Submitted:
04 March 2026
Posted:
09 March 2026
You are already at the latest version
Abstract
Neutrophils, once viewed as short-lived and functionally uniform antimicrobial cells, are now recognized as highly plastic immune regulators with context-dependent roles in cancer. Among circulating neutrophil subsets, low-density neutrophils (LDNs) have emerged as a clinically relevant population in oncology due to their distinct phenotypic, potent immunosuppressive activity, and strong association with disease progression. LDNs are rare in healthy individuals but markedly expanded in patients with solid tumors.LDNs display enhanced activation, increased expression of immunoregulatory molecules such as programmed death‑ligand 1, elevated reactive oxygen species production, and a heightened propensity for neutrophil extracellular trap formation, collectively impairing cytotoxic T cell function and promoting tumor immune evasion.Although LDNs have been described across multiple malignancies, their biological and clinical relevance has been most characterized in breast and lung cancers. This review synthesizes current knowledge on neutrophil biology, emphasizing the clinical and mechanistic features of LDNs, with a focus on these two tumor types.In breast cancer, increased LDNs abundance correlates with metastatic disease, poor response to neoadjuvant chemotherapy, immunosuppression, accelerated progression, and reduced survival. In lung cancer, elevated LDNs frequencies similarly associate with advanced disease stage, unfavorable clinical outcomes, and distinct surface marker profiles distinguishing them from high-density neutrophils.Mechanistic studies indicate that tumor-derived cytokines and chronic inflammation drive LDNs expansion, functional reprogramming, and tissue trafficking, linking systemic neutrophil dysregulation to tumor progression.Finally, we highlight emerging biomarkers, unresolved challenges, and therapeutic strategies targeting neutrophil plasticity and LDN-mediated immunosuppression, underscoring their potential as actionable components in precision oncology.
Keywords:
low density neutrophils (LDNs)
; breast cancer
; lung cancer
; immunosuppression
; biomarkers
Summary Sentence
Low-density neutrophils are a distinct circulating subset that expands in breast and lung cancer, driving tumor progression, therapy resistance, and metastasis through heightened ROS production, NET formation, and promotion of immune evasion; their increased abundance consistently correlates with poor patient outcomes.
Key Concepts
- Historically regarded as bystanders, neutrophils are now well-established key players in cancer
- In cancer, LDNs exhibit activated, immunosuppressive phenotypes that promote tumor progression.
- In breast cancer, elevated LDNs correlate with metastatic disease, poor therapeutic response, farter disease progression and worse prognosis.
- In lung cancer, increased LDN levels are also associated with advanced disease stage and poorer clinical outcomes.
- LDNs are emerging as accessible biomarkers and promissing therapeutic targets in precision oncology.
Open Questions
- Which molecular pathways drive the generation, expansion, and persistence of LDNs in cancer?
- How can LDN subpopulations be consistently defined and standardized across studies?
- Which LDN associated markers most accurately predict prognosis or treatment response?
- How should LDN quantification be incorporated into clinical decision making in breast and lung cancer?
- Through which mechanisms can LDN expansion or function be therapeutically modulated?
Introduction
Neutrophils are the most abundant circulating leukocytes in human blood and represent a frontline component of the innate immune system [1,2]. Traditionally, they have been recognized for their pivotal roles in host defense, including rapid recruitment to sites of infection or injury, phagocytosis of pathogens, generation of reactive oxygen species (ROS), and release of antimicrobial peptides and neutrophil extracellular traps (NETs). These classical functions underscore their importance in maintaining immune homeostasis and preventing microbial dissemination [3,4,5,6].
Once considered short-lived, homogeneous cells solely dedicated to pathogen clearance [7], neutrophils are now known to be highly plastic and phenotypically diverse, capable of adapting to a broad range of environment-dependent signals. This functional versatility has placed neutrophils at the forefront of cancer research [8].
In the context of cancer, neutrophils exhibit a dual role, capable of either promoting or suppressing tumor progression. Depending on the tumor microenvironment (TME) and systemic cues, they can facilitate tumor growth through immunosuppression, promotion of angiogenesis and metastasis or contribute to anti-tumor immunity by directly killing malignant cells and supporting adaptive immune responses [9].
Within the TME, at least two distinct populations of tumor-associated neutrophils (TANs) have been described: the anti-tumor N1 phenotype, characterized by cytotoxic and tumor inhibiting activity, and the pro-tumor N2 phenotype, which supports tumor growth, angiogenesis, and immune evasion [10,11]. This renewed understanding of neutrophil biology has opened new possibilities for cancer therapy. However, critical challenges remain, particularly in distinguishing between pathological and beneficial neutrophil populations, making it difficult to target or modulate these cells without unintended effects.
Systemically, similar to the N1 and N2 subsets observed in the TME, two distinct populations of circulating neutrophils have been identified based on density: high-density neutrophils (HDNs), corresponding to conventional mature neutrophils and low-density neutrophils (LDNs), a heterogeneous population displaying altered phenotypes and functions frequently associated with pathological conditions. [12,13,14].
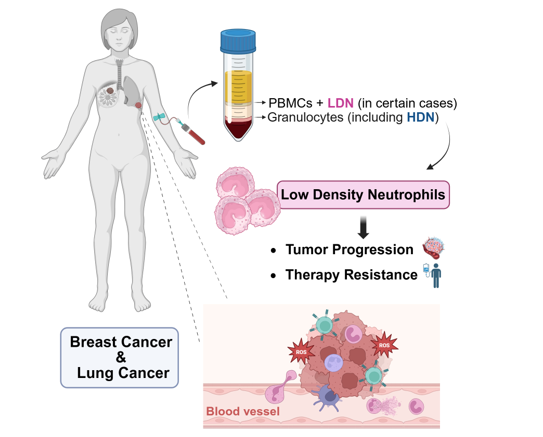
Within this landscape, LDNs have emerged as a particularly relevant subset in immune-oncology. These cells co-purify with peripheral blood mononuclear cells (PBMC) during density gradient centrifugation and are nearly absent in healthy individuals, but markedly enriched in cancer patients [12,15,16]. LDNs have been described across multiple solid tumors, including hepatocellular carcinoma, melanoma, colorectal, breast and lung cancers, with breast and lung malignancies providing the most comprehensive characterization of their association with disease progression and therapeutic response.
In breast cancer, elevated circulating LDNs correlate with metastatic disease and poor response to neoadjuvant chemotherapy [12]. Notably, in the metastatic BC, increased LDN levels were associated with faster disease progression and reduced overall survival [17]. Accordingly, increased circulating LDN abundance also inversely correlated with activated cytotoxic T lymphocytes (CTLs) and positively correlated with regulatory T cells (Tregs), reflecting their immunosuppressive, tumor-promoting role [12,17]. At the cellular level, LDNs exhibit elevated expression of programmed death-ligand 1 (PD-L1), matrix metalloproteinase-9 (MMP-9), and the activation markers CD11b and CD66b [12,17]. Together with the enhanced production of reactive oxygen species (ROS), increased neutrophil extracellular trap (NET) formation and arginase activity [12,13,17], these features provide mechanistic support for their capacity to suppress antitumor immunity and facilitate metastatic dissemination.
