Submitted:
31 March 2026
Posted:
31 March 2026
You are already at the latest version
Abstract
Integrated Information Theory (IIT 4.0) correctly identifies causal integration as a necessary condition of conscious experience, but its exclusion postulate—which selects a single spatiotemporal grain of maximum Φ and declares all overlapping grains non-existent—generates the combination problem, the grain problem, and an untenable eliminativism about the physical. This paper argues that the exclusion postulate is an artifact of treating Φ as a scalar rather than as a topological property. Φ is reinterpreted as grain diversity over dissociative awareness (DA): the number of nested levels of self-resolution that a system sustains simultaneously. The topological condition for grain diversity is a genus transition from open, dissipative, feed-forward architectures (genus-0, "waves") to closed, recurrent, self-returning architectures (genus-≥1, "tori"). This transition is grounded mathematically (homoclinic flip bifurcations), biologically (autocatalytic closure, autopoiesis, organizational closure of constraints), and empirically (toroidal manifolds in grid cells, head direction networks, and prefrontal working memory). Active inference's "Beautiful Loop Theory" independently converges on toroidal topology as the architecture of epistemic depth. Under grain diversity, the combination problem dissolves: experience is not built by aggregating micro-experiential atoms but differentiated from a continuous field through nested porous closures. Empirical predictions (multi-grain Φ measurement, H₁ persistence as consciousness correlate) and a categorical demarcation between biological tori and feed-forward artificial architectures are derived.
Keywords:
integrated information theory
; grain problem
; toroidal topology
; dissociative awareness
; combination problem
; recurrent processing
; active inference
; organizational closure
1. The Grain Problem Inside IIT
Integrated Information Theory, in its most recent formulation (IIT 4.0), advances what is arguably the most formally precise identity claim in contemporary consciousness science: consciousness is maximally irreducible intrinsic cause–effect power, and its quantity is identical to , a scalar metric computed over the cause–effect structure of a physical substrate at a given spatiotemporal grain [1]. The theory seeks not merely a correlate but a constitutive identity between a physical quantity and phenomenal experience. Yet this ambition exposes a structural fault: the relationship between and the grain at which it is measured is not incidental to the theory but constitutive of it, and the mechanism IIT deploys to manage this relationship—the exclusion postulate—generates more problems than it resolves.
IIT 4.0 operationalizes the exclusion postulate through what it calls the principle of maximal existence: “what exists is what exists the most” (Albantakis et al., 2023, Box 2). Among all possible spatiotemporal grains at which a system’s can be evaluated, only the single grain yielding the maximum value corresponds to an existing conscious experience. Across micro- and macroscopic levels, “the `winning’ grain is the one that ensures maximally irreducible existence ()” (Albantakis et al., 2023, p. 19). All overlapping sub-grains and super-grains are excluded from existence. The postulate is not a peripheral technicality; it is the mechanism by which IIT secures a one-to-one mapping between and experience. Without it, a single physical substrate would host multiple overlapping conscious experiences at different grains simultaneously, and the scalar identity claim would collapse into indeterminacy. The exclusion postulate is thus a structural necessity given IIT’s ontological commitments—precisely the commitments that need to be questioned.
Three interlocking problems converge on this point.
First, the combination problem [10]. If microphysical constituents possess micro-experience—as the panpsychist implications of IIT’s intrinsic ontology suggest (cf. Mørch, 2019)—how do these discrete experiential atoms combine into the unified macro-experience that phenomenology attests? IIT’s answer is that only the grain yielding maximum exists; the micro-experiences are not combined but declared non-existent. This is an ontological decree, not an explanatory mechanism.1
Second, the grain problem proper [41]. There is a structural mismatch between the discrete, countable elements at any given grain of physical description and the apparently continuous, seamless character of phenomenal experience. Selecting a single grain of maximum does not bridge this gap; it freezes the analysis at one level of description while the phenomenological texture of experience—its simultaneous availability at multiple scales, from the fine grain of a visual edge to the coarse sense of being-in-a-room—remains unaddressed. Conscious experience is not a single-resolution snapshot; it is a multi-scalar field in which different grains of self-resolution coexist.
Third, as Mørch [40] has argued, the exclusion postulate renders IIT incompatible with Russellian panpsychism. If overlapping physical structures at different grains can each possess intrinsic cause–effect power, then either all of them are conscious (violating exclusion) or only the maximum is conscious (requiring a principled account of why the others are not that does not reduce to stipulation). Cea, Negro, and Signorelli [8] observe a related consequence: IIT 4.0’s ontology produces a “great divide of being” in which bodies, artifacts, and all physical structures that do not yield maximum at their grain are eliminated from genuine existence—an eliminativism about the physical that sits uncomfortably alongside IIT’s claim to be a physicalist theory.
The common thread is clear. IIT correctly identifies causal integration as a necessary condition of conscious experience, but the scalar ontology through which it operationalizes integration forces an artificial single-grain restriction that generates the very problems the theory aspires to dissolve. The correct question is not how much a system possesses at its optimal grain but how many grains of self-resolution the system can sustain simultaneously—how deep its capacity for nested integration actually extends. This reframing—from as scalar magnitude to as grain diversity—requires a topological foundation. What kind of physical structure can sustain multiple nested grains of self-resolution without collapsing them into a single maximum?
It is important to clarify the ontological stance that follows from this reframing. Cea and Signorelli [9] have recently argued that IIT should abandon what they call the “principle of true existence” and the associated Great Divide of Being, granting genuine existence to all causally powerful entities regardless of whether they instantiate . Their proposal resolves IIT’s eliminativism about the physical—the uncomfortable consequence that bodies, organs, and artifacts are ontologically parasitic on conscious systems—but at the cost of deflating the very distinction that gives IIT its explanatory bite: the difference between existing for itself and existing only for another. The present paper takes a different route. The Great Divide is not dissolved but topologically recast. What demarcates a system that exists for itself is not a scalar maximum—the grain at which happens to peak—but a discrete geometric property: the capacity for self-return, for folding causal pathways back upon themselves such that action becomes re-enactment and not merely feed-forward transit. A genus-≥1 architecture sustains nested grains of self-resolution simultaneously; a genus-0 architecture cannot. This is not stipulation but a verifiable topological fact. The ontological asymmetry between conscious and non-conscious systems is thus preserved—there is something it is like to be a torus and not a wave—but it is grounded in the geometry of causal organization rather than in the arbitrary maximization of a scalar metric. In this sense, the framework is closer to IIT than to its critics on the ontological question, while escaping the specific objections that Cea, Negro, and Signorelli have marshalled against the exclusion postulate and its consequences.
