Submitted:
05 April 2026
Posted:
06 April 2026
You are already at the latest version
Abstract
A single paragraph of about 200 words maximum. For research articles, abstracts should give a pertinent overview of the work. We strongly encourage authors to use the following style of structured abstracts, but without headings: (1) Background: Place the question addressed in a broad context and highlight the purpose of the study; (2) Methods: briefly describe the main methods or treatments applied; (3) Results: summarize the article’s main findings; (4) Conclusions: indicate the main conclusions or interpretations. The abstract should be an objective representation of the article and it must not contain results that are not presented and substantiated in the main text and should not exaggerate the main conclusions.
Keywords:
atrial fibrillation
; humanin
; MOTS-c
; mitochondria
1. Introduction
Atrial fibrillation (AF) , as the most prevalent clinical arrhythmia [1], often cause severe consequences such as stroke and heart failure (HF), thereby imposing a substantial societal burden [2]. The incidence of AF increases dramatically with age. Atrial electrical and structural remodeling are core pathophysiological mechanisms underlying the initiation and perpetuation of AF [3]. The current recommended therapies, including rate and rhythm control treatment and anticoagulation, fail to improve the outcomes and life quality of patients [4]. Even with extensive explorations, comprehensive mechanistic insightand targeted research to design precise interventions are still needed.
Atrial fibrosis is the central, dominant component of the structural remodeling process [3,5,6,7,8]. The hyperactivation, proliferation of fibroblasts, and excessive deposition of extracellular matrix, driven by factors like transforming growth factor-β and angiotensin II (AngII), are direct causes of fibrosis [3,8]. Previous studies confirmed the pivotal role of the renin-angiotensin-aldosterone system (RAAS) in this process. Inhibition of the angiotensin-converting enzyme suppress atrial fibrosis and the development of persistent AF in an animal model [9]. Meanwhile, clinical studies reported a lower incidence of AF in selected patient populations treated with RAAS inhibitors [10]. Further supporting this, a sub analysis of the EMPHASIS-HF trial showed that treatment with the mineralocorticoid receptor antagonist eplerenone was associated with a lower incidence of new-onset AF compared to placebo, underscoring its potential for AF prevention [11]. Furthermore, the mineralocorticoid receptor blocker (MRB) eplerenone (EPL) reduced the AF burden by attenuating structural remodeling in experimental sheep AF models [12]. Despite these advances, therapeutic responses are often incomplete, prompting the search for complementary or alternative targets within the fibrotic cascade.
Recently, mitochondrial dysfunction has been increasingly recognized as a key contributor to the pathogenesis of AF [13,14,15,16]. The rapid atrial rate during AF leads to energy depletion and excessive production of reactive oxygen species (ROS), triggering oxidative stress, inflammatory responses, and apoptosis, which ultimately exacerbate electrical and structural deterioration [14,15,16,17]. Consequently, targeting mitochondrial function has emerged as a promising therapeutic direction [16].
Humanin (HN) and MOTS-c are recently identified microp eptides encoded by mitochondrial DNA [18,19]. Notably, bioinformatic analyses have identified 13 sequences (MT-RNR2L1–MT-RNR2L13) in the nuclear genome that are highly homologous to the mitochondrial MT-RNR2 gene, at least 10 of which can be transcribed into mRNA and potentially translated into HN-like peptides [20]. Preclinical studies have demonstrated their broad cytoprotective capacities. HN protects endothelial cells from oxidized low-density lipoprotein (ox-LDL)-induced oxidative stress and apoptosis [21]. In aged mice, administration of HNG ameliorates myocardial fibrosis and oxidative stress [22]. In cerebral ischemia models, HNG reduces infarction area and pro-inflammatory cytokine expression [23]. Similarly, MOTS-c mitigates cardiac dysfunction, inflammation, and oxidative stress in transverse aortic constriction (TAC)-induced murine HF models [24] and enhances cardiac mechanical efficiency in exercised rats [25]. Recent study revealed the protective effects of MOTS-c in rat models of acute lung injury following cardiac ischemia [26]. However, whether and how humanin (HN) and MOTS-cattenuate AF remains elusive.
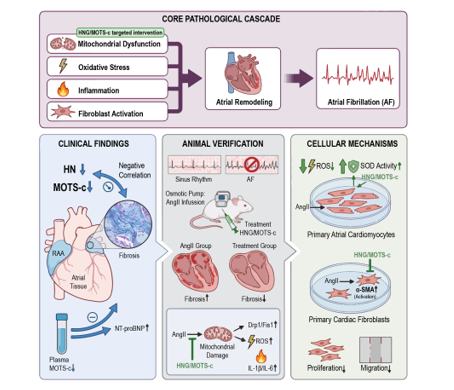
In the present study, we firstly evaluated the expressions of HN and MOTS-c in AF cardiac tissues as well as the plasma of patients, and determined the correlation of these peptides with the severity of AF. Then we explored their role and possible mechanism in which HN and MOTS-c exert their role in murine AF models and cell models. We found that 1) HN and MOTS-c expression was downregulated in human AF atrial tissue and negatively correlated with atrial fibrosis, while plasma MOTS-c levels were decreased and inversely associated with NT-proBNP; 2) Administration of HNG or MOTS-c reduced AF susceptibility and attenuated structural remodeling in an AngII-induced mouse model; 3) The peptides exerted their protective effects by preserving mitochondrial integrity, reducing oxidative stress and inflammation in atrial myocytes, and directly suppressing pro-fibrotic responses in atrial fibroblasts.
2. Material and Methods
2.1. Human Samples
Plasma samples were obtained from elderly residents (age >60 years) undergoing routine health examinations at the Beicai Community Health Center in Shanghai. Atrial fibrillation (AF) was defined by both a documented history of AF and its presence on a standard 12-lead electrocardiogram obtained during the examination. For each identified AF patient, one control subject with sinus rhythm (SR) was selected using 1:1 propensity score matching based on age, sex, hypertension, diabetes mellitus, and left ventricular ejection fraction. Participants meeting any of the following criteria were excluded: (1) History of myocardial infarction or catheter ablation for AF; (2) Chronic inflammatory diseases, as determined by medical history and white blood cell count; (3) Diagnosis of malignancy; (4) Severe hepatic or renal dysfunction; (5) Psychiatric disorders that could compromise the reliability of questionnaire-based information or informed consent. Initially, 40 matched pairs were enrolled. However, one plasma sample tube from an AF patient was found empty during processing, leading to the exclusion of that AF patient and its matched SR control. Consequently, 39 matched pairs (n=39 per group) were included in the final analysis. All plasma samples were processed on the day of collection and immediately stored at -80°C in the Biobank of Shanghai East Hospital until subsequent assay. Plasma levels of humanin and MOTS-c were measured using ELISA kits (EH9261, human humanin (MT-RNR2), and EH5056, human mitochondrial-derived peptide MOTS-c, FineTest, Wuhan Fine Biotech Co., Ltd., Wuhan, Hubei, China) as described previously [27]. The study was approved by the Ethics Committee of Shanghai East Hospital. Clinical trial registration number: NCT06673615. Informed consents were obtained from all enrolled participants. Table S1 provides details on the baseline characteristics of the study population.
Human right atrial tissue samples were harvested from patients (>18 years) who underwent elective open-heart surgery at Shanghai East Hospital. Clinical data for each patient were retrieved from electronic medical records. All patients provided written informed consent before undergoing cardiac surgery. Patients were categorized into the atrial fibrillation (AF) group (including paroxysmal and persistent AF) and the sinus rhythm (SR) group based on their clinical chart diagnoses. Atrial tissues from 13 individuals (6 with sinus rhythm and 7 with AF) were collected for histological and immunohistochemical evaluations. Exclusion criteria encompassed a diagnosis of post-operative AF, a history of autoimmune disorders, or systemic inflammatory conditions. All procedures involving human tissues and clinical data were reviewed and approved by the Ethics Committee of Shanghai East Hospital (approval number: 2025YS-159). Patient baseline characteristics are listed in Table S2.
2.2. Experimental Animals
All animal procedures complied with the guidelines of the Chinese National Institutes of Health and Animal Care, and received approval from the Animal Care Ethics Committee of Tongji University School of Medicine (approval No. TJBB09125103). Animals were maintained under controlled temperature, humidity, and a 12-hour light/dark cycle. All animals that finished the study protocol were included in the final data analysis, with nopost-allocation exclusions. The study established humane exclusion criteria, including severe illness, inability to consume food or water, or weight loss over 20%. None of the animals met these criteria during the course of the experiment.