Similar patterns have been observed in lung cancer, where LDNs levels are significantly higher in advanced disease compared with early-stage tumors, healthy controls, or patients with stable chronic obstructive pulmonary disease. High frequencies of LDNs (>10%) are strongly associated with poorer prognosis [16]. Furthermore, high-dimensional cytometry studies in non-small cell lung carcinoma (NSCLC) have identified novel distinctive surface markers, including CD36, CD41, CD61, and CD226, which in NSCLC, distinguish LDNs from the regular HDN, reinforcing their uniqueness and potential as diagnostic or therapeutic targets [18].
Given their growing clinical relevance, this review synthesizes current knowledge on LDNs biology with a particular focus on breast and lung cancers - two of the most prevalent and deadly malignancies worldwide [19], integrating clinical associations, mechanistic insights, and emerging therapeutic strategies, to elucidate the contribution of LDNs to cancer progression and to evaluate their potential as biomarkers and targets in precision oncology.
1. Overview of Neutrophils
1.1. Basic Characteristics and Lifespan
Neutrophils are polymorphonuclear granulocytes and constitute 50-70% of all circulating leukocytes, representing the most abundant white cell population in human peripheral blood. They are essential players in innate immunity [3]. These cells derive from a large granulocytic leukocyte group, and are morphologically characterized by contains a segmented nucleus with usually three to five lobes and cytoplasm rich in specialized granules that store antimicrobial molecules [20]. Traditionally, neutrophils were viewed as short-lived cells, based on ex vivo observations suggesting a lifespan of approximately eight to twelve hours in blood circulation and one day in tissues. However, recent in vivo labeling studies indicate a lifespan of five days, challenging previous beliefs [21].
1.2. Effector Mechanisms: Phagocytosis, Degranulation and NETosis
Neutrophils protect their host by detecting pathogens and tissue damage through a wide range of receptors, including selectins, integrins, chemokine receptors and pattern recognition receptors (PRRs) such as toll-like receptors (TLRs), C-type lectin receptors, and nucleotide-binding oligomerization domain-like receptors (NOD-like receptors). These receptors guide neutrophil migration through a series of adhesion steps, leading neutrophils to inflamed or infected tissues where they act through phagocytosis, degranulation, NETosis or cytokine secretion [4].
1.2.1. Phagocytosis
Pathogens are recognized through PRRs, Fcγ receptors, and complement receptor 3. Upon activation complement receptor 3 facilitates pathogen detection via opsonization, enhancing pathogen capture, and enabling neutrophils to adhere to endothelial surfaces [4].
After entering the cell, pathogens are sequestered into immature phagosomes that mature by fusing with secretory vesicles and cytoplasmic granules. This maturation is associated with the formation of the nicotinamide adenine dinucleotide phosphate (NADPH) oxidase complex that mediates superoxide radical production. These are transformed into hydrogen peroxide and later into hypochlorous acid, which is a potent antimicrobial agent [4].
1.2.2. Degranulation
PRRs not only induce phagocytosis but also play a role in neutrophil degranulation. While the two processes run independently, both rely on neutrophil cytoplasmic granules. Degranulation consists of an extracellular release of granule contents. In doing so, neutrophils release primary (azurophilic), secondary (specific), and tertiary (gelatinase) granules, packaging a variety of antimicrobial peptides and enzymes. Myeloperoxidase (MPO), neutrophil elastase (NE), proteinase 3, cathepsins, defensins, and others are stored in primary granules. The secondary granules contain lactoferrin, cathelicidin, and neutrophil gelatinase-associated lipocalin. The tertiary granules contain gelatinase, arginase-1, and ficolin I [4]. These mediators contribute to pathogen killing but may also cause collateral tissue damage if excessively released [22].
1.2.3. NETosis
If phagocytosis and degranulation are insufficient, neutrophils have another trick up their sleeves: NETosis, a type of cell death, entirely distinct from apoptosis and necrosis, that results in the release of NETs. NETs are composed of decondensed chromatin fibers decorated with antimicrobial proteins (like NE and MPO) that immobilize and neutralize invading microorganisms. Additionally, NETs propagate inflammation by recruiting other immune cells and amplifying interferon responses [23]. There are two different mechanisms of NETs formation. The classical or lytic pathway involves NADPH oxidase-dependent generation of ROS and MPO, NE, and protein-arginine deiminase 4 (PAD4) activation [23]. This cascade leads to chromatin decondensation, plasma membrane rupture, and, finally, to cell death, resulting in NETs release [4]. Alternatively, a non-lytic NADPH oxidase-independent pathway relies on calcium influx via ionophores, activating PAD4 without complete membrane rupture, which enables neutrophils to release DNA while retaining their viability [23].
1.3. Recruitment and Resolution of Inflammation
Neutrophils are the first immune cells attracted to sites of tissue injury or infection. Their mobilization from the bone marrow into the circulation is predominantly controlled by granulocyte colony-stimulating factor (G-CSF), released in response to inflammation. When tissue homeostasis is broken, macrophages, mast cells and stromal cells sense the breach and unleash pro-inflammatory factors, such as interleukin-1β (IL-1β), tumor necrosis factor (TNF-α) and chemotactic elements, that bring over neutrophils to participate and be activated by it [24]. Recruitment begins with initial neutrophil adhesion to the endothelium through selectins and then proceeds to firm adhesion, directed by integrins. This interaction mediates trans endothelial migration through the vascular endothelium by both paracellular and transcellular routes [25]. Once in the tissue, neutrophils migrate along chemotactic gradients formed by chemoattractants such as C-X-C motif chemokine ligand (CXCL)1 and interleukin-8 (IL-8), as well as complement activation fragments, toward sites of inflammation where they exert their effector functions [26].
Although they are essential for an effective host defense, unregulated or sustained neutrophil activation can lead to the development of diseases such as acute organ injury, atherosclerosis, and autoimmune diseases like rheumatoid arthritis. Neutrophils produce a wide variety of pro-inflammatory mediators (multiple cytokines, chemokines, prostaglandins, and leukotrienes) that have the capacity to potentiate inflammatory reactions [4,10]. When the activity is excessive or uncontrolled, it will result in damage to collateral tissues. As a result, proper resolution of inflammation depends on the control of neutrophil function and its efficient removal, involving the process of apoptosis and subsequent phagocytosis by macrophages in order to restore immune homeostasis, avoiding chronic inflammation [26,27].
2. Development of Neutrophils
2.1. Hematopoiesis in the Bone Marrow
Neutrophils are generated through a tightly regulated process of hematopoiesis, more precisely, through a pathway termed granulopoiesis, which occurs primarily within the bone marrow. This process begins with hematopoietic stem cells (HSCs), which, under the influence of both intrinsic transcriptional programs and extrinsic signals, give rise to granulocyte-monocyte progenitors (GMPs) [20].