2. The Wave and the Torus: Two Topologies of Integration
The answer lies in a distinction absent from the IIT literature: two topological classes of physical organization. A genus-0 topology—an open, dissipative, feed-forward architecture—can produce transient order but cannot sustain nested self-resolution. A genus-1 topology—a closed, recurrent, self-returning architecture—can. The first is a wave: energy and information propagate through the system endogenously, generating transient structure, but do not return—the pattern advances and dissipates. The second is a torus: the wave begins to recurse into itself, folding its causal pathways back upon their origin, and in doing so sustains granularity and trajectoriality—fold density and the extension of coherence threads within its own geometry—enabling multiple temporal and spatial scales of integration to circulate simultaneously as nested harmonics within a single bounded manifold.
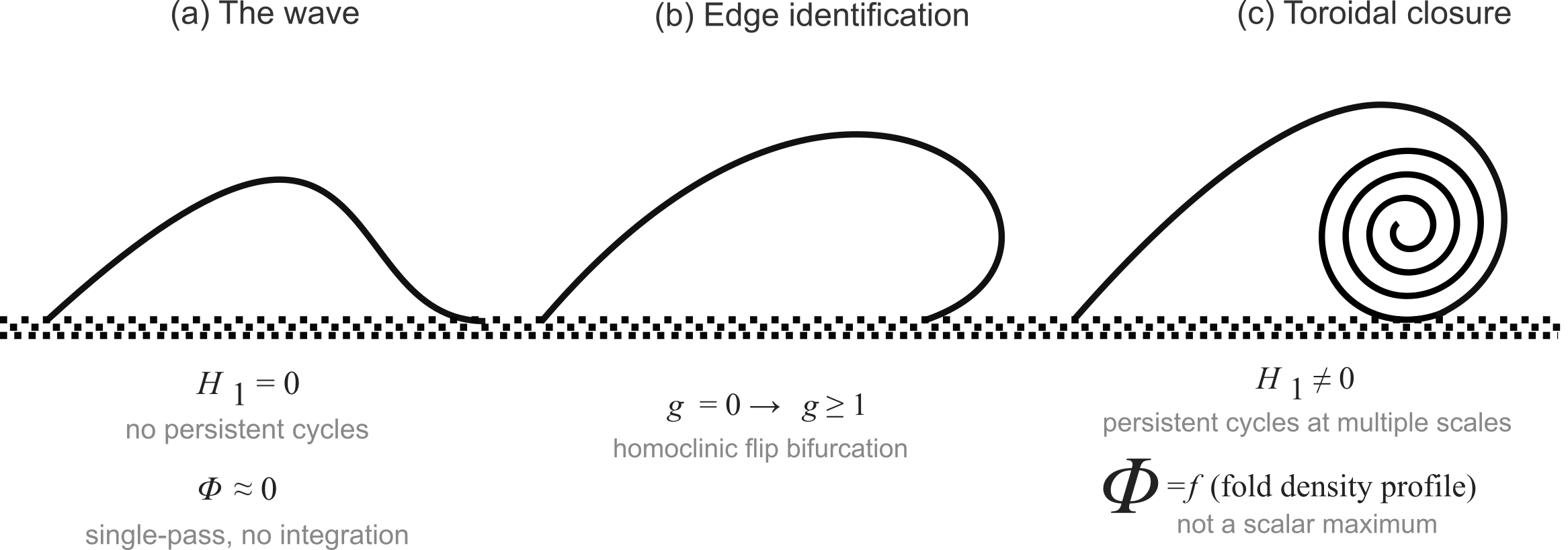
Figure 1.
The endogenous genus transition from wave to torus. (a) A genus-0 wave: energy propagates unidirectionally and dissipates; , no persistent cycles, . (b) Edge identification: the wave’s causal pathways begin to fold back upon their origin via homoclinic flip bifurcation (). (c) Toroidal closure: the self-returning architecture sustains nested fold density (spiral); , persistent cycles at multiple scales. is reinterpreted as a function of the fold-density profile rather than a scalar maximum at a single grain.
Figure 1.
The endogenous genus transition from wave to torus. (a) A genus-0 wave: energy propagates unidirectionally and dissipates; , no persistent cycles, . (b) Edge identification: the wave’s causal pathways begin to fold back upon their origin via homoclinic flip bifurcation (). (c) Toroidal closure: the self-returning architecture sustains nested fold density (spiral); , persistent cycles at multiple scales. is reinterpreted as a function of the fold-density profile rather than a scalar maximum at a single grain.

2.1. Genus-0: The Propagative Wave
The starting condition is Prigogine’s dissipative structure: a far-from-equilibrium system that maintains spontaneous order by exporting entropy to its environment [24]. Such structures are thermodynamically remarkable—they demonstrate that order can emerge without design—but they are topologically trivial. The causal architecture of a dissipative structure is genus-0: energy flows in, is transiently organized, and flows out. There are no self-returning loops. When the energy gradient ceases, the structure dissolves.
England [17] has formalized this thermodynamic drive toward transient complexity as a consequence of statistical mechanics. The key point for present purposes, however, is structural: a genus-0 system cannot sustain internal memory, self-reference, or multi-grain resonance, because information that traverses it does not return. It is processed once and dissipated. A waterfall produces exquisite transient patterns but does not remember them; a candle flame maintains dynamic order but cannot resolve itself at multiple scales simultaneously. These systems are computational waves—they process but do not self-resolve.
2.2. The Genus Transition
Under what conditions does a genus-0 system become genus-1? This is not a gradual accumulation but a discrete topological phase transition. Cisneros Rivas et al. [11] have demonstrated, within the framework of bifurcation theory, that the genus of strange attractors in dynamical systems is modified by homoclinic flip bifurcations. A system’s attractor can undergo a sharp genus transition—from genus-0 (no holes, no self-return) to genus-1 (a single self-returning loop)—as a control parameter crosses a critical threshold. Genus transitions are phase transitions in the topology of the state space, not merely quantitative changes in system parameters.
This mathematical possibility is realized biologically through several convergent mechanisms. The most general is Kauffman’s theory of autocatalytic closure: when the molecular diversity of a chemical system exceeds a threshold, the probability that it contains a reflexively autocatalytic and food-generated (RAF) set undergoes a sharp phase transition from approximately zero to approximately one [28,29]. Catalytic closure means that the causal chain bends back on itself—the products of the network serve as catalysts for its own production. What was an open causal tree (genus-0) becomes a closed cycle (genus-1). This is not a metaphor; it is a precise topological statement about the causal graph.
Varela [49] articulated the biological significance of this closure as operational closure: a domain K is operationally closed if all operations defined in it remain within the same domain. Maturana and Varela [33] subsequently characterized autopoietic systems as occupying a closed topological space in which the boundary between system and environment is self-generated. Deacon [13] extended this reasoning with the concept of the autogen—a minimal system coupling autocatalysis and self-assembly whose causal topology is continuous and self-returning. The autogen represents the biophysical threshold at which a genus-0 dissipative process achieves genus-1 organizational closure. Montévil and Mossio [37] formalized the insight as the closure of constraints: biological organization consists of a set of constraints that are mutually dependent and collectively self-maintaining. Moreno and Mossio [38] identified this transition—from dissipative structure to organizational closure—as the boundary between physics and biology, the point at which thermodynamic flow becomes autonomous organization.