2.3. Ang II Induced Atrial Fibrillation Murine Models with Peptide Treatment
Thirty-two male C57BL/6J mice (7 weeks old, 20–25 g) were purchased from GemPharmatech Co., Ltd. (Jiangsu, China; License No. SCXK (Su) 2023-0009). The mice were housed under specific-pathogen-free (SPF) conditions in the Animal Facility of Shanghai East Hospital, Tongji University School of Medicine. After one week of acclimatization, adult male C57BL/6J mice at 8 weeks of age were randomly assigned to four groups: Control group (n = 9), AngII group (n = 8), AngII + HNG group (n = 8), and AngII + MOTS-c group (n = 7). Mice were infused subcutaneously with saline or Ang II (T8560, TargetMol, USA) at a dose of 3 mg/kg/d via osmotic mini-pumps (2004w, RWD, China) for 3 weeks as described previously [28]. Concurrently, mice in the treatment groups received intraperitoneal injections of either HNG (HY-P3993A, MCE, USA) at 4 mg/kg/d or MOTS-c (TP2312, TargetMol, USA) at 3 mg/kg/d, five days per week, for the same 3-week duration. Control animals received same dose of sterile PBS vehicle injections on the same schedule.
2.4. Isolation and Culture of Primary Rat Atrial Cells
Primary rat atrial myocytes (NRAMs) and fibroblasts were isolated from the atrial tissue of 1-3-day-old Sprague-Dawley rats using a differential adhesion protocol, as previously described with modifications [29]. Briefly, the hearts were rapidly excised, and the atria were separated, minced into 1-2 mm³ fragments, and subjected to sequential digestion with 0.25% trypsin at 37°C. Dissociated cells were collected by centrifugation at 1500 g for 5 min and resuspended in DMEM (Biological Industries, Israel) supplemented with 10% fetal bovine serum (FBS, Biological Industries) and 1% penicillin/streptomycin (Beyotime, China). To separate cell types, the total cell suspension was plated in a culture dish and incubated for 45 minutes, allowing fibroblasts to adhere. The supernatant containing the non-adherent cardiac myocytes was then gently transferred and seeded into 6-well plates at a density of 3×10⁵ cells per well in DMEM containing 10% FBS for subsequent experiments. The adherent fibroblasts were cultured in the original dish for 48 hours, after which they were trypsinized, counted, and seeded into 6-well plates at a density of 3×10⁵ cells per well in DMEM containing 10% FBS for expansion prior to experiments.
2.5. Drug Treatment Protocol
For in vitro experiments, both cell types were pretreated with either 10 μmol/L HNG (HY-P3993A, MCE) or 5 μmol/L MOTS-c (TP2312, TargetMol) for 1 hour, followed by co-treatment with 1 μmol/L Ang II (T8560, TargetMol) for 48 hours. During the 48-hour treatment period, fibroblasts were maintained in DMEM containing 2% FBS, while cardiomyocytes were kept in DMEM with 10% FBS. All cells were cultured at 37°C in a humidified atmosphere of 5% CO2.
2.6. Histology and Immunohistochemistry
Atrial tissue samples from human and mouse sources were fixed in paraffin and sectioned at 5–7 μm thickness. Deparaffinization was performed using xylene, followed by rehydration through a graded alcohol series. Sections were then stained with hematoxylin solution, rinsed under running water, and differentiated with 1% acid alcohol. To evaluate atrial interstitial fibrosis, Masson’s trichrome and picrosirius red staining were employed. For immunofluorescence analysis, 8 μm-thick paraffin sections were subjected to antigen retrieval with citrate buffer and incubated with primary antibodies overnight at 4°C. Subsequently, sections were labeled with Alexa Fluor-conjugated secondary antibodies. Nuclear counterstaining was performed using DAPI, while the extracellular matrix was visualized with wheat germ agglutinin (WGA, Thermo Fisher Scientific, USA). Representative images were obtained using a Zeiss microscope, and fibrotic regions or antibody-positive areas were quantified with ImageJ software.
2.7. Measurement of Intracellular Reactive Oxygen Species (ROS)
Intracellular ROS levels were assessed using dihydroethidium (DHE) staining combined with both fluorescence microscopy and flow cytometry [29,30]. For both approaches, cells were incubated with 4µM DHE (Thermo Fisher Scientific) at 37°C for 20 minutes in the dark. For microscopic analysis, cells were washed with PBS after incubation. DHE fluorescence (indicative of superoxide oxidation) was visualized with Leica DM4B fluorescence microscope (excitation/emission: ~550/570nm) and quantified with ImageJ software (v1.8.0). For flow cytometric analysis, cells were harvested after DHE staining, washed twice with PBS by centrifugation at 800 rpm for 5 minutes at room temperature, and then resuspended in 200µL of PBS. The fluorescence intensity of oxidized DHE was immediately measured on a flow cytometer (Beckman cytoFLEX) using the PE channel (excitation: 488 nm; emission: 585 nm) and analyzed with FlowJo software (v10.8.1).
2.8. Western Blot Analysis
Protein lysates were prepared from cultured cells and atrial tissues using RIPA buffer (Beyotime). Equal amounts of protein (20–40 μg) were resolved via SDS-PAGE and electro transferred onto PVDF membranes (Millipore, USA). The membrane was blocked for 1 hour at room temperature, then incubated with primary antibody overnight at 4°C. After incubation with appropriate secondary antibodies, protein signals were detected using enhanced chemiluminescence reagents (Epizyme Biotech, China) and imaged with a ChemiDoc MP system (Bio-Rad Laboratories, USA). The primary antibodies used were as follows: CollagenI (66761-1-1g, protein tech, China), α-SMA (A17910, ABclonal, China), Drp1 (A21968, ABclonal), Fis1 (32525, cell signaling), GAPDH (A19056, ABclonal), β-actin (AC026, ABclonal) and horseradish peroxidase-labeled secondary antibodies (Beyotime, China).
2.9. Quantitative Real-Time Polymerase Chain Reaction
Total RNA was isolated from cultured cells and atrial tissue samples employing the SteadyPure Quick RNA Extraction Kit (Accurate Biotechnology, China). Complementary DNA was synthesized using the HiScript IIIRT SuperMix for qPCR (Vazyme, Nanjing, China). All subsequent amplifications were conducted using aQuantStudio™ 6 Flex Real-Time PCR Systems (ThermoFisher Scientific) and ChamQ Universal SYBR qPCR Master Mix (Vazyme). Relative mRNA levels were calculated by the 2−ΔΔCtmethod, with β-actin serving as the internal reference gene. Primer sequences are provided in Table S3.
2.10. Superoxide Dismutase (SOD)
Cellular samples from different experimental groups were analyzed to determine superoxide dismutase (SOD) activity levels. The measurements were performed with commercial SOD assay kits (Beyotime), following the standardized protocols provided by the manufacturer.
2.11. Transesophageal Programmed Electrical Stimulation
Transesophageal programmed electrical stimulation was performed to assess atrial fibrillation (AF) inducibility [31,32]. Mice were anesthetized via intraperitoneal administration of tribromoethanol (2.5%, 0.02 mL/g; Aibei Biotechnology Co., Ltd, Nanjing, China). Under stable anesthesia, a 1.1F octapolar electrophysiology catheter (Millar Instruments, Houston, TX) was advanced transesophageally and positioned at the level of the left atrium for subsequent programmed stimulation. Each mouse underwent the same atrial burst-pacing protocol three times. AF was defined as a period of rapid, irregular atrial activity lasting >1 second. Mice were classified as AF-positive when atrial fibrillation was elicited in a minimum of two out of three pacing trials.T he incidence of inducible AF was calculated as the percentage of AF-positive mice in each group. To ensure animal welfare, any AF episode was terminated by gentle tactile stimulation if it exceeded 10 minutes. Electrophysiological signals were recorded and analyzed using LabChart Pro software (v7.3, ADInstruments). Throughout the procedure and analysis, the operator was blinded to the treatment groups of the mice.