At the core of intrinsic programming, HSCs utilize unique patterns of chromatin accessibility and transcription factor binding, called cistrome, to sequentially activate or silence lineage-specific genes. Cells pass through a spectrum of lineage priming, gradually accumulating epigenetic and transcriptional marks that reinforce fate decisions [28,29].
This intrinsic pathway is dynamically responsive to external signals. Cytokines, such as G-CSF, produced by marrow stromal cells and macrophages are the principal extrinsic drivers: they activate signalling cascades such as Janus kinase/signal transducer and activator of transcription (JAK/STAT) and mitogen-activated protein kinase (MAPK), amplifying or redirecting transcriptional activity according to the demand for granulocytes. Inflammation, microbial products and metabolic cues further modulate the transcriptional program, ensuring adaptation to physiological and pathological states [30,31].
MicroRNAs and other epigenetic modifiers (like DNA methylation) also contribute to fine-tune gene expression, shaping the progenitor cell landscape and conferring plasticity to the developmental process [28,29].
Granulopoiesis is orchestrated by a hierarchy of transcription factors that ensure accurate lineage commitment, proliferation and maturation of neutrophil precursors [28,29,31].
PU box-binding protein 1 (PU.1) is a pivotal factor in early lineage restriction: high levels promote monocyte development (often at the expense of neutrophil differentiation), whereas intermediate levels permit granulocyte fate [30]. GATA-binding protein 1 (GATA-1) has an antagonistic role to PU.1, being chiefly involved in erythroid and megakaryocytic lineages: its suppression is necessary for effective neutrophil commitment [30].
Several members of CCAAT/Enhancer Binding Protein (CEBP) family are involved in granulopoiesis, particularly in neutropoiesis [28]. CEBPα is responsible for catalysing commitment to the neutrophil pathway at the GMPs and pro-neutrophil stages, driving expression of myeloid and early neutrophil genes; also, regulates primary granule protein transcription. Growth factor independent 1 (Gfi-1) operates downstream of CEBPα, repressing monocytic fate via inhibition of Early Growth Response (EGR) factors and their NGFI-A Binding (NAB) protein cofactors; its presence is critical for upregulating CEBPε, the master regulator of terminal maturation. This transcription factors are essential for non-dividing late precursors, orchestrating the formation of secondary granules and halting proliferation at the metamyelocyte stage. CEBPβ acts predominantly during emergency granulopoiesis, favouring the expansion and accelerated maturation of neutrophil precursors in response to systemic stress [32]. Finally, Runt-related transcription factor 1 (RUNX1) and Krüppel-like factor 6 (KLF6) mediate the final steps of nuclear segmentation and expression of chemotactic receptors, marking the transition to mature neutrophils [29,31].
The commitment of GMPs to the neutrophil lineage is further facilitated by G-CSF, which drives both homeostatic and emergency granulopoiesis [33].
Under physiological conditions, granulopoiesis is a steady and tightly controlled process driven by periodic cues and subject to negative feedback from neutrophil clearance, maintaining a large marrow reserve [28,31]. This process is called homeostatic granulopoiesis. On the other hand, emergency granulopoiesis is activated in response to systemic infection or inflammation, being largely triggered by upsurges in G-CSF, pro-inflammatory cytokines (IL-1β, IL-6 and TNF-α) and direct PRRs signalling on HSCs. This process is distinguished by a rapid expansion of myeloid-biased progenitors, an acceleration of neutrophil maturation (mainly CEBPβ-driven), a premature release of immature and mature neutrophils (which can exhibit distinct immune regulatory functions or heightened antimicrobial profiles) and by mobilization of extra-medullary hematopoiesis (in the spleen or liver) if marrow output is insufficient [29].
2.2. Maturation Stages and Phenotypic Markers
Neutrophil maturation takes place in the bone marrow and follows a sequential progression through morphologically and functionally distinct stages, each transition being characterized by progressive nuclear segmentation, granule formation and dynamic shifts in surface marker expression [29,31,34].
HSCs and GMPs express, on their surface, CD34, CD117 and CD33. When they evolve into early precursors (promyelocytes and myelocytes), they acquire CD66b, CD33, CD15 and CD49d; these cells are characterised by a round nucleus with slight indentation. During intermediate stages (metamyelocytes and bands), nucleus evolve from kidney to rod shape; these cells express, on their surface, CD16 and gradually lose CD49d. The transition from myelocytes to metamyelocytes separates two different steps: mitotic pool (when cells can undergo self-renewing proliferation and differentiation) and post-mitotic pool (Figure 1) [35,36].
Finally, a mature neutrophil presents a segmented nucleus and the highest granule content; on the surface, expresses high concentration of CD16 and CD10, while definitely loses CD49d. After they leave the bone marrow, they differentiate into their different phenotypes (Figure 1) [35,36].
Transcriptional drivers are tightly coupled to granule biogenesis: CEBPα and GFI-1 support primary granule formation at early stages, while CEBPε orchestrates secondary and tertiary granule production, critical for effector function acquisition. Mature neutrophils display full migratory and antimicrobial machinery, whereas precursors, particularly those released during emergency granulopoiesis, may retain proliferative or unique regulatory capacities [30,31].
2.3. Mobilization to Circulation
The release of neutrophils from the bone marrow into peripheral blood is a highly controlled process, governed by chemokine gradients and the interplay of retention and egress signals. C-X-C chemokine receptor (CXCR) 4 and its ligand CXCL12 retain developing neutrophils in the marrow, whereas expression of CXCR2 and its ligands (CXCL1 and CXCL2) promote mobilization into the bloodstream. G-CSF triggers neutrophil egress by altering CXCL12-CXCR4 signalling and promoting the expression of CXCR2 ligands [31,34].
During homeostasis, only a minority of total body neutrophils circulate at any given time, with most residing in bone marrow reserves. Upon infection, inflammation or G-CSF administration, accelerated mobilization (emergency granulopoiesis) leads to a detectable "left shift" with increased circulating immature forms. Mobilized neutrophils, including immature subsets, possess distinct phenotypic and functional profiles, reflecting both their maturation state and recent environmental reprogramming encountered within the marrow or during trafficking [29,34].
Following their release, neutrophils circulate briefly before migrating into tissues in response to inflammatory cues. The subsequent fate includes participation in immune defence, functional specialization upon tissue entry and eventual clearance by macrophages either in tissues or through return to the bone marrow [31,34].
3. Neutrophils in the Immune System
Under homeostatic conditions, neutrophils are continuously generated in the bone marrow and released into the bloodstream, where they circulate for a short lifespan and migrate into tissues, constantly patrolling for signs of infection or damage [3,34]. Their rapid recruitment to sites of inflammation, mediated by adhesion, rolling, and transmigration across the endothelium in response to chemokines and pathogens/damage-associated molecular patterns, places neutrophils as the first line of defense of the innate immune system [2,37,38].
