to intestinal permeability and endotoxin mediated inflammation [53,55].
Microbiome mediated metabolism appears to play an important role in curcumin bioactivity [53,56]. Although curcumin demonstrates relatively poor systemic bioavailability, microbial biotransformation may generate metabolites with distinct biological properties [56,57]. Certain microbial enzymes convert curcumin into tetrahydro-curcumin and related derivatives that may retain anti-inflammatory and antioxidant activities [58].
Curcumin may also influence epithelial barrier integrity through modulation of tight junction proteins and inflammatory cytokines [58,59]. Reduced intestinal permeability may theoretically decrease systemic exposure to pro-inflammatory microbial products capable of activating nuclear factor kappa B and interleukin-6 associated signaling pathways [53,60].
Despite encouraging experimental findings, translational interpretation remains limited by variability in formulations, absorption profiles, dosing strategies, and study methodology [52,61]. Consequently, the biological relevance of microbiome mediated curcumin effects in human prostate cancer requires further investigation [61].
4.2. Epigallocatechin-3-Gallate and Microbial Metabolism
Epigallocatechin-3-gallate, the major catechin present in green tea, has been widely studied for its antioxidant and anti-inflammatory properties [62,63]. Experimental studies suggest that EGCG may inhibit androgen receptor signaling, suppress PI3K/AKT activation, reduce oxidative stress, and modulate inflammatory cytokine production in prostate cancer models [63].
The interaction between EGCG and the microbiome appears particularly important because intestinal bacteria participate extensively in catechin metabolism [64]. Microbial enzymes convert EGCG into smaller phenolic metabolites that may exhibit altered absorption and biological activity [63]. These metabolites may influence oxidative stress responses, inflammatory signaling, and epithelial homeostasis [65].
EGCG supplementation has additionally been associated with changes in microbial composition, including increased abundance of bacteria linked to short chain fatty acid production and reductions in inflammatory taxa in some experimental studies [63]. Through these effects, EGCG may indirectly influence systemic inflammatory tone and metabolic regulation [65].
Several mechanisms have been proposed to explain EGCG mediated microbiome interactions, including modulation of microbial enzyme activity, alteration of bile acid metabolism, and effects on intestinal oxidative stress [63,66]. However, human data remain limited, and interindividual variability in catechin metabolism represents an important challenge [63,66].
Differences in diet, microbial diversity, genetic background, and concurrent medication exposure may substantially influence biological responses to EGCG[67]. These factors underscore the complexity of translating microbiome related natural product research into consistent clinical application [63,67].
4.3. Resveratrol and Microbiome Associated Signaling.
Resveratrol is a polyphenolic stilbene found in grapes, berries, peanuts, and red wine [68]. Experimental evidence suggests that resveratrol may influence proliferation, apoptosis, mitochondrial function, oxidative stress responses, and inflammatory signaling in prostate cancer cells [69]. Proposed mechanisms include suppression of nuclear factor kappa B activation, modulation of STAT3 signaling, inhibition of PI3K/AKT pathways, and regulation of sirtuin associated metabolic processes [68,69].
Resveratrol also demonstrates important interactions with the gut microbiome [70]. Several microbial species metabolize resveratrol into derivatives with altered biological activity and absorption characteristics [70]. In turn, resveratrol supplementation may influence microbial diversity, reduce intestinal inflammation, and alter metabolic signaling pathways [69,70].
Animal studies have reported that resveratrol may increase bacterial populations associated with anti-inflammatory effects and epithelial barrier maintenance while reducing microbial taxa linked to endotoxin production and chronic inflammation [70]. Through these effects, resveratrol may contribute indirectly to reduced systemic inflammatory signaling and oxidative stress [71].
Resveratrol associated modulation of bile acid metabolism and mitochondrial function has additionally generated interest in its potential influence on host metabolic homeostasis [72]. Since metabolic reprogramming represents an important component of prostate cancer progression, microbiome mediated metabolic regulation may be biologically relevant [71,72].
4.4. Sulforaphane, Quercetin, and Other Phytochemicals
Sulforaphane, an isothiocyanate derived primarily from cruciferous vegetables such as broccoli, has demonstrated anti-inflammatory, antioxidant, and epigenetic regulatory properties [74]. Sulforaphane activates nuclear factor erythroid 2 related factor 2 signaling, thereby enhancing cellular antioxidant defenses and reducing oxidative stress associated damage [74]. Experimental evidence additionally suggests that sulforaphane may inhibit histone deacetylase activity, modulate androgen receptor signaling, and suppress inflammatory pathways in prostate cancer models [75].
Emerging evidence indicates that sulforaphane may also alter gut microbial composition and influence microbial metabolite production [76]. Interactions between cruciferous vegetable intake, microbial metabolism, and glucosinolate conversion may affect the generation of biologically active isothiocyanates [77].
Quercetin, a flavonoid present in fruits, onions, tea, and several medicinal plants, has similarly demonstrated antioxidant and anti-inflammatory activities [78]. Experimental studies suggest that quercetin may influence androgen receptor signaling, apoptosis, mitochondrial function, and inflammatory cytokine production [79]. Quercetin may additionally alter microbial diversity and reduce oxidative stress associated intestinal inflammation [78]. Other phytochemicals including lycopene, genistein, berberine, apigenin, and luteolin have also demonstrated varying degrees of microbiome associated biological activity [80]. Many of these compounds influence overlapping signaling pathways involving nuclear factor kappa B, STAT3, PI3K/AKT, oxidative stress regulation, and epigenetic remodeling [81]. Several natural products have demonstrated the ability to influence prostate cancer-associated signaling pathways through modulation of inflammatory responses, oxidative stress, androgen receptor signaling, microbiome dynamics, and epigenetic regulation. The principal phytochemicals and their proposed biological actions are summarized in Table 1.
Importantly, these compounds frequently exhibit modest direct pharmacological potency when compared with conventional anticancer drugs [81]. However, their potential biological significance may derive from cumulative and multi-target regulatory effects across interconnected physiological systems [81].
4.5. Challenges in Microbiome Targeted Natural Product Research
Despite increasing scientific interest, several major limitations continue to affect microbiome related natural product research in prostate cancer [82]. One of the principal challenges involves substantial interindividual variability in microbiome composition and metabolic activity [81,82]. Differences in diet, ethnicity, medication exposure, age, obesity, geography, and host genetics may significantly influence microbial responses to natural products [82].
Another important limitation relates to bioavailability and pharmacokinetics [83]. Many phytochemicals demonstrate poor intestinal absorption, rapid metabolism, and limited systemic distribution [80]. In some cases, microbiome associated metabolism may enhance biological activity, whereas in others it may reduce therapeutic potential [83].
Experimental heterogeneity also complicates interpretation [84]. Many studies employ different formulations, extraction methods, dosing regimens, microbial sequencing approaches, and experimental models [84]. Consequently, direct comparison across studies remains difficult [81,84]. In addition, causal relationships between microbiome modulation and tumor associated outcomes remain incompletely established. Altered microbial composition may represent a consequence rather than a cause of metabolic or inflammatory changes associated with cancer progression [85]. These limitations highlight the need for more integrated and mechanistically rigorous investigation combining microbiome analysis, metabolomics, epigenetic profiling, and clinical translational studies [83,85].
5. Natural Products and Androgen Receptor Signaling
Androgen receptor signaling remains the dominant molecular driver of prostate cancer progression and therapeutic resistance [6,7]. Even in advanced disease, persistent activation of androgen receptor-associated transcriptional programs continues to support proliferation, metabolic adaptation, survival, and resistance under conditions of androgen deprivation [6,9]. Consequently, suppression of androgen receptor signaling remains the principal therapeutic strategy in advanced prostate cancer management [10,11]. However, adaptive mechanisms including androgen receptor amplification, splice variant formation, intratumoral steroidogenesis, and pathway cross-talk frequently limit the long-term efficacy of hormonal therapies [11,12,13,16].
Natural products have attracted considerable scientific interest because many phytochemicals appear capable of influencing multiple components of androgen receptor-associated signaling networks simultaneously [27,28,29,30,31]. Unlike highly selective receptor antagonists, several plant-derived compounds exhibit broader regulatory effects involving inflammatory signaling, oxidative stress responses, epigenetic modulation, mitochondrial function, and metabolic pathways that intersect with androgen receptor biology [27,28,29]. Although most available evidence remains preclinical, experimental findings suggest that selected phytochemicals may influence both canonical and noncanonical androgen receptor signaling mechanisms relevant to prostate cancer progression [27,87].
5.1. Direct Modulation of Androgen Receptor Activity
Several natural products have demonstrated the ability to suppress androgen receptor expression, interfere with receptor activation, or alter androgen receptor-mediated transcriptional signaling in experimental prostate cancer models [27,88]. Curcumin is among the most extensively investigated compounds in this context [27,88]. Experimental studies suggest that curcumin may reduce androgen receptor expression, impair receptor binding to androgen response elements, and suppress prostate-specific antigen transcription [50,52]. Curcumin-mediated inhibition of androgen receptor signaling has additionally been associated with reduced proliferation and increased apoptotic signaling in prostate cancer cells [27,50].
EGCG has similarly demonstrated inhibitory effects on androgen receptor signaling [63,66]. Experimental evidence suggests that EGCG may reduce androgen receptor expression, suppress prostate-specific antigen production, and inhibit androgen receptor-dependent cell cycle progression [63,66]. Resveratrol has also been reported to influence androgen receptor signaling through modulation of inflammatory pathways, oxidative stress responses, and mitochondrial metabolism [69,72].
Other phytochemicals including genistein, quercetin, luteolin, apigenin, and lycopene have demonstrated varying degrees of androgen receptor-associated regulatory activity in experimental systems [74,75,76]. However, the magnitude and reproducibility of these effects differ substantially across models, concentrations, and experimental conditions [84].
5.2. Natural Products and Castration-Resistant Prostate Cancer
One of the most clinically relevant areas of investigation involves the potential influence of natural products on mechanisms associated with castration-resistant prostate cancer [8,34]. Castration-resistant disease develops through adaptive molecular changes that permit sustained androgen receptor signaling despite intensive androgen suppression [8,34].
Curcumin has been reported to suppress AR-V7-associated signaling in selected experimental studies [27]. Proposed mechanisms include downregulation of androgen receptor transcription, modulation of receptor coactivators, and suppression of inflammatory pathways involved in ligand-independent receptor activation [52,57,61]. Resveratrol has similarly demonstrated inhibitory effects on androgen receptor splice variant-associated transcriptional activity in some preclinical models [29,30,72].
Genistein and quercetin have additionally demonstrated inhibitory effects on androgen-independent prostate cancer cell growth through modulation of PI3K/AKT signaling, apoptosis, oxidative stress pathways, and inflammatory signaling [79,86,88]. Nevertheless, many preclinical investigations employ concentrations substantially higher than those achievable through dietary exposure or standard supplementation [32,33]. Consequently, the biological significance of these mechanisms in clinical settings remains incompletely established [32,33]. As depicted in Figure 3, natural products exert multitarget biological effects in prostate cancer through modulation of androgen receptor signaling, inflammatory pathways, oxidative stress responses, epigenetic regulation, apoptosis, and metabolic adaptation.
5.3. Cross-Talk between Androgen Receptor Signaling and Survival Pathways
Androgen receptor signaling interacts extensively with inflammatory, metabolic, and survival-associated signaling pathways [7,8,9,10]. This interconnected biology may partially explain why phytochemicals capable of influencing multiple pathways simultaneously continue to attract scientific interest [27,28].
The PI3K/AKT/mTOR pathway represents one of the most important adaptive survival pathways in prostate cancer [15,16]. Experimental studies suggest that curcumin, EGCG, resveratrol, quercetin, and sulforaphane may suppress PI3K/AKT signaling while simultaneously influencing androgen receptor activity [15,27,31]. MAPK signaling also contributes to inflammatory cross-talk, stress adaptation, and androgen receptor stabilization under therapeutic pressure [89]. Several natural compounds have demonstrated inhibitory effects on ERK, JNK, and p38 MAPK activation in experimental systems [27,28,29,30].
5.4. Effects on Lipid Metabolism and Metabolic Adaptation
Metabolic reprogramming is a defining feature of advanced prostate cancer and is closely linked to persistent androgen receptor activity [19,48]. Several natural products may influence lipid metabolic pathways linked to androgen receptor signaling [27,28,90]. Curcumin has demonstrated inhibitory effects on lipid synthesis-associated enzymes and sterol regulatory element-binding proteins [27], whereas EGCG and resveratrol may influence mitochondrial metabolism, oxidative stress regulation, and AMP-activated protein kinase signaling [63,68,69].
5.5. Translational Challenges and Clinical Relevance
Despite substantial mechanistic interest, several limitations continue to affect interpretation of studies investigating natural products and androgen receptor signaling [27,86,88]. Much of the available evidence derives from experimental systems that may not fully replicate the complexity of human prostate cancer biology [27,30,86]. Bioavailability, rapid metabolism, microbiome variability, and tumor heterogeneity further complicate translational interpretation [27,91]. Current evidence therefore supports cautious evaluation rather than simplified assumptions regarding direct therapeutic efficacy.
