Submitted:
30 April 2026
Posted:
30 April 2026
You are already at the latest version
Abstract
Scorpion venom peptides, with their stable disulphide backbone, compact structural framework, and highly selective regulation of ion channels, have long been regarded as important molecular probes in neuropharmacology. However, recent studies have revealed their potential for regulating oxidative stress, anti-inflammation, and neuroprotection, making them a new research frontier. This article focuses on “scorpion venom peptides as drugs,” constructing an integrated knowledge framework from structural classification to clinical translation. First, scorpion venom peptides are systematically classified based on cysteine arrangement patterns and three-dimensional folding topology, and their structure-activity relationships are summarised. Based on this, the molecular mechanisms by which scorpion venom peptides regulate ion channels are systematically analysed. In addition, we review the emerging pharmacological activities of scorpion venom peptides. Of particular note, the representative molecule SVHRSP has shown multi-target synergistic antioxidant and neuroprotective activity in models of Parkinson’s disease. This article also systematically evaluates the application of engineering strategies, including cyclisation modification, nanodelivery, recombinant expression, and AI-assisted optimisation, to overcome the translational bottlenecks in the development of scorpion venom peptides. By integrating molecular structure, redox regulation mechanisms, and translational medicine perspectives, this review aims to provide a theoretical basis and practical pathways for scorpion venom peptides as precision therapeutic molecules for oxidative stress-related diseases.
Keywords:
scorpion venom peptides
; oxidative stress
; Nrf2-ARE pathway
; NADPH oxidase (NOX2)
; neuroprotection
1. Introduction
Animal venom is an important source of bioactive molecules for structural optimisation in modern drug development, and some venom-derived compounds have inspired the development of clinically applicable drugs [1]. Among these natural resources, scorpion venom has attracted much attention due to its diverse range of peptide compounds with high targeting specificity, compact conformation, and potent pharmacological activity. In particular, scorpion venom peptides possess a stable disulphide backbone, well-defined ion channel affinity, and significant functional diversity, making them ideal templates for peptide-based drug development [2]. Scorpion venom peptides are secreted by specialised venom glands and are typically small molecules with molecular weights between 1 and 10 kDa. Many scorpion venom peptides are rich in cysteine, and their three-dimensional conformation is stabilised by multiple disulphide bonds; others are linear, non-disulphide peptides with amphiphilic or membrane-active properties. These structural features endow scorpion venom peptides with a variety of biological functions, including regulating ion channels, recognising membrane-associated proteins, disrupting microbial membranes, and modulating inflammatory signalling pathways [3,4].
Historically, research on scorpion venom peptides has primarily focused on neurotoxicology and ion channel pharmacology. Many classic scorpion venom peptides have become important molecular probes for elucidating the structure and gating mechanisms of voltage-gated sodium (NaV), potassium (KV), and calcium (CaV) ion channels. In recent years, advances in venomomics, transcriptomics, and structural biology have greatly expanded this field, revealing that scorpion venom peptides are not limited to classic neurotoxins but also have broader therapeutic potential for pain management, cancer diagnosis and treatment, anti-infective therapy, immunomodulation, and intervention in oxidative stress-related diseases [5,6].
Of particular interest is the emerging role of scorpion venom peptides in oxidative stress-related diseases. Oxidative stress is a crucial pathological basis for Parkinson’s disease, Alzheimer’s disease, epilepsy, cerebral ischemia-reperfusion injury, and tumor progression, characterised by excessive reactive oxygen species (ROS), mitochondrial dysfunction, and persistent activation of inflammatory signals. Previous studies have shown that some scorpion venom peptides can exert cytoprotective effects by inhibiting NADPH oxidase (NOX2) activation, reducing ROS accumulation, blocking the NF-κB/NLRP3 inflammatory axis, and improving mitochondrial homeostasis. This suggests that scorpion venom peptides are not only ion channel ligands but may also be multifunctional therapeutic molecules with both membrane-targeting regulatory and antioxidant activities [7,8].
Despite these advances, the clinical translation of scorpion venom peptides remains challenging. Key limitations include insufficient subtype selectivity, limited metabolic stability, poor tissue delivery, potential immunogenicity, and high manufacturing costs [9]. Therefore, a systematic understanding of their structural principles, molecular mechanisms, and engineering solutions is crucial for the development of therapeutics [10].
While some reviews have summarized certain toxin families or therapeutic applications, significant knowledge gaps remain. First, most previous articles primarily categorized toxins by molecular target (e.g., NaV or Kv toxins), with limited cross-family comparisons based on conserved cysteine skeletons, thus obscuring shared and differentiated recognition strategies among different peptides [11]. Furthermore, conversion barriers such as poor in vivo stability, immunogenicity risks, and limited blood-brain barrier penetration of scorpion venom peptides have not been fully assessed [12,13]. Moreover, recent advances in AI-assisted structure prediction and synthetic biology-supported production have not been fully integrated into the current review framework.
Based on this, this review systematically summarizes the research progress of scorpion venom peptides, focusing on the “structural scaffold, functional mechanism, translational application” framework. This article will focus on discussing the structural diversity of major peptide families and their molecular interactions with targets such as NaV, Kv, and TRP. It will further elucidate their emerging pharmacological activities in areas such as antioxidation, anti-inflammation, antitumor, anti-infection, and neuroprotection, and evaluate engineering strategies such as cyclization modification, nanodelivery, recombinant expression, and artificial intelligence optimization. By integrating perspectives from molecular pharmacology, oxidative stress biology, and translational medicine, this article aims to provide a theoretical foundation and practical pathway for the precise development of scorpion venom-derived peptide drugs.
2. Structural Classification and Molecular Architecture
The structural diversity of scorpion venom peptides underlies their functional specificity. Based on the arrangement of cysteine residues and disulphide bond linkages, scorpion venom peptides can be systematically classified into several structural families, among which the CSα/β (cysteine-stabilised α-helix/β-sheet), CSα/α, and CSβ families constitute the core architecture of the venom group [14] (Figure 1). These compact structural motifs endow venoms with high thermal stability, proteolytic resistance, and evolutionary adaptability, making them common templates for venom diversification. In addition to these disulphide-rich peptides, scorpion venom also contains linear non-disulphide peptides with membrane activity or antibacterial properties. These peptide families together constitute a structurally coherent yet functionally diverse group of venom peptides. This section summarises the main classification systems of scorpion venom peptides, their characteristic three-dimensional folded structures, and the latest advances in structural biology, which have deepened our understanding of the skeleton-function relationship (Table 1).
2.1. Cysteine-Based Scaffold Classification System
2.1.1. CSα/β Family: Classic α-Helix-β-Fold Double-Chain Structure
The CSα/β sheet is the most widely distributed structural motif in scorpion venom peptides. It is typically stabilised by four disulphide bonds formed by eight cysteine residues and consists of a tightly packed α-helix and a triple-stranded antiparallel β-sheet [14]. This scaffold structure is dominant in sodium channel toxins and contains two main functional groups: α-scorpion venom (α-NaTxs) and β-scorpion venom (β-NaTxs).
α-NaTxs bind to receptor site 3 of voltage-gated sodium (NaV) channels, delaying rapid inactivation and thus prolonging the duration of sodium current. A representative member is AaH II from the Australian scorpion (Androctonus australis). Conversely, β-NaTxs bind to receptor site 4, shifting channel activation towards a more hyperpolarised potential, thereby altering the voltage threshold for channel opening; CsVII from the gorilla scorpion (Centruroides sculpturatus) is a classic example. Although the pharmacological effects of α- and β-toxins differ, crystallographic and NMR studies indicate that they share a conserved CSα/β core structure, with functional differences primarily depended by surface electrostatic interactions and conformational changes in the exposed ring that participate in receptor recognition. Recent studies have further highlighted the adaptability of this scaffold. The newly discovered β-toxins Chirp7 and Chirp9 from Centruroides hirsutipalpus interact with the S3-S4 paddle motif of NaV channels, providing new structural templates for the development of channel-selective probes and modulators [15].
2.1.2. CSα/α Family: Potassium Channel Toxins
The CSα/α family comprises α-KTx peptides. These toxins typically contain a C-C-CC-C-C cysteine sequence folded into two short, stable α-helices linked by disulphide bonds. Depending on the cyclic structure and helical orientation, the resulting topology is described as “two-finger” or “three-finger” shaped. Members of this family selectively block the outer vestibule of voltage-gated potassium (Kv) channels, and more than 30 sequence-based subfamilies have been identified [14]. Scorpion venom (α-KTx1.1) was one of the first scorpion potassium channel toxins to be structurally characterised, providing a basic model for pore blockage in which Lys27 protrudes into the channel pore to prevent ion permeation [16]. Subsequent studies have shown that this family possesses broader functional diversity. Agitoxin-2 (α-KTx3.1) can bind to the surfaces of multiple extracellular channels simultaneously, while BmTx3 (α-KTx6.1) is selective for Kv4 channel subtypes [17].
Besides their value as pharmacological tools, α-KTx scaffolds also have growing translational applications. A recently discovered Kv1.3 blocker, Meuk7-3, isolated from Mesobuthus crucittii, showed improved affinity and developability after computer optimization, indicating the potential application of CSα/α peptides in immunomodulatory drug design [14]. Similarly, Cvill6 and Cvill7 from Centruroides villegasi exhibit high activity against the Kv1.2 channel, with Cvill7 showing picomolar activity and significant subtype selectivity, supporting the potential of this family in treating channel disorders such as epilepsy [18].
2.1.3. CSβ Family of Non-Disulphide Bonded Linear Peptides
The CSβ family is represented by chlortoxin, a compact peptide stabilised by a C-C-C-C framework of four cysteine residues. Its overall folding consists primarily of three antiparallel β-sheets, lacking the typical α-helix. This structure enables it to form high-affinity interactions with membrane-associated targets, including matrix metalloproteinase-2 (MMP-2) and annexin A2, which explains its long-standing interest as a glioma-targeting ligand in imaging and drug delivery applications [19]. The structural diversity of chlortoxin-like peptides continues to expand. In 2025, researchers identified a novel chlortoxin-like insecticidal peptide, Ce1, from the venom of the Egyptian scorpion (Compsobuthus egyptiensis). Ce1 consists of 36 amino acid residues and four disulphide bonds, but unlike typical chlortoxins, it employs a mixed folding structure consisting of one α-helix and three antiparallel β-sheets. This discovery broadens the known structural spectrum of the CSβ family and provides a new evolutionary solution for selective predatory toxicity [20]. In addition to disulphide-rich toxins, scorpion venom also contains a large number of cysteine-free linear peptides. A typical example is Pandinin 1 from the emperor scorpion (Pandinus imperator), which exerts its antibacterial activity by disrupting microbial cell membranes through the formation of amphiphilic α-helices. A recent study comparing Pandinin 1, Pandinin 2, and Pandinin 3 against multidrug-resistant Klebsiella pneumoniae showed that Pandinin 1 and Pandinin 2 produced rapid bactericidal effects within one hour, with minimum inhibitory concentrations of 6-25 μM. Mechanistic analysis indicated that Pandinin 1, Pandinin 2, and Pandinin 3 directly interact with lipopolysaccharides in the bacterial outer membrane [21]. Short-chain linear scorpion venom peptides typically contain 9-19 amino acid residues and are attracting increasing attention due to their ease of synthesis and relatively low production costs. These molecules, along with disulphide-rich toxins, demonstrate two complementary evolutionary strategies in scorpion venom: broad-spectrum membrane defence and highly selective receptor and highly selective targeting of receptors and ion channels.
