Submitted:
17 April 2026
Posted:
21 April 2026
You are already at the latest version
Abstract
The renin–angiotensin–aldosterone system (RAAS) is a central regulator of blood pressure and fluid homeostasis. However, its dysregulation contributes to the development of cardiovascular and chronic kidney diseases, including hypertension, diabetes, and metabolic disorders. The identification of the prorenin receptor (PRR) has expanded the understanding of RAAS, revealing functions beyond its classical role in angiotensin II (Ang II) generation.
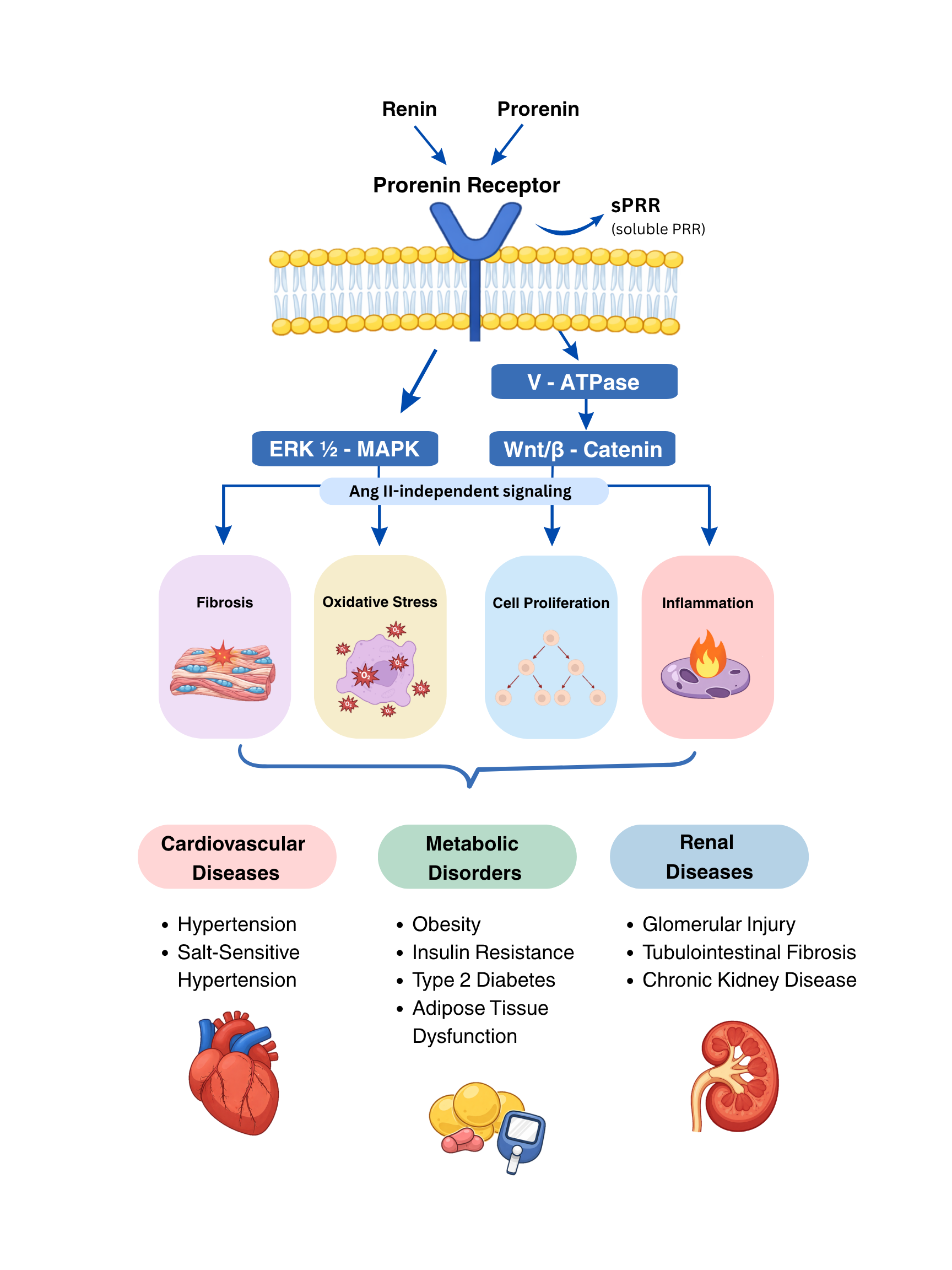
In this review, we provide an updated and integrative overview of PRR biology, emphasizing its multifunctional roles in both Ang II–dependent and independent signaling. PRR also functions as an accessory component of the vacuolar H⁺-ATPase and participates in key intracellular pathways, including ERK1/2-MAPK, PI3K/Akt, and Wnt/β-catenin. Through these mechanisms, PRR contributes to cardiovascular remodeling, renal inflammation and fibrosis, metabolic dysregulation, and angiogenesis.
Emerging evidence further identifies the soluble form of PRR (sPRR) as a biologically active circulating factor with endocrine-like properties. Clinical and experimental studies suggest that sPRR serves as both a biomarker and a mediator linking tissue RAAS activation to systemic cardiorenal and metabolic disease progression.
Collectively, this review highlights PRR as a central molecular hub that integrates extracellular hormonal signals with intracellular metabolic and inflammatory pathways, underscoring its relevance in the pathophysiology of cardiovascular, renal, and metabolic diseases.
Keywords:
renin-angiotensin-aldosterone system
; hypertension
; chronic kidney disease
; intracellular signaling
1. Introduction
The renin–angiotensin–aldosterone system (RAAS) is a key regulator of blood pressure (BP) and extracellular fluid homeostasis [1]. When inappropriately activated, however, RAAS signaling contributes to the development and progression of cardiovascular disease (CVD) and chronic kidney disease (CKD), including hypertension, diabetes mellitus, and metabolic disorders. The cloning of the receptor for renin and prorenin—now known as the prorenin receptor (PRR)—more than twenty-five years ago [2] has stimulated fundamental investigations regarding its role in CV and renal physiology and its contribution to systemic and tissue RAAS-mediated pathophysiological processes.