6. Epigenetic and microRNA Regulation
Epigenetic dysregulation is increasingly recognized as a major contributor to prostate cancer initiation, progression, therapeutic resistance, and tumor plasticity [20,21]. Unlike genetic mutations, epigenetic alterations do not change the DNA sequence itself but instead regulate how genes are expressed, silenced, or reactivated within different biological contexts [20,21]. These mechanisms include DNA methylation, histone modification, chromatin remodeling, enhancer reprogramming, and noncoding RNA-mediated regulation [20,21].
In prostate cancer, epigenetic regulation interacts closely with androgen receptor signaling, inflammatory pathways, oxidative stress responses, metabolic adaptation, and microbiome-associated signaling [20,21]. These interconnected mechanisms are particularly important in advanced disease, where tumor cells adapt to therapeutic pressure through transcriptional flexibility and lineage plasticity [20,21]. Several natural products have been reported to influence epigenetic pathways in experimental models, although their translational significance remains incompletely established [27,28,29].
6.1. DNA Methylation and Transcriptional Silencing
DNA methylation is among the most extensively studied epigenetic mechanisms in prostate cancer [20,92]. Aberrant methylation of CpG-rich promoter regions contributes to transcriptional silencing of tumor suppressor genes involved in apoptosis, oxidative stress regulation, DNA repair, and cell cycle control [20,93].
One of the most characteristic epigenetic alterations in prostate cancer is hypermethylation of GSTP1, which impairs antioxidant defense mechanisms and increases vulnerability to oxidative DNA damage [20]. Additional genes affected by abnormal methylation include PTEN, APC, RASSF1A, and CDH1 [20,94,95]. DNA methyltransferases such as DNMT1 and DNMT3A help maintain these abnormal methylation patterns and may contribute to disease progression and therapeutic resistance [20,96].
Several natural products have demonstrated the ability to influence DNA methylation-associated pathways in experimental systems [27,33,97]. Curcumin may reduce DNA methyltransferase activity and partially restore tumor suppressor gene expression [27]. Genistein and EGCG have similarly demonstrated methylation-related regulatory effects [27,63,98]. However, most findings derive from preclinical studies employing concentrations that may not be physiologically achievable in humans [27,32,88].
6.2. Histone Modification, Chromatin Remodeling, and EZH2
Histone proteins regulate chromatin organization and determine whether genomic regions remain transcriptionally active or repressed [20,21]. Histone acetylation generally promotes transcriptional activation by relaxing chromatin structure, whereas deacetylation favors chromatin compaction and gene repression [20].
Histone deacetylases are frequently dysregulated in prostate cancer and contribute to androgen receptor signaling, epithelial–mesenchymal transition, inflammatory activation, and resistance biology [20,21]. Sulforaphane is among the best studied phytochemicals in this context and has demonstrated histone deacetylase inhibitory activity in prostate cancer models [27,75,99]. Butyrate, a microbiome-derived short-chain fatty acid, also functions as an endogenous histone deacetylase inhibitor, thereby linking gut microbial metabolism with host epigenetic regulation [42,100].
EZH2, a catalytic component of polycomb repressive complex 2, represents another major epigenetic regulator in aggressive prostate cancer [101,102]. Overexpression of EZH2 has been associated with lineage plasticity, metastatic progression, stemness-associated signaling, and resistance to androgen receptor-targeted therapy [101,102]. Experimental evidence suggests that curcumin and resveratrol may influence EZH2-associated signaling pathways in selected cancer models [27,50,52].
6.3. microRNAs and Post-Transcriptional Regulation
MicroRNAs are small noncoding RNAs that regulate gene expression through post-transcriptional repression of target messenger RNAs [103]. Dysregulated microRNA networks contribute to proliferation, apoptosis resistance, invasion, angiogenesis, androgen receptor signaling, and therapeutic resistance in prostate cancer [103,104]. microRNA-21 is among the most extensively studied oncogenic microRNAs in prostate cancer and has been associated with inflammatory signaling, apoptosis resistance, and activation of PI3K/AKT pathways through suppression of PTEN-associated signaling [105,106]. Other oncogenic microRNAs including microRNA-141, microRNA-221, and microRNA-375 have also been implicated in progression and metastatic behavior [107].
Conversely, tumor suppressive microRNAs such as microRNA-34a and members of the microRNA-200 family may inhibit epithelial–mesenchymal transition, stemness-associated signaling, and invasive behavior [108]. Curcumin, resveratrol, genistein, and EGCG have demonstrated the ability to modulate selected microRNA-associated pathways in experimental systems [27,28,29,50]. However, microRNA responses remain highly context dependent and influenced by tumor genotype, metabolic state, inflammatory signaling, and experimental conditions [32,33].
6.4. Crosstalk among Epigenetics, Androgen Receptor Signaling, and the Microbiome
Epigenetic regulation interacts extensively with androgen receptor signaling, inflammatory pathways, and microbiome-associated metabolism [20,21]. Androgen receptor binding to chromatin depends on enhancer accessibility, histone modification, and recruitment of transcriptional co-regulators [6,7,109]. In castration-resistant prostate cancer, epigenetic adaptation may allow tumor cells to maintain androgen receptor-associated transcriptional programs despite androgen deprivation [12,34].
The gut microbiome may additionally influence epigenetic regulation through production of short-chain fatty acids, bile acid derivatives, and inflammatory metabolites capable of modifying chromatin-associated signaling pathways [23,24]. Because many phytochemicals undergo microbial biotransformation, the biological effects of natural products may reflect combined interactions among dietary compounds, microbial metabolism, host inflammatory signaling, and epigenetic regulation [28,29,47,53]. Epigenetic regulators and dysregulated microRNA networks contribute substantially to prostate cancer progression, therapeutic resistance, and tumor plasticity. Selected natural products may influence several of these pathways through modulation of DNA methylation, histone remodeling, inflammatory signaling, and post-transcriptional regulation ( see Table 2).
6.5. Translational Perspectives and Current Limitations
Epigenetic pathways are attractive therapeutic targets because they are potentially reversible [20,37]. However, translating experimental epigenetic findings into clinically meaningful interventions remains difficult [32,33]. Epigenetic signaling is highly context dependent, and broad modulation of chromatin-associated enzymes may produce complex effects across tumor cells, stromal compartments, immune populations, and normal tissues [20].
For natural products, additional challenges include poor bioavailability, rapid metabolism, microbiome-dependent biotransformation, inconsistent formulations, and limited tissue-level pharmacodynamic data [32,33]. Most evidence linking phytochemicals to epigenetic modulation in prostate cancer remains preclinical [27,110]]. Few clinical investigations have systematically evaluated epigenetic endpoints such as DNA methylation patterns, histone modifications, enhancer activity, or microRNA signatures in relation to phytochemical exposure [32,110].
Current evidence therefore supports cautious but scientifically relevant interpretation. Natural products may influence epigenetic and microRNA-associated pathways within interconnected biological systems involving inflammation, oxidative stress, endocrine signaling, and microbiome metabolism [20,111]. However, further translational research integrating molecular profiling, microbiome analysis, epigenetic biomarkers, and clinically meaningful endpoints will be necessary to determine whether these mechanisms can be translated into effective strategies for prostate cancer prevention or therapy.
7. Tumor Microenvironment and Immune Modulation
Prostate cancer progression is shaped not only by intrinsic alterations in the tumor cell but also by continuous interactions with the surrounding tumor microenvironment. This environment includes immune cells, cancer-associated fibroblasts, endothelial cells, extracellular matrix components, adipocytes, soluble cytokines, growth factors, and metabolic mediators that collectively influence proliferation, angiogenesis, invasion, immune escape, and therapeutic responsiveness [6,19,112,113]. In advanced disease, the microenvironment often evolves toward a chronically inflammatory and immunosuppressive state, creating conditions that support tumor persistence under therapeutic pressure [113,114].
This microenvironmental remodeling does not occur independently of the molecular pathways discussed earlier. Androgen receptor signaling, PI3K/AKT activation, oxidative stress, lipid metabolic reprogramming, epigenetic plasticity, and microbiome-associated inflammation all converge within the tumor microenvironment [15,16,17,18,20]. Natural products are relevant in this context because many phytochemicals have been reported to modulate inflammatory signaling, oxidative stress, immune cell function, and stromal activation in experimental systems [27,28,29]. However, these effects should be interpreted cautiously, as immune and stromal responses are highly context dependent and may vary according to tumor stage, host metabolism, microbiome composition, and treatment exposure [24,32,33].
7.1. Chronic Inflammation and Cytokine Signaling
Chronic inflammation is a recognized contributor to prostate carcinogenesis and progression [18,21,45]. Persistent inflammatory signaling can promote oxidative DNA damage, angiogenesis, epithelial-mesenchymal transition, stromal activation, and resistance to apoptosis [18]. Cytokines such as interleukin-6, tumor necrosis factor-α, transforming growth factor-β, and interleukin-1β are frequently implicated in tumor-promoting inflammatory networks [115,116].
Interleukin-6 is particularly relevant because it activates STAT3 signaling and may enhance androgen receptor activity under androgen-deprived conditions [116]. This interaction provides a mechanistic bridge between inflammation and castration-resistant growth. NF-κB signaling further supports inflammatory transcriptional programs that promote survival, angiogenesis, immune suppression, and metabolic adaptation [40,116]. These pathways may be amplified by oxidative stress and microbial products derived from gut dysbiosis, which can increase systemic inflammatory tone [18,22].
Curcumin, epigallocatechin gallate, resveratrol, quercetin, and sulforaphane have demonstrated inhibitory effects on inflammatory mediators such as NF-κB, STAT3, interleukin-6, and tumor necrosis factor-α in experimental models [27,28,29,111]. These effects may be relevant to prostate cancer biology because inflammatory cytokines interact with androgen receptor signaling and survival pathways [116]. Nevertheless, clinical translation remains uncertain because cytokine networks are adaptive and influenced by multiple host and tumor variables [117].
7.2. Tumor-Associated Macrophages and Myeloid Cells
Tumor-associated macrophages are major regulators of immune and stromal behavior in prostate cancer [118]. Depending on cytokine exposure, oxygen tension, metabolic conditions, and stromal signals, macrophages may acquire phenotypes ranging from inflammatory and tumor-restrictive to immunosuppressive and tumor-supportive [113,114,118]. In advanced prostate cancer, macrophage populations often contribute to angiogenesis, extracellular matrix remodeling, invasion, and therapeutic resistance [119].
Myeloid-derived suppressor cells also contribute to immune dysfunction by inhibiting cytotoxic T-cell activity and promoting immunosuppressive signaling through arginase activity, reactive oxygen species, and anti-inflammatory cytokines [120]. Myeloid cell–derived mediators can activate STAT3, NF-κB, and PI3K/AKT pathways, thereby reinforcing the signaling networks that sustain tumor progression [15,121].
Several phytochemicals may influence myeloid signaling in experimental systems [27,28,29]. Curcumin and resveratrol have been reported to reduce inflammatory macrophage activation through suppression of NF-κB and oxidative stress pathways [27,29]. Sulforaphane and quercetin may also affect redox-regulated inflammatory responses and macrophage-associated cytokine production [27,74,75]. However, macrophage biology is highly heterogeneous, and experimental findings cannot be directly translated into assumptions of clinical immune restoration [32,33].
7.3. T Cells, Immune Dysfunction, and Immune Evasion
Prostate cancer is often considered an immunologically less responsive tumor compared with melanoma or lung cancer, but immune dysfunction remains important in disease progression [122]. Reduced cytotoxic T-cell infiltration, increased regulatory T-cell activity, impaired antigen presentation, and immunosuppressive stromal signaling collectively contribute to immune evasion [122,123]. These features may partly explain the limited efficacy of immune checkpoint inhibitors in unselected prostate cancer populations [123]. The gut microbiome may further influence systemic and tumor-associated immunity [22,23]. Short-chain fatty acids and other microbial metabolites can affect T-cell differentiation, macrophage function, cytokine production, and immune tolerance [24,25]. Therefore, microbiome-associated signaling may indirectly shape the immune landscape of prostate cancer, particularly in the context of inflammation, obesity, aging, and treatment exposure [22,23,24,25].
Natural products may influence selected immune pathways through anti-inflammatory, antioxidant, and microbiome-associated mechanisms [27, 32, 50. However, broad claims that phytochemicals “boost” anticancer immunity should be avoided. Immune modulation may be beneficial or harmful depending on cellular context, tumor stage, treatment setting, and the balance between inflammatory suppression and antitumor immune activation [27,32,50]. Multiple interconnected signaling pathways link gut microbial dysbiosis, inflammatory signaling, epigenetic regulation, oxidative stress, immune modulation, and tumor microenvironment remodeling during prostate cancer progression. The principal molecular interactions involved in these processes are summarized in Table 3.
7.4. Stromal Remodeling, Angiogenesis, and Extracellular Matrix Interactions
Cancer-associated fibroblasts and extracellular matrix remodeling contribute to prostate cancer invasion, metastasis, and therapeutic resistance [35,124]. Stromal fibroblasts produce cytokines, chemokines, matrix metalloproteinases, and growth factors that promote epithelial–mesenchymal transition, angiogenesis, and tumor cell migration [35,125]. Transforming growth factor-β is a central mediator of stromal activation and fibrosis and may contribute to immune suppression and metastatic dissemination [124,125].
Angiogenesis is also influenced by inflammatory signaling, oxidative stress, hypoxia, and stromal–tumor interactions [16,18,125]. Vascular remodeling supplies nutrients and oxygen to expanding tumors while also influencing immune cell trafficking and drug delivery. In advanced disease, abnormal vascular and stromal architecture may contribute to therapeutic resistance [16,125].
