Submitted:
25 March 2026
Posted:
26 March 2026
Read the latest preprint version here
Abstract
Fatty acids are central to cardiac physiology, serving as both primary energy substrates and precursors for bioactive lipid mediators that shape myocardial structure and function. Essential n-3 and n-6 polyunsaturated fatty acids (PUFAs) are of particular interest because they give rise to prostaglandins, leukotrienes, and a diverse oxylipin network that regulates coronary tone, inflammation, thrombosis, and tissue remodeling in the heart. In parallel, non essential saturated, monounsaturated, and trans fatty acids modulate cardiomyocyte metabolism, membrane organization, and receptor microdomains, thereby influencing how these mediator pathways are engaged in health and disease. Clinically, n--3 long chain PUFAs such as eicosapentaenoic acid and docosahexaenoic acid have been associated with reduced cardiovascular mortality and more favorable post-ischemic remodeling, yet high-dose supplementation has also been linked to a modestly increased risk of atrial fibrillation. Conversely, diets enriched in industrial trans fats and excessive long-chain saturated fats promote dyslipidemia, endothelial dysfunction, and pro arrhythmic remodeling, and are consistently associated with higher rates of coronary artery disease, heart failure, and sudden cardiac death.
At the mechanistic level, cardiac fatty acid handling is governed by coordinated uptake via CD36 and fatty acid transport proteins, mitochondrial β oxidation pathways, and nuclear receptor signaling through peroxisome proliferator-activated receptors, which together determine substrate preference, mitochondrial function, and oxidative stress. Superimposed on these core metabolic processes, cyclooxygenase, lipoxygenase, and cytochrome P450 epoxygenase pathways convert arachidonic acid and n 3 PUFAs into distinct repertoires of prostanoids, leukotrienes, hydroxyeicosatetraenoic acids, epoxyeicosatrienoic acids, and specialized pro resolving mediators that critically influence myocardial inflammation, fibrosis, electrophysiology, and repair. This review synthesizes experimental and clinical evidence on how specific fatty acid species and their oxylipin derivatives contribute to cardiac physiology and pathology, with emphasis on lipotoxic cardiomyopathy, heart failure phenotypes, ischemia–reperfusion injury, and arrhythmogenesis. We also evaluate interventional strategies—including dietary patterns, essential fatty acid supplementation, and pharmacological modulation of fatty acid uptake or oxidation—to optimize cardiac fatty acid and oxylipin metabolism. By framing fatty acids primarily through the lens of essential fatty acid biology and addresses key gaps in linking mechanistic lipid mediator pathways to cardiac outcomes.
Keywords:
fatty acids
; cardiac physiology
; cardiovascular diseases
; lipotoxicity
; N-3 fatty acids
; trans fats
; dietary interventions
; cardiac metabolism
1. Introduction
Fatty acids play a crucial role in cardiac physiology, serving as both essential energy substrates and precursors for bioactive lipid mediators that regulate myocardial function [1]. Among these, the essential n-3 and n-6 polyunsaturated fatty acids (PUFAs) are of particular interest because they give rise to prostaglandins, leukotrienes, and a broad oxylipin network that modulates vascular tone, inflammation, thrombosis, and tissue remodeling in the heart [1]. At the same time, non-essential saturated, monounsaturated, and trans fatty acids shape cardiomyocyte metabolism and membrane composition, thereby influencing how these mediator pathways are activated in health and disease [2,3].
Clinically, specific fatty acid classes exert divergent and sometimes paradoxical effects. N-3 long-chain PUFAs such as eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) can lower overall cardiovascular mortality, yet high-dose supplementation has been associated with a modest increase in atrial fibrillation risk. In contrast, diets rich in industrial trans fats and excessive long-chain saturated fats promote dyslipidemia, endothelial dysfunction, and pro-arrhythmic remodeling, increasing the incidence of coronary artery disease, heart failure, and sudden cardiac death [4,5,6]. These outcome differences reflect not only bulk lipid levels but also distinct patterns of downstream eicosanoids and specialized pro-resolving mediators derived from arachidonic acid (AA), EPA, and DHA.
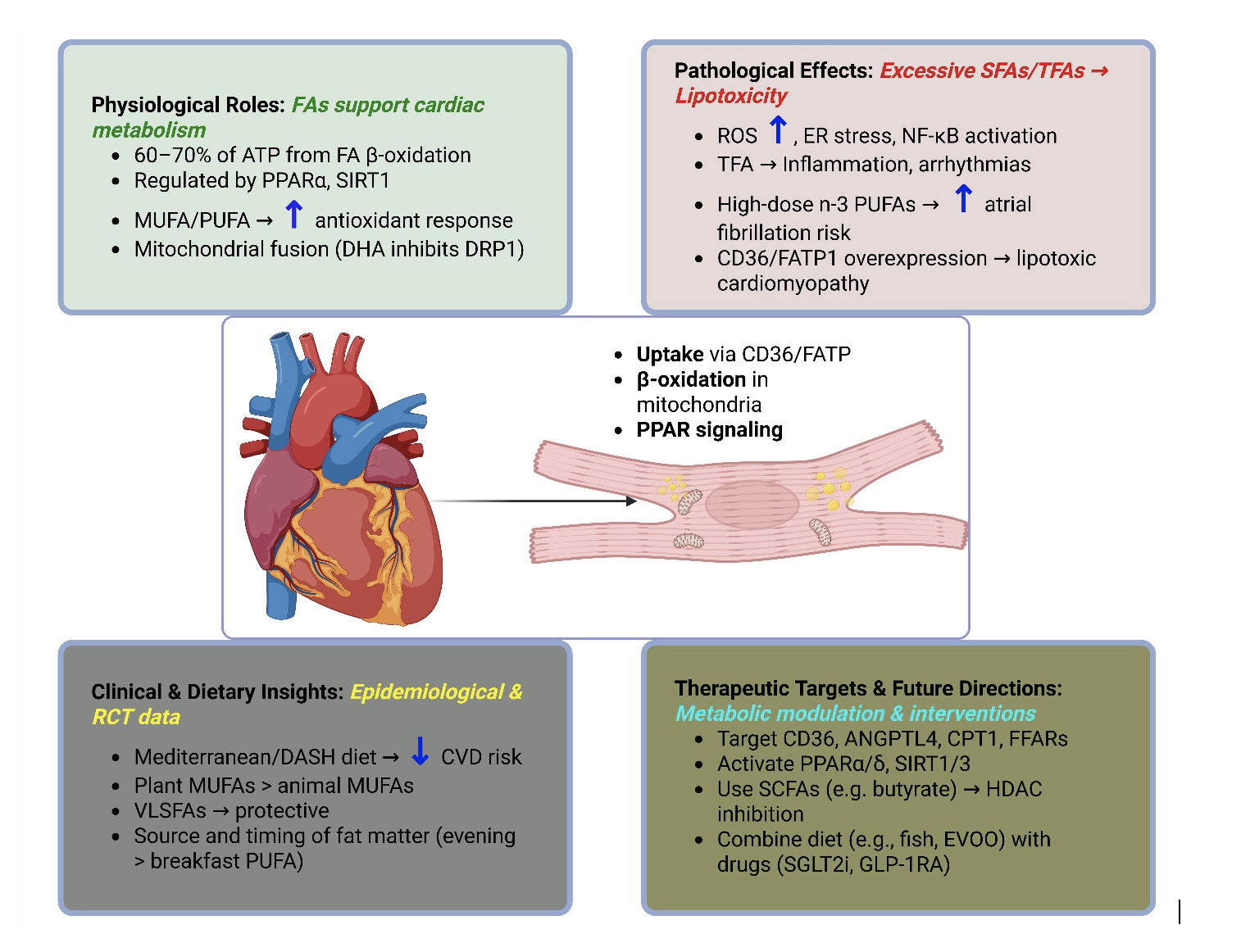
At a molecular level, fatty acids enter the heart via coordinated transport and metabolic systems. Uptake into cardiomyocytes is facilitated by transporters such as CD36 and fatty acid transport proteins (FATPs), followed by activation via acyl-CoA synthetases and mitochondrial import for β-oxidation [7,8]. In the healthy adult myocardium, approximately 60–70% of adenosine triphosphate (ATP) production arises from fatty acid oxidation, underscoring the central role of lipid substrates in sustaining contractile work. Beyond serving as fuels, fatty acids and their oxylipin metabolites act as ligands for nuclear receptors such as peroxisome proliferator-activated receptors (PPARs), thereby regulating transcriptional programs that govern substrate preference, mitochondrial biogenesis, oxidative stress responses, and inflammatory tone. Conversely, chronic overload of long-chain fatty acids (particularly in obesity and diabetes) can precipitate lipotoxic cardiomyopathy, characterized by accumulation of toxic lipid intermediates, mitochondrial dysfunction, and activation of inflammatory and apoptotic signaling pathways.
Given this dualistic nature, a mechanistic understanding of how fatty acids—especially essential n-3 and n-6 series—are converted into prostaglandins, leukotrienes, and related oxylipins, and how these mediators influence cardiac structure and function, is critical for developing targeted dietary and pharmacological interventions. In line with the journal’s focus, the present review is structured around the axis “essential fatty acids → lipid mediator pathways → cardiac outcomes,” while also considering how other lipid classes and dietary patterns modulate this axis. In this context, we synthesize current evidence on (1) the physiological roles of distinct fatty acid species in myocardial energy production and lipid-mediator signaling; (2) the molecular mechanisms linking fatty acid and oxylipin dysregulation to lipotoxicity, inflammation, oxidative stress, and arrhythmogenesis; (3) clinical and epidemiological data relating dietary fatty acid profiles and EFA-derived mediators to cardiovascular outcomes; and (4) interventional strategies—including dietary modifications, nutraceutical supplementation, and molecular therapies—aimed at optimizing cardiac fatty acid and oxylipin metabolism. By explicitly integrating essential fatty acid biology and eicosanoid/oxylipin pathways that directly regulate cardiac inflammation, metabolism, vascular tone, and remodeling.
2. Types of Fatty Acids: Structure, Sources and Cardiac Relevance
The biological roles and cardiovascular effects of fatty acids are fundamentally governed by their chemical structure—particularly chain length, degree of saturation, and double bond geometry (Figure 1). This classification is especially relevant to Prostaglandins, Leukotrienes and Essential Fatty Acids because only the essential n-3 and n-6 polyunsaturated fatty acids (PUFAs)—linoleic acid (LA, 18:2n-6) and α-linolenic acid (ALA, 18:3n-3)—serve as precursors for prostaglandins, thromboxanes, leukotrienes, lipoxins, and specialized pro-resolving mediators (SPMs) that regulate cardiac inflammation, vascular tone, thrombosis, and remodeling [9]. Non-essential saturated fatty acids (SFAs), monounsaturated fatty acids (MUFAs), and trans fatty acids (TFAs) lack these enzymatic conversion pathways but modulate myocardial pools of essential fatty acids (EFAs), membrane microdomains, and lipotoxic stress, thereby shaping oxylipin biosynthesis and signaling.
SFAs, lacking double bonds, are not a monolithic entity with respect to cardiovascular risk. Palmitic acid (C16:0), abundant in palm oil and animal fats, frequently associates with adverse outcomes as it bypasses desaturation, accumulating as ceramides and diacylglycerols that drive insulin resistance, endoplasmic reticulum stress, and atherogenesis via TLR4/NF-κB activation [10,11,12]. This SFA overload can also reduce membrane incorporation of n-6/n-3 EFAs, skewing arachidonic acid (AA)-derived prostanoid/leukotriene production toward inflammation. In contrast, stearic acid (C18:0) is rapidly desaturated by stearoyl-CoA desaturase-1 (SCD1) into oleic acid, minimizing ceramide formation and exerting neutral effects on LDL-cholesterol [13,14]; substitution studies show palmitate replacement with stearate lowers LDL by 10-15% [15]. Very-long-chain SFAs (here defined as C20-C24, though nomenclature varies) enrich cardiac sphingolipids and may influence lipid raft organization of oxylipin receptors, though direct cardiac data remain limited [9].
MUFAs such as oleic acid (C18:1n-9), abundant in olive oil, characterize cardioprotective Mediterranean diets and improve lipid profiles while suppressing NLRP3 inflammasome activation [16,17]. Unlike EFAs, MUFAs do not directly form oxylipins but compete with LA/ALA for desaturase/elongase enzymes (FADS1/2, ELOVL), potentially optimizing tissue n-6/n-3 ratios and reducing AA availability for pro-inflammatory series-2 prostaglandins [18,19]. Oleic acid also stabilizes mitochondrial membranes and enhances PPARα-driven fatty acid oxidation, indirectly supporting the metabolic milieu for EFA-derived mediator production [19].
Essential PUFAs subdivide into n-6 (LA → AA) and n-3 (ALA → EPA/DHA) families that cannot be synthesized de novo and must be dietary precursors for the journal’s core mediators [8,9,20]. N-6 LA (seed oils) elongates/desaturates to AA (20:4n-6), the canonical substrate for COX-2-derived PGE2/TXB2 (vasoconstrictive/pro-thrombotic) and 5-LOX-derived LTB4/C4 (chemoattractant/neutrophil-activating) [21,22]. Excess dietary n-6 relative to n-3 (>10:1 in Western diets) promotes AA dominance and myocardial inflammation post-ischemia [23].
N-3 ALA (flaxseed, walnuts) converts inefficiently (~5-10%) to EPA (20:5n-3) and DHA (22:6n-3) from marine sources, yielding less inflammatory series-3 prostanoids (PGE3/TXB3), 5-LOX-derived resolvins/protectins/maresins, and CYP-derived epoxyeicosatrienoic acids that oppose AA products [24]. EPA/DHA incorporation into sarcolemma alters lipid rafts, ion channel function (Na+/Ca2+), and GPR120 signaling to suppress NF-κB while promoting resolution; optimal cardiac benefits occur at tissue n-6/n-3 ratios <4:1 [25]. Clinical trials show high-dose EPA/DHA reduces cardiovascular mortality but raises atrial fibrillation risk, likely via electrophysiological remodeling [25].
Industrial TFAs (elaidic acid, 18:1n-9t) from partial hydrogenation are non-essential, uniquely atherogenic, and elevate LDL/HDL ratios while promoting endothelial dysfunction. TFAs disrupt membrane order, potentially mislocalizing COX/LOX enzymes or GPCR oxylipin receptors in lipid rafts, though direct cardiac mediator data are sparse; global bans reflect their consistent CVD hazard independent of EFA pathways [26,27].
Dietary fatty acids enter circulation via chylomicrons/VLDL, undergo lipoprotein lipase (LPL)-mediated hydrolysis, and enter cardiomyocytes via CD36/FATPs to fuel β-oxidation (~60-70% myocardial ATP) while establishing EFA pools for phospholipase A2-mediated release and COX/LOX/CYP conversion into oxylipins [9]. Thus, dietary composition directly shapes not only energy homeostasis but also the myocardial substrate landscape for prostaglandin, leukotriene, and SPM biosynthesis, which governs cardiac health and disease.
3. Fatty Acids in Cardiac Physiology
Fatty acids sustain cardiac contractile function both as primary energy substrates (~60-70% myocardial ATP via β-oxidation) and as precursors for oxylipins that modulate excitation-contraction coupling, inflammation, and bioenergetics [1,28,29]. This section examines how essential n-3/n-6 PUFAs and other fatty acids support physiological cardiac work, with emphasis on how their membrane incorporation and phospholipase A2-mediated release establish substrate pools for COX/LOX/CYP-derived prostaglandins, leukotrienes, and SPMs. Non-essential fatty acids provide metabolic context by influencing the incorporation of EFA into sarcolemma/phospholipids and receptor signaling domains.
3.1. Role of Fatty Acids in Cardiac Metabolism
The myocardium exhibits metabolic flexibility but preferentially oxidizes long-chain fatty acids (LCFAs, e.g., palmitate) for efficient ATP production during fasting or moderate workloads. Oxidative metabolism generates ~95% of cardiac energy, with fatty acid β-oxidation predominating over glycolysis even in heart failure, where ketones/lactate utilization increases [28]. Essential PUFAs contribute minimally to bulk ATP (~1-5% oxidation rate) but critically maintain cardiolipin composition and mitochondrial cristae structure, while their sn-2 phospholipid esterification enables rapid PLA2-mediated release for oxylipin biosynthesis under stress [9].
3.2. Fatty Acids as Energy Sources for the Heart
Dietary lipids circulate as chylomicron/VLDL triglycerides, undergo endothelial LPL hydrolysis, and deliver non-esterified fatty acids (NEFAs) bound to albumin for cardiac uptake (Figure 2). During insulin suppression (fasting/exercise), adipocyte hormone-sensitive lipase (HSL) releases additional NEFAs, with insulin normally inhibiting this flux to favor glucose [30]. β-oxidation of LCFAs yields 2.8x more ATP/mole oxygen than glucose, explaining myocardial LCFA preference; however, n-3 PUFAs like DHA resist complete oxidation and instead enrich mitochondrial membranes to optimize electron transport chain efficiency [9,31].
3.3. Mechanisms of Fatty Acid Uptake and Oxidation in Cardiac Cells
Cardiac NEFA uptake occurs via CD36/FATPs in coronary endothelial cells, with trans-endothelial transfer to cardiomyocytes facilitated by FABPpm (Figure 2). CD36, PPAR-regulated and critical for 60-80% of LCFA flux, shows unique cardiac upregulation during fasting, unlike skeletal muscle/adipose tissue [8,28,32,33,34]. Cytosolic FABPs shuttle acyl-CoA esters to mitochondria, where CPT1 converts them to acyl-carnitines for β-oxidation, a rate-limiting step inhibited by malonyl-CoA (ACC product) [8]. LPL deficiency impairs TG uptake but spares basal function via NEFA compensation; stress-exposed LPL-null hearts fail due to ATP substrate limitation [33].
PPARα (abundant in cardiomyocytes) transcriptionally coordinates this system by activating CD36, FABPs, CPT1, and MCAD while KLF15 enhances PPARα expression [35,36,37,38]. AMPK reciprocally promotes fatty acid oxidation (FAO) by phosphorylating ACC (relieving CPT1 inhibition) during energy stress [35]. Essential n-3 PUFAs uniquely activate PPARα/δ and GPR120/FFAR4 to enhance FAO capacity while their oxylipin metabolites (e.g., EETs from CYP2J) vasodilate coronaries to match substrate delivery to demand [39].
3.4. Impact of Different Types of Fatty Acids on Cardiac Function
Under physiological conditions, distinct fatty acids contribute specialized roles beyond bulk energetics (Figure 3):
SFAs: palmitate/stearate provide efficient ATP, but excess induces ceramide-driven lipotoxicity; stearate’s rapid SCD1→oleic conversion minimizes this risk (Li et al., 2022).
MUFAs: oleic acid suppresses NLRP3 inflammasome, stabilizes mitochondria, and competes with n-6 EFAs for desaturases to optimize AA/EPA balance [45,46].
PUFAs (n-3): EPA/DHA incorporate into sarcolemma (altering Na+/Ca2+ channel kinetics, reducing excitability), activate GPR120/NF-κB suppression, generate resolvins/protectins via 5-LOX, and stabilize cardiolipin to prevent mtROS—effects explaining anti-arrhythmic/anti-fibrotic actions [29,47,48,49].
PUFAs (n-6): AA sustains baseline prostanoid production (PGE2 for vascular tone) but excess favors LTB4-driven inflammation when n-6/n-3 >10:1 [9].
TFAs: minimal physiological role; disrupt membrane order and impair ion channel function in myocytes. For example, HDAC inhibitors have been shown to attenuate pathological gene programs in cardiac fibroblasts and myocytes, reducing hypertrophy and fibrosis [50].