2.12. Cell Proliferation Assay (CCK-8)
Cell proliferation was assessed using the Cell Counting Kit-8 (CCK-8, Dojindo, Japan) according to the manufacturer's instructions. Primary rat atrial fibroblasts were seeded into 96-well plates and cultured overnight. After the indicated treatments for 48 hours, 10 µL of CCK-8 solution was added to each well and incubated for an additional 1 hours at 37°C. Absorbance was measured at 450 nm using a microplate reader.
2.13. Wound Healing Assay
Cell migration was evaluated using a wound healing assay. Primary rat atrial fibroblasts were seeded into 6-well plates and cultured until reaching approximately 90% confluence. A linear scratch wound was created across the cell monolayer using a sterile 200 µL pipette tip. Wound closure was photographed at 0, 24, and 48 hours using an inverted microscope. The wound area was quantified using ImageJ software, and the migration rate was calculated as the percentage of wound closure relative to the 0-hour time point.
2.14. Bioinformatic Analysis of Spatial Transcriptomic Data
The spatial transcriptomic dataset GSE261363 was downloaded from the Gene Expression Omnibus (GEO) database (http://www.ncbi.nlm.nih.gov/geo/, accessed on 14 October 2024). We processed and normalized the dataset using the GEOquery package (v2.66.0) in R [33]. To identify differentially expressed genes (DEGs), the limma package (version 4.4.1) was applied [34]. Genes with p-values < 0.05 and log-fold changes > 1 were considered significantly differentially expressed. Volcano plots and heatmaps were generated using R packages (‘ggplot2’v3.5.1and ‘pheatmap’v1.0.12) [35]. For each donor, six distinct tissue regions were analyzed, resulting in 12 columns per group in the heatmap.
2.15. RNA Sequencing of Primary Cardiac Fibroblasts
Primary rat atrial fibroblasts were treated with AngII alone, AngII combined with HNG, or AngII combined with MOTS-c for 48 h, with four biological replicates per group. Total RNA was extracted using TRIzol reagent (Invitrogen, USA) following the manufacturer’s instructions. RNA integrity was assessed, and qualified RNA samples were used for library preparation. Sequencing libraries were constructed using the Hieff NGS Ultima Dual-mode mRNA Library Prep Kit (Yeasen Biotechnology, China) according to the manufacturer’s protocol. The prepared libraries were sequenced on a DNBSEQ platform (BGI, China) to generate paired-end reads. The raw RNA-seq data have been deposited in the Gene Expression Omnibus (GEO) database under accession number GSE322635. Principal component analysis (PCA) and Gene Ontology (GO) enrichment analysis were performed using the online analysis platform (https://cloud.tsingke.com.cn, accessed on 13 July 2025).
2.16. Outcome Measures
The study evaluated outcomes encompassing electrophysiological, histopathological, molecular, and cellular parameters. Electrophysiological assessments focused on atrial fibrillation (AF) inducibility using transesophageal burst pacing, where AF was defined as sustained irregular atrial activity exceeding 1 second. Histopathological endpoints included atrial fibrosis quantified by Picrosirius red and Masson’s trichrome staining, cardiomyocyte hypertrophy determined by wheat germ agglutinin (WGA) staining, and mitochondrial morphology examined via transmission electron microscopy. Molecular measurements consisted of protein and mRNA expression levels of Humanin (HN) and MOTS-c in human atrial tissues and mouse atria, along with markers of mitochondrial fission (Drp1, Fis1), pro-inflammatory cytokines (IL-1β, IL-6), myocardial stress (Nppa, Acta1, Myh7, Sesn2), and fibroblast activation (α-SMA, Col1a1). Cellular endpoints included reactive oxygen species (ROS) generation, superoxide dismutase (SOD) activity in primary cardiomyocytes, and fibroblast proliferation (CCK-8 assay) and migration (wound healing assay).
As this study was designed as an exploratory mechanistic investigation, a single primary outcome measure was not predefined for sample size calculation. Rather, treatment effects were assessed based on the consistency of findings across multiple independent outcomes spanning electrophysiological, histopathological, molecular, and cellular domains.
2.17. Data Analysis and Statistical Methods
Statistical analyses were performed with GraphPad Prism software (version 8.0). Data for continuous variables are expressed as mean ± standard deviation (SD). The normality of data distribution for each dataset was evaluated using the Shapiro-Wilk test. For comparisons between two groups of normally distributed data, an unpaired two-tailed Student’s t test was applied. Comparisons among more than two groups of normally distributed data were conducted by one-way analysis of variance (ANOVA), followed by Tukey post hoc analysis for multiple comparisons. Non-normally distributed data were analyzed using the Mann-Whitney U test (two groups) or the Kruskal–Wallis test with Dunn’s post hoc correction (more than two groups). Associations between continuous variables were assessed using Pearson's correlation analysis for normally distributed data or Spearman's rank correlation for non-normally distributed data. A two-sided P value of less than 0.05 was considered statistically significant.
3. Results
3.1. Both Expressions of HN and MOTS-c Are Downregulated in Human Atrial Fibrillation Tissue and Negatively Correlate with Fibrosis
To investigate the potential role of mitochondrial peptides including HN and MOTS-cin AF, we first analyzed their gene expressions in human atrial tissues using public GEO database. Bioinformatic analysis of spatial transcriptomic data revealed significant downregulation of the nuclear gene encoding Humanin (HN) in AF atrial myocardial tissues compared to sinus rhythm (SR) controls (Figure 1A). This downregulation was further illustrated in a heatmap of the same dataset (Figure 1B). Consistent with the transcriptomic findings, IHC and IF staining further confirmed the substantial reductions of both HN (Figure 1C, D) and MOTS-c (Figure 1C,D) protein levels in atrial appendage of AF patients, compared to the SR control.
Next, Masson's trichrome and Picrosirius red staining showed a marked increase of collagen deposition in the atria of AF patients (Figure 1E). Importantly, quantitative correlation analysis demonstrated a significant negative relationship between the levels of both HN and MOTS-c and the degree of fibrosis quantified by both Masson's trichrome (Figure 2A, B) and Picrosirius red staining (Figure 1C, D). These results indicated that the downregulation of mitochondrial peptides HN and MOTS-c in the human AF atrium is closely associated with the progression or severity of of atrial fibrosis.
3.2. Plasma MOTS-c Level Decreased in AF Patients and Inversely Correlates with NT-proBNPlevel (N-Terminal pro-B-Type Natriuretic Peptide)
We also measured the levels of these peptides in plasma. As shown by figure 3A, MOTS-c concentration was significantly lower in AF patients than that in SR controls (Figure 3A). Interestingly, plasma HN levels were slightly elevated in AF patients (Figure 3B). A positive correlation was observed between plasma levels of HN and MOTS-c (Figure 3B). Furthermore, MOTS-c expressions exhibited a significant inverse correlation with NT-proBNP levels (Figure 3E), while no significant correlation was found between HN and NT-proBNP (Figure 3D). These results suggested that plasma MOTS-c, but not HN, may serve as a biomarker reflecting AF-related pathological stress.
3.3. Administration of HNG or MOTS-c Attenuates AngII-Induced Atrial Fibrillation Susceptibility and Structural Remodeling in Mice
To determine the therapeutic potential of these peptides, we employed an AngII-infused murine AF model (experimental timeline, Figure 4A). (Gly14)-Humanin (HNG), a synthetic and more potent analogue of HN with a glycine-for-serine substitution at position 14was utilized [36]. AngII treatment significantly increased AF inducibility upon transesophageal electrical pacing compared to the control group, as evidenced by representative electrocardiograms (Figure 4B) and summary data (Figure 4C). This pro-arrhythmic effect was markedly attenuated by concurrent treatment with either HNG (a potent HN analogue) or MOTS-c (Figure 4B, C).
Histological examination revealed that AngII infusion induced pronounced atrial fibrosis, which was ameliorated by both HNG and MOTS-c treatments (Figure 4D). Furthermore, Wheat Germ Agglutinin (WGA) staining showed that cardiomyocyte hypertrophy induced by AngII was also suppressed by either peptide administration (Figure 4E). At the molecular level, the upregulated mRNA expressions of myocardial stress markers, including Nppa, Nppb, Acta1, Myh7 [37], and Sesn2 [38] in AngII-treated mice were significantly blunted in the HNG and MOTS-c treatment groups (Figure 4F). These data demonstrated that inter peritoneal administration of HNG and MOTS-c can effectively protect the AngII treated mice from AF susceptibility as well as mitigate the cardiac structural remodeling.