Curcumin, resveratrol, sulforaphane, and quercetin have been reported to influence transforming growth factor-β signaling, matrix metalloproteinase activity, oxidative stress, and epithelial–mesenchymal transition-associated pathways in experimental systems [27,28,29]. These findings are biologically plausible, but the clinical significance of stromal modulation by natural products remains insufficiently established [126].
7.5. Integrated Perspective on Microenvironmental Modulation
The prostate tumor microenvironment is best understood as an integrated biological system rather than a collection of isolated cell types. Inflammation, oxidative stress, stromal activation, immune dysfunction, metabolic adaptation, androgen receptor signaling, and microbiome-associated mediators continuously interact during disease progression [16,28]. These interactions may create feedback loops that sustain tumor growth and therapeutic resistance.
Natural products may influence several components of this network simultaneously, including cytokine signaling, redox balance, macrophage activation, epithelial–mesenchymal transition, and microbiome-derived inflammatory signals [27,28,29]. Their potential relevance therefore lies less in direct cytotoxicity and more in their capacity to modulate biological context. However, tumor heterogeneity, pharmacokinetic limitations, microbiome variability, and differences in immune status make clinical responses difficult to predict [27,28,29]. Future studies should integrate immune profiling, microbiome analysis, metabolomics, and pharmacodynamic endpoints to clarify whether microenvironmental modulation by natural products can become clinically meaningful.
8. Translational and Clinical Evidence
Although mechanistic studies provide a strong biological rationale for investigating natural products in prostate cancer, clinical translation remains limited and uneven [28,29,127]. Many phytochemicals demonstrate effects on androgen receptor signaling, inflammatory pathways, oxidative stress, apoptosis, metabolic adaptation, and epigenetic regulation in experimental models [27,28,86]. However, human studies often produce variable results because of differences in formulations, dosing regimens, patient populations, disease stage, dietary background, microbiome composition, follow-up duration, and clinical endpoints [27,91,128].
A realistic interpretation is therefore essential. Natural products should not be positioned as substitutes for established prostate cancer therapies. Their most plausible role, if validated, may be as adjunctive biological modulators that influence inflammation, metabolism, oxidative stress, microbiome function, or treatment tolerance within broader therapeutic strategies [27,129].
8.1. Curcumin: Clinical and Translational Findings
Curcumin has been widely studied because of its reported effects on NF-κB, STAT3, oxidative stress, apoptosis, androgen receptor signaling, and epigenetic regulation [27,50,51,52]. These mechanisms provide a strong preclinical rationale for prostate cancer research. Some clinical and translational studies have evaluated curcumin as a supplement in men with prostate cancer or as an adjunct to radiotherapy and hormonal therapy [27,52]. Reported outcomes have included changes in inflammatory biomarkers, oxidative stress indicators, quality of life parameters, and prostate-specific antigen kinetics [27,52].
Despite these signals, interpretation remains limited. Curcumin has poor aqueous solubility, limited intestinal absorption, rapid metabolism, and low systemic bioavailability [50,52,130]. Studies also vary substantially in formulation, dose, duration, and patient selection [27,50]. Enhanced formulations, including nanoparticles, liposomes, phospholipid complexes, and piperine-containing preparations, may improve exposure but require more rigorous comparative evaluation [27,52,130]. At present, curcumin is best viewed as a biologically active compound with promising mechanistic effects but insufficient evidence for direct anticancer efficacy in prostate cancer [27,52,130]. Although several natural products demonstrate promising mechanistic activity in experimental models of prostate cancer, clinical translation remains variable because of differences in bioavailability, study design, patient heterogeneity, and treatment endpoints. Major translational and clinical investigations involving selected phytochemicals are summarized in Table 4.
8.2. Green Tea Catechins and Epigallocatechin Gallate
Green tea catechins, particularly epigallocatechin gallate, have been investigated in prostate cancer prevention and progression because of their antioxidant, anti-inflammatory, and antiproliferative effects [27,63,135]. Preclinical studies suggest that EGCG may modulate androgen receptor signaling, PI3K/AKT activity, oxidative stress, and cell cycle regulation [27,63,134]. Clinical studies have produced mixed results. Some trials and observational studies suggest potential effects on prostate cancer risk, prostate-specific antigen kinetics, or oxidative stress markers, whereas others show limited benefit [27,131,135]. Differences in catechin dose, extract composition, treatment duration, baseline dietary exposure, and microbial metabolism may contribute to inconsistent outcomes [27,62,131]. Green tea catechins therefore remain biologically interesting, but current evidence does not support definitive therapeutic recommendations.
8.3. Resveratrol and Metabolic Regulation
Resveratrol has been explored because of its effects on oxidative stress, mitochondrial function, inflammatory signaling, apoptosis, and metabolic regulation [27,68,71]. In prostate cancer models, resveratrol may influence androgen receptor-associated signaling, PI3K/AKT activity, AMPK signaling, and mitochondrial adaptation [68,134].
Human evidence remains limited. Most clinical studies involving resveratrol focus on safety, metabolic biomarkers, inflammatory mediators, or systemic oxidative stress rather than definitive oncological outcomes [27,71,134]. Rapid metabolism and low circulating concentrations of the parent compound remain major translational barriers [68,134]. The interaction between resveratrol, gut microbial metabolism, and host metabolic state may be particularly relevant but remains insufficiently studied in prostate cancer [71,134].
8.4. Soy Isoflavones, Lycopene, and Dietary Patterns
Soy isoflavones, particularly genistein, have attracted interest because of their endocrine-regulatory properties and potential effects on androgen receptor signaling, epigenetic regulation, and inflammatory pathways [27,98]. Epidemiological studies have suggested possible associations between higher soy intake and lower prostate cancer risk in some populations, although causality remains difficult to establish [27,29,136].
Lycopene, a carotenoid abundant in tomatoes, has also been evaluated because of its antioxidant properties and possible association with reduced prostate cancer risk [27,28,137]. However, intervention studies remain inconsistent, and dietary effects are difficult to isolate from broader lifestyle patterns, body composition, metabolic health, and microbiome composition [27,28,137].
8.5. Combination Strategies and Adjunctive Approaches
Natural products may have greater translational relevance as adjunctive modulators rather than standalone therapies [27,28,138]. Experimental studies suggest that curcumin, epigallocatechin gallate, quercetin, sulforaphane, and resveratrol may enhance sensitivity to radiotherapy, chemotherapy, or androgen receptor-targeted treatment by modulating inflammatory signaling, oxidative stress, apoptosis, and adaptive survival pathways [27,28,29,30,31,138]. However, potential interactions with conventional therapies must be evaluated carefully. Phytochemicals may alter drug absorption, hepatic enzyme activity, transporters, oxidative stress responses, and systemic pharmacokinetics [32,33]. Therefore, rational combination strategies require careful pharmacological, safety, and biomarker-driven evaluation before clinical recommendations can be made.
8.6. Limitations of Current Clinical Evidence
The clinical evidence base remains limited by small cohorts, heterogeneous study designs, variable formulations, inconsistent dosing schedules, short follow-up periods, and indirect endpoints [27,32,33,51]. Changes in prostate-specific antigen kinetics, inflammatory biomarkers, or oxidative stress indicators may provide useful biological information, but they do not necessarily translate into improved progression-free survival, metastasis-free survival, or overall survival [27,28,139].
Another major limitation is the lack of integrated mechanistic endpoints. Few clinical studies simultaneously examine pharmacokinetics, tissue exposure, microbiome composition, metabolite generation, inflammatory markers, epigenetic changes, and clinical outcomes [32,33,139]. Without such integration, it remains difficult to determine whether observed biomarker changes reflect meaningful biological modulation. Future trials should include standardized formulations, clear patient stratification, longer follow-up, clinically relevant endpoints, and multi-omics biomarker assessment. Until such evidence becomes available, natural products should be discussed as investigational biological modulators rather than established therapeutic agents in prostate cancer.
9. Challenges and Knowledge Gaps
Despite growing scientific interest in the role of natural products, microbiome-associated signaling, and epigenetic regulation in prostate cancer, several major conceptual and translational challenges remain unresolved [27,32,33]. Much of the current literature demonstrates important mechanistic plausibility; however, the transition from experimental observations to clinically meaningful therapeutic application remains limited [32]. The complexity of prostate cancer biology, together with variability in microbiome composition, host metabolism, pharmacokinetics, and tumor heterogeneity, complicates interpretation of available evidence [16,19,27,138].
Importantly, many natural products exert broad multitarget biological effects rather than highly selective pharmacological actions [27,32,33]. While this systems-level activity may be biologically advantageous, it also creates substantial difficulty in defining precise mechanisms of action and identifying reproducible therapeutic endpoints.
9.1. Biological Heterogeneity of Prostate Cancer
Prostate cancer encompasses diverse molecular, endocrine, metabolic, and immunological phenotypes [6,16,19]. Significant variability exists in androgen receptor signaling intensity, inflammatory activity, genomic instability, lipid metabolism, epigenetic regulation, and tumor microenvironment composition. Consequently, biological responses to natural products are unlikely to be uniform across patients or disease stages [32,33].
Aggressive metastatic and castration-resistant tumors frequently demonstrate adaptive signaling networks involving androgen receptor splice variants, PI3K/AKT activation, inflammatory signaling, and epigenetic plasticity [16,140]. These complex interactions reduce the likelihood that individual phytochemicals will exert consistent therapeutic effects across heterogeneous tumor populations.
9.2. Variability of the Gut Microbiome
The gut microbiome represents a highly dynamic ecosystem influenced by diet, obesity, medications, ethnicity, aging, host genetics, and environmental exposure [22,23,141]. Microbiome composition may therefore differ substantially between individuals and populations, contributing to variability in microbial metabolism of dietary compounds and phytochemicals [142]. Because many natural products undergo microbiome-associated biotransformation, interindividual differences in microbial composition may alter bioavailability, metabolite generation, and biological responsiveness [24,27]. These factors complicate reproducibility across experimental and clinical studies [32,33].
In addition, causal relationships between microbiome alterations and prostate cancer progression remain incompletely defined [24,25]. Microbial dysbiosis may represent either a contributor to disease biology or a secondary consequence of tumor progression, dietary changes, obesity, or treatment exposure [24].
9.3. Bioavailability and Pharmacokinetic Limitations
One of the most important translational barriers involves the poor bioavailability of many phytochemicals [32,33]. Curcumin, resveratrol, quercetin, and several related compounds demonstrate limited aqueous solubility, rapid intestinal and hepatic metabolism, and low systemic exposure following oral administration [32,50,52]. Variability in extraction methods, formulations, purity, and delivery systems further complicates interpretation of experimental and clinical findings [32,33]. In many preclinical studies, concentrations substantially exceed levels achievable in human tissues under normal physiological conditions [32].
9.4. Limitations of Current Experimental Models
Most available evidence derives from in vitro experiments and animal models that may not fully replicate the biological complexity of human prostate cancer [16,142]. Cell line systems often lack the endocrine, immune, stromal, and microbiome-associated interactions present in vivo [142,143].
Similarly, many experimental models fail to adequately reproduce tumor heterogeneity, chronic inflammatory states, metabolic adaptation, microbiome diversity, and therapeutic selection pressures observed in advanced human disease [16,142,143]. Consequently, translation of experimental observations into clinically meaningful outcomes remains uncertain [143]. Despite substantial mechanistic progress, major translational and methodological challenges continue to limit the clinical application of natural products and microbiome-targeted strategies in prostate cancer. The principal knowledge gaps and unresolved issues are summarized in Table 5.
9.5. Complexity of Multitarget Biological Effects
Natural products frequently influence multiple interconnected signaling pathways simultaneously, including androgen receptor signaling, PI3K/AKT activation, inflammatory pathways, oxidative stress responses, mitochondrial metabolism, immune regulation, and epigenetic remodeling [11,15,16]. Although this broad activity may be biologically relevant, it complicates mechanistic interpretation and identification of dominant therapeutic pathways [3,8,32].
In some contexts, multitarget biological effects may produce adaptive or compensatory responses that are difficult to predict. Furthermore, interactions among microbiome-associated metabolites, endocrine signaling, host immunity, and metabolic state may modify biological responses in ways that remain poorly understood [3,63,144].
9.6. Need for Systems Biology and Precision Approaches
Current evidence increasingly supports the need for systems-level investigation rather than reductionist single-pathway approaches [24,27]. Future progress will require integration of molecular profiling, microbiome analysis, metabolomics, epigenetic mapping, pharmacokinetics, and immune characterization [20,145].
Precision oncology approaches may help identify patient subsets more likely to benefit from specific dietary or phytochemical interventions based on microbiome composition, metabolic phenotype, inflammatory state, or molecular signaling profiles [24,146]. Such strategies remain largely exploratory but represent important future directions for translational prostate cancer research.
10. Future Directions and Emerging Perspectives
Future prostate cancer research will likely move increasingly toward integrated systems-biology frameworks that recognize the dynamic interactions among endocrine signaling, metabolism, inflammation, immunity, microbiome regulation, and epigenetic adaptation. Within this evolving paradigm, natural products may ultimately prove most relevant not as isolated therapeutic alternatives, but as biologically active modulators within broader physiological and therapeutic networks.
