Submitted:
17 March 2026
Posted:
18 March 2026
You are already at the latest version
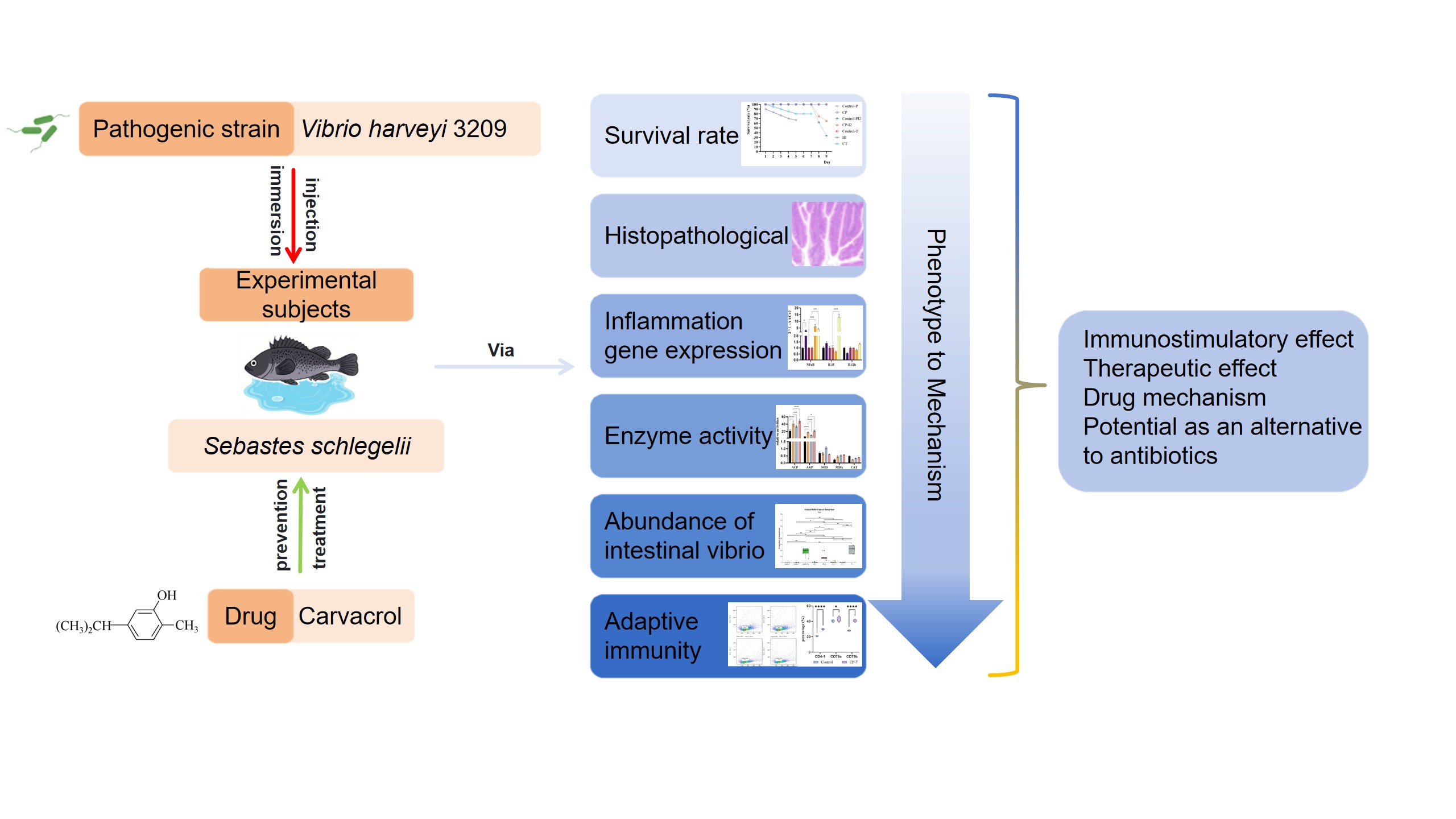
Abstract
Results indicated that pre-treatment with carvacrol significantly increased the survival rate of infected Sebastes schlegelii. Conversely, post-infection administration alleviated intestinal pathological damage. Carvacrol regulated host immunity by modulating the transcription of immune-related genes NF-κB/RelA and IL-15. It had no significant effects on SOD, MDA and CAT, suggesting that the oxidative defense pathway was not involved. Analysis of intestinal Vibrio pathogen load confirmed that carvacrol could inhibit the growth and colonization of intestinal Vibrio, thereby maintaining microbial homeostasis. Immunohistochemistry and peripheral blood flow cytometry showed that carvacrol enhanced the adaptive immunity of fish by increasing the proportions of CD4‑1⁺ T cells and CD79a/CD79b⁺ B cells in tissues. In conclusion, carvacrol enhances the resistance of S. schlegelii against V. harveyi by inhibiting pathogenic bacteria, improving intestinal morphological structure, reducing pathogenic bacterial load to maintain microbial homeostasis, and enhancing the adaptive immunity of the organism. This study provides a theoretical basis and data support for the substitution of antibiotics and the development of green feed additives in aquaculture.
Keywords:
mariculture
; chinese herbal monomer
; antibiotic substitutes
; Sebastes schlegelii
; Vibrio harveyi
; immunity
1. Introduction
Aquaculture has emerged as the dominant source of global aquatic product production. Nevertheless, the frequent occurrence of diseases has led to a significant reduction in the yield of certain aquaculture species and a subsequent decline in economic returns. Bacterial diseases represent the predominant category of ailments in aquaculture (van Gelderen, Debnath et al. 2025).
Vibrio is one of the most harmful pathogens to aquatic animals in the marine environment, causing massive deaths of aquaculture animals (Bloomfield, Palau et al. 2025, Zhang, Qiu et al. 2025). Species such as Vibrio harveyi, Vibrio alginolyticus, and Vibrio vulnificus pose considerable risks to the health of farmed fish (Manchanayake, Mohamad et al. 2025). V. harveyi, a Gram-negative marine bacterium, is widely distributed in the environment and affects a variety of marine vertebrates and invertebrates (Jeong-hyeon, Gyeong-mi et al. 2007). Fish infected with V. harveyi can display a diverse range of clinical symptoms, including growth retardation, spiral or irregular movement, darkening of body surface pigmentation, the appearance of ulcers and blood clots on the body surface, an increase in the number of mucus cells, eye damage or cloudiness, bleeding in the oral area and around internal organs, and other related manifestations. In severe instances, it can result in the large-scale mortality of farmed fish (Strem, Meiri-Ashkenazi et al. 2023, Wan, Da et al. 2023, Yang, Cheng et al. 2023).