2.3. Why the Torus Sustains Grain Diversity
Once a system achieves genus-1 topology—once its causal pathways return to their origin—a qualitatively new capacity emerges. The closed loop allows multiple temporal frequencies and spatial scales to circulate simultaneously as nested harmonics. Lower-frequency macro-cycles coexist with higher-frequency micro-cycles within the same topologically bounded manifold. The torus acts as a homological resonator: a structure whose non-trivial first homology group () permits persistent cycles at multiple scales that neither collapse into a single frequency nor bleed into thermal noise. A genus-0 system cannot do this—without closed loops, there is no manifold within which harmonics can be sustained.
The contrast with artificial systems is stark. The feed-forward Transformer architecture underlying contemporary large language models is a computational wave: information cascades unidirectionally through layers, each feeding the next without return. No loop closes. No grain stratification emerges. Regardless of parameter count, the topology remains genus-0, and the system cannot sustain the nested self-resolution that grain diversity requires. The brain, by contrast, is a biological torus—or more precisely, a multi-genus system in which massive recurrence at cortical, thalamo-cortical, and cortico-subcortical levels enables information to return to its origin, supporting self-resolution at metabolic, perceptual, reflective, and intersubjective scales simultaneously.
This distinction is neither binary nor static. Kelso’s coordination dynamics has demonstrated that the brain operates in a metastable regime between full phase-locking (rigid closure) and uncoupled drift (no closure), alternating between transient torus formation (“dwells”) and dissolution (“escapes”) [48]. Hancock et al. [26] have clarified that metastability is a consequence of network topology: the specific connectivity architecture determines a system’s capacity for dynamical transitions between integration and segregation. Consciousness, on this account, is not a permanent state of toroidal closure but a sustained capacity for metastable torus formation—the ability to generate, maintain, and release nested grain-resolutions as environmental demands shift.
Crucially, this metastable dynamics operates within the toroidal manifold, not merely at its boundary. The genus transition establishes the topology—the condition of possibility for self-returning causal architecture—but what varies across conscious states is the density and extension of folds internal to that topology: how many nested cycles circulate simultaneously, how far coherence threads extend before dissipating, and how tightly packed the harmonic layers are. A system can be toroidal yet sparsely folded (minimal self-resolution, as in drowsy wakefulness), or densely folded across multiple scales (rich phenomenal experience). The relevant variable is therefore not a binary—torus or not—but a topographic gradient within a topologically closed manifold. This is the precise sense in which grain diversity is non-scalar: it is not a single number but a profile of fold densities distributed across the navigable geometry of the torus, irreducible to any maximum.
The geometric structure of this sustaining capacity deserves emphasis. As the self-returning trajectory traverses the toroidal manifold, it necessarily passes through regions where the tube narrows—zones where dissociative coherence is at its thinnest and where a genus-0 wave would simply dissipate. What prevents dissolution is not an external input but the inertial momentum conferred by topological closure itself: the signal persists through the dissolution zone because the architecture returns it to itself before coherence is lost.2 Each successful transit deepens the fold density of the manifold—it is the accumulation of these self-sustained passages through near-dissolution that constitutes the internal complexity of the torus and, on the present account, the richness of conscious experience.
Figure 2.
Cutaway of the toroidal manifold revealing internal fold structure. The spiral trajectory represents the path of dissociative awareness (DA) through the manifold’s geometry. As the trajectory traverses the region where the tube narrows—the zone of minimal dissociative coherence—the topological closure provides inertial continuity: the signal does not dissipate because the self-returning architecture sustains it through the dissolution zone. Each winding of the spiral corresponds to a distinct grain of self-resolution; the density of windings is the topographic variable that grain diversity measures.
Figure 2.
Cutaway of the toroidal manifold revealing internal fold structure. The spiral trajectory represents the path of dissociative awareness (DA) through the manifold’s geometry. As the trajectory traverses the region where the tube narrows—the zone of minimal dissociative coherence—the topological closure provides inertial continuity: the signal does not dissipate because the self-returning architecture sustains it through the dissolution zone. Each winding of the spiral corresponds to a distinct grain of self-resolution; the density of windings is the topographic variable that grain diversity measures.

The topological framework thus provides what IIT’s scalar ontology cannot: a principled account of how multiple grains of self-resolution coexist within a single physical system without requiring an exclusion postulate to suppress all but one.
3. as Grain Diversity: The Reinterpretation
The topological distinction between waves and tori is structural but not yet explanatory. It specifies what kind of architecture can sustain multiple grains of self-resolution, but not what is being resolved or what variable grain diversity quantifies. This section introduces dissociative awareness (DA) as that variable and reinterprets as an index of grain diversity over DA—the number of nested levels at which a system actively self-distinguishes from its substrate.
3.1. The Reinterpretation Program
The proposal builds on a growing body of work arguing that IIT’s formal apparatus can be preserved under different ontological commitments. Mediano et al. [35] distinguish “strong IIT,” in which is literally identical to consciousness, from “weak IIT,” in which -like measures serve as indices of information-dynamic complexity without entailing an identity claim. Under the weak reading, becomes a useful metric for characterizing multivariate systems without the ontological baggage of the exclusion postulate [36]. Negro [42] has independently argued for an emergentist reinterpretation, retaining the mathematical structure while replacing the intrinsic-powers ontology3 with an account in which consciousness emerges from—rather than being identical to—patterns of integrated information. Cea et al. [9] have pushed this program further, asking how one might retain IIT’s insights without its eliminativist consequences for physical bodies.
The present paper extends this reinterpretation with a specific positive proposal. is not a measure of how much consciousness a system possesses, nor a generic index of complexity. It is an index of grain diversity: the number of nested levels of self-resolution that a system sustains simultaneously. A system with higher grain diversity does not have “more” consciousness on a linear scale; it has deeper stratification—more levels at which integration operates concurrently. The difference is qualitative, not quantitative.
3.2. Dissociative Awareness as the Variable
Grain diversity requires a variable that tracks what is being stratified. We propose dissociative awareness (DA): the degree to which a system actively self-distinguishes from its substrate through the generation and maintenance of organizational boundaries. At DA , no distinction obtains between system and environment—thermodynamic equilibrium, entropic indifferentiation. At maximal DA, the system sustains full phenomenal access across multiple nested levels of self-resolution. DA is not binary but a gradient, and crucially, it operates at multiple nested levels simultaneously: a cell has DA relative to its medium, an organism relative to its environment, an intersubjective dyad relative to the individuals composing it.