10.1. Precision Oncology and Molecular Stratification
One of the most important future priorities involves molecular stratification of prostate cancer [146,147,148,149]. Advances in genomics, transcriptomics, metabolomics, and epigenetic profiling may help identify biologically distinct tumor subtypes characterized by specific androgen receptor signatures, inflammatory states, metabolic dependencies, or microbiome-associated features [147,148,150].
Such approaches could facilitate more individualized evaluation of dietary interventions and phytochemical responsiveness [147,148,149,150]. For example, tumors characterized by strong inflammatory signaling or oxidative stress-associated pathways may respond differently to selected natural compounds than tumors dominated primarily by endocrine resistance mechanisms [3,151].
10.2. Microbiome-Informed Therapeutic Strategies
The gut microbiome represents a particularly important area for future translational investigation [22,25]. Advances in metagenomics, metabolomics, and microbial functional analysis may improve understanding of how intestinal bacteria influence androgen metabolism, immune regulation, inflammatory signaling, and therapeutic responsiveness [152,153].
Future strategies may involve combinations of dietary modification, probiotics, prebiotics, microbiome-targeted interventions, and phytochemical supplementation designed to influence microbial metabolite production and systemic inflammatory tone [24,25,27]. However, these approaches require careful validation because microbiome manipulation may produce context-dependent and potentially unpredictable biological effects [24,25].
10.3. Multi-Omics and Systems Biology Integration
Future progress will likely depend on integration of multi-omics technologies including genomics, epigenomics, metabolomics, proteomics, microbiomics, and immune profiling [24,145]. These approaches may help clarify how natural products interact with interconnected biological systems rather than isolated signaling pathways [145].
Systems biology frameworks may additionally improve understanding of adaptive signaling networks involved in castration resistance, metabolic reprogramming, and tumor microenvironment remodeling [15,16,154]. Such integration is particularly important because prostate cancer progression reflects dynamic pathway interactions rather than single molecular events.
10.4. Advanced Formulations and Drug Delivery Systems
Improving bioavailability remains essential for translational progress [27,32]. Nanoparticle delivery systems, liposomal formulations, phospholipid complexes, polymer-based carriers, and microbiome-responsive delivery strategies may enhance stability, absorption, tissue distribution, and pharmacokinetic performance of selected phytochemicals [32,33].
10.5. Combination Strategies and Integrative Therapeutic Models
Natural products may ultimately demonstrate greatest relevance as adjunctive biological modulators integrated with conventional therapeutic approaches [27,28,30,32]. Experimental evidence suggests that selected phytochemicals may influence inflammatory signaling, oxidative stress responses, and adaptive survival pathways associated with resistance to hormonal therapy, chemotherapy, or radiotherapy [27,28,30,32].
Future translational studies should therefore explore rational combination strategies rather than simplistic standalone approaches [32]. However, potential interactions with drug metabolism, hepatic enzyme activity, and systemic pharmacokinetics must be evaluated carefully [32,33]. Emerging evidence supports a systems-level model in which diet, microbiome dynamics, microbial metabolites, epigenetic regulation, immune signaling, and tumor microenvironment interactions collectively shape prostate cancer progression and therapeutic responsiveness as illustrated in Figure 4.
10.6. Humanistic and Preventive Perspectives
Beyond therapeutic intervention, future research should also consider broader preventive and humanistic dimensions of prostate cancer biology [27,28]. Lifestyle, diet, metabolic health, obesity, inflammation, and microbiome composition interact continuously throughout aging and may influence long-term disease susceptibility [18,22,24,27].
Natural products should therefore not be viewed solely through a pharmaceutical framework, but also within broader contexts involving nutrition, lifestyle medicine, metabolic regulation, and preventive health [27,28,30]. Such perspectives may prove particularly important in early disease prevention and survivorship research.
11. Conclusions
Prostate cancer progression reflects a highly interconnected biological process involving androgen receptor signaling, inflammatory pathways, metabolic adaptation, oxidative stress, immune dysregulation, epigenetic remodeling, and microbiome-associated signaling. Increasing evidence suggests that these processes function as integrated regulatory networks rather than isolated molecular events. Within this framework, the gut microbiome has emerged as a potentially important regulator of systemic inflammation, endocrine metabolism, immune homeostasis, and epigenetic signaling relevant to prostate cancer biology.
Natural products including curcumin, epigallocatechin gallate, resveratrol, sulforaphane, quercetin, genistein, and related phytochemicals demonstrate the capacity to influence multiple components of these interconnected systems in experimental models. Reported effects include modulation of androgen receptor activity, PI3K/AKT and MAPK signaling, inflammatory cytokine production, oxidative stress responses, microbial metabolism, and epigenetic regulation. However, substantial translational challenges remain, including poor bioavailability, microbiome variability, tumor heterogeneity, and limited clinical validation.
Current evidence therefore supports a cautious systems-level interpretation rather than simplified assumptions regarding direct anticancer efficacy. Natural products may ultimately prove most relevant not as isolated therapeutic alternatives, but as biologically active modulators within broader integrative strategies linking metabolism, microbiome dynamics, endocrine signaling, inflammation, and tumor biology. Future progress will require rigorously designed translational studies integrating molecular profiling, microbiome analysis, pharmacokinetics, and clinically meaningful outcomes to clarify the therapeutic significance of these interconnected pathways in prostate cancer.
Funding
The authors have not received any funding for this study.
Acknowledgments
The authors are thankful to the Deanship of Graduate Studies and Scientific Research at the University of Bisha for supporting this work through the Fast-Track Research Support Program.
Conflicts of Interest
The authors declare no conflict of interest.
Declaration
All the authors hereby declare that they qualify for the authorship, have reviewed and approved the manuscript for submission.
Ethical Approval
The institutional ethical approval is not applicable.
References
- Bray, F.; Laversanne, M.; Sung, H.; et al. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2024, 74(3), 229–263. [Google Scholar] [CrossRef]
- Siegel, R.L.; Giaquinto, A.N.; Jemal, A. Cancer statistics, 2024. CA Cancer J. Clin. 2024, 74(1), 12–49. [Google Scholar] [CrossRef] [PubMed]
- Cicchetti, R.; Basconi, M.; Litterio, G.; Orsini, A.; Mascitti, M.; Digiacomo, A.; Salzano, G.; Tătaru, O.S.; Ferro, M.; Giulioni, C.; et al. Therapeutic Advances in Metastatic Prostate Cancer: A Journey from Standard of Care to New Emerging Treatment. Int. J. Mol. Sci. 2025, 26(23), 11665. [Google Scholar] [CrossRef]
- Wang, L.; Lu, B.; He, M.; Wang, Y.; Wang, Z.; Du, L. Prostate Cancer Incidence and Mortality: Global Status and Temporal Trends in 89 Countries From 2000 to 2019. Front Public Health Published. 2022, 10, 811044. [Google Scholar] [CrossRef]
- Schafer, E.J.; Laversanne, M.; Sung, H.; et al. Recent Patterns and Trends in Global Prostate Cancer Incidence and Mortality: An Update. Eur. Urol. 2025, 87(3), 302–313. [Google Scholar] [CrossRef]
- Sandhu, S.; Moore, C.M.; Chiong, E.; Beltran, H.; Bristow, R.G.; Williams, S.G. Prostate cancer. Lancet 2021, 398(10305), 1075–1090. [Google Scholar] [CrossRef]
- Quistini, A.; Chierigo, F.; Fallara, G.; et al. Androgen Receptor Signalling in Prostate Cancer: Mechanisms of Resistance to Endocrine Therapies. Res. Rep. Urol. Published. 2025, 17, 211–223. [Google Scholar] [CrossRef]
- Henry, I.; Foreman, R.; Balachandran, L.; Mortimer, E.; Asim, M. Emerging Therapeutic Approaches to Engage the Androgen Receptor for the Treatment of Castration-Resistant Prostate Cancer. Cancers 2025, 17(23), 3755. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Cheng, D.; Li, P. Androgen receptor dynamics in prostate cancer: from disease progression to treatment resistance. Front Oncol. Published. 2025, 15, 1542811. [Google Scholar] [CrossRef] [PubMed]
- Crawford, E.D.; Heidenreich, A.; Lawrentschuk, N.; et al. Androgen-targeted therapy in men with prostate cancer: evolving practice and future considerations. Prostate Cancer Prostatic Dis. 2019, 22(1), 24–38. [Google Scholar] [CrossRef]
- Huang, F.; Li, K.; Shevach, J.W.; Wang, Q. Emerging therapies to overcome antiandrogen resistance and beyond in lethal prostate cancer. J. Natl. Cancer Cent. Published. 2025, 6(1), 42–57. [Google Scholar] [CrossRef]
- Raychaudhuri, R.; Lin, D.W.; Montgomery, R.B. Prostate Cancer: A Review. JAMA 2025, 333(16), 1433–1446. [Google Scholar] [CrossRef]
- Ning, S.; Armstrong, C.M.; Xing, E.; et al. LX1 Dual Targets AR Variants and AKR1C3 in Advanced Prostate Cancer Therapy. Cancer Res. 2024, 84(21), 3617–3628. [Google Scholar] [CrossRef]
- Shin, W.S.; Han, S.H.; Jo, K.W.; Cho, Y.; Kim, K.T. Pinostilbene inhibits full-length and splice variant of androgen receptor in prostate cancer. Sci. Rep. Published. 2023, 13(1), 16663. [Google Scholar] [CrossRef]
- Pungsrinont, T.; Kallenbach, J.; Baniahmad, A. Role of PI3K-AKT-mTOR Pathway as a Pro-Survival Signaling and Resistance-Mediating Mechanism to Therapy of Prostate Cancer. Int. J. Mol. Sci. 2021, 22(20), 11088. [Google Scholar] [CrossRef]
- Wasim, S.; Lee, S.Y.; Kim, J. Complexities of Prostate Cancer. Int. J. Mol. Sci. Published. 2022, 23(22), 14257. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, W.; Kong, P.; et al. New insights into lipid metabolism and prostate cancer (Review). Int. J. Oncol. 2023, 62(6), 74. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Xiao, Y.; Miao, J.; Zhang, X.; Liu, M.; Zhu, L.; Liu, H.; Shen, X.; Wang, J.; Xie, B.; et al. Oxidative Stress and Inflammation: Drivers of Tumorigenesis and Therapeutic Opportunities. Antioxidants 2025, 14(6), 735. [Google Scholar] [CrossRef]
- Sabbah, A.; Delouya, G.; Laskine, M.; Taussky, D. Metabolic Plasticity in Prostate Cancer: The Warburg Effect and Its Clinical Relevance. Am. J. Clin. Oncol. 2025, 48(11), 566–571. [Google Scholar] [CrossRef] [PubMed]
- Enikeeva, K.; Rafikova, G.; Sharifyanova, Y.; Mulyukova, D.; Vanzin, A.; Pavlov, V. Epigenetics as a Key Factor in Prostate Cancer. Adv. Biol. (Weinh) 2024, 8(5), e2300520. [Google Scholar] [CrossRef]
- Rafikova, G.; Gilyazova, I.; Enikeeva, K.; Pavlov, V.; Kzhyshkowska, J. Prostate Cancer: Genetics, Epigenetics and the Need for Immunological Biomarkers. Int. J. Mol. Sci. Published. 2023, 24(16), 12797. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Motiwala, Z.; Corona-Avila, I.; Makhanasa, D.; Alkahalifeh, L.; Khan, M.W. The Gut Microbiome and Its Multifaceted Role in Cancer Metabolism, Initiation, and Progression: Insights and Therapeutic Implications. Technol. Cancer Res. Treat. 2025, 24, 15330338251331960. [Google Scholar] [CrossRef]