Antibiotics serve as one of the primary means for the prevention and treatment of bacterial diseases in aquatic animals. However, the improper utilization of antibiotics in aquaculture promotes drug resistance in pathogenic bacteria, which not only hinders their eradication but also further reduces the success rates of farming. Therefore, it is of great necessity to develop effective antibiotic alternatives that can inhibit pathogenic bacteria (Ma, Chen et al. 2022, Milijasevic, Veskovic-Moracanin et al. 2024). Currently, apart from the use of antibiotics, the primary strategies to reduce aquaculture losses caused by these pathogens include researching and developing Chinese herbal medicines, probiotics (Pan and Yan 2020, Wu, Jiang et al. 2021), antimicrobial peptides (Bhat, Khangembam et al. 2023, Prazdnova, Zaikina et al. 2025) and vaccines(Imtiaz, Anwar et al. 2024, Tayyab, Islam et al. 2025). Chinese Herbal Medicines (CHM) have attracted extensive attention due to their natural origin, low toxicity, and minimal side-effects. Additionally, they can effectively prevent and treat bacterial and viral diseases, enhance immunity, and improve antioxidant capacity (Zhu, Yao et al. 2023, Wang, Huang et al. 2024, Xu, Li et al. 2024). Nevertheless, traditional Chinese herbal medicines (TCHMs) present problem such as complex compositions, poor drug stability, and potential drug interactions (Yu, Sun et al. 2023). Therefore, it is highly necessary to develop TCHM monomers with well-defined drug mechanisms and targeted therapeutic effects. A multitude of studies have demonstrated that Chinese herbal essential oils (EOs) exhibit antimicrobial activity and can enhance the growth performance, immunity, and antioxidant activity of organisms (Alagawany, Farag et al. 2020, Deng, Deng et al. 2024, Hirose, Horiyama et al. 2024). Carvacrol is the primary component of essential oils derived from Cinnamomum cassia Presl and Origanum vulgare L (Zhou, Ji et al. 2007). It exerts an effective inhibitory effect on some pathogenic bacteria in aquaculture, including Vibrio cholerae, Vibrio parahaemolyticus, V. alginolyticus, Vibrio anguillarum, and V. harveyi (Coelho, Rosa et al. 2021, Das, Chourashi et al. 2021, Rossi, Esteban et al. 2021). Furthermore, carvacrol can enhance immunity, improve growth performance and digestive capacity, modulate intestinal microbiota, and boost antioxidant activity (Ahmadifar, Mansour et al. 2014, Wang, Mai et al. 2022).
However, there are relatively few studies on the preventive and therapeutic effects of carvacrol in marine fish against Vibrio infection, as well as its mechanism of action. Therefore, in this study, S. schlegelii was selected as the experimental subject. Through a comprehensive analysis of the changes in apparent symptoms, survival rate, intestinal pathological sections, enzyme activity, quantitative expression of inflammatory genes, pathogenic microbial load after the action of V. harveyi and carvacrol, and by investigating the effects of carvacrol preventive administration on the proportion changes of CD4-1+ T cells, CD79a+ B cells, and CD79bb+ B cells in key immune tissues and peripheral blood. This research systematically explores the preventive and immunity-enhancing effects of carvacrol on juvenile S. schlegelii without pathogenic bacteria infection, as well as its therapeutic effects after pathogenic bacteria infection. Our findings aim to provide a novel antibiotic-alternative strategy for controlling V. harveyi in marine fish farming, while also offering references for the subsequent development of new fishery drugs.
2. Materials and Methods
2.1. Fish Management, Sampling, Survival Rate Calculation and Pathological Observation
The aquaculture farm is located in Rizhao, Shandong Province. Feeding methods, sampling time-points and steps are shown in Figure 1. The carvacrol employed in the experiment was purchased from Aladdin Biochemical Technology Co., Ltd. (Shanghai, China). The serial number of the V. harveyi strain utilized used is V3209. It was isolated and identified from diseased S. schlegelii with body lesion and is stored in the laboratory of Yellow Sea Fishery Research Institute. Seven fish were randomly chosen per group. Their foregut, midgut, hindgut, and liver were dissected on a tray, each for specific analyses: the foregut for microbial diversity analysis; the midgut for gene expression detection, enzyme activity measurement, and pathological sectioning; the hindgut for intestinal metabolite determination; and the liver for enzyme activity assay.
Intestinal samples designated for pathological sections were immersed in a fixative solution for 16 h and subsequently transferred to 75% ethanol. Meanwhile, all the remaining intestinal and liver tissues were placed into cryovials, rapidly frozen in liquid nitrogen for 2 h, and then stored at -80 ℃ for future analysis.
Throughout the V. harveyi challenge period in the experiment, the apparent symptoms of the experimental fish were continuously recorded. Moreover, the daily mortality of the experimental fish was documented, and the survival rate was calculated using the following formula:
During the sampling process, the apparent symptoms of the experimental fish were carefully observed. Then, intestinal tissues were placed in an ASP300S Tissue Processor (LEICA, Germany) to undergo dehydration, clearing, and paraffin infiltration. Subsequently, an EG1150H Paraffin Embedding Station (LEICA, Germany) was employed to embed the tissues. The trimmed paraffin blocks were sectioned with an RM2235 Rotary Microtome (LEICA, Germany) to a thickness of 5 µm, and at least three non-consecutive sections were cut from each paraffin-embedded block. The cut tissue sections were transferred onto glass slides, and 30% alcohol was gently added dropwise from the side to flatten the paraffin sections. The slides were then transferred to an HI1220 Slide Dryer (LEICA, Germany) for section spreading at 45 ℃. After the sections were fully flattened, they were dried at 60 ℃ for 10 hours. Following the slide drying step, the tissue sections were stained with hematoxylin and eosin using an AUTO SSTAINER XL Staining Machine (LEICA, Germany).
2.2. Carvacrol Treatment and Intestinal Pathogen Load in Sebastes schlegelii
Microbial diversity sequencing can be classified into two approaches: DNA extraction sequencing and RNA extraction followed by reverse transcription to cDNA for sequencing. These two sequencing analyses were carried out by Shanghai Meiji Biomedical Technology Co., Ltd. A methodological comparison was conducted using these two methods to determine microbial diversity.
2.3. Effects of Carvacrol on Non-Specific Immunity of Sebastes schlegelii Under Vibrio harveyi Stress
The expression levels of intestinal inflammation-related genes and enzyme activities in Sebastes schlegelii from the Control group and each treatment group were determined to explore the effect of the drug on non-specific immunity under pathogenic bacterial stress. The midgut tissue was retrieved from -80 ℃ storage and immediately placed in a foam-insulated container filled with liquid nitrogen for rapid freezing. The tissue was then ground into a fine powder using liquid nitrogen and a mortar. Total RNA was extracted using the FastPure Cell/Tissue Total RNA Isolation Kit V2. After measuring the RNA concentration with a spectrophotometer, the RNA was reverse-transcribed into cDNA using the HiScript III RT SuperMix for qPCR (+gDNA wiper) kit. Finally, quantitative analysis was performed using the Taq Pro Universal SYBR qPCR Master Mix. All the kits used were purchased from Vazyme Biotech Co., Ltd. (Nanjing, China). Three genes were selected for Real-time Quantitative PCR (q-PCR), with RPL-17 chosen as the reference gene. Gene sequences were searched for using NCBI, and primers were designed using Primer Premier 5. Primers for q-PCR are presented in Table 1.
