Submitted:
09 June 2026
Posted:
11 June 2026
You are already at the latest version
Abstract
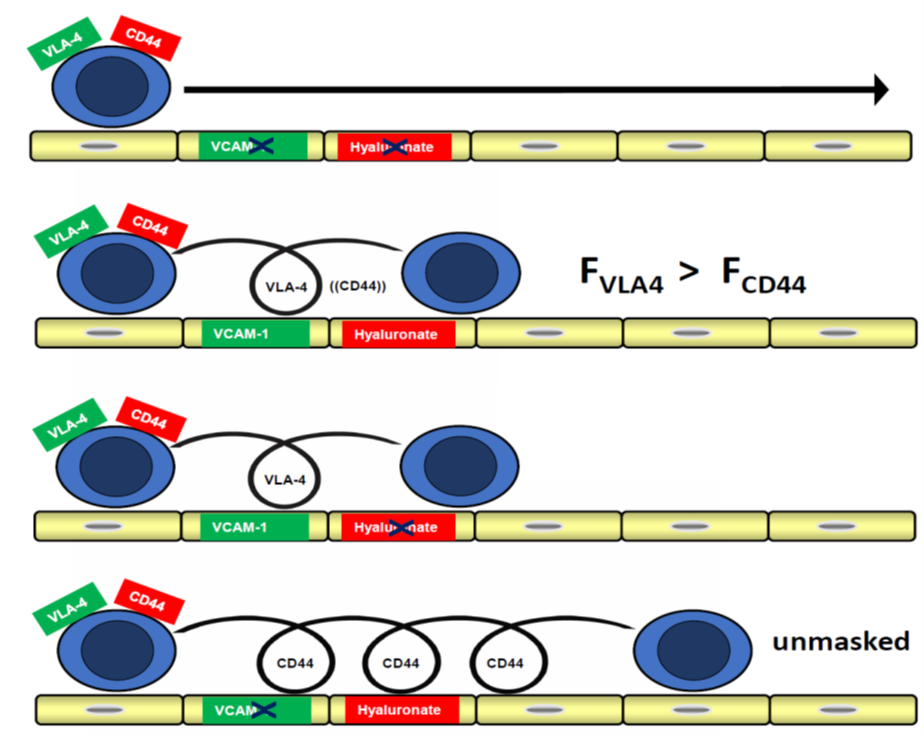
Leukocyte recruitment from blood into tissues involves sequential adhesive steps, including rolling and integrin-dependent arrest. VLA-4 can support firm adhesion and, in some settings, rolling interactions, whereas CD44–hyaluronan interactions have also been implicated in leukocyte rolling. Here, we used adhesion assays and parallel-plate flow chamber experiments to analyze CD44–hyaluronan-dependent monocyte interactions on ECV304 monolayers and to compare them with α4-integrin-sensitive adhesion on endothelial monolayers. WEHI 78/24 monocytoid cells interacted with ECV304 monolayers in a CD44- and hyaluronan-dependent manner, whereas adhesion to HMEC-1 and bEnd.3 monolayers was sensitive to α4-integrin blockade. Blocking CD44, adding soluble hyaluronan, or treating ECV304 monolayers with hyaluronidase reduced adhesion and rolling. Mixed primary human monocyte preparations also showed CD44-dependent adhesion and rolling on ECV304 monolayers. ECV304 cells are interpreted here not as endothelial cells, but as T24-derived, hyaluronidase-sensitive cellular monolayers useful for functional analysis of CD44–hyaluronan-dependent interactions. These findings support a substrate-dependent functional hierarchy in which CD44–hyaluronan-dependent monocyte rolling becomes detectable when α4-integrin-dependent adhesion is not dominant, while emphasizing the cell-model-based nature of the assay.