- Lachance, G.; Robitaille, K.; Laaraj, J.; et al. The gut microbiome-prostate cancer crosstalk is modulated by dietary polyunsaturated long-chain fatty acids. Nat. Commun. Published. 2024, 15(1), 3431. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Dong, T.; Rong, X.; et al. Microbiome in prostate cancer: pathogenic mechanisms, multi-omics diagnostics, and synergistic therapies. J. Cancer Res. Clin. Oncol. Published. 2025, 151(6), 178. [Google Scholar] [CrossRef]
- Huang, J.; Zhu, X.-H.; Trotman, L.C.; Tsao, C.-K. The Role of Gut Microbiome in Prostate Cancer: Current Evidence and Emerging Opportunities. Cancers 2026, 18(6), 998. [Google Scholar] [CrossRef]
- Pernigoni, N.; Zagato, E.; Calcinotto, A.; et al. Commensal bacteria promote endocrine resistance in prostate cancer through androgen biosynthesis. Science 2021, 374(6564), 216–224. [Google Scholar] [CrossRef]
- Pratama, F.; Novitasari, D.; Mardianingrum, R.; Holik, H.A.; Ikram, N.K.K.; Muchtaridi, M. Bioactive Natural Products Targeting Androgen Receptor Signaling in Prostate Cancer: A Systematic Review. Cancers Published. 2026, 18(5), 786. [Google Scholar] [CrossRef]
- Wu, J.; Ji, H.; Li, T.; et al. Targeting the prostate tumor microenvironment by plant-derived natural products. Cell Signal. 2024, 115, 111011. [Google Scholar] [CrossRef]
- Zhang, M.; Zhong, J.; Yu, M.; et al. Natural product-mediated autophagy targeting for prostate cancer: A novel potential therapeutic strategy. Phytomedicine 2026, 152, 157790. [Google Scholar] [CrossRef]
- Vrânceanu, M.; Galimberti, D.; Banc, R.; et al. The Anticancer Potential of Plant-Derived Nutraceuticals via the Modulation of Gene Expression. Plants Published. 2022, 11(19), 2524. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Oceguera Nava, E.I.; Ashong, D.; Chen, G.; Chen, Q.H. Natural Products Targeting the Androgen Receptor Signaling Pathway: Therapeutic Potential and Mechanisms. Curr. Issues Mol. Biol. Published. 2025, 47(9), 780. [Google Scholar] [CrossRef]
- Metri, N.A.; Mandl, A.; Paller, C.J. Harnessing nature's therapeutic potential: A review of natural products in prostate cancer management. Urol. Oncol. 2025, 43(4), 221–243. [Google Scholar] [CrossRef]
- Qian, R.; Ge, J.; Fan, N.; et al. Natural Products in Cancer Prevention and Therapy: Current Challenges and Future Directions. MedComm (2020) Published. 2026, 7(2), e70585. [Google Scholar] [CrossRef]
- Labrecque, M.P.; Coleman, I.M.; Brown, L.G.; et al. Molecular profiling stratifies diverse phenotypes of treatment-refractory metastatic castration-resistant prostate cancer. J. Clin. Invest. Published. 2019, 129(10), 4492–4505. [Google Scholar] [CrossRef] [PubMed]
- Quistini, A.; Chierigo, F.; Fallara, G.; et al. Androgen Receptor Signalling in Prostate Cancer: Mechanisms of Resistance to Endocrine Therapies. Res. Rep. Urol. Published. 2025, 17, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xu, X.; Fang, S.; Shi, X.; Yang, Y. Inhibition of nuclear import suppresses androgen receptor action and overcomes resistance in prostate cancer. BMC Cancer Published. 2026, 26(1), 428. [Google Scholar] [CrossRef]
- Militaru, F.C.; Militaru, V.; Crisan, N.; et al. Molecular basis and therapeutic targets in prostate cancer: A comprehensive review. Biomol. Biomed. Published. 2023, 23(5), 760–771. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.J.; Asim, M. Unravelling the Role of Kinases That Underpin Androgen Signalling in Prostate Cancer. Cells 2022, 11(6), 952. [Google Scholar] [CrossRef]
- Potrykus, M.; Czaja-Stolc, S.; Stankiewicz, M.; Kaska, Ł.; Małgorzewicz, S. Intestinal Microbiota as a Contributor to Chronic Inflammation and Its Potential Modifications. Nutrients 2021, 13(11), 3839. [Google Scholar] [CrossRef]
- Tong, Y.; Yu, C.; Chen, S.; Zhang, X.; Yang, Z.; Wang, T. Trans-anethole exerts protective effects on lipopolysaccharide-induced acute jejunal inflammation of broilers via repressing NF-κB signaling pathway. Poult. Sci. 2023, 102(2), 102397. [Google Scholar] [CrossRef]
- Ide, H.; Lu, Y.; Yu, J.; et al. Testosterone promotes DNA damage response under oxidative stress in prostate cancer cell lines. Prostate 2012, 72(13), 1407–1411. [Google Scholar] [CrossRef]
- Kalkan, A.E.; BinMowyna, M.N.; Raposo, A.; et al. Beyond the Gut: Unveiling Butyrate's Global Health Impact Through Gut Health and Dysbiosis-Related Conditions: A Narrative Review. Nutrients Published. 2025, 17(8), 1305. [Google Scholar] [CrossRef]
- Du, Y.; He, C.; An, Y.; Huang, Y.; Zhang, H.; Fu, W.; Wang, M.; Shan, Z.; Xie, J.; Yang, Y.; et al. The Role of Short Chain Fatty Acids in Inflammation and Body Health. Int. J. Mol. Sci. 2024, 25(13), 7379. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Meng, Q.; Chen, Y.; et al. Interaction between gut microbiota and immunity in health and intestinal disease. Front Immunol. Published. 2025, 16, 1673852. [Google Scholar] [CrossRef] [PubMed]
- Oseni, S.O.; Naar, C.; Pavlović, M.; et al. The Molecular Basis and Clinical Consequences of Chronic Inflammation in Prostatic Diseases: Prostatitis, Benign Prostatic Hyperplasia, and Prostate Cancer. Cancers Published. 2023, 15(12), 3110. [Google Scholar] [CrossRef]
- Gonçalves, A.C.; Pinto, A.R.; Cima, A.; Olo-Fontinha, E.; Martins, J.C.L.; Garcia, J.; Lemos, A.; Saavedra, M.J.; Pintado, M.M.; Alves, M.J. Plant-Derived Bioactive Compounds: One Health Perspective. Appl. Sci. 2026, 16(1), 327. [Google Scholar] [CrossRef]
- Pacyga, K.; Tabiś, A.; Pacyga, P. Medicinal Plants for a Healthy Gut Microbiome: Scientific Insights into Modern Herbal Applications. Int. J. Mol. Sci. Published. 2025, 26(22), 10875. [Google Scholar] [CrossRef] [PubMed]
- Jyoti, Dey P. Mechanisms and implications of the gut microbial modulation of intestinal metabolic processes. npj Metab. Health Dis. Published. 2025, 3(1), 24. [Google Scholar] [CrossRef]
- Yao, Y.; Zhu, Y.; Chen, K.; et al. Microbiota in cancer: current understandings and future perspectives. Signal Transduct. Target Ther. Published. 2026, 11(1), 39. [Google Scholar] [CrossRef]
- Ameer, S.F.; Mohamed, M.Y.; Elzubair, Q.A.; Sharif, E.A.M.; Ibrahim, W.N. Curcumin as a novel therapeutic candidate for cancer: can this natural compound revolutionize cancer treatment? Front Oncol. Published. 2024, 14, 1438040. [Google Scholar] [CrossRef]
- Hashem, S.; Ali, T.A.; Akhtar, S.; et al. Targeting cancer signaling pathways by natural products: Exploring promising anti-cancer agents. BioMed Pharmacother. 2022, 150, 113054. [Google Scholar] [CrossRef]
- Esmaeli, M.; Dehghanpour Dehabadi, M. Curcumin in prostate cancer: a systematic review of molecular mechanisms and nanoformulated therapeutic strategies. BMC Cancer Published. 2025, 25(1), 1609. [Google Scholar] [CrossRef]
- Balaji, S.; Jeyaraman, N.; Jeyaraman, M.; et al. Impact of curcumin on gut microbiome. World J. Exp. Med. Published. 2025, 15(1), 100275. [Google Scholar] [CrossRef] [PubMed]
- Lamichhane, G.; Olawale, F.; Liu, J.; Lee, D.-Y.; Lee, S.-J.; Chaffin, N.; Alake, S.; Lucas, E.A.; Zhang, G.; Egan, J.M.; et al. Curcumin Mitigates Gut Dysbiosis and Enhances Gut Barrier Function to Alleviate Metabolic Dysfunction in Obese, Aged Mice. Biology 2024, 13(12), 955. [Google Scholar] [CrossRef]
- Servida, S.; Piontini, A.; Gori, F.; Tomaino, L.; Moroncini, G.; De Gennaro Colonna, V.; La Vecchia, C.; Vigna, L. Curcumin and Gut Microbiota: A Narrative Overview with Focus on Glycemic Control. Int. J. Mol. Sci. 2024, 25(14), 7710. [Google Scholar] [CrossRef]
- Zhu, J.; He, L. The Modulatory Effects of Curcumin on the Gut Microbiota: A Potential Strategy for Disease Treatment and Health Promotion. Microorganisms 2024, 12(4), 642. [Google Scholar] [CrossRef]
- Magini, A.; Datti, A. Curcumin Between Pleiotropic Potential and Translational Constraints. Int. J. Mol. Sci. 2026, 27(5), 2212. [Google Scholar] [CrossRef]
- Zeng, A.; Quan, Y.; Tao, H.; Dai, Y.; Song, L.; Zhao, J. The Role of Tetrahydrocurcumin in Tumor and Neurodegenerative Diseases Through Anti-Inflammatory Effects. Int. J. Mol. Sci. 2025, 26(8), 3561. [Google Scholar] [CrossRef] [PubMed]
- Konaktchieva, M.; Stojchevski, R.; Hadzi-Petrushev, N.; Gagov, H.; Konakchieva, R.; Mitrokhin, V.; Kungulovski, G.; Mladenov, M.; Avtanski, D. Curcumin and Tetrahydrocurcumin as Multi-Organ Modulators of the Adipose Tissue–Gut–Liver Axis: Mechanistic Insights, Therapeutic Potential, and Translational Challenges. Pharmaceuticals 2025, 18(12), 1791. [Google Scholar] [CrossRef] [PubMed]
- Di Vincenzo, F.; Del Gaudio, A.; Petito, V.; Lopetuso, L.R.; Scaldaferri, F. Gut microbiota, intestinal permeability, and systemic inflammation: a narrative review. Intern Emerg. Med. 2024, 19(2), 275–293. [Google Scholar] [CrossRef]
- Akter, K.; Gul, K.; Mumtaz, S. Revisiting Curcumin in Cancer Therapy: Recent Insights into Molecular Mechanisms, Nanoformulations, and Synergistic Combinations. Curr. Issues Mol. Biol. 2025, 47(9), 716. [Google Scholar] [CrossRef]
- Mokra, D.; Adamcakova, J.; Mokry, J. Tea Polyphenol (-)-Epigallocatechin-3-Gallate (EGCG): A Time for a New Player in the Treatment of Respiratory Diseases? Antioxidants Published. 2022, 11(8), 1566. [Google Scholar] [CrossRef]
- Capasso, L.; De Masi, L.; Sirignano, C.; Maresca, V.; Basile, A.; Nebbioso, A.; Rigano, D.; Bontempo, P. Epigallocatechin Gallate (EGCG): Pharmacological Properties, Biological Activities and Therapeutic Potential. Molecules 2025, 30(3), 654. [Google Scholar] [CrossRef]
- Liu, Z.; de Bruijn, W.J.C.; Bruins, M.E.; Vincken, J.P. Reciprocal Interactions between Epigallocatechin-3-gallate (EGCG) and Human Gut Microbiota In Vitro. J. Agric. Food Chem. 2020, 68(36), 9804–9815. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.M.; Gul, H.; Khan, S.; et al. Green tea polyphenol epigallocatechin-3-gallate mediates an antioxidant response via Nrf2 pathway in heat-stressed poultry: A review. Poult. Sci. 2025, 104(5), 105071. [Google Scholar] [CrossRef]
- Yang, J.; Chen, W.; Chen, J.; Xie, D.; Wang, Y.; Zhou, J. Tea Polyphenol Epigallocatechin Gallate and the Gut-Health Axis: Unraveling Structural Characteristics, Metabolic Pathways, and Systemic Benefits. Adv. Nutr. 2025, 16(12), 100545. [Google Scholar] [CrossRef]
- Dovey, Z.; Horowitz, A.; Waingankar, N. The influence of lifestyle changes (diet, exercise and stress reduction) on prostate cancer tumour biology and patient outcomes: A systematic review. BJUI Compass Published. 2023, 4(4), 385–416. [Google Scholar] [CrossRef]
- Bejenaru, L.E.; Biţă, A.; Belu, I.; Segneanu, A.-E.; Radu, A.; Dumitru, A.; Ciocîlteu, M.V.; Mogoşanu, G.D.; Bejenaru, C. Resveratrol: A Review on the Biological Activity and Applications. Appl. Sci. 2024, 14(11), 4534. [Google Scholar] [CrossRef]
- Jang, J.Y.; Im, E.; Kim, N.D. Mechanism of Resveratrol-Induced Programmed Cell Death and New Drug Discovery against Cancer: A Review. Int. J. Mol. Sci. Published. 2022, 23(22), 13689. [Google Scholar] [CrossRef] [PubMed]
- Yılmaz, A.; Park, H.J.; Ahn, E.M.; Bae, J. Dietary Polyphenols in Aging: A Systems-Level Perspective on Mitochondrial Quality Control and Microbiome Interactions. Int. J. Mol. Sci. Published. 2026, 27(9), 3930. [Google Scholar] [CrossRef]
- de Sá Coutinho, D.; Pacheco, M.T.; Frozza, R.L.; Bernardi, A. Anti-Inflammatory Effects of Resveratrol: Mechanistic Insights. Int. J. Mol. Sci. Published. 2018, 19(6), 1812. [Google Scholar] [CrossRef]
- Ge, Y.; Zhang, L.; Chen, W.; Sun, M.; Liu, W.; Li, X. Resveratrol Modulates the Redox Response and Bile Acid Metabolism to Maintain the Cholesterol Homeostasis in Fish Megalobrama amblycephala Offered a High-Carbohydrate Diet. Antioxidants 2023, 12(1), 121. [Google Scholar] [CrossRef] [PubMed]