2.2. Three-Dimensional Structural Features and Dynamic Properties
2.2.1. Conformational Constraints of Disulphide Bonds
Disulphide bonds are key determinants of the folding and stability of scorpion venom peptides. In the classic cysteine-stabilised α/β (CSα/β) motif, two disulphide bonds link the α-helix to the C-terminal antiparallel β-sheet, forming the most common compact scaffold in scorpion venom. A complete CSα/β fold typically contains three disulphide bonds, at least two of which anchor the helix to the β-sheet, forming a rigid hydrophobic core. Comparative studies have shown that this scaffold is not unique to scorpions and is also conserved in insect defensins and plant γ-sulfur proteins, highlighting its evolutionary success as a functional template [22]. However, disulphide bond connections are not absolutely conserved. Saucedo et al. described two potassium channel-blocking peptides from the scorpion Tityus that retain the typical six-cysteine spacing but employ different pairing patterns. This rearrangement is accompanied by a complete recombination of secondary structures, forming a cysteine-stabilised helix-loop-helix fold, revealing unexpected structural plasticity in the venom repertoire [23]. Atypical pairing patterns have also been reported in some centruroid sodium channel toxins, further expanding the known structural map.
A complete disulphide bond network also significantly contributes to thermostability. The degradation-derived peptide BmK86-P1 isolated from medicinal scorpion material retains its native fold over a wide temperature range (20-95 °C), indicating strong thermostability, which is closely related to the preservation of disulphide bond connections [24]. Mutational analysis of leucine toxin I further shows that the loss of key disulphide bonds reduces α-helix content and bioactivity. Although two native bridges still maintain a near-native folded structure, all three disulphide bonds are essential for complete thermodynamic stability [22].
2.2.2. Functional Surfaces and Molecular Recognition
Scorpion venom recognition of ion channels depends on finely arranged surface properties. For sodium channel toxins, α-toxins and β-toxins bind to different receptor sites: α-toxins interact with receptor site 3 and inhibit rapid inactivation, while most β-toxins bind to receptor site 4 and shift voltage-dependent activation to a more hyperpolarised potential [25]. Electrostatic analysis shows that many sodium channel toxins exhibit asymmetric charge distributions. Positively charged regions (typically with theoretical isoelectric points above 8.5) promote initial attraction to negatively charged residues in voltage-sensing domains or extracellular loops, while hydrophobic regions stabilise subsequent docking. Crystal structure analysis of Ts1 revealed a single-sided cluster of residues, previously confirmed by chemical modification and mutagenesis, defining a possible channel recognition surface where charged and nonpolar side chains jointly determine selectivity [26].
2.2.3. Post-Translational Modifications Expand Chemical Diversity
Scorpion peptides undergo various post-translational modifications (PTMs), significantly broadening their chemical and functional range. C-terminal amidation is one of the most common modifications, crucial for both ion channel toxins and antimicrobial peptides. Transcriptomic analysis of the venom glands identified a dual-enzyme α-amidation system consisting of a membrane-bound bifunctional peptidylglycine α-amidation monooxygenase and an associated soluble enzyme, providing the molecular basis for scorpion peptide amidation [27]. Functionally, C-terminal amidation enhances the activity of various toxins. In the BmKAS of Mesobuthus martensii, amidation increases its affinity for insect voltage-gated sodium channels [28]. A similar phenomenon was observed in the Ts1 of Tityus serrulatus, where amidation affects its biological activity and proteolytic stability [29]. N-terminal pyroglutamylation is another important post-translational modification. Ts1 (also known as γ-toxin) is a typical example. Although pyroglutamylation is less frequent in scorpions than in cone snails or snake venom, it can protect the N-terminus from exopeptidases and regulate its interaction with targets [30]. A review of engineered scorpion antimicrobial peptides further indicates that C-terminal amidation and specific proline substitutions can enhance protease resistance and stabilize amphiphilic helical structures, while hydroxyproline may enhance local conformational preference [31].
2.3. Advances in Structural Biology Methodology
2.3.1. X-Ray Crystallography and Nuclear Magnetic Resonance Spectroscopy
X-ray crystallography remains the primary method for determining high-resolution toxin structures. The Protein Data Bank (PDB) had indexed approximately 1,200 scorpion venom peptide structures, about one-third of which had a resolution better than 2.0 Å [32]. Nuclear magnetic resonance spectroscopy has unique advantages in capturing conformational dynamics in solution. Recent ¹⁵N relaxation experiments revealed the picosecond to nanosecond timescale motion of the scorpion venom peptide BmKK2 in free state and membrane-simulated environments, confirming conformational exchange behavior in its functional ring region [33]. This dynamic information provides crucial atomic-level evidence for understanding the initial events of toxin-membrane interactions.
2.3.2. The Cryo-Electron Microscopy Structural Revolution
Since the “resolution revolution” of 2017, cryo-electron microscopy has become a core technology for resolving the structure of scorpion venom peptide-membrane protein complexes. In 2020, Clairfeuille et al. resolved the complex structure of α-scorpion venom LqhIII and the NaV1.5 channel at a resolution of 3.3 Å, demonstrating for the first time at the atomic level how α-scorpion venom binds to the S3-S4 linker region of the voltage-sensing domain IV, exerting its effect by hindering channel inactivation. In 2021, Wisedchaisri et al., by designing a NaVAb/NaV1.7 chimeric channel, resolved the structural details of the tarantula venom Huwentoxin-IV, capturing the resting state of NaV1.7 at a resolution of 2.8 Å, revealing that key lysine residues in the venom act as “stingers” penetrating deep into the S3-S4 linker region of the voltage receptor, locking the channel in a resting state [34]. These works validate and extend the gated modified toxin action model proposed in electrophysiological studies, providing an atomic-level template for the rational design of ion channel-targeted drugs.
2.3.3. Computational Structural Biology and Artificial Intelligence
Molecular dynamics simulations effectively compensate for the lack of dynamic information in static experimental structures. All-atom molecular dynamics (MD) simulations revealed the stepwise insertion behavior of the scorpion venom peptide ADWX-1 during its binding to the Kv1.3 channel, predicting the transient interactions between aromatic residues of the toxin and the hydrophobic ring in the channel pore region, dynamic details difficult to capture directly through crystallography [35]. The release of AlphaFold2 in 2021 revolutionized the scorpion venom peptide structure prediction paradigm: its prediction confidence (pLDDT > 90) for the CSαβ and CSαα family toxin backbones approached the precision of NMR experiments [36]. In 2024, AlphaFold3 further achieved end-to-end prediction of toxin-protein complexes, but its confidence in predicting the extracellular loop region of channels with multiple glycosylation sites remains low, a key region for scorpion venom peptide binding. Therefore, its applicability to ion channel complexes containing multiple transmembrane helices still needs systematic validation with experimental data [37].
2.4. An Integrated Perspective on Structure-Function Relationships
The CSαβ and CSαα scaffolds of scorpion venom peptides exhibit profound modularity in both evolution and function. Although they share a similar disulphide bond core, they achieve distinctly different target recognition strategies through differentiated combinations of secondary structures: the CSαβ scaffold utilizes the functional surface recognition channel between the α-helix and β-sheet, while the CSαα scaffold mainly relies on the spatial configuration of the double α-helix and the connecting loop region to function. The first insecticidal peptide with a CSαα scaffold, LaIT5, identified in 2024 from the scorpion Liocheles australasiae, has a double α-helix and two consecutive histidine residues that are crucial for its insecticidal activity and are non-toxic to mammals, demonstrating the potential of the CSαα scaffold as a template for the development of novel biopesticides [38]. At the molecular engineering level, customized functional modification has been successfully achieved through domain transplantation between different CSαβ toxins. In 2025, Montero-Dominguez et al. grafted the domain of CeII8 from non-lethal β-scorpion venom onto the backbone of the lethal β-scorpion venom toxin CssII, yielding the chimeric peptide rCssII-Del-D23A-TCD. This variant retains affinity for NaV1.7 but loses toxicity toward NaV1.6 and exhibits analgesic activity comparable to the μ-opioid receptor agonist DAMGO in inflammatory and neuropathic pain models [39]. In the future, the deep integration of cryo-electron microscopy, artificial intelligence prediction, and high-throughput mutation screening is expected to accelerate the transformation of scorpion venom peptides from natural molecules into precision therapeutic tools targeting specific ion channel diseases.
3. Ion Channel Modulation: Mechanisms and Selectivity
The main pharmacological value of scorpion venom peptides lies in their ability to regulate ion channels with high affinity and, in many cases, exhibit significant subtype selectivity. Voltage-gated sodium channels (NaV), voltage-gated potassium channels (KV), and transient receptor potential (TRP) channels are core regulators of electrical signal transduction in excitable tissues and are therefore major molecular targets of scorpion venom [2,3]. Studies of these peptide-channel interactions not only elucidate the toxin’s mechanism of action but also provide valuable templates for designing therapeutic ligands (Figure 2).
3.1. Voltage-Gated Sodium Channel Regulation Mechanism
Scorpion venom peptides are the most representative regulators of NaV channels, mainly divided into α-toxins and β-toxins. Both share the CSα/β backbone, but their sites of action and gating effects differ. α-toxins primarily bind to the extracellular S3–S4 loop (receptor site 3) of the NaV channel DIV domain, restricting the outward movement of the DIV voltage sensor S4, delaying rapid inactivation and prolonging channel open time, leading to sustained sodium ion influx and membrane depolarization. AaH II, LqhαIT, and Ts3 are classic examples, with Ts3 significantly reducing gating charge movement, thereby further confirming its function by restricting voltage-sensor movement [41,42]. In contrast, β-toxins primarily bind to the S3–S4 region of the DII domain (receptor site 4), stabilizing the voltage sensor in an activated conformation, shifting the channel activation curve towards hyperpolarization, and lowering the opening threshold, thereby enhancing sodium current at lower membrane potentials [43]. Although the two types of toxins have different mechanisms, both can increase neuronal excitability, which is a key molecular basis for scorpion sting pain and neurotoxicity [11].