Keywords:
1. Introduction
2. Results
2.1. Distinct α4-Integrin– and CD44-Dependent Adhesion Pathways Are Revealed on Different Cellular Monolayers
2.2. Rolling Interactions on ECV304 Monolayers Are Weaker than on bEnd.3 Monolayers
2.3. CD44 Mediates Rolling and Arrest of WEHI 78/24 Cells on ECV304 Monolayers via Hyaluronan
2.4. WEHI 78/24 Cells Bind Soluble Hyaluronan via CD44
2.5. Surface-Associated Hyaluronan on ECV304 Monolayers Supports Adhesion of WEHI 78/24 Cells
2.6. Primary Human Monocytes Use CD44 to Bind Hyaluronan-Presenting ECV304 Monolayers
2.7. Primary Human Monocytes Also Roll on ECV304 Monolayers via CD44 Under Low Shear
3. Discussion
4. Materials and Methods
4.1. Cells and Reagents
4.2. Monolayer Activation
4.3. Hyaluronidase Treatment
4.4. Flow Cytometry
4.5. Static and Shaking Adhesion Assays
4.6. Monocyte–Hyaluronate Binding Assay
4.7. Laminar Flow Assays
4.8. Statistics
4.9. Study Approval and Human Samples
4.10. AI Tool Disclosure
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Butcher, E.C. Leukocyte-endothelial cell recognition: Three (or more) steps to specificity and diversity. Cell 1991, 67, 1033-1036. [CrossRef]
- Springer, T.A. Traffic signals for lymphocyte recirculation and leukocyte emigration: the multistep paradigm. Cell 1994, 76, 301-314. [CrossRef]
- Sher, B.T.; Bargatze, R.; Holzmann, B.; Gallatin, W.M.; Matthews, D.; Wu, N.; Picker, L.; Butcher, E.C.; Weissman, I.L. Homing receptors and metastasis. Adv Cancer Res 1988, 51, 361-390. [CrossRef]
- Gosslar, U.; Jonas, P.; Luz, A.; Lifka, A.; Naor, D.; Hamann, A.; Holzmann, B. Predominant role of alpha 4-integrins for distinct steps of lymphoma metastasis. Proceedings of the National Academy of Sciences 1996, 93, 4821-4826. [CrossRef]
- Altevogt, P.; Hubbe, M.; Ruppert, M.; Lohr, J.; von Hoegen, P.; Sammar, M.; Andrew, D.P.; McEvoy, L.; Humphries, M.J.; Butcher, E.C. The alpha 4 integrin chain is a ligand for alpha 4 beta 7 and alpha 4 beta 1. J Exp Med 1995, 182, 345-355. [CrossRef]
- Aigner, S.; Ruppert, M.; Hubbe, M.; Sammar, M.; Sthoeger, Z.; Butcher, E.C.; Vestweber, D.; Altevogt, P. Heat stable antigen (mouse CD24) supports myeloid cell binding to endothelial and platelet P-selectin. Int Immunol 1995, 7, 1557-1565. [CrossRef]
- Hubbe, M.; Kowitz, A.; Schirrmacher, V.; Schachner, M.; Altevogt, P. L1 adhesion molecule on mouse leukocytes: regulation and involvement in endothelial cell binding. Eur J Immunol 1993, 23, 2927-2931. [CrossRef]
- Aruffo, A.; Stamenkovic, I.; Melnick, M.; Underhill, C.B.; Seed, B. CD44 is the principal cell surface receptor for hyaluronate. Cell 1990, 61, 1303-1313. [CrossRef]
- DeGrendele, H.C.; Estess, P.; Picker, L.J.; Siegelman, M.H. CD44 and its ligand hyaluronate mediate rolling under physiologic flow: a novel lymphocyte-endothelial cell primary adhesion pathway. J Exp Med 1996, 183, 1119-1130. [CrossRef]
- Goldstein, L.A.; Zhou, D.F.; Picker, L.J.; Minty, C.N.; Bargatze, R.F.; Ding, J.F.; Butcher, E.C. A human lymphocyte homing receptor, the hermes antigen, is related to cartilage proteoglycan core and link proteins. Cell 1989, 56, 1063-1072. [CrossRef]
- Stamenkovic, I.; Amiot, M.; Pesando, J.M.; Seed, B. A lymphocyte molecule implicated in lymph node homing is a member of the cartilage link protein family. Cell 1989, 56, 1057-1062. [CrossRef]
- Campbell, J.J.; Hedrick, J.; Zlotnik, A.; Siani, M.A.; Thompson, D.A.; Butcher, E.C. Chemokines and the arrest of lymphocytes rolling under flow conditions. Science 1998, 279, 381-384. [CrossRef]
- Geissmann, F.; Manz, M.G.; Jung, S.; Sieweke, M.H.; Merad, M.; Ley, K. Development of monocytes, macrophages, and dendritic cells. Science 2010, 327, 656-661. [CrossRef]
- Ley, K.; Laudanna, C.; Cybulsky, M.I.; Nourshargh, S. Getting to the site of inflammation: the leukocyte adhesion cascade updated. Nat Rev Immunol 2007, 7, 678-689. [CrossRef]
- Martin-Padura, I.; Lostaglio, S.; Schneemann, M.; Williams, L.; Romano, M.; Fruscella, P.; Panzeri, C.; Stoppacciaro, A.; Ruco, L.; Villa, A.; et al. Junctional adhesion molecule, a novel member of the immunoglobulin superfamily that distributes at intercellular junctions and modulates monocyte transmigration. J Cell Biol 1998, 142, 117-127. [CrossRef]
- Merad, M.; Martin, J.C. Pathological inflammation in patients with COVID-19: a key role for monocytes and macrophages. Nat Rev Immunol 2020, 20, 355-362. [CrossRef]
- Schneemann, M.; Schoedon, G.; Linscheid, P.; Walter, R.; Blau, N.; Schaffner, A. Nitrite generation in Interleukin-4--treated human macrophage cultures does not involve the nitric oxide synthase pathway. Journal of Infectious Diseases 1997, 175, 130-135. [CrossRef]
- Xu, H.; Manivannan, A.; Liversidge, J.; Sharp, P.F.; Forrester, J.V.; Crane, I.J. Involvement of CD44 in leukocyte trafficking at the blood-retinal barrier. J Leukoc Biol 2002, 72, 1133-1141.
