Submitted:
27 March 2026
Posted:
30 March 2026
You are already at the latest version
Abstract
The gut microbiota plays a crucial role in digestion, metabolism, immunity, and inflammation. Dietary components, including food additives, significantly influence its composition and function. With the widespread use of food additives in processed foods and increasing consumption of such products, concerns arise regarding their long-term effects on gut health. This review explores the relationship between food additives and the gut microbiota, focusing on their potential role in gut dysbiosis and disease. Recent research highlights how specific additives including preservatives may disrupt microbial balance, alter metabolic pathways, and contribute to inflammation-related disorders. While some additives may have minimal effects, others could negatively impact gut health. This review focused on Carboxymethyl Cellulose (CMC) and Carrageenan (CGN) and how they can potentially create an inflammatory-prone environment. Evidence indicates that both CMC and CGN can alter the microbiota, promoting pro-inflammatory profiles, although the effects depend on dose, exposure time, and model system. These findings raise concerns regarding long-term consumption and highlight the need for harmonised methodologies and regulatory reassessment.
Keywords:
carrageenan
; carboxymethyl cellulose
; gut microbiota
; inflammation
1. Introduction
The human gut microbiota is a complex ecosystem of microorganisms colonizing the digestive tract and plays a crucial role in digestion, metabolism, immune function, and disease prevention [1]. Firmicutes and Bacteroidota dominate the gut microbiome, with the genera Clostridium and Bacteroides as key representatives [2]. Maintaining a balanced microbial composition is essential for health, as dysbiosis—an imbalance in gut microbiota—has been linked to various conditions, including obesity, colorectal cancer, and type II diabetes, among other chronic diseases [3,4]. Multiple external factors influence the gut microbiota, including diet, lifestyle, and medications [5]. One growing concern is the effect of food additives—substances that enhance visual aspect, flavour, taste, texture, and shelf-life— on microbial composition [5]. While regulatory bodies consider many additives safe, emerging research suggests that some may disrupt gut homeostasis, potentially contributing to inflammation and disease [4,5]. Despite increasing interest in the gut microbiome, the impact of food additives remains understudied compared to macronutrients and other dietary components.
This review examines current evidence on the interactions between food additives and the gut microbiota, highlighting their potential role in disease progression. By summarising existing literature and identifying research gaps, this work aims to provide insights into the broader implications of food additives on human health.
2. The Human Gut Microbiota Composition and Interaction
The gut microbiota is a thriving ecosystem of microorganisms that performs intricate processes that profoundly influence our health and well-being, changing with the host. Therefore, the gut microbiota evolves into a diverse community of microorganisms. According to numerous studies, the human gut microbiota contains more than 1000 microbial species [1,5]. The composition of an individual’s gut microbiota can vary significantly from person to person, influenced by factors such as genetics, diet, age, and environment; it also differs along the gastrointestinal tract, leading to the assembly of specific communities [6]. The main phyla in the gut microbiota are Bacteroidota, Firmicutes, Fusobacteria, Proteobacteria, Cyanobacteria, Verrucomicrobia, and Actinobacteria [6]. The most common species of these phyla are presented in the figure below (Figure 1).
The Human Microbiome Project Consortium identified pathogenic species in healthy individuals using shotgun metagenomic data and unique marker sequences. This analysis, which encompasses the largest cohort and most diverse collection of clinically relevant body habitats to date, offers significant insights into the complexity of human-associated microbial communities [8,12]. Although several pathogenic species (Figure 2) may also be present in healthy individuals, their presence alone does not necessarily indicate a pathological condition. Instead, evaluating shifts in their relative abundance is essential when interpreting gut microbiota alterations associated with serious diseases. Both increases and decreases in these taxa may reflect significant changes in microbial community structure and function. Therefore, examining their relative proportions, ecological interactions, and collective influence on the microbial ecosystem is crucial for accurately characterizing gut microbiome dynamics. The human gut microbiota is largely dominated by the phyla Firmicutes and Bacteroidota. The phylum Firmicutes includes more than 200 genera, such as Lactobacillus, Bacillus, Clostridium, Enterococcus, and Ruminococcus, with many members belonging to the class Clostridia, which constitutes a substantial proportion of this phylum. In contrast, the phylum Bacteroidota is mainly represented by the dominant genera Prevotella and Bacteroides [8,12]. Furthermore, according to Mariat et al. (2009), the dominant genera characterising the adult faecal microbiota include Clostridium cluster IV (e.g., Faecalibacterium), Clostridium cluster XIVa (e.g., Roseburia, Eubacterium), Bacteroides, and Bifidobacterium. In contrast, genera such as Lactobacillus, Escherichia/Shigella (representing Enterobacteriaceae), Desulfovibrio, Sporomusa, Atopobium, and other less abundant members of Clostridium clusters XI, XIVb, and XVIII constitute subordinate components of the microbiota [8].
The abundance of Firmicutes in the gut microbiota of healthy individuals ranges from 11% to 95%, while that of Bacteroidota varies between 0.6% to 86.6%. Thus, the relative abundance of these phyla is highly variable even among subjects within the same population [2]. This variability is expected, as the gut microbiota is influenced by factors such as diet, physical activity, food additives and contaminants, and antibiotic use [9]. Bacteroidota play a crucial role in protecting against pathogens and supplying nutrients to other microbial residents, thereby contributing to the stability of the immune system. They are primarily responsible for producing short-chain fatty acids (SCFAs), mainly acetate and propionate, which serve as potent anti-inflammatory mediators by reducing the release of pro-inflammatory cytokines from neutrophils and macrophages [9].
Moreover, Bacteroidota can thrive over a wide pH range and modify the nutritional landscape by synthesising compounds such as fucosylated glycoproteins or by liberating fucose and sialic acid residues from glycoproteins, which can then be utilised by other microorganisms, including pathogens [11]. The Firmicutes phylum tends to be stable under neutral or slightly acidic conditions, adapting to the specific gastrointestinal tract regions in which it resides. Firmicutes are fundamental to various metabolic pathways in the gut, encompassing the metabolism of dietary nutrients, bile acids, and host-derived compounds. Their ability to metabolise a wide range of substrates significantly influences nutrient absorption, energy balance, and the production of bioactive molecules [13]. It has also been observed that the balance between these two phyla has been associated with the maintenance of intestinal homeostasis, with imbalances being linked to several pathologies, including obesity, colorectal cancer, and type II diabetes [12].
Such dysbiosis is typically characterized by shifts in the abundance of specific microbial taxa. For example, increases in genera such as Enterococcus, Fusobacterium, Parvimonas, Gemella, and Leptotrichia, as well as members of the family Enterobacteriaceae have frequently been reported in disease states. In contrast, beneficial genera such as Lachnospira, Clostridium, Bifidobacterium, Faecalibacterium, Blautia and Roseburia are often reduced. Alterations in the abundance of these taxa, either through overgrowth or depletion, can disrupt microbial community stability and contribute to pathologies associated with gut dysbiosis (Figure 3).
3. Gut Microbiota and Its Impacts on Human Health
According to the literature, the gut microbiota contributes significantly to human physiology through its metabolic activities [6,14]. These microorganisms decompose complex carbohydrates and proteins by producing a variety of enzymes essential for nutrient metabolism [6]. In addition to its metabolic functions, the gut microbiota plays a pivotal role in modulating the human immune system. It regulates nutrient absorption and host metabolism and influences gene expression and cellular physiology, thereby modulating immune responses [15].
This intricate connection supports innate and adaptive immune functions and is closely linked to overall human health. Moreover, its involvement in the digestion and absorption of nutrients via fermentation and colonic decomposition has been well documented [7]. For instance, the relative abundance of Ruminococcus 2, Faecalibacterium, and Akkermansia is associated with increased neutrophil rates, and the total abundance of Faecalibacterium correlates with neutrophil dynamics, underscoring the critical role of the gut microbiota in immune function [7]. Furthermore, dysbiosis of the gut microbiota is implicated in various diseases, including inflammatory bowel disease (IBD), irritable bowel syndrome (IBS), diabetes, obesity, cancer, cardiovascular problems, and central nervous system disorders. Therefore, understanding the mechanisms by which the microbiome contributes to these conditions is paramount.