NaV1.7 is the most studied subtype in pain management; loss-of-function mutations can lead to congenital analgesia, whereas gain-of-function mutations can cause hereditary pain syndromes [44]. Therefore, scorpion venom-derived NaV1.7 modulators are considered important lead molecules for non-opioid analgesics. OD1 can efficiently target NaV1.7, but also affects NaV1.4 and NaV1.6, suggesting that high selectivity between homologous isoforms remains a bottleneck in development [39,45]. Recently, the N18W variant was obtained through targeted modification of key sites in DKK2, which enhanced analgesic activity while reducing its effect on NaV1.4 and NaV1.5, demonstrating that the therapeutic window can be significantly improved through limited residue optimization based on the conserved CSα/β backbone [46].
3.2. Regulation of Voltage-Gated Potassium Channels
AKv channels are another class of targets that have been studied most extensively by scorpion venom peptides, especially the α-KTx family. Its classic mechanism is “external pore blockade”: the functional lysine side chain on the toxin’s surface is inserted into the outer vestibule of the channel and approaches the selectivity filter, thereby directly blocking K⁺ permeation. Structural studies show that this process usually does not significantly alter the overall conformation of the filter, but substituting key lysine residues significantly reduces affinity and accelerates dissociation, which is also an important reason why many scorpion venom peptides have nanomolar to picomolar potency [40,47]. In addition, not all Kv toxins rely on pore blockade mechanisms. Some κ-KTx peptides stabilize the non-conductive state of the channel by binding to the extracellular surface and adjacent gating domains, thereby achieving allosteric inhibition. HsTx1, a representative Kv1.3 inhibitor, possesses picomolar activity and has become an important template for optimizing immunomodulatory peptides [40].
Kv subtype selectivity arises from the fine complementarity of the toxin-channel interface. The protonation state of vestibular histidine residues outside Kv1.3 modulates margatoxin affinity, while a single amino acid substitution can significantly alter the selectivity of charybdotoxin analogs for Kv1.2 [48,49]. CboK7 (α-KTx2.24) from Centruroides bonito also exhibits a significantly higher affinity for Kv1.2 than for Kv1.3 [50]. From a transformational perspective, Kv1.3 is involved in the activation of effector memory T cells and macrophages, and is closely related to inflammation and oxidative stress. BmKK2 inhibition of Kv1.3 can downregulate NF-κB/NLRP3 signaling, suggesting that scorpion venom peptides have anti-inflammatory and immunomodulatory potential [51].
3.3. Other Ion Channel Targets
Besides NaV and KV, scorpion venom peptides can also act on many other channels. The high-conductivity calcium-activated potassium channel BK (KCa1.1) is one of the classic targets. Iberi toxin (IbTX, α-KTx1.3) inhibits K⁺ conductance by binding to the outer pore region; its potency is affected by the composition of the accessory subunits and the cellular environment. BK channels containing the β4 subunit show significantly decreased sensitivity to IbTX [52,53,54]. In terms of sensory transduction, TRPV1 is an important receptor for thermal and chemical pain. BmP01 from the East Asian pincers scorpion is the first scorpion venom peptide confirmed to directly activate TRPV1 and induce an acute pain response, suggesting that scorpion sting pain originates not only from aberrant NaV activation but also involves the TRP pathway [11,55].
Furthermore, the acid-sensitive ion channel ASIC1a is involved in acidosis and ischemic nerve injury. Although the classic inhibitory peptide PcTx1 is derived from spider venom, its mechanism of binding to the inter-subunit interface and stabilizing a non-conductive conformation provides an important comparative model for understanding how venom peptides achieve highly selective inhibition through extracellular allosteric changes [56]. These studies indicate that the regulation of ion channels by venom peptides is far more than a single-pore-blocking mode; it encompasses multiple mechanisms, including gating capture, allosteric stabilization, and auxiliary subunit-dependent regulation.
3.4. Integrated Determinants of Selectivity
The high selectivity of scorpion venom peptides stems from the synergistic effect of multiple structural factors. First, surface electrostatic complementarity determines initial recognition efficiency. Most NaV-active toxins are strongly alkaline, which is more conducive to binding to negatively charged voltage-sensing regions, whereas Kv toxins typically have a more balanced charge distribution, making them more suitable for recognition in the outer pore region [57]. Second, the local flexibility of the exposed loop region endows the toxin with induced-fit ability, enabling it to adapt to highly homologous but subdivided channel subtypes [58]. Furthermore, the membrane lipid environment can influence the orientation, accumulation, and contact efficiency of toxins on the membrane surface, especially for gated toxins [59].
Overall, the CSα/β and CSα/α scorpion venom peptide scaffolds demonstrate how a finite structural framework can evolve into diverse recognition strategies. In the future, combining cryo-electron microscopy, electrophysiology, computational design, and directed evolution technologies will further promote the transformation of scorpion venom peptides from molecular probes into highly selective analgesics, immunomodulators, and anti-inflammatory drug candidates.
4. Beyond Ion Channels: Emerging Targets and Pharmacological Profiles
Although scorpion venom peptides were initially thought to be ion channel modulators, their pharmacological spectrum has now been shown to be much broader. Recent studies have found that scorpion venom peptides have activity in areas such as pain control, tumor treatment, antibacterial therapy, and neuroprotection. In particular, thermostable peptides from Mesobuthus martensii and their synthetic derivatives have shown protective effects in experimental models of Parkinson’s disease, Alzheimer’s disease, and epilepsy, suggesting that scorpion venom peptides can act simultaneously on inflammation, oxidative stress, mitochondrial homeostasis, and glial cell signaling networks, rather than being limited to excitatory membrane channels [60,61,62] (Figure 3).
4.1. Analgesic Mechanisms and Targets
Scorpion venom remains an important natural resource for the development of novel analgesics. NaV1.7, NaV1.8, and NaV1.9 are closely associated with congenital analgesia, erythromelalgia, and paroxysmal pain syndrome, thereby making them key targets for non-opioid analgesics. Active peptides in scorpion venom and other arachnid venoms provide abundant templates for the design of highly selective analgesic ligands [63]. Mechanistically, α-NaTx prolongs channel opening time by delaying inactivation, while β-NaTx lowers the activation threshold; both enhance the excitability of nociceptive neurons and pain transmission [11]. Current research has shifted from broad-spectrum NaV regulation to optimizing NaV1.7 subtype selectivity, aiming to reduce off-target risks to the heart and skeletal muscle while maintaining analgesic activity [45]. Structurally, most analgesic-related scorpion venom peptides retain the CSα/β backbone, while surface charge distribution and active ring conformation determine their subtype preference.
4.2. Anticancer Mechanisms and Targets
Chlortoxin is one of the most representative molecules in scorpion venom antitumor research. This CSβ family peptide is rich in β-sheet content and can recognize glioma-related molecules, such as MMP-2 and annexin A2, with high affinity. After binding, it induces complex internalization, disrupts Ca²⁺ homeostasis, and remodels the cytoskeleton, thereby inhibiting tumor cell migration and invasion [64]. Based on its tumor-targeting properties, chlortoxin has been widely used in brain tumor delivery systems, improving nanocarriers’ ability to cross the blood-brain barrier and the efficiency of lesion accumulation, and has been extended to fluorescence-guided surgery, multimodal imaging, radionuclide delivery, siRNA transport, and chlortoxin-guided CAR-T therapy [19]. In addition to chlortoxin, many other scorpion venom peptides also exhibit direct cytotoxicity. Smp24 can induce mitochondrial apoptosis in HepG2 cells and inhibit tumor growth in vivo [65]. Other venom components may exert their effects through tumor immune remodeling. For example, the F1 component of the European scorpion (Mesobuthus eupeus) can transform IL-4-polarized M2 macrophages into an M1-like phenotype, and the conditioned medium of treated macrophages can inhibit the proliferation and migration of CT-26 colon cancer cells [66]. These findings suggest that scorpion venom peptides may possess both direct tumor-killing and immunomodulatory effects. Recent reviews also indicate that oncology is one of the most promising translational research directions in scorpion venom pharmacology [67].
4.3. Antimicrobial Activity
Scorpion venom contains a large number of non-disulfide-linked antimicrobial peptides, which are increasingly seen as alternatives to traditional antibiotics. Many of these peptides possess broad-spectrum antimicrobial activity and may exert less selective pressure for resistance than classic small-molecule drugs. Their emergence has greatly expanded the pharmacological properties of scorpion venom beyond ion channel regulation [3,67]. A systematic review published in 2022 summarized the structural diversity of scorpion venom peptides with antimicrobial, antifungal, and antiparasitic properties and concluded that these molecules are valuable templates for the design of next-generation anti-infective drugs [68]. Most antimicrobial scorpion venom peptides are linear, cysteine-free molecules, such as the pandinine family. Unlike disulphide-rich neurotoxins, these peptides typically form amphiphilic α-helices upon contact with cell membranes and disrupt microbial envelopes through carpet-like or barrel-like mechanisms. Pandinin 1 and Pandinin 2 have been reported to increase bacterial membrane permeability and induce intracellular leakage [2,69]. Recent reviews further indicate that scorpion venom peptides may also inhibit fungi and certain viruses, highlighting their potential as multifunctional anti-infective agents [67].
4.4. Neuroprotective Effects
Among recently identified scorpion venom-derived molecules, SVHRSP has emerged as a representative multi-target neuroprotective peptide. Current evidence suggests its beneficial effects primarily manifest in three interrelated pathways (Figure 3). First, SVHRSP inhibits NLRP3 inflammasome-mediated neuroinflammation in microglia. Studies by Zhang et al. have shown that this peptide inhibits inflammasome assembly and caspase-1 activation, thereby reducing the maturation and release of IL-1β and related cytokines. Pharmacological inhibition or gene knockout of NLRP3 significantly attenuates this protective effect, indicating that this pathway is a major downstream target [70]. A review on venom peptides regulating the NLRP3 inflammasome also highlights the anti-inflammatory potential of scorpion venom peptides [71]. Second, SVHRSP attenuates oxidative stress and the TLR4/NF-κB signaling pathway. This peptide has been reported to remodel gut microbiota composition and reduce circulating and intracranial LPS/HMGB1 levels, thereby limiting activation of the TLR4/NF-κB pathway in the brain [72]. Simultaneously, it can inhibit NOX2 assembly, reduce reactive oxygen species production, and enhance the activity of endogenous antioxidant enzymes [73,74]. Furthermore, SVHRSP may regulate unconventional ion channel targets in glial cells. Studies have shown that it can reduce Nav1.6 expression in microglia, inhibit sodium current and intracellular Ca²⁺ accumulation, and attenuate activation of the MAPK pathway [75]. In summary, these findings support SVHRSP as a multi-mechanism neuroprotective lead compound and elucidate how scorpion peptides can be applied to the treatment of complex neurological diseases (Figure 4).