The PRR is widely expressed in several tissues in humans and rodents, including the heart, brain, placenta, kidney, vasculature, and adipose tissues, to name a few [2,3,4]. The PRR critically regulates CV function through both Angiotensin II (Ang II)–dependent and –independent mechanisms. In the kidney, PRR is highly expressed along the nephron, where it influences intrarenal RAAS activity and the handling of water and sodium [5,6]. Through these actions, it contributes to the regulation of BP, acid-base homeostasis, and remodeling processes [7].
Emerging clinical evidence suggests that the circulating form of the soluble PRR (sPRR) plays a role in CV, renal, and metabolic disorders. Elevated plasma sPRR levels have been reported in patients with type 2 diabetes, hypertension, and CKD, and correlate with markers of renal dysfunction and CV risk [8,9]. The sPRR may influence hepatic glucose production, adipose tissue inflammation, and systemic RAAS activity, thereby linking PRR signaling to broader metabolic disturbances [9]. In this review, we provide an update of the contributions of PRR in CVD, vascular development, metabolic diseases, and renal inflammation.
2. Prorenin Receptor in Cardiovascular, Metabolic and Renal Diseases
The PRR, also known as Atp6ap2, is a multifunctional receptor linking the RAAS with intracellular signaling pathways that regulate inflammation, fibrosis, and metabolic homeostasis [10]. PRR was initially identified as a receptor that binds both prorenin and renin, with its role dependent on their activity/activation. Further studies found that PRR enhances catalytic conversion of angiotensinogen to Angiotensin I (Ang I), leading to increased local formation of Ang II in tissues fully independent of circulating renin activity [10]. In addition to this enzymatic amplification of tissue RAAS, PRR activation triggers Ang II-independent intracellular signaling cascades, including activation of ERK1/2, p38 MAPK, and PI3K/Akt pathways. These signaling events stimulate transcription of profibrotic mediators such as transforming growth factor-β (TGF-β), plasminogen activator inhibitor-1 (PAI-1), and extracellular matrix proteins, implicating PRR in tissue remodeling and fibrosis [10,11].
Beyond its role in RAAS signaling, PRR functions as an accessory protein of the vacuolar H+-ATPase (V-ATPase), which is essential for intracellular acidification, vesicular trafficking, and autophagy [3]. Through this interaction, PRR regulates lysosomal function and cellular metabolism. Disruption of the PRR–V-ATPase pathway impairs intracellular pH and protein degradation—processes increasingly recognized as key contributors to metabolic dysfunction and cellular stress [13]. These findings highlight PRR as a molecular hub integrating extracellular hormonal signals with intracellular metabolic pathways. In cardiomyocytes, PRR is closely associated with the V-ATPase and the ryanodine receptor, positioning it at the interface of intracellular calcium handling, lysosomal function, and downstream signaling pathways that influence myocardial contractility and structural integrity [13].
Cardiac PRR expression is upregulated in pathological states including diabetes, myocardial infarction, and heart failure, supporting its involvement in maladaptive cardiac remodeling [10,14]. Both low and high sodium diets increase myocardial prorenin/renin and PRR expression, leading to enhanced fibrotic remodeling under conditions of extreme sodium imbalance [15,16]. Despite these associations, the role of cardiac PRR in systemic BP regulation remains unclear. Constitutive deletion of PRR in cardiomyocytes results in lethal heart failure due to impaired lysosomal acidification, underscoring its essential role in cardiomyocyte homeostasis [13]. In contrast, global or cardiomyocyte-specific PRR overexpression does not significantly alter baseline BP or cardiac morphology. Adenovirus-mediated PRR overexpression in adult rat hearts induces Ang II–independent activation of intracellular signaling pathways and reduces left ventricular ejection fraction, indicating that PRR can directly promote cardiac dysfunction independent of systemic RAAS activation [17]. These findings suggest that cardiac PRR critically regulates myocardial remodeling and function rather than exerting a dominant influence on hemodynamic control.
In vascular smooth muscle cells (VSMCs), PRR expressed at the cell surface plays a more direct role in BP regulation and vascular remodeling [2,18]. VSMC-specific human PRR overexpression in transgenic rats induces hypertension and tachycardia, whereas PRR deficiency in VSMCs leads to aortic sclerosis without significant changes in BP, highlighting its context-dependent actions within the vasculature. PRR activation in VSMCs stimulates cellular proliferation, ERK1/2 signaling, and the expression of inflammatory and profibrotic mediators, including PAI-1, reinforcing its contribution to vascular injury and remodeling [18]. Collectively, these data indicate that while cardiac PRR predominantly influences myocardial structure and function, vascular PRR plays a more prominent role in the regulation of BP and the progression of vascular pathology, positioning PRR as a key molecular link between hemodynamic stress and CV remodeling.
In the kidney, PRR is abundantly expressed in collecting duct intercalated cells, podocytes, and mesangial cells, where it contributes to both renal sodium handling and glomerular integrity [19,20]. The PRR plays a critical role in nephrogenesis, as its deletion in nephron progenitor cells results in reduced nephron number, podocyte abnormalities, and proteinuria [21]. The PRR is thought to regulate kidney development partly through Wnt/β-catenin signaling, which coordinates ureteric bud branching and nephron formation. In the mature kidney, PRR contributes to the regulation of water and sodium homeostasis. PRR activation increases intrarenal RAAS and epithelial sodium channel (ENaC) activity, promoting sodium retention and the development of hypertension [4,19,22]. In models of diabetic nephropathy, PRR expression is markedly upregulated in glomerular cells, where it stimulates oxidative stress, extracellular matrix accumulation, and inflammatory signaling [10]. These findings, along with evidence that PRR activates Wnt/β-catenin signaling, highlight its critical role in driving renal fibrosis and promoting epithelial-mesenchymal transition during the progression of CKD.