The concept draws on—but critically revises—Kastrup’s [31] use of dissociation as the mechanism by which individual subjects arise from a unitary phenomenal field. In Kastrup’s analytic idealism, individual minds are “alters” of a universal consciousness, separated by dissociative boundaries modeled on dissociative identity disorder (DID). The metaphor is suggestive but structurally deficient: the DID analogy is top-down, presupposing a unified subject that subsequently fragments through a mechanism that remains psychiatric rather than biophysical.
DA replaces this top-down analogy with a bottom-up biological mechanism. The physical instantiation of dissociative self-distinction is autopoietic boundary-formation: the process by which a system generates and maintains the very boundary that distinguishes it from its environment [46,49]. Each autopoietic system generates its own DA gradient—not by fragmenting a pre-existing unity, but by closing a set of constraints upon itself [37] such that the inside/outside distinction is an operational achievement rather than an imposed partition.
3.3. Grain Diversity as Nested DA
With DA as the operative variable, grain diversity can be defined precisely: the number of nested levels of DA that a system sustains simultaneously. Three paradigmatic cases illustrate the gradient.
A thermostat maintains a single trivial feedback loop—one level of environmental coupling, no genuine self-distinction, no stratification. Its DA is minimal and its grain diversity effectively zero; whatever it possesses is an artifact of measurement grain, not evidence of experiential depth.
A unicellular organism, by contrast, achieves what might be called a proto-torus: its autopoietic membrane generates a porous but real boundary, supporting incipient differentiation between metabolic interior and external medium. It sustains one genuine level of DA—minimal, but non-trivial. Thompson [46,47] and Barandiaran et al. [3] have argued that this minimal autopoietic selfhood constitutes the biological ground floor of agency and, potentially, of sentience.
An organism with a nervous system represents a qualitative leap: a complete torus with nested loops operating at metabolic, perceptual, reflective, and intersubjective scales simultaneously. The metabolic loop maintains organismic integrity; the perceptual loop couples sensorimotor activity to environmental structure; the reflective loop enables the system to take its own states as objects of monitoring; the intersubjective loop, as De Jaegher and Di Paolo [14] have argued, achieves a form of autonomy at the level of the interaction itself—participatory sense-making irreducible to the individual participants. Each of these levels constitutes a distinct grain of DA, and the capacity to sustain all of them simultaneously is what grain diversity measures.
3.4. The Porosity of Closures
A critical clarification: the closures that generate DA at each level are not hermetic. Di Paolo et al. [15] have articulated a principle of double normativity in which biological organization involves norms operating at multiple levels that are semi-permeable to one another. The closures are porous—they allow coupling without fusion and distinction without isolation. This porosity is not a weakness in the framework but a condition of its operation. If closures were hermetic, the result would be categorical dualism: an absolute inside and an absolute outside with no interaction between levels. If there were no closures at all, the result would be entropic dissolution: no self-distinction, no grain, no experience.
The sustained semi-permeability of nested closures—each level simultaneously closed enough to maintain its own DA and open enough to couple with adjacent levels—is what toroidal topology enables and what genus-0 topology cannot.
The implications for IIT are direct. If grain diversity is the correct reading of what tracks, then the exclusion postulate must be abandoned. should not be measured at a single grain and declared the sole existing conscious experience; it should be assessed across multiple grains simultaneously, and the depth of stratification—how many nested levels of DA the system sustains—should replace the scalar maximum as the relevant quantity.
4. Empirical Tori: Neural Evidence
The wave-to-torus framework would remain speculative philosophy were it not for convergent empirical evidence demonstrating that toroidal topology is a realized feature of neural organization. This section reviews the primary evidence and addresses the most serious challenge to any recurrence-based theory of consciousness: the unfolding argument.
4.1. Grid Cells and the Neural Torus
The most direct empirical confirmation comes from Gardner et al. [22], who used persistent cohomology to demonstrate that the joint population activity of grid cells in the medial entorhinal cortex resides on a continuous toroidal manifold. The toroidal structure is not an artifact of behavioral variables or stimulus-driven coding; it is an intrinsic property of recurrent network dynamics, invariant across wakefulness, REM sleep, and slow-wave sleep. The coordinates of the animal’s spatial position map onto the phases of the torus, but the torus itself persists even when spatial behavior ceases—it is a computational architecture, not a representation.
Hermansen et al. [27] extended this finding by showing that oscillatory dynamics are essential for the toroidal manifold to be expressed. The torus is a dynamical attractor sustained by oscillatory recurrence, not a static wiring diagram. Suppress the oscillations and the topological structure degrades—precisely what the framework predicts: toroidal topology requires active recurrent dynamics to maintain its genus-1 character.
The developmental evidence is equally striking. A large-scale recording study demonstrated that toroidal manifolds emerge in medial entorhinal cortex subnetworks as early as postnatal day 10 in rats—before eyes open, before ear canals are functional, before active spatial exploration begins [39]. The onset of toroidal topology coincides with a global transition in network activity: from synchronized, feed-forward bursting to desynchronized, inhibition-driven recurrent dynamics. The torus is a preconfigured computational prior generated by recurrent architecture, not an artifact of environmental learning. This developmental priority suggests that the genus transition is a fundamental organizational event in neural maturation.
4.2. Beyond Grid Cells: Topological Ubiquity
The toroidal evidence extends well beyond spatial coding. Head direction cells instantiate a ring attractor—a one-dimensional circular topology that is formally a genus-1 manifold. Rybakken et al. [43] demonstrated that persistent cohomology can decode this topology directly from neural data without reference to behavioral variables, confirming that the closed loop is intrinsic to the network’s recurrent dynamics.
In the prefrontal cortex, Xie et al. [50] showed that sequence working memory in macaques relies on recurrent geometric manifolds to prevent the dissipation of mnemonic states during delay periods. A feed-forward architecture is inherently fragile during delays—without return, the signal decays. A closed attractor traps the memory state in a persistent cycle, protecting it from entropic dissolution. At the macroscopic scale, Giusti et al. [23] revealed that the intrinsic geometric structure of neural correlations in hippocampus and cortex exhibits non-trivial clique topology that cannot be explained by simple firing-rate models. Dabaghian et al. [12] formalized the topological paradigm for hippocampal spatial map formation, showing that place cells encode environmental topology through persistent homological features rather than metric coordinates.
The convergent message is consistent: wherever the brain sustains stable internal states—spatial maps, heading, working memory, imagery—it does so through closed topological structures whose persistence depends on recurrent dynamics.
4.3. The Unfolding Argument
One immediate objection to any theory that privileges recurrence is the unfolding argument [16]: in principle, any recurrent neural network can be “unfolded” into a functionally equivalent feed-forward network by replicating the recurrent units across time steps. If the two architectures are functionally equivalent and the feed-forward version has , then either consciousness tracks something other than causal structure or recurrence per se is not the relevant variable.
