Submitted:
07 April 2026
Posted:
09 April 2026
You are already at the latest version
Abstract
Copper supplementation in European pig production selects for the pandemic multidrug-resistant (MDR) monophasic Salmonella Typhimurium ST34 clone through Salmonella Genomic Island 4, which co-carries copper/silver resistance and antimicrobial resistance determinants on the same mobile genetic element — a paradigmatic example of non-antibiotic co-selective pressure connecting animal health, food safety, and public health under the One Health paradigm. EU Regulation 2025/179, which mandates whole-genome sequencing for foodborne outbreak investigation from August 2026, will generate an unprecedented genomic data infrastructure across Europe; this infrastructure contains, as an inherent by-product of sequencing, the metal resistance markers that would enable quantification of such co-selective pressures — a capability that extends beyond the outbreak detection purpose for which it was conceived. Multi-dimensional genomic risk assessment frameworks integrating virulence, environmental persistence, and antimicrobial resistance into quantitative actionable indices represent a pathway to realise this opportunity, connecting copper reduction in animal feed (Regulation 2018/1039) with genomic surveillance (Regulation 2025/179) along a One Health continuum from farm to patient.
Keywords:
antimicrobial resistance
; copper co-selection
; Salmonella Typhimurium ST34
; whole-genome sequencing
; One Health
; food safety surveillance
; metal resistance
; EU regulation
Copper as a One Health Co-Selective Pressure
Antimicrobial resistance (AMR) is recognised as a One Health challenge in which agricultural practices, food safety, and human medicine are interconnected. In this context, non-antibiotic selective pressures — heavy metals, biocides, disinfectants — have received increasing attention as drivers of co-selection and co-resistance. Copper represents perhaps the most thoroughly documented case.
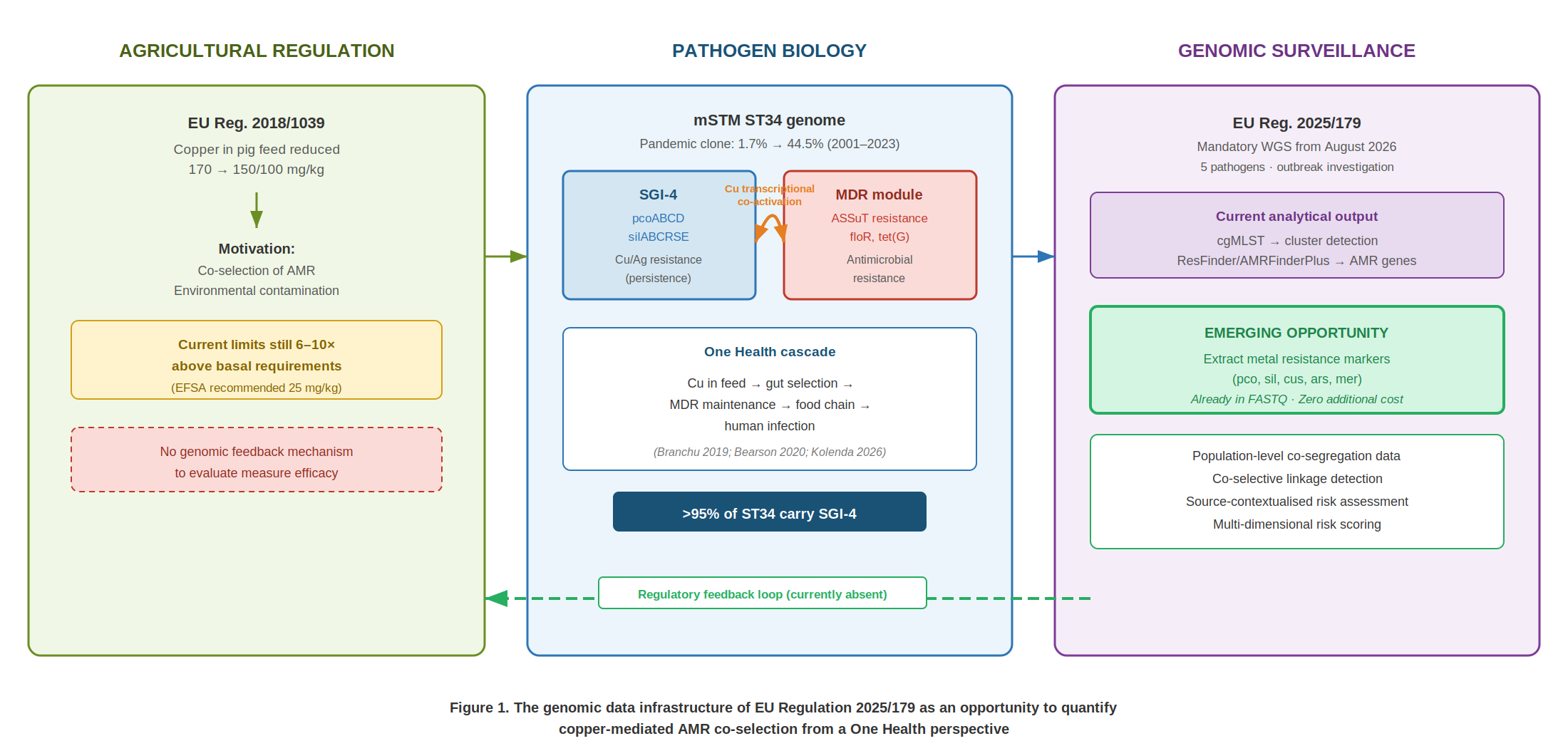
The monophasic variant of Salmonella enterica serovar Typhimurium sequence type 34 (mSTM ST34) has undergone extraordinary global expansion, rising from 1.7% of human S. Typhimurium isolates in 2001–2005 to 44.5% in 2016–2023, with the sharpest increase observed in Europe (Chen et al. 2025). This pandemic clone, strongly associated with pig production, carries a characteristic integrative conjugative