- Kogut, Ł.; Puchalski, C.; Katryńska, D.; Zaguła, G. The Multidirectional Biological Activity of Resveratrol: Molecular Mechanisms, Systemic Effects and Therapeutic Potential-A Review. Nutrients Published. 2026, 18(2), 313. [Google Scholar] [CrossRef]
- Kaiser, A.E.; Baniasadi, M.; Giansiracusa, D.; et al. Sulforaphane: A Broccoli Bioactive Phytocompound with Cancer Preventive Potential. Cancers Published. 2021, 13(19), 4796. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.Y.; Kim, D.; Lee, N.K.; Im, E.; Kim, N.D. Sulforaphane in Cancer Prevention and Therapy: A State-of-the-Art Review of Epidemiological Evidence, Molecular Mechanisms, and Translational Challenges. Int. J. Mol. Sci. 2026, 27(4), 2028. [Google Scholar] [CrossRef]
- Bouranis, J.A.; Beaver, L.M.; Choi, J.; et al. Composition of the Gut Microbiome Influences Production of Sulforaphane-Nitrile and Iberin-Nitrile from Glucosinolates in Broccoli Sprouts. Nutrients Published. 2021, 13(9), 3013. [Google Scholar] [CrossRef]
- Bouranis, J.A.; Beaver, L.M.; Ho, E. Metabolic Fate of Dietary Glucosinolates and Their Metabolites: A Role for the Microbiome. Front Nutr. Published. 2021, 8, 748433. [Google Scholar] [CrossRef] [PubMed]
- Alharbi, H.O.A.; Alshebremi, M.; Babiker, A.Y.; Rahmani, A.H. The Role of Quercetin, a Flavonoid in the Management of Pathogenesis Through Regulation of Oxidative Stress, Inflammation, and Biological Activities. Biomolecules 2025, 15(1), 151. [Google Scholar] [CrossRef]
- Ward, A.B.; Mir, H.; Kapur, N.; Gales, D.N.; Carriere, P.P.; Singh, S. Quercetin inhibits prostate cancer by attenuating cell survival and inhibiting anti-apoptotic pathways. World J. Surg. Oncol. Published. 2018, 16(1), 108. [Google Scholar] [CrossRef]
- Carrera-Quintanar, L.; López Roa, R.I.; Quintero-Fabián, S.; Sánchez-Sánchez, M.A.; Vizmanos, B.; Ortuño-Sahagún, D. Phytochemicals That Influence Gut Microbiota as Prophylactics and for the Treatment of Obesity and Inflammatory Diseases. Mediat. Inflamm. Published. 2018, 2018, 9734845. [Google Scholar] [CrossRef]
- Kaur, S.; Ahmad, S.S.; Raja, V.; Mahajan, P.; Abdi, G. Deciphering the cancer chemopreventive potential of phytochemicals: an integrative perspective on molecular targets and signaling networks. Cancer Cell Int. Published. 2026, 26(1), 127. [Google Scholar] [CrossRef]
- Trecarten, S.; Fongang, B.; Liss, M. Current Trends and Challenges of Microbiome Research in Prostate Cancer. Curr. Oncol. Rep. 2024, 26(5), 477–487. [Google Scholar] [CrossRef]
- Santhiravel, S.; Bekhit, A.E.A.; Mendis, E.; et al. The Impact of Plant Phytochemicals on the Gut Microbiota of Humans for a Balanced Life. Int. J. Mol. Sci. Published. 2022, 23(15), 8124. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Kim, S.J.; Gil, W.J.; Yang, C.S. Exploring the therapeutic potential of phytochemicals: challenges and strategies for clinical translation. Phytomedicine 2025, 145, 157090. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Fang, Y.; Lyu, Z.; et al. Microbiome modulation of tumorigenesis and immune responses. J. BioMed Sci. Published. 2026, 33(1), 4. [Google Scholar] [CrossRef]
- Wu, S.; Oceguera Nava, E.I.; Ashong, D.; Chen, G.; Chen, Q.-H. Natural Products Targeting the Androgen Receptor Signaling Pathway: Therapeutic Potential and Mechanisms. Curr. Issues Mol. Biol. 2025, 47(9), 780. [Google Scholar] [CrossRef]
- Jameel, M.; Fatma, H.; Nadtochii, L.A.; Siddique, H.R. Molecular Insight into Prostate Cancer: Preventive Role of Selective Bioactive Molecules. Life 2023, 13(10), 1976. [Google Scholar] [CrossRef]
- Mazurakova, A.; Samec, M.; Koklesova, L.; et al. Anti-prostate cancer protection and therapy in the framework of predictive, preventive and personalised medicine - comprehensive effects of phytochemicals in primary, secondary and tertiary care. EPMA J. Published. 2022, 13(3), 461–486. [Google Scholar] [CrossRef]
- Lee, S.; Rauch, J.; Kolch, W. Targeting MAPK Signaling in Cancer: Mechanisms of Drug Resistance and Sensitivity. Int. J. Mol. Sci. Published. 2020, 21(3), 1102. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wang, R.; Yu, W.; Shi, C.; Wang, X.; Liu, A.; Zhang, L. Natural Products in the Metabolic and Endocrine Modulation of Polycystic Ovary Syndrome: Current Perspectives. Nutrients 2026, 18(6), 964. [Google Scholar] [CrossRef]
- Narra, F.; Galgani, G.; Harris, C.B.; Moreno, D.A.; Núñez-Gómez, V. Bioavailability, Human Metabolism, and Dietary Interventions of Glucosinolates and Isothiocyanates: Critical Insights and Future Perspectives. Foods 2025, 14(16), 2876. [Google Scholar] [CrossRef] [PubMed]
- Barletta, F.; Bandini, M.; Cirulli, G.O.; et al. DNA methylation alterations in prostate cancer: from diagnosis to treatment. Transl. Androl. Urol. 2025, 14(2), 454–462. [Google Scholar] [CrossRef]
- Bouyahya, A.; Mechchate, H.; Oumeslakht, L.; et al. The Role of mitic Modifications in Human Cancers and the Use of Natural Compounds as Epidrugs: Mechanistic Pathways and Pharmacodynamic Actions. Biomolecules Published. 2022, 12(3), 367. [Google Scholar] [CrossRef]
- Huang, Y.; Mao, J.; Li, X. Emerging biomarkers in prostate cancer diagnosis and treatment: Insights into genetic, RNA and metabolic markers (Review). Int. J. Oncol. 2026, 68(2), 15. [Google Scholar] [CrossRef]
- Zhu, Y.; Liu, K.; Zhu, H. Targeting epigenetic methylation: emerging diagnosis and therapeutic strategies in cancer. Exp. Hematol. Oncol. Published. 2026, 15(1), 27. [Google Scholar] [CrossRef]
- Marei, H.E. Epigenetic regulators in cancer therapy and progression. npj Precis Oncol. Published. 2025, 9(1), 206. [Google Scholar] [CrossRef]
- Shankar, S.; Kumar, D.; Srivastava, R.K. Epigenetic modifications by dietary phytochemicals: implications for personalized nutrition. Pharmacol. Ther. 2013, 138(1), 1–17. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A.M.; Al-Alem, U.; Ali, M.M.; Bosland, M.C. Genistein increases estrogen receptor beta expression in prostate cancer via reducing its promoter methylation. J. Steroid Biochem Mol. Biol. 2015, 152, 62–75. [Google Scholar] [CrossRef]
- Gibbs, A.; Schwartzman, J.; Deng, V.; Alumkal, J. Sulforaphane destabilizes the androgen receptor in prostate cancer cells by inactivating histone deacetylase 6. Proc. Natl. Acad. Sci. U S A 2009, 106(39), 16663–16668. [Google Scholar] [CrossRef]
- Li, L.; Zhao, S.; Xiang, T.; Feng, H.; Ma, L.; Fu, P. Epigenetic connection between gut microbiota-derived short-chain fatty acids and chromatin histone modification in kidney diseases. Chin. Med. J. (Engl) Published. 2022, 135(14), 1692–1694. [Google Scholar] [CrossRef] [PubMed]
- Jacobi, J.J.; Wadosky, K.M.; Jaiswal, N.; et al. EZH2 Suppression Diversifies Prostate Cancer Lineage Variant Evolution and Lacks Efficacy in Inhibiting Disease Progression. Cancer Res. 2026, 86(4), 889–908. [Google Scholar] [CrossRef]
- Singh, R.; Venkadakrishnan, V.B.; Imada, E.; et al. Crosstalk between EZH2 and DNA methylation mediates neuroendocrine prostate cancer lineage plasticity. Nat. Commun. Published. 2026, 17(1), 2992. [Google Scholar] [CrossRef]
- Mir, R.; Waza, A.A.; Mir, M.M. MicroRNAs as Biomarkers for the Diagnosis and Prognosis of Different Diseases and Cancer. Exp. Suppl. 2026, 115, 199–213. [Google Scholar] [CrossRef]
- Rezaei, S.; Jafari Najaf Abadi, M.H.; Bazyari, M.J.; Jalili, A.; Kazemi Oskuee, R.; Aghaee-Bakhtiari, S.H. Dysregulated microRNAs in prostate cancer: In silico prediction and in vitro validation. Iran. J. Basic Med. Sci. 2024, 27(5), 611–620. [Google Scholar] [CrossRef]
- Yuan, F.; Hu, Y.; Lei, Y.; Jin, L. Recent progress in microRNA research for prostate cancer. Discov. Oncol. Published. 2024, 15(1), 480. [Google Scholar] [CrossRef]
- Gan, L.; Zheng, L.; Zou, J.; et al. MicroRNA-21 in urologic cancers: from molecular mechanisms to clinical implications. Front Cell Dev. Biol. Published. 2024, 12, 1437951. [Google Scholar] [CrossRef]
- Andl, T.; Ganapathy, K.; Bossan, A.; Chakrabarti, R. MicroRNAs as Guardians of the Prostate: Those Who Stand before Cancer. What Do We Really Know about the Role of microRNAs in Prostate Biology? Int. J. Mol. Sci. Published. 2020, 21(13), 4796. [Google Scholar] [CrossRef]
- Mansoori, B.; Silvestris, N.; Mohammadi, A.; et al. miR-34a and miR-200c Have an Additive Tumor-Suppressive Effect on Breast Cancer Cells and Patient Prognosis. Genes Published. 2021, 12(2), 267. [Google Scholar] [CrossRef]
- Stelloo, S.; Bergman, A.M.; Zwart, W. Androgen receptor enhancer usage and the chromatin regulatory landscape in human prostate cancers. Endocr. Relat. Cancer 2019, 26(5), R267–R285. [Google Scholar] [CrossRef]
- Zaczek, A.; Rodacka, A. Investigating the Epigenetic Therapeutic Potential of Natural Compounds in Cancer. Int. J. Mol. Sci. 2025, 26(21), 10776. [Google Scholar] [CrossRef]
- Cord, D.; Rîmbu, M.C.; Iordache, M.P.; et al. Phytochemicals as Epigenetic Modulators in Chronic Diseases: Molecular Mechanisms. Molecules Published. 2025, 30(21), 4317. [Google Scholar] [CrossRef]
- Almazrouei, K.M.; Mishra, V.; Pandya, H.; Sambhav, K.; Bhavsar, S.N. Tumor Microenvironment and Its Role in Cancer Progression: An Integrative Review. Cureus Published. 2025, 17(9), e92707. [Google Scholar] [CrossRef]
- Wang, Y.; Luo, S.; Dong, H.; Huang, R.P. Unraveling tumor cell-tumor microenvironment crosstalk through antibody array technologies (Review). Int. J. Oncol. 2025, 67(4), 81. [Google Scholar] [CrossRef]
- Liu, Y.; Yu, Y.; Chen, Y.; Zhuang, J.; Sun, C. Tumor Immunosenescence Driven by Chronic Inflammation: Mechanisms, Microenvironment Remodeling and Therapeutic Strategies. Aging Dis. Published. 2025, 17(3), 1347–1370. [Google Scholar] [CrossRef]
- Hirano, T. IL-6 in inflammation, autoimmunity and cancer. Int. Immunol. 2021, 33(3), 127–148. [Google Scholar] [CrossRef]
- Nishida, A.; Andoh, A. The Role of Inflammation in Cancer: Mechanisms of Tumor Initiation, Progression, and Metastasis. Cells Published. 2025, 14(7), 488. [Google Scholar] [CrossRef]
- Wang, Y.; Lin, A.; Liu, Z.; Cheng, Q.; Zhang, J.; Luo, P. Tumor Microenvironment Onmyoji: Cytokines with Dual Protumor and Antitumor Roles. Cancer Commun. (Lond) Published. 2026, 46, 0008. [Google Scholar] [CrossRef]
- Han, C.; Deng, Y.; Xu, W.; et al. The Roles of Tumor-Associated Macrophages in Prostate Cancer. J. Oncol. Published. 2022, 2022, 8580043. [Google Scholar] [CrossRef]
- Chai, Z.; Dong, X.; Li, S.; et al. Macrophage-derived extracellular vesicles in the remodeling of the prostate cancer immune microenvironment and therapeutic resistance. J. Transl. Med. Published. 2026, 24(1), 553. [Google Scholar] [CrossRef]
- Lu, J.; Luo, Y.; Rao, D.; et al. Myeloid-derived suppressor cells in cancer: therapeutic targets to overcome tumor immune evasion. Exp. Hematol. Oncol. Published. 2024, 13(1), 39. [Google Scholar] [CrossRef]
- Nishida, A.; Andoh, A. The Role of Inflammation in Cancer: Mechanisms of Tumor Initiation, Progression, and Metastasis. Cells 2025, 14(7), 488. [Google Scholar] [CrossRef]
- Flores-Islas, A.K.; Rico-Fuentes, C.; Sierra-Díaz, E.; García-Chagollán, M.; Pereira-Suárez, A.L.; Zepeda-Nuño, J.S.; Moreno-Ortiz, J.M.; Ramírez-de-Arellano, A. Immunotherapeutic Strategies for Prostate Cancer: A Comprehensive Review. Cancers 2026, 18(2), 255. [Google Scholar] [CrossRef]
- Tufail, M.; Jiang, C.H.; Li, N. Immune evasion in cancer: mechanisms and cutting-edge therapeutic approaches. Signal Transduct. Target Ther. Published. 2025, 10(1), 227. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Liu, J.; Qian, H.; Zhuang, Q. Cancer-associated fibroblasts: from basic science to anticancer therapy. Exp. Mol. Med. 2023, 55(7), 1322–1332. [Google Scholar] [CrossRef] [PubMed]