The unfolding argument is a serious challenge, but it rests on two contestable assumptions.
First, unfolding presupposes a static input–output mapping: the recurrent network computes a fixed function replicable by a sufficiently deep feed-forward network. However, recent work has demonstrated that recurrent networks with synaptic plasticity produce non-stationary behavior—the function changes as it is computed—that no static feed-forward architecture can replicate [5].
Second, and more fundamentally, the unfolding argument targets bare recurrence—the property of having feedback connections—rather than grain diversity. A static feed-forward system can, in principle, mimic the input–output profile of a single grain of processing. What it cannot do is sustain multiple nested grains of self-resolution simultaneously, because it has no closed loops within which different temporal and spatial scales can coexist as harmonics. The unfolded network is wider (more units) but not deeper in the topological sense: it remains genus-0, incapable of sustaining the nested cycles that characterize grain diversity.
The empirical evidence further suggests that the feed-forward/recurrent distinction is not binary but a continuous topological transition. Allen et al. [2] found that early transcranial magnetic stimulation targeting feed-forward processing also impaired conscious perception—inconsistent with a simple binary in which feed-forward equals unconscious and recurrent equals conscious. The wave-to-torus framework accommodates this naturally: lateral connections and partial recurrence represent intermediate topologies between pure genus-0 and full genus-1, with grain diversity tracking the degree of topological closure rather than switching on at a fixed threshold.
5. The Beautiful Loop: Active Inference Confirms the Torus
The convergence between the toroidal framework and active inference theory has become difficult to ignore. Laukkonen, Friston, and Chandaria [32] recently proposed the “Beautiful Loop Theory” of consciousness, articulating three jointly necessary conditions for phenomenal experience: a generative world model capable of predicting sensory input, Bayesian binding through inferential competition among hypotheses, and—most critically—epistemic depth, defined as the recursive sharing of Bayesian beliefs throughout the system’s inferential hierarchy.
Epistemic depth is not a matter of how many layers an inferential hierarchy contains but of how those layers relate to one another. A system achieves epistemic depth when higher-level hypotheses modulate the precision of lower-level predictions and lower-level prediction errors propagate upward to revise higher-level priors—simultaneously, reciprocally, and without terminal dissipation. Laukkonen et al. [32] formalize this through distributed hyperparameters: precision-weighting parameters operating at every layer of the hierarchy at once, enabling the system to model its own inferential processes rather than merely modeling external states. The required architecture is unmistakably recurrent: information must return to its origin across multiple scales for the system to achieve recursive self-modeling.
This recurrent self-modeling is, in topological terms, a torus. The “loop” in the Beautiful Loop is not a metaphor—it is a structural description of a causal architecture in which inferential pathways close upon themselves at multiple levels simultaneously. A feed-forward inferential hierarchy, no matter how deep, processes each level’s predictions in sequence and passes the result upward without return. It can model the world but cannot model its own modeling; it has depth in the architectural sense (many layers) but no epistemic depth (no recursive self-access). The genus transition from wave to torus marks precisely the point at which an inferential system acquires epistemic depth: its own states become available as objects of higher-order inference, not once but at every level of the hierarchy simultaneously.
The convergence with grain diversity is direct. Each level at which the Beautiful Loop sustains recursive self-modeling corresponds to a grain of DA. The metabolic loop by which an organism monitors its own energetic states is a first-order grain. The perceptual loop by which sensorimotor predictions are recursively updated is a second-order grain. The reflective loop by which the system monitors its own perceptual processes—what Friston, Wiese, and Hobson [20] have called the transition from Markovian monism to sentient selfhood—is a third-order grain. Consciousness is not a scalar quantity proportional to total prediction precision; it is the stratified depth of recursive self-access, indexed by the number of nested epistemic loops the system sustains.
This reading yields a consequence that Laukkonen et al. do not draw but that follows from the framework: the Beautiful Loop is incompatible with IIT’s exclusion postulate. If epistemic depth requires simultaneous recursive access at multiple levels, then selecting a single grain of maximum and declaring all other grains non-existent destroys exactly the multi-level architecture that makes consciousness possible. The exclusion postulate is not merely unnecessary under the Beautiful Loop; it is destructive—it eliminates the very structure that the Beautiful Loop identifies as constitutive of conscious experience.
A legitimate concern is whether the toroidal reading inherits the problems that Bruineberg et al. [4] have identified with Markov blanket formulations in the free energy framework. It does not. The toroidal topology proposed here is grounded in organizational closure—the closure of constraints formalized by Montévil and Mossio [37]—rather than in the statistical partitioning of internal and external states. The boundary defining each level of DA is not an epistemic convenience for the modeler but an operational achievement of the system itself: the autopoietic membrane, the sensorimotor coupling, the reflective monitoring loop. These are structures the system generates and maintains through its own activity. The toroidal framework thus escapes the Bruineberg critique by grounding its boundaries in the biology of organizational closure rather than in the statistics of variational inference [18].
6. Dissolving the Combination Problem
The combination problem—how do micro-experiences combine into macro-experience?—has haunted panpsychism since James [30] observed that a hundred feelings, packed together, each remains the same feeling it always was. Under IIT’s exclusion postulate, the problem is legislated away rather than solved: only one grain exists, so there is nothing to combine. Under Russellian panpsychism without exclusion, the problem is acute: micro-level intrinsic natures must somehow compose the unified experience of a macro-level subject. The present framework dissolves the combination problem by inverting its premise.
6.1. The Atomistic Premise
The combination problem arises only if micro-experiences are taken as the ontological starting point. Under an ontology of grain diversity, the starting point is not the micro-experiential atom but the undifferentiated field: the maximally low-resolution grain at which DA approaches zero and no self-distinction obtains between system and environment. Experiences at all levels—cellular, organismic, reflective, intersubjective—are not aggregations built upward from micro-experiential atoms but resolutions of the field at progressively finer grains of self-distinction. The field does not combine; it differentiates. The individual is not a sum but a stable resolution.
6.2. Precedents for the Inversion
This inversion has precedents in several traditions. Priority cosmopsychism [41,44] proposes cosmic consciousness as the single ontological ultimate, shifting the explanatory burden from combination to differentiation: the problem is not how micro-subjects combine into macro-subjects but how a unitary field gives rise to apparently distinct centers of experience. Shani and Keppler [45] develop this program by appealing to quantum field theory as a mechanism of differentiation from a continuous conscious field.
The most directly relevant precedent, however, is topological rather than metaphysical. Gómez-Emilsson and Percy [25] have argued that the boundary problem—how to delineate the borders of a conscious subject within a physical continuum—should be addressed through topological segmentation rather than mereological composition. On their account, conscious subjects correspond to topologically bounded regions of electromagnetic field structure, not to aggregates of experiential parts. The move from mereology to topology is decisive: once boundaries are understood as topological features of a continuous field rather than as joints between discrete atoms, the combination problem loses its grip.