Figure 4.
Schematic representation of the shift from healthy microbiota to dysbiosis, contributing to chronic inflammation and the development of gastrointestinal and metabolic diseases. Created with Biorender.
Figure 4.
Schematic representation of the shift from healthy microbiota to dysbiosis, contributing to chronic inflammation and the development of gastrointestinal and metabolic diseases. Created with Biorender.

i. Inflammatory Bowel Disease and Irritable Bowel Syndrome
IBD and IBS are chronic gastrointestinal disorders that affect the gastrointestinal tract (GIT). In the case of IBD, the main forms include Crohn’s disease and ulcerative colitis. Individuals with IBD often exhibit dysbiosis, characterized by alterations in the abundance and diversity of specific microbial taxa in the gut [16]. IBS is a common GIT disorder that affects the large intestine (colon), resulting in abnormal intestinal motility and low-grade inflammation. Several studies have linked these syndromes to gut microbiota dysbiosis, which may contribute to the intestinal inflammation observed particularly in IBD and, to a lesser extent, in IBS [16,20].
Therefore, it is essential to understand which alterations in the microbiota are associated with the development of these conditions. In both diseases, the gut microbiota is primarily composed of the phyla Firmicutes and Bacteroidota. In IBS specifically, an increased abundance of genera such as Enterococcus and members of the order Bacillales has been observed, along with an overall increase in Bacteroidota and a relative reduction in Firmicutes. This reduction notably includes Faecalibacterium prausnitzii, a butyrate-producing bacterium known for its anti-inflammatory properties. Consequently, this decrease may lead to reduced concentrations of SCFA [16,20]. Nevertheless, in IBD patients the impact of dysbiosis often depends on the location within the GIT. Microbial communities typically show higher phylogenetic diversity in the large intestine compared with the small intestine. In the small intestine, there is a higher abundance of Lactobacillus, while members of the family Lachnospiraceae are present at lower levels compared with the colon.
Throughout the GIT, patients with IBD exhibit decreased levels of species such as Blautia faecis, Roseburia inulinivorans, Ruminococcus torques, and Clostridium lavalense. Conversely, an increased abundance of Desulfovibrio, a genus of sulfate-reducing bacteria, has been observed. These bacteria can damage intestinal epithelial cells and promote mucosal inflammation. Additionally, a subtle increase in Proteobacteria—particularly Escherichia coli strains with adherent-invasive characteristics (AIEC)—has been associated with enhanced intestinal mucosal pro-inflammatory responses [16,19,21]. A significant decrease in the abundance of Faecalibacterium and Bifidobacterium has been observed exclusively in IBS, with the gut microbiota predominantly dominated by Enterobacteriaceae. This shift is associated with a higher prevalence of pro-inflammatory phylotypes, including Clostridium cocleatum (88%), Clostridium thermosuccinogenes (85%), Coprobacillus catenaformis (91%), Ruminococcus bromii-like (91%), and Ruminococcus torques (93%) [17,20].
ii. Colorectal Cancer
Colorectal cancer (CRC) occurs when cells grow in the large intestine, making it one of the most common cancers worldwide [22,23]. There is reason to believe that gut dysbiosis is related to the progression of cancer in light of several studies on CRC being associated with the pathogenetic bacterium, various being identified as carcinogens [24]. According to tissue and stool assays, it is possible to identify those genera, such as, Fusobacterium, Parvimonas, Gemella, Leptotrichia, Enterococcus, Escherichia/ Shigella, Klebsiella, Streptococcus, and Peptostreptococcus are enriched, and anti-inflammatory F. prausnitzii, Bifidobacterium, Faecalibacterium, and Blautia have a lower abundance in CRC patients [21,24]. Furthermore, enterotoxigenic Bacteroides fragilis (ETBF), which encodes B. fragilis metalloprotease toxin (BFT) to induce diarrhoea in most reports, and Fusobacterium nucletum, which has proven to be invasive, are highly expressed in CRC tissue when compared with healthy tissue [24].
On a separate issue, CRC patients also exhibit significantly lower SCFA abundance and SCFA-producing bacteria, which may be driven by a reduction in Firmicutes and an increase in Bacteroidota, leading to the development and progression of CRC [25].
iii. Diabetes Type 2
Research suggests that the composition and function of the gut microbiota may contribute to the development and progression of Type 2 diabetes (T2DM). While healthy individuals have a high abundance of butyrate-producing bacteria, patients with T2DM exhibit reductions primarily in Clostridium coccoides, Clostridium leptum, Akkermansia muciniphila, and Faecalibacterium prausnitzii. Conversely, they show increased levels of several pathogenic bacteria, including Clostridium hathewayi, Clostridium symbiosum, Escherichia coli, Bacteroides vulgatus, Veillonella denticariosi, and Lactobacillus. Furthermore, the genera Bifidobacterium, Bacteroides, Faecalibacterium, Akkermansia, and Roseburia are negatively associated with T2DM, whereas the genera Ruminococcus, Fusobacterium, and Blautia are positively associated with the disease [26,27,28]. Moreover, when analysing individuals by glucose tolerance status, T2DM patients have a higher abundance of Lactobacillus species and a lower abundance of Clostridium species than individuals with normal glucose tolerance. While Lactobacillus has been positively associated with fasting glucose, Clostridium species have been negatively correlated with it, suggesting that these same bacterial taxa are associated with the development of T2DM [27,28].
iv. Obesity
According to several studies, obesity can be influenced by the imbalance of the Firmicutes/ Bacteroidota ratio at the phylum level, showing a low amount of Firmicutes, such as Blautia hydrogenotrophica, Coprococcus catus, Eubacterium ventriosum, Ruminococcus bromii, and Ruminococcus obeum, with a prevalence of Prevotella in obese groups [20,29]. Moreover, since Methanobacteriales, Lactobacillus, Bifidobacteria genera, and Christensenellaceae and Akkermansia families are used as probiotics to help this condition, it is essential to analyse their reduction, as it can be associated with obesity advance [19].
4. Food Additives Related to Gut Health Issues
Nowadays, our products are often far from their natural state, mainly presented in altered forms. This is due to the pressure to ensure specific quality, convenience and shelf-life parameters that must be maintained to increase food products’ time-to-market. Food additives are crucial players, as they are intentionally incorporated into products to achieve specific goals, such as preserving food, enhancing colour, flavour and improving texture, or extending shelf-life. According to the Codex Alimentarius, an additive is defined as any substance that would not be usually used as an ingredient but is applied in food for manufacture, processing, preparation, treatment, packing, packaging, transport, or holding purposes, which may or may not maintain or improve its nutritional qualities [30]. Since they have become normalised in human consumption, the potential health issues they may cause have been studied. Some of them were blocked from use in food products due to their hazardous effects, which have the potential to cause gut dysbiosis. Considering this, research on this topic has been conducted to understand the impact of additives (still circulating) on our health [5].
Many food additives could have effects like those discussed, but the additives listed below (Table 1) are currently under research by the European Food Safety Authority [5]. Among these additives, Carrageenan (E407) and Carboxymethyl cellulose (E466) have been shown to pose potential risks to the gut microbiota, and it is essential to deepen knowledge of their possible toxicity and to understand how they may affect the human gut microbiota, potentially leading to more severe conditions due to dysbiosis.