The midgut segment of S. schlegelii was retrieved from -80 ℃ storage and promptly transferred to a foam-insulated container filled with liquid nitrogen for rapid freezing, aiming to preserve tissue integrity and enzymatic activity. The activities of total protein (TP), alkaline phosphatase (AKP), acid phosphatase (ACP), catalase (CAT), and superoxide dismutase (SOD), along with the content of malondialdehyde (MDA), were precisely determined following the manufacturer’s instructions provided with the assay kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China).
2.4. Regulatory Effect of Carvacrol Prophylactic Treatment on Specific Immunity in Sebastes schlegelii
The antibacterial effect and non-specific immune response of the drug were investigated in previous experiments. In this section, the changes in the specific immunity of fish after 7-day prophylactic administration of carvacrol were verified using antibodies that were prepared previously in the laboratory and have been confirmed to specifically recognize CD4-1, CD79a, and CD79b molecules.
Immunohistochemistry was performed on the liver, spleen, and kidney of Sebastes schlegelii. The previously prepared CD4-1, CD79a, and CD79b antibodies were used as the primary antibody, while the HRP-labeled goat anti-rabbit antibody as the secondary antibody. This was carried out to analyze the changes in CD4-1+ T cells and CD79a+/CD79b+ B cells so as to evaluate effects on specific immunity. Tissues were fixed for 24 h, then paraffin-sectioned and dewaxed. Endogenous peroxidase was blocked with 3% H2O2 for 25 min in the dark. Sections were blocked with 3% BSA for 30 min, incubated with primary and secondary antibodies, visualized with DAB, counterstained with hematoxylin, dehydrated, mounted, and observed microscopically. The number and proportion of positive cells were recorded.
Six fish were randomly selected from each experimental group. Before blood collection, 4 mL of single-cell suspension medium was aspirated into a 5 mL syringe, and blood was collected from the caudal vein into sterile 15-mL centrifuge tubes. Leukocytes were isolated using a commercial fish whole-blood leukocyte isolation kit within 2 h after collection. In a 10 mL centrifuge tube, 5 mL of isolation solution was gently added to 3 mL of blood diluted with single-cell suspension medium, avoiding high-drop addition to prevent layer disruption. Centrifugation was performed at 700×g for 33 min (acceleration = 1, deceleration = 0) using a swinging-bucket or angle rotor. After centrifugation, four layers were observed (from top to bottom): pale yellow single-cell suspension medium, white opaque membranous leukocyte layer, colorless transparent isolation layer, and red blood cell pellet. The leukocyte layer was carefully collected into 1.5-mL centrifuge tubes; residual erythrocytes were lysed with red blood cell lysis buffer provided by the kit to avoid data interference.
Five to eight volumes of lysis buffer were added to the cell pellet. Subsequently, the mixture was gently mixed and incubated at 4 °C for 1.5 min. After centrifugation at 2000 rpm for 7 min at 4 °C, the red supernatant was discarded. The pellet was resuspended and washed with 1% bovine serum albumin—phosphate—buffered saline (BSA-PBS), and then centrifuged again at 2000 rpm for 7 min. The supernatant was removed, and the leukocytes were retained for subsequent use.
Washed leukocytes were incubated with a validated homemade lymphocyte surface marker primary antibody (1:10,000) at room temperature with shaking at 100 rpm for 1 h. After centrifugation and washing with 1% BSA-PBS, the cells were incubated with 488 HRP-conjugated secondary antibody (1:1000) in the dark (aluminum foil-wrapped) for 1 h. The samples were washed, filtered through a cell strainer, and analyzed within 30 min.
Antibody-labeled cells were resuspended in PBS at a concentration ranging from 1 × 106–1 × 107 cells/mL and analyzed using a Beckman CytoFLEX flow cytometer (Beckman Coulter, Inc.) with the fluorescein isothiocyanate (FITC) fluorescence channel. Data acquisition was terminated after 10,000 cells were recorded for each sample, and data were stored for subsequent analysis.
2.5. Statistical Analysis
Statistical analyses were performed using SPSS (IBM SPSS Statistics 27.0.1) and the Meiji Cloud Platform (Shanghai Meiji Biomedical Technology Co., Ltd.). One-way Analysis of Variance (ANOVA) and Kruskal-Wallis test were applied to determine statistically significant differences among treatment groups.
The use of experimental animals and the study protocol were reviewed and approved by the Ethics Committee of Yellow Sea fishery research institute, Chinese Academy of Fishery Sciences.
3. Result
3.1. Effects of Carvacrol on Survival and Intestinal Histopathology of Fish Infected with Vibrio harveyi
After 7-day feeding period with a blank diet, the experimental fish challenged with V. harveyi via injection for 2 days (Control-PI2) showed extensive intestinal hemorrhage. The experimental fish in the group challenged with V. harveyi via immersion for 5 days (HI) exhibited symptoms such as body surface exhibited ulceration, intestinal hemorrhage, ascites, and liver atrophy (Figure 2).
The survival rates of each group during the experiment are presented in Figure 3. In both the prevention phase and the treatment phase, the survival rates of the two blank control groups and the group fed with carvacrol-supplemented feed for 7 days (Control-P, Control-T, CP-7) remained 100% throughout the entire experiment. In the prevention phase, the survival rate of experimental fish in the group that was fed with blank feed for 7 days and then subjected to an injection challenge with V. harveyi for 2 days (Control-PI2) was 33.33%. In contrast, the survival rate of the group that was fed with carvacrol-supplemented feed for 7 days before the 2-days V. harveyi injection challenge (CP-I2) reached 65%. In the treatment experiment, the survival rate of experimental fish that were immersed in V. harveyi for 5 days (HI) without carvacrol treatment was 66.67%. The final survival rate of experimental fish in the carvacrol treatment group (CT) after HI challenge remained stable at 80%.
Intestinal histology revealed that in the two control groups (Control-P and Control-T), fish maintained well-organized intestinal villi that were dense, structurally intact, and free from rupture, fusion, or atrophy. The single-layer columnar epithelial cells appeared plump with intense cytoplasmic eosinophilia, and their nuclei were regularly aligned at the basal region. Goblet cell density remained normal, with no obvious suppression of mucin secretion. The submucosal connective tissue showed no edema or inflammatory cell infiltration. Smooth muscle fibers in the muscular layer were orderly arranged without hypertrophy or necrosis, and the serosal layer remained structurally intact with no adhesion or exudation. (Figure 4). In the unprotected V. harveyi challenge group (Control-PI2), extensive fissure-like necrotic regions were present in the intestinal mucosa. These cavities arose from ischemic or inflammatory necrosis and lysis of mucosal tissues, accompanied by marked infiltration of lymphocytes, macrophages, and other inflammatory cells, together with interstitial edema. By contrast, only mild inflammatory cell infiltration was observed in the intestines of fish in the carvacrol-pretreated challenge group (CP-I2). In the treatment experiment, fish subjected to 5 days of V. harveyi immersion challenge (HI) displayed severe morphological damage to intestinal villi, including structural disruption and loss of typical finger-like projections. Columnar epithelial cells were disorganized, with evident vacuolation, focal necrosis, and shedding, leading to interrupted cellular continuity. Goblet cell numbers were decreased and unevenly distributed. The lamina propria exhibited massive inflammatory cell infiltration, accompanied by severe interstitial edema, loosening, and local hyperemia. Fish in the 4-day carvacrol treatment group (CT-4) exhibited numerous goblet cells within the intestinal epithelium. In the 7-day carvacrol treatment group (CT-7), only mild nuclear pyknosis, inflammatory cell infiltration, and edema were detected, with no notable hemorrhage or tissue necrosis (Figure 4).