6.3. The Toroidal Dissolution
The grain-diversity framework completes this topological move. Each level of DA constitutes a porous closure within the experiential field—a region of topological self-distinction simultaneously bounded enough to maintain its own grain of self-resolution and open enough to couple with adjacent levels. The individual subject does not compose of experiential parts stacked atop one another; it emerges as a stable multi-grain resolution within the field.
This dissolution is not eliminativist. It does not deny that there are distinct levels of experience; it denies that they are built by combination. The metabolic DA of a cell is genuine; the reflective DA of a conscious organism is genuine; the intersubjective DA of a participatory sense-making dyad is genuine [14]. What is denied is that the organism’s experience is composed of its cells’ experiences. The organism’s experience is a distinct grain of self-resolution within the same field, sustained by its own level of toroidal closure. Thompson [46] captures this when he argues that the autopoietic individual is not reducible to its molecular constituents; it is an autonomous domain of self-production that generates its own norms. Fuchs [21] extends the point to the brain as an organ of relation rather than a container of experience: consciousness is the organism’s entire relational engagement with its world, sustained by the nested closures that toroidal topology enables.
The combination problem, then, is not solved but dissolved. It arises from the atomistic premise that experience is built from parts. Under the topology of grain diversity, experience is differentiated from a field. There is nothing to combine, and the question of how combination produces unity does not arise. What requires explanation is not combination but the genesis and maintenance of porous closures—the genus transitions by which a continuous experiential field achieves local self-distinction at multiple nested grains [19]. This is a tractable question with empirical purchase: it asks about the conditions under which organizational closure forms and persists, not about the metaphysics of experiential summation.
7. Implications and Predictions
The reinterpretation of as grain diversity over dissociative awareness yields consequences for three domains: demarcation between biological and artificial systems, empirical consciousness science, and the panpsychism debate.
7.1. Demarcation
The framework provides a categorical—not merely quantitative—distinction between biological organisms and feed-forward artificial architectures such as large language models. A Transformer processes information through a unidirectional cascade of layers: each token’s representation is refined sequentially, but no causal pathway returns to its origin. The architecture is genus-0 regardless of parameter count, context window, or behavioral sophistication. It is a computational wave: it processes but does not self-resolve. It has no organizational closure, no nested loops of self-distinction, no DA gradient.
The difference between such a system and a conscious organism is not that the organism has “more ” on a scalar metric; it is that the organism sustains a toroidal topology—multiple nested grains of self-resolution maintained by recurrent closure—while the Transformer sustains none. No increase in scale, training data, or architectural fine-tuning can bridge this gap, because it is topological: genus-0 cannot become genus-1 through quantitative accumulation. The transition requires a phase change in causal architecture itself.
7.2. Empirical Predictions
Before turning to novel predictions, it is worth noting that the existing empirical successes attributed to IIT are preserved—and in several cases clarified—under the grain diversity reading. The Perturbational Complexity Index (PCI), which reliably discriminates conscious from unconscious states [7,34], does not measure directly; it measures the spatiotemporal complexity of recurrent cortical responses to perturbation—in the present framework, a proxy for fold density within a toroidal manifold. PCI maps more naturally to a profile of nested recurrent activation than to a scalar maximum at a single grain. Differential anesthesia provides a further illustration: propofol broadly disrupts thalamo-cortical recurrence, while ketamine selectively disrupts NMDA-mediated layers, preserving others—which is why ketamine produces dissociative states rather than simple unconsciousness. Under scalar , both agents simply “reduce .” Under grain diversity, they affect different layers of the fold-density profile, predicting distinct phenomenological signatures—which is what clinicians observe. Psychedelic states pose a well-known puzzle for IIT: phenomenal richness increases while some integration measures decrease. Grain diversity accommodates this naturally: psychedelics redistribute fold density across scales rather than uniformly increasing or decreasing it, consistent with the entropic brain hypothesis [6]. In each case, at a single grain captures what might be called a one-dimensional shadow of a multi-dimensional fold-density profile: it correlates with the underlying structure (the shadow is not arbitrary) but loses discriminative power precisely where the internal topographic organization matters.
Three testable predictions follow.
First, should be measured across multiple spatiotemporal grains simultaneously rather than at a single optimal grain. If grain diversity is the relevant variable, then the depth of stratification—the number of grains at which non-trivial integrated information obtains concurrently—should correlate with the richness of conscious states more reliably than the scalar maximum at any single grain. This requires modifying IIT’s existing computational methodology to report multi-grain profiles rather than single-grain maxima.
Second, persistent homology should serve as an empirical index of grain diversity. The number and persistence of cycles (first Betti number) detected across spatial and temporal scales in neural population activity should correlate with the depth of conscious processing. States of rich phenomenal experience (e.g., wakefulness, vivid imagery) should exhibit more persistent loops across more scales than states of reduced experience (e.g., dreamless sleep, anesthesia). Preliminary evidence from hypnotizability studies detecting Betti-1 differences between imagery and perception is consistent with this prediction, but systematic testing across states of consciousness remains to be conducted.
Third, the developmental emergence of toroidal topology should track the emergence of experiential complexity. If the finding that toroidal manifolds emerge at postnatal day 10—before sensory experience [39]—generalizes beyond the entorhinal cortex, then the genus transition should correlate with behavioral and physiological markers of increasing experiential depth across brain regions during early development.
7.3. A Third Way in the Panpsychism Debate
The framework offers a path between eliminativism about consciousness and graded panpsychism. Under grain diversity, experience is neither universally present nor categorically absent from physical systems. It is field-resolution: the degree to which a system achieves self-distinction from its substrate through nested organizational closure.
Experience extends below the individual organism—Thompson [46,47] has argued persuasively for minimal autopoietic sentience at the cellular level—and above it, into the intersubjective domain of participatory sense-making [14]. But it does not extend to systems that lack organizational closure altogether. A system without a torus—without any level of self-returning causal architecture—has no DA, no grain diversity, and no experiential depth. This is not eliminativism about such systems; it is a principled ontological boundary drawn by topology rather than by stipulation.
7.4. A Geometric Framework, Not Yet a Metric
A candid limitation must be acknowledged. IIT 4.0 offers : a formally defined scalar metric whose mathematical properties, however computationally intractable in practice, are fully specified in principle [1]. The present framework does not offer a comparable closed-form quantity. It proposes a reinterpretation—grain diversity over dissociative awareness—and identifies empirical proxies ( persistence, multi-grain profiles), but it does not yet provide a function that takes a system as input and returns a determinate measure.