Furthermore, specific fatty acids can modulate microRNA (miRNA) expression. Treatment with DHA can alter the expression of miRNAs related to lipid metabolism and angiogenesis in cardiomyocytes (e.g., miR-107, miR-223), while saturated fat consumption deregulates miRNAs implicated in cardiac hypertrophy and fibrosis [51,52].
Short-chain FFAs (C2-C6) preferentially activate FFAR2 (GPR43) and FFAR3 (GPR41), while FFAR1 (GPR40) processes sensing of medium- and long-chain FFAs (> C6) [53,54]. GPR120 is a medium- and long-chain FFA-sensing GPCR that has also been identified, in addition to FFAR1 [53,55] (Da, Micha & Mozaffarian, 2008).
These baseline functions establish myocardial resilience, while pathological EFA/oxylipin shifts (explored next) precipitate dysfunction. Membrane n-3 enrichment particularly lowers resting potential and action potential duration via direct channel modulation, reducing arrhythmia susceptibility independent of metabolism [49].
4. Molecular Pathways and Emerging Research in Fatty Acid-Cardiac Interactions
Fatty acids regulate cardiac physiology through transcriptional, receptor-mediated, inflammatory, and mitochondrial pathways, with essential n-3/n-6 PUFAs providing substrates for COX/LOX/CYP-derived prostaglandins (PGs), leukotrienes (LTs), and SPMs that fine-tune excitation-contraction coupling, bioenergetics, and stress responses [9]. This section examines these pathways with explicit focus on oxylipin biosynthesis and signaling—core to PLEFA’s scope—while SFA/MUFA/TFA effects are contextualized as modulators of EFA pools and membrane microdomains. Each pathway links back to cardiac functional outcomes (arrhythmia, contractility, fibrosis).
4.1. Transcriptional and Nuclear Receptor Pathways
Fatty acids modulate cardiac physiology through distinct molecular pathways, with SFAs and UFAs exerting differential effects on key signaling cascades:
PPAR Regulation
A subfamily of nuclear hormone receptors, known as PPARs, utilizes ligand-activated transcription factors to regulate various biological functions [59]. Alpha (α), beta (β)/delta (δ), and gamma (γ) are the three PPAR isoforms that exhibit differential expression across different tissues [60]. The primary functions of PPAR-α and PPAR-γ are to control glucose homeostasis, insulin sensitivity, and lipid metabolism; their agonists are used to treat hyperlipidemia and type 2 diabetes. In contrast, PPAR-β/δ helps control FAO, glucose homeostasis, lipid metabolism, and inflammation. Its agonists are used to treat cardiovascular and metabolic disorders [59].
Fatty acids serve as endogenous ligands for PPARs, modulating gene expression related to FAO (e.g., CPT1, FABPs, ACOX1), lipid transport, and glucose handling (Li et al., 2024). To control the transcription of target genes, PPARs move into the nucleus after ligand binding, where they heterodimerize with the retinoid X receptor and bind to peroxisome proliferator response elements, specific DNA sequences upstream of target genes [61]. Notably, PPARα regulates cholesterol synthesis via SREBP-2 and suppresses inflammatory gene expression through NF-κB antagonism [62]. n-3 PUFAs (EPA/DHA) are particularly potent PPARα/γ ligands, while their COX-derived metabolite 15d-PGJ2 further activates PPARγ anti-inflammatory signaling in cardiomyocytes (König et al., 2007). PPARs transcription is regulated by factors such as Kruppel-like factors KLF5, KLF4, and KLF15. KLF5 binds to the PPARα promoter, KLF4 regulates mitochondrial biogenesis, and KLF15 associates with PPARα [36].
AMP-Activated Protein Kinase (AMPK) Energy Sensing
As a cellular energy sensor, AMPK is traditionally activated by a decline in energy status, which is indicated by increases in the ratios of AMP to ATP and ADP to ATP. After activation, AMPK promotes ATP-producing catabolic pathways and inhibits energy-consuming processes to restore energy homeostasis [63]. It phosphorylates ACC to relieve the inhibition of CPT1, ultimately decreasing fatty acid synthesis and enhancing FAO, which, in turn, activates SREBP-1c to suppress de novo lipogenesis [64,65].
FFAs can act both as activators and inhibitors of AMPK. For instance, nitrated oleic acid can activate AMPK in endothelial cells, and increased intake of industrial foods and frequent meals appear to increase SFAs, which inactivate AMPK (Canbolat & Cakıroglu, 2023; Wu et al., 2012). In cardiomyocytes, n-6 derived LTB4 inhibits AMPK via p38MAPK while n-3 derived EETs activate it, protecting against ischemia-reperfusion injury [29,66].
Sirtuin Modulation
During the past 20 years, seven isoforms (SIRT1–7) of Sirtuins (SIRTs), which are highly conserved NAD+ (nicotinamide adenine dinucleotide) dependent deacetylases and ADP-ribosyl transferase, have been identified in mammals [67,68]. SIRT3 can increase PGC-1α gene expression [69]. On the other hand, in the liver, increased FAO and gluconeogenesis result from SIRT1 activation by resveratrol, NAD+, fasting, and calorie restriction, through direct deacetylation of PGC-1α, forkhead box protein O1 (FOXO1), and Target of Rapamycin complex 2 (TORC2) [70].
In white adipocytes, SIRT1 stimulates fat mobilization by inhibiting PPAR-γ through docking with its cofactors, SMRT (silence mediator of retinoid and thyroid hormone receptors) and NCoR (nuclear receptor co-repressor [71]. SIRT1 can also directly deacetylate SREBP, and alterations in SIRT1 activity can affect target gene expression, protein stability, and SREBP ubiquitination [72]. Oleic acid is found to modulate rates of FAO by activating the SIRT1-PGC1α transcriptional complex through stimulating the “cAMP/protein kinase A” pathway [73]. Additionally, in the 5/6 Nephrectomy Rat Model, n-3 fatty acids have been shown to activate PGC-1α via deacetylation by upregulating SIRT1/3 [74].
4.2. Inflammatory and Epigenetic Networks
Nuclear Factor Kappa B (NF-κB) Inflammatory Cascade
Fatty acids are potent modulators of inflammatory and epigenetic pathways within cardiac and associated cells. A key inflammatory cascade involves the nuclear factor kappa B (NF-κB) pathway. In adipocytes and macrophages, metabolic stress sensors and signals from toll-like receptors (TLRs) activate the IKK complex, which in turn regulates the expression of NF-κB-driven inflammatory genes such as TNFα, IL-1β, and IL-6 [75]. These cytokines directly promote cardiac fibroblast activation and collagen deposition, driving adverse remodeling post-MI. These cytokines stimulate lipolysis in adipocytes, leading to the release of free fatty acids (FFAs) into the circulation (Edwards & Mohiuddin, 2023; Yan [56]. In the cardiovascular context, HDAC inhibition attenuates pathological gene programs in cardiac fibroblasts and myocytes. For example, HDAC inhibitors have been shown to attenuate pathological gene programs in cardiac fibroblasts and myocytes, reducing hypertrophy and fibrosis [50].
Furthermore, specific fatty acids can modulate microRNA (miRNA) expression. Treatment with DHA can alter the expression of miRNAs related to lipid metabolism and angiogenesis in cardiomyocytes (e.g., miR-107, miR-223), while saturated fat consumption deregulates miRNAs implicated in cardiac hypertrophy and fibrosis [51]. These miRNA shifts directly influence SERCA2a expression and Ca2+ handling in failing myocardium.
4.3. Membrane Receptors and Lipid Signaling
The heart relies on fatty acid-derived signaling for contraction, survival, and adaptation. Key mechanisms include:
G-protein-Coupled Receptor (GPCR) Activation by FFAs
GPCRs, sometimes referred to as seven-transmembrane receptors, are essential membrane proteins expressed on almost every cell type in the body [78]. Free fatty acids (FFAs) act as ligands that bind and activate free fatty acid receptors (FFARs), which are themselves GPCRs [52].
Short-chain FFAs (C2-C6) preferentially activate FFAR2 (GPR43) and FFAR3 (GPR41), while FFAR1 (GPR40) processes sensing of medium- and long-chain FFAs (> C6) [53,54]. GPR120 is a medium- and long-chain FFA-sensing GPCR that has also been discovered in addition to FFAR1 [53,55]. FFAR2 can promote cardiomyocyte hypertrophy by using ERK1/2 to activate signal transducer and activator of transcription 3 (STAT3), and GATA4 and GPR120 activation has been associated with better insulin homeostasis [78,79]. n-3 PUFAs preferentially activate GPR120 to inhibit NF-κB, while PGE2/EP receptors (from amino acid) modulate contractility, and LT receptors drive fibrosis [29].
Lipid Raft Modulation
The plasma membrane’s lipid rafts are microdomains rich in sphingolipids and cholesterol that orchestrate and control a range of signaling pathways. Ion channel regulatory proteins and signaling molecules are abundant in lipid rafts, which are also found in cardiac myocytes [80]. Multiple channel types (such as voltage-gated Na+, K+, and Ca2+ channels) and even various isoforms of a single channel are expressed by the majority of cardiovascular system cells, and each channel contributes differently to excitability [81,82].
The effect of natural FFAs on membrane lipid structure depends on their length and degree of unsaturation [83]. For phospholipid bilayers, long-chain SFAs raise the gel-to-fluid phase (Lβ-to-Lα) transition temperature (also called the melting temperature, or Tm). At the same time, short-chain or cis-unsaturated fatty acids lower the Tm [83]. Lipids that contain unsaturated fatty acids increase membrane fluidity because double bonds cause the fatty acid chains to kink, making it harder for them to pack together [84]. Thus, these fatty acids alter the lipid structure of membranes by altering their fluidity, phase behavior, permeability, fusion, lateral pressure, and flip-flop dynamics [85]. DHA partitions into cardiac lipid rafts to modulate Na+/Ca2+ channel function and reduce the risk of ectopy, while EPA localizes to non-raft regions to optimize anti-arrhythmic effects [49].
Calcium Handling
The connection between the electrical signals that flow through the heart and the contraction of the myocytes to pump blood is made possible by calcium (Ca2+), which is a crucial regulator of cardiomyocyte function [86]. During contraction and relaxation, Ca2+ is released and reabsorbed through the sarcoplasmic reticulum (SR), an organelle that stores Ca2+ [87]. A subtype of sarcoplasmic/endoplasmic reticulum Ca2+ ATPase (SERCA) that is expressed in the heart, SERCA2a mediates the contraction of cardiomyocytes and the re-entry of Ca2+ into the SR through the cytoplasm.
According to an in vitro study conducted at 35 °C, the PUFA content of SR membranes was found to influence cardiac function by modifying SERCA activity in the hearts of Syrian hamsters (Mesocricetus auratus) that hibernate and those that do not [88]. SERCA activity was adversely impacted by the amount of docosahexaenoic acid (DHA; C22:6 n-3), but it increased significantly as the percentage of LA (C18:2 n-6) in SR phospholipid increased. Another in vitro test revealed that, following incubation with phosphatidylcholine-containing SFA, membrane order (stiffness) increased, thereby compromising SERCA activity [89,90]. When the rate of Ca2+ reuptake via SERCA2a decreases, a condition known as Ca2+ overload occurs, which causes the ventricles to relax slowly or not at all, resulting in diastolic dysfunction [90,91].
4.4. Mitochondrial Function and Dynamics
Fatty Acid Oxidation (FAO) Entry
The outer mitochondrial membrane contains the canonical isoforms Carnitine palmitoyl transferase (CPT) 1A and CPT1B, which carry LCFA into the mitochondria for β-oxidation [92]. In brief, the heart can absorb fatty acids through diffusion or FAT/FATP transporters (Lopaschuk et al., 2010b). Fatty acyl-CoA synthase (FACS) inside cardiomyocytes esterifies fatty acids that are bound to fatty acid binding proteins to fatty acyl-CoA. CPT1 catalyzes the formation of acyl-carnitine, the rate-limiting step in mitochondrial β-oxidation that regulates fatty acid entry into the mitochondria. After being transported into the mitochondria, CPT 2 transforms the acylcarnitine back into fatty acyl-CoA, the majority of which enters the fatty acid β-oxidation cycle (Lopaschuk et al., 2010b). n-3 derived EETs from CYP2J2 vasodilate coronaries to match FAO substrate delivery to cardiac workload [29].
Reactive Oxygen Species (ROS)
During aerobic respiration, mitochondria are the primary consumers of molecular oxygen in cells [94]. A tiny amount of the oxygen is partially reduced by the mitochondrial electron-transport chain (ETC), which results in the production of mitochondrial ROS (mtROS), even though the majority of the oxygen consumed is reduced to water, creating a proton-motive force in the process that propels the synthesis of ATP [94]. These mtROS molecules can damage mitochondrial DNA, proteins, and lipids by inducing oxidative stress [95]. Conversely, n-3 FAs were also found to enhance antioxidant defense against ROS [96]. Treatment with n-3 PUFA prevents dilated cardiomyopathy (DCM) in mice primarily through ROS suppression and mitochondrial protection, though direct pro-apoptotic effects remain uncertain in human disease [97].
Mitochondrial Dynamics
Large GTPases that are members of the dynamin superfamily are primarily responsible for regulating mitochondrial dynamics in mammals [98]. Specifically, dynamin-related-like protein 1 (DRP1) promotes mitochondrial fission, whereas mitofusin 1 and 2 (MFN1 and MFN2) and optic atrophy 1 (OPA1) are necessary for mitochondrial fusion [98,99,100,101,102].
4.5. Emerging Research Frontiers in Cardiac Lipid Biology
Membrane Lipid Remodeling and Microdomain Signaling
n-3 PUFA Incorporation
EPA/DHA incorporate into membrane phospholipid bilayers, modulate membrane fluidity, and disrupt lipid raft-dependent signaling (e.g., TLR4/NADPH oxidase) [103]. DHA preferentially partitions into lipid rafts, whereas EPA localizes to non-raft regions, which explains their differential anti-inflammatory effects [104]. This differential partitioning influences the localization of COX/LOX receptors and SPM production in cardiac membranes.
Cardiolipin (CL) Dynamics
Distinct membrane lipids, such as CL, also play pivotal roles. CL is a mitochondria-specific phospholipid (∼15–20% of inner membrane lipids) that in the heart is unusually homogeneous—largely tetralinoleoyl (all 18:2) in composition [105]. It anchors the electron-transport supercomplexes and maintains cristae architecture; perturbations of CL quantity or acyl-chain composition dramatically impair mitochondrial bioenergetics and dynamics [105]. Beyond energy metabolism, injured cardiomyocytes externalize oxidized CL, which serves as a potent damage-associated molecular pattern that activates innate immune receptors and triggers inflammation [106]. In sum, both the fatty acyl chains of energy substrates and the polar headgroup lipids of membranes integrate to regulate mitochondrial function, calcium handling, and inflammatory signaling in the myocardium.
Very Long-Chain SFA (VLSFA) Paradox
VLSFAs (C>24 saturated fatty acids) and their ceramide/sphingomyelin derivatives are inversely correlated with heart failure risk, likely via sphingolipid-mediated membrane stabilization of oxylipin receptors [107], suggesting these lipids favor healthy aging of the myocardium.
Mitochondrial Plasticity and Metabolic Flexibility
Fission-Fusion Balance
n-3 PUFAs: n-3 PUFAs markedly influence mitochondrial dynamics and calcium homeostasis. DHA has been shown to stabilize mitochondrial networks under stress by inhibiting the activation of the fission protein Drp1 (decreasing phospho-Drp1 at Ser616), thereby preventing stress-induced fragmentation, preserving ATP production, and reducing mtROS in cardiac ischemia models [108].
Lipotoxicity: The accumulation of lipids in cardiomyocytes is also associated with increased oxidative stress [109]. For example, long-term (>8h) palmitate exposure increases ROS production, accompanied by reticulum loss and a pattern suggesting increased mitochondrial fission via activation of the fission protein Drp1 (increased phospho-Drp1 at Ser616), resulting in apoptosis [108,110].
Alternative Fuels
Microbiota-derived SCFAs (acetate, propionate, and butyrate) enter the circulation and serve as alternative fuels; in fact, these may contribute to myocardial energetics under specific pathological conditions, although quantitative estimates and relevance to lipid mediator signaling remain uncertain [50]. Thus, dietary and microbial lipids together tune mitochondrial metabolism and calcium signaling in cardiomyocytes.
Chrononutrition and Circadian Lipid Metabolism
Meal Timing
Chrononutrition (which examines the relationship between the timing of food intake and the body’s circadian rhythms) has emerged as a key variable: recent cohort analyses indicate that the circadian timing of fat intake influences cardiovascular risk. For instance, Evening PUFA intake lowers CVD mortality (HR 0.85), while breakfast PUFA increases risk (HR 1.30) [111]. These associations suggest circadian regulation of COX-2/SPM biosynthesis, though RCTs are needed to establish causality. One speculative explanation is that evening n-3 PUFA intake may better align with circadian regulation of SPM biosynthesis, whereas morning intake could interfere with diurnal patterns of arachidonic-acid mobilization; however, direct evidence linking meal timing to cardiac oxylipin flux is currently lacking.
Time-Restricted Feeding
Time-restricted feeding aligns nutrient intake with circadian lipid oxidation cycles, improving plasma triglycerides, HDL, and myocardial FAO gene expression in metabolic syndrome models [112].
Gut-Heart Axis Modulation (PLEFA-Compliant)
SCFAs as Oxylipin Adjuvants
Although SCFAs are not lipid mediators in the classical prostaglandin or leukotriene sense, they may indirectly influence cardiac inflammatory tone by modifying epigenetic and metabolic contexts in which oxylipin signaling operates. Microbial SCFAs have been shown to exert epigenetic effects: butyrate and propionate inhibit host HDACs [50]. This HDAC inhibition has a functional impact: it attenuates pathological gene programs in cardiac fibroblasts and myocytes. Indeed, studies show that HDAC inhibitors (including SCFAs such as butyrate and valproate) reduce hypertrophy and fibrosis by altering chromatin at growth-factor target genes. SCFAs have been reported to dampen oxidative and proinflammatory signaling, thereby indirectly influencing the inflammatory milieu in which oxylipin signaling operates [50]. Collectively, SCFAs may modulate the cellular context in which n-3–derived specialized pro-resolving mediators exert cardioprotective effects, without directly participating in oxylipin biosynthesis.
TMAO Counteraction
The gut–heart axis is also a significant research frontier: shifts in microbiota composition alter circulating lipids (low SCFAs, high TMAO) that modulate vascular tone and inflammation. Trials in mice have shown that restoring SCFAs (via fiber or acetate) lowers blood pressure and attenuates cardiac remodeling [50,113].
Direct evidence linking gut-derived signals to myocardial oxylipin production or prostaglandin/leukotriene-mediated cardiac outcomes remains limited.
Omics-Driven Discoveries
Lipidomics
Integrative lipidomic and transcriptomic approaches are mapping the cardiac lipidome and its genetic regulation. For example, a recent “lipidome atlas” of the developing heart revealed that postnatal enrichment of DHA-containing phosphatidylcholines and phosphatidylethanolamines coincides with upregulation of specific acyltransferases (Lpcat3, Agpat3) [114], pinpointing molecular drivers of membrane remodeling. In adult studies, combined omics profiling is uncovering lipid signatures and gene modules that differentiate healthy versus diseased myocardium. In sum, emerging chronobiology and multi-omics tools are deepening our mechanistic understanding of how diverse fatty acids regulate cardiac function and adaptation [111,114].