Classically, neutrophil antimicrobial activity is mediated by phagocytosis, degranulation, and the release of NETs. While these mechanisms are essential for pathogen control, their dysregulation can promote collateral tissue damage and contribute to chronic inflammation, autoimmunity, and thrombosis [2].
For decades, neutrophils were regarded as homogeneous, short-lived, terminally differentiated cells whose role was restricted to pathogen elimination [39]. However, advances in molecular biology, single-cell approaches, and gene-editing technologies have dramatically reshaped this view. It is now well established that neutrophils are functionally versatile and heterogeneous, with diverse phenotypes depending on maturation stage, activation status, and environmental cues. Rather than passive end-stage effectors, neutrophils are transcriptionally active cells capable of dynamically altering their gene expression and functional programs [38].
This versatility extends beyond pathogen killing into immunomodulation. Neutrophils produce a broad range of pro-inflammatory cytokines, such as IL-1β, TNF-α, and IL-12, which amplify innate and adaptive responses, but they also contribute to the resolution of inflammation by clearing apoptotic cells and releasing anti-inflammatory mediators like transforming growth factor-β (TGF-β) [40]. Through these mediators, neutrophils orchestrate the recruitment and activation of macrophages, natural killer cells, dendritic cells, and T and B lymphocytes [41,42]. Moreover, under specific conditions they can upregulate major histocompatibility complex (MHC) class II and function as atypical antigen-presenting cells, or conversely suppress T cell proliferation via arginase-1, ROS, or immune checkpoint ligands such as PD-L1 [43].
Altogether, neutrophils have emerged as a plastic, context-dependent cell population at the crossroads of innate and adaptive immunity. Their ability to switch between pro- and anti-inflammatory programs, and to acquire specialized functions depending on physiological or pathological conditions, has redefined them as key players not only in host defense but also in autoimmunity, chronic inflammatory disorders, and cancer [42,43].
4. Neutrophils in Cancer
The presence of immune cells in the tumor immune microenvironment (TIME), including neutrophils, was initially interpreted as a host attempt to eradicate malignant cells. This interpretation has since evolved into a more complex framework, as cancer-associated inflammation is now recognized as a hallmark of cancer [44], with neutrophils emerging as central regulators of this process [45].
In line with this paradigm shift, neutrophils are increasingly appreciated as highly plastic cells capable of adopting either tumor-promoting or tumor-suppressive functions in response to contextual cues. Clinically, this functional versatility is reflected by the frequent occurrence of neutrophilia and elevated neutrophil-to-lymphocyte ratio (NLR) in cancer patients, particularly those with advanced disease, where both parameters consistently associate with poor prognosis across multiple cancer types, including breast, colorectal and lung cancers, as well as melanoma [12,45,46,47,48]. Although the mechanisms driving tumor-induced neutrophilia remain incompletely defined, accumulating evidence implicates tumor-derived granulopoietic and chemotactic factors, such as granulocyte-macrophage colony-stimulating factor (GM-CSF), G-CSF, IL-1β, IL-6, CXCL1, CXCL2, and CXCL8 (IL-8), in sustaining a chronic pro-inflammatory and tumor-permissive systemic environment [45,49].
These systemic alterations are paralleled by an increasing appreciation of neutrophil heterogeneity in cancer. Distinct neutrophil subsets have been identified both within the TME and in circulation, differing in their maturation status, activation profiles, and suppressive capacity. While TANs exert localized effects within the TIME, circulating neutrophils are more readily accessible and may serve as indicators of tumor-driven systemic immune remodeling [50]. Given the continuous trafficking of neutrophils to and from the tumor niche, their phenotypic and functional characteristics can provide valuable insights into intratumoral dynamics and disease evolution [49,51]. Nevertheless, most studies to date have predominantly focused on TANs or relied heavily on preclinical animal models, with only more recent clinical investigations addressing the contribution of systemic neutrophils to cancer. Importantly, emerging evidence, including our own findings [12], highlights the relevance of profiling circulating neutrophil subsets to better understand their role in tumor progression, therapeutic resistance, and patient outcomes [49,52,53,54,55,56,57].
At the tissue level, neutrophils display remarkable functional adaptability while retaining their classical roles in acute inflammation. In response to local microenvironmental signals, TANs can be polarized toward either antitumor or pro-tumor phenotypes. Pro-tumorigenic TANs actively promote tumor progression through the secretion of pro-angiogenic mediators, such as vascular endothelial growth factor (VEGF) A, as well as matrix metalloproteinases (MMPs), thereby facilitating neovascularization, extracellular matrix remodeling, and tumor cell invasion [58]. Beyond these direct effects on tumor biology, TANs also exert profound influence on adaptive immune responses, ultimately shaping tumor growth trajectories and modulating responses to anticancer therapies.
Mechanistically, TGF-β signaling has emerged as a key driver of neutrophil polarization toward immunosuppressive phenotypes, conferring protection to cancer cells from immune surveillance and reducing the efficacy of both conventional and immune-based therapies [59]. More broadly, disruption of neutrophil homeostasis in cancer encompassing their production, activation, trafficking, and clearance favors the accumulation of neutrophil populations that support tumor growth and immune evasion. Collectively, these observations position neutrophils as dynamic regulators of tumor-associated inflammation and immunity, reinforcing their relevance as both prognostic biomarkers and promising therapeutic targets in cancer.
4.1. TANs
The phenotype and function of TANs are largely dictated by tumor-derived factors, which drive neutrophil polarization toward either anti- or pro-tumoral states.
The duality of TANs function was first formalized in 2009 by Fridlender et al., who proposed a polarization model in which TGF-β promoted a pro-tumoral neutrophil phenotype (N2), whereas type I interferons (e.g., IFN-β) favored anti-tumoral polarization (N1). N1 TANs exhibit a mature phenotype, short lifespan, and hypersegmented nuclei, and they exert anti-tumor activity through multiple mechanisms: via antibody-dependent cellular cytotoxicity, release of ROS and reactive nitrogen species (RNS), and expression of death-inducing ligands such as TNF-related apoptosis-inducing ligand (TRAIL) and TNF-α N1 TANs also act as immune activators, producing cytokines (IL-12, TNF-α, GM-CSF) and chemokines (CCL3, CXCL9, CXCL10) that recruit dendritic cells and CTLs, thereby bridging innate and adaptive immunity [60]. Remarkably, neutrophils are also capable of antigen presentation, further amplifying T cell responses in early disease, N1 TANs can even limit metastasis formation by killing or restraining circulating tumor cells (Figure 2) [61].
In contrast, N2 TANs, which are typically characterized by circular nuclei [50], promote tumor progression and metastasis through multiple mechanisms. They contribute to genomic instability by releasing ROS and RNS that damage DNA [49], and directly sustain tumor proliferation via growth factors [epidermal growth factor (EGF), hepatocyte growth factor (HGF), oncostatin M] [62], cytokines (IL-6, IL-8, TNF-α), and proteases such as NE [49]. By releasing MMP-9 and elastase, N2 TANs remodel the extracellular matrix and promote invasion, while secreting angiogenic factors (VEGF, prokineticin 2) to stimulate vascularization [49,62]. Importantly, N2 TANs are potent immunosuppressors, characterized by high expression of CCL17 (which recruits Tregs to the tumor site) [63], arginase-1 (depleting arginine essential for T cell proliferation and survival), and PD-L1 (directly impairing T cell activation) [49]. Collectively, these mechanisms foster an immunosuppressive TIME that facilitates tumor immune evasion and progression (Figure 2).