2.1. Prorenin Receptor Signaling in Metabolic Homeostasis and Disease
The PRR in adipose tissue modulates biology and obesity-associated metabolic alterations. PRR expression is increased in adipose tissue under metabolic stress, including high-fat diet–induced obesity. PRR activation promotes adipocyte hypertrophy, inflammatory cytokine production, and oxidative stress, thereby contributing to adipose tissue dysfunction and systemic insulin resistance [23,24]. In addition, PRR has been implicated in the regulation of adipocyte metabolism through its association with the V-ATPase complex and Wnt/β-catenin signaling-pathways that are critical for adipocyte differentiation and energy homeostasis [23,25].
Growing evidence indicates that the soluble form of prorenin receptor (sPRR), generated by proteolytic cleavage of the membrane-bound receptor, is a circulating factor potentially involved in metabolic diseases. Elevated circulating levels of sPRR are present in patients with obesity, type 2 diabetes, and metabolic syndrome, suggesting that this form may serve as a potential biomarker or mediator of metabolic dysfunction [8]. The sPRR also influences glucose metabolism, adipose tissue inflammation, and insulin signaling, linking PRR activity to systemic metabolic regulation [23,26,27]. Collectively, these findings support the concept that PRR functions as an important molecular integrator connecting RAAS, cellular metabolism, and inflammatory pathways, thereby contributing to the development and progression of metabolic disorders.
2.2. Prorenin Receptor as a Regulator of Endothelial Signaling, Angiogenesis, and Vascular Remodeling
The PRR is a key modulator in developmental angiogenesis. Endothelial cell–specific deletion of PRR in mice leads to severe defects in retinal vascular development characterized by impaired endothelial cell migration, loss of tip cell polarity, and defective sprouting angiogenesis [28]. These abnormalities reflect dysregulation of genes governing cell migration, polarity, extracellular matrix organization, and angiogenic signaling, and indicate that PRR is critical for proper vascular network formation.
In addition to its role in endothelial signaling and angiogenesis, the Atp6ap2 gene has been increasingly recognized as an important contributor to vascular remodeling, a key pathological process underlying hypertension and CVD. Independent from Ang II, the activation of PRR by renin or prorenin initiates intracellular signaling pathways—including ERK1/2–MAPK, PI3K/Akt, and Wnt/β-catenin—that regulate VSMC proliferation, migration, and extracellular matrix production. [2,28,29]. These processes promote structural changes within the vascular wall, including media thickening, increased collagen deposition, and reduced vascular compliance, which contribute to elevated peripheral resistance and sustained hypertension.
During oxidative stress, the PRR mediates vascular injury by driving maladaptive vascular remodeling. PRR signaling can enhance the production of reactive oxygen species (ROS) through activation of NADPH oxidase pathways, leading to endothelial dysfunction and vascular inflammation [27,30]. These processes facilitate leukocyte recruitment, extracellular matrix reorganization, and vascular wall hypertrophy. PRR-associated signaling through V-ATPase and Wnt/β-catenin pathways may regulate gene programs involved in vascular cell growth and tissue remodeling, linking PRR to both developmental vascular patterning and pathological vascular remodeling in adult tissues [25,28]. These findings position PRR as a central molecular regulator of vascular remodeling, integrating RAAS-dependent and independent signaling that contribute to the structural and functional changes in hypertension and other CVD. Collectively, we propose that PRR functions as a molecular hub that integrates metabolic, developmental, and signaling pathways to regulate endothelial cell behavior, angiogenic remodeling, and vascular development.
2.3. Prorenin Receptor Pathogenic Feedback Loops and Chronic Kidney Disease
In addition to classical renin production by juxtaglomerular cells, renin and its precursor prorenin are synthesized by the distal nephron segments [6]. Prorenin produced by the collecting duct is regulated by hormones acting on the distal nephron rather than by BP [6,31]. Binding of PRR to renin and prorenin increases local renin enzymatic activity, activates prorenin, and augments intratubular Ang II formation. Increased distal prorenin production and secretion into the lumen has been demonstrated in several experimental hypertensive animal models, including chronic Ang II–infusion, 2-kidney 1-clip (2K1C) Goldblatt hypertension, and transgenic mice with renin deficiency in the collecting duct [6,19,31,32]. Likewise, these findings are present in mice with high fat diet-induced type II diabetes and rats with streptozotocin-induced type 1 diabetes [7,33].
The PRR activation by its natural agonists, renin and prorenin, promotes kidney fibrosis and inflammation through various pathways independent of the RAAS [6,10]. Its activation directly triggers intracellular signal transduction via ERK1/2 and upregulation of profibrotic genes, including TGF-β1, fibronectin, collagen I, and PAI-1 [7,34,35]. Likewise, PRR exerts a positive feedback action on the Wnt/β-catenin signaling, which exacerbates renal fibrosis and inflammation in hypertensive nephropathy [36]. This crosstalk establishes a feed-forward loop that sustains inflammatory and profibrotic signaling over time and accelerates maladaptive tissue remodeling. Consistent with this pathogenic role, experimental inhibition or knockdown of PRR in ischemia–reperfusion kidney injury models significantly attenuates renal fibrosis and further support its role as the key amplifier of fibrosis [10].
The PRR has gained significant attention as a potential therapeutic target for CV and renal diseases. The handle region peptide (HRP) was the first PRR-blocking peptide identified; however, its specificity in inhibiting PRR signaling has been widely debated [37]. In contrast, the PRR decoy inhibitor PRO20—composed of the first 20 amino acids of the prorenin pro-segment—has produced more consistent and less controversial results. Its efficacy has been validated in multiple rodent models of hypertension, fluid, and electrolyte imbalances [30,38,39,40,41]. These studies indicate that PRO20 disrupts the interaction between the pro-segment of prorenin and PRR without altering PRR expression, thus reducing inflammation and injury in models of Ang II-dependent hypertension and diabetic nephropathy.