3.4. HNG and MOTS-c Preserve Mitochondrial Integrity and Alleviate Oxidative Stress and Inflammation
We next explored the mechanisms underlying the protective effects of these two peptides. Electron microscopy revealed severe mitochondrial ultrastructural damage in the atria of AngII-treated mice, which was not ably prevented by HNG and MOTS-c treatment (Figure 5A). In addition, AngII infusion led to increased mRNA and protein expression of mitochondrial fission proteins Drp1 and Fis1, indicating aberrant mitochondrial dynamics. This effect was reversed by both peptide treatments (Figure 5B-D). Concurrently, the elevated mRNA levels of pro-inflammatory cytokines IL-1β and IL-6 in the AngII group were reduced following HNG and MOTS-c administration (Figure 5F). Immunofluorescence staining and quantification of F4/80 confirmed a reduction of macrophage infiltration into the heart of the treatment groups compared to the AF heart (Figure 5E).
In primary rat atrial cardiomyocytes, AngII challenge for 48 hours significantly increased oxidative stress, as detected by elevated ROS levels in flow cytometry and DHE staining, and decreased SOD activity. Pre-treatment with either HNG or MOTS-c effectively mitigated these changes (Figure 5G-I). Collectively, these findings indicated that the protective efficacy of HNG and MOTS-c in AF models may attribute to their function to preserve mitochondrial integrity, suppress inflammatory responses, and reduce oxidative stress in the heart.
3.5. HNG and MOTS-c Inhibit AngII-Induced Activation, Proliferation, and Migration of Cardiac Fibroblasts
Since atrial fibrosis is a hallmark of AF, we then investigated the direct effects of the peptides on cardiac fibroblasts. In primary rat cardiac fibroblasts, AngII stimulation significantly upregulated the mRNA and protein expression of activation markers α-SMA and Col1a1 (Figure 6A-C). This profibrotic activation response was suppressed by pre-treatment with HNG or MOTS-c. CCK-8 assays demonstrated that AngII-enhanced fibroblast proliferation was also inhibited by both peptides (Figure 6E). Finally, a wound-healing assay showed that the increased migratory capacity of fibroblasts induced by AngII was markedly reduced by addition of HNG and MOTS-c (Figure 6D). These results provided direct evidence that HNG and MOTS-c exert potent anti-fibrotic effects by attenuating the pro-fibrotic phenotype of cardiac fibroblasts.
To gain a comprehensive understanding of how HNG and MOTS-c modulate fibroblast phenotype at the transcriptional level, we performed RNA sequencing on primary cardiac fibroblasts treated with AngII, AngII+HNG, or AngII+MOTS-c. Principal component analysis (PCA) of the global gene expression data revealed a clear separation between AngII-treated fibroblasts and those co-treated with either HNG (Figure 6E) or MOTS-c (Figure 6F), indicating that both peptides profoundly altered the AngII-induced transcriptional landscape.
Gene Ontology (GO) enrichment analysis of the differentially expressed genes identified distinct biological processes influenced by each peptide. Compared to the AngII group, HNG treatment predominantly reversed genes associated with signal transduction, immune response, and cell-substrate adhesion (Figure 6G). In contrast, MOTS-c treatment primarily normalized the expression of genes involved in various metabolic processes (Figure 6H). These transcriptomic findings suggested that while both peptides effectively counteract AngII-driven fibroblast activation, they may exert their functions through overlapping yet distinct mechanisms, with HNG mainly affecting signaling and adhesion pathways and MOTS-c acting on metabolic reprogramming.
4. Discussion
Previous observations suggested a complex profile of HN and MOTS-c in peripheral blood and tissues in different clinical settings. It was reported that plasma HN levels were lower in patients with angina and myocardial infarction [39]. In a large cohort comprising 693 individuals across a wide age range (21–113 years), plasma HN levels were reported to increase with age, yet this elevation was associated with worsened metabolic and functional parameters in the elderly [40]. In chronic kidney disease, circulating HN is elevated while its muscle expression is reduced, and circulating HN correlates positively with inflammatory mediators; meanwhile, MOTS-c level decreased in both compartments [41]. Qin et al. reported that lower circulating endogenous MOTS-c levels were associated with impaired coronary endothelial function in patients [42]. Another study revealed the lower level of MOTS-c in coronary artery disease (CAD) patients and a strong correlation between MOTS-c levels and CAD [43]. These results suggested a common theme of MOTS-c deficiency in cardiovascular pathology. Other studies identified both peptides as exercise-sensitive myokines, with their levels increasing in both blood stream and muscle following physical activity [25,44,45]. Our data reveal that atrial tissues from AF patients exhibit markedly reduced levels of HN and MOTS-c, which negatively correlate with fibrotic burden. This local deficiency suggests that both peptides may participate in the pathogenesis of AngII-induced cardiac fibrosis and AF. We also found that plasma MOTS-c level decreases in AF patients’ plasma and negatively correlates with NT-proBNP production. These results indicated that plasma MOTS-c level might be a marker for cardiac injury. Despite these findings, their expression patterns, as well as the value in predicting the prognosis of diseases in atrial fibrillation need further exploration. The slight elevation of plasma HN in our AF cohort, however, presents an apparent paradox, mirroring the discordant pattern observed in chronic kidney disease where circulating HN rises despite local tissue reduction [41].
Large amount of evidence confirmed that mitochondrial dysfunction, oxidative stress, and inflammation contribute to AF progression. Previous studies confirmed the cardioprotective potential of mitochondrial-derived peptides including HN and MOTS-c in some pathologic conditions. For instance, HN was shown to mitigate endothelial apoptosis [21] and age-related fibrosis [22]. While MOTS-c can ameliorate pressure-overload heart failure [24]. The present study demonstrated that administration of both HN and MOTS-c can reduce AF inducibility, attenuate atrial fibrosis and hypertrophy induced by Ang II, suggesting their cardiac protective role in AF. Moreover, treatment with these peptides mitigate mitochondrial dysfunction, oxidative stress, and inflammation. Both peptides preserve mitochondrial ultrastructure and normalize the expression of fission proteins Drp1 and Fis1, which were reported to play important roles in stabilizing mitochondrial dynamics [46,47]. Notably, the upregulation of Drp1 aligns with recent single-cell RNA sequencing data from human AF atrial tissues, which revealed increased Drp1 expression across diverse cellular subpopulations [48]. By stabilizing mitochondrial dynamics, HNG and MOTS-c disrupt the consequent vicious cycle of oxidative stress and inflammation—key drivers of AF progression. The attenuation of this cycle is evidenced by reduced ROS, increased SOD activity, and downregulated IL-1β and IL-6, which is consistent with the anti-inflammatory effects of HNG reported in cerebral ischemia models [23]. Therefore, the improved mitochondrial function, downregulated oxidative stress, and reduced inflammation may attribute to the efficacy of administration of HN and MOTS-c in murine AF models.
Fibroblast activation plays a central role in the development of atrial fibrosis during atrial fibrillation [49]. The present data firstly revealed that both peptides directly inhibited AngII-induced transformation of fibroblasts into activated myofibroblasts, as well as their proliferation and migration. In murine models, administration of HNG and MOTS-c significantly attenuate fibrosis, suggesting their direct role on fibrosis. On the other hand, HNG and MOTS-c may also attenuate fibrosis in part by alleviating oxidative stress and inflammation, as well as improving the cardiomyocyte environment.
Several limitations of our study should be acknowledged. The human sample size was limited, and larger cohort studies are warranted to validate plasma MOTS-c as a biomarker for AF. We evaluated the direct role of HNG and MOTS-c on cardiomyocyte and fibroblast, whether and how both peptides play their roles on other cells in the heart need further explorations. Although we identified several downstream effectors, the specific receptor(s) and immediate signaling events triggered by HN and MOTS-c in the heart remain incompletely defined. Further research is needed to map the precise receptor-mediated signaling pathways.