Additionally, NETs released by TANs are increasingly recognized as critical mediators of metastasis (Figure 2). NETs can trap circulating tumor cells, remodel the extracellular matrix, release proteases and growth factors, awaken dormant tumor cells, and shield metastatic seeds from immune clearance. Components of NETs, such as MPO, can also directly impair T cell function, reinforcing their immunosuppressive role [64,65].
It is important to note, however, that the N1/N2 paradigm is largely based on murine models and in vitro polarization studies. Human TANs are more difficult to access, and single-cell transcriptomic studies now suggest that neutrophils in cancer exist along a continuum of plasticity rather than as discrete N1 or N2 subsets [66]. While the N1/N2 framework remains conceptually useful, a deeper understanding of the signals shaping TANs phenotypes in human tumors will be critical for translating these insights into therapeutic strategies.
4.2. Circulating Neutrophils in Cancer
Systemically, cancer patients often present neutrophilia, frequently accompanied by relative lymphopenia, resulting in an elevated NLR. This parameter, derived from routine blood counts, provides an indirect measure of the balance between pro-tumor inflammation (driven by neutrophils) and antitumor immunity (driven by lymphocytes) [67]. Elevated NLR has been reported across multiple cancer types, including breast, lung, colorectal, and hepatocellular carcinoma, and has emerged as one of the most robust and widely studied systemic immune biomarkers in oncology [68].
Consistent evidence links high NLR values with advanced disease stage, poor overall survival, and reduced responses to therapy. Meta-analyses further validate its prognostic value across solid tumors, though the optimal cutoff varies between studies [69,70,71]. Importantly, dynamic changes in NLR during treatment can also be informative: patients whose NLR decreases upon chemotherapy tend to achieve better outcomes compared with those who maintain persistently high NLR [72]. Nevertheless, because NLR reflects both neutrophil expansion and lymphocyte depletion, it provides only an overview of systemic immune status and does not capture the functional diversity of circulating neutrophils.
To overcome this limitation, researchers have turned to a more detailed characterization of neutrophil subsets. By distinguishing populations based on density and functional properties, it is possible to uncover immunosuppressive or antitumoral activities that are not apparent from total neutrophil counts or NLR alone [73].
Indeed, similar to intratumoral neutrophils, circulating neutrophils represent a heterogeneous population, which can be subdivided based on density. Under homeostatic conditions, neutrophils sediment in the granulocytic fraction of density gradients and are referred to as HDNs or normal-density neutrophils (NDNs). In contrast, during pathological conditions, including cancer, autoimmune diseases, infection, and even physiological conditions such as pregnancy, a distinct fraction of neutrophils appears in the PBMC layer, termed LDNs (Figure 3) [12,13,34].
5. LDNs
5.1. Definition and Historical Discovery
The historical discovery of LDNs dates to 1986, when Hacbarth and Kajdacsy-Balla first described "low buoyant density neutrophils" contaminating PBMC fractions from patients with systemic lupus erythematosus and rheumatoid arthritis during routine density separation. Initially dismissed as artifacts, these cells were systematically characterized in the early 2010s across chronic inflammatory diseases, infections (e.g., Mycobacterium tuberculosis), and cancer, with key studies documenting their expansion, pro-inflammatory NETosis propensity, and T cell suppressive functions [74,75].
5.2. Distinctive Density, Phenotype, and Function Compared to HDNs
Morphologically, HDNs are predominantly mature cells, while LDNs include both mature and immature forms [13,76]. Phenotypically, both populations share classical markers such as CD15, CD66b, and CD11b, but LDNs often exhibit higher expression of activation markers (CD66b, CD11b), suggesting a more activated or primed state [12,77]. Moreover, high-dimensional flow cytometry in lung cancer, identified novel surface markers that distinguish LDNs from HDNs, including CD36, CD41, CD61, and CD226, all of which were significantly upregulated in LDNs [18]. Nevertheless, despite partial phenotypic overlaps between LDNs and HDNs, density-based remains the most reliable approach to distinguish these populations in functional studies [12,77].
Functionally, HDNs and LDNs mirror the N1/N2 dichotomy: HDNs display anti-tumoral properties, including cytotoxic activity and immune stimulation, whereas LDNs are enriched in cancer patients and display immunosuppressive, pro-tumoral features (Figure 2) [12,17]. LDNs suppress T cell proliferation and activation, facilitate tumor progression, and are associated with resistance to therapies such as chemotherapy and immune checkpoint blockade [12,13,17,78]. Accordingly, increased frequencies of LDNs correlate with advanced disease, aggressive tumor phenotypes, and poor clinical outcomes [12,18,77,79,80].
5.3. Origin, Development, and Heterogeneity of LDNs
The origin of LDNs remains debated. They may represent immature neutrophils prematurely released from the bone marrow under emergency granulopoiesis, or mature neutrophils reprogrammed into a suppressive phenotype by systemic and tumor-derived factors. Murine studies suggest that HDNs can convert into LDNs under TGF-β signaling, acquiring immunosuppressive features, though this conversion has not been confirmed in tumor-free settings [13,34,81,82]. This plasticity underscores the idea that neutrophils should be viewed as a dynamic spectrum of states rather than as fixed subtypes [83,84,85].
Another unresolved question is whether LDNs and granulocytic or polymorphonuclear myeloid-derived suppressor cells (G-MDSCs or PMN-MDSCs) are distinct entities or overlapping populations. Both expand under pathological conditions, share morphological and phenotypic similarities, and exert immunosuppressive effects. Some authors propose that G-MDSCs represent a functional subset within the broader LDNs pool, differing mainly in activation state and immunosuppressive ability within cancer context [34,86].
Altogether, these uncertainties highlight the need for precise characterization of neutrophil subsets in humans. Nonetheless, the consistent association of LDNs with disease progression and therapeutic resistance makes them attractive candidates as biomarkers and as therapeutic targets [12,53,55,78,79]. Strategies aimed at reducing LDNs accumulation, reprogramming them toward anti-tumoral functions, or blocking their recruitment to the TIME are currently under investigation and hold promises for improving overall survival.
5.4. Activation and Metabolic Properties of LDNs
LDNs are a subset of neutrophils characterized by sustained activation and enhanced effector potential. Across pathological contexts, including cancer and chronic inflammation, LDNs exhibit features of persistent activation that distinguish them from conventional HDNs. Importantly, their functional specialization is increasingly recognized as being tightly linked to profound metabolic adaptations that support their survival and activity under conditions of environmental stress [78].