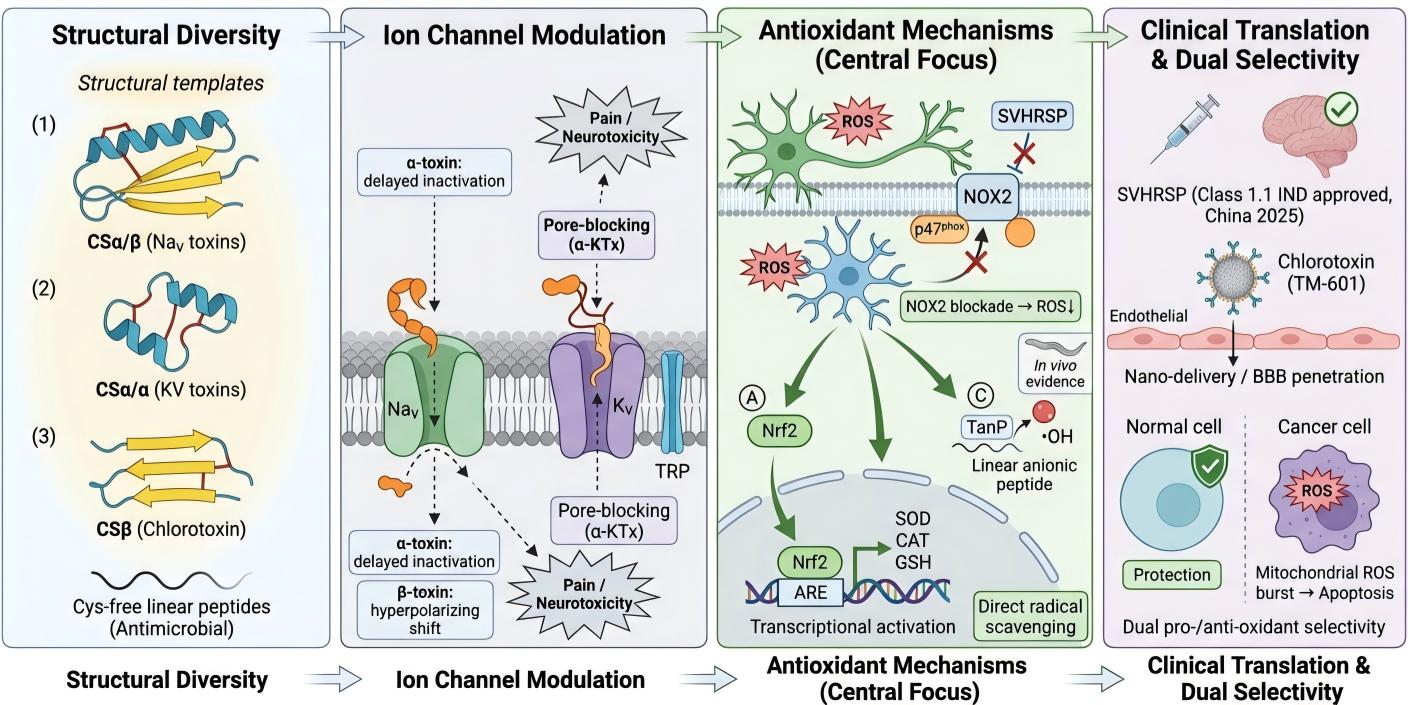
4.5. Antioxidant Mechanism of Scorpion Venom Peptides
In recent years, the antioxidant activity of scorpion venom peptides has evolved from a co-effect of ion channel regulation to an independent research direction. Existing evidence suggests that scorpion venom peptides exert their antioxidant effects through multiple pathways, including activating the Nrf2-ARE signaling axis, targeting and inhibiting NADPH oxidase (NOX2), regulating the endogenous antioxidant enzyme system, and directly scavenging free radicals, forming a multi-level, multi-target oxidative stress regulatory network [72].
In the Nrf2-ARE pathway, the representative synthetic peptide SVHRSP, in an Alzheimer’s disease model exacerbated by PM2.5 exposure, alleviated cognitive impairment by upregulating Nrf2 expression and inhibiting endoplasmic reticulum stress and neuronal pyroptosis. SVHRSP can also alleviate PM2.5 exposure-induced neuronal necrosis and apoptosis by regulating the long non-coding RNA Gm6410 [76]. Furthermore, in a Caenorhabditis elegans model, SVHRSP reduced ROS levels, increased SOD-3 activity, and upregulated the expression of antioxidant-related genes such as ctl-1, egl-1, and cat-2, thereby further validating the ability of scorpion venom peptides to activate endogenous antioxidant defense systems across species [77]. These findings indicate that scorpion venom peptides can establish a systemic defense against oxidative stress at the transcriptional level.
In terms of NADPH oxidase targeting inhibition, SVHRSP demonstrated precise regulation of NOX2 in a Parkinson’s disease model. Mechanistic studies showed that SVHRSP reduces reactive oxygen species (ROS) generation at its source by blocking the translocation of the cytoplasmic subunit p47phox to the cell membrane, thereby inhibiting the assembly and activation of the NOX2 holoenzyme. Knockdown of NOX2 expression via siRNA significantly weakened the inhibitory effects of SVHRSP on LPS- and rotenone-induced pro-inflammatory cytokine gene expression and related neurotoxicity, validating NOX2 as its core target [78].
Scorpion venom peptides can also regulate the activity of endogenous antioxidant enzyme systems. Smp24 not only inhibited tumor growth in a mouse model of solid Ehrlich ascites carcinoma but also significantly increased the levels of superoxide dismutase (SOD), catalase (CAT), and glutathione (GSH), while decreasing malondialdehyde (MDA) and nitric oxide (NO) levels [79]. Furthermore, some linear anionic peptides (such as TanP) can directly scavenge free radicals, and their effects are independent of intracellular signaling pathways, providing new insights for developing structurally concise antioxidant candidate molecules [80].

Table 2.
Pharmacological activities and clinical translation status of representative scorpion venom peptides.
Table 2.
Pharmacological activities and clinical translation status of representative scorpion venom peptides.
| Mechanism Type | Peptide | Target /Pathway |
Key Experimental Evidence | Citations |
|---|---|---|---|---|
| Nrf2-ARE pathway activation | SVHRSP | Nrf-2; p38 MAPK; Lnc Gm6410 |
Upregulates Nrf-2 expression in PM2.5-exposed AD model, alleviating ER stress and neuronal pyroptosis; Regulates Lnc Gm6410 to mitigate necroptosis; Reduces ROS, increases SOD-3 activity, and upregulates ctl-1, egl-1, cat-2 expression in C. elegans | [77] |
| NOX2 targeted inhibition | SVHRSP | NOX2; p47phox membrane translocation |
Blocks membrane translocation of p47phox, preventing NOX2 assembly and activation; NOX2 siRNA knockdown significantly attenuates anti-inflammatory and neuroprotective effects of SVHRSP | [78] |
| Endogenous antioxidant enzyme regulation | Smp24 | SOD; CAT; GSH; MDA; NO | Increases SOD, CAT and GSH levels while reducing MDA and NO content in solid-Ehrlich carcinoma mouse model | [69] |
| Direct free radical scavenging | TanP | Free radical direct scavenging | Scavenges free radicals independent of intracellular signaling pathways | [67] |
| Pro-oxidant / antioxidant dual selectivity | S6540, Androcin 18-1 | Mitochondrial ROS; PI3K/Akt | Induces excessive mitochondrial ROS generation and triggers apoptosis in tumor cells; tends to inhibit oxidative stress in normal cells | [61,81] |
It is noteworthy that the relationship between scorpion venom peptides and reactive oxygen species is not a one-way “inhibition.” In tumor cells, some scorpion venom peptides (such as S6540 and Androcin 18-1) induce apoptosis by promoting excessive mitochondrial ROS production, exhibiting dual selectivity for “pro-oxidation-antioxidation.” This difference may stem from differences in membrane potential, metabolic state, and surface charge distribution between normal and tumor cells, providing a theoretical basis for the targeted application of scorpion venom peptides in tumor therapy [61,81]. In summary, the antioxidant mechanisms of scorpion venom peptides can be attributed to three main pathways: transcriptional activation of the Nrf2-ARE pathway, targeted inhibition of ROS-generating enzymes such as NOX2, and direct free-radical scavenging by linear peptides. Elucidating these mechanisms not only expands the pharmacological scope of scorpion venom peptides but also provides new molecular tools and therapeutic strategies for the precise intervention of oxidative stress-related diseases (such as neurodegenerative diseases, inflammation-related injuries, and tumors).
5. Peptide Engineering and Therapeutic Development
5.1. Molecular Optimization and Delivery Strategies
Despite the high affinity and subtype selectivity of natural scorpion venom peptides for ion channel targets, their development as therapeutics has long been hampered by three major bottlenecks: rapid proteolytic degradation, potential immunogenicity, and limited tissue penetration, particularly the difficulty in crossing the blood-brain barrier (BBB). Recent advances in peptide engineering, drug delivery, and synthetic biotechnology have begun to address these obstacles (Table 3).
Improving metabolic stability is a primary goal. Cyclic skeletalization and terminal modification are commonly used methods. Cyclic analogs of BmKTX significantly prolonged resistance to enzymatic degradation while maintaining potent Kv1.3 inhibitory activity, demonstrating that conformational restriction can maintain biological activity while reducing protease sensitivity [82]. Introducing non-natural amino acids provides a complementary strategy. Replacing key residues of scorpion venom with D-amino acid analogs significantly improved its plasma stability in non-human primates and prolonged its in vivo half-life, while maintaining immunosuppressive efficacy [83]. Half-life extension can also be achieved through fusion engineering. Integrating engineered scorpion venom peptide motifs into Fc-based multivalent constructs yielded a selective Kv1.3 inhibitor with prolonged systemic exposure and sustained inhibition of IL-2 secretion in cynomolgus monkey T cells, supporting the platform’s potential for treating autoimmune diseases [84].