5. Gut Modulation by Carrageenan and Carboxymethyl Cellulose: Insights and Implications
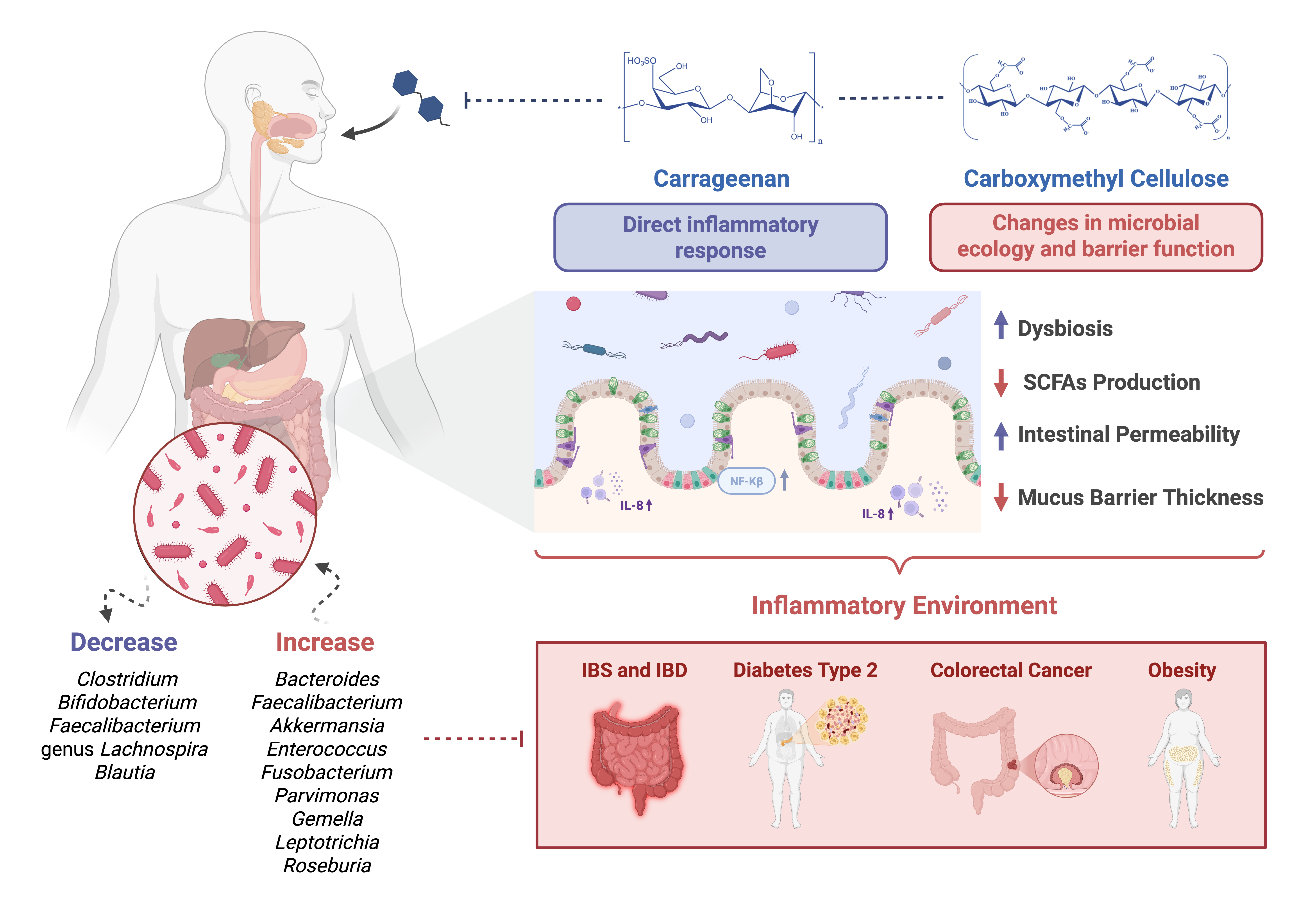
The literature reviewed highlights the distinct effects of carrageenan (CGN) and carboxymethyl cellulose (CMC) on gut health. Studies indicate that both additives can alter gut microbiota composition, disrupt the intestinal barrier, and promote inflammatory responses. However, the specific mechanisms and severity of these effects vary depending on factors such as dosage, exposure duration, and host susceptibility. The following sections summarise key findings on alterations in the microbiota, gut permeability, and immune responses associated with CGN and CMC consumption.
i. Carrageenan
CGN is a sulphated polysaccharide widely used in the food industry for its gelling, thickening and stabilising properties. It is an emulsifier derived from red seaweed, commonly used in beverages to stabilise emulsions [30]. Due to its extensive use, increasing evidence has raised concerns regarding its impact on intestinal health, particularly through interactions with the gut microbiota and the induction of inflammatory responses [75]. Importantly, these effects appear to be largely dose-dependent, reflecting exposure levels rather than the compound’s intrinsic toxicity [30]. Several studies indicate that food emulsifiers, including CGN, may reduce mucus barrier thickness and increase intestinal permeability, thereby promoting gastrointestinal tract inflammation [7]. Such promotion of pro-inflammatory signalling pathways, notably through the activation of NF-κB and increased production of inflammatory mediators such as IL-8, is supposedly due to interactions in gut microbial composition and metabolism, rather than a direct interaction with epithelial cells [76].
Research on CGN primarily focuses on its two main types: iota-(i-)CGN and kappa-(k-)CGN, which differ in their gelling properties. While i-CGN forms soft, heat-reversible gels, k-CGN forms strong, heat-stable gels. Due to safety concerns, multiple studies have investigated the potential adverse effects of CGN on gut health (Table 2). In in vivo studies, the duration of experiments varies significantly, ranging from 90 days [32] to lifelong administration. CGN concentrations applied in experiments range from 0.1% to 25% (w/v). High concentrations have been associated with severe outcomes such as hepatic cirrhosis and, in extreme cases, death [31]. Studies using concentrations between 0.1% and 5.0% have reported ulcerative lesions, diarrhoea, and bowel damage [35,36,37]. While in vitro research on CGN is limited, existing studies display a wide range of concentrations and testing durations. The longest trials in cell models lasted 96 hours [38], with CGN administration at 1.0% (w/v) and the lowest dose at 0.1% (w/v). Most studies report CGN-induced inflammation [39]. Animal studies provide the most robust evidence regarding the adverse intestinal effects of carrageenan exposure. Using data collected and shown in Table 2, the consumption report heterogeneous outcomes that depend on dose, CGN type, species, and exposure duration.
Adverse effects were mainly observed at very high dietary concentrations (> 15-25%), including mortality in mice and hepatic alterations in rats, conditions that are considered non-physiological and not representative of realistic dietary exposure [35]. At moderate levels (< 5%), the majority of subchronic and chronic studies in rodents, pigs, and non-human primates reported no significant effects on growth, behaviour, organ weights, or intestinal morphology. However, mild gastrointestinal effects, such as soft stools or transient diarrhoea, were occasionally observed [39,43,44].
Studies using mg/kg body weight doses, closer to estimated human intake levels, generally showed no compound-related toxicity. However, some evidence suggests that λ-CGN may alter gut microbiota composition and metabolic activity without overt clinical disease [31,48,52]. Overall, the animal studies evidence indicates that potential adverse effects of CGN are dose-dependent and species-specific, with limited toxicological relevance at exposure levels.
Human evidence on CGN indicates limited but relevant findings. Clinical data suggest that consumption of food-grade CGN may aggravate disease activity and shorten relapse time in patients with ulcerative colitis in remission, indicating a potential risk for susceptible populations [53,54,55].
In contrast, in vitro studies using human intestinal cell lines generally show no cytotoxicity, oxidative stress, or intestinal permeability, although CGN has been shown to activate pro-inflammatory signalling pathways (TLR-4) [56]. Overall, human data does not demonstrate overt toxicity in healthy models but suggests a possible pro-inflammatory effect under specific pathological conditions, highlighting the need for further controlled studies in humans.
Across various experimental conditions, most studies indicated that CGN fosters an inflammatory environment [38,40,41]. However, some studies have found no significant effects [43,44,45]. As summarised in Table 2, these results underscore the need for personalised guidance on CGN intake and further investigation into its gastrointestinal health and effects.
ii. Carboxymethyl Cellulose
CMC is a water-soluble polysaccharide derived from cellulose, widely used as a food emulsifier for improving texture and stability [56]. In recent years, increasing attention has been given to its potential effects on gastrointestinal health, particularly on intestinal inflammation and alterations in the gut microbiota (Table 3).