Transcriptomics
5. Pathological Effects of Fatty Acids
Fatty acids exert differential effects on cardiac health depending on their structure and metabolic context. While unsaturated fatty acids, particularly n-3 PUFAs, are often cardioprotective, excessive intake of SFAs and TFAs is closely associated with detrimental cardiac outcomes. This section outlines key pathological mechanisms by which fatty acids contribute to CVD.
5.1. Lipotoxicity and Metabolic Stress
Excess fatty acid influx overwhelms cardiac β-oxidation capacity, leading to toxic lipid intermediates (ceramides, diacylglycerols) that impair insulin signaling, activate PKCθ, and trigger apoptosis via ER stress/UPR pathways [115]. Palmitate is particularly prone to accumulation as ceramide through de novo synthesis via serine palmitoyltransferase, resulting in activation of protein phosphatase 2A, dephosphorylation of Akt, nuclear translocation of FoxO3a, and induction of pro-apoptotic mediators including Bim and Bad [10]. This lipotoxic cardiomyopathy manifests as diastolic dysfunction, systolic decline, and fibrosis, exacerbated by diabetes/obesity, where CD36 upregulation amplifies LCFA uptake [33]. In contrast, n-3 polyunsaturated fatty acids attenuate ceramide accumulation by activating PPARα-dependent peroxisomal oxidation pathways, including upregulation of ACOX2 (Li et al., 2022). Additionally, EPA-derived PGE3 can competitively antagonize arachidonic acid–derived PGE2 signaling at EP2 and EP4 receptors, thereby suppressing TGF-β–driven cardiac fibroblast activation [103]. In parallel, LTB4 (n-6 derived 5-LOX) exacerbates macrophage infiltration and collagen deposition post-injury (Kimura et al., 2020).
5.2. Fatty Acid Profiles in Heart Failure Phenotypes
Systolic failure (known as Heart failure with reduced ejection fraction (HFrEF)) is marked by reduced FAO, increased glycolysis via PDK4, impaired CPT1 (malonyl-CoA ↑), and ceramide buildup [115]. PPARα/PGC-1α repression reduces mitochondrial biogenesis; therapeutic FAO partial inhibition (trimetazidine, ranolazine) restores coupling efficiency [118]. Paradoxically in diastolic impairment (Heart failure with preserved ejection fraction (HFpEF)), fatty acid oxidation can paradoxically increase despite fibrosis and stiffness, with palmitate activating NLRP3 and interleukin-1 beta, contributing to diastolic dysfunction (Li et al., 2022). n-3 trials show mixed EF preservation but consistent ↓sudden death [119]. HFrEF shows n-6/n-3 >15:1 with elevated AA-derived HETEs and LTB4; HFpEF shows low EPA and DHA levels correlating with fibrosis severity [120]. Resolvin D1 infusion improves diastolic parameters in preclinical HFpEF [121].
5.3. Ischemia-Reperfusion Injury and Arrhythmia
During acute ischemia, AA release increases, causing a COX-2-mediated imbalance of thromboxane A2 and prostacyclin, which promotes coronary vasoconstriction and thrombosis [122,123,124]; reperfusion elevates LTB4, activating neutrophils and contributing to microvascular no-reflow [125,126]. n-3 acute infusion produces protectin D1, reducing infarct size by 30-50% via ALX/FPR2-mediated neutrophil apoptosis [127,128]. DHA sarcolemma incorporation shortens APD via direct modulation of NaV1.5/SERCA2a [29,47,48,49], whereas high-dose EPA or DHA can increase the risk of atrial fibrillation through IKur hyperpolarization [4]. Trans fatty acids disrupt connexin-43 localization, causing conduction heterogeneity [26,27].
5.4. Atherosclerosis and Vascular Interactions
Cardiac capillary ECs express COX-2 and 5-LOX, and the resulting PGE2 and LTB4 promote monocyte adhesion and facilitate LPL-mediated long-chain fatty acid delivery to foam cells [129,130]. n-3 SPMs (e.g., maresin-1) activate macrophage efferocytosis, thereby improving plaque stability [103]. Perivascular adipose tissue releases palmitate, which activates cardiac TLR4 and drives concentric left ventricular remodeling [131].
These findings underscore the importance of dietary modulation (Table 1) —specifically, reducing SFA and TFA intake and increasing n-3 PUFA consumption—as a strategy for preventing and mitigating cardiac disease.
6. Clinical Implications and Epidemiological Studies
6.1. Saturated Fatty Acids (SFAs)
Among the four major fatty acid classes, trans fats and most SFAs elevate coronary heart disease (CHD) risk by increasing serum cholesterol levels [132,133]. A 14-year follow-up study (n=939 incidents) revealed that longer-chain SFAs (12:0–18:0) were independently associated with increased CHD risk, whereas short-to-medium-chain SFAs (4:0–10:0) were not [134]. Interestingly, very-long-chain SFAs (VLSFAs; >C24), e.g., lignoceric acid, demonstrate cardioprotective effects—linked to lower risks of heart failure, CHD, atrial fibrillation, and mortality when present at higher concentrations [135].
The EPIC-CVD case-cohort study (n=385,747; 10,529 CHD cases) demonstrated that SFA effects depend critically on food sources:
Harmful sources: Red meat (HR: 1.07; 95% CI: 1.02–1.12) and butter (HR: 1.02; 95% CI: 1.00–1.04) [136].
Neutral/beneficial sources: Yogurt (HR: 0.93; 95% CI: 0.88–0.99), cheese (HR: 0.98; 95% CI: 0.96–1.00), and fish (HR: 0.87; 95% CI: 0.75–1.00) [136].
This source-dependent risk is further supported by Dutch cohort data, which show that higher palmitic acid intake and substitution of SFA with animal protein elevate CHD risk [137]. Plant-derived SFAs may raise LDL-C (3.43–9.18 mg/dL) but also increase HDL-C (0.94–1.89 mg/dL), yielding neutral cardiovascular effects when substituted for plant-based UFAs [138]. Notably, a comprehensive review (2010–2021) found no consistent correlation between naturally occurring SFA consumption and CVD risk [139].
These source-dependent effects likely reflect differential impacts on the risk of essential fatty acid metabolism [137]. Red meat and butter increase palmitic acid, which displaces AA from membrane phospholipids, leading to elevated pro-thrombotic thromboxane A2 and prostaglandin E2, whereas fish/dairy provide VLSFAs that stabilize oxylipin receptor microdomains [138,140,141].
6.2. Unsaturated Fatty Acids (UFAs)
UFAs demonstrate complex, sometimes contradictory cardiometabolic effects. A 12-year study of 1,807 ischemic heart disease (IHD) cases paradoxically found that replacing SFAs with cis MUFAs, PUFAs, or animal protein increased the risk of IHD (HR per 5% energy: 1.27–1.37) [142]. However, meta-analyses indicate MUFAs reduce total and hemorrhagic stroke risk [143], though not CVD mortality [144]. Replacing SFAs with UFAs/carbohydrates/protein reduces CVD events by 21% [145], with MUFAs and PUFAs similarly improving triglycerides, LDL-C, and blood pressure—though MUFAs show superior nighttime systolic BP reduction [146], a critical CVD risk factor [147].
These UFA benefits reflect n-6/n-3 balance optimization. Marine n-3 PUFAs (EPA/DHA) generate anti-arrhythmic series-3 prostaglandins (PGE3) and resolvins that compete with AA-derived pro-thrombotic TXB2/LTB4, explaining sudden death reduction despite inconsistent total CVD effects [132,148,149,150,151,152,153].
6.3. Food Source Considerations
The source of fatty acids substantially modulates their cardiovascular impact. International guidelines emphasize:
Beneficial: Plant-based foods (whole grains, fruits, vegetables), sea fish, fermented dairy, lean meats [154].
Detrimental: Processed meats, sugar-sweetened beverages, butter, cream, refined starches [154].
Whole-food approaches (e.g., Mediterranean diet) reduce CVD mortality more effectively than isolated nutrient modifications [155].
6.4. Timing of Intake
Emerging evidence highlights meal timing as a key modulator of UFA effects. In a cohort of 30,136 adults, high breakfast intake of PUFAs (HR: 1.30; 95% CI: 1.13–1.50), MUFAs (HR: 1.28; 95% CI: 1.13–1.45), or total UFAs (HR: 1.35; 95% CI: 1.17–1.57) increased CVD mortality. Conversely, dinner consumption of MUFAs/total UFAs reduced all-cause mortality, while PUFAs lowered both CVD and all-cause mortality [111]. However, these observations are derived from association studies and do not establish causality, as residual confounding and reverse causation cannot be excluded. Evening n-3 intake aligns with peak SPM biosynthesis; breakfast disrupts diurnal AA mobilization → ↑pro-thrombotic eicosanoids [159,160].
6.5. Reconciling Controversies: A Unifying Perspective
The relationship between fatty acids and CVD is not monolithic. SFA effects depend critically on chain length (with VLSFAs being protective) and food matrix (dairy sources neutral/beneficial vs. meat sources harmful). UFAs generally confer benefits, but outcomes are influenced by type (n-3 > n-6 PUFAs), source (plant > animal), and timing (dinner > breakfast). These nuances explain apparent contradictions in the literature and underscore the importance of considering fatty acids within their dietary context rather than as isolated nutrients. Clinical heterogeneity reflects tissue n-6/n-3 ratios, which shape the oxylipin repertoire (pro-thrombotic series-2 vs. anti-arrhythmic series-3 and SPMs) [161]. VLSFAs stabilize receptor microdomains; meal timing modulates circadian PLA2/COX-2 activity.
7. Interventional Studies
Evidence-based dietary patterns consistently demonstrate cardioprotective effects, although their benefits are multifaceted and cannot be attributed solely to their fatty acid composition [162,163,164]. The Mediterranean diet is characterized by a high intake of extra-virgin olive oil (EVOO), nuts, legumes, whole grains, and vegetables. In the PREDIMED trial, supplementing with EVOO or nuts reduced major cardiovascular events compared to a low-fat control diet [162]. EVOO delivers oleic acid and polyphenols that optimize the competition between linoleic acid and alpha-linolenic acid, resulting in tissue n-6 to n-3 ratios ~4:1, increasing resolvins and downregulating LTB4, which helps explain the 30% reduction in cardiovascular events observed in PREDIMED [161,162,165].
The DASH diet, which emphasizes fruits, vegetables, low-fat dairy, and reduced sodium, robustly lowers blood pressure and improves lipid profiles [163,164]. ALA enrichment, combined with low SFA, increases circulating EPA, promoting series-3 PGs and complementing potassium-mediated vasodilation [166,167].
Similarly, plant-based diets rich in whole foods and fiber are associated with lower LDL-C levels, reduced inflammation, and a lower incidence of coronary heart disease [168,169,170]. LA/ALA synergy drives n-6 to n-3 ratios <5:1, reducing TXB2/thrombosis despite limited DHA conversion (Yang et al., 2014).
However, several clinical trials have investigated the effect of n-3 PUFAs on CVD outcomes (Table 2). The mechanisms underlying the success of these diets are complex and synergistic. While the high content of MUFAs and PUFAs in the Mediterranean diet likely contributes to improved lipid profiles and reduced inflammation, other components—such as fiber, polyphenols, antioxidants, and the low glycemic load—play significant and potentially additive roles in reducing oxidative stress, improving endothelial function, and modulating the gut microbiome [173]. Therefore, the cardioprotection offered by these patterns is best viewed as an emergent property of the entire dietary matrix, rather than a consequence of any single nutrient class—though tissue n-6/n-3 ratios and oxylipin profiles provide mechanistic biomarkers for their efficacy [174].
8. Future Directions
As detailed, recent advances in membrane lipid remodeling (e.g., VLSFAs) and gut-heart axis signaling provide novel targets for intervention.
8.1. Future Therapeutic Directions
Modulation of fatty acid uptake represents a logical strategy to prevent lipotoxicity in the heart. In cardiomyocytes, LCFAs enter the cell via LPL cleavage products and surface transporters, including FAT/CD36, FABPpm, and members of the FATP family [175]. Notably, transgenic mice with cardiac-specific overexpression of FATP1 exhibit massive lipid uptake and develop lipotoxic cardiomyopathy [175]. At the same time, FAT/ CD36 deficiency shifts myocardial substrate use toward glucose and can protect against lipid overload. Conversely, ANGPTL4—an LPL inhibitor induced by PPARδ—limits lipid uptake into cells [175] and has been shown to mitigate lipid accumulation in muscle and heart.
However, therapeutic inhibition of FAT/CD36 may not be without risk: FAT/CD36 facilitates rapid fatty acid uptake during cardiac stress, and its absence may impair myocardial energy homeostasis under increased workload [176,177,178,179]. Thus, careful targeting—such as partial or context-dependent modulation rather than complete inhibition—may be necessary to avoid compromising cardiac performance.
FFAR4/GPR120 agonists—n-3 PUFAs, inhibit NF-κB through β-arrestin and sequester NLRP3 in cardiomyocytes [180,181,182,183].
Once inside the cell, fatty acids must be handled by mitochondrial and enzymatic machinery to avoid toxic buildup. CPT1 controls mitochondrial entry of long-chain fatty acids; its inhibition by malonyl-CoA (produced by acetyl-CoA carboxylase, ACC) provides a checkpoint for FAO. Thus, ACC inhibitors or malonyl-CoA decarboxylase activators can enhance CPT1 activity and boost β-oxidation, improving energy output. At the gene level, PPAR nuclear receptors orchestrate these pathways: PPARα (with coactivator PGC-1α) upregulates genes for fatty acid transport and β-oxidation (including CPT1), whereas PPARδ similarly promotes oxidative metabolism. Activators of PPARα and PPARδ, as well as SIRT1 and SIRT3, increase CPT1 and MCAD expression while reducing SREBP-1c, thereby accelerating FAO [72,184].
Additionally, Table 3 includes the target and effects of every USFDA-approved medication listed in DrugBank [185]. Targets including 3-hydroxy-3-methylglutaryl coenzyme A (HMG-CoA) reductase, angiotensin-converting enzyme (ACE), beta-1 adrenergic receptor, bile acid, antithrombin-III (ATIII), PPARα, and P2Y purinoceptor 12 are all clinically relevant. Statins (HMG-CoA reductase inhibitors), PCSK9 inhibitors, SGLT2 inhibitors, GLP-1 receptor agonists, and PPARα agonists remain cornerstone therapies for lipid and glucose modulation, inflammation reduction, and cardiovascular risk mitigation [186]. These combined interventions—spanning fatty acid transporters, metabolic enzymes, nuclear receptors, and immune sensors—form a coherent therapeutic framework to prevent lipotoxic cardiac injury and improve substrate handling in CVD.
8.2. Innovative Dietary Strategies
Several evidence-based dietary patterns have been shown to be cardioprotective. For example, the randomized PREDIMED trial showed that a Mediterranean-style diet (rich in vegetables, fruits, whole grains, and extra-virgin olive oil or nuts) significantly reduced the risk of major cardiovascular events compared to a low-fat control diet (Estruch et al., 2018b). Consumption of EVOO and nuts results in tissue n-6 to n-3 ratios ~4:1, which increases resolvin production [187,188].
Short-term trials in healthy older adults have also confirmed that a Mediterranean diet modestly lowers blood pressure and improves endothelial function [189]. Likewise, the DASH (Dietary Approaches to Stop Hypertension) eating plan—emphasizing fruits, vegetables, low-fat dairy, and reduced sodium—robustly lowers systolic blood pressure and favorably modulates lipid homeostasis and other metabolic risk factors [164]. Plant-based dietary patterns (vegetarian or vegan) similarly confer cardiovascular benefits: multiple studies have found that higher intake of plant foods is associated with reductions in total and LDL cholesterol and improvements in inflammatory profiles. For instance, vegetarians tend to consume less saturated fat and have lower LDL-C than omnivores [190], contributing to lower atherosclerotic risk, and recent reviews have concluded that healthy plant-based diets reduce cardiovascular morbidity and mortality in diverse populations [190].
Beyond overall patterns, the quality and source of dietary fat appear to influence outcomes. In cohort analyses, replacing SFA from red and processed meats with SFA from dairy sources (especially cheese) has been linked with lower CVD risk [191]. These observations suggest that not all SFAs have identical effects—for example, dairy-derived SFAs may be less atherogenic than meat-derived SFAs.
More broadly, replacing dietary SFAs with unsaturated fats (monounsaturated and polyunsaturated) generally improves lipid profiles: meta-analyses show that substitution of SFAs with PUFAs lowers LDL-C and the total/HDL cholesterol ratio. In fact, one metabolic ward study indicated that a balanced fatty acid mix (roughly SFA:MUFA: PUFA in a 1:1.3:1 ratio) produced greater reductions in LDL-C without depressing HDL-C [192]. This underlies recommendations to emphasize high-MUFA foods (like olive oil and nuts) and adequate PUFA (especially n-3 and n-6) to optimize the LDL/HDL balance.
Emerging concepts in diet timing and composition are also of interest. Chrononutrition—the study of meal timing about circadian biology—suggests that restricting the daily eating window may improve cardiometabolic health. In a trial of patients with metabolic syndrome (most of whom were on statins and antihypertensives), adopting a 10-hour self-selected feeding window (TRE) for 12 weeks led to weight loss, lower blood pressure, and reductions in “atherogenic” lipid fractions (e.g., triglycerides) [193]. Thus, TRE can complement standard medical care by improving risk factors independently of calorie counting. Likewise, seafood-rich diets garner attention for their n-3 PUFA content. Oily fish, such as salmon, mackerel, herring, and sardines, provide 1–2.5 g of EPA+DHA per serving [194], and regular fish intake has been associated with lower levels of circulating triglycerides and inflammatory cytokines. (For example, epidemiologic studies report that higher fish consumption correlates with reduced C-reactive protein and other vascular inflammation markers.) Together, data on dietary fats and timing suggest that both what one eats and when one eats can modulate lipids, blood pressure, endothelial function, and inflammatory status.
Finally, integrating a diet with pharmacotherapy may enhance cardiovascular prevention. There is growing evidence that combining n-3 PUFA supplementation with statin therapy yields incremental benefit. A recent meta-analysis of imaging studies found that patients receiving both high-dose EPA/DHA and a statin (compared to statin alone) had significantly slower progression of coronary plaque, thicker fibrous caps, and lower high-sensitivity C-reactive protein (hs-CRP) levels [195]. In other words, n-3s appeared to stabilize atherosclerotic lesions beyond the effects of statins alone, without adverse changes in HDL or LDL levels. Mechanistically, both statins and n-3 PUFAs share anti-inflammatory and plaque-modulating pathways (for example, each promotes the production of pro-resolving lipid mediators) [194]. These synergistic effects translate into improved lipid profiles (notably lower triglycerides) and attenuated vascular inflammation when diet and drug are combined.
In sum, a comprehensive cardioprotective diet emphasizes vegetables, fruits, whole grains, legumes, nuts, fish, and unsaturated fats, while moderating refined carbohydrates and saturated fats—with special consideration of fat sources (dairy vs. meat) and fat types (MUFA vs. PUFA) [191,192,196]. Emerging strategies, such as time-restricted feeding and increased intake of oily fish, further improve traditional risk factors (blood pressure, dyslipidemia, endothelial function, and inflammatory biomarkers) [193,194]. When combined with optimal pharmacotherapy, these dietary approaches can produce additive cardiometabolic benefits. For instance, co-administering n-3 PUFAs with statins yields greater reductions in triglycerides, C-reactive protein, and plaque vulnerability than statins alone [194,195]. Collectively, these data support a multifaceted dietary prescription (e.g., Mediterranean/DASH/plant-based patterns with source-specific fat guidance and consideration of meal timing) to maximize cardiovascular prevention and treatment outcomes.