2.4. Prorenin Receptor Contributes to Systemic and Renal Inflammation
Pathologic upregulation of PRR is strongly associated with enhanced renal and systemic inflammation in disease states including diabetes, hypertension, and renal ischemia. Increased PRR expression correlates with activation of ERK1/2 and p38 MAPK, as well as proinflammatory transcription factors such as NF-κB [35,40,42]. Activation of these pathways increases inflammatory mediators including TNF-α, IL-1β, MCP-1, and COX-2 [36,43], and promote immune cell recruitment and progressive renal injury independent of the RAAS. Although PRR overexpression in healthy animals does not independently induce hypertension or renal injury, it functions as a pathological amplifier to exacerbate inflammation and fibrosis under disease conditions [17]. In the 2K1C model of renovascular hypertension, PRR expression is selectively upregulated in the ischemic kidney, which is accompanied by marked increases in TNF-α, IL-1β, MCP-1, COX-2, phosphorylated NF-κB, and collagen type I, despite comparable elevations in intrarenal Ang II levels [10]. Furthermore, targeted intrarenal knockdown of PRR using shRNA significantly attenuates inflammatory cytokine expression, NF-κB activation, fibrosis, and albuminuria without altering BP or renal Ang II levels [35]. In addition, PRR inhibition can reduce inflammation and fibrosis in heart and liver disease models, supporting a broader role for PRR in systemic inflammatory signaling across organs [23,44].
3. The Soluble Prorenin Receptor: A Circulating Link Between the RAAS and Cardiorenal System
The discovery of the soluble form of the PRR (sPRR) has expanded the conceptual framework of PRR biology from a membrane-restricted receptor to a molecule capable of mediating local and systemic signaling effects. Functionally, sPRR retains the ability to bind prorenin and renin, thereby facilitating activation of the RAAS in extracellular compartments [6,45]. Beyond its role in RAAS activation, growing evidence also indicates that sPRR can directly modulate intracellular signaling pathways associated with inflammation, oxidative stress, and fibrosis [46,47]. The sPRR is produced by proteolytic cleavage of the full-length receptor by serine-proteases, including site-1 protease (SP-1), furin, and A Disintegrin And Metalloproteinase 19 (ADAM19) [26,45]. The released sPRR is detectable in plasma, cerebrospinal fluid, urine, and extracellular media [45,47]. It retains the ability to bind renin and prorenin, enhancing angiotensinogen conversion to Ang I and promoting downstream Ang II formation [47]. Although once thought to be just a by-product of receptor processing, growing evidence shows that sPRR is biologically active and has endocrine-like properties, enabling it to affect cellular signaling and systemic physiology.
In contrast to plasma renin activity, systemic sPRR levels may reflect circulating and tissue RAAS activation and indicate risk of CV and renal diseases. Increased circulating concentrations of sPRR have been found in individuals with hypertension, heart failure, and type 2 diabetes, correlating with indices of disease severity and target-organ damage [8,48]. Uremic toxins further potentiate PRR-dependent fibrosis. Indoxyl Sulfate (IS), a protein-bound uremic toxin that accumulates during CKD, upregulates PRR expression in mesangial cells and induces TGF-β, type IV collagen, and fibronectin expression via ERK1/2 activation. These effects are markedly suppressed by PRR knockdown [49]. IS also increases matrix metalloproteinase-9 (MMP9) and tissue inhibitor of metalloproteinase-1 (TIMP-1) expression, promoting ECM remodeling and mesangial expansion [34]. Clinically, circulating sPRR levels correlate with serum IS and histopathological indices of mesangial injury in CKD, supporting the concept that sPRR is a potential biomarker of mesangial fibrosis [34]. The fact that sPRR can directly promote fibrotic and oxidative stress responses via PI3K-Akt/β-catenin and NOX4-dependent pathways further implicate PRR signaling as a pathogenic driver of chronic tissue fibrosis [42,50].
In the kidney, sPRR plays a key role in the regulation of sodium and water balance. The sPRR stimulates sodium reabsorption in the distal nephron, partly through activation of ENaC in collecting duct epithelial cells. By this mechanism, it contributes to enhanced sodium retention and to the development of hypertension [19,51]. Additionally, sPRR may amplify intrarenal RAAS activity, creating a positive feedback loop that sustains renal inflammation and fibrosis in CKD. Elevated plasma sPRR levels have been reported in patients with CKD and diabetic nephropathy, suggesting that sPRR may serve as both a mediator and a biomarker of renal injury [8,52,53].
In the vasculature, sPRR promotes endothelial dysfunction and vascular remodeling by enhancing oxidative stress and inflammatory signaling [52]. In metabolic tissues such as the liver and adipose tissue, sPRR may influence glucose metabolism and insulin sensitivity, potentially linking PRR signaling to broader met.abolic dysregulation [23].
From a clinical perspective, circulating sPRR has gained attention as a potential biomarker for cardiorenal disease progression. Measurement of plasma sPRR levels may provide insights into the activation of tissue RAAS and associated pathological processes, including fibrosis, inflammation, and sodium retention. Because sPRR reflects both PRR expression and proteolytic processing, it may serve as an integrative indicator of PRR pathway activation across multiple organs. Although sPRR is not currently adopted in the clinical practice, accumulating data supports its value to help identify patients at risk for CVD, progressive kidney disease, resistant hypertension, or adverse cardiovascular outcomes.
4. Conclusions
Over the past two decades, the PRR has been established as a multifunctional regulator that extends far beyond its functions within the RAAS. The PRR is an important molecular hub integrating RAAS-dependent and -independent signaling pathways, influencing diverse cellular processes including endothelial signaling, angiogenesis, vascular remodeling, metabolic regulation, and renal homeostasis. Through PRR-dependent activation of intracellular pathways such as ERK1/2-MAPK, PI3K/Akt, and Wnt/β-catenin, and its interaction with the V-ATPase complex; PRR regulates cellular proliferation, inflammation, oxidative stress, and fibrosis—key mechanisms underlying the pathogenesis of CV, metabolic, and renal diseases.