element designated Salmonella Genomic Island 4 (SGI-4), present in over 95% of the ST34 clade (Branchu et al. 2019). SGI-4 encodes copper and silver resistance determinants (sil and pco gene clusters) that confer enhanced survival under the copper concentrations encountered in the pig intestinal tract, particularly under anaerobic conditions, where the minimum inhibitory concentrations for SGI-4-negative strains fall within the range measured in manure effluent from farms using copper-supplemented diets (Branchu et al. 2019).
The causal chain has recently been established in a mechanistically compelling manner. Kolenda, Kingsley and colleagues demonstrated through on-farm shotgun metagenomics that therapeutic copper levels in feed reduce key intestinal competitors of Salmonella — including Bifidobacterium, Escherichia, and Lactobacillus — creating an ecological niche that selectively favours SGI-4-bearing ST34 (Kolenda et al. 2026). Complementarily, Bearson and colleagues documented that copper exposure transcriptionally induces not only the metal tolerance genes within SGI-4, but also mercury tolerance genes located on the physically separate MDR module encoding aminoglycoside, sulphonamide, and tetracycline resistance (Bearson et al. 2020). Copper, therefore, does not merely select for copper-resistant Salmonella: it transcriptionally activates the maintenance of MDR determinants on a separate genetic element within the same genome. This cascade — animal feed → intestinal selection → MDR maintenance → foodborne transmission → human infection — is intrinsically One Health.
The Genomic Infrastructure of Regulation 2025/179: an Emerging Opportunity
The European Union has acted at both ends of this chain. Regulation 2018/1039, effective from August 2019, reduced maximum permitted copper levels in piglet feed from 170 to 150/100 mg/kg, motivated explicitly by concerns over AMR co-selection and environmental contamination, although the levels in force remain 6–10 times above basal nutritional requirements (EFSA 2016). At the surveillance end, Regulation 2025/179, applicable from August 2026, mandates whole-genome sequencing (WGS) for isolates of five key foodborne pathogens (Salmonella enterica, Listeria monocytogenes, Escherichia coli, Campylobacter jejuni and C. coli) obtained during foodborne outbreak investigations, with mandatory transmission of raw reads, genome assemblies, and typing results to EFSA.