9. Conclusions
Longer-chain saturated fatty acids (SFAs; C14:0–18:0), especially those found in red meat, butter, and palm oil, are consistently associated with increased CVD risk, particularly CHD. These associations reflect that palmitate displacement of AA from membrane phospholipids is associated with increased pro-thrombotic TXA2/PGE2 production [103]. Although the associations between other SFAs and UFAs and specific CVD outcomes remain inconsistent, this likely reflects residual confounding and heterogeneity in food sources, metabolism, and clinical context.
Importantly, not all SFA-rich foods are equal. While processed meats, butter, and cream should be minimized—especially in individuals with diabetes or elevated CVD risk—fermented dairy (e.g., yogurt, cheese) and seafood (e.g., sardines) often exhibit neutral or even beneficial cardiometabolic effects due to their complex nutrient matrices. VLSFAs from dairy/fish stabilize oxylipin receptor microdomains while seafood delivers EPA/DHA → series-3 PG/SPM cardioprotection [197,198,199].
Replacing SFAs with PUFAs, particularly those found in whole foods such as fish, nuts, and olive oil, remains a cornerstone dietary strategy for CVD prevention. EPA/DHA substitution, associated with a lower tissue n-6/n-3 ratio, has been linked to reduced dominance of AA-derived LTB4 and TXB2 signaling and may contribute to the observed reduction in cardiovascular events [174]. Evidence-based patterns such as the Mediterranean and DASH diets—rich in these components—should be prioritized across populations, including those with established CVD. In contrast, dietary strategies such as the ketogenic diet (KD) and intermittent fasting (IF)—which fall outside the core scope of this review—have been proposed to influence cardiometabolic outcomes, but their effects on cardiac EFA pools and oxylipin signaling remain insufficiently explored.
Whole foods should be prioritized over supplements; for instance, sardines and other oily fish provide n-3 PUFAs alongside beneficial nutrients such as calcium, potassium, and selenium, making them superior to isolated supplements. While n-3 PUFA supplementation may be beneficial in some cases, it is limited by unclear dosing thresholds, bioavailability issues, and potential risks (e.g., an increased risk of atrial fibrillation at high doses).
Key Take-Home Messages:
- Prioritize whole foods: Choose fish (especially sea fish like sardines, salmon etc), nuts, and extra-virgin olive oil over supplements.
- Avoid industrial trans fats (iTFAs) and minimize consumption of processed meats, refined carbohydrates, sugar-sweetened beverages, and excessive sodium.
- Replace saturated fats with polyunsaturated fats from natural sources, not refined seed oils or processed alternatives.
- Adopt established dietary patterns, such as the Mediterranean or DASH diets, which have consistently been shown to lower CVD risk.
- Use supplements judiciously, only where whole-food alternatives are insufficient or contraindicated.
- Support emerging dietary approaches (e.g., KD, IF) with well-designed randomized trials before broad implementation.
Future research should focus on refining supplementation protocols, investigating synergistic whole-food combinations, and expanding clinical trials of promising but understudied dietary interventions.
Tables

Table 3.
Summary table for USFDA-approved drugs against Cardiovascular problems [185].
Table 3.
Summary table for USFDA-approved drugs against Cardiovascular problems [185].
| Targets | Inhibitors (drug bank ID) | Effects |
| Angiotensin-converting enzyme (ACE) inhibitor | Perindopril/ DB00790 |
Used in combination with Atorvastatin (DB01076) to prevent CVD events [200]. |
| Ramipril/ DB00178 |
Reduction of cardiovascular mortality, MI [201]. | |
| Quinapril/ DB00881 |
Treat hypertension, congestive heart failure [202]. | |
| Lisinopril/ DB00722 |
Treat hypertension, heart failure, and acute MI [203]. | |
| Trandolapril/ DB00519 |
Treat hypertension, congestive heart failure, and improve survival following a MI [204]. | |
| Fosinopril/ DB00492 |
Used to treat mild to moderate hypertension, congestive heart failure [205]. | |
| Enalapril/ DB00584 |
Used to treat mild to moderate hypertension, congestive heart failure [205]. | |
| Acyl-CoA:1,2-diacylglycerol acyltransferase (DGAT) inhibitor | Icosapent ethyl/ DB08887 |
Reduce the risk of MI, stroke, coronary revascularization, elevated triglycerides (≥150 mg/dL) and established cardiovascular disease [174]. |
| Aldo-keto reductase family 1 member C2 inhibitor | Ursodeoxycholic acid/ DB01586 | Reduces cholesterol levels in the blood [206]. |
| Antithrombin-III inhibitor | Fondaparinux/ DB00569 |
Prevent venous thromboembolism to improve survival following MI [207]. |
| Apolipoprotein C-III (APOC3, apoC-III) inhibitor | Olezarsen/ DB18728 | Reduce triglyceride levels in adults with familial chylomicronemia syndrome [208]. |
| ATP citrate lyase (ACLY) inhibitor |
Bempedoic acid/ DB11936 | Reduces cholesterol levels, Prevents MI [209]. |
| β-tubulin inhibitor | Colchicine/ DB01394 | Cardiovascular mortality, Coronary revascularization, MI, Stroke [210,211]. |
| Beta-1 adrenergic receptor agonist | Dobutamine/ DB00841 |
Treat cardiac decompensation [212]. |
| Beta-1 adrenergic receptor antagonist | Nebivolol/ DB04861 |
Treat hypertension and aid in the management of heart failure [213]. |
| Bisoprolol/ DB00612 |
Prevent MI and heart failure and treat mild to moderate hypertension [214]. | |
| Carvedilol/ DB01136 |
Treat mild to severe heart failure, left ventricular dysfunction after MI [215]. | |
| Metoprolol/ DB00264 |
Treat heart failure, MI [216]. | |
| Propranolol/ DB00571 |
Used to treat hypertension, MI [217]. | |
| Atenolol/ DB00335 |
Secondary prevention of MI [218]. | |
| Bile acid sequestrant | Cholestyramine/DB01432 | Reduce elevated serum cholesterol in patients with primary hypercholesterolemia [219]. |
| Colesevelam/ DB00930 |
Used to lower LDL-C in adults with hyperlipidemia and pediatric patients with heterozygous familial hypercholesterolemia [220]. | |
| Colestipol/ DB00375 |
Used as an adjunct to diet and exercise to reduce LDL-C cholesterol levels in patients with primary hypercholesterolemia [219]. | |
| COX-1 inhibitor | Aspirin/ DB00945 |
Reducing the risk of major adverse cardiovascular events [221]. |
| Endothelin-1 (ET-1) antagonist | Bosentan/ DB00559 |
Used to treat pulmonary arterial hypertension [222]. |
| Fatty acid binding protein 1 (FABP-1), [Peroxisome proliferator receptor alpha (PPAR-alpha) agonist] |
Fenofibric acid/ DB13873 | Treat severe hypertriglyceridemia, primary hypercholesterolemia, or mixed dyslipidemia [223]. |
| Glucagon-like peptide 1 receptor agonist | Semaglutide/ DB13928 |
Reduces the risk of major adverse cardiovascular events in selected adults [224]. |
| Liraglutide/ DB06655 |
Prevention of cardiovascular complications associated with diabetes and obesity [225]. | |
| Hepatocyte diacylglycerol acyltransferase-2 inhibitor | Niacin/ DB00627 |
Treat hyperlipidemia, dyslipidemia, hypertriglyceridemia, and reduce the risk of MIs [226]. |
| Plasminogen activator | Tenecteplase/ DB00031 |
Used in the emergency treatment of MI [227]. |
| mRNA that codes for apolipoprotein B—100 (apoB-100) | Mipomersen/ DB05528 |
Used for the treatment of homozygous familial hypercholesterolemia [228]. |
| Microsomal triglyceride transfer protein (MTP) inhibitor | Lomitapide/ DB08827 |
Used in homozygous familial hypercholesterolemia (HoFH) patients to reduce low-density lipoprotein cholesterol (LDL-C), total cholesterol (TC), apolipoprotein B (apo B), and non-high-density lipoprotein cholesterol (non-HDL-C) [219]. |
| Mineralocorticoid receptor antagonist | Finerenone/ DB16165 |
Used to treat Cardiovascular mortality, non-fatal MI [229]. |
| Niemann-Pick C1 (NPC1)-like intracellular cholesterol transporter 1 inhibitor, Sterol O-acyltransferase 1 inhibitor |
Ezetimibe/ DB00973 |
Used to lower total cholesterol, LDL-C, Apo-B, and non-HDL-C in primary hyperlipidemia and familial cholesterolemia [230]. |
| Proprotein convertase subtilisin/kexin type 9 (PCSK9) inhibitor | Evolocumab/ DB09303 | Coronary revascularization, MI [231]. |
| Protease-activated receptor 1 (PAR-1) antagonist | Vorapaxar/ DB09030 |
Reducing the number of thrombotic cardiovascular events in patients with a history of MI (MI) or peripheral arterial disease (PAD) [232]. |
| Potassium/sodium hyperpolarization-activated cyclic nucleotide-gated channel 2 inhibitor | Ivabradine/ DB09083 |
Reduce the risk of chronic heart failure [233]. |
| Potentiates antithrombin-III (ATIII) inhibitor | Dalteparin/ DB06779 |
Prophylaxis of Cardiovascular event ischemic complications of unstable angina and non-Q-wave MI [234]. |
| Enoxaparin/ DB01225 |
Prophylaxis of Cardiovascular event ischemic complications of unstable angina and non-Q-wave MI [235]. | |
| Platelet glycoprotein (GP) IIb/IIIa receptor antagonist | Tirofiban/ DB00775 |
Prevents Cardiovascular event [236]. |
| Peroxisome proliferator activated receptor alpha (PPARα) agonist | Fenofibrate/ DB01039 |
Used to lower LDL-C, total-C, triglycerides, and Apo B, while increasing HDL-C in hypercholesterolemia, dyslipidemia, and hypertriglyceridemia [237]. |
| Gemfibrozil/ DB01241 |
Reduction of serum triglyceride levels in high-risk patients with hyperlipidemia [238]. | |
| Peroxisome proliferator activated receptor alpha (PPARα) activator | N-3 fatty acids/ DB11133 |
Prevention of recurrent events after MI in addition to treatment of hypertriglyceridemia [239]. |
| P2Y Purinoceptor 12 antagonist | Ticagrelor/ DB08816 |
Used to lower the rate of cardiovascular events such as heart attack and stroke [240]. |
| Prasugrel/ DB06209 |
Used to reduce risk of thrombotic cardiovascular events in unstable angina or non-ST-elevation MI (NSTEMI) [241]. | |
| Proprotein convertase subtilisin/kexin type 9 (PCSK9) inhibitor | Alirocumab/ DB09302 |
Used as an adjunct to manage heterozygous familial hypercholesterolemia or clinical atherosclerotic cardiovascular disease in patients who require additional lowering of LDL-cholesterol (LDL-C) [242]. |
| Recombinant tissue plasminogen activator (rt-PA) | Alteplase/ DB00009 |
Used for emergency treatment of MI, ischemic stroke [243]. |
| SGLT1 and, or SGLT2 inhibitor | Sotagliflozin/ DB12713 |
Used to treat cardiovascular mortality and heart failure [244]. |
| Type-1 angiotensin II receptor antagonist | Irbesartan/ DB01029 |
Treat congestive heart failure [245]. |
| Transthyretin (TTR) inhibitor | Acoramidis/ DB17999 |
Treatment of Cardiomyopathy caused by transthyretin mediated amyloidosis [246]. |
| 3-hydroxy-3-methylglutaryl coenzyme A (HMG-CoA) reductase inhibitor | Rosuvastatin/ DB01098 | Reduces the risk of CVD including heart attacks and stroke [247]. |
| Lovastatin /DB00227 |
Reduces the risk of CVD including heart attacks and stroke [247]. | |
| Atorvastatin/ DB01076 |
Reduces the risk of CVD including myocardial infarction (MI) and stroke [247]. | |
| Fluvastatin/ DB01095 |
Reduces the risk of CVD including MI and stroke [248]. | |
| Pitavastatin/ DB08860 |
Reduces the risk of CVD including MI and stroke [249]. | |
| Pravastatin/ DB00175 |
Reduces the risk of CVD including MI and stroke [247]. | |
| Simvastatin/ DB00641 |
Used to lower lipid levels and reduce the risk of cardiovascular events including MI and stroke [250]. |
Abbreviations
AA: arachidonic acid; ACC: acetyl-CoA carboxylase; AMPK: AMP-activated protein kinase; ADF: alternate day fasting; ANGPTL: angiopoietin-like protein; ALA: α-linolenic acid; ATP: adenosine triphosphate; ATGL: adipose triglyceride lipase; EVOO: extra virgin olive oil; ETC: electron-transport chain; CHD: coronary heart disease; CPT: carnitine palmitoyltransferase; CL: cardiolipin; CVD: cardiovascular disease; DHA: docosahexaenoic acid; DCM: dilated cardiomyopathy; DRP1: dynamin-related-like protein 1; ER: endoplasmic reticulum; EFA: essential fatty acid; EPA: eicosapentaenoic acid; FATP1: fatty acid transport protein 1; FAT/CD36: fatty acid translocase; FABPpm: plasma membrane fatty acid-binding protein; FABP: fatty acid-binding protein; FAO: fatty acid oxidation; FFAR: free fatty acid receptor; FOXO1: forkhead box protein O1; GPCR: G-protein coupled receptor; GLUT1: Glucose transporter 1; HDAC: histone deacetylase; HFpEF: heart failure with preserved ejection fraction; HFrEF: HFrtEF: heart failure with reduced ejection fraction; HDL: high-density lipoprotein; HSL: hormone-sensitive lipase; IF: intermittent fasting; IHD: ischemic heart disease; KD: ketogenic diet; KLF: Kruppel-like factor; LA: Linolic acid; LDL: low-density lipoprotein; LPL: lipoprotein lipase; LCFA: long-chain fatty acid; MFN1: mitofusin 1; MI: myocardial infarction; MCP-1: monocyte chemoattractant protein-1; MUFA: monounsaturated fatty acid; NEFA: non-esterified fatty acid; NF-κB: nuclear factor kappa-B; OPA1: optic atrophy 1; PUFA: polyunsaturated fatty acid; PPAR: peroxisome proliferator-activated receptor; ROS: reactive oxygen species; SERCA: Sarcoplasmic/Endoplasmic Reticulum Ca2+-ATPase; SIRT: sirtuin; SFA: Saturated fatty acid; SPM: specialized pro-resolving mediator; STAT3: signal transducer and activator of transcription 3; TAG: triglyceride; TFA: trans fatty acid; TLR: toll-like receptor; TG: triglyceride; TNFα: tumor necrosis Factor-alpha; TORC 2: target of Rapamycin complex 2; UPR: unfolded protein response; VLDL: Very low-density lipoprotein; VLSFA: very long-chain SFAs; TRE: Time-restricted eating; RCT: randomized controlled trial;
References
- Duttaroy, A.K.; Mallick, R. Comparative Effects of Fatty Acid and Glucose in Cardiac Remodeling. In Cellular, Molecular, and Environmental Contribution in Cardiac Remodeling; Elsevier, 2024; pp. 89–100.
- Stanley, W.C.; Recchia, F.A.; Lopaschuk, G.D. Myocardial Substrate Metabolism in the Normal and Failing Heart. Physiol. Rev. 2005, 85.
- Spector, A.A.; Yorek, M.A. Membrane Lipid Composition and Cellular Function. J. Lipid Res. 1985, 26.
- Calder, P.C. Marine Omega-3 Fatty Acids and Inflammatory Processes: Effects, Mechanisms and Clinical Relevance. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2015, 1851.
- Mozaffarian, D.; Wu, J.H.Y. Omega-3 Fatty Acids and Cardiovascular Disease: Effects on Risk Factors, Molecular Pathways, and Clinical Events. J. Am. Coll. Cardiol. 2011, 58.
- Elagizi, A.; Lavie, C.J.; O’keefe, E.; Marshall, K.; O’keefe, J.H.; Milani, R. V. An Update on Omega-3 Polyunsaturated Fatty Acids and Cardiovascular Health. Nutrients 2021, 13.
- Yoshida, A.; Sekine, W.; Homma, J.; Sekine, H.; Itoyama, Y.Y.; Sasaki, D.; Matsuura, K.; Kobayashi, E.; Shimizu, T. Development of Appropriate Fatty Acid Formulations to Raise the Contractility of Constructed Myocardial Tissues. Regen. Ther. 2022, 21. [CrossRef]
- Mallick, R.; Basak, S.; Duttaroy, A.K. Fatty Acids and Evolving Roles of Their Proteins in Neurological, Cardiovascular Disorders and Cancers. Prog. Lipid Res. 2021, 83.
- Balta, I.; Stef, L.; Pet, I.; Iancu, T.; Stef, D.; Corcionivoschi, N. Essential Fatty Acids as Biomedicines in Cardiac Health. Biomedicines 2021, 9.
- Listenberger, L.L.; Han, X.; Lewis, S.E.; Cases, S.; Farese, R. V.; Ory, D.S.; Schaffer, J.E. Triglyceride Accumulation Protects against Fatty Acid-Induced Lipotoxicity. Proceedings of the National Academy of Sciences 2003, 100, 3077–3082. [CrossRef]
- Annevelink, C.E.; Sapp, P.A.; Petersen, K.S.; Shearer, G.C.; Kris-Etherton, P.M. Diet-Derived and Diet-Related Endogenously Produced Palmitic Acid: Effects on Metabolic Regulation and Cardiovascular Disease Risk. J. Clin. Lipidol. 2023, 17.
- Glass, C.K.; Olefsky, J.M. Inflammation and Lipid Signaling in the Etiology of Insulin Resistance. Cell Metab. 2012, 15.
- Emken, E. Metabolism of Dietary Stearic Acid Relative to Other Fatty Acids in Human Subjects. Am. J. Clin. Nutr. 1994, 60, 1023S-1028S. [CrossRef]
- BRUCE, J.S.; SALTER, A.M. Metabolic Fate of Oleic Acid, Palmitic Acid and Stearic Acid in Cultured Hamster Hepatocytes. Biochemical Journal 1996, 316, 847–852. [CrossRef]
- van Rooijen, M.; Mensink, R. Palmitic Acid Versus Stearic Acid: Effects of Interesterification and Intakes on Cardiometabolic Risk Markers—A Systematic Review. Nutrients 2020, 12, 615. [CrossRef]
- Shahidi, F.; Senanayake, S.P.J.N. Fatty Acids. In International Encyclopedia of Public Health; Elsevier, 2008; pp. 594–603.
- Fan, H.; Wang, Y.; Ren, Z.; Liu, X.; Zhao, J.; Yuan, Y.; Fei, X.; Song, X.; Wang, F.; Liang, B. Mediterranean Diet Lowers All-Cause and Cardiovascular Mortality for Patients with Metabolic Syndrome. Diabetol. Metab. Syndr. 2023, 15, 107. [CrossRef]
- Gonzalez-Soto, M.; Mutch, D.M. Diet Regulation of Long-Chain PUFA Synthesis: Role of Macronutrients, Micronutrients, and Polyphenols on Δ-5/Δ-6 Desaturases and Elongases 2/5. Advances in Nutrition 2021, 12.
- Salsinha, A.S.; Socodato, R.; Relvas, J.B.; Pintado, M. The Pro- and Antiinflammatory Activity of Fatty Acids. In Bioactive Lipids; Elsevier, 2023; pp. 51–75.