Compared with other food additives, toxicological and physiological data on CMC are relatively limited, and the available studies exhibit substantial heterogeneity in experimental design. Reported studies on CMC remain limited, experimental concentrations range from 0.1% to 10.0% [57,58].
Most in vivo studies in mice indicate that dietary exposure to 1.0% CMC induces chronic low-grade inflammation and colitis, accompanied by alterations in microbiota localisation, composition, and pro-inflammatory potential [63,64,65].
These findings suggest that CMC may disrupt host-microbiota interactions, contributing to intestinal inflammatory responses. However, not all studies have reported adverse outcomes; safety trials using higher concentrations (2.5-10.0%) did not observe statistically significant treatment-related effects on evaluated physiological parameters, highlighting inconsistencies across experimental models and endpoints [59,60,61,66].
Findings from in vitro and ex vivo models further support a potential microbiota-mediated mechanism. Studies using human intestinal cell lines (HT29-MTX and HEpG2) reported a strong pro-inflammatory profile following CMC exposure [69,70]. At the same time, microbiota-focused systems demonstrated persistent alterations in microbiota composition and increased pro-inflammatory potential at concentrations as low as 0.1% [68,69].
iii. Comparative Perspective and Critical Assessment
The continuous consumption of food additives such as CGN and CMC, particularly amid rising processed food intake, has been linked to multiple gut health issues. Both exhibit distinct but overlapping mechanisms of interaction with gut microbiota and host intestinal physiology.
CGN shows a more pronounced association with direct inflammatory responses, whereas CMC predominantly affects microbial ecology and intestinal barrier function. Nevertheless, these effects create an environment that promotes inflammation, disrupts lipid metabolism and alters gastrointestinal health.
The major limitation across studies is the lack of harmonised experimental designs, particularly regarding dosage, exposure, duration, and microbial assessment methods. Additionally, most available data derive from simplified models that may not fully capture the complexity of human dietary patterns and gut ecosystems.
A key aspect of gut modulation and potential dysbiosis associated with CGN and CMC exposure is the alteration of microbial populations, particularly shifts within the Bacteroidota and Firmicutes phyla. Reported changes included increased abundance of genera such as Bacteroides, Faecalibacterium, Akkermansia, Enterococcus, Fusobacterium, Parvimonas, Gemella, Leptotrichia, and Roseburia, as well as representatives of the genera Enterobacter (within the Enterobacteriaceae family) and Bacillus (from the Bacillales order). Conversely, decreases have been noted in genera such as Clostridium, Bifidobacterium, Faecalibacterium, Blautia, and members of the genus Lachnospira (from the Lachnospiraceae family) [69,70], which are clustered with intestinal homeostasis.
Specific bacterial species implicated in CGN- and CMC-associated dysbiosis, which may favour inflammatory processes, are summarised in Table 4. These taxa warrant prioritised attention in future mechanistic and human intervention studies, as they represent key microbial markers linking food additive exposure to inflammation-derived gut dysfunction.
These species require prioritised attention when studying the effects of additives on the gut microbiota, as they can create an environment that supports inflammation and are directly associated with the consumption of these additives. Overall, the converging evidence highlights the need for standardised, microbiota-centred risk assessment frameworks and underscores the importance of evaluating food additives not only in isolation but also within the context of realistic dietary patterns and long-term consumption.
6. Hands-On Exploration of Carrageenan and Carboxymethyl Cellulose Effects In Vitro
As an initial screening step for evaluating the effects of CNG and CMC, in vitro simulation models are highly suitable and widely adopted. These models offer several advantages, including reduced labour and time requirements, diminished reliance on in vivo experimentation, the ability to simulate controlled gastrointestinal conditions, and fewer ethical concerns than animal or human studies.
In vitro systems enable the assessment of digestibility, fermentability, and microbiota modulation, providing insight into whether CGN and CMC promote beneficial microbial activity or contribute to dysbiosis. These models facilitate the assessment of a product’s nutritional value, the digestion process, and their effects on microbial populations. Typically, this approach encompasses preliminary fermentability assessments, followed by simulated gastrointestinal digestion and colonic fermentation, allowing the profiling of bacterial populations and their metabolic outputs [71].
i. Fermentation Assay
Fermentability assays are essential for evaluating whether CGN and CMC can be metabolised by gut microbiota and how they influence microbial growth and activity. Key parameters assessed include bacterial proliferation, organic acid production, pH variation, and ammonia formation. These indicators provide valuable information on microbial metabolic responses and ecosystem stability [71]. Although there have been studies on the impact these additives in vivo, and in vitro models, there is a gap of knowledge on their impact on prolonged intake.
This can be achieved through either batch (Fed-batch) or continuous fermentation systems. Continuous systems allow long-term, steady-state operation with controlled microbial growth. In contrast, fed-batch systems enable modulation of substrate availability and reduce risks of substrate inhibition or excessive metabolite accumulation. Nevertheless, fed-batch systems are widely applied in studies investigating gut microbiota fermentation. In this approach, fresh substrate or test compounds are periodically introduced into the system while fermentation proceeds continuously. This design allows for repeated sampling through the collection of aliquots at defined intervals, enabling the monitoring of microbial activity and metabolic changes over time. Consequently, fed-batch fermentation facilitates the evaluation of the temporal effects of the tested sample on gut microbiota composition and metabolic outputs across an extended experimental period [71,72,73].
ii. Sample Selection
Human faecal samples are commonly used as proxies for gut microbiota due to their convenience, non-invasiveness, feasibility, and rich microbial communities. This approach offers a pragmatic alternative to delving into the microbial ecosystems of the gastrointestinal tract, sidestepping more intrusive methods. Even though they cannot fully represent the gut microbiota, experts believe faeces are more representative of luminal microorganisms than the mucosa-associated gut microbiota. Stool samples still provide a good representation of bacterial communities, as 55-60% of them are bacteria [64,65]. To study the dysbiosis caused by CGN and CMC consumption, faecal samples are typically collected from healthy donors, enabling the identification of additive-induced alterations in microbial balance and metabolic activity under controlled conditions.
iii. Additive Supplementation
Previous fermentation studies were conducted over a 48-h fermentation period. To investigate the effects of prolonged exposure to these additives, it would be valuable to collect samples and renew the nutritional medium at this time point, thereby enabling an extension of the fermentation period [71,73]. This approach could provide an innovative strategy to address existing gaps in knowledge regarding the effects of additives on the microbiome.
iv. Metabolite Analysis
Metabolite profiling is a critical component of in vitro fermentation studies, as it reflects functional changes in microbial activity [71,73]. One of the primary roles of the human gut microbiota is to metabolise undigested carbohydrates, proteins, and peptides into essential SCFAs, including acetate, propionate, and butyrate, as well as intermediate metabolites, such as succinate, lactate, and formate, which are branched-chain fatty acids (BCFAs) obtained from proteolytic fermentation.
v. Human GUT Simulation
Advanced in vitro gastrointestinal models, using previously the INFOGEST protocol, replicate key physicochemical and physiological conditions of the human GIT. These models integrate the study of gut digestion and absorption processes with the assessment of nutrient bioavailability and bioaccessibility [73]. The process can be broken down into three sequential steps: (1) simulating gastrointestinal digestion, (2) mimicking intestinal absorption, and (3) performing colonic fermentation studies. The application of INFOGEST, with further in vitro gastrointestinal models, can provide data on bacterial quantification, organic acid production, and NH4+ measurement [71].
Application of such dynamic systems allows simultaneous evaluation of bacterial composition, metabolite production, and ammonia levels, making them particularly relevant for studying chronic exposure scenarios and additive–microbiota interactions. Nevertheless, despite their increased physiological relevance, these models remain simplifications of the human gut and should be interpreted in conjunction with in vivo evidence.