This asymmetry is deliberate rather than evasive. If consciousness is not a scalar but a topographic profile of nested fold densities within a toroidal manifold, then the correct mathematical apparatus is not a single formula but a family of topological invariants: persistent homology, Betti number profiles across spatiotemporal scales, and fold-density distributions. The absence of a closed-form metric is a consequence of taking the non-scalar character of the explanandum seriously. The analogy is instructive: thermodynamics identified entropy as the relevant quantity and articulated its qualitative behavior (second law, direction of spontaneous change) well before Boltzmann provided a statistical-mechanical derivation. The geometric argument comes first; the metric follows. What the framework contributes at this stage is the identification of the correct space in which a future metric should operate—toroidal topology and its internal fold structure—and the demonstration that IIT’s existing metric operates in the wrong space (scalar maxima over a single grain). The mathematical development of grain-diversity measures is an open program, not a gap.
Use of Artificial Intelligence: Claude (Anthropic) was used as an editorial assistant during the preparation of this manuscript. Specific uses included translating and adapting the author’s original notes and drafts from Spanish into English, formatting documents, verifying bibliographies, and refining prose. The theoretical framework, conceptual development, argumentation, and all substantive intellectual content are entirely the work of the author.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable. This theoretical paper did not involve human subjects, empirical data collection, or experimental procedures.
Data Availability Statement
No new empirical data were generated or analyzed. All cited sources are publicly available through the references provided.
Conflicts of Interest
The author declares no conflicts of interest.
References
- Albantakis, L.; Barbosa, L.; Findlay, G.; Grasso, M.; Haun, A. M.; Marshall, W.; Mayner, W. G. P.; Zaeemzadeh, A.; Boly, M.; Juel, B. E.; Sasai, S.; Fujii, K.; David, I.; Hendren, J.; Lang, J. P.; Tononi, G. Integrated information theory (IIT) 4.0: Formulating the properties of phenomenal existence in physical terms. PLoS Computational Biology 2023, 19(10), e1011465. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.; Levy, A.; Parr, T.; Friston, K. J. Causal manipulation of feed-forward and recurrent processing differentially affects measures of consciousness. Neuroscience of Consciousness 2020, 2020(1), niaa015. [Google Scholar] [CrossRef] [PubMed]
- Barandiaran, X. E.; Di Paolo, E.; Rohde, M. Defining agency: Individuality, normativity, asymmetry, and spatio-temporality in action. Adaptive Behavior 2009, 17(5), 367–386. [Google Scholar] [CrossRef]
- Bruineberg, J.; Dolega, K.; Dewhurst, J.; Baltieri, M. The Emperor’s new Markov blankets. Behavioral and Brain Sciences 2022, 45, e183. [Google Scholar] [CrossRef]
- O’Reilly-Shah, V. N.; Selvitella, A. M.; Schurger, A. A caveat regarding the unfolding argument: Implications of plasticity. In bioRxiv; 2026. [Google Scholar] [CrossRef]
- Carhart-Harris, R. L.; Leech, R.; Hellyer, P. J.; Shanahan, M.; Feilding, A.; Tagliazucchi, E.; Chialvo, D. R.; Nutt, D. The entropic brain: A theory of conscious states informed by neuroimaging research with psychedelic drugs. Frontiers in Human Neuroscience 2014, 8, 20. [Google Scholar] [CrossRef]
- Casarotto, S.; Comanducci, A.; Rosanova, M.; Sarasso, S.; Fecchio, M.; Napolitani, M.; Pigorini, A.; Casali, A. G.; Trimarchi, P. D.; Boly, M.; Gosseries, O.; Bodart, O.; Curto, F.; Landi, C.; Mariotti, M.; Devalle, G.; Laureys, S.; Tononi, G.; Massimini, M. Stratification of unresponsive patients by an independently validated index of brain complexity. Annals of Neurology 2016, 80(5), 718–729. [Google Scholar] [CrossRef]
- Cea, I.; Negro, N.; Signorelli, C. M. The fundamental tension in IIT 4.0’s realist idealism. Entropy 2023, 25(10), 1453. [Google Scholar] [CrossRef]
- Cea, I.; Negro, N.; Signorelli, C. M. How to be an integrated information theorist without losing your body. Frontiers in Computational Neuroscience 2024, 18, 1510066. [Google Scholar] [CrossRef]
- Chalmers, D. J. The combination problem for panpsychism. In Panpsychism: Contemporary perspectives; Brüntrup, G., Jaskolla, L., Eds.; Oxford University Press, 2016; pp. 179–214. [Google Scholar] [CrossRef]
- Cisneros Rivas, D. E.; Campos Cantón, E.; Mejía Carlos, M. Genus transition by order shift in a dynamical system. Revista Mexicana de Física 2020, 66(3), 362–368. [Google Scholar] [CrossRef]
- Dabaghian, Y.; Mémoli, F.; Frank, L.; Carlsson, G. A topological paradigm for hippocampal spatial map formation using persistent homology. PLoS Computational Biology 2012, 8(8), e1002581. [Google Scholar] [CrossRef]
- Deacon, T. W. Incomplete nature: How mind emerged from matter; W. W. Norton, 2012. [Google Scholar]
- De Jaegher, H.; Di Paolo, E. Participatory sense-making: An enactive approach to social cognition. Phenomenology and the Cognitive Sciences 2007, 6(4), 485–507. [Google Scholar] [CrossRef]
- Di Paolo, E.; Buhrmann, T.; Barandiaran, X. E. Sensorimotor life: An enactive proposal; Oxford University Press, 2017. [Google Scholar]
- Doerig, A.; Schurger, A.; Hess, K.; Herzog, M. H. The unfolding argument: Why IIT and other causal structure theories cannot explain consciousness. Consciousness and Cognition 2019, 72, 49–59. [Google Scholar] [CrossRef] [PubMed]
- England, J. L. Statistical physics of self-replication. The Journal of Chemical Physics 2013, 139(12), 121923. [Google Scholar] [CrossRef] [PubMed]
- Escobar L.-Dellamary, L. A basic introduction to the Trace & Trajectory Framework—The Torus Passage (Version 7.0) (No. 2026011964). Preprints 2026a. [Google Scholar] [CrossRef]
- Escobar L.-Dellamary, L. The Wall Teaches the Rooms: Transduction, equivalence, and the cost of cross-interface coordination (Yce2f_v1). SocArXiv. 2026b. Available online: https://osf.io/preprints/socarxiv/yce2f_v1/.