Delivery innovation is particularly important for central nervous system applications. Chlorine-functionalized nanoparticles have attracted significant attention for glioblastoma treatment because chlorine can promote receptor-mediated trans-blood-brain-barrier transport while maintaining tumor-targeting specificity [85]. Physical modulation of the blood-brain barrier (BBB) is another option: focused ultrasound combined with microbubbles can transiently and reversibly increase BBB permeability in specific brain regions, thereby enhancing peptide delivery [86]. Non-invasive receptor-mediated transport strategies also show promising potential. Angiopep-2, which can cross the BBB via LRP1-mediated endocytosis, has been proposed as a carrier to increase the exposure of peptide drugs such as chlortoxin conjugates in the central nervous system [87]. Pharmacokinetic studies have further confirmed that SVHRSP can cross the blood-brain barrier and be detected in cerebrospinal fluid following systemic administration [88]. Biomimetic carriers, such as exosomes, may offer advantages including low immunogenicity and good tissue penetration. In glioma models, drug-loaded exosomes achieved selective intracranial accumulation, supporting their future use in delivering scorpion peptides [89].
5.2. Clinical Translation and Patent Landscape
Among scorpion venom peptide-derived drugs, synthetic chlortoxin TM-601 has the most mature clinical application. Early-phase I/II clinical trials used intracavitary injection of 131I-TM-601 to treat patients with recurrent high-grade gliomas, demonstrating good tolerability, limited systemic toxicity, and a relatively long tumor retention time after local administration [90]. Subsequent studies increasingly emphasize its value as a targeted ligand in imaging and payload delivery, rather than as an independent cytotoxic drug.
In analgesic drug development, NaV1.7-selective peptide inhibitors remain a focus of research. A review summarized engineering strategies for converting venom peptides into NaV1.7 antagonists and emphasized the importance of scaffold-specific optimization [91]. However, due to the high structural homology of NaV channel subtypes, the risk of off-target effects on cardiac or skeletal muscle sodium channels is increased, making translational applications still very difficult. This issue is not purely theoretical: a recent study of a spider venom-derived NaV1.7 inhibitor found that it exhibited unexpected activity against Kv4.2/4.3 channels, accompanied by cardiotoxicity, highlighting the need for more extensive selectivity analysis during lead compound optimization [91].
Patent activity over the past decade indicates growing interest in scorpion peptide therapies, particularly for treating neurological disorders. Among these, the thermostable peptide derivative SVHRSP, derived from the East Asian scorpion, has accumulated a relatively complete intellectual property portfolio, covering extraction methods, synthetic peptide composition, and therapeutic uses in epilepsy, Parkinson’s disease, and Alzheimer’s disease. Based on this platform, SVHRSP injections are expected to receive regulatory approval in China in 2025, entering clinical evaluation as a Class 1.1 innovative drug for rapid control of epileptic seizure clusters. This milestone marks the increasing maturity of scorpion peptide drug translational development.
6. Challenges and Future Perspectives
Despite significant progress in scorpion venom peptide research, several challenges remain in transitioning them from laboratory to clinical applications. One major issue stems from the rapid development of omics-based discovery technologies. High-throughput sequencing and integrated proteomics-transcriptomics workflows can now identify hundreds of putative venom components from a single species. For example, a combined transcriptomic and proteomic analysis of Centruroides possanii identified 244 full-length transcripts, including 109 homologous to ion-channel toxins [92]. However, the biological functions of many predicted peptides remain unknown. Therefore, establishing standardized workflows for high-throughput functional annotation, target identification, and early safety screening is crucial. A second challenge is druggability. Natural peptides are often readily proteolytic and may exhibit limited pharmacokinetic exposure. While cyclization, sequence minimization, and the introduction of non-natural residues can improve stability, these modifications may also introduce novel epitopes and increase immunogenicity risk. MHC-related peptide proteomics has recently become an effective method for preclinical immunogenicity assessment of biologics and can be incorporated into scorpion peptide lead compound optimization schemes earlier [93].
The inherent multi-target properties of many scorpion peptides present both opportunities and challenges. Off-target activities across ion channel subtypes or irrelevant receptors may narrow the therapeutic window. Conversely, multi-target pharmacology may be advantageous in complex diseases. SVHRSP is one example: reported mechanisms include inhibition of NADPH oxidase-dependent oxidative stress [94], inhibition of NLRP3 inflammasome-driven neuroinflammation [70], and regulation of the microglial Nav1.6 signaling pathway [75]. This regulation of synergistic pathways may be particularly relevant to neurodegenerative diseases.
Emerging technologies are expected to accelerate the progress of related research. Structural prediction tools can now simulate the folding structures of various toxins and toxin-target interaction interfaces with considerable accuracy [95,96]. Experimental benchmark tests show that the predicted structures are in close agreement with the resolved structures of representative venom peptides, such as chlortoxin and melitin-related backbones [97]. Generative design platforms, including ProteinMPNN and diffusion-based methods, are gradually enabling the de novo synthesis of target-binding peptides [98]. The integration of prediction, automated synthesis, and iterative screening is expected to significantly shorten the R&D cycle.
Production processes are also constantly improving. Heterologous expression systems such as Pichia pastoris have successfully produced active Kv1.3 inhibitors, including Vm24, Anuroctoxin, and Ts6, in practical quantities [99]. Cell-free protein synthesis platforms may further facilitate the rapid screening of toxic or highly engineered variants [100]. Beyond their therapeutic applications, scorpion venom peptides remain highly valuable research probes. For example, BmKK2 has been used to elucidate the Kv1.3-dependent pathway linking the NF-κB signaling pathway and NLRP3 activation in macrophages [101]. Such mechanistic studies often precede drug discovery.
Future clinical development is likely to move towards precision medicine. A patient’s genotype, channel expression profile, immune status, and disease subtype can all influence responses to toxin-derived drugs. With improvements in target stratification and delivery technologies, scorpion venom peptides hold promise for evolving from specialized pharmacological tools into clinically applicable precision therapies for pain, cancer, and neurological disorders.
7. Conclusions
In summary, scorpion venom peptides have evolved from classical ion channel probes to a versatile platform for redox-based therapeutics. This review establishes that their antioxidant activity operates through three convergent mechanisms: transcriptional activation of the Nrf2-ARE axis, upregulating endogenous enzymes (SOD, CAT, GSH); direct inhibition of NOX2 assembly by blocking p47phox membrane translocation, curtailing ROS generation at its source; and radical scavenging by cysteine-free linear peptides independent of cellular signaling. The synthetic peptide SVHRSP exemplifies this multi-pathway synergy, showing neuroprotection in Parkinson’s, Alzheimer’s, and epilepsy models via NOX2 suppression, NLRP3 inflammasome inhibition, and Nrf2 nuclear translocation, a combination that has now entered Phase 1.1 clinical evaluation in China.
Despite these advances, major translational gaps remain, including off-target effects due to subtype selectivity, metabolic instability, immunogenicity, and limited blood–brain barrier penetration. Moreover, most antioxidant claims are based on cellular or rodent models, with limited human data on redox biomarkers (e.g., plasma 8-OHdG, GSH/GSSG ratio). Future efforts should integrate AI-guided peptide engineering (e.g., cyclization, D-amino acid substitution), nanoparticle or exosome delivery, and harmonized clinical readouts of oxidative stress. The dual pro-/anti-oxidant selectivity observed between normal and cancer cells also warrants deeper investigation into its therapeutic window.