- Mohebi-Nejad, A.; Bikdeli, B. Omega-3 Supplements and Cardiovascular Diseases. Tanaffos 2014, 13, 6–14.
- Masud Parvez, G.M.; Akanda, K.M. Foods and Arthritis: An Overview. In Bioactive Food as Dietary Interventions for Arthritis and Related Inflammatory Diseases; Elsevier, 2019; pp. 3–22.
- Wolf, C.; Steller, R.N. Introduction to the Nutrients and Their Association with Common Gastrointestinal Disorders. Physician Assist. Clin. 2022, 7.
- Poli, A.; Agostoni, C.; Visioli, F. Dietary Fatty Acids and Inflammation: Focus on the n-6 Series. Int. J. Mol. Sci. 2023, 24.
- Anderson, B.M.; Ma, D.W.L. Are All N-3 Polyunsaturated Fatty Acids Created Equal? Lipids Health Dis. 2009, 8.
- Djuricic, I.; Calder, P.C. N-3 Fatty Acids (EPA and DHA) and Cardiovascular Health—Updated Review of Mechanisms and Clinical Outcomes. Curr. Atheroscler. Rep. 2025, 27.
- Delgado, G.E.; Kleber, M.E. Trans Fatty Acids and Mortality. In The Molecular Nutrition of Fats; Elsevier, 2019; pp. 335–345.
- Gyamfi, D.; Ofori Awuah, E.; Owusu, S. Classes, Nomenclature, and Functions of Lipids and Lipid-Related Molecules and the Dietary Lipids. In The Molecular Nutrition of Fats; Elsevier, 2019; pp. 3–16.
- Da Dalt, L.; Cabodevilla, A.G.; Goldberg, I.J.; Norata, G.D. Cardiac Lipid Metabolism, Mitochondrial Function, and Heart Failure. Cardiovasc. Res. 2023, 119.
- O’Connell, T.D.; Mason, R.P.; Budoff, M.J.; Navar, A.M.; Shearer, G.C. Mechanistic Insights into Cardiovascular Protection for Omega-3 Fatty Acids and Their Bioactive Lipid Metabolites. European Heart Journal, Supplement 2020, 22.
- Dufau, J.; Recazens, E.; Bottin, L.; Bergoglio, C.; Mairal, A.; Chaoui, K.; Marques, M.A.; Jimenez, V.; García, M.; Wang, T.; et al. Nuclear Hormone-Sensitive Lipase Regulates Adipose Tissue Mass and Adipocyte Metabolism. Cell Metab. 2025, 37. [CrossRef]
- Herbst, E.A.F.; Paglialunga, S.; Gerling, C.; Whitfield, J.; Mukai, K.; Chabowski, A.; Heigenhauser, G.J.F.; Spriet, L.L.; Holloway, G.P. Omega-3 Supplementation Alters Mitochondrial Membrane Composition and Respiration Kinetics in Human Skeletal Muscle. Journal of Physiology 2014, 592. [CrossRef]
- Abumrad, N.A.; Goldberg, I.J. CD36 Actions in the Heart: Lipids, Calcium, Inflammation, Repair and More? Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2016, 1861.
- Bharadwaj, K.G.; Hiyama, Y.; Hu, Y.; Huggins, L.A.; Ramakrishnan, R.; Abumrad, N.A.; Shulman, G.I.; Blaner, W.S.; Goldberg, I.J. Chylomicron- and VLDL-Derived Lipids Enter the Heart through Different Pathways: In Vivo Evidence for Receptor- and Non-Receptor-Mediated Fatty Acid Uptake. Journal of Biological Chemistry 2010, 285. [CrossRef]
- Hames, K.C.; Vella, A.; Kemp, B.J.; Jensen, M.D. Free Fatty Acid Uptake in Humans with CD36 Deficiency. Diabetes 2014, 63. [CrossRef]
- Fillmore, N.; Hou, V.; Sun, J.; Springer, D.; Murphy, E. Cardiac Specific Knock-down of Peroxisome Proliferator Activated Receptor α Prevents Fasting-Induced Cardiac Lipid Accumulation and Reduces Perilipin 2. PLoS One 2022, 17. [CrossRef]
- Prosdocimo, D.A.; John, J.E.; Zhang, L.; Efraim, E.S.; Zhang, R.; Liao, X.; Jain, M.K. KLF15 and PPAR α Cooperate to Regulate Cardiomyocyte Lipid Gene Expression and Oxidation. PPAR Res. 2015, 2015. [CrossRef]
- Tien, Y.T.; Chang, M.H.; Chu, P.Y.; Lin, C.S.; Liu, C.H.; Liao, A.T. Downregulation of the KLF4 Transcription Factor Inhibits the Proliferation and Migration of Canine Mammary Tumor Cells. Veterinary Journal 2015, 205. [CrossRef]
- Drosatos, K.; Pollak, N.M.; Pol, C.J.; Ntziachristos, P.; Willecke, F.; Valenti, M.C.; Trent, C.M.; Hu, Y.; Guo, S.; Aifantis, I.; et al. Cardiac Myocyte KLF5 Regulates Ppara Expression and Cardiac Function. Circ. Res. 2016, 118. [CrossRef]
- Kucharski, M.; Kaczor, U. PPARα and PPARγ as Main Regulators of Fatty Acid Metabolism. Postepy Hig. Med. Dosw. 2018, 72.
- Savarese, G.; Becher, P.M.; Lund, L.H.; Seferovic, P.; Rosano, G.M.C.; Coats, A.J.S. Global Burden of Heart Failure: A Comprehensive and Updated Review of Epidemiology. Cardiovasc. Res. 2022, 118.
- Li, Z.; Lei, H.; Jiang, H.; Fan, Y.; Shi, J.; Li, C.; Chen, F.; Mi, B.; Ma, M.; Lin, J.; et al. Saturated Fatty Acid Biomarkers and Risk of Cardiometabolic Diseases: A Meta-Analysis of Prospective Studies. Front. Nutr. 2022, 9.
- Maki, K.C.; Dicklin, M.R.; Kirkpatrick, C.F. Saturated Fats and Cardiovascular Health: Current Evidence and Controversies. J. Clin. Lipidol. 2021, 15.
- Elnwasany, A.; Ewida, H.A.; Menendez-Montes, I.; Mizerska, M.; Fu, X.; Kim, C.-W.; Horton, J.D.; Burgess, S.C.; Rothermel, B.A.; Szweda, P.A.; et al. Reciprocal Regulation of Cardiac β-Oxidation and Pyruvate Dehydrogenase by Insulin. Journal of Biological Chemistry 2024, 300, 107412. [CrossRef]
- Yarmohammadi, F.; Hayes, A.W.; Karimi, G. Natural and Chemical Compounds as Protective Agents against Cardiac Lipotoxicity. Biomedicine and Pharmacotherapy 2022, 145.
- Silva-Soto, M.; Carrillo-Fernández, P.; Saez Lancellotti, E.; Medina-Jiménez, E.; Mogaburo Alba, J.; Catena-Granados, N.; López-Carmona, M.; Pérez-Belmonte, L.; Prieto Lain, N.; Gómez Hernández, A.; et al. Extra Virgin Olive Oil Phenolic Compounds: Modulating Mitochondrial Function and Protecting Against Chronic Diseases—A Narrative Review. Nutrients 2025, 17, 1443. [CrossRef]
- Seifi, K.; Rezaei, M.; Yansari, A.T.; Zamiri, M.J.; Riazi, G.H.; Heidari, R. Short Chain Fatty Acids May Improve Hepatic Mitochondrial Energy Efficiency in Heat Stressed-Broilers. J. Therm. Biol. 2020, 89. [CrossRef]
- Endo, J.; Arita, M. Cardioprotective Mechanism of Omega-3 Polyunsaturated Fatty Acids. J. Cardiol. 2016, 67.
- Mallick, R.; Basak, S.; Duttaroy, A.K. Docosahexaenoic Acid,22:6n-3: Its Roles in the Structure and Function of the Brain. International Journal of Developmental Neuroscience 2019, 79. [CrossRef]
- Kang, J.X. Reduction of Heart Rate by Omega-3 Fatty Acids and the Potential Underlying Mechanisms. Front. Physiol. 2012, 3 OCT.
- Matacchione, G.; Piacenza, F.; Pimpini, L.; Rosati, Y.; Marcozzi, S. The Role of the Gut Microbiota in the Onset and Progression of Heart Failure: Insights into Epigenetic Mechanisms and Aging. Clin. Epigenetics 2024, 16, 175. [CrossRef]
- Macdonald-Ramos, K.; Martínez-Ibarra, A.; Monroy, A.; Miranda-Ríos, J.; Cerbón, M. Effect of Dietary Fatty Acids on MicroRNA Expression Related to Metabolic Disorders and Inflammation in Human and Animal Trials. Nutrients 2021, 13, 1830. [CrossRef]
- Ahn, S.; Kaipparettu, B.A. G-Protein Coupled Receptors in Metabolic Reprogramming and Cancer. Pharmacol. Ther. 2025, 270, 108849. [CrossRef]
- Kaemmerer, E.; Plum, P.; Klaus, C.; Weiskirchen, R.; Liedtke, C.; Adolf, M.; Schippers, A.; Wagner, N.; Reinartz, A.; Gassler, N. Fatty Acid Binding Receptors in Intestinal Physiology and Pathophysiology. World J. Gastrointest. Pathophysiol. 2010, 1, 147–153. [CrossRef]
- Fredriksson, R.; Höglund, P.J.; Gloriam, D.E.I.; Lagerström, M.C.; Schiöth, H.B. Seven Evolutionarily Conserved Human Rhodopsin G Protein-Coupled Receptors Lacking Close Relatives. FEBS Lett. 2003, 554, 381–388. [CrossRef]
- Hirasawa, A.; Tsumaya, K.; Awaji, T.; Katsuma, S.; Adachi, T.; Yamada, M.; Sugimoto, Y.; Miyazaki, S.; Tsujimoto, G. Free Fatty Acids Regulate Gut Incretin Glucagon-like Peptide-1 Secretion through GPR120. Nat. Med. 2005, 11, 90–94. [CrossRef]
- Davie, J.R. Inhibition of Histone Deacetylase Activity by Butyrate. Journal of Nutrition 2003, 133. [CrossRef]
- Wu, Y.; Dong, Y.; Song, P.; Zou, M.H. Activation of the AMP-Activated Protein Kinase (AMPK) by Nitrated Lipids in Endothelial Cells. PLoS One 2012, 7, e31056. [CrossRef]
- Canbolat, E.; Cakıroglu, F.P. The Importance of AMPK in Obesity and Chronic Diseases and the Relationship of AMPK with Nutrition: A Literature Review. Crit. Rev. Food Sci. Nutr. 2023, 63, 449–456. [CrossRef]
- Monsalve, F.A.; Pyarasani, R.D.; Delgado-Lopez, F.; Moore-Carrasco, R. Peroxisome Proliferator-Activated Receptor Targets for the Treatment of Metabolic Diseases. Mediators Inflamm. 2013, 2013. [CrossRef]
- Kota, B.P.; Huang, T.H.W.; Roufogalis, B.D. An Overview on Biological Mechanisms of PPARs. Pharmacol. Res. 2005, 51, 85–94. [CrossRef]
- Shu, L.; Chan, A.; Wells, R.A. Cross-Talk between PPARs and the Partners of RXR: A Molecular Perspective. 2009, 9. [CrossRef]
- König, B.; Koch, A.; Spielmann, J.; Hilgenfeld, C.; Stangl, G.I.; Eder, K. Activation of PPARα Lowers Synthesis and Concentration of Cholesterol by Reduction of Nuclear SREBP-2. Biochem. Pharmacol. 2007, 73, 574–585. [CrossRef]
- Steinberg, G.R.; Hardie, D.G. New Insights into Activation and Function of the AMPK. Nature Reviews Molecular Cell Biology 2022 24:4 2022, 24, 255–272. [CrossRef]
- Jung, E.J.; Kwon, S.W.; Jung, B.H.; Oh, S.H.; Lee, B.H. Role of the AMPK/SREBP-1 Pathway in the Development of Orotic Acid-Induced Fatty Liver. J. Lipid Res. 2011, 52, 1617–1625. [CrossRef]
- Li, N.; Li, X.; Ding, Y.; Liu, X.; Diggle, K.; Kisseleva, T.; Brenner, D.A. SREBP Regulation of Lipid Metabolism in Liver Disease, and Therapeutic Strategies. Biomedicines 2023, Vol. 11, Page 3280 2023, 11, 3280. [CrossRef]
- Myers, M.N.; Chirivi, M.; dos Santos Neto, J.M.; Parales-Girón, J.; Worden, L.C.; Lock, A.L.; Contreras, G.A. Dynamics of Oxylipin Biosynthesis in Systemic Inflammation: Insights from a Large Animal Model of Endotoxemia. Front. Immunol. 2025, 16. [CrossRef]
- Verdin, E.; Hirschey, M.D.; Finley, L.W.S.; Haigis, M.C. Sirtuin Regulation of Mitochondria: Energy Production, Apoptosis, and Signaling. Trends Biochem. Sci. 2010, 35, 669–675. [CrossRef]
- Singh, C.K.; Chhabra, G.; Ndiaye, M.A.; Garcia-Peterson, L.M.; MacK, N.J.; Ahmad, N. The Role of Sirtuins in Antioxidant and Redox Signaling. Antioxid. Redox Signal. 2018, 28, 643–661. [CrossRef]
- Han, Y.; Zhou, S.; Coetzee, S.; Chen, A. Sirt4 and Its Roles in Energy and Redox Metabolism in Health, Disease and during Exercise. Front. Physiol. 2019, 10. [CrossRef]
- Purushotham, A.; Schug, T.T.; Li, X. SIRT1 Performs a Balancing Act on the Tight-Rope toward Longevity. Aging 2009, 1, 669–673. [CrossRef]
- Picard, F.; Kurtev, M.; Chung, N.; Topark-Ngarm, A.; Senawong, T.; De Oliveira, R.M.; Leid, M.; McBurney, M.W.; Guarente, L. Sirt1 Promotes Fat Mobilization in White Adipocytes by Repressing PPAR-γ. Nature 2004, 429, 771–776. [CrossRef]
- Walker, A.K.; Yang, F.; Jiang, K.; Ji, J.Y.; Watts, J.L.; Purushotham, A.; Boss, O.; Hirsch, M.L.; Ribich, S.; Smith, J.J.; et al. Conserved Role of SIRT1 Orthologs in Fasting-Dependent Inhibition of the Lipid/Cholesterol Regulator SREBP. Genes Dev. 2010, 24. [CrossRef]
- Lim, J.H.; Gerhart-Hines, Z.; Dominy, J.E.; Lee, Y.; Kim, S.; Tabata, M.; Xiang, Y.K.; Puigserver, P. Oleic Acid Stimulates Complete Oxidation of Fatty Acids through Protein Kinase A-Dependent Activation of SIRT1-PGC1α Complex. Journal of Biological Chemistry 2013, 288, 7117–7126. [CrossRef]
- Son, S.H.; Lee, S.M.; Lee, M.H.; Son, Y.K.; Kim, S.E.; An, W.S. Omega-3 Fatty Acids Upregulate SIRT1/3, Activate PGC-1α via Deacetylation, and Induce Nrf1 Production in 5/6 Nephrectomy Rat Model. Mar. Drugs 2021, 19, 182. [CrossRef]
- Tornatore, L.; Thotakura, A.K.; Bennett, J.; Moretti, M.; Franzoso, G. The Nuclear Factor Kappa B Signaling Pathway: Integrating Metabolism with Inflammation. Trends Cell Biol. 2012, 22, 557–566. [CrossRef]
- Yang, X.; Zhang, X.; Heckmann, B.L.; Lu, X.; Liu, J. Relative Contribution of Adipose Triglyceride Lipase and Hormone-Sensitive Lipase to Tumor Necrosis Factor-α (TNF-α)-Induced Lipolysis in Adipocytes. Journal of Biological Chemistry 2011, 286, 40477–40485. [CrossRef]
- Edwards, M.; Mohiuddin, S.S. Biochemistry, Lipolysis. StatPearls 2023.
- Cho, Y.Y.; Kim, S.; Kim, P.; Jo, M.J.; Park, S.E.; Choi, Y.; Jung, S.M.; Kang, H.J. G-Protein-Coupled Receptor (GPCR) Signaling and Pharmacology in Metabolism: Physiology, Mechanisms, and Therapeutic Potential. Biomolecules 2025, Vol. 15, Page 291 2025, 15, 291. [CrossRef]
- Gao, H.; Tian, K.; Feng, X.; Yan, M.; Gao, C.; Jiang, Y.; Zhu, C.; Zhu, H.; Liu, X.; Peng, Y. Free Fatty Acid Receptor 2 Promotes Cardiomyocyte Hypertrophy by Activating STAT3 and GATA4. Food Science and Human Wellness 2022, 11, 405–417. [CrossRef]
- Das, M.; Das, D.K. Lipid Raft in Cardiac Health and Disease. Curr. Cardiol. Rev. 2009, 5, 105. [CrossRef]
- O’Connell, K.M.S.; Martens, J.R.; Tamkun, M.M. Localization of Ion Channels to Lipid Raft Domains within the Cardiovascular System. Trends Cardiovasc. Med. 2004, 14, 37–42. [CrossRef]
- Maguy, A.; Hebert, T.E.; Nattel, S. Involvement of Lipid Rafts and Caveolae in Cardiac Ion Channel Function. Cardiovasc. Res. 2006, 69, 798–807. [CrossRef]
- Epand, R.M.; Epand, R.F.; Ahmed, N.; Chen, R. Promotion of Hexagonal Phase Formation and Lipid Mixing by Fatty Acids with Varying Degrees of Unsaturation. Chem. Phys. Lipids 1991, 57, 75–80. [CrossRef]
- Cooper, G.M. Cell Membranes. 2000.