Within TME, these general features of LDNs acquire particular relevance. For instance, in metastatic breast cancer, LDNs display a highly activated phenotype compared to HDNs, characterized by elevated expression of activation and degranulation markers. In parallel, LDNs upregulate immunoregulatory molecules including PD-L1 and TLR4, the latter being associated with prolonged neutrophil survival and the secretion of a proliferation-inducing ligand (APRIL), a cytokine implicated in tumor proliferation and disease progression [17,87,88].
This activated state translates into robust effector functions. LDNs secrete pro-inflammatory cytokines such as IL-8 and TNF-α and release high levels of granule-associated enzymes, including MPO, NE, and MMPs. These mediators contribute to extracellular matrix remodeling, angiogenesis, and immune evasion, thereby facilitating tumor invasion and metastatic dissemination. Elevated expression of MMP-9, VEGF, and MPO further underscores the contribution of LDNs to the dynamic remodeling of the TME [17,87,88,89].
Crucially, the capacity of LDNs to sustain these energetically demanding effector functions relies on extensive metabolic reprogramming. LDNs exhibit marked metabolic flexibility, engaging glycolysis, oxidative phosphorylation, and lipid metabolism to maintain ROS production and NETs formation, even under nutrient-deprived conditions typical of the TME [90,91,92,93]. This metabolic adaptability is also evident in neutrophils transitioning from the high-density to the low-density state, which display increased ROS generation, reduced phagocytic capacity, and lower expression of CD39 and CD73, features associated with enhanced immunosuppressive activity and tumor-promoting functions [94].
NETosis constitutes a central functional output closely linked to the metabolic state of LDNs. NETs derived from LDNs display higher MPO intensity than those formed by HDNs. This enhanced NETotic capacity has been associated with metastatic progression, immune suppression, and resistance to therapy [12].
Within the heterogeneous LDNs compartment, immature LDNs (iLDNs) emerge as a metabolically distinct and particularly pro-metastatic subset. iLDNs exhibit superior global bioenergetic capacity compared to HDNs, enabling them to sustain metabolically demanding functions such as NETosis under conditions of metabolic stress. In breast cancer models, iLDNs rely on mitochondrial metabolism and amino acid catabolism, particularly glutamate and proline, to maintain NETs formation when glucose availability is limited, thereby promoting metastatic colonization of distant organs, including the liver and lungs [78,90,95]. Tumor-derived G-CSF has been shown to be necessary, though not sufficient, for iLDNs mobilization, and its knockdown significantly reduces both iLDNs levels and metastatic burden [78].
In contrast, mature LDNs (mLDNs) lack comparable metabolic plasticity and fail to promote metastatic dissemination, highlighting neutrophil maturation state as a key determinant of pro-tumorigenic activity. In addition to amino acid metabolism, lipid metabolic pathways also contribute to LDNs function, as lung mesenchymal cells can induce lipid accumulation in neutrophils, providing metabolic support that enhances tumor cell survival and proliferation at metastatic sites [90].
Collectively, these metabolic adaptations enable LDNs to maintain ROS production, NETosis, and immunosuppressive activity despite the hostile metabolic landscape of the TME, to where these cells could be recruited. LDNs-mediated arginase release further contributes to local arginine depletion, impairing T cell activation and reinforcing immune suppression, a mechanism in vitro reverted by arginase inhibition with Nω-hydroxy-nor-L-arginine (nor-NOHA) [17]. Clinically, LDNs accumulation has been associated with poor outcomes across several cancers, including breast cancer, where it correlates with reduced overall survival, resistance to neoadjuvant chemotherapy, faster disease progression, diminished cytotoxic T cell activity, and increased regulatory T cell populations [12,88], underscoring the pathological relevance of LDNs metabolic reprogramming in tumor progression.
5.5. Interactions of LDNs with Other Cells in Immune Responses and Cancer
LDNs also display remarkable plasticity in their interactions with other immune cells. These interactions occur both within innate and adaptive immunity and play critical roles in inflammation, immune regulation, and cancer progression.
A characteristic feature of LDNs is their increased expression of P-selectin glycoprotein ligand-1 (PSGL-1), which facilitates the formation of neutrophil-platelet aggregates [96,97]. This crosstalk enhances the release of NETs, thereby amplifying inflammatory damage and contributing to tissue injury. In cancer, platelet-LDNs cooperation can facilitate the establishment of pre-metastatic niches by enhancing NETs-mediated trapping of circulating tumor cells and supporting pro-thrombotic, tumor-permissive conditions (Figure 4) [96,97].
LDNs also establish reciprocal communication with macrophages. In systemic lupus erythematosus, for example, LDNs-derived NETs activate the NOD-, LRR- and pyrin domain-containing protein 3 (NLRP3) inflammasome in macrophages, leading to the secretion of inflammatory mediators such as TNF-α, IL-6, IL-10, IL-1β, and IL-18, and creating a feed-forward inflammatory loop. In in vitro experiments, conditioned medium derived from LDN isolated from BC patients also promoted an anti-inflammatory myeloid phenotype [17]. In other contexts, such as alcohol-induced LDNs, these cells express “don’t-eat-me” signals that inhibit macrophage phagocytosis, which contributes to immune evasion (Figure 4) [81].
The interaction of LDNs with T cells is particularly complex and context-dependent. In many pathological conditions, including cancer, LDNs act as potent suppressors of T cell responses through mechanisms such as hydrogen peroxide production, arginine depletion, or PD-L1 expression, thereby impairing T cell activation and effector functions [81]. Recent evidence further demonstrates that tumor-associated LDNs directly disrupt T cell receptor signaling by impairing early phosphorylation events, which reduces both CD4⁺ and CD8⁺ T cell activation and favors an immunosuppressive environment (Figure 4). These inhibitory effects correlate with transcriptional profiles enriched in immunosuppressive and pro-metastatic signatures, reinforcing the role of LDNs as key mediators of tumor immune escape [15]. However, in certain settings such as neonatal immunity, these cells can instead promote T cell proliferation and enhance cytokine secretion, including IL-6 and interferon gamma (IFN-γ) (Figure 4). This duality highlights the importance of the developmental and activation state of LDNs in shaping their functional outcome on adaptive immune responses [81].
Although substantial progress has been made in characterizing the crosstalk of LDNs with platelets, macrophages, and T cells, some gaps remain. For instance, the role of LDNs in modulating B cell responses is less explored, even though they have been reported to activate B cells in autoimmune diseases (Figure 4). In addition, the extent to which LDNs maturity influences their interactions with other immune cells in cancer remains poorly defined. These aspects highlight the need for further research into therapeutic strategies that could target LDNs functions, including the inhibition of PSGL-1, the blockade of PD-L1, the neutralization of “don’t-eat-me” signals, or the prevention of NETs formation, all of which may provide novel opportunities to counteract LDNs-mediated immunosuppression in cancer [81].