7. Conclusions and Future Perspectives
This review highlights the growing body of evidence indicating that widely used food additives, such as CGN and CMC, can influence gut microbiota composition, intestinal barrier integrity, and inflammatory responses. Although these additives are generally regarded as safe based on traditional toxicological endpoints, emerging data suggest that their chronic consumption, particularly within diets rich in ultra-processed foods, may have unintended consequences for gut health.
CGN and CMC exhibit distinct yet converging mechanisms of action. CGN has been more consistently associated with direct pro-inflammatory effects, including activation of innate immune signalling pathways and epithelial stress responses. In contrast, CMC primarily affects microbial spatial organisation and mucus barrier integrity, indirectly promoting dysbiosis and low-grade inflammation. Despite these mechanistic differences, both additives contribute to an intestinal environment that favours microbial imbalance, barrier dysfunction, and inflammatory susceptibility. The mechanism behind its inflammatory potential is still under investigation, but current evidence suggests that CMC alters gut microbial diversity and promotes colitis-like symptoms in experimental models. Despite growing concerns regarding the health implications of CGN and CMC, gaps remain in our understanding of their long-term effects, particularly in human populations. Further studies are needed to clarify safe consumption thresholds and to assess potential cumulative effects from prolonged exposure.
A critical limitation across the current literature is the predominance of in vitro and animal studies, alongside significant heterogeneity in experimental design, including differences in the type of additive, molecular characteristics, dosage, and exposure duration. Human data remains scarce and highly variable, underscoring the need for well-designed, long-term intervention studies that account for individual variability in the gut microbiota composition and its susceptibility.
The observed shifts in microbiota composition suggest that CGN and CMC consumption may contribute to gut dysbiosis by fostering an inflammatory environment. The dominance of Bacteroidota and Firmicutes, along with the reduction of beneficial bacteria such as Bifidobacterium and Faecalibacterium, is typical in conditions associated with gut microbiota imbalance. These changes are particularly concerning, given the role of these bacterial communities in maintaining intestinal homeostasis, modulating immune responses, and producing SCFAs that support gut health. Understanding these mechanisms is crucial for assessing the long-term implications of emulsifier consumption on human health.
From a regulatory and industrial perspective, these findings emphasise the need to re-evaluate current safety frameworks, incorporating gut microbiota modulation, chronic exposure, and realistic dietary patterns as key assessment parameters. Integrating advanced in vitro gut models with human microbiome data may offer a valuable pathway toward more predictive and physiologically relevant safety evaluations.
In conclusion, while CGN and CMC remain technologically valuable additives, their widespread and long-term consumption warrants greater scientific scrutiny. Future research should prioritise harmonised methodologies, mechanistic clarity, and human-relevant models to define safe use conditions better and support evidence-based dietary guidance in the context of modern food systems.
Author Contributions
“Conceptualisation, D.A.C. and M.P.; methodology, E.R.C.; formal analysis, A.L.P.F.; investigation, A.L.P.F.; resources, M.P.; data curation, D.A.C; E.R.C.; writing—original draft preparation, A.L.P.F.; writing—review and editing A.L.P.F.; D.A.C.; E.R.C.; M.P, All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by National Funds from FCT - Fundação para a Ciência e a Tecnologia through projects UID/50016/2025 and LA/P/0076/2020 (https://doi.org/10.54499/LA/P/0076/2020). Ezequiel R. Coscueta had the support from the FCT through an Assistant Researcher contract (DOI: 10.54499/2023.08679.CEECIND/CP2855/CT0005). Débora A. Campos had the support from the FCT through and Junior Researcher contract with the reference: 2020.02639.CEECIND.
Data Availability Statement
All the data is available in the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| CGN | Carrageenan |
| CMC | Carboxymethyl Cellulose |
| SCFA | Short Chain Fatty Acid |
References
- Adak, A.; Khan, M.R. An Insight into Gut Microbiota and Its Functionalities. Cellular and Molecular Life Sciences 2019, 76, 473–493.
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The Firmicutes/Bacteroidetes Ratio: A Relevant Marker of Gut Dysbiosis in Obese Patients? Nutrients 2020, 12.
- Meeting and World Health Organization Meeting and World Health Organization . 2007.
- Gomez-Arango, L.F.; Barrett, H.L.; Wilkinson, S.A.; Callaway, L.K.; McIntyre, H.D.; Morrison, M.; Dekker Nitert, M. Low Dietary Fiber Intake Increases Collinsella Abundance in the Gut Microbiota of Overweight and Obese Pregnant Women. Gut Microbes 2018, 9, 189–201. [CrossRef]
- Abiega-Franyutti, P.; Freyre-Fonseca, V. Chronic Consumption of Food-Additives Lead to Changes via Microbiota Gut-Brain Axis. Toxicology 2021, 464.
- Zhou, X.; Qiao, K.; Wu, H.; Zhang, Y. The Impact of Food Additives on the Abundance and Composition of Gut Microbiota. Molecules 2023, 28.
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What Is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7. [CrossRef]
- Mariat, D.; Firmesse, O.; Levenez, F.; Guimarǎes, V.D.; Sokol, H.; Doré, J.; Corthier, G.; Furet, J.P. The Firmicutes/Bacteroidetes Ratio of the Human Microbiota Changes with Age. BMC Microbiol. 2009, 9. [CrossRef]
- Zafar, H.; Saier, M.H. Gut Bacteroides Species in Health and Disease. Gut Microbes 2021, 13. [CrossRef]
- Wexler, A.G.; Goodman, A.L. An Insider’s Perspective: Bacteroides as a Window into the Microbiome. Nat. Microbiol. 2017, 2.
- Stojanov, S.; Berlec, A.; Štrukelj, B. The Influence of Probiotics on the Firmicutes/Bacteroidetes Ratio in the Treatment of Obesity and Inflammatory Bowel Disease. Microorganisms 2020, 8, 1715. [CrossRef]
- Bä ckhed, F.; Ding, H.; Wang, T.; Hooper, L. V; Young Koh, G.; Nagy, A.; Semenkovich, C.F.; Gordon, J.I. The Gut Microbiota as an Environmental Factor That Regulates Fat Storage; 2004;
- Cao, Y.; Liu, H.; Qin, N.; Ren, X.; Zhu, B.; Xia, X. Impact of Food Additives on the Composition and Function of Gut Microbiota: A Review. Trends Food Sci. Technol. 2020, 99, 295–310.
- Curtis, H. et al. Structure, Function and Diversity of the Healthy Human Microbiome. Nature 2012, 486, 207–214. [CrossRef]
- Belkaid, Y.; Hand, T.W. Role of the Microbiota in Immunity and Inflammation. Cell 2014, 157, 121–141.
- Frank, D.N., S.A.A.L., F.R.A., B.E.C., H.N., & P.N.R. Molecular-Phylogenetic Characterization of Microbial Community Imbalances in Human Inflammatory Bowel Diseases. 2007.
- Jacob, A., M.J. and 28. N.S.H. Frontmatter. In The Gut Microbiota in Health and Disease; Wiley, 2023.