- Ibarguren, M.; López, D.J.; Escribá, P. V. The Effect of Natural and Synthetic Fatty Acids on Membrane Structure, Microdomain Organization, Cellular Functions and Human Health. Biochimica et Biophysica Acta (BBA)—Biomembranes 2014, 1838, 1518–1528. [CrossRef]
- Fearnley, C.J.; Llewelyn Roderick, H.; Bootman, M.D. Calcium Signaling in Cardiac Myocytes. Cold Spring Harb. Perspect. Biol. 2011, 3, a004242. [CrossRef]
- Zhihao, L.; Jingyu, N.; Lan, L.; Michael, S.; Rui, G.; Xiyun, B.; Xiaozhi, L.; Guanwei, F. SERCA2a: A Key Protein in the Ca2+ Cycle of the Heart Failure. Heart Fail. Rev. 2020, 25, 523–535. [CrossRef]
- Giroud, S.; Frare, C.; Strijkstra, A.; Boerema, A.; Arnold, W.; Ruf, T. Membrane Phospholipid Fatty Acid Composition Regulates Cardiac SERCA Activity in a Hibernator, the Syrian Hamster (Mesocricetus Auratus). PLoS One 2013, 8. [CrossRef]
- Li, Y.; Ge, M.; Ciani, L.; Kuriakose, G.; Westover, E.J.; Dura, M.; Covey, D.F.; Freed, J.H.; Maxfield, F.R.; Lytton, J.; et al. Enrichment of Endoplasmic Reticulum with Cholesterol Inhibits Sarcoplasmic-Endoplasmic Reticulum Calcium ATPase-2b Activity in Parallel with Increased Order of Membrane Lipids: Implications for Depletion of Endoplasmic Reticulum Calcium Stores and Apoptosis in Cholesterol-Loaded Macrophages. Journal of Biological Chemistry 2004, 279, 37030–37039. [CrossRef]
- Yamamoto, T.; Endo, J.; Kataoka, M.; Matsuhashi, T.; Katsumata, Y.; Shirakawa, K.; Yoshida, N.; Isobe, S.; Moriyama, H.; Goto, S.; et al. Decrease in Membrane Phospholipids Unsaturation Correlates with Myocardial Diastolic Dysfunction. PLoS One 2018, 13, e0208396. [CrossRef]
- Asp, M.L.; Martindale, J.J.; Heinis, F.I.; Wang, W.; Metzger, J.M. Calcium Mishandling in Diastolic Dysfunction: Mechanisms and Potential Therapies. Biochimica et Biophysica Acta (BBA)—Molecular Cell Research 2013, 1833, 895–900. [CrossRef]
- Rodríguez-Rodríguez, R.; Fosch, A.; Garcia-Chica, J.; Zagmutt, S.; Casals, N. Targeting Carnitine Palmitoyltransferase 1 Isoforms in the Hypothalamus: A Promising Strategy to Regulate Energy Balance. J. Neuroendocrinol. 2023, 35, e13234. [CrossRef]
- Lopaschuk, G.D.; Ussher, J.R.; Folmes, C.D.L.; Jaswal, J.S.; Stanley, W.C. Myocardial Fatty Acid Metabolism in Health and Disease. Physiol. Rev. 2010, 90, 207–258. [CrossRef]
- Tirosh, O.; Ilan, E.; Anavi, S.; Ramadori, G.; Madar, Z. Nutritional Lipid-Induced Oxidative Stress Leads to Mitochondrial Dysfunction Followed by Necrotic Death in FaO Hepatocytes. Nutrition 2009, 25, 200–208. [CrossRef]
- Jain, A.; Casanova, D.; Padilla, A.V.; Paniagua Bojorges, A.; Kotla, S.; Ko, K.A.; Samanthapudi, V.S.K.; Chau, K.; Nguyen, M.T.H.; Wen, J.; et al. Premature Senescence and Cardiovascular Disease Following Cancer Treatments: Mechanistic Insights. Front. Cardiovasc. Med. 2023, 10. [CrossRef]
- Heshmati, J.; Morvaridzadeh, M.; Maroufizadeh, S.; Akbari, A.; Yavari, M.; Amirinejad, A.; Maleki-Hajiagha, A.; Sepidarkish, M. Omega-3 Fatty Acids Supplementation and Oxidative Stress Parameters: A Systematic Review and Meta-Analysis of Clinical Trials. Pharmacol. Res. 2019, 149, 104462. [CrossRef]
- Li, Q.; Yu, Q.; Na, R.; Liu, B. Omega-3 Polyunsaturated Fatty Acids Prevent Murine Dilated Cardiomyopathy by Reducing Oxidative Stress and Cardiomyocyte Apoptosis. Exp. Ther. Med. 2017, 14, 6152. [CrossRef]
- Kyriakoudi, S.; Drousiotou, A.; Petrou, P.P. When the Balance Tips: Dysregulation of Mitochondrial Dynamics as a Culprit in Disease. Int. J. Mol. Sci. 2021, 22, 4617. [CrossRef]
- Smirnova, E.; Griparic, L.; Shurland, D.L.; Van der Bliek, A.M. Dynamin-Related Protein Drp1 Is Required for Mitochondrial Division in Mammalian Cells. Mol. Biol. Cell 2001, 12, 2245–2256. [CrossRef]
- Santel, A.; Fuller, M.T. Control of Mitochondrial Morphology by a Human Mitofusin. J. Cell Sci. 2001, 114, 867–874. [CrossRef]
- Chen, H.; Detmer, S.A.; Ewald, A.J.; Griffin, E.E.; Fraser, S.E.; Chan, D.C. Mitofusins Mfn1 and Mfn2 Coordinately Regulate Mitochondrial Fusion and Are Essential for Embryonic Development. Journal of Cell Biology 2003, 160, 189–200. [CrossRef]
- Hoppins, S.; Lackner, L.; Nunnari, J. The Machines That Divide and Fuse Mitochondria. Annu. Rev. Biochem. 2007, 76, 751–780. [CrossRef]
- Sherratt, S.C.R.; Juliano, R.A.; Copland, C.; Bhatt, D.L.; Libby, P.; Mason, R.P. EPA and DHA Containing Phospholipids Have Contrasting Effects on Membrane Structure. J. Lipid Res. 2021, 62, 100106. [CrossRef]
- Pal, A.; Metherel, A.H.; Fiabane, L.; Buddenbaum, N.; Bazinet, R.P.; Shaikh, S.R. Do Eicosapentaenoic Acid and Docosahexaenoic Acid Have the Potential to Compete against Each Other? Nutrients 2020, 12, 1–12. [CrossRef]
- Fuentes, J.; Morcillo, P. The Role of Cardiolipin in Mitochondrial Function and Neurodegenerative Diseases. Cells 2024, 13, 609. [CrossRef]
- Lin, J.; Duan, J.; Wang, Q.; Xu, S.; Zhou, S.; Yao, K. Mitochondrial Dynamics and Mitophagy in Cardiometabolic Disease. Front. Cardiovasc. Med. 2022, 9.
- Lemaitre, R.N.; King, I.B. Very Long-Chain Saturated Fatty Acids and Diabetes and Cardiovascular Disease. Curr. Opin. Lipidol. 2022, 33.
- Wang, C.; Han, D.; Feng, X.; Hu, L.; Wu, J. Docosahexaenoic Acid Alleviates LPS-Induced Cytotoxicity in HL-1 Cardiac Cells via Improving Stress-Induced Mitochondrial Fragmentation. Heliyon 2023, 9. [CrossRef]
- Elezaby, A.; Sverdlov, A.L.; Tu, V.H.; Soni, K.; Luptak, I.; Qin, F.; Liesa, M.; Shirihai, O.S.; Rimer, J.; Schaffer, J.E.; et al. Mitochondrial Remodeling in Mice with Cardiomyocyte-Specific Lipid Overload. J. Mol. Cell. Cardiol. 2015, 79, 275–283. [CrossRef]
- Tsushima, K.; Bugger, H.; Wende, A.R.; Soto, J.; Jenson, G.A.; Tor, A.R.; McGlauflin, R.; Kenny, H.C.; Zhang, Y.; Souvenir, R.; et al. Mitochondrial Reactive Oxygen Species in Lipotoxic Hearts Induces Post-Translational Modifications of AKAP121, DRP1 and OPA1 That Promote Mitochondrial Fission. Circ. Res. 2017, 122, 58. [CrossRef]
- Gao, J.; Li, C.; Chen, H.; Li, Z.-H.; You, F.-F.; Song, W.-Q.; Zhong, W.-F.; Chen, P.-L.; Yang, J.; Huang, Q.-M.; et al. Effect of Meal-Timing on the Association of Unsaturated Fatty Acids with All-Cause and Cardiovascular Mortality among Adults: A Prospective Cohort Study with 10-Year Follow-Up. Nutrients 2024, 16, 2071. [CrossRef]
- Manoogian, E.N.C.; Zadourian, A.; Lo, H.C.; Gutierrez, N.R.; Shoghi, A.; Rosander, A.; Pazargadi, A.; Ormiston, C.K.; Wang, X.; Sui, J.; et al. Feasibility of Time-Restricted Eating and Impacts on Cardiometabolic Health in 24-h Shift Workers: The Healthy Heroes Randomized Control Trial. Cell Metab. 2022, 34, 1442-1456.e7. [CrossRef]
- Pakhomov, N.; Baugh, J.A. The Role of Diet-Derived Short-Chain Fatty Acids in Regulating Cardiac Pressure Overload. Am. J. Physiol. Heart Circ. Physiol. 2021, 320.
- Miao, H.; Li, B.; Wang, Z.; Mu, J.; Tian, Y.; Jiang, B.; Zhang, S.; Gong, X.; Shui, G.; Lam, S.M. Lipidome Atlas of the Developing Heart Uncovers Dynamic Membrane Lipid Attributes Underlying Cardiac Structural and Metabolic Maturation. Research 2022, 2022. [CrossRef]
- Finck, B.N.; Kelly, D.P. Peroxisome Proliferator-Activated Receptor α (PPARα) Signaling in the Gene Regulatory Control of Energy Metabolism in the Normal and Diseased Heart. J. Mol. Cell. Cardiol. 2002, 34, 1249–1257. [CrossRef]
- Kimura, I.; Ichimura, A.; Ohue-Kitano, R.; Igarashi, M. Free Fatty Acid Receptors in Health and Disease. Physiol. Rev. 2020, 100, 171–210. [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-ΚB Signaling in Inflammation. Signal Transduct. Target. Ther. 2017, 2, 1–9. [CrossRef]
- Beauloye, V.; Zech, F.; Mong, H.T.T.; Clapuyt, P.; Maes, M.; Brichard, S.M. Determinants of Early Atherosclerosis in Obese Children and Adolescents. Journal of Clinical Endocrinology and Metabolism 2007, 92. [CrossRef]
- Jaca, A.; Durão, S.; Harbron, J. Omega-3 Fatty Acids for the Primary and Secondary Prevention of Cardiovascular Disease. South African Medical Journal 2020, 110. [CrossRef]
- Eclov, J.A.; Qian, Q.; Redetzke, R.; Chen, Q.; Wu, S.C.; Healy, C.L.; Ortmeier, S.B.; Harmon, E.; Shearer, G.C.; O’Connell, T.D. EPA, Not DHA, Prevents Fibrosis in Pressure Overload-Induced Heart Failure: Potential Role of Free Fatty Acid Receptor 4. J. Lipid Res. 2015, 56. [CrossRef]
- Kain, V.; Ingle, K.A.; Colas, R.A.; Dalli, J.; Prabhu, S.D.; Serhan, C.N.; Joshi, M.; Halade, G. V. Resolvin D1 Activates the Inflammation Resolving Response at Splenic and Ventricular Site Following Myocardial Infarction Leading to Improved Ventricular Function. J. Mol. Cell. Cardiol. 2015, 84. [CrossRef]
- Lipsky, P.E.; Brooks, P.; Crofford, L.J.; DuBois, R.; Graham, D.; Simon, L.S.; Van De Putte, L.B.A.; Abramson, S.B. Unresolved Issues in the Role of Cyclooxygenase-2 in Normal Physiologic Processes and Disease. Arch. Intern. Med. 2000, 160.
- Cheng, Y.; Austin, S.C.; Rocca, B.; Koller, B.H.; Coffman, T.M.; Grosser, T.; Lawson, J.A.; FitzGerald, G.A. Role of Prostacyclin in the Cardiovascular Response to Thromboxane A2. Science (1979). 2002, 296. [CrossRef]
- Mehta, J.; Mehta, P.; Feldman, R.L. Severe Intracoronary Thromboxane Release Preceding Acute Coronary Artery Occlusion. Prostaglandins Leukot. Med. 1982, 8.
- Barone, F.C.; Schmidt, D.B.; Hillegass, L.M.; Price, W.J.; White, R.F.; Feuerstein, G.Z.; Clark, R.K.; Lee, E. V.; Griswold, D.E.; Sarau, H.M. Reperfusion Increases Neutrophils and Leukotriene B4 Receptor Binding in Rat Focal Ischemia. Stroke 1992, 23. [CrossRef]
- Kadletz, M.; Dignan, R.J.; Loesser, K.E.; Hess, M.L.; Wechsler, A.S. Ischemia and Activated Neutrophils Alter Coronary Microvascular but Not Epicardial Coronary Artery Reactivity. Journal of Thoracic and Cardiovascular Surgery 1994, 108. [CrossRef]
- Zhang, P.; Wang, J.; Wang, X.; Wang, L.; Xu, S.; Gong, P. Protectin D1 Alleviates Myocardial Ischemia/Reperfusion Injury by Regulating PI3K/AKT Signaling Pathway. J. Cardiovasc. Transl. Res. 2024, 17. [CrossRef]
- Li, X.W.; Li, C.L.; Liang, W.D.; Bi, Y.T.; Chen, M.H.; Dong, S. Protectin D1 Promotes Resolution of Inflammation in a Murine Model of Lipopolysaccharide-Induced Acute Lung Injury via Enhancing Neutrophil Apoptosis. Chin. Med. J. (Engl). 2014, 127. [CrossRef]
- Colas, R.A.; Shinohara, M.; Dalli, J.; Chiang, N.; Serhan, C.N. Identification and Signature Profiles for Pro-Resolving and Inflammatory Lipid Mediators in Human Tissue. Am. J. Physiol. Cell Physiol. 2014, 307. [CrossRef]
- Friedrich, E.B.; Tager, A.M.; Liu, E.; Pettersson, A.; Owman, C.; Munn, L.; Luster, A.D.; Gerszten, R.E. Mechanisms of Leukotriene B4-Triggered Monocyte Adhesion. Arterioscler. Thromb. Vasc. Biol. 2003, 23. [CrossRef]
- Hara, T.; Sata, M. Roles of Perivascular Adipose Tissue in the Pathogenesis of Atherosclerosis ― an Update on Recent Findings. Front. Physiol. 2024, 15.
- Wahrburg, U. What Are the Health Effects of Fat? Eur. J. Nutr. 2004, 43 Suppl 1. [CrossRef]
- WHITE, B. Dietary Fatty Acids. Am. Fam. Physician 2009, 80, 345–350.
- Hu, F.B.; Stampfer, M.J.; Manson, J.A.E.; Ascherio, A.; Colditz, G.A.; Speizer, F.E.; Hennekens, C.H.; Willett, W.C. Dietary Saturated Fats and Their Food Sources in Relation to the Risk of Coronary Heart Disease in Women. Am. J. Clin. Nutr. 1999, 70, 1001–1008. [CrossRef]
- Lemaitre, R.N.; King, I.B. Very Long-Chain Saturated Fatty Acids and Diabetes and Cardiovascular Disease. Curr. Opin. Lipidol. 2022, 33, 76–82. [CrossRef]
- Steur, M.; Johnson, L.; Sharp, S.J.; Imamura, F.; Sluijs, I.; Key, T.J.; Wood, A.; Chowdhury, R.; Guevara, M.; Jakobsen, M.U.; et al. Dietary Fatty Acids, Macronutrient Substitutions, Food Sources and Incidence of Coronary Heart Disease: Findings from the EPIC-CVD Case-Cohort Study across Nine European Countries. J. Am. Heart Assoc. 2021, 10, 19814. [CrossRef]
- Praagman, J.; De Jonge, E.A.L.; Kiefte-De Jong, J.C.; Beulens, J.W.J.; Sluijs, I.; Schoufour, J.D.; Hofman, A.; Van Der Schouw, Y.T.; Franco, O.H. Dietary Saturated Fatty Acids and Coronary Heart Disease Risk in a Dutch Middle-Aged and Elderly Population. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 2011–2018. [CrossRef]
- Unhapipatpong, C.; Shantavasinkul, P.C.; Kasemsup, V.; Siriyotha, S.; Warodomwichit, D.; Maneesuwannarat, S.; Vathesatogkit, P.; Sritara, P.; Thakkinstian, A. Tropical Oil Consumption and Cardiovascular Disease: An Umbrella Review of Systematic Reviews and Meta Analyses. Nutrients 2021, 13.
- Valk, R.; Hammill, J.; Grip, J. Saturated Fat: Villain and Bogeyman in the Development of Cardiovascular Disease? Eur. J. Prev. Cardiol. 2022, 29, 2312–2321. [CrossRef]
- Rucker, D.; Dhamoon, A.S. Physiology, Thromboxane A2; 2019;
- Wone, B.W.M.; Donovan, E.R.; Cushman, J.C.; Hayes, J.P. Metabolic Rates Associated with Membrane Fatty Acids in Mice Selected for Increased Maximal Metabolic Rate. Comparative Biochemistry and Physiology—A Molecular and Integrative Physiology 2013, 165. [CrossRef]
- Praagman, J.; Beulens, J.W.; Alssema, M.; Zock, P.L.; Wanders, A.J.; Sluijs, I.; van der Schouw, Y.T. The Association between Dietary Saturated Fatty Acids and Ischemic Heart Disease Depends on the Type and Source of Fatty Acid in the European Prospective Investigation into Cancer and Nutrition–Netherlands Cohort. Am. J. Clin. Nutr. 2016, 103, 356–365. [CrossRef]
- Mehrabani, A.; Jalalzadeh, M.; Jannati, N.; Lotfi, K.; Arzhang, P.; Azadbakht, L. Association Between Monounsaturated Fatty Acid Intake and Risk of Total Stroke and Its Subtypes: A Systematic Review and Dose–Response Meta-Analysis of Prospective Cohort Studies. Nutr. Rev. 2024. [CrossRef]
- Lotfi, K.; Salari-Moghaddam, A.; Yousefinia, M.; Larijani, B.; Esmaillzadeh, A. Dietary Intakes of Monounsaturated Fatty Acids and Risk of Mortality from All Causes, Cardiovascular Disease and Cancer: A Systematic Review and Dose-Response Meta-Analysis of Prospective Cohort Studies. Ageing Res. Rev. 2021, 72, 101467. [CrossRef]
- Hooper, L.; Martin, N.; Jimoh, O.F.; Kirk, C.; Foster, E.; Abdelhamid, A.S. Reduction in Saturated Fat Intake for Cardiovascular Disease. Cochrane Database of Systematic Reviews 2020, 2020. [CrossRef]
- Sheashea, M.; Xiao, J.; Farag, M.A. MUFA in Metabolic Syndrome and Associated Risk Factors: Is MUFA the Opposite Side of the PUFA Coin? Food Funct. 2021, 12, 12221–12234. [CrossRef]
- Yamada, M.H.; Fujihara, K.; Kodama, S.; Sato, T.; Osawa, T.; Yaguchi, Y.; Yamamoto, M.; Kitazawa, M.; Matsubayashi, Y.; Yamada, T.; et al. Associations of Systolic Blood Pressure and Diastolic Blood Pressure With the Incidence of Coronary Artery Disease or Cerebrovascular Disease According to Glucose Status. Diabetes Care 2021, 44, 2124–2131. [CrossRef]
- Cao, X.; Xia, J.; Zhou, Y.; Wang, Y.; Xia, H.; Wang, S.; Liao, W.; Sun, G. The Effect of MUFA-Rich Food on Lipid Profile: A Meta-Analysis of Randomized and Controlled-Feeding Trials. Foods 2022, 11, 1982. [CrossRef]
- Li, Y.; Hruby, A.; Bernstein, A.M.; Ley, S.H.; Wang, D.D.; Chiuve, S.E.; Sampson, L.; Rexrode, K.M.; Rimm, E.B.; Willett, W.C.; et al. Saturated Fats Compared With Unsaturated Fats and Sources of Carbohydrates in Relation to Risk of Coronary Heart Disease. J. Am. Coll. Cardiol. 2015, 66, 1538–1548. [CrossRef]
- Critselis, E.; Tsiampalis, T.; Damigou, E.; Georgousopoulou, E.; Barkas, F.; Chrysohoou, C.; Skoumas, J.; Pitsavos, C.; Liberopoulos, E.; Tsioufis, C.; et al. High Fish Intake Rich in N-3 Polyunsaturated Fatty Acids Reduces Cardiovascular Disease Incidence in Healthy Adults: The ATTICA Cohort Study (2002-2022). Front. Physiol. 2023, 14. [CrossRef]
- Park, G.-H.; Cho, J.-H.; Lee, D.; Kim, Y. Association between Seafood Intake and Cardiovascular Disease in South Korean Adults: A Community-Based Prospective Cohort Study. Nutrients 2022, 14, 4864. [CrossRef]
- Petersen, K.S.; Maki, K.C.; Calder, P.C.; Belury, M.A.; Messina, M.; Kirkpatrick, C.F.; Harris, W.S. Perspective on the Health Effects of Unsaturated Fatty Acids and Commonly Consumed Plant Oils High in Unsaturated Fat. British Journal of Nutrition 2024, 132, 1039–1050. [CrossRef]
- Bork, C.S.; Lundbye-Christensen, S.; Venø, S.K.; Lasota, A.N.; Schmidt, E.B.; Overvad, K. Plant N-3 PUFA Intake May Lower the Risk of Atherosclerotic Cardiovascular Disease Only among Subjects with a Low Intake of Marine n-3 PUFAs. Eur. J. Nutr. 2022, 61, 557–559. [CrossRef]
- Nestel, P.J.; Mori, T.A. Dietary Patterns, Dietary Nutrients and Cardiovascular Disease. Rev. Cardiovasc. Med. 2022, 23. [CrossRef]
- Wang, Y.; Liu, B.; Han, H.; Hu, Y.; Zhu, L.; Rimm, E.B.; Hu, F.B.; Sun, Q. Associations between Plant-Based Dietary Patterns and Risks of Type 2 Diabetes, Cardiovascular Disease, Cancer, and Mortality—a Systematic Review and Meta-Analysis. Nutr. J. 2023, 22, 46. [CrossRef]
- Rizzo, G.; Baroni, L.; Lombardo, M. Promising Sources of Plant-Derived Polyunsaturated Fatty Acids: A Narrative Review. Int. J. Environ. Res. Public Health 2023, 20.