6. Dynamic Changes of LDNs During Human Cancer
6.1. Variation in Frequency, Phenotype, and Functional Relevance Across Cancer Stages
LDNs exhibit dynamic changes in frequency, phenotype, and function throughout tumor progression. In early-stage cancer, LDNs are generally rare, but their proportion increases markedly as the disease advances [12,18,52]. For instance, in advanced lung cancer, LDNs can constitute a median of 7% of circulating neutrophils, with some patients exhibiting levels as high as 80%, compared to less than 1% in early-stage disease and healthy individuals [18,52]. Similar trends are observed in breast cancer, where LDNs are significantly enriched in patients with metastatic disease. Notably, among non-metastatic patients, those who fail to respond to neoadjuvant chemotherapy present higher pre-treatment LDNs levels, suggesting their potential as a predictive biomarker [12].
Phenotypically, LDNs are heterogeneous, particularly in advanced cancer, where immature and mature subsets coexist [13,78]. However, transcriptomic analyses, reveal that iLDNs are enriched in migration-related gene signatures, including genes involved in transendothelial migration [98]. Functionally, neutrophils shift during tumor progression: HDNs predominate in early stages, supporting antitumor responses, whereas LDNs expand in advanced disease, often becoming the major circulating neutrophil population. This transition highlights the plasticity of neutrophils, which can adopt either antitumor or protumor roles depending on environmental cues, suggesting that LDNs may evolve from HDNs or vice versa [13].
This plasticity is further shaped by tumor-derived factors such as CXCL1 and TGF-β, which drive the conversion of HDNs into LDNs via the CXCL1–CXCR2 axis, a pathway also involved in tumor-directed migration. LDNs exhibit enhanced infiltration into tumor tissue compared to HDNs, both ex vivo and in vivo, reflecting superior migratory capacity and recruitment to the TME [94].
Moreover, as cancer progresses, LDNs acquire a more immunosuppressive and pro-metastatic phenotype, with the highest levels and most pronounced functional changes observed in metastatic and high-burden disease [12,52]. iLDNs become particularly relevant during metastatic progression, suggesting a systemic neutrophil adaptation that supports metastasis, especially in metabolically stressed environments such as the liver [78,98].
Clinically, LDNs levels correlate with prognosis and therapeutic response. Indeed, the presence of LDNs has been identified as predictive of poor outcomes, with higher levels associated with worse overall survival, reduced progression-free survival (PFS), poor response to chemotherapy, and increased metastatic burden [17,99,100]. Persistent high neutrophil counts and immature neutrophil phenotypes, including LDNs, are also linked to early immunotherapy failure namely in lung, colorectal, gastric and breast cancers [99,100,101]. Moreover, LDNs exhibit impaired phagocytosis, increased ROS production, and a higher propensity for NETs formation, which protects circulating tumor cells, facilitates adhesion and migration, and supports pre-metastatic niche establishment in distant organs [102,103].
Taken together, these observations illustrate that LDNs are not only markers of tumor burden but also active contributors to tumor progression, metastasis, and immune evasion, emphasizing their importance as both prognostic biomarkers and potential therapeutic targets.
6.2. LDNs in Breast Cancer: From Subtype Associations to Clinical Outcomes
LDNs have emerged as key players in breast cancer progression, contributing to both tumor promoting and immunosuppressive functions (Figure 5), as consistently demonstrated across multiple studies summarized in Table 1. Their recruitment and expansion in circulation are associated with poorer prognosis, particularly in aggressive disease subtypes such as triple-negative breast cancer (TNBC) and HER2-positive tumors [88,104]. Mechanistic studies demonstrate that breast cancer cells can directly enhance neutrophil survival and protumorigenic activity [88], reinforcing the systemic expansion of LDNs observed in patients. Recent analyses further reveal that LDNs display transcriptional programs enriched for interferon stimulated genes, NETosis-related pathways, and immunosuppressive mediators such as arginase-1, PD-L1, and TGF-β responsive signatures, supporting their role as active drivers of tumor progression [17,105].
The systemic impact of LDNs expansion is reflected in alterations of the NLR, which is frequently elevated in patients with higher LDNs frequencies. An increased NLR has been consistently linked to poor PFS reduced overall survival (OS), and higher metastatic risk, particularly in TNBC and HER2-positive subtypes [104,106]. These clinical associations align with observations from large scale transcriptomic datasets, where neutrophil-related inflammatory signatures correlate with aggressive tumor biology and early metastatic dissemination [107].
Phenotypic heterogeneity among circulating neutrophils is well documented in breast cancer. Early animal models first highlighted the expansion of LDNs in aggressive tumors, and more recent human studies have confirmed these findings. In early-stage breast cancer, alterations in circulating neutrophil populations, including increased LDNs proportion, are detectable from diagnosis. Distinct subtype dependent differences have been described: TNBC patients exhibit marked expansion of LDNs characterized by reduced L-selectin expression (CD62Llow), altered intracellular signaling, prolonged lifespan, and enhanced immunosuppressive potential, whereas hormone-receptor positive (HR+) tumors display opposite trends with lower LDNs abundance and less activated phenotypes [108]. These observations are consistent with multiomic profiling showing that TNBC associated LDNs are enriched for inflammatory and interferon driven transcriptional programs, while HR+ tumors exhibit comparatively muted neutrophil activation [105,107].
Beyond subtype associations, LDNs have gained attention as predictors of therapeutic response. Elevated baseline LDNs levels have been associated with reduced pathological complete response rates in patients undergoing neoadjuvant chemotherapy, and dynamic increases in LDNs during treatment correlate with residual disease and chemoresistance [12]. Their immunosuppressive activity, mediated through T cell inhibition, ROS, and NETs-mediated physical exclusion of immune cells, also contributes to diminished responses to immune checkpoint blockade, particularly in TNBC [17,105]. Emerging evidence further suggests that neutrophil related inflammatory signatures may modulate the efficacy of HER2targeted therapies, raising the possibility that LDNs quantification could refine existing predictive models [107].
Clinically, high LDNs levels consistently associate with adverse outcomes across breast cancer cohorts. Increased LDNs abundance correlates with shorter PFS and OS, higher rates of early visceral metastasis, and greater likelihood of treatment resistance [12,17,104,105,106,107]. Mechanistic studies support these observations, demonstrating that LDNs facilitate metastasis by promoting NETs mediated trapping of circulating tumor cells, enhancing angiogenesis, and secreting pro-invasive factors that support tumor cell dissemination and colonization [12].
Together, these findings highlight the relevance of LDNs as both biomarkers and functional mediators of breast cancer progression. As interest grows in integrating neutrophil biology into clinical decision-making, future research should prioritize standardizing LDNs quantification, dissecting their ontogeny and heterogeneity, and exploring therapeutic strategies aimed at modulating LDNs activity. Incorporating LDNs related metrics into predictive algorithms for chemotherapy, immunotherapy, and targeted therapy may ultimately improve BC patient stratification and open new avenues for improved therapeutic intervention.
6.3. LDNs in Lung Cancer: Clinical Relevance and Immunoregulatory Functions
Similar to other solid cancers, LDNs have gained increasing attention as a distinct neutrophil subset with major implications for lung cancer biology (Figure 5), particularly in NSCLC, as summarized in Table 1.