Looking forward, scorpion venom peptides represent a promising, nature-inspired scaffold for precision intervention in oxidative stress-driven diseases, from neurodegeneration to ischemia and malignancy. With continued innovation in structural biology, synthetic biology, and delivery technologies, these molecules are poised to move from venom to valuable medicines in the antioxidant armamentarium.
Author Contributions
Conceptualization, L.C. and J.Z.; investigation, M.W., H.L. and Y. X.; writing-original draft preparation, L.C.; writing-review and editing S.L., Y. G. and J.Z.; funding acquisition, L.C., S.L. and J.Z.. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Liaoning Provincial Key R&D Program for Industrial Technology Innovation: 2024JH2/102400054. Liaoning Provincial Natural Science Foundation General Project (2025-MS-246). The Innovation and Entrepreneurship Plan of Science and Technology Talent of Dalian: 2022RG09. Dalian Medical University Young Talent Cultivation Fund (xqnky202412).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Guan, C.G.; Torres, M.D.T.; Li, S.F.; de la Fuente-nunez, C. Computational exploration of global venoms for antimicrobial discovery with Venomics artificial intelligence. Nat Commun 2025, 16. [CrossRef]
- Xia, Z.Q.; He, D.G.; Wu, Y.L.; Kwok, H.F.; Cao, Z.J. Scorpion venom peptides: Molecular diversity, structural characteristics, and therapeutic use from channelopathies to viral infections and cancers. Pharmacol Res 2023, 197. [CrossRef]
- Mendes, L.C.; Viana, G.M.M.; Nencioni, A.L.A.; Pimenta, D.C.; Beraldo-Neto, E. Scorpion Peptides and Ion Channels: An Insightful Review of Mechanisms and Drug Development. Toxins 2023, 15. [CrossRef]
- Xia, Z.Q.; Xie, L.X.; Li, B.; Lv, X.Y.; Zhang, H.Z.; Cao, Z.J. Antimicrobial Potential of Scorpion-Venom-Derived Peptides. Molecules 2024, 29. [CrossRef]
- Pashmforoosh, N.; Baradaran, M. Peptides with Diverse Functions from Scorpion Venom: A Great Opportunity for the Treatment of a Wide Variety of Diseases. Iran Biomed J 2023, 27, 84-99. [CrossRef]
- Kamau, P.M.; Zhong, J.; Yao, B.; Lai, R.; Luo, L. Bioactive peptides from scorpion venoms: therapeutic scaffolds and pharmacological tools. Chin J Nat Medicines 2023, 21, 19-35. [CrossRef]
- Barnham, K.J.; Masters, C.L.; Bush, A.I. Neurodegenerative diseases and oxidative stress. Nature Reviews Drug Discovery 2004, 3, 205-214. [CrossRef]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat Rev Mol Cell Bio 2020, 21, 363-383. [CrossRef]
- Abdallnasser Amen, R.; Atef Essmat, R.; Farid, A.; Abdel-Rahman, M.A.; El-Sherif, A.A.; Zhang, Y. Scorpion venom as a natural peptide source for innovative therapeutic solutions: A comprehensive review of its potential in emerging medical frontiers. Toxicon 2025, 268. [CrossRef]
- Liu, M.; Svirskis, D.; Proft, T.; Loh, J.; Yin, N.; Li, H.; Li, D.; Zhou, Y.; Chen, S.; Song, L.; et al. Progress in peptide and protein therapeutics: Challenges and strategies. Acta Pharm Sin B 2025, 15, 6342-6381. [CrossRef]
- He, D.G.; Lei, Y.N.; Qin, H.X.; Cao, Z.J.; Kwok, H.F. Deciphering Scorpion Toxin-Induced Pain: Molecular Mechanisms and Ion Channel Dynamics. International Journal of Biological Sciences 2025, 21, 2921-2934. [CrossRef]
- Liu, M.; Svirskis, D.; Proft, T.; Loh, J.; Yin, N.; Li, H.; Li, D.; Zhou, Y.; Chen, S.; Song, L.; et al. Progress in peptide and protein therapeutics: Challenges and strategies. Acta Pharmaceutica Sinica B 2025, 15, 6342-6381. [CrossRef]
- Rafferty, J.; Nagaraj, H.; P. McCloskey, A.; Huwaitat, R.; Porter, S.; Albadr, A.; Laverty, G. Peptide Therapeutics and the Pharmaceutical Industry: Barriers Encountered Translating from the Laboratory to Patients. Current Medicinal Chemistry 2016, 23, 4231-4259. [CrossRef]
- Shariati, S.; Mafakher, L.; Shirani, M.; Baradaran, M. Unveiling new Kv1.3 channel blockers from scorpion venom: Characterization of Meuk7–3 and in silico design of its analogs for enhanced affinity and therapeutic potential. International Journal of Biological Macromolecules 2025, 319. [CrossRef]
- Huang, Y.; Kamau, P.M.; Wang, J.M.; Gao, M.Y.; Li, B.W. Scorpion Venom Neurotoxins: Molecular Diversity, Mechanisms, and Drug Scaffolds. Toxins 2026, 18. [CrossRef]
- Aoki-Shioi, N.; Nomura, S.; Tanaka, Y.; Hirose, S. Ion Channel-Targeting Toxins: Structural Mechanisms of Activation, Inhibition, and Therapeutic Potential. Toxins 2025, 17. [CrossRef]
- Vacher, H.; Diochot, S.; Bougis, P.E.; Martin-Eauclaire, M.F.; Mourre, C. Kv4 channels sensitive to BmTX3 in rat nervous system: autoradiographic analysis of their distribution during brain ontogenesis. Eur J Neurosci 2006, 24, 1325-1340. [CrossRef]
- Shakeel, K.; Naseem, M.U.; Olamendi-Portugal, T.; Zamudio, F.Z.; Possani, L.D.; Panyi, G. Cvill6 and Cvill7: Potent and Selective Peptide Blockers of Kv1.2 Ion Channel Isolated from Mexican Scorpion Centruroides villegasi. Toxins (Basel) 2025, 17. [CrossRef]
- Ramsoomair, D.; Ramsoomair, C.K.; Daftari, M.; Himic, V.; Shlobin, N.A.; Wang, S.E.; Ivan, M.E.; Komotar, R.J.; Shah, A.H. Translating Venom to Medicine: A Comprehensive Review on the Role of Chlorotoxin in Glioblastoma Diagnosis and Therapy. Molecular Cancer Therapeutics 2025, 24, 1867-1877. [CrossRef]
- Megaly, A.M.A.; Nakamichi, R.; Wakayu, M.; Nakagawa, Y.; Abdel-Wahab, M.; Miyashita, M. Identification of a novel insecticidal chlorotoxin-like peptide from the venom of the Compsobuthus egyptiensis scorpion. Toxicon 2025, 267. [CrossRef]
- Giugliano, R.; Della Marca, R.; Chianese, A.; Monti, A.; Donadio, F.; Esposito, E.; Doti, N.; Zannella, C.; Galdiero, M.; De Filippis, A. The inhibitory potential of three scorpion venom peptides against multidrug-resistant Klebsiella pnemoniae. Frontiers in Microbiology 2025, 16. [CrossRef]
- Zhu, S.; Gao, B.; Tytgat, J. Phylogenetic distribution, functional epitopes and evolution of the CSαβ superfamily. Cellular and Molecular Life Sciences 2005, 62, 2257-2269. [CrossRef]
- Saucedo, A.L.; Flores-Solis, D.; Rodriguez de la Vega, R.C.; Ramirez-Cordero, B.; Hernandez-Lopez, R.; Cano-Sanchez, P.; Noriega Navarro, R.; Garcia-Valdes, J.; Coronas-Valderrama, F.; de Roodt, A.; et al. New tricks of an old pattern: structural versatility of scorpion toxins with common cysteine spacing. J Biol Chem 2012, 287, 12321-12330. [CrossRef]
- Qin, C.; Yang, X.; Zuo, Z.; Yang, L.; Yang, F.; Cao, Z.; Chen, Z.; Wu, Y. BmK86-P1, a New Degradation Peptide with Desirable Thermostability and Kv1.2 Channel-Specific Activity from Traditional Chinese Scorpion Medicinal Material. Toxins 2021, 13. [CrossRef]
- Catterall, W.A.; Cestèle, S.; Yarov-Yarovoy, V.; Yu, F.H.; Konoki, K.; Scheuer, T. Voltage-gated ion channels and gating modifier toxins. Toxicon 2007, 49, 124-141. [CrossRef]
- Martin-Eauclaire, M.F.; Bougis, P.E.; de Lima, M.E. Ts1 from the Brazilian scorpion: A half-century of studies on a multifunctional beta like-toxin. Toxicon 2018, 152, 106-120. [CrossRef]
- Delgado-Prudencio, G.; Possani, L.D.; Becerril, B.; Ortiz, E. The Dual α-Amidation System in Scorpion Venom Glands. Toxins 2019, 11. [CrossRef]
- Adegoke, V.A.; Dongol, Y.; Gonzalez, T.; Song, A.; Clark, R.J.; Lewis, R.J.; Conibear, A.C.; Rosengren, K.J.; Aguilar, M. Proline hydroxylation and C-terminal amidation in µ-conotoxins increase structural stability and potency at sodium channels. Australian Journal of Chemistry 2025, 78. [CrossRef]
- Möller, D.S.; van der Walt, M.; Oosthuizen, C.; Serian, M.; Serem, J.C.; Lorenz, C.D.; Mason, A.J.; Bester, M.J.; Gaspar, A.R.M. Improving the Activity and Selectivity of a Scorpion-Derived Peptide, A3a, against through Rational Design. Acs Omega 2025, 10, 4699-4710. [CrossRef]
- Du, C.; Li, H.; Liu, C.; Fan, H. Understanding of the postgerminative development response to salinity and drought stresses in cucumber seeds by integrated proteomics and transcriptomics analysis. Journal of Proteomics 2021, 232. [CrossRef]
- Zeng, L.; Sun, Y.; Zhang, H.; Yi, X.; Du, R.; Chen, Z.; Wang, Q. Scorpion venom peptides enhance immunity and survival in Litopenaeus vannamei through antibacterial action against Vibrio parahaemolyticus. Frontiers in Immunology 2025, 16. [CrossRef]
- Xia, Z.; He, D.; Wu, Y.; Kwok, H.F.; Cao, Z. Scorpion venom peptides: Molecular diversity, structural characteristics, and therapeutic use from channelopathies to viral infections and cancers. Pharmacol Res 2023, 197. [CrossRef]
- Zhang, N.; Li, M.; Chen, X.; Wang, Y.; Wu, G.; Hu, G.; Wu, H. Solution structure of BmKK2, a new potassium channel blocker from the venom of chinese scorpion Buthus martensi Karsch. Proteins: Structure, Function, and Bioinformatics 2004, 55, 835-845. [CrossRef]
- Clairfeuille, T.; Cloake, A.; Infield, D.T.; Llongueras, J.P.; Arthur, C.P.; Li, Z.R.; Jian, Y.; Martin-Eauclaire, M.-F.; Bougis, P.E.; Ciferri, C.; et al. Structural basis of α-scorpion toxin action on Nav channels. Science 2019, 363. [CrossRef]
- Chen, R.; Chung, S.-H. Computational Studies of Venom Peptides Targeting Potassium Channels. Toxins 2015, 7, 5194-5211. [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583-+. [CrossRef]