5. Conclusion
Mitochondrial-derived peptides Humanin and MOTS-c are downregulated in atria of patients with AF and negatively associated with atrial fibrosis. Administration of HNG or MOTS-c protect the heart from AF susceptibility through preservation of mitochondrial integrity, attenuation of oxidative stress and inflammation, and direct suppression of pro-fibrotic signaling in cardiac fibroblasts. Therefore,HN and MOTS-c may act as promising novel therapeutic agents for the treatment of atrial fibrillation.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1 Baseline characteristics of the study population for plasma samples; Table S2 Baseline characteristics of the patients for atrial biopsies: Table S3 Primer sequences used in this study.
Author Contributions
Conceptualization, Y.L. and X.Z.; methodology, Y.L.; validation, Y.L., J.X. and Y.J.; formal analysis, J.X. and D.C.; investigation, Y.S.; resources, X.S., M.G. and Y.D.; data curation, Y.L.; writing—original draft preparation, Y.L.; writing—review and editing, X.Z.; visualization, Y.L.; supervision, W.H.; project administration, W.H.; funding acquisition, W.H. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (Grant No. 82370331, entitled "The role and mechanism of CACNA2D2/ISYNA1 endoplasmic reticulum axis in atrial fibrosis of atrial fibrillation", 2024–2027), the Shanghai Health Commission (Grant No. 202440054), and the Key Discipline Construction Project of Shanghai Pudong New Area Health Commission (Grant No. PWZxk2022-20).
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Ethics Committee, Shanghai East Hospital (protocol code 2025YS-159, date of approval: 16 June 2025) for studies involving human samples. All animal procedures conformed to the guidelines of the Chinese National Institutes of Health and Animal Care, with ethical approval granted by the Tongji University School of Medicine Animal Care Ethics Committee (approval number TJBB09125103; date: 11 September 2025). The community-based clinical study was registered at ClinicalTrials.gov (registration number: NCT06673615, date of registration: 5 November 2024).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The RNA sequencing data of primary cardiac fibroblasts generated in this study have been deposited in the Gene Expression Omnibus (GEO) database under accession number GSE322635. The data are publicly available at the time of publication. The dataset GSE261363 is obtained from the GEO database (http://www.ncbi.nlm.nih.gov/geo/, accessed on 14 October 2024). Other data supporting the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
During the preparation of this manuscript, the authors used AI-assisted language editing tools for the purposes of language polishing and formatting assistance. The authors have reviewed and edited the output and take full responsibility for the content of this publication.
Conflicts of Interest
The authors confirm that there are no conflicts of interest to disclose.
Abbreviations
The following abbreviations are used in this manuscript:
| AF | Atrial fibrillation |
| α-SMA | Alpha-smooth muscle actin |
| AngII | Angiotensin II |
| ANOVA | Analysis of variance |
| CAD | Coronary artery disease |
| CCK-8 | Cell Counting Kit-8 |
| Col1a1 | Collagen type I alpha 1 chain |
| DAPI | 4′,6-diamidino-2-phenylindole |
| DEGs | Differentially expressed genes |
| DHE | Dihydroethidium |
| DMEM | Dulbecco's Modified Eagle Medium |
| DNA | Deoxyribonucleic acid |
| Drp1 | Dynamin-related protein 1 |
| ECG | Electrocardiogram |
| ELISA | Enzyme-linked immunosorbent assay |
| EPL | Eplerenone |
| FBS | Fetal bovine serum |
| Fis1 | Mitochondrial fission protein 1 |
| GEO | Gene Expression Omnibus |
| GO | Gene Ontology |
| HNG | Gly14-Humanin (Humanin analogue) |
| HN | Humanin |
| IF | Immunofluorescence |
| IHC | Immunohistochemistry |
| IL-1β | Interleukin-1 beta |
| IL-6 | Interleukin-6 |
| MDPs | Mitochondrial-derived peptides |
| MI | Myocardial infarction |
| MOTS-c | Mitochondrial open reading frame of the 12S rRNA-c |
| MRB | Mineralocorticoid receptor blocker |
| mRNA | Messenger RNA |
| MTRNR2L | MT-RNR2-like (nuclear humanin genes) |
| NRAMs | Neonatal rat atrial myocytes |
| NT-proBNP | N-terminal pro-brain natriuretic peptide |
| ox-LDL | Oxidized low-density lipoprotein |
| PAGE | Polyacrylamide gel electrophoresis |
| PBS | Phosphate-buffered saline |
| PCA | Principal component analysis |
| PCR | Polymerase chain reaction |
| PVDF | Polyvinylidene difluoride |
| qPCR | Quantitative polymerase chain reaction |
| RAAS | Renin-angiotensin-aldosterone system |
| RIPA | Radioimmunoprecipitation assay |
| RNA | Ribonucleic acid |
| ROS | Reactive oxygen species |
| SD | Standard deviation |
| SDS | Sodium dodecyl sulfate |
| SOD | Superoxide dismutase |
| SR | Sinus rhythm |
| TAC | Transverse aortic constriction |
| WGA | Wheat germ agglutinin |
References
- Marek-Iannucci, S.; Ozdemir, A.B.; Moreira, D.; Gomez, A.C.; Lane, M.; Porritt, R.A.; Lee, Y.; Shimada, K.; Abe, M.; Stotland, A.; et al. Autophagy-mitophagy induction attenuates cardiovascular inflammation in a murine model of Kawasaki disease vasculitis. J. Clin. Investig. 2021, 6. [CrossRef]
- Andrade, J., et al., The clinical profile and pathophysiology of atrial fibrillation: relationships among clinical features, epidemiology, and mechanisms.Circ Res, 2014. 114(9): p. 1453-68.
- Dzeshka, M.S., et al., Cardiac Fibrosis in Patients With Atrial Fibrillation: Mechanisms and Clinical Implications.J Am Coll Cardiol, 2015. 66(8): p. 943-59.
- Hindricks, G.; Potpara, T.; Dagres, N.; Arbelo, E.; Bax, J.J.; Blomstrӧm-Lundqvist, C.; Boriani, G.; Castella, M.; Dan, G.-A.; Dilaveris, P.E.; et al. 2020 ESC Guidelines for the diagnosis and management of atrial fibrillation developed in collaboration with the European Association for Cardio-Thoracic Surgery (EACTS). Russ. J. Cardiol. 2021, 26, 4701. [CrossRef]
- de Groot, N.M., et al., Electropathological substrate of longstanding persistent atrial fibrillation in patients with structural heart disease: epicardial breakthrough.Circulation, 2010. 122(17): p. 1674-82.
- Krishnan, A.; Chilton, E.; Raman, J.; Saxena, P.; McFarlane, C.; Trollope, A.F.; Kinobe, R.; Chilton, L. Are Interactions between Epicardial Adipose Tissue, Cardiac Fibroblasts and Cardiac Myocytes Instrumental in Atrial Fibrosis and Atrial Fibrillation?. Cells 2021, 10, 2501. [CrossRef]
- Zhu, P.; Li, H.; Zhang, A.; Li, Z.; Zhang, Y.; Ren, M.; Zhang, Y.; Hou, Y. MicroRNAs sequencing of plasma exosomes derived from patients with atrial fibrillation: miR-124-3p promotes cardiac fibroblast activation and proliferation by regulating AXIN1. J. Physiol. Biochem. 2021, 78, 85–98. [CrossRef]
- Karakasis, P.; Theofilis, P.; Vlachakis, P.K.; Korantzopoulos, P.; Patoulias, D.; Antoniadis, A.P.; Fragakis, N. Atrial Fibrosis in Atrial Fibrillation: Mechanistic Insights, Diagnostic Challenges, and Emerging Therapeutic Targets. Int. J. Mol. Sci. 2024, 26, 209. [CrossRef]
- Anné, W.; Willems, R.; Holemans, P.; Beckers, F.; Roskams, T.; Lenaerts, I.; Ector, H.; Heidbüchel, H. Self-terminating AF depends on electrical remodeling while persistent AF depends on additional structural changes in a rapid atrially paced sheep model. J. Mol. Cell. Cardiol. 2007, 43, 148–158. [CrossRef]
- Ehrlich, J.R.; Hohnloser, S.H.; Nattel, S. Role of angiotensin system and effects of its inhibition in atrial fibrillation: clinical and experimental evidence. Eur. Hear. J. 2005, 27, 512–518. [CrossRef]
- Swedberg, K., et al., Eplerenone and atrial fibrillation in mild systolic heart failure: results from the EMPHASIS-HF (Eplerenone in Mild Patients Hospitalization And SurvIval Study in Heart Failure) study.J Am Coll Cardiol, 2012. 59(18): p. 1598-603.