The explicit and legitimate purpose of Regulation 2025/179 is the epidemiological detection of clusters and the identification of outbreak sources. The analytical pipelines supporting this objective are centred on two capabilities: high-resolution typing through core-genome MLST (cgMLST) for the detection of clonal relationships, and identification of acquired antimicrobial resistance genes through tools such as ResFinder or AMRFinderPlus.
However, the implementation of this regulation will generate something that transcends its immediate objective: a continental-scale infrastructure of complete genomic data. Every FASTQ file generated under this regulation will contain, in addition to the information required for cgMLST and AMR detection, the complete sequence of all metal resistance genes present in the isolate — pcoA, pcoD, silA, silC, arsB, merA, among others. This information, intrinsic to whole-genome sequencing, will be available at no additional sequencing or laboratory cost.
Herein lies a significant opportunity. The same infrastructure that enables outbreak detection contains the data necessary to quantify co-selective pressures at the population level — precisely the pressures that motivated the copper reduction of Regulation 2018/1039. Realising this opportunity requires only expanding the bioinformatic processing of data that are already being generated.
From Gene Cataloguing to Multi-Dimensional Risk Assessment
A key aspect that this opportunity highlights is the inherent limitation of one-dimensional approaches to genomic risk assessment. Two isolates with identical AMR gene profiles (e.g., blaTEM-1, aadA2, sul1, tet(A)) are classified identically by current pipelines. Yet if one additionally carries SGI-4, its probability of persisting in copper-exposed environments and of maintaining its MDR determinants through copper-mediated transcriptional co-activation is quantifiably greater (Bearson et al. 2020). The selective pressure maintaining MDR in that isolate is not antibiotics — it is a metal legally permitted as a feed additive. An approach that only catalogues AMR genes cannot distinguish these fundamentally different risk profiles.
This type of limitation underscores the value of multi-dimensional genomic assessment frameworks, in which WGS information is processed across multiple biological axes simultaneously — not only resistance, but also virulence and environmental persistence — to generate integrated risk assessments (Gamboa 2026). Under such approaches, metal resistance determinants contribute to a persistence dimension that, when co-elevated with antimicrobial resistance markers in the same genome, signals co-selective maintenance — a risk that current one-dimensional approaches do not capture. This multi-component integration capability is especially relevant in the One Health context, where cross-environmental selective pressures (farm, food, patient) require analytical frameworks capable of handling multiple biological dimensions simultaneously.
Opportunities for Integrated Genomic Surveillance
The temporal convergence between the regulatory reduction of copper (2018/1039) and mandatory WGS (2025/179) opens several concrete opportunities worth identifying.
First, routine extraction of metal resistance determinants (pco, sil, cus, ars, mer operons) from the WGS data already being generated would enable, at a continental scale, quantification of the prevalence of metal–AMR co-segregation and its temporal evolution. This information does not currently exist at the population level.
Second, identification of the co-localisation of these determinants with AMR genes on the same mobile genetic element (genomic island, plasmid, ICE) would allow distinction between incidental co-occurrence and co-selective linkage — a fundamental distinction for risk assessment.
Third, contextualisation with the source metadata that Regulation 2025/179 itself requires to be reported (including source and sampling circumstances) would add an essential ecological layer: an mSTM ST34 isolate bearing SGI-4 from a porcine source carries a different risk interpretation than the same genotype isolated from poultry, given the specificity of copper selective pressure in swine production.
Finally, aggregated metal resistance prevalence data obtained from genomic surveillance could feed back into the review cycles of Regulation 2018/1039, providing population-level genomic evidence on the efficacy of current copper limits in reducing co-selective pressure — a regulatory feedback loop that does not currently exist.