Emerging studies suggest that sPRR may act as a circulating mediator, linking tissue RAAS activity to systemic metabolic and cardiorenal dysfunction. The discovery of sPRR has significantly broadened the understanding of PRR biology. Rather than functioning solely as a membrane receptor, the PRR now appears to operate through both membrane-bound and soluble signaling mechanisms, allowing communication between organs such as the kidney, heart, vasculature, and metabolic tissues. New insights into PRR biology offer the potential for novel therapeutic interventions to treat cardiometabolic and cardiorenal diseases by targeting PRR-associated pathways. Further research will be necessary to clarify the tissue-specific functions of PRR and to determine how modulation of PRR/sPRR signaling can be translated into effective clinical strategies. Future studies into the regulation of sPRR generation, signaling mechanisms, and clinical associations will be crucial for determining whether targeting the PRR–sPRR axis may provide new diagnostic or therapeutic opportunities in CV, metabolic, and renal diseases
Author Contributions
M.C.P., conceived and designed the manuscript; M.C.P., A.S.M-B., S.E.T., J.D.P., Z.V., B.A., B.C., contributed with some previously generated experimental data and drafted the manuscript; A.S.M-B., prepared the figure; and A.S.M-B., and M.C.P., edited and approved final version of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
M.C.P., was partially supported by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP 2017/17027-0) and the Carol Lavin-Bernick Faculty Grant Award from Tulane University. J.D.P., was a recipient of the The Ramadhyani-Bourgeois MD Student Endowed Award.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study.
Acknowledgments
The authors thank Tulane University School of Medicine for institutional support.
Conflicts of Interest
The authors declare that there are no potential conflicts of interest relating to this manuscript.
Abbreviations
| ADAM19 | A Disintegrin And Metalloproteinase 19 |
| Akt | Protein kinase B |
| Ang I | Angiotensin I |
| Ang II | Angiotensin II |
| AP-1 | Activator protein-1 |
| ATP6AP2 | ATPase H+-transporting accessory protein 2 (gene encoding PRR) |
| BP | Blood pressure |
| CKD | Chronic kidney disease |
| COX-2 | Cyclooxygenase-2 |
| CREB | cAMP response element-binding protein |
| CVD | Cardiovascular disease |
| ECM | Extracellular matrix |
| ENaC | Epithelial sodium channel |
| ERK1/2 | Extracellular signal-regulated kinase 1/2 |
| HSP27 | Heat shock protein 27 |
| HRP | Handle region peptide |
| IL-1β | Interleukin-1 beta |
| IS | Indoxyl sulfate |
| MAPK | Mitogen-activated protein kinase |
| MCP-1 | Monocyte chemoattractant protein-1 |
| MMP9 | Matrix metalloproteinase-9 |
| NF-κB | Nuclear factor κB |
| NOX4 | NADPH oxidase 4 |
| p38 MAPK | p38 mitogen-activated protein kinase |
| PAI-1 | Plasminogen activator inhibitor-1 |
| PI3K | Phosphoinositide 3-kinase |
| PRR | Prorenin receptor |
| PRO20 | PRR decoy inhibitor derived from the first 20 amino acids of the prorenin pro-segment |
| RAAS | Renin–angiotensin–aldosterone system |
| ROS | Reactive oxygen species |
| sPRR | Soluble prorenin receptor |
| SP-1 | Site-1 protease |
| TGF-β | Transforming growth factor-β |
| TIMP1 | Tissue inhibitor of metalloproteinase-1 |
| TNF-α | Tumor necrosis factor-α |
| V-ATPase | Vacuolar H+-ATPase |
| VSMC | Vascular smooth muscle cell |
| Wnt/β-catenin | Wnt/β-catenin signaling pathway |
| 2K1C | Two-kidney, one-clip (Goldblatt hypertension model) |
References
- Crowley, S.D.; Navar, L.G.; Prieto, M.C.; Gurley, S.B.; Coffman, T.M. Kidney Renin–Angiotensin System: Lost in a RAS Cascade. Hypertension 2024, 81, 682–686. [Google Scholar] [CrossRef]
- Nguyen, G.; Delarue, F.; Burcklé, C.; Bouzhir, L.; Giller, T.; Sraer, J.-D. Pivotal Role of the Renin/Prorenin Receptor in Angiotensin II Production and Cellular Responses to Renin. J. Clin. Invest. 2002, 109, 1417–1427. [Google Scholar] [CrossRef] [PubMed]
- Advani, A.; Kelly, D.J.; Cox, A.J.; White, K.E.; Advani, S.L.; Thai, K.; Connelly, K.A.; Yuen, D.; Trogadis, J.; Herzenberg, A.M.; et al. The (Pro)Renin Receptor: Site-Specific and Functional Linkage to the Vacuolar H+ -ATPase in the Kidney. Hypertension 2009, 54, 261–269. [Google Scholar] [CrossRef]
- Gonzalez, A.A.; Lara, L.S.; Luffman, C.; Seth, D.M.; Prieto, M.C. Soluble Form of the (pro)Renin Receptor Is Augmented in the Collecting Duct and Urine of Chronic Angiotensin II-Dependent Hypertensive Rats. Hypertension 2011, 57, 859–864. [Google Scholar] [CrossRef] [PubMed]