3.2. Changes in Intestinal Vibrio Load in Sebastes schlegelii Under Pathogen Infection and Carvacrol Treatment
Conventional intestinal microbiota sequencing and reverse-transcription intestinal microbiota sequencing were conducted on experimental fish from different treatment groups. The microorganisms identified at the genus level in each group were presented through community bar charts and box plots of differential genera between groups (Figure 5A-B). These plots were employed to evaluate the compositional differences between the entire intestinal microbiota and the currently bioactive intestinal microbiota of the experimental fish. The inter-group differences in Vibrio, an important pathogenic genus, are shown in Figure 5C-D.
Vibrio, the predominant pathogen in this study, exhibited intestinal abundance in each experimental fish group as determined by the two sequencing approaches, as summarized in Table 2. The results indicated that in the prophylactic trial, the intestinal Vibrio loads in fish from the carvacrol-pretreated V. harveyi challenge group (CP-I2) were significantly reduced by both detection methods when compared with those in the direct challenge group without pretreatment (Control-PI2). In the therapeutic trial, the DNA extraction and sequencing method revealed that the intestinal Vibrio loads in fish from the 4-day treatment group (CT-4) were already markedly decreased relative to those in the immersion challenge group (HI). Meanwhile, the RNA extraction and reverse transcription sequencing method showed that the intestinal Vibrio loads in fish from the 7-day carvacrol treatment group were also substantially reduced (CT-7).
3.3. Changes in Non-Specific Immunity of Experimental Fish in Each Treatment Group
The expression levels of midgut immune genes (NFκB, IL15, and IL12b) and hepatic enzyme activities (ACP, AKP, SOD, CAT, MDA) in experimental fish from different groups are shown in Figure 6. The results indicated that the intestinal expression of NFκB in the CP-7 group was significantly upregulated compared with that in the Control-P. In contrast, there were no significant differences in the expression levels of IL15 and IL12b between the two groups.
In the experimental fish that received carvacrol prevention and subsequently challenge with V. harveyi for 1 day or 2 day (CP-I1 and CP-I2), the expression of NFκB was significantly upregulated compared to that in the experimental fish from the control group directly subjected to 1 day or 2 day V. harveyi challenge (Control-PI1 and Control-PI2 groups, respectively). Additionally, the expression level of IL15 in the CP-I2 group was significantly upregulated compared with that in the Control-PI2 group.
The expression level of NFκB in the group challenged with V. harveyi via immersion for 5 days (HI) was significantly upregulated compared to that in the Control-T group. In contrast, the expression level of NFκB after 4-day carvacrol treatment (CT-4) was significantly downregulated compared to the HI group. There was no significant difference in the expression level of IL15 between HI and Control-T. However, the expression levels of IL15 in the CT-4 and CT-7 groups were significantly upregulated when compared to that in the HI group. Regarding IL12b, its expression level in the HI group was significantly upregulated compared to that in the Control-T group, yet the expression levels of IL12b in the CT-4 and CT-7 groups were significantly downregulated compared to that in the HI group.
The activities of AKP, ACP, MDA, CAT and SOD in intestinal tissues sampled under different treatment methods and at different time points are shown in Figure 4C-D. In the prevention phase (Figure 4C), the phosphatase activities (ACP and AKP) of experimental fish in the CP-7 group were significantly upregulated compared to those in the Control-P group. The SOD and CAT activities in the CP-7 group were upregulated relative to the Control-P group, although the difference was not significant. The MDA content in the CP-7 group was downregulated compared to the Control-P group, and this difference was also not statistically significant. The ACP activity in the CP-I1 group was significantly upregulated compared to the Control-PI1 group, while the ACP activity in the CP-I2 group was upregulated compared to theControl-PI2 group, but the difference was not significant. The AKP activities in the CP-I1 and CP-I2 groups were significantly upregulated compared to the Control-PI1 and Control-PI2 groups, respectively. The SOD and CAT activities in the CP-I1 and CP-I2 groups were upregulated compared to their corresponding control groups (Control-PI1 and Control-PI2), but the differences were not significant. The MDA contents in the CP-I1 and CP-I2 groups were downregulated compared to their corresponding control groups, but the differences were not significant. In the treatment section (Figure 4D), the activities of ACP and AKP in experimental fish of the HI group were upregulated compared to those in the control group (Control-T). After carvacrol treatment, the activities of ACP and AKP exhibited a trend of significant downregulation first, followed by significant upregulation. There were no significant differences in the activities of SOD, MDA, and CAT among all groups.
3.4. Expression of Specific Immune Cells in Tissues and Peripheral Blood After Prophylactic Administration of Carvacrol
After 7-day prophylactic administration of carvacrol, immunohistochemical assays were performed on the liver, spleen, and head kidney of experimental fish. The percentages of CD4-1+ T cells and CD79a/CD79b+ B cells in the relevant tissues were determined by calculating the ratio of positive cells to total cells in the sections, so as to investigate the effect of carvacrol on specific immunity in experimental fish.
As shown in Figure 7, compared with the blank control group, the ratios of CD4-1+ T cells and CD79a/CD79b+ B cells in the liver, CD79a/CD79b+ B cells in the spleen, and CD4-1+ T cells and CD79a/CD79b+ B cells in the head kidney were significantly higher in the fish that were fed carvacrol for prophylaxis.
Flow cytometry was used to sort and count peripheral blood leukocytes of Sebastes schlegelii to further investigate the enhancement of adaptive immunity by carvacrol. After lymphocyte gating and doublet exclusion, sorting was performed via the FITC fluorescence channel, and the results are shown in Figure 8. As shown in the figure, the percentages of CD4-1+ T cells, CD79a+ B cells, and CD79b+ B cells in peripheral blood leukocytes of Sebastes schlegelii in the 7-day carvacrol prophylaxis group (CP-7) were significantly higher than those in the blank control group.