- Takemoto, Y.; Ramirez, R.J.; Kaur, K.; Salvador-Montañés, O.; Ponce-Balbuena, D.; Ramos-Mondragón, R.; Ennis, S.R.; Guerrero-Serna, G.; Berenfeld, O.; Jalife, J. Eplerenone Reduces Atrial Fibrillation Burden Without Preventing Atrial Electrical Remodeling. JACC 2017, 70, 2893–2905. [CrossRef]
- Murphy, E., et al., Mitochondrial Function, Biology, and Role in Disease: A Scientific Statement From the American Heart Association.Circ Res, 2016. 118(12): p. 1960-91.
- Bukowska, A.; Schild, L.; Keilhoff, G.; Hirte, D.; Neumann, M.; Gardemann, A.; Neumann, K.H.; Röhl, F.-W.; Huth, C.; Goette, A.; et al. Mitochondrial Dysfunction and Redox Signaling in Atrial Tachyarrhythmia. Exp. Biol. Med. 2008, 233, 558–574. [CrossRef]
- Lin, P.-H.; Lee, S.-H.; Su, C.-P.; Wei, Y.-H. Oxidative damage to mitochondrial DNA in atrial muscle of patients with atrial fibrillation. Free. Radic. Biol. Med. 2003, 35, 1310–1318. [CrossRef]
- Júnior, A.d.S.M.; de França-E-Silva, A.L.G.; de Oliveira, J.M.; da Silva, D.M. Developing Pharmacological Therapies for Atrial Fibrillation Targeting Mitochondrial Dysfunction and Oxidative Stress: A Scoping Review. Int. J. Mol. Sci. 2023, 25, 535. [CrossRef]
- Xie, W., et al., Mitochondrial oxidative stress promotes atrial fibrillation.Sci Rep, 2015. 5: p. 11427.
- Hashimoto, Y.; Niikura, T.; Tajima, H.; Yasukawa, T.; Sudo, H.; Ito, Y.; Kita, Y.; Kawasumi, M.; Kouyama, K.; Doyu, M.; et al. A rescue factor abolishing neuronal cell death by a wide spectrum of familial Alzheimer's disease genes and Aβ. Proc. Natl. Acad. Sci. USA 2001, 98, 6336–6341. [CrossRef]
- Lee, C.; Zeng, J.; Drew, B.G.; Sallam, T.; Martin-Montalvo, A.; Wan, J.; Kim, S.-J.; Mehta, H.; Hevener, A.L.; de Cabo, R.; et al. The Mitochondrial-Derived Peptide MOTS-c Promotes Metabolic Homeostasis and Reduces Obesity and Insulin Resistance. Cell Metab. 2015, 21, 443–454. [CrossRef]
- Bodzioch, M.; Lapicka-Bodzioch, K.; Zapala, B.; Kamysz, W.; Kiec-Wilk, B.; Dembinska-Kiec, A. Evidence for potential functionality of nuclearly-encoded humanin isoforms. Genomics 2009, 94, 247–256. [CrossRef]
- Bachar, A.R.; Scheffer, L.; Schroeder, A.S.; Nakamura, H.K.; Cobb, L.J.; Oh, Y.K.; Lerman, L.O.; Pagano, R.E.; Cohen, P.; Lerman, A. Humanin is expressed in human vascular walls and has a cytoprotective effect against oxidized LDL-induced oxidative stress. Cardiovasc. Res. 2010, 88, 360–366. [CrossRef]
- Qin, Q.; Mehta, H.; Yen, K.; Navarrete, G.; Brandhorst, S.; Wan, J.; Delrio, S.; Zhang, X.; Lerman, L.O.; Cohen, P.; et al. Chronic treatment with the mitochondrial peptide humanin prevents age-related myocardial fibrosis in mice. Am. J. Physiol. Circ. Physiol. 2018, 315, H1127–H1136. [CrossRef]
- Peng, T.; Wan, W.; Wang, J.; Liu, Y.; Fu, Z.; Ma, X.; Li, J.; Sun, G.; Ji, Y.; Lu, J.; et al. The Neurovascular Protective Effect of S14G-Humanin in a Murine MCAO Model and Brain Endothelial Cells. IUBMB Life 2018, 70, 691–699. [CrossRef]
- Zhong, P.; Peng, J.; Hu, Y.; Zhang, J.; Shen, C. Mitochondrial derived peptide MOTS-c prevents the development of heart failure under pressure overload conditions in mice. J. Cell. Mol. Med. 2022, 26, 5369–5378. [CrossRef]
- Yuan, J.; Wang, M.; Pan, Y.; Liang, M.; Fu, Y.; Duan, Y.; Tang, M.; Laher, I.; Li, S. The mitochondrial signaling peptide MOTS-c improves myocardial performance during exercise training in rats. Sci. Rep. 2021, 11, 1–11. [CrossRef]
- Lu, P.; Li, X.; Li, B.; Li, X.; Wang, C.; Liu, Z.; Ji, Y.; Wang, X.; Wen, Z.; Fan, J.; et al. The mitochondrial-derived peptide MOTS-c suppresses ferroptosis and alleviates acute lung injury induced by myocardial ischemia reperfusion via PPARγ signaling pathway. Eur. J. Pharmacol. 2023, 953, 175835. [CrossRef]
- Kavak, A.G.; Karslioglu, I.; Saracaloglu, A.; Demiryürek, S.; Demiryürek, A.T. Impact of Radiation Therapy on Serum Humanin and MOTS-c Levels in Patients with Lung or Breast Cancer. Curr. Radiopharm. 2024, 17, 229–237. [CrossRef]
- Li, J.; Wang, S.; Zhang, Y.-L.; Bai, J.; Lin, Q.-Y.; Liu, R.-S.; Yu, X.-H.; Li, H.-H. Immunoproteasome Subunit β5i Promotes Ang II (Angiotensin II)–Induced Atrial Fibrillation by Targeting ATRAP (Ang II Type I Receptor–Associated Protein) Degradation in Mice. Hypertension 2019, 73, 92–101. [CrossRef]
- Li, D.-S.; Xue, G.-L.; Yang, J.-M.; Li, C.-Z.; Zhang, R.-X.; Tian, T.; Li, Z.; Shen, K.-W.; Guo, Y.; Liu, X.-N.; et al. Knockout of interleukin-17A diminishes ventricular arrhythmia susceptibility in diabetic mice via inhibiting NF-κB-mediated electrical remodeling. Acta Pharmacol. Sin. 2021, 43, 307–315. [CrossRef]
- Han, W.; Du, C.; Zhu, Y.; Ran, L.; Wang, Y.; Xiong, J.; Wu, Y.; Lan, Q.; Wang, Y.; Wang, L.; et al. Targeting Myocardial Mitochondria-STING-Polyamine Axis Prevents Cardiac Hypertrophy in Chronic Kidney Disease. JACC: Basic Transl. Sci. 2022, 7, 820–840. [CrossRef]
- Schrickel, J.W.; Bielik, H.; Yang, A.; Schimpf, R.; Shlevkov, N.; Burkhardt, D.; Meyer, R.; Grohé, C.; Fink, K.; Tiemann, K.; et al. Induction of atrial fibrillation in mice by rapid transesophageal atrial pacing. Basic Res. Cardiol. 2002, 97, 452–460. [CrossRef]
- Murphy, M.B.; Kim, K.; Kannankeril, P.J.; Subati, T.; Van Amburg, J.C.; Barnett, J.V.; Murray, K.T. Optimizing transesophageal atrial pacing in mice to detect atrial fibrillation. Am. J. Physiol. Circ. Physiol. 2022, 322, H36–H43. [CrossRef]
- Davis, S.; Meltzer, P.S. GEOquery: a bridge between the Gene Expression Omnibus (GEO) and BioConductor. Bioinformatics 2007, 23, 1846–1847. [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [CrossRef]
- Hao, D.; Yang, X.; Li, Z.; Xie, B.; Feng, Y.; Liu, G.; Ren, X. Screening core genes for minimal change disease based on bioinformatics and machine learning approaches. Int. Urol. Nephrol. 2024, 57, 655–671. [CrossRef]
- Chin, Y.-P.; Keni, J.; Wan, J.; Mehta, H.; Anene, F.; Jia, Y.; Lue, Y.-H.; Swerdloff, R.; Cobb, L.J.; Wang, C.; et al. Pharmacokinetics and Tissue Distribution of Humanin and Its Analogues in Male Rodents. Endocrinology 2013, 154, 3739–3744. [CrossRef]
- Wang, W.; Li, E.; Zou, J.; Qu, C.; Ayala, J.; Wen, Y.; Islam, S.; Weintraub, N.L.; Fulton, D.J.; Liang, Q.; et al. Ubiquitin Ligase RBX2/SAG Regulates Mitochondrial Ubiquitination and Mitophagy. Circ. Res. 2024, 135, e39–e56. [CrossRef]