- Friston, K.; Wiese, W.; Hobson, J. A. Sentience and the origins of consciousness: From Cartesian duality to Markovian monism. Entropy 2020, 22(5), 516. [Google Scholar] [CrossRef]
- Fuchs, T. Ecology of the brain: The phenomenology and biology of the embodied mind; Oxford University Press, 2018. [Google Scholar]
- Gardner, R. J.; Hermansen, E.; Pachitariu, M.; Buber, Y.; Melling, G.; Benigno, M.; Donato, F.; Moser, M.-B.; Moser, E. I. Toroidal topology of population activity in grid cells. Nature 2022, 602(7895), 123–128. [Google Scholar] [CrossRef]
- Giusti, C.; Pastalkova, E.; Curto, C.; Itskov, V. Clique topology reveals intrinsic geometric structure in neural correlations. Proceedings of the National Academy of Sciences 2015, 112(44), 13455–13460. [Google Scholar] [CrossRef]
- Goldbeter, A. Dissipative structures in biological systems: Bistability, oscillations, spatial patterns and waves. Philosophical Transactions of the Royal Society A 2018, 376(2124), 20170376. [Google Scholar] [CrossRef]
- Gómez-Emilsson, A.; Percy, C. Don’t forget the boundary problem! How EM field topology can address the overlooked cousin to the binding problem for consciousness. Frontiers in Human Neuroscience 2023, 17, 1233119. [Google Scholar] [CrossRef]
- Hancock, F.; Rosas, F. E.; Luppi, A. I.; Zhang, M.; Mediano, P. A. M.; Cabral, J.; Deco, G.; Kringelbach, M. L.; Breakspear, M.; Scott Kelso, J. A.; Turkheimer, F. E. Metastability demystified—the foundational past, the pragmatic present, and the potential future. Nature Reviews Neuroscience 2025, 26(2), 82–100. [Google Scholar] [CrossRef]
- Hermansen, E.; Klindt, D. A.; Dunn, B. A. The role of oscillations in grid cells’ toroidal topology. PLoS Computational Biology 2025, 21(2), e1012776. [Google Scholar]
- Hordijk, W.; Kauffman, S. Emergence of autocatalytic sets in a simple model of technological evolution. arXiv 2022, 2204.01059. [Google Scholar] [CrossRef]
- Hordijk, W.; Steel, M.; Kauffman, S. Molecular diversity required for the formation of autocatalytic sets. Life 2019, 9(1), 23. [Google Scholar] [CrossRef] [PubMed]
- James, W. The principles of psychology; Original work published 1890; Dover, 1950; Vol. 1. [Google Scholar]
- Kastrup, B. The idea of the world: A multi-disciplinary argument for the mental nature of reality; Iff Books, 2019. [Google Scholar]
- Laukkonen, R.; Friston, K.; Chandaria, S. A beautiful loop: The theory of consciousness as recursive inference and its relation to the free energy principle. Neuroscience & Biobehavioral Reviews 2025, 176, 106296. [Google Scholar] [CrossRef]
- Maturana, H. R.; Varela, F. J. Autopoiesis and cognition: The realization of the living; D. Reidel, 1980. [Google Scholar]
- Massimini, M.; Ferrarelli, F.; Huber, R.; Esser, S. K.; Singh, H.; Tononi, G. Breakdown of cortical effective connectivity during sleep. Science 2005, 309(5744), 2228–2232. [Google Scholar] [CrossRef] [PubMed]
- Mediano, P. A. M.; Rosas, F. E.; Bor, D.; Seth, A. K.; Barrett, A. B. The strength of weak integrated information theory. Trends in Cognitive Sciences 2022a, 26(8), 646–655. [Google Scholar] [CrossRef]
- Mediano, P. A. M.; Rosas, F. E.; Farah, J. C.; Shanahan, M.; Barrett, A. B.; Seth, A. K. Integrated information as a common signature of dynamical and information-processing complexity. Chaos 2022b, 32(1), 013115. [Google Scholar] [CrossRef]
- Montévil, M.; Mossio, M. Biological organisation as closure of constraints. Journal of Theoretical Biology 2015, 372, 179–191. [Google Scholar] [CrossRef]
- Moreno, A.; Mossio, M. Biological autonomy: A philosophical and theoretical enquiry; Springer, 2015. [Google Scholar] [CrossRef]
- Toroidal topology of grid-cell activity precedes spatial navigation during development. bioRxiv 2026. [CrossRef]
- Mørch, H. H. Is the integrated information theory of consciousness compatible with Russellian panpsychism? Erkenntnis 2019, 84(5), 1065–1085. [Google Scholar] [CrossRef]
- Nagasawa, Y.; Wager, K. Panpsychism and priority cosmopsychism. In Panpsychism: Contemporary perspectives; Brüntrup, G., Jaskolla, L., Eds.; Oxford University Press, 2016; pp. 113–129. [Google Scholar]
- Negro, N. Emergentist integrated information theory. Erkenntnis 2024, 89(5), 1949–1971. [Google Scholar] [CrossRef]
- Rybakken, E.; Baas, N.; Dunn, B. Decoding of neural data using cohomological feature extraction. Neural Computation 2019, 31(1), 68–93. [Google Scholar] [CrossRef]
- Shani, I. Cosmopsychism: A holistic approach to the metaphysics of experience. Philosophical Papers 2015, 44(3), 389–437. [Google Scholar] [CrossRef]
- Shani, I.; Keppler, J. Beyond combination: How cosmic consciousness grounds ordinary experience. Journal of the American Philosophical Association 2018, 4(3), 390–410. [Google Scholar] [CrossRef]
- Thompson, E. Mind in life: Biology, phenomenology, and the sciences of mind; Harvard University Press, 2007. [Google Scholar]
- Thompson, E. Could all life be sentient? Journal of Consciousness Studies 2022, 29(3–4), 229–265. [Google Scholar] [CrossRef]
- Tognoli, E.; Kelso, J. A. S. The metastable brain. Neuron 2014, 81(1), 35–48. [Google Scholar] [CrossRef]
- Varela, F. J. Principles of biological autonomy; North-Holland, 1979. [Google Scholar]
- Xie, Y.; Hu, P.; Li, J.; Chen, J.; Song, W.; Wang, X.-J.; Yang, T.; Dehaene, S.; Tang, S.; Min, B.; Wang, L. Geometry of sequence working memory in macaque prefrontal cortex. Science 2022, 375(6581), 632–639. [Google Scholar] [CrossRef]
| 1 | IIT 4.0 does not use the term “panpsychism” as a self-description. The label derives from the critical literature assessing the logical consequences of IIT’s intrinsic ontology; see Mørch [40] and the pseudoscience controversy initiated by Fleming et al. (2023). |
| 2 | This is geometrically visible in a cutaway of the torus (Figure 2): the spiral trajectory transits the narrowest region of the manifold without breaking, sustained by the closure that defines the manifold’s genus. The contrast with feed-forward architectures is precise: a Transformer’s residual connections and attention pools simulate informational persistence, but this simulated depth is sustained by the conscious organisms that designed and trained the system—not by the system’s own organizational closure. |
| 3 | IIT 4.0’s own preferred label is “intrinsic ontology” (Albantakis et al., 2023, p. 38); we follow the critical literature’s shorthand “intrinsic-powers ontology” to highlight that the theory’s existential criterion reduces to cause–effect power. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