- Hollingsworth, J.W.; Li, Z.; Brass, D.M.; Garantziotis, S.; Timberlake, S.H.; Kim, A.; Hossain, I.; Savani, R.C.; Schwartz, D.A. CD44 regulates macrophage recruitment to the lung in lipopolysaccharide-induced airway disease. Am J Respir Cell Mol Biol 2007, 37, 248-253. [CrossRef]
- Eibl, R.H. Chemokine-Independent VLA-4/VCAM-1-Mediated Rolling and Arrest of B16 Melanoma Cells Under Shear. Int J Mol Sci 2026, 27. [CrossRef]
- Jutila, M.A.; Lewinsohn, D.M.; Berg, E.L.; Butcher, E.C. Homing Receptors in Lymphocyte, Neutrophil, and Monocyte Interaction with Endothelial Cells. 1990, 1990; pp. 227-235.
- Jain, R.K. Determinants of tumor blood flow: a review. Cancer Res 1988, 48, 2641-2658.
- Jain, R.K. Transport of molecules, particles, and cells in solid tumors. Annu Rev Biomed Eng 1999, 1, 241-263. [CrossRef]
- Jain, R.K. Normalizing tumor vasculature with anti-angiogenic therapy: a new paradigm for combination therapy. Nat Med 2001, 7, 987-989. [CrossRef]
- Maniotis, A.J.; Folberg, R.; Hess, A.; Seftor, E.A.; Gardner, L.M.; Pe’er, J.; Trent, J.M.; Meltzer, P.S.; Hendrix, M.J. Vascular channel formation by human melanoma cells in vivo and in vitro: vasculogenic mimicry. Am J Pathol 1999, 155, 739-752. [CrossRef]
- Shirakawa, K.; Kobayashi, H.; Heike, Y.; Kawamoto, S.; Brechbiel, M.W.; Kasumi, F.; Iwanaga, T.; Konishi, F.; Terada, M.; Wakasugi, H. Hemodynamics in vasculogenic mimicry and angiogenesis of inflammatory breast cancer xenograft. Cancer Res 2002, 62, 560-566.
- Seftor, R.E.; Hess, A.R.; Seftor, E.A.; Kirschmann, D.A.; Hardy, K.M.; Margaryan, N.V.; Hendrix, M.J. Tumor cell vasculogenic mimicry: from controversy to therapeutic promise. Am J Pathol 2012, 181, 1115-1125. [CrossRef]
- Bao, X.; Ran, J.; Kong, C.; Wan, Z.; Wang, J.; Yu, T.; Ruan, S.; Ding, W.; Xia, L.; Zhang, D. Pan-cancer analysis reveals the potential of hyaluronate synthase as therapeutic targets in human tumors. Heliyon 2023, 9, e19112. [CrossRef]
- Paget, S. The distribution of secondary growths in cancer of the breast. The Lancet 1889, 133, 571-573. [CrossRef]
- Gunthert, U.; Hofmann, M.; Rudy, W.; Reber, S.; Zoller, M.; Haussmann, I.; Matzku, S.; Wenzel, A.; Ponta, H.; Herrlich, P. A new variant of glycoprotein CD44 confers metastatic potential to rat carcinoma cells. Cell 1991, 65, 13-24. [CrossRef]
- Eibl, R.H.; Pietsch, T.; Moll, J.; Skroch-Angel, P.; Heider, K.H.; von Ammon, K.; Wiestler, O.D.; Ponta, H.; Kleihues, P.; Herrlich, P. Expression of variant CD44 epitopes in human astrocytic brain tumors. Journal of Neuro-Oncology 1995, 26, 165-170. [CrossRef]
- Chambers, A.F.; Groom, A.C.; MacDonald, I.C. Dissemination and growth of cancer cells in metastatic sites. Nat Rev Cancer 2002, 2, 563-572. [CrossRef]
- Montesano, R.; Pepper, M.S.; Möhle-Steinlein, U.; Risau, W.; Wagner, E.F.; Orci, L. Increased proteolytic activity is responsible for the aberrant morphogenetic behavior of endothelial cells expressing the middle T oncogene. Cell 1990, 62, 435-445. [CrossRef]
- Takahasi, K.; Sawasaki, Y. Rare spontaneously transformed human endothelial cell line provides useful research tool. In Vitro Cell Dev Biol - Animal 1992, 28, 380-382. [CrossRef]
- Dirks, W.G.; MacLeod, R.A.; Drexler, H.G. ECV304 (endothelial) is really T24 (bladder carcinoma): cell line cross- contamination at source. In Vitro Cell Dev Biol Anim 1999, 35, 558-559. [CrossRef]
- Gil’iano, N.I.; Semenova, E.G.; Fedortseva, R.F.; Konevega, L.V. [Characteristics of the spontaneously transformed human endothelial cell line ECV304. II. Functional responses of the ECV304 cells]. Tsitologiia 2008, 50, 576-584.