- Ramkumar, N.; Stuart, D.; Peterson, C.S.; Hu, C.; Wheatley, W.; Min Cho, J.; Symons, J.D.; Kohan, D.E. Loss of Soluble (Pro)Renin Receptor Attenuates Angiotensin-II Induced Hypertension and Renal Injury. Circulation Research 2021, 129, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Prieto, M.C.; Gonzalez, A.A.; Visniauskas, B.; Navar, L.G. The Evolving Complexity of the Collecting Duct Renin-Angiotensin System in Hypertension. Nat Rev Nephrol 2021, 17, 481–492. [Google Scholar] [CrossRef]
- Gogulamudi, V.R.; Arita, D.Y.; Bourgeois, C.R.T.; Jorgensen, J.; He, J.; Wimley, W.C.; Satou, R.; Gonzalez, A.A.; Prieto, M.C. High Glucose Induces Trafficking of Prorenin Receptor and Stimulates Profibrotic Factors in the Collecting Duct. Sci Rep 2021, 11, 13815. [Google Scholar] [CrossRef]
- Visniauskas, B.; Arita, D.Y.; Rosales, C.B.; Feroz, M.A.; Luffman, C.; Accavitti, M.J.; Dawkins, G.; Hong, J.; Curnow, A.C.; Thethi, T.K.; et al. Sex Differences in Soluble Prorenin Receptor in Patients with Type 2 Diabetes. Biol Sex Differ 2021, 12, 33. [Google Scholar] [CrossRef]
- Wang, B.; Jie, H.; Wang, S.; Dong, B.; Zou, Y. The Role of (pro)Renin Receptor and Its Soluble Form in Cardiovascular Diseases. Front. Cardiovasc. Med. 2023, 10, 1086603. [Google Scholar] [CrossRef]
- Ichihara, A.; Yatabe, M.S. The (pro)Renin Receptor in Health and Disease. Nat Rev Nephrol 2019, 15, 693–712. [Google Scholar] [CrossRef]
- Nakagawa, T.; Suzuki-Nakagawa, C.; Watanabe, A.; Asami, E.; Matsumoto, M.; Nakano, M.; Ebihara, A.; Uddin, M.N.; Suzuki, F. Site-1 Protease Is Required for the Generation of Soluble (pro)Renin Receptor. The Journal of Biochemistry 2017, 161, 369–379. [Google Scholar] [CrossRef]
- Figueiredo, M.; Daryadel, A.; Sihn, G.; Müller, D.N.; Popova, E.; Rouselle, A.; Nguyen, G.; Bader, M.; Wagner, C.A. The (pro)Renin Receptor (ATP6ap2) Facilitates Receptor-Mediated Endocytosis and Lysosomal Function in the Renal Proximal Tubule. Pflugers Arch 2021, 473, 1229–1246. [Google Scholar] [CrossRef] [PubMed]
- Kinouchi, K.; Ichihara, A.; Sano, M.; Sun-Wada, G.-H.; Wada, Y.; Kurauchi-Mito, A.; Bokuda, K.; Narita, T.; Oshima, Y.; Sakoda, M.; et al. The (Pro)Renin Receptor/ATP6AP2 Is Essential for Vacuolar H+ -ATPase Assembly in Murine Cardiomyocytes. Circulation Research 2010, 107, 30–34. [Google Scholar] [CrossRef]
- Li, L.; Cui, Y.; Liu, Y.; Li, H.; Su, Y.; Li, S.; Wang, L.; Zhao, Y.; Wang, S.; Yan, F.; et al. ATP6AP2 Knockdown in Cardiomyocyte Deteriorates Heart Function via Compromising Autophagic Flux and NLRP3 Inflammasome Activation. Cell Death Discov. 2022, 8, 161. [Google Scholar] [CrossRef]
- Hayakawa, Y.; Aoyama, T.; Yokoyama, C.; Okamoto, C.; Komaki, H.; Minatoguchi, S.; Iwasa, M.; Yamada, Y.; Kawamura, I.; Kawasaki, M.; et al. High Salt Intake Damages the Heart through Activation of Cardiac (Pro) Renin Receptors Even at an Early Stage of Hypertension. PLoS ONE 2015, 10, e0120453. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, C.; Hayakawa, Y.; Aoyama, T.; Komaki, H.; Minatoguchi, S.; Iwasa, M.; Yamada, Y.; Kanamori, H.; Kawasaki, M.; Nishigaki, K.; et al. Excessively Low Salt Diet Damages the Heart through Activation of Cardiac (pro) Renin Receptor, Renin-Angiotensin-Aldosterone, and Sympatho-Adrenal Systems in Spontaneously Hypertensive Rats. PLoS ONE 2017, 12, e0189099. [Google Scholar] [CrossRef]
- Moilanen, A.-M.; Rysä, J.; Serpi, R.; Mustonen, E.; Szabò, Z.; Aro, J.; Näpänkangas, J.; Tenhunen, O.; Sutinen, M.; Salo, T.; et al. (Pro)Renin Receptor Triggers Distinct Angiotensin II-Independent Extracellular Matrix Remodeling and Deterioration of Cardiac Function. PLoS ONE 2012, 7, e41404. [Google Scholar] [CrossRef]
- Ma, J.; Li, Y.; Yang, X.; Liu, K.; Zhang, X.; Zuo, X.; Ye, R.; Wang, Z.; Shi, R.; Meng, Q.; et al. Signaling Pathways in Vascular Function and Hypertension: Molecular Mechanisms and Therapeutic Interventions. Sig Transduct Target Ther 2023, 8, 168. [Google Scholar] [CrossRef]
- Prieto, M.C.; Reverte, V.; Mamenko, M.; Kuczeriszka, M.; Veiras, L.C.; Rosales, C.B.; McLellan, M.; Gentile, O.; Jensen, V.B.; Ichihara, A.; et al. Collecting Duct Prorenin Receptor Knockout Reduces Renal Function, Increases Sodium Excretion, and Mitigates Renal Responses in ANG II-Induced Hypertensive Mice. American Journal of Physiology-Renal Physiology 2017, 313, F1243–F1253. [Google Scholar] [CrossRef] [PubMed]