- Duvall, M.G.; Levy, B.D. DHA- and EPA-Derived Resolvins, Protectins, and Maresins in Airway Inflammation. Eur. J. Pharmacol. 2016, 785. [CrossRef]
- Wang, Z.; Yuan, C.; Huang, T.; Lu, B. Early Nutritional Interventions for Chronic Low-Grade Inflammation. Trends in Endocrinology and Metabolism 2025.
- Browning, L.M.; Walker, C.G.; Mander, A.P.; West, A.L.; Gambell, J.; Madden, J.; Calder, P.C.; Jebb, S.A. Compared with Daily, Weekly n-3 PUFA Intake Affects the Incorporation of Eicosapentaenoic Acid and Docosahexaenoic Acid into Platelets and Mononuclear Cells in Humans. Journal of Nutrition 2014, 144. [CrossRef]
- Lindqvist, H.M.; Winkvist, A.; Gjertsson, I.; Calder, P.C.; Armando, A.M.; Quehenberger, O.; Coras, R.; Guma, M. Influence of Dietary N-3 Long Chain Polyunsaturated Fatty Acid Intake on Oxylipins in Erythrocytes of Women with Rheumatoid Arthritis. Molecules 2023, 28. [CrossRef]
- Calder, P.C. Omega-3 Fatty Acids and Inflammatory Processes: From Molecules to Man. Biochem. Soc. Trans. 2017, 45.
- Estruch, R.; Ros, E.; Salas-Salvadó, J.; Covas, M.-I.; Corella, D.; Arós, F.; Gómez-Gracia, E.; Ruiz-Gutiérrez, V.; Fiol, M.; Lapetra, J.; et al. Primary Prevention of Cardiovascular Disease with a Mediterranean Diet Supplemented with Extra-Virgin Olive Oil or Nuts. New England Journal of Medicine 2018, 378. [CrossRef]
- Filippou, C.; Tatakis, F.; Polyzos, D.; Manta, E.; Thomopoulos, C.; Nihoyannopoulos, P.; Tousoulis, D.; Tsioufis, K. Overview of Salt Restriction in the Dietary Approaches to Stop Hypertension (DASH) and the Mediterranean Diet for Blood Pressure Reduction. Rev. Cardiovasc. Med. 2022, 23. [CrossRef]
- Onwuzo, C.; Olukorode, J. o; Omokore, O.A.; Odunaike, O.S.; Omiko, R.; Osaghae, O. w; Sange, W.; Orimoloye, D.A.; Kristilere, H.O.; Addeh, E.; et al. DASH Diet: A Review of Its Scientifically Proven Hypertension Reduction and Health Benefits. Cureus 2023. [CrossRef]
- de Souza, P.A.L.; Marcadenti, A.; Portal, V.L. Effects of Olive Oil Phenolic Compounds on Inflammation in the Prevention and Treatment of Coronary Artery Disease. Nutrients 2017, 9.
- Bercea, C.I.; Cottrell, G.S.; Tamagnini, F.; McNeish, A.J. Omega-3 Polyunsaturated Fatty Acids and Hypertension: A Review of Vasodilatory Mechanisms of Docosahexaenoic Acid and Eicosapentaenoic Acid. Br. J. Pharmacol. 2021, 178.
- S, B.; L, R.; P, S.; KS, L. Novel SGC Stimulators and SGC Activators for the Treatment of Heart Failure. Handb. Exp. Pharmacol. 2017, 243.
- Satija, A.; Bhupathiraju, S.N.; Spiegelman, D.; Chiuve, S.E.; Manson, J.E.; Willett, W.; Rexrode, K.M.; Rimm, E.B.; Hu, F.B. Healthful and Unhealthful Plant-Based Diets and the Risk of Coronary Heart Disease in U.S. Adults. J. Am. Coll. Cardiol. 2017, 70, 411–422. [CrossRef]
- Gan, Z.H.; Cheong, H.C.; Tu, Y.-K.; Kuo, P.-H. Association between Plant-Based Dietary Patterns and Risk of Cardiovascular Disease: A Systematic Review and Meta-Analysis of Prospective Cohort Studies. Nutrients 2021, 13, 3952. [CrossRef]
- Choi, Y.; Larson, N.; Steffen, L.M.; Schreiner, P.J.; Gallaher, D.D.; Duprez, D.A.; Shikany, J.M.; Rana, J.S.; Jacobs, D.R. Plant-Centered Diet and Risk of Incident Cardiovascular Disease During Young to Middle Adulthood. J. Am. Heart Assoc. 2021, 10. [CrossRef]
- Yang, Q.; Cao, W.; Zhou, X.; Cao, W.; Xie, Y.; Wang, S. Anti-Thrombotic Effects of α-Linolenic Acid Isolated from Zanthoxylum Bungeanum Maxim Seeds. BMC Complement. Altern. Med. 2014, 14. [CrossRef]
- Singer, P.; Berger, I.; Moritz, V.; Förster, D.; Taube, C. N-6 and N-3 PUFA in Liver Lipids, Thromboxane Formation and Blood Pressure from SHR during Diets Supplemented with Evening Primrose, Sunflowerseed or Fish Oil. Prostaglandins Leukot. Essent. Fatty Acids 1990, 39. [CrossRef]
- Guasch-Ferré, M.; Liu, X.; Malik, V.S.; Sun, Q.; Willett, W.C.; Manson, J.A.E.; Rexrode, K.M.; Li, Y.; Hu, F.B.; Bhupathiraju, S.N. Nut Consumption and Risk of Cardiovascular Disease. J. Am. Coll. Cardiol. 2017, 70. [CrossRef]
- Bhatt, D.L.; Steg, P.G.; Miller, M.; Brinton, E.A.; Jacobson, T.A.; Ketchum, S.B.; Doyle, R.T.; Juliano, R.A.; Jiao, L.; Granowitz, C.; et al. Cardiovascular Risk Reduction with Icosapent Ethyl for Hypertriglyceridemia. New England Journal of Medicine 2019, 380, 11–22. [CrossRef]
- Goldberg, I.J.; Trent, C.M.; Schulze, P.C. Lipid Metabolism and Toxicity in the Heart. Cell Metab. 2012, 15.
- Shu, H.; Peng, Y.; Hang, W.; Nie, J.; Zhou, N.; Wang, D.W. The Role of CD36 in Cardiovascular Disease. Cardiovasc. Res. 2022, 118.
- Nakatani, K.; Masuda, D.; Kobayashi, T.; Sairyo, M.; Zhu, Y.; Okada, T.; Naito, A.T.; Ohama, T.; Koseki, M.; Oka, T.; et al. Pressure Overload Impairs Cardiac Function in Long-Chain Fatty Acid Transporter CD36-Knockout Mice. Int. Heart J. 2019, 60, 159–167. [CrossRef]
- Umbarawan, Y.; Syamsunarno, M.R.A.A.; Koitabashi, N.; Obinata, H.; Yamaguchi, A.; Hanaoka, H.; Hishiki, T.; Hayakawa, N.; Sano, M.; Sunaga, H.; et al. Myocardial Fatty Acid Uptake through CD36 Is Indispensable for Sufficient Bioenergetic Metabolism to Prevent Progression of Pressure Overload-Induced Heart Failure. Sci. Rep. 2018, 8, 12035. [CrossRef]
- Kim, T.T.; Dyck, J.R.B. The Role of CD36 in the Regulation of Myocardial Lipid Metabolism. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2016, 1861.
- Zhang, M.J.; Karachenets, S.; Gyberg, D.J.; Puccini, S.; Healy, C.L.; Wu, S.C.; Shearer, G.C.; O’Connell, T.D. Free Fatty Acid Receptor 4 in Cardiac Myocytes Ameliorates Ischemic Cardiomyopathy 2024.
- Anbazhagan, A.N.; Priyamvada, S.; Gujral, T.; Bhattacharyya, S.; Alrefai, W.A.; Dudeja, P.K.; Borthakur, A. A Novel Anti-Inflammatory Role of GPR120 in Intestinal Epithelial Cells. Am. J. Physiol. Cell Physiol. 2016, 310. [CrossRef]
- Park, M.H.; Lee, M.; Nam, G.; Kim, M.; Kang, J.; Choi, B.J.; Jeong, M.S.; Park, K.H.; Han, W.H.; Tak, E.; et al. N,N′-Diacetyl-p-Phenylenediamine Restores Microglial Phagocytosis and Improves Cognitive Defects in Alzheimer’s Disease Transgenic Mice. Proc. Natl. Acad. Sci. U. S. A. 2019, 116. [CrossRef]
- Shewale, S. V.; Brown, A.L.; Bi, X.; Boudyguina, E.; Sawyer, J.K.; Alexander-Miller, M.A.; Parks, J.S. In Vivo Activation of Leukocyte GPR120/FFAR4 by PUFAs Has Minimal Impact on Atherosclerosis in LDL Receptor Knockout Mice. J. Lipid Res. 2017, 58. [CrossRef]
- Hirschey, M.D.; Shimazu, T.; Goetzman, E.; Jing, E.; Schwer, B.; Lombard, D.B.; Grueter, C.A.; Harris, C.; Biddinger, S.; Ilkayeva, O.R.; et al. SIRT3 Regulates Mitochondrial Fatty-Acid Oxidation by Reversible Enzyme Deacetylation. Nature 2010, 464, 121–125. [CrossRef]
- Knox, C.; Wilson, M.; Klinger, C.M.; Franklin, M.; Oler, E.; Wilson, A.; Pon, A.; Cox, J.; Chin, N.E. (Lucy); Strawbridge, S.A.; et al. DrugBank 6.0: The DrugBank Knowledgebase for 2024. Nucleic Acids Res. 2024, 52, D1265–D1275. [CrossRef]
- Mallick, R.; Bhowmik, P.; Duttaroy, A.K. Targeting Fatty Acid Uptake and Metabolism in Cancer Cells: A Promising Strategy for Cancer Treatment. Biomedicine & Pharmacotherapy 2023, 167, 115591. [CrossRef]
- Spite, M.; Clària, J.; Serhan, C.N. Resolvins, Specialized Proresolving Lipid Mediators, and Their Potential Roles in Metabolic Diseases. Cell Metab. 2014, 19.
- Al-Shaer, A.E.; Buddenbaum, N.; Shaikh, S.R. Polyunsaturated Fatty Acids, Specialized pro-Resolving Mediators, and Targeting Inflammation Resolution in the Age of Precision Nutrition. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2021, 1866. [CrossRef]
- Davis, C.R.; Hodgson, J.M.; Woodman, R.; Bryan, J.; Wilson, C.; Murphy, K.J. A Mediterranean Diet Lowers Blood Pressure and Improves Endothelial Function: Results from the MedLey Randomized Intervention Trial. American Journal of Clinical Nutrition 2017, 105. [CrossRef]
- Salehin, S.; Rasmussen, P.; Mai, S.; Mushtaq, M.; Agarwal, M.; Hasan, S.M.; Salehin, S.; Raja, M.; Gilani, S.; Khalife, W.I. Plant Based Diet and Its Effect on Cardiovascular Disease. Int. J. Environ. Res. Public Health 2023, 20.
- Vogtschmidt, Y.D.; Soedamah-Muthu, S.S.; Imamura, F.; Givens, D.I.; Lovegrove, J.A. Replacement of Saturated Fatty Acids from Meat by Dairy Sources in Relation to Incident Cardiovascular Disease: The European Prospective Investigation into Cancer and Nutrition (EPIC)-Norfolk Study. Am. J. Clin. Nutr. 2024, 119, 1495–1503. [CrossRef]
- Hayes, K.C. Dietary Fat and Heart Health: In Search of the Ideal Fat. Asia Pac. J. Clin. Nutr. 2002, 11 Suppl 7.
- Wilkinson, M.J.; Manoogian, E.N.C.; Zadourian, A.; Lo, H.; Fakhouri, S.; Shoghi, A.; Wang, X.; Fleischer, J.G.; Navlakha, S.; Panda, S.; et al. Ten-Hour Time-Restricted Eating Reduces Weight, Blood Pressure, and Atherogenic Lipids in Patients with Metabolic Syndrome. Cell Metab. 2020, 31. [CrossRef]
- Djuricic, I.; Calder, P.C. Omega-3 (n-3) Fatty Acid–Statin Interaction: Evidence for a Novel Therapeutic Strategy for Atherosclerotic Cardiovascular Disease. Nutrients 2024, 16, 962. [CrossRef]
- Fan, H.; Zhou, J.; Yuan, Z. Meta-Analysis Comparing the Effect of Combined Omega-3 + Statin Therapy Versus Statin Therapy Alone on Coronary Artery Plaques. American Journal of Cardiology 2021, 151.
- Estruch, R.; Ros, E.; Salas-Salvadó, J.; Covas, M.-I.; Corella, D.; Arós, F.; Gómez-Gracia, E.; Ruiz-Gutiérrez, V.; Fiol, M.; Lapetra, J.; et al. Primary Prevention of Cardiovascular Disease with a Mediterranean Diet Supplemented with Extra-Virgin Olive Oil or Nuts. New England Journal of Medicine 2018, 378. [CrossRef]
- Plewes, M.R.; Burns, P.D.; Graham, P.E.; Bruemmer, J.E.; Engle, T.E. Influence of Omega-3 Polyunsaturated Fatty Acids from Fish Oil or Meal on the Structure of Lipid Microdomains in Bovine Luteal Cells. Anim. Reprod. Sci. 2018, 193. [CrossRef]
- Cottin, S.C.; Sanders, T.A.; Hall, W.L. The Differential Effects of EPA and DHA on Cardiovascular Risk Factors. In Proceedings of the Proceedings of the Nutrition Society; 2011; Vol. 70.
- McManus, S.; Tejera, N.; Awwad, K.; Vauzour, D.; Rigby, N.; Fleming, I.; Cassidy, A.; Minihane, A.M. Differential Effects of EPA versus DHA on Postprandial Vascular Function and the Plasma Oxylipin Profile in Men. J. Lipid Res. 2016, 57. [CrossRef]
- [Fixed Combination Atorvastatin-Perindopril (Lipercosyl®) for Substitution Treatment of Cardiovascular Risk Management]—PubMed Available online: https://pubmed.ncbi.nlm.nih.gov/33543860/ (accessed on 5 June 2025).
- Teo, K.K.; Mitchell, L.B.; Pogue, J.; Bosch, J.; Dagenais, G.; Yusuf, S. Effect of Ramipril in Reducing Sudden Deaths and Nonfatal Cardiac Arrests in High-Risk Individuals without Heart Failure or Left Ventricular Dysfunction. Circulation 2004, 110, 1413–1417. [CrossRef]
- Sedman, A.J.; Posvar, E. Clinical Pharmacology of Quinapril in Healthy Volunteers and in Patients with Hypertension and Congestive Heart Failure. Angiology 1989, 40, 360–369. [CrossRef]
- Simpson, K.; Jarvis, B. Lisinopril: A Review of Its Use in Congestive Heart Failure. Drugs 2000, 59, 1149–1167. [CrossRef]
- Peters, D.C.; Noble, S.; Plosker, G.L. Trandolapril. An Update of Its Pharmacology and Therapeutic Use in Cardiovascular Disorders. Drugs 1998, 56, 871–893. [CrossRef]
- Zannad, F.; Chati, Z.; Guest, M.; Plat, F.; Baille, N.; Baillie, S.; Tyneside, N.; Baksi, A.; Bloomfield, P.; Bocadamo, I.; et al. Differential Effects of Fosinopril and Enalapril in Patients with Mild to Moderate Chronic Heart Failure. Am. Heart J. 1998, 136, 672–680. [CrossRef]
- Woollett, L.A.; Buckley, D.D.; Yao, L.; Jones, P.J.H.; Granholm, N.A.; Tolley, E.A.; Heubi, J.E. Effect of Ursodeoxycholic Acid on Cholesterol Absorption and Metabolism in Humans. J. Lipid Res. 2003, 44, 935–942. [CrossRef]
- Eikelboom, J.W.; Quinlan, D.J.; O’Donnell, M. Major Bleeding, Mortality, and Efficacy of Fondaparinux in Venous Thromboembolism Prevention Trials. Circulation 2009, 120, 2006–2011. [CrossRef]
- Stroes, E.S.G.; Alexander, V.J.; Karwatowska-Prokopczuk, E.; Hegele, R.A.; Arca, M.; Ballantyne, C.M.; Soran, H.; Prohaska, T.A.; Xia, S.; Ginsberg, H.N.; et al. Olezarsen, Acute Pancreatitis, and Familial Chylomicronemia Syndrome. New England Journal of Medicine 2024, 390, 1781–1792. [CrossRef]
- Albosta, M.; Grant, J.K.; Michos, E.D. Bempedoic Acid: Lipid Lowering for Cardiovascular Disease Prevention. Heart Int. 2023, 17, 27. [CrossRef]
- Chang, Y.C.; Yeh, L.C.; Lin, T.T.; Huang, C.C.; Hung, C.S.; Yang, Y.Y.; Chuang, S.L.; Lin, L.Y.; Lin, L.C.; Kao, H.L.; et al. Colchicine to Prevent Cardiovascular Death after an Acute Myocardial Infarction. Journal of the Formosan Medical Association 2024. [CrossRef]
- Al-Atta, A.; Kuzemczak, M.; Alkhalil, M. Colchicine for the Prevention of Ischemic Stroke. Brain Circ. 2021, 7, 187–193. [CrossRef]
- Ahmad, T.; Manohar, S.A.; Stencel, J.D.; Le Jemtel, T.H. Dobutamine in the Management of Advanced Heart Failure. Journal of Clinical Medicine 2024, Vol. 13, Page 3782 2024, 13, 3782. [CrossRef]
- Olawi, N.; Krüger, M.; Grimm, D.; Infanger, M.; Wehland, M. Nebivolol in the Treatment of Arterial Hypertension. Basic Clin. Pharmacol. Toxicol. 2019, 125, 189–201. [CrossRef]
- Wang, Y. [Retracted] The Efficacy and Safety of Bisoprolol in the Treatment of Myocardial Infarction with Cardiac Insufficiency. Comput. Math. Methods Med. 2022, 2022, 3098726. [CrossRef]
- Packer, M.; Colucci, W.S.; Sackner-Bernstein, J.D.; Liang, C.S.; Goldscher, D.A.; Freeman, I.; Kukin, M.L.; Kinhal, V.; Udelson, J.E.; Klapholz, M.; et al. Double-Blind, Placebo-Controlled Study of the Effects of Carvedilol in Patients with Moderate to Severe Heart Failure: The PRECISE Trial. Circulation 1996, 94, 2793–2799. [CrossRef]
- Yan, C.; Duan, G.; Zhang, Y.; Wu, F.X.; Pan, Y.; Wang, J. Predicting Drug-Drug Interactions Based on Integrated Similarity and Semi-Supervised Learning. IEEE/ACM Trans. Comput. Biol. Bioinform. 2022, 19, 168–179. [CrossRef]
- Laviolette, S.R.; Nalepa, I.; Zelek-Molik, A.; Szeleszczuk, Ł.; Fr, D. Propranolol versus Other Selected Drugs in the Treatment of Various Types of Anxiety or Stress, with Particular Reference to Stage Fright and Post-Traumatic Stress Disorder. International Journal of Molecular Sciences 2022, Vol. 23, Page 10099 2022, 23, 10099. [CrossRef]
- Mahmood, A.; Eqan, M.; Pervez, S.; Javed, R.; Ullah, R.; Islam, A.; Khan, A.; Baig, A.A.; Kumar, G.; Abaid-Ullah, M.; et al. Drugs Resistance in Heart Diseases. Biochemistry of Drug Resistance 2021, 295–334. [CrossRef]
- Feingold, K.R. Cholesterol Lowering Drugs. Endotext 2024.