4. Discussion
As shown in the survival rate results, feeding with carvacrol can increased the survival rate of S. schlegelii after immersion infection with V. harveyi. Pathological tissue sections indicated that treating with carvacrol after V. harveyi infection could effectively alleviate phenomena such as aggregation of intestinal inflammatory cells, atrophy and shedding of intestinal villus, hemorrhage of intestinal tissue, and pyknosis of intestinal cell nuclear. These experimental results demonstrate that carvacrol can assist S. schlegelii in resisting the infection of V. harveyi.
NF-κB initiates and regulates the innate immune response to infection by modulating the expression of cytokines (Pittet, Quinton et al. 2011) . Existing studies have clearly demonstrated that IL-15 can maintain the homeostatic proliferation of memory CD8+ T cells and the persistence of CD8+ memory T cells. Moreover, it can also sustain the proliferative capacity of T cells and NK cells in vertebrates via the STAT5 signaling pathway (Drake, Kaur et al. 2016, Clark, Burrack et al. 2019). The interleukin-12b (IL-12b) gene encodes the p40 chain of IL-12 (Xu, Pei et al. 2013). Recent genomic research has identified IL12b as a susceptibility locus for inflammatory bowel disease (IBD) (Márquez, Mendoza et al. 2008). The findings of this study show that carvacrol intervention modifies the intestinal expression levels of NF-κB, IL-15 and IL-12b in experimental fish challenged with V. harveyi, suggesting that carvacrol might exert a regulatory influence on the host inflammatory response.
The results of enzyme activity expression showed that both V. harveyi and carvacrol could influence the activities of ACP and AKP in the intestine of S. schlegelii, yet they had no significant impact on SOD, MDA and CAT. This finding suggests that the protective effect of carvacrol against V. harveyi stress is mediated via a non-oxidative defense mode.
The results obtained through conventional DNA extraction and sequencing encompass viable, dead, and dormant microbes in the intestine, making this approach suitable for profiling the species composition and relative proportions of intestinal microorganisms. In contrast, the reverse transcription method involves isolating labile intestinal RNA and reverse-transcribing it into complementary DNA (cDNA) before sequencing. This strategy solely reflects the composition of metabolically active intestinal microbes, thus facilitating the analysis of the activity status and functional potential of the intestinal microbial community. Analysis revealed that Vibrio was the microbial species with significant differences in the intestine of experimental fish among groups as determined by both methods. The abundance of Vibrio in the challenged groups (HI, Control-PI2) measured by the two methods was significantly higher than that in the Control-P and Control-T groups, suggesting that both a 5-day immersion challenge with V. harveyi and a 2-day injection challenge significantly increased the intestinal Vibrio abundance of S. schlegelii. In the prevention section, the relative abundance of Vibrio in the CP-I2 group was substantially lower than that in the Control-PI2 group. In the treatment section, the results of the conventional method demonstrated that the Vibrio abundance in the CT-4 and CT-7 groups was significantly lower than that in the HI group; the reverse transcription method showed that the Vibrio abundance in the CT-7 group was significantly lower than that in the HI group. Consistent findings from the two relative quantification methods for pathogenic bacteria demonstrated that carvacrol could effectively inhibit the proliferation of Vibrio in the intestine of Sebastes schlegelii and improve host resistance against pathogenic bacterial invasion.
Regarding the effect on immunity, administration of carvacrol significantly increased the expression of CD4-1+ T cells and CD79a/CD79b+ B cells in visceral tissues of experimental fish. As key components of the immune system, CD4+ T cells play a central role in coordinating adaptive immune responses (Maisey, Montero et al. 2016, Zhu and Zhu 2020). CD79 molecules are specific to B cells and are expressed during most developmental stages. Encoded by the CD79a and CD79b genes, they form a heterodimeric signaling unit within the B-cell receptor complex, responsible for transmitting signals for B-cell activation, proliferation, and differentiation(Huang, Takata et al. 2011, Huse, Bai et al. 2022). The proportions of helper T cells and B lymphocytes in the total cell population can be determined via specific binding of CD4-1, CD79a, and CD79b antibodies to cell surface antigens, which further reflects the activation of cellular and humoral immunity in the organism. Immunohistochemical (IHC) results showed that in fish receiving 7 days of carvacrol prophylaxis, the percentage of CD4-1+ T cells was significantly elevated in all detected tissues except the spleen, where no significant difference was observed compared with the control group. The percentages of CD79a/CD79b+ B cells were significantly increased in the liver, spleen, and head kidney of all treatment groups, and the positive cell trends detected by CD79a and CD79b were consistent. Flow cytometric analysis of peripheral blood revealed that carvacrol significantly elevated the proportions of CD4-1+ T cells and CD79a/CD79b+ B cells in peripheral blood of experimental fish. In conclusion, dietary administration of 500 mg/kg carvacrol can activate the expression of T cells and B cells in experimental fish, thereby enhancing the efficiency of cellular and humoral immune responses against pathogenic stress.
5. Conclusions
In this study, S. schlegelii was selected as the experimental subject to thoroughly explore the preventive and immune-enhancing effects of carvacrol on uninfected juvenile S. schlegelii as well as its therapeutic effect following pathogen infection. The results demonstrated that the administration of carvacrol could enhance the resistance of S. schlegelii to V. harveyi stress in farming environments, consequently improving the survival rates. This enhancement was accomplished through multiple mechanisms, including the improvement of intestinal histological morphology, the regulation of the inflammation-related genes expression and phosphatase activity, the reconstruction of the intestinal microbial community, the reduction of Vibrio abundance, and the enhancement of the adaptive immunity, as well as the increase in the proportions of CD4-1+ T cells and CD79a/CD79b+ B cells in key immune organs and peripheral blood. Collectively, these findings offer empirical data to support the healthy cultivation of S. schlegelii and promote the subsequent screening of alternative agents to substitute antibiotics.
Author Contributions
T.W.: Data curation, Validation. Formal analysis, Methodology, Writing-original draft. Y.Y.: Investigation, Methodology, Software. C.W.: Investigation, Validation. Y.W.: Funding acquisition, Methodology, Formal analysis. Z.Z.: Methodology, Formal analysis, Software. X.R.: Funding acquisition, Formal analysis. M.L.: Formal analysis, Methodology. J.G.: Data curation, Software. B.L.: Data curation, Methodology. J.W.: Methodology. K.H.: Formal analysis, Methodology. Z.Z.: Conceptualization, Writing-review & editing, Methodology, Software, Funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Key Research and Development Program [grant numbers 2023YFD2400704]; the Key Research and Development Program of Guangxi Province [grant number GUIKENONG-AB2506910047]; and the Central Public-Interest Scientific Institution Basal Research Fund [grant number 20603022025012 and 2023TD29].
Institutional Review Board Statement
The research in this manuscript has been conducted under Institutional Animal Care and Use Committee, IACUC (approval code: YSFRI-2024059 and approval date: 7 April 2024).
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding author.