- Abramson, J.; Adler, J.; Dunger, J.; Evans, R.; Green, T.; Pritzel, A.; Ronneberger, O.; Willmore, L.; Ballard, A.J.; Bambrick, J.; et al. Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature 2024, 630, 493-500. [CrossRef]
- Miyashita, M.; Mitani, N.; Iwamoto, F.; Hirota, M.; Nakagawa, Y. Discovery of a Novel Insecticidal Peptide with a Cystine-Stabilized α-Helix/α-Helix Motif from the Venom of Scorpion Liocheles australasiae. Molecules 2024, 30. [CrossRef]
- Montero-Dominguez, P.A.; Restano-Cassulini, R.; Magaña-Ávila, L.C.; Almanza, A.; Mercado, F.; Corzo, G. Design of antinociceptive peptide by grafting domains between scorpion β-neurotoxins. Bioorganic Chemistry 2025, 162. [CrossRef]
- Wai, D.C.C.; Naseem, M.U.; Mocsár, G.; Reddiar, S.B.; Pan, Y.J.; Csoti, A.; Hajdu, P.; Nowell, C.; Nicolazzo, J.A.; Panyi, G.; et al. Fluorescent Peptide Toxin for Selective Visualization of the Voltage-Gated Potassium Channel K1.3. Bioconjugate Chem 2022. [CrossRef]
- Attali, B.; Ma, Z.; Kong, J.; Gordon, D.; Gurevitz, M.; Kallen, R.G. Direct Evidence that Scorpion α-Toxins (Site-3) Modulate Sodium Channel Inactivation by Hindrance of Voltage-Sensor Movements. PLoS ONE 2013, 8. [CrossRef]
- Catterall, W.A.; Swanson, T.M. Structural Basis for Pharmacology of Voltage-Gated Sodium and Calcium Channels. Mol Pharmacol 2015, 88, 141-150. [CrossRef]
- Gurevitz, M., Zhorov, B. S. and Dong, K. Allosteric interactions among voltage-sensor modules of sodium channels probed by scorpion toxin modifiers. Journal of Neurobiology and Physiology 2022, 4. [CrossRef]
- Bennett, D.L.H.; Woods, C.G. Painful and painless channelopathies. The Lancet Neurology 2014, 13, 587-599. [CrossRef]
- Xin, K.; Sun, R.; Xiao, W.; Lu, W.; Sun, C.; Lou, J.; Xu, Y.; Chen, T.; Wu, D.; Gao, Y. Short Peptides from Asian Scorpions: Bioactive Molecules with Promising Therapeutic Potential. Toxins 2025, 17. [CrossRef]
- Liu, Y.Y.; Liu, Y.Y.; Liu, Y.X.; Cui, Y.; Meng, T.; Song, Y.B.; Zhao, F. From traditional medicine to targeted therapy: Structure-activity relationship-guided optimization of scorpion toxin DKK2 for pain-associated sodium channel blockade. J Ethnopharmacol 2025, 352. [CrossRef]
- Bennett, D.L.H.; Woods, C.G. Painful and painless channelopathies. Lancet Neurol 2014, 13, 587-599. [CrossRef]
- Gigolaev, A.M.; Pinheiro, E.; Peigneur, S.; Tytgat, J.; Vassilevski, A.A. KV1.2-Selective Peptide with High Affinity. J Evol Biochem Phys+ 2022, 58, 2048-2057. [CrossRef]
- Gigolaev, A.M.; Kuzmenkov, A.I.; Peigneur, S.; Tabakmakher, V.M.; Pinheiro-Junior, E.L.; Chugunov, A.O.; Efremov, R.G.; Tytgat, J.; Vassilevski, A.A. Tuning Scorpion Toxin Selectivity: Switching From K1.1 to K1.3. Front Pharmacol 2020, 11. [CrossRef]
- Shakeel, K.; Olamendi-Portugal, T.; Naseem, M.U.; Becerril, B.; Zamudio, F.Z.; Delgado-Prudencio, G.; Possani, L.D.; Panyi, G. Of Seven New K+ Channel Inhibitor Peptides of Centruroides bonito, α-KTx 2.24 Has a Picomolar Affinity for Kv1.2. Toxins 2023, 15. [CrossRef]
- Short, B. A scorpion toxin takes the sting out of T cell activation. J Gen Physiol 2022, 154. [CrossRef]
- Meredith, A.L. BK Channelopathies and -Linked Disease Models. Annual Review of Physiology 2024, 86, 277-300. [CrossRef]
- Chen, Y.H.; Markov, N.; Gigon, L.; Hosseini, A.; Yousefi, S.; Stojkov, D.; Simon, H.U. The BK Channel Limits the Pro-Inflammatory Activity of Macrophages. Cells-Basel 2024, 13. [CrossRef]
- Ancatén-González, C.; Segura, I.; Alvarado-Sánchez, R.; Chávez, A.E.; Latorre, R. Ca2+- and Voltage-Activated K+ (BK) Channels in the Nervous System: One Gene, a Myriad of Physiological Functions. International Journal of Molecular Sciences 2023, 24. [CrossRef]
- Hakim, M.; Jiang, W.; Luo, L.; Li, B.; Yang, S.; Song, Y.; Lai, R. Scorpion Toxin, BmP01, Induces Pain by Targeting TRPV1 Channel. Toxins 2015, 7, 3671-3687. [CrossRef]
- Heusser, S.A.; Borg, C.B.; Colding, J.M.; Pless, S.A. Conformational decoupling in acid-sensing ion channels uncovers mechanism and stoichiometry of PcTx1-mediated inhibition. Elife 2022, 11. [CrossRef]
- Jiang, D.; Tonggu, L.; Gamal El-Din, T.M.; Banh, R.; Pomès, R.; Zheng, N.; Catterall, W.A. Structural basis for voltage-sensor trapping of the cardiac sodium channel by a deathstalker scorpion toxin. Nat Commun 2021, 12. [CrossRef]
- Sumino, A.; Sumikama, T.; Uchihashi, T.; Oiki, S. High-speed AFM reveals accelerated binding of agitoxin-2 to a K+ channel by induced fit. Science Advances 2019, 5. [CrossRef]
- Lee, S.-Y.; MacKinnon, R. A membrane-access mechanism of ion channel inhibition by voltage sensor toxins from spider venom. Nature 2004, 430, 232-235. [CrossRef]
- Zhu, Q.; Du, Y.; Nomura, Y.; Gao, R.; Cang, Z.; Wei, G.-W.; Gordon, D.; Gurevitz, M.; Groome, J.; Dong, K. Charge substitutions at the voltage-sensing module of domain III enhance actions of site-3 and site-4 toxins on an insect sodium channel. Insect Biochem Molec 2021, 137. [CrossRef]
- Zhang, X.M.; Tu, D.Z.; Li, S.; Li, N.; Li, D.L.; Gao, Y.; Tian, L.; Liu, J.N.; Zhang, X.; Hong, J.S.; et al. A novel synthetic peptide SVHRSP attenuates dopaminergic neurodegeneration by inhibiting NADPH oxidase-mediated neuroinflammation in experimental models of Parkinson’s disease. Free Radical Biology and Medicine 2022, 188, 363-374. [CrossRef]
- Xiao, Q.; Zhang, Z.P.; Hou, Y.B.; Qu, D.X.; Tang, L.L.; Chen, L.J.; Li, G.Y.; Ji, Y.H.; Tao, J.; Zhu, Y.D. Anti-epileptic/pro-epileptic effects of sodium channel modulators from Buthus martensii Karsch. Sheng Li Xue Bao 2022, 74, 621-632.
- Wang, X.; Luo, H.; Peng, X.; Chen, J. Spider and scorpion knottins targeting voltage-gated sodium ion channels in pain signaling. Biochemical Pharmacology 2024, 227. [CrossRef]
- Aftabizadeh, M.; Bawa, L.; Wang, S.; Brewster, B.; Wang, D.; Wang, X.; Brown, C.; Barish, M. Exth-10. Exploration of a Novel Toxin-Incorporating Car T Cell: How Does Chlorotoxin Recognize Glioblastoma Cells? Neuro-Oncology 2021, 23, vi165-vi165. [CrossRef]
- Nguyen, T.; Guo, R.Y.; Chai, J.W.; Wu, J.N.; Liu, J.F.; Chen, X.; Abdel-Rahman, M.A.; Xia, H.; Xu, X.Q. Smp24, a Scorpion-Venom Peptide, Exhibits Potent Antitumor Effects against Hepatoma HepG2 Cells via Multi-Mechanisms In Vivo and In Vitro. Toxins 2022, 14. [CrossRef]
- Xu, L.P.; Zhang, Y.; Dai, Q.D.; Lin, N.; Guan, T.R.; Song, X.W.; Hong, S.H. Scorpion venom polypeptide governs alveolar macrophage M1/M2 polarization to alleviate pulmonary fibrosis. Tissue Cell 2022, 79. [CrossRef]
- Amen, R.A.; Essmat, R.A.; Farid, A.; Abdel-Rahman, M.A.; El-Sherif, A.A.; Zhang, Y.H. Scorpion venom as a natural peptide source for innovative therapeutic solutions: A comprehensive review of its potential in emerging medical frontiers. Toxicon 2025, 268. [CrossRef]
- Rincón-Cortés, C.A.; Bayona-Rojas, M.A.; Reyes-Montaño, E.A.; Vega-Castro, N.A. Antimicrobial Activity Developed by Scorpion Venoms and Its Peptide Component. Toxins 2022, 14. [CrossRef]
- Nasr, S.; Borges, A.; Sahyoun, C.; Nasr, R.; Roufayel, R.; Legros, C.; Sabatier, J.M.; Fajloun, Z. Scorpion Venom as a Source of Antimicrobial Peptides: Overview of Biomolecule Separation, Analysis and Characterization Methods. Antibiotics-Basel 2023, 12. [CrossRef]
- Zhang, Y.; Li, S.; Hou, L.; Wu, M.; Liu, J.; Wang, R.; Wang, Q.; Zhao, J. NLRP3 mediates the neuroprotective effects of SVHRSP derived from scorpion venom in rotenone-induced experimental Parkinson’s disease model. J Ethnopharmacol 2023, 312. [CrossRef]
- Balde, A.; Benjakul, S.; Nazeer, R.A. A review on NLRP3 inflammasome modulation by animal venom proteins/peptides: mechanisms and therapeutic insights. Inflammopharmacology 2025, 33, 1013-1031. [CrossRef]
- Chen, M.D.; Zhang, Y.; Hou, L.Y.; Zhao, Z.R.; Tang, P.Y.; Sun, Q.Q.; Zhao, J.; Wang, Q.S. SVHRSP protects against rotenone-induced neurodegeneration in mice by inhibiting TLR4/NF-κB-mediated neuroinflammation via gut microbiota. Npj Parkinsons Dis 2025, 11. [CrossRef]
- Li, S.; Guo, Z.Y.; Liu, J.N.; Ma, Y.; Zhang, X.M.; Hou, L.Y.; Wang, Q.H.; Jiang, W.W.; Wang, Q.S. CD11b-NOX2 mutual regulation-mediated microglial exosome release contributes to rotenone-induced inflammation and neurotoxicity in BV2 microglia and primary cultures. Free Radical Biology and Medicine 2024, 224, 436-446. [CrossRef]
- Wang, Q.H.; Liu, J.N.; Zhang, Y.; Li, Z.; Zhao, Z.R.; Jiang, W.W.; Zhao, J.; Hou, L.Y.; Wang, Q.S. Microglial CR3 promotes neuron ferroptosis via NOX2-mediated iron deposition in rotenone-induced experimental models of Parkinson’s disease. Redox Biol 2024, 77. [CrossRef]
- Li, X.; Wu, X.; Li, N.; Li, D.; Sui, A.; Khan, K.; Ge, B.; Li, S.; Li, S.; Zhao, J. Scorpion venom heat-resistant synthesized peptide ameliorates 6-OHDA-induced neurotoxicity and neuroinflammation: likely role of Na(v) 1.6 inhibition in microglia. Br J Pharmacol 2021, 178, 3553-3569. [CrossRef]
- Qin, C.; Li, D.; Zhang, J.; Yin, Z.; Li, F. Scorpion Venom Heat-Resistant Synthetic Peptide Alleviates Neuronal Necroptosis in Alzheimer’s Disease Model by Regulating Lnc Gm6410 Under PM2.5 Exposure. International Journal of Molecular Sciences 2025, 26. [CrossRef]