- Butt, W.Z.; Yee, J.K. The Role of Non-Statin Lipid-Lowering Medications in Youth with Hypercholesterolemia. Curr. Atheroscler. Rep. 2022, 24, 379–389. [CrossRef]
- Calderone, D.; Greco, A.; Ingala, S.; Agnello, F.; Franchina, G.; Scalia, L.; Buccheri, S.; Capodanno, D. Efficacy and Safety of Aspirin for Primary Cardiovascular Risk Prevention in Younger and Older Age: An Updated Systematic Review and Meta-Analysis of 173,810 Subjects from 21 Randomized Studies. Thromb. Haemost. 2022, 122, 445–455. [CrossRef]
- Kuang, H. yu; Li, Q.; Du, H. an; Chen, M.; Yin, Y. hui Efficacy and Safety of Long-Term Oral Bosentan in Different Types of Pulmonary Arterial Hypertension: A Systematic Review and Meta-Analysis. American Journal of Cardiovascular Drugs 2021, 21, 181–191. [CrossRef]
- Biswas, K.; Tiwari, A.; Jadhav, P.; Goel, A.; Chanukya, G. V Rosuvastatin and Fenofibrate Combination in The Treatment of Mixed Hyperlipidemia: A Narrative Review. Journal of Current Medical Research and Opinion 2021, 4, 867–877. [CrossRef]
- Lusk, J.B.; Glover, L.S.; Soneji, S.; Granger, C.B.; O’Brien, E.; Pagidipati, N. Cardiovascular Event Reduction among a US Population Eligible for Semaglutide per the SELECT Trial. Am. Heart J. 2024, 276, 110–114. [CrossRef]
- Drucker, D.J. Prevention of Cardiorenal Complications in People with Type 2 Diabetes and Obesity. Cell Metab. 2024, 36, 338–353. [CrossRef]
- Suda, N.; Leon-Martinez, D.; Peter, P.R.; Flannery, C.A.; Irani, R.A. Management of Severe Hypertriglyceridemia in Pregnancy With Niacin: Reevaluating Safety and Therapeutic Benefits. Case Rep. Endocrinol. 2025, 2025, 2644678. [CrossRef]
- Forry, J.; Chappell, A.; Alexander, E.; Allen, J. Tenecteplase: A Review of Its Pharmacology and Uses. AACN Adv. Crit. Care 2023, 34, 77–83. [CrossRef]
- Chambergo-Michilot, D.; Alur, A.; Kulkarni, S.; Agarwala, A. Mipomersen in Familial Hypercholesterolemia: An Update on Health-Related Quality of Life and Patient-Reported Outcomes. Vasc. Health Risk Manag. 2022, 18, 73–80. [CrossRef]
- Vaduganathan, M.; Filippatos, G.; Claggett, B.L.; Desai, A.S.; Jhund, P.S.; Henderson, A.; Brinker, M.; Kolkhof, P.; Schloemer, P.; Lay-Flurrie, J.; et al. Finerenone in Heart Failure and Chronic Kidney Disease with Type 2 Diabetes: The FINE-HEART Pooled Analysis of Cardiovascular, Kidney, and Mortality Outcomes. Nat. Med. 2024, 30, 3758–3764. [CrossRef]
- Kiaei, A.A.; Boush, M.; Abadijou, S.; Momeni, S.; Safaei, D.; Bahadori, R.; Salari, N.; Mohammadi, M. Recommending Drug Combinations Using Reinforcement Learning Targeting Genes/Proteins Associated with Heterozygous Familial Hypercholesterolemia: A Comprehensive Systematic Review and Net-Work Meta-Analysis. 2023. [CrossRef]
- Oyama, K.; Furtado, R.H.M.; Fagundes, A.; Zelniker, T.A.; Tang, M.; Kuder, J.; Murphy, S.A.; Hamer, A.; Wang, H.; Keech, A.C.; et al. Effect of Evolocumab on Complex Coronary Disease Requiring Revascularization. J. Am. Coll. Cardiol. 2021, 77, 259–267. [CrossRef]
- Chaudhary, R.; Mohananey, A.; Sharma, S.P.; Singh, S.; Singh, A.; Kondur, A. Improving Outcomes in Cardiovascular Diseases: A Review on Vorapaxar. Cardiol. Rev. 2022, 30, 241–246. [CrossRef]
- Bryan Richard, S.; Huang, B.; Liu, G.; Yang, Y.; Luo, S. Impact of Ivabradine on the Cardiac Function of Chronic Heart Failure Reduced Ejection Fraction: Meta-Analysis of Randomized Controlled Trials. Clin. Cardiol. 2021, 44, 463–471. [CrossRef]
- Chen, W.-H.; Lau, C.-P.; Lau, C.A. Recent Advances in Antithrombin Therapy for Acute Coronary Syndromes. Journal of the Hong Kong College of Cardiology 2022, 7, 109–118. [CrossRef]
- Imbalzano, E.; Orlando, L.; Dattilo, G.; Gigliotti De Fazio, M.; Camporese, G.; Russo, V.; Perrella, A.; Bernardi, F.F.; Di Micco, P. Update on the Pharmacological Actions of Enoxaparin in Nonsurgical Patients. Medicina 2024, Vol. 60, Page 156 2024, 60, 156. [CrossRef]
- Wang, J.; Zou, D. Tirofiban-Induced Thrombocytopenia. Ann. Med. 2023, 55, 2233425. [CrossRef]
- Ganda, O.P. Triglyceride-Rich Lipoproteins, Remnant-Cholesterol, and Atherosclerotic Cardiovascular Disease. Curr. Opin. Lipidol. 2023, 34, 105–113. [CrossRef]
- Patel, K.; Patil, J.; Girase, T.; Tatiya, A.; Patil, D. An Overview of the Applications of Gemfibrozil Nano-Formulation in Hyperlipidemia. Materials Proceedings 2023, Vol. 14, Page 45 2023, 14, 45. [CrossRef]
- Kalstad, A.A.; Myhre, P.L.; Laake, K.; Tveit, S.H.; Schmidt, rik B.; Smith, P.; Nilsen, D.W.T.; Tveit, A.; Fagerland, M.W.; Solheim, S.; et al. Effects of N-3 Fatty Acid Supplements in Elderly Patients after Myocardial Infarction: A Randomized, Controlled Trial. Circulation 2021, 143, 528–539. [CrossRef]
- Sanderson, N.C.; Parker, W.A.E.; Storey, R.F. Ticagrelor: Clinical Development and Future Potential. Rev. Cardiovasc. Med. 2021, 22, 373–394. [CrossRef]
- Mitsis, A.; Gragnano, F. Myocardial Infarction with and without ST-Segment Elevation: A Contemporary Reappraisal of Similarities and Differences. Curr. Cardiol. Rev. 2020, 17, 49–58. [CrossRef]
- Santos, R.D.; Wiegman, A.; Caprio, S.; Cariou, B.; Averna, M.; Poulouin, Y.; Scemama, M.; Manvelian, G.; Garon, G.; Daniels, S. Alirocumab in Pediatric Patients With Heterozygous Familial Hypercholesterolemia: A Randomized Clinical Trial. JAMA Pediatr. 2024, 178, 283–293. [CrossRef]
- Murphy, L.R.; Hill, T.P.; Paul, K.; Talbott, M.; Golovko, G.; Shaltoni, H.; Jehle, D. Tenecteplase Versus Alteplase for Acute Stroke: Mortality and Bleeding Complications. Ann. Emerg. Med. 2023, 82, 720–728. [CrossRef]
- Pitt, B.; Bhatt, D.L.; Szarek, M.; Cannon, C.P.; Leiter, L.A.; McGuire, D.K.; Lewis, J.B.; Riddle, M.C.; Voors, A.A.; Metra, M.; et al. Effect of Sotagliflozin on Early Mortality and Heart Failure-Related Events: A Post Hoc Analysis of SOLOIST-WHF. JACC Heart Fail. 2023, 11, 879–889. [CrossRef]
- Yoo, Y.G.; Lim, M.J.; Kim, J.S.; Jeong, H.E.; Ko, H.; Shin, J.Y. Risk of Myocardial Infarction, Heart Failure, and Cerebrovascular Disease with the Use of Valsartan, Losartan, Irbesartan, and Telmisartan in Patients. Medicine (United States) 2023, 102, E36098. [CrossRef]
- Gillmore, J.D.; Judge, D.P.; Cappelli, F.; Fontana, M.; Garcia-Pavia, P.; Gibbs, S.; Grogan, M.; Hanna, M.; Hoffman, J.; Masri, A.; et al. Efficacy and Safety of Acoramidis in Transthyretin Amyloid Cardiomyopathy. New England Journal of Medicine 2024, 390, 132–142. [CrossRef]
- Liu, T.; Zuo, R.; Wang, J.; Huangtao, Z.; Wang, B.; Sun, L.; Wang, S.; Li, B.; Zhu, Z.; Pan, Y. Cardiovascular Disease Preventive Effects of Aspirin Combined with Different Statins in the United States General Population. Sci. Rep. 2023, 13, 1–7. [CrossRef]
- Cai, T.; Abel, L.; Langford, O.; Monaghan, G.; Aronson, J.K.; Stevens, R.J.; Lay-Flurrie, S.; Koshiaris, C.; McManus, R.J.; Richard Hobbs, F.D.; et al. Associations between Statins and Adverse Events in Primary Prevention of Cardiovascular Disease: Systematic Review with Pairwise, Network, and Dose-Response Meta-Analyses. BMJ 2021, 374, 1537. [CrossRef]
- Grinspoon, S.K.; Fitch, K. V.; Zanni, M. V.; Fichtenbaum, C.J.; Umbleja, T.; Aberg, J.A.; Overton, E.T.; Malvestutto, C.D.; Bloomfield, G.S.; Currier, J.S.; et al. Pitavastatin to Prevent Cardiovascular Disease in HIV Infection. New England Journal of Medicine 2023, 389, 687–699. [CrossRef]
- Yang, X.H.; Zhang, B.L.; Cheng, Y.; Fu, S.K.; Jin, H.M. Statin Use and the Risk of CVD Events, Stroke, and All-Cause Mortality in Patients with Diabetes: A Systematic Review and Meta-Analysis. Nutrition, Metabolism and Cardiovascular Diseases 2022, 32, 2470–2482. [CrossRef]
Figure 1.
Classification of Fatty Acids by Hydrocarbon Chain Characteristics. Fatty acids are classified based on the nature of their hydrocarbon chains into saturated (SFA) and unsaturated fatty acids (UFA). UFAs include cis and trans isomers. Cis UFAs are subdivided into monounsaturated (MUFA, with one double bond) and polyunsaturated fatty acids (PUFA, with two or more double bonds), which include essential fatty acids (EFAs) such as omega-3 and omega-6. SFAs are further categorized by chain length into very long-chain (VLCFA), long-chain (LCFA), medium-chain (MCFA), and short-chain fatty acids (SCFA). Representative examples are listed for each category.
Figure 1.
Classification of Fatty Acids by Hydrocarbon Chain Characteristics. Fatty acids are classified based on the nature of their hydrocarbon chains into saturated (SFA) and unsaturated fatty acids (UFA). UFAs include cis and trans isomers. Cis UFAs are subdivided into monounsaturated (MUFA, with one double bond) and polyunsaturated fatty acids (PUFA, with two or more double bonds), which include essential fatty acids (EFAs) such as omega-3 and omega-6. SFAs are further categorized by chain length into very long-chain (VLCFA), long-chain (LCFA), medium-chain (MCFA), and short-chain fatty acids (SCFA). Representative examples are listed for each category.

Figure 2.
Fatty Acid Uptake and Metabolism in Cardiomyocytes. This figure illustrates the pathways of fatty acid uptake, intracellular trafficking, and metabolic processing in cardiomyocytes. Circulating fatty acids, derived from lipoproteins (chylomicrons, VLDL, LDL) or in non-esterified form, enter cardiomyocytes via various transporters including CD36, FATP1/4, and FABPpm. Once inside, fatty acids are activated to acyl-CoA by ACSL1 and directed toward mitochondrial β-oxidation via CPT1/2 for ATP generation through the Krebs cycle. Alternatively, acyl-CoAs are stored as triacylglycerols (TAG) in lipid droplets or converted into signaling molecules like ceramides and eicosanoids. Regulatory pathways involving transcription factors (PPARs, FOXO1), enzymes (DGAT1, HSL, AGTL), and receptors (VEGFR1, Apelin receptor) coordinate lipid uptake, storage, and oxidation in response to metabolic demands.
Figure 2.
Fatty Acid Uptake and Metabolism in Cardiomyocytes. This figure illustrates the pathways of fatty acid uptake, intracellular trafficking, and metabolic processing in cardiomyocytes. Circulating fatty acids, derived from lipoproteins (chylomicrons, VLDL, LDL) or in non-esterified form, enter cardiomyocytes via various transporters including CD36, FATP1/4, and FABPpm. Once inside, fatty acids are activated to acyl-CoA by ACSL1 and directed toward mitochondrial β-oxidation via CPT1/2 for ATP generation through the Krebs cycle. Alternatively, acyl-CoAs are stored as triacylglycerols (TAG) in lipid droplets or converted into signaling molecules like ceramides and eicosanoids. Regulatory pathways involving transcription factors (PPARs, FOXO1), enzymes (DGAT1, HSL, AGTL), and receptors (VEGFR1, Apelin receptor) coordinate lipid uptake, storage, and oxidation in response to metabolic demands.

Figure 3.
Mechanisms of lipotoxicity-induced mitochondrial dysfunction and cardiomyopathy. Excess fatty acid uptake through CD36, VLDLR, and MCT1 leads to lipotoxicity, triggering ER stress, UPR, and mitochondrial dysfunction. Mitochondrial impairment—marked by reduced fatty acid oxidation, ATP production, and calcium handling—results in increased ROS generation. ROS cause oxidative damage, inflammation, and imbalance in mitochondrial dynamics, ultimately leading to cell death and the development of cardiomyopathy.
Figure 3.
Mechanisms of lipotoxicity-induced mitochondrial dysfunction and cardiomyopathy. Excess fatty acid uptake through CD36, VLDLR, and MCT1 leads to lipotoxicity, triggering ER stress, UPR, and mitochondrial dysfunction. Mitochondrial impairment—marked by reduced fatty acid oxidation, ATP production, and calcium handling—results in increased ROS generation. ROS cause oxidative damage, inflammation, and imbalance in mitochondrial dynamics, ultimately leading to cell death and the development of cardiomyopathy.

Table 1.
Summary of pathogenic mechanisms & CVD outcomes by fatty acids.
| Fatty Acid Types | Key Pathogenic Mechanisms | Associated CVD Outcomes |
| Saturated FAs (SFAs)(e.g., palmitate, stearate) | • ER stress → UPR activation (PERK/IRE1 → CHOP) → caspase-3–mediated apoptosis • Mitochondrial dysfunction (ceramide accumulation, impaired electron transport) • Lipid intermediate buildup (ceramides, DAG) → insulin resistance, fibrosis |
• Hypertrophic cardiomyopathy • Left ventricular dysfunction • Increased risk of heart failure • Atherosclerosis progression |
| Trans FAs (TFAs)(e.g., industrial elaidic acid) | • NF-κB–mediated inflammation (↑ TNF-α, IL-6) • NADPH oxidase → ↑ superoxide → oxidative damage • Endothelial dysfunction (↓ eNOS), macrophage infiltration • Impaired phospholipid membrane integrity → pro-arrhythmic remodeling |
• Accelerated atherosclerosis • Coronary artery disease • Exacerbated ischemia-reperfusion injury • Higher incidence of arrhythmias |
| n-3 PUFAs(e.g., EPA, DHA) | • Low-Moderate Dose (1–2 g/day):—↓ NF-κB activation, ↓ proinflammatory eicosanoids—↑ antioxidant enzymes (SOD, catalase)—Membrane stabilization → improved ion channel function • High Dose (>3 g/day):—Altered ion currents → prolonged repolarization—Potential imbalance of ROS if excess incorporation |
• ↓ Major adverse cardiovascular events (MACE) at moderate intake • ↓ post-MI inflammation • ↓ sudden cardiac death • ↑ risk of atrial fibrillation at high dose |
| MUFAs (Plant-Derived)(e.g., oleic acid) | • ↑ HDL, ↓ oxidized LDL • ↑ eNOS activity → vasodilation • Redirect SFA-induced lipids into triglycerides rather than ceramides • Anti-inflammatory via NF-κB suppression |
• ↓ CVD mortality • ↓ progression of atherosclerosis • Improved diastolic function |
| MUFAs (Animal-Derived)(e.g., palmitoleic acid) | • May contribute to low-grade inflammation in insulin-resistant states • In conditions of metabolic syndrome, can be converted to ceramides (via salvage pathways) → mild ER stress • Less favorable lipid profile modulation compared with plant MUFAs |
• Neutral to slightly ↑ CVD risk when part of high-SFA diet • Effects largely depend on overall dietary pattern |
Table 2.
Key Clinical Trials of n-3 PUFA Supplementation and CVD Outcomes.
| Intervention | Key Trials | Patient Population | CVD Benefit | Risks / Limitations |
| Icosapent ethyl (4 g/day) | REDUCE-IT (2018) | CVD/diabetes + high triglecerides | ↓ 25% MACE; ↓ MI, stroke, CV death |
|
| EPA+DHA (4 g/day, CA) | STRENGTH (2020) | High CVD risk + low HDL/high triglecerides | No significant CV benefit |
|
| EPA+DHA (1 g/day, EE) | ASCEND, VITAL (2019) | Primary prevention (no CVD) | No significant primary prevention benefit |
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