- Zahid, M.A.; Abdelsalam, S.S.; Raïq, H.; Parray, A.; Korashy, H.M.; Zeidan, A.; Elrayess, M.A.; Agouni, A. Sestrin2 as a Protective Shield against Cardiovascular Disease. Int. J. Mol. Sci. 2023, 24, 4880. [CrossRef]
- Cai, H.; Cao, P.; Sun, W.; Shao, W.; Li, R.; Wang, L.; Zou, L.; Forno, E.; Muzumdar, R.; Gong, Z.; et al. Circulating humanin is lower in coronary artery disease and is a prognostic biomarker for major cardiac events in humans. Biochim. et Biophys. Acta (BBA) - Gen. Subj. 2022, 1866, 130010. [CrossRef]
- Conte, M.; Ostan, R.; Fabbri, C.; Santoro, A.; Guidarelli, G.; Vitale, G.; Mari, D.; Sevini, F.; Capri, M.; Sandri, M.; et al. Human Aging and Longevity Are Characterized by High Levels of Mitokines. Journals Gerontol. Ser. A 2018, 74, 600–607. [CrossRef]
- Liu, C.; Gidlund, E.-K.; Witasp, A.; Qureshi, A.R.; Söderberg, M.; Thorell, A.; Nader, G.A.; Barany, P.; Stenvinkel, P.; von Walden, F. Reduced skeletal muscle expression of mitochondrial-derived peptides humanin and MOTS-C and Nrf2 in chronic kidney disease. Am. J. Physiol. Physiol. 2019, 317, F1122–F1131. [CrossRef]
- Qin, Q.; Delrio, S.; Wan, J.; Widmer, R.J.; Cohen, P.; Lerman, L.O.; Lerman, A. Downregulation of circulating MOTS-c levels in patients with coronary endothelial dysfunction. Int. J. Cardiol. 2018, 254, 23–27. [CrossRef]
- Yasar, E.; Cakmak, T.; Bayramoglu, A.; Karaku, Y.; Tekin, S.; Sekerci, G.; Turkoglu, C. MOTS-c as a predictor of coronary lesions and complexity in patients with stable coronary artery disease. 2022, 26, 5676–5682. [CrossRef]
- Woodhead, J.S.T.; D’sOuza, R.F.; Hedges, C.P.; Wan, J.; Berridge, M.V.; Cameron-Smith, D.; Cohen, P.; Hickey, A.J.R.; Mitchell, C.J.; Merry, T.L. High-intensity interval exercise increases humanin, a mitochondrial encoded peptide, in the plasma and muscle of men. J. Appl. Physiol. 2020, 128, 1346–1354. [CrossRef]
- Reynolds, J.C.; Lai, R.W.; Woodhead, J.S.T.; Joly, J.H.; Mitchell, C.J.; Cameron-Smith, D.; Lu, R.; Cohen, P.; Graham, N.A.; Benayoun, B.A.; et al. MOTS-c is an exercise-induced mitochondrial-encoded regulator of age-dependent physical decline and muscle homeostasis. Nat. Commun. 2021, 12, 1–11. [CrossRef]
- Tilokani, L.; Nagashima, S.; Paupe, V.; Prudent, J. Mitochondrial dynamics: overview of molecular mechanisms. Essays Biochem. 2018, 62, 341–360. [CrossRef]
- Quiles, J.M. and B. Gustafsson Å, The role of mitochondrial fission in cardiovascular health and disease. Nat Rev Cardiol, 2022. 19(11): p. 723-736.
- Li, Y., et al., Ibrutinib Promotes Atrial Fibrillation by Disrupting A-Kinase Anchoring Protein 1-Mediated Mitochondrial Quality Surveillance in Cardiomyocytes. Research (Wash D C), 2024. 7: p. 0509.
- Harada, M.; Nattel, S. Implications of Inflammation and Fibrosis in Atrial Fibrillation Pathophysiology. Card. Electrophysiol. Clin. 2021, 13, 25–35. [CrossRef]
Figure 1.
Both HN and MOTS-c levels decreased in atrial tissues of atrial fibrillation (AF). (A) Reanalysis of spatial transcriptomic data (GEO dataset GSE261363) showing downregulated expression of the nuclear-encoded humanin genes (MTRNR2L1, MTRNR2L5) in atrial myocardialtissue from donors with atrial fibrillation (AF, n=2 donors) compared to those with sinus rhythm (SR, n=2 donors). (B) Heatmap generated from the same dataset (GSE261363), displaying the expression patterns of the top 20 differentially expressed genes. Six distinct tissue regions were analyzed per donor, resulting in 12 columns per group (SR, AF). (C) Representative immunohistochemical (IHC) staining and quantification of HN and MOTS-c protein expression in human right atrial appendage tissues from patients with SR and AF. Scale bars, 100 μm. (D) Representative immunofluorescence (IF) staining and quantification of HN and MOTS-c in human atrial tissues from SR and AF patients. Scale bars, 100 μm. (E) Representative Masson’s trichrome staining and Picrosirius red staining of human atrial tissues, showing increased collagen deposition in AF. Scale bars,100 μm. Data in C, D were analyzed via the Student t tests. Data of Masson’s trichrome staining in E were analyzed via the Mann-Whitney U tests. Data of Picrosirius red staining in E wasanalyzed via the Student t tests.
Figure 1.
Both HN and MOTS-c levels decreased in atrial tissues of atrial fibrillation (AF). (A) Reanalysis of spatial transcriptomic data (GEO dataset GSE261363) showing downregulated expression of the nuclear-encoded humanin genes (MTRNR2L1, MTRNR2L5) in atrial myocardialtissue from donors with atrial fibrillation (AF, n=2 donors) compared to those with sinus rhythm (SR, n=2 donors). (B) Heatmap generated from the same dataset (GSE261363), displaying the expression patterns of the top 20 differentially expressed genes. Six distinct tissue regions were analyzed per donor, resulting in 12 columns per group (SR, AF). (C) Representative immunohistochemical (IHC) staining and quantification of HN and MOTS-c protein expression in human right atrial appendage tissues from patients with SR and AF. Scale bars, 100 μm. (D) Representative immunofluorescence (IF) staining and quantification of HN and MOTS-c in human atrial tissues from SR and AF patients. Scale bars, 100 μm. (E) Representative Masson’s trichrome staining and Picrosirius red staining of human atrial tissues, showing increased collagen deposition in AF. Scale bars,100 μm. Data in C, D were analyzed via the Student t tests. Data of Masson’s trichrome staining in E were analyzed via the Mann-Whitney U tests. Data of Picrosirius red staining in E wasanalyzed via the Student t tests.

Figure 2.
Correlations of HN and MOTS-C expression with atrial fibrosis degree in human atrial tissues. Correlation analysis between quantitative fibrosis area [from Masson’s staining (A,B) and Picrosirius red staining (C,D)] and the protein expression levels of HN (A,C) or MOTS-c (B,D) (quantified from IHC). Spearman rank correlation coefficient quantifies these correlations.
Figure 2.
Correlations of HN and MOTS-C expression with atrial fibrosis degree in human atrial tissues. Correlation analysis between quantitative fibrosis area [from Masson’s staining (A,B) and Picrosirius red staining (C,D)] and the protein expression levels of HN (A,C) or MOTS-c (B,D) (quantified from IHC). Spearman rank correlation coefficient quantifies these correlations.

Figure 3.