References
- Ahmadifar, E.; Mansour, M. R.; Amirkolaie, A. K.; Rayeni, M. F. Growth efficiency, survival and haematological changes in great sturgeon (Huso huso Linnaeus, 1758) juveniles fed diets supplemented with different levels of thymol-carvacrol. Animal Feed Science and Technology 2014, 198, 304–308. [Google Scholar] [CrossRef]
- Alagawany, M.; Farag, M. R.; Salah, A. S.; Mahmoud, M. A. The role of oregano herb and its derivatives as immunomodulators in fish. Reviews in Aquaculture 2020, 12(4), 2481–2492. [Google Scholar] [CrossRef]
- Bhat, R. A. H.; Khangembam, V. C. C.; Pant, V.; Tandel, R. S.; Pandey, P. K.; Thakuria, D. Antibacterial Potential of a De-Novo Designed Peptide Against Bacterial Fish Pathogens. International Journal of Peptide Research and Therapeutics 2023, 29(5). [Google Scholar] [CrossRef]
- Bloomfield, S. J.; Palau, R.; Janecko, N.; Baker-Austin, C.; Mather, A. E. Association of antimicrobial resistant Vibrio and species pathogenic to humans with aquacultured seafood. Food Microbiology 2025, 132. [Google Scholar] [CrossRef] [PubMed]
- Clark, S. E.; Burrack, K. S.; Jameson, S. C.; Hamilton, S. E.; Lenz, L. L. NK Cell IL-10 Production Requires IL-15 and IL-10 Driven STAT3 Activation. Frontiers in Immunology 2019, 10. [Google Scholar] [CrossRef]
- Coelho, J. D.; Rosa, K. V.; Rocha, J. S.; Ramirez, N. C. B.; Maraschin, M.; Vieira, F. D. IN VITRO ANTIMICROBIAL ACTIVITY OF CARVACROL AGAINST SHRIMP PATHOGENS AND ITS USE AS FEED ADDITIVE FOR THE PACIFIC WHITE SHRIMP; <u>Boletim Do Instituto De Pesca</u> <b>47</b>, 2021. [Google Scholar] [CrossRef]
- Das, S.; Chourashi, R.; Mukherjee, P.; Kundu, S.; Koley, H.; Dutta, M.; Mukhopadhyay, A. K.; Okamoto, K.; Chatterjee, N. S. Inhibition of growth and virulence of Vibrio cholerae by carvacrol, an essential oil component of Origanum spp. Journal of Applied Microbiology 2021, 131(3), 1147–1161. [Google Scholar] [CrossRef]
- Deng, H.; Deng, Y.; Song, T.; Pang, L.; Zhu, S.; Ren, Z.; Guo, H.; Xu, Z.; Zhu, L.; Geng, Y.; Ouyang, P.; He, R.; Deng, J. Evaluation of the activity and mechanisms of oregano essential oil against PRV in vivo and in vitro. Microbial Pathogenesis 2024, 194. [Google Scholar] [CrossRef]
- Drake, A.; Kaur, M.; Iliopoulou, B. P.; Phennicie, R.; Hanson, A.; Chen, J. Z. Interleukins 7 and 15 Maintain Human T Cell Proliferative Capacity through STAT5 Signaling. Plos One 2016, 11(11). [Google Scholar] [CrossRef]
- Hirose, S.; Horiyama, S.; Morikami, A.; Fujiwara, K.; Tsukagoshi, H. Eugenol and basil essential oil as priming agents for enhancing Arabidopsis immune response. Bioscience Biotechnology and Biochemistry 2024, 89(1), 41–50. [Google Scholar] [CrossRef]
- Huang, X. G.; Takata, K.; Sato, Y.; Tanaka, T.; Ichimura, K.; Tamura, M.; Oka, T.; Yoshino, T. Downregulation of the B-cell receptor signaling component CD79b in plasma cell myeloma: A possible post transcriptional regulation. Pathology International 2011, 61(3), 122–129. [Google Scholar] [CrossRef]
- Huse, K.; Bai, B. Y.; Hilden, V. I.; Bollum, L. K.; Våtsveen, T. K.; Munthe, L. A.; Smeland, E. B.; Irish, J. M.; Wälchli, S.; Myklebust, J. H. Mechanism of CD79A and CD79B Support for IgM plus B Cell Fitness through B Cell Receptor Surface Expression. Journal of Immunology 2022, 209(10), 2042–2053. [Google Scholar] [CrossRef]
- Imtiaz, N.; Anwar, Z.; Waiho, K.; Shi, C.; Mu, C.; Wang, C.; Wu, Q. A review on aquaculture adaptation for fish treatment from antibiotic to vaccine prophylaxis. Aquaculture International 2024, 32(3), 2643–2668. [Google Scholar] [CrossRef]
- Jeong-hyeon, C.; Gyeong-mi, W.; Sae-bom, S.; Hyo-jin, P.; Soon-kyu, B.; Bae-ik, L.; Jong-ha, L.; I-cheong, K.; Su-il, P. Pathogenicity of Vibrio harveyi to black rockfish, Sebastes schlegeli. Journal of Fish Pathology 2007, 20(2), 99–108. [Google Scholar]
- Ma, P.; Chen, Y.; Shao, N.; Zhang, J.; Wu, Y.; Liu, R. Synergistic Combination of Biodegradable Peptide Polymer and Curcumin as Promising Antibiotic Substitution in Aquaculture to Alleviate the Global Challenge of Antimicrobial Resistance. Chinese Journal of Chemistry 2022, 40(24), 2947–2955. [Google Scholar] [CrossRef]
- Maisey, K.; Montero, R.; Corripio-Miyar, Y.; Toro-Ascuy, D.; Valenzuela, B.; Reyes-Cerpa, S.; Sandino, A. M.; Zou, J.; Wang, T. H.; Secombes, C. J.; Imarai, M. Isolation and Characterization of Salmonid CD4+ T Cells. Journal of Immunology 2016, 196(10), 4150–4163. [Google Scholar] [CrossRef]
- Manchanayake, T.; Mohamad, A.; Amir-Danial, Z.; Abu Nor, N.; Yong-Kit, C.; Nazarudin, M. F.; Nor, R. M.; Hasnan, Q.; Zamri-Saad, M.; Amal, M. N. A.; Ina-Salwany, M. Y.; Salleh, A. Oral adjuvanted Vibrio vaccine enhances antibody production and lysozyme activity in the serum and mucus of marine-cultured red hybrid tilapia (Oreochromis sp.) against V. harveyi and V. alginolyticus. Fish & Shellfish Immunology 2025, 165. [Google Scholar] [CrossRef] [PubMed]
- Márquez, A.; Mendoza, J. L.; Toxonera, C.; Díaz-Rubio, M.; de la Concha, E. G.; Urcelay, E.; Martínez, A. IL23R and IL12B polymorphisms in Spanish IBD patients:: No evidence of interaction. Inflammatory Bowel Diseases 2008, 14(9), 1192–1196. [Google Scholar] [CrossRef]
- Milijasevic, M.; Veskovic-Moracanin, S.; Milijasevic, J. B.; Petrovic, J.; Nastasijevic, I. Antimicrobial Resistance in Aquaculture: Risk Mitigation within One Health Context. Preprints 2024. [Google Scholar] [CrossRef]