- Wang, Y.-Z.; Guo, S.-Y.; Kong, R.-L.; Sui, A.-R.; Wang, Z.-H.; Guan, R.-X.; Supratik, K.; Zhao, J.; Li, S. Scorpion Venom Heat–Resistant Synthesized Peptide Increases Stress Resistance and Extends the Lifespan of Caenorhabditis elegans via the Insulin/IGF-1-Like Signal Pathway. Front Pharmacol 2022, 13. [CrossRef]
- Zhang, X.; Tu, D.; Li, S.; Li, N.; Li, D.; Gao, Y.; Tian, L.; Liu, J.; Zhang, X.; Hong, J.-S.; et al. A novel synthetic peptide SVHRSP attenuates dopaminergic neurodegeneration by inhibiting NADPH oxidase-mediated neuroinflammation in experimental models of Parkinson’s disease. Free Radical Biology and Medicine 2022, 188, 363-374. [CrossRef]
- Fawzy, B.S.; Nafie, M.S.; Ali, I.A.I.; El-Baz, L.M.F.; Xu, X.; Abdel-Rahman, M.A. Scorpion Venom Peptide Smp24 Revealed Apoptotic and Antiangiogenic Activities in Solid-Ehrlich Carcinoma Bearing Mice. International Journal of Peptide Research and Therapeutics 2023, 29. [CrossRef]
- Dahiya, R.; Goyal, K.; Sharma, K.; Rawat, A.; Sharma, V.; Mathur, P. Scorpion venom as a molecular treasure: emerging bioactive compounds and translational therapeutic insights. Archives of Toxicology 2025, 100, 437-450. [CrossRef]
- Duan, K.; Qin, C.; Zhang, Y.; Li, D.; Zhang, J.; Li, F. Scorpion venom heat-resistant synthetic peptide regulates Nrf-2 to alleviate neuronal pyroptosis in an AD model induced by endoplasmic reticulum stress under PM2.5 exposure. Toxicon 2025, 265. [CrossRef]
- Zheng, Q.; Na, R.; Yang, L.; Yu, H.; Zhao, X.; Huang, X. The binding process of BmKTX and BmKTX-D33H toward to Kv1.3 channel: a molecular dynamics simulation study. Journal of Biomolecular Structure and Dynamics 2020, 39, 2788-2797. [CrossRef]
- Edwards, W.; Fung-Leung, W.P.; Huang, C.C.; Chi, E.; Wu, N.; Liu, Y.; Maher, M.P.; Bonesteel, R.; Connor, J.; Fellows, R.; et al. Targeting the Ion Channel Kv1.3 with Scorpion Venom Peptides Engineered for Potency, Selectivity, and Half-life. Journal of Biological Chemistry 2014, 289, 22704-22714. [CrossRef]
- Rashid, M.H.; Huq, R.; Tanner, M.R.; Chhabra, S.; Khoo, K.K.; Estrada, R.; Dhawan, V.; Chauhan, S.; Pennington, M.W.; Beeton, C.; et al. A potent and Kv1.3-selective analogue of the scorpion toxin HsTX1 as a potential therapeutic for autoimmune diseases. Sci Rep-Uk 2014, 4. [CrossRef]
- Boltman, T.; Meyer, M.; Ekpo, O. Diagnostic and Therapeutic Approaches for Glioblastoma and Neuroblastoma Cancers Using Chlorotoxin Nanoparticles. Cancers 2023, 15. [CrossRef]
- Durham, P.G.; Butnariu, A.; Alghorazi, R.; Pinton, G.; Krishna, V.; Dayton, P.A. Current clinical investigations of focused ultrasound blood-brain barrier disruption: A review. Neurotherapeutics 2024, 21. [CrossRef]
- Zhang, Z.; Li, J.; Wang, Y.; Tang, C.; Zhou, Y.; Li, J.; Lu, X.; Wang, Y.; Ma, T.; Xu, H.; et al. Angiopep-2 conjugated biomimetic nano-delivery system loaded with resveratrol for the treatment of methamphetamine addiction. International Journal of Pharmaceutics 2024, 663. [CrossRef]
- Wang, T.; Wang, S.W.; Zhang, Y.; Wu, X.F.; Peng, Y.; Cao, Z.; Ge, B.Y.; Wang, X.; Wu, Q.; Lin, J.T.; et al. Scorpion Venom Heat-Resistant Peptide (SVHRP) Enhances Neurogenesis and Neurite Outgrowth of Immature Neurons in Adult Mice by Up-Regulating Brain-Derived Neurotrophic Factor (BDNF). Plos One 2014, 9. [CrossRef]
- Batrakova, E.V.; Kim, M.S. Using exosomes, naturally-equipped nanocarriers, for drug delivery. J Control Release 2015, 219, 396-405. [CrossRef]
- Mamelak, A.N.; Rosenfeld, S.; Bucholz, R.; Raubitschek, A.; Nabors, L.B.; Fiveash, J.B.; Shen, S.; Khazaeli, M.B.; Colcher, D.; Liu, A.; et al. Phase I single-dose study of intracavitary-administered iodine-131-TM-601 in adults with recurrent high-grade glioma. J Clin Oncol 2006, 24, 3644-3650. [CrossRef]
- Neff, R.A.; Wickenden, A.D. Selective Targeting of Nav1.7 with Engineered Spider Venom-Based Peptides. Channels 2021, 15, 179-193. [CrossRef]
- Abramson, J.; Adler, J.; Dunger, J.; Evans, R.; Green, T.; Pritzel, A.; Ronneberger, O.; Willmore, L.; Ballard, A.J.; Bambrick, J.; et al. Accurate structure prediction of biomolecular interactions with AlphaFold 3 (vol 630, pg 493, 2024). Nature 2024, 636, E4-E4. [CrossRef]
- Dauparas, J.; Anishchenko, I.; Bennett, N.; Bai, H.; Ragotte, R.J.; Milles, L.F.; Wicky, B.I.M.; Courbet, A.; de Haas, R.J.; Bethel, N.; et al. Robust deep learning-based protein sequence design using ProteinMPNN. Science 2022, 378, 49-55. [CrossRef]
- Ren, C.-Z.; Wu, Z.-T.; Wang, W.; Tan, X.; Yang, Y.-H.; Wang, Y.-K.; Li, M.-L.; Wang, W.-Z. SIRT1 exerts anti-hypertensive effect via FOXO1 activation in the rostral ventrolateral medulla. Free Radical Biology and Medicine 2022, 188, 1-13. [CrossRef]
- Radhakrishna, K.; Rajshekhar, P.K.; Arshitha, R.; Patil, K.; Dhannura, S.; Gowd, K.H. Dissecting oxidative folding of conotoxins using 3D structures of cysteine mutants predicted by AlphaFold 3: A case study of α-conotoxin RgIA, χ-conotoxin CMrVIA and ω-conotoxin MVIIA-Gly. Toxicon 2025, 262. [CrossRef]
- Callaway, E. ‘It will change everything’: DeepMind’s AI makes gigantic leap in solving protein structures. Nature 2020, 588, 203-204. [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583-589. [CrossRef]
- Wang, F.; Wang, Y.; Feng, L.; Zhang, C.; Lai, L. Target-Specific De Novo Peptide Binder Design with DiffPepBuilder. Journal of Chemical Information and Modeling 2024, 64, 9135-9149. [CrossRef]
- Borrego, J.; Naseem, M.U.; Sehgal, A.N.A.; Panda, L.R.; Shakeel, K.; Gaspar, A.; Nagy, C.; Varga, Z.; Panyi, G. Recombinant Expression in Pichia pastoris System of Three Potent Kv1.3 Channel Blockers: Vm24, Anuroctoxin, and Ts6. Journal of Fungi 2022, 8. [CrossRef]
- Thornton, E.L.; Boyle, J.T.; Laohakunakorn, N.; Regan, L. Cell-Free Protein Synthesis as a Method to Rapidly Screen Machine Learning-Generated Protease Variants. Acs Synth Biol 2025, 14, 1710-1718. [CrossRef]
- Wang, Z.; Sang, M.; Zhang, Y.; Chen, S.; Li, S.; Chen, Y.; Xu, E.; Zhou, Q.; Xu, W.; Zhao, C.; et al. BmKK2, a thermostable Kv1.3 blocker from Buthus martensii Karsch (BmK) scorpion, inhibits the activation of macrophages via Kv1.3-NF-κB- NLRP3 axis. J Ethnopharmacol 2023, 314. [CrossRef]
Figure 1.
Topological schematic of the three major structural scaffolds of scorpion venom peptides.

Figure 2.
Molecular mechanisms of voltage-gated sodium channel modulation by scorpion venom peptides.
Figure 2.
Molecular mechanisms of voltage-gated sodium channel modulation by scorpion venom peptides.

Figure 3.
Multi-target neuroprotective mechanism of SVHRSP.

Figure 4.
Schematic diagram of the antioxidant mechanisms of scorpion venom peptides.

Table 1.
Major structural families and functional characteristics of scorpion venom peptides.
| Structural family | disulphide pattern | 3D folding feature | Representative member | Target/mechanism | Citations |
|---|---|---|---|---|---|
| CSα/β | 8 Cys, 4 disulphide bonds |
α-helix + three-stranded antiparallel β-sheet | AaH II (α-NaTx), CsVII (β-NaTx) | NaV channels: α-toxin slows inactivation; β-toxin shifts activation voltage | [15] |
| CSα/α | C-C-CC-C-C | Two α-helices (“two-finger” or “three-finger”) | Charybdotoxin (α-KTx 1.1), HsTx1 (κ-KTx) | KV channels: physical pore blocking or voltage sensor trapping | [14,40] |
| CSβ | C-C-C-C | Three-stranded antiparallel β-sheet (no α-helix) | Chlorotoxin | MMP-2/Annexin A2, glioma targeting | [19] |
| Cysteine-free linear peptide | No disulphide bond | Amphipathic α-helix | Pandinin-1 | Membrane disruption (“carpet” or “barrel-stave” model) | [21] |
Table 3.
Pharmacological activities and clinical translation status of representative scorpion venom peptides.
Table 3.
Pharmacological activities and clinical translation status of representative scorpion venom peptides.
| Molecule | Structural family | Primary target | Pharmacological activity | Clinical/translation status | Citations |
|---|---|---|---|---|---|
| OD1 | CSα/β (α-like) | NaV1.7 | Analgesia | Preclinical (engineering for subtype selectivity) | [39] |
| DKK2-N18W | CSα/β | NaV1.7 | Analgesia | Preclinical (effective in formalin pain model) | [45,46] |
| Charybdotoxin (ChTx) | CSα/α (α-KTx 1.1) | KV1.3 | Immunosuppression | Tool molecule; engineered for autoimmune diseases | [48] |
| Cvill7 | CSα/α (α-KTx 2) | KV1.2 | Anti-epileptic potential | Lead candidate (450-fold selective over KV1.3) | [18] |
| BmKK2 | CSα/α | KV1.3 | Anti-inflammatory (NF-κB-NLRP3 inhibition) | Preclinical (from traditional Chinese medicine) | [70] |
| Chlorotoxin (TM-601) | CSβ | MMP-2 | Glioma-targeted diagnosis and therapy | Phase I/II trials (intracavitary); shifted to imaging/delivery | [19] |
| SVHRSP | Synthetic derivative (heat-resistant peptide) | Multi-target (NLRP3, TLR4, NOX2, Nav1.6) | Neuroprotection (Parkinson’s, Alzheimer’s, epilepsy) | NMPA Class 1.1 IND approved (2025) | [70,72,75] |
| Pantinin-1/2 | Cysteine-free linear peptide | Bacterial outer membrane LPS | Anti-MDR Klebsiella pneumoniae | Preclinical (MIC 6-25 μM) | [21] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