- Quaglio, A.E.V.; Grillo, T.G.; Oliveira, E.C.S. De; Stasi, L.C. Di; Sassaki, L.Y. Gut Microbiota, Inflammatory Bowel Disease and Colorectal Cancer. World J. Gastroenterol. 2022, 28, 4053–4060. [CrossRef]
- Krogius-Kurikka, L.; Lyra, A.; Malinen, E.; Aarnikunnas, J.; Tuimala, J.; Paulin, L.; Mäkivuokko, H.; Kajander, K.; Palva, A. Microbial Community Analysis Reveals High Level Phylogenetic Alterations in the Overall Gastrointestinal Microbiota of Diarrhoea-Predominant Irritable Bowel Syndrome Sufferers. BMC Gastroenterol. 2009, 9, 95. [CrossRef]
- Center, M.M.; Jemal, A.; Ward, E. International Trends in Colorectal Cancer Incidence Rates. Cancer Epidemiology, Biomarkers & Prevention 2009, 18, 1688–1694. [CrossRef]
- Karsa, L. v.; Lignini, T.A.; Patnick, J.; Lambert, R.; Sauvaget, C. The Dimensions of the CRC Problem. Best Pract. Res. Clin. Gastroenterol. 2010, 24, 381–396. [CrossRef]
- Gao, R.; Gao, Z.; Huang, L.; Qin, H. Gut Microbiota and Colorectal Cancer. European Journal of Clinical Microbiology & Infectious Diseases 2017, 36, 757–769. [CrossRef]
- Hou, H.; Chen, D.; Zhang, K.; Zhang, W.; Liu, T.; Wang, S.; Dai, X.; Wang, B.; Zhong, W.; Cao, H. Gut Microbiota-Derived Short-Chain Fatty Acids and Colorectal Cancer: Ready for Clinical Translation? Cancer Lett. 2022, 526, 225–235. [CrossRef]
- Wu, W.; Zhou, J.; Xuan, R.; Chen, J.; Han, H.; Liu, J.; Niu, T.; Chen, H.; Wang, F. Dietary κ-Carrageenan Facilitates Gut Microbiota-Mediated Intestinal Inflammation. Carbohydr. Polym. 2022, 277, 118830. [CrossRef]
- Gurung, M.; Li, Z.; You, H.; Rodrigues, R.; Jump, D.B.; Morgun, A.; Shulzhenko, N. Role of Gut Microbiota in Type 2 Diabetes Pathophysiology. EBioMedicine 2020, 51, 102590. [CrossRef]
- Zhou, Z.; Sun, B.; Yu, D.; Zhu, C. Gut Microbiota: An Important Player in Type 2 Diabetes Mellitus. Front. Cell. Infect. Microbiol. 2022, 12. [CrossRef]
- Castaner, O.; Goday, A.; Park, Y.-M.; Lee, S.-H.; Magkos, F.; Shiow, S.-A.T.E.; Schröder, H. The Gut Microbiome Profile in Obesity: A Systematic Review. Int. J. Endocrinol. 2018, 2018, 1–9. [CrossRef]
- Carocho, M.; Barreiro, M.F.; Morales, P.; Ferreira, I.C.F.R. Adding Molecules to Food, Pros and Cons: A Review on Synthetic and Natural Food Additives. Compr. Rev. Food Sci. Food Saf. 2014, 13, 377–399. [CrossRef]
- Necas, J., & B. Carrageenan: A Review. Vet. Med. (Praha). 2013.
- Collins, T.F.X.; Black, T.N.; Prew, J.H. Long-Term Effects of Calcium Carrageenan in Rats—I. Effects on Reproduction. Food Cosmet. Toxicol. 1977, 15, 533–538. [CrossRef]
- Grasso, P., G.S.D., B.K.R., & W.M.G. STUDIES ON DEGRADED CARRAGEENAN IN RATS AND GUINEA-PIGS. Pergamon Press 1975.
- Grasso, P., S.M., C.F.M.B., & G.S.D. Studies on Carrageenan and Large-Bowel Ulceration in Mammals. Pergamon Press 1973.
- Marcus, R. Carrageenan-Induced Ulceration of the Large Intestine in the Guinea Pig. In Gut 1971.
- Borthakur, A.; Bhattacharyya, S.; Anbazhagan, A.N.; Kumar, A.; Dudeja, P.K.; Tobacman, J.K. Prolongation of Carrageenan-Induced Inflammation in Human Colonic Epithelial Cells by Activation of an NFκB-BCL10 Loop. Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease 2012, 1822, 1300–1307. [CrossRef]
- McKim, J.M.; Wilga, P.C.; Pregenzer, J.F.; Blakemore, W.R. The Common Food Additive Carrageenan Is Not a Ligand for Toll-Like- Receptor 4 (TLR4) in an HEK293-TLR4 Reporter Cell-Line Model. Food and Chemical Toxicology 2015, 78, 153–158. [CrossRef]
- Benitz, K.-F., G.L., & C.F. Intestinal Effects of Carrageenans in the Rhesus Monkey. Pergamon Press 1973.
- Mcgill, H.C.; Mcmahan, C.A.; Wigodsky, H.S.; Sprinz, H. Carrageenan in Formula and Infant Baboon Development. Gastroenterology 1977, 73, 512–517. [CrossRef]
- Poulsen, E. Short-Term Peroral Toxicity of Undegraded Carrageenan in Pigs; Pergamon Press, 1973; Vol. 11;
- McGill, H.C.; McMahan, C.A.; Wigodsky, H.S.; Sprinz, H. Carrageenan in Formula and Infant Baboon Development. Gastroenterology 1977, 73, 512–517. [CrossRef]
- Bhattacharyya, S.; Shumard, T.; Xie, H.; Dodda, A.; Varady, K.A.; Feferman, L.; Halline, A.G.; Goldstein, J.L.; Hanauer, S.B.; Tobacman, J.K. A Randomized Trial of the Effects of the No-Carrageenan Diet on Ulcerative Colitis Disease Activity. Nutr. Healthy Aging 2017, 4, 181–192. [CrossRef]
- McKim, J.M. Food Additive Carrageenan: Part I: A Critical Review of Carrageenan in Vitro Studies, Potential Pitfalls, and Implications for Human Health and Safety. Crit. Rev. Toxicol. 2014, 44, 211–243. [CrossRef]
- Rhee, M.; Pittz, E.P.; Abraham, R. Effect of Combinations of Irideae Carrageenan and Cellulose on the Absorption of Some Nutrients from the Alimentary Tract of Rats. Ecotoxicol. Environ. Saf. 1981, 5, 1–14. [CrossRef]
- Tomarelli et al. Tomarelli1974. 1974.
- Grasso, P.; Sharratt, M.; Carijanini, F.M.B.; Gangolli, S.D. Studies on Carrageenan and Large-Bowel Ulceration in Mammals; Pergamon Press, 1973; Vol. 11;
- Marcus, R. Carrageenan-Induced Ulceration of the Large Intestine in the Guinea Pig; 1971; Vol. 12;
- Nilson et al. Nilson1959. 1959.
- Abraham, R.; Benitz, K.-F.; Mankes, R.; Rosenblum, I. Chronic and Subchronic Effects of Various Forms of Carrageenan in Rats. Ecotoxicol. Environ. Saf. 1985, 10, 173–183. [CrossRef]
- Benitz, K.-F.; Golberg, L.; Couuton, F. Intestinal Effects of Carrageenans in the Rhesus Monkey (Macaca Mulatta)*; Pergamon Press, 1973; Vol. 11;
- Necas, J.; Bartosikova, L. Carrageenan: A Review; Vol. 58;
- Martino, J.V.; Van Limbergen, J.; Cahill, L.E. The Role of Carrageenan and Carboxymethylcellulose in the Development of Intestinal Inflammation. Front. Pediatr. 2017, 5.