- Pan, T.; Yan, M. The screening of traditional Chinese herbs on nonspecific immune response and protection of Pacific white shrimp (Litopenaeus vannamei) from Vibrio harveyi infection. Aquaculture International 2020, 28(2), 767–776. [Google Scholar] [CrossRef]
- Pittet, L. A.; Quinton, L. J.; Yamamoto, K.; Robson, B. E.; Ferrari, J. D.; Algül, H.; Schmid, R. M.; Mizgerd, J. P. Earliest Innate Immune Responses Require Macrophage RelA during Pneumococcal Pneumonia. American Journal of Respiratory Cell and Molecular Biology 2011, 45(3), 573–581. [Google Scholar] [CrossRef] [PubMed]
- Prazdnova, E.; Zaikina, A.; Neurov, A.; Mazanko, M.; Ranjan, A.; Rudoy, D. Bacillibactin, a Potential Bacillus-Based Antibacterial Non-Ribosomal Peptide: In Silico Studies for Targeting Common Fish Pathogens. International Journal of Molecular Sciences 2025, 26(12). [Google Scholar] [CrossRef] [PubMed]
- Rossi, B.; Esteban, M. A.; Garcia-Beltran, J. M.; Giovagnoni, G.; Cuesta, A.; Piva, A.; Grilli, E. Antimicrobial Power of Organic Acids and Nature-Identical Compounds against Two Vibrio spp.: An In Vitro Study. Microorganisms 2021, 9(5). [Google Scholar] [CrossRef]
- Strem, R.; Meiri-Ashkenazi, I.; Segal, N. a.; Ehrlich, R.; Shashar, N.; Sharon, G. Evaluation of Flathead Grey Mullets (Mugil cephalus) Immunization and Long-Term Protection against Vibrio harveyi Infection Using Three Different Vaccine Preparations. International Journal of Molecular Sciences 2023, 24(9). [Google Scholar] [CrossRef]
- Tayyab, M.; Islam, W.; Waqas, W.; Zhang, Y. Probiotic-Vaccine Synergy in Fish Aquaculture: Exploring Microbiome-Immune Interactions for Enhanced Vaccine Efficacy. Biology-Basel 2025, 14(6). [Google Scholar] [CrossRef]
- van Gelderen, T. A.; Debnath, P.; Joly, S.; Bertomeu, E.; Duncan, N.; Furones, D.; Ribas, L. Gonadal miRNomes and transcriptomes in infected fish reveal sexually dimorphic patterns of the immune response. Functional & Integrative Genomics 2025, 25(1). [Google Scholar] [CrossRef]
- Wan, X.; Da, F.; Lin, G.; Huang, Y.; Cai, S.; Jian, J. Transcriptome Analysis of the Cultured Hybrid Grouper (♀Epinephelus fuscoguttatus x♂E. lanceolatus) Immunized with Vibrio harveyi formalin-killed cells vaccine (FKC) combined with chitosan oligosaccharide. Israeli Journal of Aquaculture-Bamidgeh 2023, 75(2). [Google Scholar] [CrossRef]
- Wang, J.; Mai, K.; Ai, Q. Conventional Soybean Meal as Fishmeal Alternative in Diets of Japanese Seabass (Lateolabrax japonicus): Effects of Functional Additives on Growth, Immunity, Antioxidant Capacity and Disease Resistance. Antioxidants 2022, 11(5). [Google Scholar] [CrossRef]
- Wang, Q.; Huang, J.; Li, Y.; Wu, S.; Zhao, L.; Pan, Y.; Kang, Y.; Liu, Z. Chinese herbal medicines mixture improved antioxidant enzymes, immunity and disease resistance to infectious hematopoietic necrosis virus infection in rainbow trout (Oncorhynchus mykiss). Aquaculture International 2024, 32(3), 3217–3232. [Google Scholar] [CrossRef]
- Wu, Q.; Jiang, Y.; Chen, E.; Mu, C.; Waiho, K. Chinese gallnut (Gall chinensis) against Vibrio parahaemolyticus: In vitro activity and the use of medicated bath method to treat infected mud crab Scylla paramamosain. Aquaculture 2021, 539. [Google Scholar] [CrossRef]
- Xu, M.; Li, H.; Wu, X.; Dai, L.; Yang, R.; Zhang, Q.; Zhang, Z.; Lei, L.; Zhang, F. Inhibitory effect of arctigenin on reovirus infection in grass carp. Aquaculture 2024, 593. [Google Scholar] [CrossRef]
- Xu, Z.; Pei, L.; Zhang, F.; Hu, X.; Gui, Y.; Wang, L.; Wu, B. A functional variant in IL12B promoter modulates its expression and associates with increased risk of allergic asthma. Genes and Immunity 2013, 14(4), 238–243. [Google Scholar] [CrossRef]
- Yang, X.; Cheng, Y.; Zhao, Y.; Zhu, H.; Zhang, Y.; Zhao, Y.; Liu, X. Comparative transcriptome analysis reveals immunoregulation mechanism in the spleen of large yellow croaker (Larimichthys crocea) in response to Vibrio harveyi infection. Aquaculture Reports 2023, 31. [Google Scholar] [CrossRef]
- Yu, B.; Sun, Z.; Li, X. C.; Qv, A.; Sohail, M.; Li, Y. L.; Xu, H.; Xiang, P. Research Progress of Novel Drug Delivery Systems of Chinese Medicine Monomers based on Natural Silk Fibroin: A Mini-Review. Current Drug Delivery 2023, 20(3), 211–222. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B. Z.; Qiu, Y. L.; Shi, C. X.; Zhang, J. Development of Multiple Real-Time Fluorescent Quantitative PCR for Vibrio Pathogen Detection in Aquaculture. Veterinary Sciences 2025, 12(4). [Google Scholar] [CrossRef]
- Zhou, F.; Ji, B. P.; Zhang, H.; Jiang, H.; Yang, Z. W.; Li, J. J.; Li, J. H.; Yan, W. J. The antibacterial effect of cinnamaldehyde, thymol, carvacrol and their combinations against the foodborne pathogen Salmonella typhimurium. Journal of Food Safety 2007, 27(2), 124–133. [Google Scholar] [CrossRef]
- Zhu, X. L.; Zhu, J. F. CD4 T Helper Cell Subsets and Related Human Immunological Disorders. International Journal of Molecular Sciences 2020, 21(21). [Google Scholar] [CrossRef]
- Zhu, Z.-X.; Yao, Y.-Y.; Ai, C.-H.; Yang, G.; Liang, X.-Y.; Liu, T.-D.; He, M.-L.; Xia, J.-H. Dietary supplementation of Patchouli oil increases the resistance against Streptococcus agalactiae infection in GIFT tilapia as revealed by transcriptome and 16s amplicon profiling. Aquaculture Reports 2023, 33. [Google Scholar] [CrossRef]
Figure 1.
Experimental fish sampling flow chart.

Figure 2.