- Peng, K.; Lu, X.; Wang, F.; Nau, A.; Chen, R.; Zhou, S.-F.; Yang, T. Collecting Duct (pro)Renin Receptor Targets ENaC to Mediate Angiotensin II-Induced Hypertension. American Journal of Physiology-Renal Physiology 2017, 312, F245–F253. [Google Scholar] [CrossRef]
- Yosypiv, I.V.; Sequeira-Lopez, M.L.S.; Song, R.; De Goes Martini, A. Stromal Prorenin Receptor Is Critical for Normal Kidney Development. Am J Physiol Regul Integr Comp Physiol 2019, 316, R640–R650. [Google Scholar] [CrossRef]
- Wang, F.; Lu, X.; Peng, K.; Fang, H.; Zhou, L.; Su, J.; Nau, A.; Yang, K.T.; Ichihara, A.; Lu, A.; et al. Antidiuretic Action of Collecting Duct (Pro)Renin Receptor Downstream of Vasopressin and PGE2 Receptor EP4. JASN 2016, 27, 3022–3034. [Google Scholar] [CrossRef]
- Gatineau, E.; Arthur, G.; Poupeau, A.; Nichols, K.; Spear, B.T.; Shelman, N.R.; Graf, G.A.; Temel, R.E.; Yiannikouris, F.B. The Prorenin Receptor and Its Soluble Form Contribute to Lipid Homeostasis. American Journal of Physiology-Endocrinology and Metabolism 2021, 320, E609–E618. [Google Scholar] [CrossRef]
- Nichols, K.; Yiannikouris, F. The Role of (Pro)Renin Receptor in the Metabolic Syndrome. CHYR 2022, 18, 117–124. [Google Scholar] [CrossRef]
- Hoffmann, N.; Peters, J. Functions of the (pro)Renin Receptor (Atp6ap2) at Molecular and System Levels: Pathological Implications in Hypertension, Renal and Brain Development, Inflammation, and Fibrosis. Pharmacological Research 2021, 173, 105922. [Google Scholar] [CrossRef] [PubMed]
- Cousin, C.; Bracquart, D.; Contrepas, A.; Corvol, P.; Muller, L.; Nguyen, G. Soluble Form of the (Pro)Renin Receptor Generated by Intracellular Cleavage by Furin Is Secreted in Plasma. Hypertension 2009, 53, 1077–1082. [Google Scholar] [CrossRef] [PubMed]
- Visniauskas, B.; Reverte, V.; Abshire, C.M.; Ogola, B.O.; Rosales, C.B.; Galeas-Pena, M.; Sure, V.N.; Sakamuri, S.S.V.P.; Harris, N.R.; Kilanowski-Doroh, I.; et al. High-Plasma Soluble Prorenin Receptor Is Associated with Vascular Damage in Male, but Not Female, Mice Fed a High-Fat Diet. American Journal of Physiology-Heart and Circulatory Physiology 2023, 324, H762–H775. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.R.; K C, R.; Blanks, A.; Li, Y.; Prieto, M.C.; Meadows, S.M. Endothelial Cell Polarity and Extracellular Matrix Composition Require Functional ATP6AP2 during Developmental and Pathological Angiogenesis. JCI Insight 2022, 7, e154379. [Google Scholar] [CrossRef]
- Sakoda, M.; Ichihara, A.; Kaneshiro, Y.; Takemitsu, T.; Nakazato, Y.; Nabi, A.H.M.N.; Nakagawa, T.; Suzuki, F.; Inagami, T.; Itoh, H. (Pro)Renin Receptor-Mediated Activation of Mitogen-Activated Protein Kinases in Human Vascular Smooth Muscle Cells. Hypertens Res 2007, 30, 1139–1146. [Google Scholar] [CrossRef]
- Luo, R.; Yang, K.; Wang, F.; Xu, C.; Yang, T. (Pro)Renin Receptor Decoy Peptide PRO20 Protects against Adriamycin-Induced Nephropathy by Targeting the Intrarenal Renin-Angiotensin System. American Journal of Physiology-Renal Physiology 2020, 319, F930–F940. [Google Scholar] [CrossRef]
- Lara, L.S.; Gonzalez, A.A.; Hennrikus, M.T.; Prieto, M.C. Hormone-Dependent Regulation of Renin and Effects on ProreninReceptor Signaling in the Collecting Duct. CHYR 2022, 18, 91–100. [Google Scholar] [CrossRef]
- Fu, Z.; Kaewsaro, K.; Soodvilai, S.; Kimball, A.; Yang, T. Collecting Duct-Derived (pro)Renin Receptor Contributes to 2-Kidney, 1-Clip-Induced Ischemic Nephropathy and Renovascular Hypertension in Mice. American Journal of Physiology-Renal Physiology 2025, 329, F46–F58. [Google Scholar] [CrossRef] [PubMed]
- Reverte, V.; Gogulamudi, V.R.; Rosales, C.B.; Musial, D.C.; Gonsalez, S.R.; Parra-Vitela, A.J.; Galeas-Pena, M.; Sure, V.N.; Visniauskas, B.; Lindsey, S.H.; et al. Urinary Angiotensinogen Increases in the Absence of Overt Renal Injury in High Fat Diet-Induced Type 2 Diabetic Mice. J Diabetes Complications 2020, 34, 107448. [Google Scholar] [CrossRef] [PubMed]
- Narumi, K.; Sato, E.; Hirose, T.; Yamamoto, T.; Nakamichi, T.; Miyazaki, M.; Sato, H.; Ito, S. (Pro)Renin Receptor Is Involved in Mesangial Fibrosis and Matrix Expansion. Sci Rep 2018, 8, 16. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.A.; Zamora, L.; Reyes-Martinez, C.; Salinas-Parra, N.; Roldan, N.; Cuevas, C.A.; Figueroa, S.; Gonzalez-Vergara, A.; Prieto, M.C. (Pro)Renin Receptor Activation Increases Profibrotic Markers and Fibroblast-like Phenotype through MAPK-Dependent ROS Formation in Mouse Renal Collecting Duct Cells. Clin Exp Pharmacol Physiol 2017, 44, 1134–1144. [Google Scholar] [CrossRef]