- Wu, W.; Zhou, J.; Xuan, R.; Chen, J.; Han, H.; Liu, J.; Niu, T.; Chen, H.; Wang, F. Dietary κ-Carrageenan Facilitates Gut Microbiota-Mediated Intestinal Inflammation. Carbohydr. Polym. 2022, 277. [CrossRef]
- Bhattacharyya, S.; Shumard, T.; Xie, H.; Dodda, A.; Varady, K.A.; Feferman, L.; Halline, A.G.; Goldstein, J.L.; Hanauer, S.B.; Tobacman, J.K. A Randomised Trial of the Effects of the No-Carrageenan Diet on Ulcerative Colitis Disease Activity. Nutr. Healthy Aging 2017, 4, 181–192. [CrossRef]
- McKim, J.M.; Wilga, P.C.; Pregenzer, J.F.; Blakemore, W.R. The Common Food Additive Carrageenan Is Not a Ligand for Toll-Like- Receptor 4 (TLR4) in an HEK293-TLR4 Reporter Cell-Line Model. Food and Chemical Toxicology 2015, 78, 153–158. [CrossRef]
- McKim, J.M. Food Additive Carrageenan: Part I: A Critical Review of Carrageenan in Vitro Studies, Potential Pitfalls, and Implications for Human Health and Safety. Crit. Rev. Toxicol. 2014, 44, 211–243. [CrossRef]
- Borthakur, A.; Bhattacharyya, S.; Anbazhagan, A.N.; Kumar, A.; Dudeja, P.K.; Tobacman, J.K. Prolongation of Carrageenan-Induced Inflammation in Human Colonic Epithelial Cells by Activation of an NFκB-BCL10 Loop. Biochim. Biophys. Acta Mol. Basis Dis. 2012, 1822, 1300–1307. [CrossRef]
- Costa, E.M.; Silva, S.; Pereira, C.F.; Ribeiro, A.B.; Casanova, F.; Freixo, R.; Pintado, M.; Ramos, Ó.L. Carboxymethyl Cellulose as a Food Emulsifier: Are Its Days Numbered? Polymers (Basel). 2023, 15, 2408. [CrossRef]
- Mondal, Md.I.H.; Yeasmin, Mst.S. Toxicity Study of Food-Grade Carboxymethyl Cellulose Synthesised from Maize Husk in Swiss Albino Mice. Int. J. Biol. Macromol. 2016, 92, 965–971. [CrossRef]
- Naimi, S.; Viennois, E.; Gewirtz, A.T.; Chassaing, B. Direct Impact of Commonly Used Dietary Emulsifiers on Human Gut Microbiota. Microbiome 2021, 9, 66. [CrossRef]
- Chassaing, B.; Van de Wiele, T.; De Bodt, J.; Marzorati, M.; Gewirtz, A.T. Dietary Emulsifiers Directly Alter Human Microbiota Composition and Gene Expression Ex Vivo Potentiating Intestinal Inflammation. Gut 2017, 66, 1414–1427. [CrossRef]
- Chassaing, B.; Koren, O.; Goodrich, J.K.; Poole, A.C.; Srinivasan, S.; Ley, R.E.; Gewirtz, A.T. Dietary Emulsifiers Impact the Mouse Gut Microbiota Promoting Colitis and Metabolic Syndrome. Nature 2015, 519, 92–96. [CrossRef]
- Zangara, M.T.; Ponti, A.K.; Miller, N.D.; Engelhart, M.J.; Ahern, P.P.; Sangwan, N.; McDonald, C. Maltodextrin Consumption Impairs the Intestinal Mucus Barrier and Accelerates Colitis Through Direct Actions on the Epithelium. Front. Immunol. 2022, 13. [CrossRef]
- Zangara, M.T.; Ponti, A.K.; Miller, N.D.; Engelhart, M.J.; Ahern, P.P.; Sangwan, N.; McDonald, C. Maltodextrin Consumption Impairs the Intestinal Mucus Barrier and Accelerates Colitis Through Direct Actions on the Epithelium. Front. Immunol. 2022, 13. [CrossRef]
- Chassaing, B.; Koren, O.; Goodrich, J.K.; Poole, A.C.; Srinivasan, S.; Ley, R.E.; Gewirtz, A.T. Dietary Emulsifiers Impact the Mouse Gut Microbiota Promoting Colitis and Metabolic Syndrome. Nature 2015, 519, 92–96. [CrossRef]
- Viennois, E.; Bretin, A.; Dubé, P.E.; Maue, A.C.; Dauriat, C.J.G.; Barnich, N.; Gewirtz, A.T.; Chassaing, B. Dietary Emulsifiers Directly Impact Adherent-Invasive E. Coli Gene Expression to Drive Chronic Intestinal Inflammation. Cell Rep. 2020, 33. [CrossRef]
- Mondal, M.I.H.; Yeasmin, M.S. Toxicity Study of Food-Grade Carboxymethyl Cellulose Synthesised from Maize Husk in Swiss Albino Mice. Int. J. Biol. Macromol. 2016, 92, 965–971. [CrossRef]
- Baran, A.; Sulukan, E.; Türkoğlu, M.; Ghosigharehagaji, A.; Yildirim, S.; Kankaynar, M.; Bolat, I.; Kaya, M.; Topal, A.; Ceyhun, S.B. Is Sodium Carboxymethyl Cellulose (CMC) Really Completely Innocent? It May Be Triggering Obesity. Int. J. Biol. Macromol. 2020, 163, 2465–2473. [CrossRef]
- Naimi, S.; Viennois, E.; Gewirtz, A.T.; Chassaing, B. Direct Impact of Commonly Used Dietary Emulsifiers on Human Gut Microbiota. Microbiome 2021, 9. [CrossRef]
- Chassaing, B.; Van De Wiele, T.; De Bodt, J.; Marzorati, M.; Gewirtz, A.T. Dietary Emulsifiers Directly Alter Human Microbiota Composition and Gene Expression Ex Vivo Potentiating Intestinal Inflammation. Gut 2017, 66, 1414–1427. [CrossRef]
- Cohen, S.M., & I.N. A Critical Review of the Toxicological Effects of Carrageenan and Processed Eucheuma Seaweed on the Gastrointestinal Tract. Toxicology 2002.
- Komisarska, P.; Pinyosinwat, A.; Saleem, M.; Szczuko, M. Carrageenan as a Potential Factor of Inflammatory Bowel Diseases. Nutrients 2024, 16, 1367. [CrossRef]
- de Carvalho, N.M.; Oliveira, D.L.; Saleh, M.A.D.; Pintado, M.; Madureira, A.R. Preservation of Human Gut Microbiota Inoculums for in Vitro Fermentations Studies. Fermentation 2021, 7. [CrossRef]
- Trilokesh, C.; Uppuluri, K.B. Biobutanol from Lignocellulosic Biomass and Microalgae: Scope, Technology, and Economics. In Sustainable Biofuels; Elsevier, 2021; pp. 163–223.
- de Carvalho, N.M.; Oliveira, D.L.; Costa, C.M.; Pintado, M.; Madureira, A.R. Can Supplemented Skim Milk (SKM) Boost Your Gut Health? Fermentation 2022, 8. [CrossRef]
- Louis, P.; Flint, H.J. Formation of Propionate and Butyrate by the Human Colonic Microbiota. Environ. Microbiol. 2017, 19, 29–41. [CrossRef]
- Jabeen, Faiza; et al. “Carrageenan: Structure, Properties and Applications with Special Emphasis on Food Science.” RSC Advances, vol. 15, no. 27, 1 Jan. 2025, pp. 22035–22062, pmc.ncbi.nlm.nih.gov/articles/PMC12203331/. [CrossRef]
- Komisarska, Paulina; et al. “Carrageenan as a Potential Factor of Inflammatory Bowel Diseases.” Nutrients, vol. 16, no. 9, 30 Apr. 2024, p. 1367, pubmed.ncbi.nlm.nih.gov/38732613/. Accessed 17 July 202. [CrossRef]
Figure 1.
Taxonomic tree diagram of the most prominent bacteria within the Bacteroidota and Firmicutes phyla on gut microbiota, adapted from [5,6].

Figure 2.
Taxonomic tree of pathogenic microorganisms present in healthy gut microbiota, adapted from [8,12].

Figure 3.
Taxonomic tree of microorganisms that (a.) increase leads to pathogeny; and those that (b.) decrease leads to pathogeny in the human gut, adapted from [9,11,12,13].


Table 1.
Food additives intended for re-evaluation work program after 2020.
| Food Additive Type | Name | EFSA Number |
|---|---|---|
| Antioxidant | Tartaric Acid | E334 |
| Sodium Tartrate | E335 | |
| Potassium Tartrate | E336 | |
| Sodic Potassium Tartrate | E337 | |
| Calcium Tartrate | E354 | |
| Metataric Acid | E353 | |
| Colorant | Calcium Carbonate | E170 |
| Vegetal Carbon | E153 | |
| Esters of Acetic Acid | E472a, E472b, E472d, E472e, E472f | |
| Texturisers | Stearyl Tartrate | E483 |
| Carrageenan Carboxymethyl Cellulose |
E407 E 466 |
Table 2.