Apparent symptoms of experimental fish in the 2-day intramuscular injection challenge group (Control-PI2) and the 5-day Vibrio harveyi immersion challenge group (HI). A-B: External appearance images of experimental fish in the Control-PI2 group and HI group; C-D: Local symptom images of experimental fish in the Control-PI2 group and HI group; E-F: Anatomical images of experimental fish in the Control-PI2 group and HI group.
Figure 2.
Apparent symptoms of experimental fish in the 2-day intramuscular injection challenge group (Control-PI2) and the 5-day Vibrio harveyi immersion challenge group (HI). A-B: External appearance images of experimental fish in the Control-PI2 group and HI group; C-D: Local symptom images of experimental fish in the Control-PI2 group and HI group; E-F: Anatomical images of experimental fish in the Control-PI2 group and HI group.

Figure 3.
Survival rates of experimental fish in different groups. Contro-P: Control group in the prevention trial. CP: Carvacrol prevention group. Control-PI2: Group with 7-day blank diet feeding followed by 2-day V.harveyi injection challenge. CP-I2: Group with 7-day feeding of 500 mg/kg carvacrol-supplemented diet followed by 2-day V. harveyi injection challenge. Control-T: Control group in the treatment trial. HI: Group subjected to 5-day V. harveyi immersion challenge. CT: Carvacrol-treated therapeutic group.
Figure 3.
Survival rates of experimental fish in different groups. Contro-P: Control group in the prevention trial. CP: Carvacrol prevention group. Control-PI2: Group with 7-day blank diet feeding followed by 2-day V.harveyi injection challenge. CP-I2: Group with 7-day feeding of 500 mg/kg carvacrol-supplemented diet followed by 2-day V. harveyi injection challenge. Control-T: Control group in the treatment trial. HI: Group subjected to 5-day V. harveyi immersion challenge. CT: Carvacrol-treated therapeutic group.

Figure 4.
Intestinal pathological sections of experimental fish in each group (20× magnification). A: Intestinal pathological section of fish in the control group of the prevention trial (Control-P); B: Intestinal pathological section of fish in the carvacrol prevention 7-day group (CP-7); C: Intestinal pathological section of fish in the group injected with V. harveyi for 2 days after 7 days of feeding with a blank diet (Control-PI2); D: Intestinal pathological section of fish in the group injected with V. harveyi for 2 days after 7-day feeding with carvacrol-supplemented diet (CP-I2); E: Intestinal pathological section of fish in the control group of the treatment trial (Control-T); F: Intestinal pathological section of fish in the group immersed with V. harveyi for 5 days (HI); G: Intestinal pathological section of fish in the carvacrol treatment 4-day group after challenge (CT-4); H: Intestinal pathological section of fish in the carvacrol treatment 7-day group after challenge (CT-7).
Figure 4.
Intestinal pathological sections of experimental fish in each group (20× magnification). A: Intestinal pathological section of fish in the control group of the prevention trial (Control-P); B: Intestinal pathological section of fish in the carvacrol prevention 7-day group (CP-7); C: Intestinal pathological section of fish in the group injected with V. harveyi for 2 days after 7 days of feeding with a blank diet (Control-PI2); D: Intestinal pathological section of fish in the group injected with V. harveyi for 2 days after 7-day feeding with carvacrol-supplemented diet (CP-I2); E: Intestinal pathological section of fish in the control group of the treatment trial (Control-T); F: Intestinal pathological section of fish in the group immersed with V. harveyi for 5 days (HI); G: Intestinal pathological section of fish in the carvacrol treatment 4-day group after challenge (CT-4); H: Intestinal pathological section of fish in the carvacrol treatment 7-day group after challenge (CT-7).

Figure 5.
A-B: Bar charts depicting the relative abundance of intestinal microbiota in the experimental fish of each group. C-D: Differences in Vibrio abundance between the groups, detected using the conventional method and reverse-transcription method respectively.
Figure 5.
A-B: Bar charts depicting the relative abundance of intestinal microbiota in the experimental fish of each group. C-D: Differences in Vibrio abundance between the groups, detected using the conventional method and reverse-transcription method respectively.

Figure 6.
Expression levels of inflammation-related genes and hepatic enzyme activities in each group. A: Expression levels of inflammation-related genes in the prevention phase. B: Expression levels of inflammation-related genes in the treatment section groups. C: Hepatic enzyme activities of experimental fish from different groups in the prevention phase. D: Hepatic enzyme activities of experimental fish from different groups in the treatment phase.
Figure 6.
Expression levels of inflammation-related genes and hepatic enzyme activities in each group. A: Expression levels of inflammation-related genes in the prevention phase. B: Expression levels of inflammation-related genes in the treatment section groups. C: Hepatic enzyme activities of experimental fish from different groups in the prevention phase. D: Hepatic enzyme activities of experimental fish from different groups in the treatment phase.

Figure 7.
Changes in the proportions of CD4-1+ T cells, CD79a+ B cells and CD79b+ B cells in various tissues of the group with 7-day carvacrol prevention.
Figure 7.
Changes in the proportions of CD4-1+ T cells, CD79a+ B cells and CD79b+ B cells in various tissues of the group with 7-day carvacrol prevention.

Figure 8.
Changes in CD4-1+ T cells, CD79a+ B cells and CD79b+ B cells in peripheral blood of experimental fish with 7-day carvacrol prevention.
Figure 8.
Changes in CD4-1+ T cells, CD79a+ B cells and CD79b+ B cells in peripheral blood of experimental fish with 7-day carvacrol prevention.


Table 1.
Target genes and their primers for q-PCR.
| Target Gene | Primer | Nucleotide Sequence 5′-3′ |
|---|---|---|
| RPL-17 | RPL17-F | AGGCGACGCACCTACCG |
| RPL17-R | CCTCTGGTTTGGGGACGA | |
| NFκB/RelA | NFκB/RelA-F | TGTCGTAGATGGGGTTGGA |
| NFκB/RelA-R | AGGAGCTGGGGAAGGTGAT | |
| IL-12b | IL-12b-F | CTCTGGCATCCTTATCAGTTCA |
| IL-12b-R | GTCTTGGTTGCTGGCGTAG | |
| IL-15 | IL-15-F | CGCCTACAATACAACTAAAGAGC |
| IL-15-R | AGATGACGGAGCATACAGCA |
Table 2.
Intestinal abundance of Vibrio in the experimental fish from each treatment group.
| Group | Vibrio Abundance Determined by DNA Extraction Method (%) | Vibrio Abundance Determined by RNA Extraction and Reverse Transcription (%) |
|---|---|---|
| Control-P | <0.06 | 42.08 |
| CP-7 | 0.32 | 38.25 |
| Control-PI2 | 46.59 | 92.54 |
| CP-I2 | 16.88 | 4.59 |
| Control-T | 0.06 | 31.12 |
| HI | 48.55 | 78.27 |
| CT-4 | 0.84 | 80.14 |
| CT-7 | 0.95 | 29.71 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