Conclusion
The implementation of EU Regulation 2025/179 represents a transformative advance in the surveillance of foodborne pathogens in Europe. Its most evident contribution will be the detection of epidemiological clusters at unprecedented resolution. But the genomic data infrastructure that this regulation creates also offers an opportunity to go further: to quantify the non-antibiotic selective pressures that maintain antimicrobial resistance along the food chain, connecting microbiological surveillance with the regulation of agricultural practices along a One Health continuum. Multi-dimensional genomic assessment frameworks integrating persistence, virulence, and resistance offer a pathway to realise this opportunity, translating the richness of WGS data into risk assessments that capture biological dimensions currently present in the FASTQ files but not yet being extracted.
Conflict of interest
The author declares no conflict of interest.
References
- Bearson, BL; Trachsel, JM; Shippy, DC; Sivasankaran, SK; Kerr, BJ; Loving, CL; Brunelle, BW; Curry, SM; Bearson, SMD. The role of Salmonella Genomic Island 4 in metal tolerance of Salmonella enterica serovar I 4,[5],12:i:- pork outbreak isolate USDA15WA-1. Genes 2020, 11, 1291. [Google Scholar] [CrossRef] [PubMed]
- Branchu, P; Charity, OJ; Bawn, M; Thilliez, G; Dallman, TJ; Petrovska, L; Kingsley, RA. SGI-4 in monophasic Salmonella Typhimurium ST34 is a novel ICE that enhances resistance to copper. Front Microbiol 2019, 10, 1118. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y; Jiang, Y; Li, X; et al. Genomic epidemiology and public health implications of zoonotic monophasic Salmonella Typhimurium ST34. Front Cell Infect Microbiol 2025, 15, 1490183. [Google Scholar]
- European Commission. Commission Implementing Regulation (EU) 2018/1039. Off J Eur Union 2018, L186, 3–4. [Google Scholar]
- European Commission (2025) Commission Implementing Regulation (EU) 2025/179 on the collection and transmission of molecular analytical data within the frame of epidemiological investigations of food-borne outbreaks. Off J Eur Union L series.
- European Food Safety Authority. Revision of the currently authorised maximum copper content in complete feed. EFSA J 2016, 14(8), 4563. [Google Scholar]
- Gamboa, J. Genomic Intelligence Framework: multi-dimensional trophic classification of Listeria monocytogenes from whole-genome sequencing data reveals amphitrophic strains as the dominant cause of food-chain persistence and clinical virulence. bioRxiv 2026. [Google Scholar] [CrossRef]
- Kolenda, R; Hassan, MM; Arrieta-Gisasola, A; Kamara, A; Ansorge, R; Sidorczuk, K; Acton, L; Thilliez, G; Baker, DJ; Burdukiewicz, M; Stares, MD; Browne, HP; Le Gall, G; Torres, RC; Chavez-Arroyo, A; Garrett, J; Stevens, MP; Lawley, TD; Baumler, AJ; La Ragione, R; Hildebrand, F; Kingsley, RA. Copper is an intestinal habitat filter affecting the gut microbiota interactions with Salmonella Typhimurium. Microbiome 2026. [Google Scholar] [CrossRef] [PubMed]
- Mourao, J; Novais, C; Machado, J; Peixe, L; Antunes, P. Metal tolerance in emerging clinically relevant multidrug-resistant Salmonella enterica serotype 4,[5],12:i:- clones circulating in Europe. Int J Antimicrob Agents 2015, 45, 610–616. [Google Scholar] [CrossRef] [PubMed]
- Petrovska, L; Mather, AE; AbuOun, M; et al. Microevolution of monophasic Salmonella Typhimurium during epidemic, United Kingdom, 2005–2010. Emerg Infect Dis 2016, 22, 617–624. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