Studies on the effects of CGN consumption.
| Type of additive | Test Type | Model | Period | Concentration | Effects | Footnote | Reference |
|---|---|---|---|---|---|---|---|
| κ/ λ-CGN from C. cripus | Animal (in vivo) |
Mouse | 70 days | 0.0, 5.0, 10 or 20% (w/v) | Mice with highest dose died. | [34] | |
| κ/ λ-CGN from C. cripus | Male and female rats | 23–143 days | 2.0, 5.0, 10, 15, or 20% (w/v) | No effects on appearance or behaviour were observed in male and female Osborne-Mendel or Sprague-Dawley rats fed 5% (w/v). | [42] | ||
| κ/ λ-CGN | Groups of 12 male and 25 female Sprague-Dawley rats | 183 days |
Diet containing 4.0% (w/v) processed, heat sterilised | There was no effect on growth rate, and the caecum and colon were normal on gross and microscopic examination. | The rat caecum is significantly larger than the human caecum, providing a greater surface area for bacterial activity and therefore increasing the potential for absorption in rodents as a result of bacterial degradation, leading to observable results. | [41] | |
| ι-CGN from E. spinosum | 10 male Wistar rats | 56 days | 5.0% (w/v) | Slight diarrhea. | [43] | ||
| Undegraded ι-CGN from E. spinosum | 10 adult male albino guineapigs | 20 days | 1.0% (w/v) | Two of four treated animals had ulcerative lesions in the caecum. The control group remained healthy. | [43] | ||
| ι-CGN from E. spinosum | 7 female guineapigs | 56 days | 5.0% (w/v) | Formation of multiple pinpoint caecal and colonic ulcerations. | [43] | ||
| κ/λ -CGN from C. cripus or G. mamillosa | Groups of 5 male and 5 female rats | Lifelong administration | 0.0, 0.1, 5.0, 15.0, or 25.0% (w/v) | Evidence of hepatic cirrhosis, only at the 25% concentration, with no effect on mortality. | [46] |
||
| κ-CGN from C. cripus | Groups of 30 male and 30 female MRC rats | Lifetime administration | 0.5, 2.5, or 5.0% (w/v) | Soft stool consistency in the begging of the experiment. | [45] | ||
| κ-CGN from C. cripus κ/ λ -CGN derived from C. cripus | Groups of 3 male and 3 female Danish Landrace pigs | 83 days | 0.0, 50.0, 200.0, or 500 mg/kg bw per day | No compound-related deaths were seen, and the behaviour, appearance, and feed intake of the animals remained normal. | In one pig receiving 200 mg/kg bw per day and two pigs receiving 500 mg/kg bw per day, areas of epithelial infolding were observed, along with infiltration of the colonic lamina propria by macrophages and lymphocytes. However, these findings were considered reversible. | [46] | |
| Male and female rhesus monkeys | 49-77 days | 1.0 and 5.0% (w/v) | There were effects of gastrointestinal disturbances at 5%. | [47] | |||
| Male and female infant baboons | 112 days | 0.0, 1.0, or 5.0% (w/v) | No effect was seen on organ or body weights, characteristics of the urine and faeces. | After death, it was possible to observe intestinal flood loss caused by λ -CGN. | [48] | ||
| ι--CGN from E. spinosum and κ- CGN from E. cottonii | Groups of 10 male and 10 female Sprague-Dawley rats | 112 days | 5.0%(w/v) | Formation of multiple pinpoint caecal and colonic ulcerations. | [49] | ||
| κ-CGN from Hypnea musciformis or Irideae crispata | Groups of 15 male and female Sprague-Dawley rats | 28-90 days | 1.0 or 5.0% (w/v) | No changes were observed in the stools of rats receiving 1% of either carrageenan. At 5% concentration, rats had loose stools. | [49] | ||
| λ-CGN | Rats | 365 days | 3,400–3,900 mg/kg (bw) per day | No observation of adverse effects. | The study focuses on reproduction effects, demonstrating that there was no difference related to the dosage, but withing each generation, the fertility decreased with consumption of CGN. | [30] | |
|
λ-CGN Equal parts of ι- and κ-CGN were incorporated in a gelatine capsule |
Guinea pigs | -- | 2.0% (w/v) | Bowel lesions first (from 2 to 6 weeks). Colonic lesions developed after 8 weeks. | [50] | ||
| Mice | 56 days | λ-CGN (1.70 mg/ kg, CGN-L, n = 16.0; 8.30 mg/kg, CGN-M, n = 16; or 41.7 mg/kg, CGN-H, n = 24) | λ-CGN may create an environment that favours inflammation by altering gut microbiota composition and gut bacterial metabolism. | [51] | |||
| Humans | 90 days | 100 mg of food- grade carrageenan | Carrageenan consumption may aggravate ulcerative colitis (UC) disease activity and reduce the interval to relapse in patients who are in clinical remission. | [52] | |||
| λ -CGN | Cell Line (in vitro) |
A Caco-2 absorption model | -- | 100, 500 and 1000 mg/mL | Could not be observed cytotoxicity and no CGN permeability. | This cell line is tumour-derived and, therefore, may not be representative of in vivo intestinal epithelium. | [54] |
| Two cell lines (HEK293 ) | 1906 days | 0.1, 1.0, and 10.0 mg/mL | No effect on oxidative stress was observed after 24 h. | The cell line used in this study differs from the human colon epithelial (NCM460) cells. | [53] | ||
| Human intestinal cells | 3 days | 1.0 μg/ml | Inflammation and colitis. Carrageenan triggers TLR-4 which mediates intestinal inflammation. | [55] |
Table 3.
Studies on the effects of CMC consumption.
| Test Type | Model | Period | Concentration | Effects | Footnote | Reference |
|---|---|---|---|---|---|---|
|
Animal (in vivo) |
Mice | 77 days | 1.0% w/w | Increased disease incidence, leading to chronic inflammation and colitis. | [62] | |
| 84 days | 1.0% w/v | Alteration of the microbiota localisation, composition, and pro-inflammatory potential. | [63] | |||
| 91 days | 1.0% w/v | It confirms the induction of low-grade inflammation. | [64] | |||
| -- | 2.5, 5.0 and 10.0% w/v | No statistically significant or treatment-related adverse effects on any of the parameters evaluated in the safety trials. | The CMC used in the studies was produced from maise husk agrowaste to meet global pharmaceutical standards. | [65] | ||
| Zebrafish embryos |
-- | The maximum CMC concentration is determined as 5000 ppm for microinjection application. | It can lead to important effects on lipid metabolism by causing changes in the expression of some genes associated with obesity. | [66] | ||
|
Cell Line (in vitro) |
MiniBioReactor Array model | -- | 0.1% w/v | Induced a lasting seemingly detrimental impact on microbiota composition and function. | [67] | |
| HT29-MTX and Hep G2 cells | -- | 1.56 and 25.0 mg/mL | Presented a strong pro-inflammatory profile. | [68] | ||
| (M-SHIME) model | -- | 1.00, 0.50, 0.25 or 0.10% w/v | Acted directly upon human microbiota to increase its pro-inflammatory potential. | [68] |
| Increase | Decrease | |
|---|---|---|
| Escherichiacoli | Enterotoxigenic Bacteroides fragilis | F. prausnitzii |
| Desulfovibrio | Clostridium hatheway | Blautia faecis |
| Clostridium cocleatum | Clostridium symbiosum | Roseburia inulinivorans |
| Clostridium thermosuccinogenes | Bacteroides vulgatus | Clostridium lavalense |
| Coprobacillus catenaformis | Veilonella denticariosi | Clostridium coccoides |
| Ruminococcus torques | Clostridium leptum | |
| Ruminococcus bromii-like bacteria | Akkermansia muciniphila | |
| Helicobactor pylori | ||
| Streptococcus bovis | ||
| Fusobacterium nucletum | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