Plasma MOTS-c level is downregulated in patients with AF and inversely associated with NT-proBNP production. (A,B) ELISA was performed to measure plasma HN (A) and MOTS-c (B) concentrations in patients with atrial fibrillation (AF; n=39) and control subjects with sinus rhythm (SR; n=39). (C) Spearman correlation analysis between plasma HN and MOTS-c levels in patients above (n=78). (C, D) Spearman correlation analysis of plasma HN (C) or MOTS-c (D) levels with NT-proBNP levels in patients above (n=78). Data in A and B were analyzed via the Mann-Whitney U tests.
Figure 3.
Plasma MOTS-c level is downregulated in patients with AF and inversely associated with NT-proBNP production. (A,B) ELISA was performed to measure plasma HN (A) and MOTS-c (B) concentrations in patients with atrial fibrillation (AF; n=39) and control subjects with sinus rhythm (SR; n=39). (C) Spearman correlation analysis between plasma HN and MOTS-c levels in patients above (n=78). (C, D) Spearman correlation analysis of plasma HN (C) or MOTS-c (D) levels with NT-proBNP levels in patients above (n=78). Data in A and B were analyzed via the Mann-Whitney U tests.

Figure 4.
HNG and MOTS-c treatment attenuates AngII-induced atrial fibrillation (AF) susceptibility and structural remodeling in mice. (A) Schematic timeline of the experimental protocol. (B) Representative electrocardiogram (ECG) tracings of burst pacing–induced AF. (C) Bar graph summarizing the incidence of inducible AF in each group. (D) Representative images of Picrosirius red-stained whole heart sections from each experimental group, alongside quantification of fibrotic area (red) in both atrial chambers. Scale bars, 2.5 mm or 100 μm. (E) Representative images of wheat germ agglutinin (WGA) staining of the mouse atria from the indicated groups, showing cardiomyocyte cross-sectional area. A total of 250 cells per mouse were measured. Scale bar, 50 μm. (F) mRNA expression levels of myocardial stress markers (Nppa, Nppb, Acta1, Myh7, Sesn2) in mouse atrial tissues. n =3 to 9 mice per group for all experiments. Data points in the graphs represent biological replicates. Data in C were analyzed via χ2 test (AF inducibility). Data in D to F were analyzed via1-way ANOVA, followed by Tukey post hoc analysis.
Figure 4.
HNG and MOTS-c treatment attenuates AngII-induced atrial fibrillation (AF) susceptibility and structural remodeling in mice. (A) Schematic timeline of the experimental protocol. (B) Representative electrocardiogram (ECG) tracings of burst pacing–induced AF. (C) Bar graph summarizing the incidence of inducible AF in each group. (D) Representative images of Picrosirius red-stained whole heart sections from each experimental group, alongside quantification of fibrotic area (red) in both atrial chambers. Scale bars, 2.5 mm or 100 μm. (E) Representative images of wheat germ agglutinin (WGA) staining of the mouse atria from the indicated groups, showing cardiomyocyte cross-sectional area. A total of 250 cells per mouse were measured. Scale bar, 50 μm. (F) mRNA expression levels of myocardial stress markers (Nppa, Nppb, Acta1, Myh7, Sesn2) in mouse atrial tissues. n =3 to 9 mice per group for all experiments. Data points in the graphs represent biological replicates. Data in C were analyzed via χ2 test (AF inducibility). Data in D to F were analyzed via1-way ANOVA, followed by Tukey post hoc analysis.

Figure 5.
HNG and MOTS-c preserve mitochondrial integrity and alleviate inflammation and oxidative stress in mouse atria and primary cardiomyocytes. (A) Representative transmission electron micrographs of mitochondria in mouse atrial cardiomyocytes from the indicated groups. Scalebar, 1 μm or 500 nm. (B-D) Representative Western blot (B) with quantitative analysis (C) and mRNA (D) of mitochondrial fission proteins (Drp1, Fis1) in mouse atrial tissues. (E) Representative immunofluorescence staining and quantification of F4/80 (macrophage marker) in mouse atrial tissues. Scale bar, 50 μm. (F) mRNA expression levels of pro-inflammatory cytokines (IL-1β, IL-6) in mouse atrial tissues. (G-I) In vitro studies on primary rat atrial cardiomyocytes. (G) Flow cytometry analysis of reactive oxygen species (ROS) using DHE probe. (H) Representative DHE fluorescence staining (red) and DAPI (blue). Scale bar, 50 μm. (I) Superoxide dismutase (SOD) activity assay. n = 3-6 per group for in vivo, n = 3 independent experiments for in vitro. Data in B to I were analyzed via1-way ANOVA, followed by Tukey post hoc analysis.
Figure 5.
HNG and MOTS-c preserve mitochondrial integrity and alleviate inflammation and oxidative stress in mouse atria and primary cardiomyocytes. (A) Representative transmission electron micrographs of mitochondria in mouse atrial cardiomyocytes from the indicated groups. Scalebar, 1 μm or 500 nm. (B-D) Representative Western blot (B) with quantitative analysis (C) and mRNA (D) of mitochondrial fission proteins (Drp1, Fis1) in mouse atrial tissues. (E) Representative immunofluorescence staining and quantification of F4/80 (macrophage marker) in mouse atrial tissues. Scale bar, 50 μm. (F) mRNA expression levels of pro-inflammatory cytokines (IL-1β, IL-6) in mouse atrial tissues. (G-I) In vitro studies on primary rat atrial cardiomyocytes. (G) Flow cytometry analysis of reactive oxygen species (ROS) using DHE probe. (H) Representative DHE fluorescence staining (red) and DAPI (blue). Scale bar, 50 μm. (I) Superoxide dismutase (SOD) activity assay. n = 3-6 per group for in vivo, n = 3 independent experiments for in vitro. Data in B to I were analyzed via1-way ANOVA, followed by Tukey post hoc analysis.

Figure 6.
HNG and MOTS-c inhibit AngII-induced activation, proliferation, and migration of primary cardiac fibroblasts. All experiments were performed on primary rat atrial fibroblasts. (A-C) Representative Western blot (A) with quantitative analysis (B) and mRNA expression (C) of fibroblast activation markers (α-SMA, Col1a1). (D) Cell migration assessed by wound healing assay. Representative images at 0 h and 48 h are shown (scale bar, 200 μm), with quantitative analysis of migration rate. (E) Cell proliferation assessed by CCK-8 assay. n = 3-6 independent experiments for in vitro. Data in Bto Ewere analyzed via1-way ANOVA, followed by Tukey post hoc analysis. (F, G) RNA sequencing analysis of primary cardiac fibroblasts. Principal component analysis (PCA) plots of global gene expression profiles for cells treated with AngII versus AngII+HNG (F) and AngII versus AngII+MOTS-c (G) (n = 4 independent samples per group). (H, I) Gene Ontology (GO) enrichment analysis of biological processes. Shown are the top enriched terms for differentially expressed genes identified between AngII-treated fibroblasts and the AngII+HNG group (H) [filtered by p < 0.05 and p.adjust (q value) < 0.05], and between AngII-treated fibroblasts and the AngII+MOTS-c group (I) [filtered by p < 0.05 and p.adjust (q value) < 0.2].
Figure 6.
HNG and MOTS-c inhibit AngII-induced activation, proliferation, and migration of primary cardiac fibroblasts. All experiments were performed on primary rat atrial fibroblasts. (A-C) Representative Western blot (A) with quantitative analysis (B) and mRNA expression (C) of fibroblast activation markers (α-SMA, Col1a1). (D) Cell migration assessed by wound healing assay. Representative images at 0 h and 48 h are shown (scale bar, 200 μm), with quantitative analysis of migration rate. (E) Cell proliferation assessed by CCK-8 assay. n = 3-6 independent experiments for in vitro. Data in Bto Ewere analyzed via1-way ANOVA, followed by Tukey post hoc analysis. (F, G) RNA sequencing analysis of primary cardiac fibroblasts. Principal component analysis (PCA) plots of global gene expression profiles for cells treated with AngII versus AngII+HNG (F) and AngII versus AngII+MOTS-c (G) (n = 4 independent samples per group). (H, I) Gene Ontology (GO) enrichment analysis of biological processes. Shown are the top enriched terms for differentially expressed genes identified between AngII-treated fibroblasts and the AngII+HNG group (H) [filtered by p < 0.05 and p.adjust (q value) < 0.05], and between AngII-treated fibroblasts and the AngII+MOTS-c group (I) [filtered by p < 0.05 and p.adjust (q value) < 0.2].

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