- González, L.O.; Eiro, N.; Fraile, M.; et al. Prostate Cancer Tumor Stroma: Responsibility in Tumor Biology, Diagnosis and Treatment. Cancers Published. 2022, 14(18), 4412. [Google Scholar] [CrossRef] [PubMed]
- Pederzoli, F.; Raffo, M.; Pakula, H.; Ravera, F.; Nuzzo, P.V.; Loda, M. Stromal cells in prostate cancer pathobiology: friends or foes? Br. J. Cancer 2023, 128(6), 930–939. [Google Scholar] [CrossRef]
- Yedjou, C.G.; Mbemi, A.T.; Noubissi, F.; et al. Prostate Cancer Disparity, Chemoprevention, and Treatment by Specific Medicinal Plants. Nutrients Published. 2019, 11(2), 336. [Google Scholar] [CrossRef]
- Bai, B.; Chen, Q.; Jing, R.; et al. Molecular Basis of Prostate Cancer and Natural Products as Potential Chemotherapeutic and Chemopreventive Agents. Front Pharmacol. Published. 2021, 12, 738235. [Google Scholar] [CrossRef]
- Izzo, A.A.; Teixeira, M.; Alexander, S.P.H.; et al. A practical guide for transparent reporting of research on natural products in the British Journal of Pharmacology: Reproducibility of natural product research. Br. J. Pharmacol. 2020, 177(10), 2169–2178. [Google Scholar] [CrossRef]
- de Waure, C.; Bertola, C.; Baccarini, G.; Chiavarini, M.; Mancuso, C. Exploring the Contribution of Curcumin to Cancer Therapy: A Systematic Review of Randomized Controlled Trials. Pharmaceutics Published. 2023, 15(4), 1275. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Pezzani, R.; Redaelli, M.; et al. Preclinical Pharmacological Activities of Epigallocatechin-3-gallate in Signaling Pathways: An Update on Cancer. Molecules Published. 2020, 25(3), 467. [Google Scholar] [CrossRef]
- Ye, M.; Tian, H.; Lin, S.; et al. Resveratrol inhibits proliferation and promotes apoptosis via the androgen receptor splicing variant 7 and PI3K/AKT signaling pathway in LNCaP prostate cancer cells. Oncol. Lett. 2020, 20(5), 169. [Google Scholar] [CrossRef]
- Springer, M.; Moco, S. Resveratrol and Its Human Metabolites-Effects on Metabolic Health and Obesity. Nutrients Published. 2019, 11(1), 143. [Google Scholar] [CrossRef]
- Kogut, Ł.; Puchalski, C.; Katryńska, D.; Zaguła, G. The Multidirectional Biological Activity of Resveratrol: Molecular Mechanisms, Systemic Effects and Therapeutic Potential—A Review. Nutrients 2026, 18(2), 313. [Google Scholar] [CrossRef]
- Mokhtari, H.; Yaghmaei, B.; Sirati-Sabet, M.; et al. Epigallocatechin-3-gallate Enhances the Efficacy of MicroRNA-34a Mimic and MicroRNA-93 Inhibitor Co-transfection in Prostate Cancer Cell Line. Iran. J. Allergy Asthma Immunol. Published. 2020, 19(6), 612–623. [Google Scholar] [CrossRef]
- Fan, Y.; Wang, M.; Li, Z.; et al. Intake of Soy, Soy Isoflavones and Soy Protein and Risk of Cancer Incidence and Mortality. Front Nutr. Published. 2022, 9, 847421. [Google Scholar] [CrossRef]
- Holzapfel, N.P.; Holzapfel, B.M.; Champ, S.; Feldthusen, J.; Clements, J.; Hutmacher, D.W. The potential role of lycopene for the prevention and therapy of prostate cancer: from molecular mechanisms to clinical evidence. Int. J. Mol. Sci. Published. 2013, 14(7), 14620–14646. [Google Scholar] [CrossRef] [PubMed]
- de Molon, R.S.; Rodrigues, J.V.S.; de Avila, E.D.; et al. Do Adjunctive Therapies with Natural Products Improve Periodontal Clinical Parameters After Non-Surgical Treatment? A Systematic Review and Meta-Analysis. Int. J. Mol. Sci. Published. 2026, 27(5), 2394. [Google Scholar] [CrossRef]
- Zhou, M.; Liang, J.; Hui, J.; Xu, J. Inflammation-related indicators have a potential to increase overall quality of the prostate cancer management: a narrative review. Transl. Androl. Urol. 2023, 12(5), 809–822. [Google Scholar] [CrossRef] [PubMed]
- Merkens, L.; Sailer, V.; Lessel, D.; et al. Aggressive variants of prostate cancer: underlying mechanisms of neuroendocrine transdifferentiation. J. Exp. Clin. Cancer Res. Published. 2022, 41(1), 46. [Google Scholar] [CrossRef]
- Masi, D.; Watanabe, M.; Clément, K. Gut microbiome and obesity care: Bridging dietary, surgical, and pharmacological interventions. Cell Rep. Med. 2026, 7(2), 102573. [Google Scholar] [CrossRef] [PubMed]
- Nascimento-Gonçalves, E.; Azevedo, T.; Medeiros, C.; Faustino-Rocha, A.I. In Vivo Prostate Cancer Modelling: From the Pre-Clinical to the Clinical Setting. Life 2026, 16(1), 111. [Google Scholar] [CrossRef]
- Namekawa, T.; Ikeda, K.; Horie-Inoue, K.; Inoue, S. Application of Prostate Cancer Models for Preclinical Study: Advantages and Limitations of Cell Lines, Patient-Derived Xenografts, and Three-Dimensional Culture of Patient-Derived Cells. Cells Published. 2019, 8(1), 74. [Google Scholar] [CrossRef]
- Bergholz, J.S.; Zhao, J.J. How Compensatory Mechanisms and Adaptive Rewiring Have Shaped Our Understanding of Therapeutic Resistance in Cancer. Cancer Res. 2021, 81(24), 6074–6077. [Google Scholar] [CrossRef]
- Resurreccion, E.P.; Fong, K.W. The Integration of Metabolomics with Other Omics: Insights into Understanding Prostate Cancer. Metabolites Published. 2022, 12(6), 488. [Google Scholar] [CrossRef]
- Mizuno, K.; Beltran, H. Future directions for precision oncology in prostate cancer. Prostate 2022, 82 Suppl 1(Suppl 1), S86–S96. [Google Scholar] [CrossRef]
- Kumar, A.A. Prostate cancer genotyping for risk stratification and precision treatment. Curr. Urol. 2024, 18(2), 87–97. [Google Scholar] [CrossRef] [PubMed]
- Iweala, E.E.J.; Nwokoro, C.P. Molecular Drivers of Prostate Cancer Metastasis: Emerging Targets for Precision Therapy. Technol. Cancer Res. Treat. 2026, 25, 15330338251408321. [Google Scholar] [CrossRef] [PubMed]
- Kwon, W.A.; Joung, J.Y. Precision Targeting in Metastatic Prostate Cancer: Molecular Insights to Therapeutic Frontiers. Biomolecules Published. 2025, 15(5), 625. [Google Scholar] [CrossRef] [PubMed]
- Robaczyńska, J.; Maj, M.; Kiljańczyk, A.; Pastuszek, B.; Reducha, E.; Nurkiewicz, A.; Kiljańczyk, M. Epigenetic and Liquid Biopsy Biomarkers in Prostate Cancer: Bridging Tumor Heterogeneity and Clinical Implementation. Cancers 2026, 18(3), 389. [Google Scholar] [CrossRef]
- Zhao, H.; Wu, L.; Yan, G.; et al. Inflammation and tumor progression: signaling pathways and targeted intervention. Signal Transduct. Target Ther. Published. 2021, 6(1), 263. [Google Scholar] [CrossRef] [PubMed]
- Bautista, J.; Cardona-Maya, W.D.; Gancino-Guevara, K.; López-Cortés, A. Reprogramming prostate cancer through the microbiome. Front. Med. 2025, 12, 1690498. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Wang, L.; Hu, H.; Shieh, A.R.-E.; Li, E.; He, D.; He, L.; Liu, Z.; Paing, T.M.; Chen, X.; et al. Composition and Function of Gut Microbiome: From Basic Omics to Precision Medicine. Genes. 2026, 17(1), 116. [Google Scholar] [CrossRef]
- Dong, S.; Li, T. Metabolic reprogramming in cancer: signaling pathways and therapeutic targets. Mol. Cancer Published. 2026, 25(1), 60. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of the principal molecular pathways involved in prostate cancer progression and transition toward castration-resistant prostate cancer (CRPC). Central androgen receptor (AR) signaling interacts with PI3K/AKT/mTOR, MAPK (ERK), NF-κB, STAT3, oxidative stress, and metabolic reprogramming pathways to regulate proliferation, survival, invasion, angiogenesis, immune evasion, and therapeutic resistance. The figure additionally highlights adaptive mechanisms including AR amplification, AR splice variants, epigenetic remodeling, inflammatory signaling, and lineage plasticity that contribute to disease progression under androgen-deprivation pressure. Crosstalk among these interconnected pathways promotes transcriptional activation and supports progression toward advanced and therapy-resistant prostate cancer phenotypes.
Figure 1.
Schematic representation of the principal molecular pathways involved in prostate cancer progression and transition toward castration-resistant prostate cancer (CRPC). Central androgen receptor (AR) signaling interacts with PI3K/AKT/mTOR, MAPK (ERK), NF-κB, STAT3, oxidative stress, and metabolic reprogramming pathways to regulate proliferation, survival, invasion, angiogenesis, immune evasion, and therapeutic resistance. The figure additionally highlights adaptive mechanisms including AR amplification, AR splice variants, epigenetic remodeling, inflammatory signaling, and lineage plasticity that contribute to disease progression under androgen-deprivation pressure. Crosstalk among these interconnected pathways promotes transcriptional activation and supports progression toward advanced and therapy-resistant prostate cancer phenotypes.

Figure 2.
Schematic overview of the relationship between gut microbiome dynamics and prostate cancer progression. Dietary patterns, obesity, medications, and lifestyle-associated factors contribute to gut microbial dysbiosis characterized by reduced microbial diversity, expansion of pathobionts, loss of beneficial bacteria, and impaired intestinal barrier integrity. Dysbiotic microbial communities generate biologically active metabolites and components including short-chain fatty acids (SCFAs), bile acid derivatives, endotoxins (lipopolysaccharides; LPS), and indole-associated metabolites that influence host inflammatory signaling, immune modulation, androgen metabolism, oxidative stress, DNA damage, and epigenetic regulation. These interconnected pathways collectively contribute to tumor growth, invasion, metastasis, therapeutic resistance, and disease progression in prostate cancer. The figure additionally highlights modifiable factors including dietary interventions, probiotics, antibiotics, physical activity, and sleep that may influence microbiome composition and systemic host responses.
Figure 2.
Schematic overview of the relationship between gut microbiome dynamics and prostate cancer progression. Dietary patterns, obesity, medications, and lifestyle-associated factors contribute to gut microbial dysbiosis characterized by reduced microbial diversity, expansion of pathobionts, loss of beneficial bacteria, and impaired intestinal barrier integrity. Dysbiotic microbial communities generate biologically active metabolites and components including short-chain fatty acids (SCFAs), bile acid derivatives, endotoxins (lipopolysaccharides; LPS), and indole-associated metabolites that influence host inflammatory signaling, immune modulation, androgen metabolism, oxidative stress, DNA damage, and epigenetic regulation. These interconnected pathways collectively contribute to tumor growth, invasion, metastasis, therapeutic resistance, and disease progression in prostate cancer. The figure additionally highlights modifiable factors including dietary interventions, probiotics, antibiotics, physical activity, and sleep that may influence microbiome composition and systemic host responses.

Figure 3.
Illustration of selected natural products and their proposed molecular targets in prostate cancer. Curcumin, epigallocatechin gallate (EGCG), resveratrol, sulforaphane, genistein, quercetin, and lycopene influence multiple interconnected signaling pathways including androgen receptor (AR) signaling, PI3K/AKT/mTOR, MAPK, NF-κB, STAT3, oxidative stress pathways, inflammatory cytokines, apoptosis-associated signaling, and histone deacetylase (HDAC)-mediated epigenetic regulation. These multitarget interactions collectively contribute to anti-proliferative activity, induction of apoptosis, suppression of inflammation, reduction of oxidative stress, inhibition of invasion and metastasis, and modulation of therapeutic resistance. Additional proposed biological actions include modulation of gut microbiome composition, angiogenesis, mitochondrial metabolism, immune responses, and microRNA-associated signaling. The figure emphasizes the systems-level and multitarget nature of phytochemical activity in prostate cancer biology.