- Brown, J.; Reading, S.J.; Jones, S.; Fitchett, C.J.; Howl, J.; Martin, A.; Longland, C.L.; Michelangeli, F.; Dubrova, Y.E.; Brown, C.A. Critical evaluation of ECV304 as a human endothelial cell model defined by genetic analysis and functional responses: a comparison with the human bladder cancer derived epithelial cell line T24/83. Lab Invest 2000, 80, 37-45. [CrossRef]
- Xiong, W.-C.; Simon, S. ECV304 Cells: An Endothelial or Epithelial Model? J Biol Chem 2011, 286, le21. [CrossRef]
- Ades, E.W.; Candal, F.J.; Swerlick, R.A.; George, V.G.; Summers, S.; Bosse, D.C.; Lawley, T.J. HMEC-1: Establishment of an Immortalized Human Microvascular Endothelial Cell Line. Journal of Investigative Dermatology 1992, 99, 683-690. [CrossRef]
- Walker, E.B.; Lanier, L.L.; Warner, N.L. Characterization and functional properties of tumor cell lines in accessory cell replacement assays. J Immunol 1982, 128, 852-859.
- de Belder, A.N.; Wik, K.O. Preparation and properties of fluorescein-labelled hyaluronate. Carbohydrate Research 1975, 44, 251-257. [CrossRef]
- Jalkanen, S.; Bargatze, R.F.; de los Toyos, J.; Butcher, E.C. Lymphocyte recognition of high endothelium: antibodies to distinct epitopes of an 85-95-kD glycoprotein antigen differentially inhibit lymphocyte binding to lymph node, mucosal, or synovial endothelial cells. The Journal of Cell Biology 1987, 105, 983-990. [CrossRef]
- Kishimoto, T.K.; Warnock, R.A.; Jutila, M.A.; Butcher, E.C.; Lane, C.; Anderson, D.C.; Smith, C.W. Antibodies Against Human Neutrophil LECAM-1 (LAM-l/Leu-8/DREG-56 antigen) and Endothelial Cell ELAM-1 Inhibit a Common CD18-Independent Adhesion Pathway In Vitro. Blood 1991, 78, 805-811. [CrossRef]
- Gallatin, W.M.; Weissman, I.L.; Butcher, E.C. A cell-surface molecule involved in organ-specific homing of lymphocytes. Nature 1983, 304, 30-34. [CrossRef]
- Miyake, K.; Weissman, I.L.; Greenberger, J.S.; Kincade, P.W. Evidence for a role of the integrin VLA-4 in lympho-hemopoiesis. Journal of Experimental Medicine 1991, 173, 599-607. [CrossRef]
- Streeter, P.R.; Berg, E.L.; Rouse, B.T.N.; Bargatze, R.F.; Butcher, E.C. A tissue-specific endothelial cell molecule involved in lymphocyte homing. Nature 1988, 331, 41-46. [CrossRef]
- Ledbetter, J.A.; Herzenberg, L.A. Xenogeneic Monoclonal Antibodies to Mouse Lymphoid Differentiation Antigens*. Immunological Reviews 1979, 47, 63-90. [CrossRef]
- Lawrence, M.B.; McIntire, L.V.; Eskin, S.G. Effect of Flow on Polymorphonuclear Leukocyte/Endothelial Cell Adhesion. Blood 1987, 70, 1284-1290. [CrossRef]
- Lawrence, M.B.; Smith, C.W.; Eskin, S.G.; McIntire, L.V. Effect of Venous Shear Stress on CD18-Mediated Neutrophil Adhesion to Cultured Endothelium. Blood 1990, 75, 227-237. [CrossRef]
- Lawrence, M.B.; Springer, T.A. Leukocytes roll on a selectin at physiologic flow rates: Distinction from and prerequisite for adhesion through integrins. Cell 1991, 65, 859-873. [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