- Lin, M.; Wang, D.; Chen, Y.; Chen, G.; Zhou, Y.; Ou, J.; Xiao, L. PRR Promotes Hypertensive Renal Injury by Activating Wnt/β-Catenin Signaling and Inflammation Infiltration in Mice. Biochim Biophys Acta Mol Basis Dis 2025, 1871, 167517. [Google Scholar] [CrossRef]
- Feldt, S.; Maschke, U.; Dechend, R.; Luft, F.C.; Muller, D.N. The Putative (Pro)Renin Receptor Blocker HRP Fails to Prevent (Pro)Renin Signaling. Journal of the American Society of Nephrology 2008, 19, 743–748. [Google Scholar] [CrossRef]
- Li, W.; Sullivan, M.N.; Zhang, S.; Worker, C.J.; Xiong, Z.; Speth, R.C.; Feng, Y. Intracerebroventricular Infusion of the (Pro)Renin Receptor Antagonist PRO20 Attenuates Deoxycorticosterone Acetate-Salt–Induced Hypertension. Hypertension 2015, 65, 352–361. [Google Scholar] [CrossRef]
- Fang, H.; Yang, T.; Zhou, B.; Li, X. (Pro)Renin Receptor Decoy Peptide PRO20 Protects against Oxidative Renal Damage Induced by Advanced Oxidation Protein Products. Molecules 2023, 28, 3017. [Google Scholar] [CrossRef]
- Larenas, P.E.; Cárdenas, P.; Aguirre-Delgadillo, M.; Moris, C.; Casarini, D.E.; Vallotton, Z.; Prieto, M.C.; Gonzalez, A.A. GLUT1 and Prorenin Receptor Mediate Differential Regulation of TGF-β and CTGF in Renal Inner Medullary Collecting Duct Cells during High Glucose Conditions. Biol Res 2024, 57, 81. [Google Scholar] [CrossRef]
- Cárdenas, P.; Cid-Salinas, C.; León, A.C.; Castillo-Geraldo, J.; De Oliveira, L.C.G.; Yokota, R.; Vallotton, Z.; Casarini, D.E.; Prieto, M.C.; Lorca, R.A.; et al. (Pro)Renin Receptor Blockade Prevents Increases in Systolic Blood Pressure, Sodium Retention, and αENaC Protein Expression in the Kidney of 2K1C Goldblatt Mice. IJMS 2025, 26, 4177. [Google Scholar] [CrossRef]
- Ramkumar, N.; Kohan, D.E. The (pro)Renin Receptor: An Emerging Player in Hypertension and Metabolic Syndrome. Kidney Int 2019, 95, 1041–1052. [Google Scholar] [CrossRef]
- Gonzalez, A.A.; Luffman, C.; Bourgeois, C.R.T.; Vio, C.P.; Prieto, M.C. Angiotensin II–Independent Upregulation of Cyclooxygenase-2 by Activation of the (Pro)Renin Receptor in Rat Renal Inner Medullary Cells. Hypertension 2013, 61, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Ellmers, L.J.; Rademaker, M.T.; Charles, C.J.; Yandle, T.G.; Richards, A.M. (Pro)Renin Receptor Blockade Ameliorates Cardiac Injury and Remodeling and Improves Function After Myocardial Infarction. J Card Fail 2016, 22, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Peng, K.; Luo, R.; Wang, F.; Yang, T. Site-1 Protease-Derived Soluble (Pro)Renin Receptor Contributes to Angiotensin II–Induced Hypertension in Mice. Hypertension 2021, 77, 405–416. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Liu, C.; Xiong, J.; Yu, J. Cardiovascular Aspects of the (pro)Renin Receptor: Function and Significance. The FASEB Journal 2022, 36, e22237. [Google Scholar] [CrossRef]
- Yang, T. Soluble (Pro)Renin Receptor in Hypertension. Nephron 2023, 147, 234–243. [Google Scholar] [CrossRef]
- Watanabe, N.; Bokuda, K.; Fujiwara, T.; Suzuki, T.; Mito, A.; Morimoto, S.; Jwa, S.C.; Egawa, M.; Arai, Y.; Suzuki, F.; et al. Soluble (Pro)Renin Receptor and Blood Pressure During Pregnancy: A Prospective Cohort Study. Hypertension 2012, 60, 1250–1256. [Google Scholar] [CrossRef]
- Yisireyili, M.; Saito, S.; Abudureyimu, S.; Adelibieke, Y.; Ng, H.-Y.; Nishijima, F.; Takeshita, K.; Murohara, T.; Niwa, T. Indoxyl Sulfate-Induced Activation of (pro)Renin Receptor Promotes Cell Proliferation and Tissue Factor Expression in Vascular Smooth Muscle Cells. PLoS One 2014, 9, e109268. [Google Scholar] [CrossRef]
- Li, Z.; Zhou, L.; Wang, Y.; Miao, J.; Hong, X.; Hou, F.F.; Liu, Y. (Pro)Renin Receptor Is an Amplifier of Wnt/β-Catenin Signaling in Kidney Injury and Fibrosis. J Am Soc Nephrol 2017, 28, 2393–2408. [Google Scholar] [CrossRef]
- Yang, T.; Gao, Z.-X.; Mao, Z.-H.; Wu, P. Soluble (pro)Renin Receptor as a Novel Regulator of Renal Medullary Na+ Reabsorption. American Journal of Physiology-Renal Physiology 2025, 328, F239–F247. [Google Scholar] [CrossRef] [PubMed]
- Visniauskas, B.; Ogola, B.O.; Kilanowski-Doroh, I.; Harris, N.R.; Diaz, Z.T.; Horton, A.C.; Blessinger, S.A.; McNally, A.B.; Zimmerman, M.A.; Arnold, A.C.; et al. Hypertension Disrupts the Vascular Clock in Both Sexes. American Journal of Physiology-Heart and Circulatory Physiology 2024, 327, H765–H777. [Google Scholar] [CrossRef] [PubMed]
- Yang, T. Potential of Soluble (pro)Renin Receptor in Kidney Disease: Can It Go beyond a Biomarker? American Journal of Physiology-Renal Physiology 2022, 323, F507–F514. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