Figure 3.
Illustration of selected natural products and their proposed molecular targets in prostate cancer. Curcumin, epigallocatechin gallate (EGCG), resveratrol, sulforaphane, genistein, quercetin, and lycopene influence multiple interconnected signaling pathways including androgen receptor (AR) signaling, PI3K/AKT/mTOR, MAPK, NF-κB, STAT3, oxidative stress pathways, inflammatory cytokines, apoptosis-associated signaling, and histone deacetylase (HDAC)-mediated epigenetic regulation. These multitarget interactions collectively contribute to anti-proliferative activity, induction of apoptosis, suppression of inflammation, reduction of oxidative stress, inhibition of invasion and metastasis, and modulation of therapeutic resistance. Additional proposed biological actions include modulation of gut microbiome composition, angiogenesis, mitochondrial metabolism, immune responses, and microRNA-associated signaling. The figure emphasizes the systems-level and multitarget nature of phytochemical activity in prostate cancer biology.

Figure 4.
Integrated systems biology framework illustrating the interconnected relationship among diet, natural products, gut microbiome composition, microbial metabolites, epigenetic regulation, host signaling pathways, tumor microenvironment dynamics, and prostate cancer progression. Dietary phytochemicals and nutritional factors influence gut microbial diversity and microbial metabolic activity, resulting in the generation of biologically active metabolites including short-chain fatty acids (SCFAs), bile acids, indoles, trimethylamine N-oxide (TMAO), and lipopolysaccharides (LPS). These metabolites interact with epigenetic mechanisms including DNA methylation, histone modification, chromatin remodeling, and microRNA regulation, thereby influencing androgen receptor signaling, inflammatory pathways, oxidative stress, metabolic adaptation, and immune responses. The resulting host responses further interact with the tumor microenvironment through immune cell infiltration, cytokine signaling, angiogenesis, stromal remodeling, and extracellular matrix interactions, collectively contributing to tumor initiation, progression, metastasis, therapeutic resistance, and recurrence. The figure additionally highlights the potential role of natural products and microbiome-targeted interventions in restoring biological balance and improving therapeutic responsiveness through personalized systems-based strategies.
Figure 4.
Integrated systems biology framework illustrating the interconnected relationship among diet, natural products, gut microbiome composition, microbial metabolites, epigenetic regulation, host signaling pathways, tumor microenvironment dynamics, and prostate cancer progression. Dietary phytochemicals and nutritional factors influence gut microbial diversity and microbial metabolic activity, resulting in the generation of biologically active metabolites including short-chain fatty acids (SCFAs), bile acids, indoles, trimethylamine N-oxide (TMAO), and lipopolysaccharides (LPS). These metabolites interact with epigenetic mechanisms including DNA methylation, histone modification, chromatin remodeling, and microRNA regulation, thereby influencing androgen receptor signaling, inflammatory pathways, oxidative stress, metabolic adaptation, and immune responses. The resulting host responses further interact with the tumor microenvironment through immune cell infiltration, cytokine signaling, angiogenesis, stromal remodeling, and extracellular matrix interactions, collectively contributing to tumor initiation, progression, metastasis, therapeutic resistance, and recurrence. The figure additionally highlights the potential role of natural products and microbiome-targeted interventions in restoring biological balance and improving therapeutic responsiveness through personalized systems-based strategies.


Table 1.
Major Natural Products Implicated in Prostate Cancer Biology and Microbiome Modulation.
| Natural Product | Principal sources | Proposed molecular targets/pathways | Microbiome-associated effects | Potential relevance in prostate cancer | References |
|---|---|---|---|---|---|
| Curcumin | Curcuma longa | NF-κB, STAT3, AR signaling, PI3K/AKT, oxidative stress, DNMTs | Modulates microbial diversity; microbial metabolism may influence activity | Anti-inflammatory, epigenetic modulation, AR suppression, oxidative stress reduction | [52,54,56,59] |
| EGCG | Green tea | AR signaling, PI3K/AKT, MAPK, oxidative stress | Microbial conversion into phenolic metabolites | Antioxidant effects, suppression of proliferation and inflammatory signaling | [27,63,66] |
| Resveratrol | Grapes, berries, peanuts | NF-κB, STAT3, AMPK, mitochondrial signaling | Alters microbial composition and intestinal inflammatory signaling | Metabolic regulation, oxidative stress modulation, apoptosis induction | [27,68,69,71,72] |
| Sulforaphane | Cruciferous vegetables | HDAC inhibition, NRF2 activation, apoptosis signaling | Possible interaction with microbiome-derived metabolism | Epigenetic regulation and antioxidant defense enhancement | [74,75,76] |
| Quercetin | Fruits, onions, tea | PI3K/AKT, NF-κB, oxidative stress | May reduce intestinal oxidative stress and inflammatory signaling | Anti-inflammatory and antiproliferative effects | [27,98,110] |
| Genistein | Soy products | AR signaling, DNMTs, inflammatory signaling | Influenced by microbial metabolism of isoflavones | Endocrine modulation and epigenetic effects | [27,98,110] |
| Lycopene | Tomatoes | Oxidative stress pathways, inflammatory mediators | Possible interaction with diet–microbiome metabolism | Antioxidant and metabolic regulatory effects | [110,137] |
| Apigenin | Parsley, celery, chamomile | Cell cycle regulation, apoptosis pathways | Limited direct microbiome data | Potential antiproliferative activity | [27,110,128] |
| Luteolin | Vegetables, herbs | NF-κB, MAPK, inflammatory pathways | Limited direct microbiome evidence | Anti-inflammatory and antioxidant effects | [27,110,128] |
| Berberine | Berberis species | AMPK, glucose metabolism, inflammatory signaling | Alters microbial composition and bile acid metabolism | Metabolic and inflammatory pathway modulation | [51,86,93,110] |
Table 2.
Major Epigenetic Regulators and microRNAs in Prostate Cancer and Their Relationship with Natural Products.
Table 2.
Major Epigenetic Regulators and microRNAs in Prostate Cancer and Their Relationship with Natural Products.
| Epigenetic regulator / microRNA | Biological role in prostate cancer | Associated pathways | Natural products reported to influence pathway | Proposed functional impact | References |
|---|---|---|---|---|---|
| GSTP1 hypermethylation | Loss of antioxidant defense | Oxidative stress, DNA damage | Curcumin, genistein | Partial restoration of tumor suppressor expression | [27,52,54,56,59,98,110] |
| DNMT1 / DNMT3A | Maintenance of aberrant DNA methylation | Epigenetic silencing | Curcumin, EGCG, genistein | Reduction of methyltransferase activity | [27,52,54,56,59,63,66,98,110] |
| HDACs | Chromatin repression, resistance biology | AR signaling, apoptosis, EMT | Sulforaphane, butyrate | Increased chromatin accessibility | [27,42,74,75,76] |
| EZH2 | Lineage plasticity, metastasis | Polycomb repression, stemness | Curcumin, resveratrol | Potential reduction in aggressive transcriptional programs | [27,52,54,56,59,68,69,71,72] |
| microRNA-21 | Oncogenic signaling | PI3K/AKT, inflammatory pathways | Curcumin, resveratrol | Reduced inflammatory and survival signaling | [27,52,54,56,59,68,69,71,72] |
| microRNA-34a | Tumor suppressive regulation | Apoptosis, EMT inhibition | Curcumin | Promotion of apoptosis and reduced invasiveness | [52,54,56,59] |
| microRNA-200 family | EMT suppression | Cell adhesion and migration | Curcumin, genistein | Reduced metastatic potential | [27,52,54,56,59,98,110] |
| AR-associated enhancer remodeling | Castration resistance | Chromatin accessibility, AR transcription | Sulforaphane, EGCG | Potential modulation of AR transcriptional output | [27,63,66,74,75,76] |
| Butyrate-mediated HDAC inhibition | Epigenetic regulation via microbiome | Histone acetylation | Dietary fiber-associated microbiome activity | Modulation of inflammatory and differentiation pathways | [47,77,82] |
Table 3.
Molecular Pathways Linking the Gut Microbiome, Epigenetic Regulation, and Prostate Cancer Progression.
Table 3.
Molecular Pathways Linking the Gut Microbiome, Epigenetic Regulation, and Prostate Cancer Progression.
| Biological component | Mechanistic contribution | Downstream effects in prostate cancer | References |
|---|---|---|---|
| Gut microbial dysbiosis | Increased inflammatory signaling and endotoxin exposure | NF-κB activation, oxidative stress, chronic inflammation | [22,23,24,25,32,41,45] |
| Short-chain fatty acids | HDAC inhibition and chromatin remodeling | Altered gene expression and immune regulation | [17,23,100] |
| Microbial androgen metabolism | Steroid precursor conversion | Sustained AR signaling and endocrine resistance | [16,26] |
| Oxidative stress pathways | DNA damage and inflammatory amplification | Genomic instability and progression | [16,17,18,21,31] |
| PI3K/AKT pathway activation | Survival signaling and metabolic adaptation | Resistance to apoptosis and therapy | [15,16,21,23] |
| STAT3 and NF-κB signaling | Inflammatory transcriptional programs | Tumor proliferation and immune suppression | [15,16,23,101] |
| EZH2 overexpression | Epigenetic silencing and lineage plasticity | Aggressive disease behavior | [36,96,100] |
| microRNA dysregulation | Post-transcriptional signaling disruption | EMT, metastasis, apoptosis resistance | [103,104,105,106] |
| Tumor microenvironment remodeling | Stromal and immune adaptation | Angiogenesis, immune escape, invasion | [112,114,117] |
Table 4.
Translational and Clinical Studies of Natural Products in Prostate Cancer.
| Natural product | Study type | Principal findings | Proposed mechanisms | Major limitations | References |
|---|---|---|---|---|---|
| Curcumin | Clinical pilot studies, adjunctive therapy investigations | Changes in inflammatory biomarkers and PSA kinetics in selected studies | NF-κB inhibition, oxidative stress reduction, AR modulation | Poor bioavailability, small cohorts, heterogeneous formulations | [52,54,56,59] |
| EGCG / Green tea catechins | Epidemiological and interventional studies | Mixed findings regarding prostate cancer risk and progression | Antioxidant effects, AR suppression, PI3K/AKT modulation | Variability in dose, metabolism, dietary exposure | [27,62,63,66] |
| Resveratrol | Early translational studies | Modulation of metabolic and inflammatory biomarkers | AMPK signaling, mitochondrial regulation, oxidative stress modulation | Rapid metabolism and low systemic exposure | [27,68,69,71,72] |
| Genistein | Pre-surgical and dietary intervention studies | Potential effects on proliferation and endocrine signaling | AR modulation, epigenetic regulation | Limited sample size and variable intake | [27,98,110] |
| Lycopene | Observational and dietary studies | Possible association with reduced risk in some populations | Antioxidant and metabolic effects | Difficulty isolating dietary variables | [110,137] |
| Sulforaphane | Early-phase translational studies | Modulation of HDAC-associated signaling | Epigenetic regulation, NRF2 activation | Limited clinical outcome data | [74,75,76] |
| Quercetin | Mainly preclinical evidence | Anti-inflammatory and antiproliferative effects | Oxidative stress modulation, PI3K/AKT inhibition | Limited human data | [27,98,110] |
| Pomegranate extracts | Pilot clinical studies | Potential slowing of PSA progression in selected cohorts | Antioxidant and anti-inflammatory pathways | Lack of definitive oncological endpoints | [27,110] |
Table 5.
Major Challenges and Knowledge Gaps in Natural Product-Based Prostate Cancer Research.
| Challenge area | Key problem | Scientific/translational implication | References |
|---|---|---|---|
| Tumor heterogeneity | Diverse molecular and metabolic phenotypes | Variable response to phytochemicals | [16,150] |
| Microbiome variability | Differences due to diet, age, obesity, medications | Inconsistent metabolite generation and responsiveness | [22,23,24,25,32,41,45] |
| Poor bioavailability | Limited absorption and rapid metabolism | Reduced clinical efficacy despite strong preclinical activity | [27,32,90] |
| Experimental model limitations | Cell lines and animal models may not reflect human disease complexity | Reduced translational predictability | [32,33,41,143] |
| Multitarget biological effects | Simultaneous modulation of multiple pathways | Difficulty identifying dominant therapeutic mechanisms | [27,33,51,61,62,63,110] |
| Lack of standardized formulations | Variable purity and extraction methods | Poor reproducibility across studies | [27,32,33,48,73,83] |
| Limited clinical endpoints | Reliance on surrogate biomarkers such as PSA | Uncertain impact on survival outcomes | 27, 32, 33, 48, 73, 128, 138 |
| Insufficient multi-omics integration | Lack of integrated microbiome, epigenetic, and metabolic profiling | Incomplete mechanistic understanding | [16,18,33,59,84] |
| Drug–phytochemical interactions | Potential pharmacokinetic and metabolic interference | Safety and efficacy concerns | [27,32,33,84,93] |
| Limited long-term human data | Short follow-up duration in most studies | Uncertain preventive and therapeutic relevance | [127, 32, 33, 93, 110 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