Clinically, elevated LDNs levels in non-small cell lung cancer (NSCLC) correlate with advanced disease stage, increased tumor burden, and reduced overall survival (OS). Proteomic and high-dimensional cytometry analyses have revealed that LDNs display distinct immune signatures enriched for tumor-promoting inflammatory pathways, extracellular matrix remodeling, angiogenesis, and immunosuppression [80,109]. Importantly, beyond their prognostic relevance, LDNs have emerged as robust predictors of resistance to immune checkpoint inhibition. In first-line NSCLC cohorts treated with programmed cell death protein 1 (PD-1)/PD-L1 blockade, high baseline LDNs frequencies were strongly associated with primary resistance and early disease progression [53]. Together, these findings indicate that LDNs function not only as minimally invasive biomarkers of poor outcome and immunotherapy resistance, but also as active contributors to tumor progression in lung cancer.
Subtype specific differences have also been reported. In a study focusing on advanced lung adenocarcinoma, Liu et al. showed that LDNs are phenotypically distinct and correlate with impaired immune status, including reduced T cell function [77]. This suggests that LDNs mediated immunosuppression may be particularly relevant in adenocarcinoma biology. Although direct comparisons across NSCLC subtypes remain limited, available evidence indicates that neutrophil driven inflammation tends to be more pronounced in squamous cell carcinoma, which may reflect a greater contribution of LDNs in this subtype. In small cell lung cancer (SCLC), systemic inflammation is typically more severe, and although LDNs were not directly quantified, neutrophil based biomarkers such as the NLR have been shown to influence immunotherapy stratification [110,111,112,113], indirectly supporting a role of LDNs mediated immunosuppression [112].
Functionally, LDNs exhibit potent immunosuppressive activity that closely resembles G-MDSCs. They suppress T cell proliferation and effector function through mechanisms such as ROS production, arginase-1 release, nutrient depletion, and secretion of immunomodulatory cytokines [77]. These suppressive mechanisms contribute to a profoundly immunosuppressive TME, characterized by impaired CD8⁺ T cell cytotoxicity and reduced antigen presentation, as reflected in the immune signatures identified in advanced lung cancer patients [80]. The immunosuppressive nature of LDNs has been consistently observed across cancer types, but in lung cancer it appears particularly impactful due to the already inflamed and neutrophil rich TME, and is further supported by proteomic analyses linking LDNs to T cell dysfunction and resistance to immunotherapy [53,109].
Collectively, these findings highlight LDNs as both biomarkers and active mediators of immune escape in lung cancer. Their ability to predict immunotherapy resistance, their association with poor prognosis, and their direct immunosuppressive functions make them compelling candidates for therapeutic intervention. As interest in this neutrophil subset grows, LDNs are increasingly recognized not only as indicators of systemic immune dysfunction but also as actionable targets capable of reshaping the tumor-immune landscape. Strategies aimed at reducing LDNs expansion, blocking their suppressive mediators, or preventing their recruitment to the tumor site may enhance the therapies efficacy. At the same time, advances in transcriptomic and proteomic profiling are refining the molecular identity of LDNs, opening opportunities to selectively target these cells without compromising essential neutrophil functions. Together, these developments position LDNs -directed therapies as a promising avenue to enhance the efficacy of immunotherapy and improve clinical outcomes for patients with lung cancer.
7. Therapeutic Potential of Targeting LDNs
For a long time, neutrophils were characterized as a homogeneous cell class with diverse functions. However, the identification of neutrophil subpopulations has allowed for the reclassification of this immune cell type, with distinct properties now described that impact cellular homeostasis. There is particular interest in the influence of these cells on carcinogenesis and on responses to oncological therapies [55]. Among the various neutrophil populations, LDNs are of particular interest in oncology research. They influence the behavior of tumor cells, immune cells responding to the tumor, and the entire TME, with implications for the efficacy or resistance to treatments such as chemotherapy or immunotherapy [13]. In various immunotherapy studies, growth factors that influence the response to this treatment in different solid neoplasms have been identified, including IL-6, IL-7, and HGF. While their inhibitory activity is better established mainly in lymphocyte populations, the actions they may perform in neutrophil populations remain to be clarified [114,115].
Importantly, neutrophil plasticity allows for the interconversion between HDNs and LDNs, a process influenced by TGF-β and the local inflammatory milieu [13]. This plasticity presents an opportunity to therapeutically reprogram neutrophils, potentially reducing the pro-tumor activity of LDNs and restoring anti-tumor immune responses.
Targeting LDNs in cancer therapy faces several challenges. Their heterogeneity and overlap of surface markers with other neutrophil subpopulations complicate selective depletion strategies. Moreover, their functional roles are context-dependent: in certain conditions, LDNs may retain residual anti-tumor functions, meaning non-specific targeting could impair beneficial immune responses. Nonetheless, multiple strategies are being explored:
- Modulation of neutrophil plasticity – targeting TGF-β or other tumor-derived factors to prevent the conversion of HDNs into pro-tumor LDNs [10].
Beyond oncology, LDNs-targeted strategies have been investigated in autoimmune and inflammatory diseases, primarily focusing on reducing NETs formation and pathological inflammation [81,97,121,122].
Overall, targeting LDNs represents a promising, yet complex, approach to cancer therapy [123]. Future strategies should aim to selectively inhibit the pro-tumorigenic and immunosuppressive functions of LDNs while preserving beneficial neutrophil activity, ideally integrating LDNs modulation into multi-modal therapeutic regimens [124,125].
8. Concluding Remarks
LDNs have emerged as a central component of cancer-related systemic inflammation, representing a dynamic and heterogeneous population with profound implications for tumor progression, immune evasion, and therapeutic resistance. Evidence from breast and lung cancers consistently demonstrates that LDNs are not passive byproducts of dysregulated granulopoiesis but active drivers of disease biology. Their enrichment in advanced stages, association with poor clinical outcomes, and capacity to suppress cytotoxic immunity underscores their relevance as both biomarkers and functional mediators of cancer progression.
Despite significant advances, several critical gaps remain. The ontogeny of LDNs, whether they arise from immature precursors, represent activated mature neutrophils, or constitute a distinct lineage, remains incompletely resolved. Similarly, the molecular cues that govern their expansion, phenotypic plasticity, and trafficking in cancer patients require deeper mechanistic exploration. High dimensional profiling has begun to reveal unique surface markers and transcriptional programs, yet a unified framework for defining LDNs subsets across cancer types is still lacking.
From a translational perspective, LDNs hold promise as minimally invasive biomarkers for early detection, disease monitoring, and prediction of therapeutic response, particularly in the context of immunotherapy. However, their clinical implementation will require standardized isolation methods, robust phenotypic signatures, and prospective validation in large patient cohorts. Therapeutically, strategies aimed at modulating neutrophil recruitment, polarization, or suppressive activity, such as CXCR2 inhibitors, NETosis blockade, or targeting PD-L1 and arginase pathways, are beginning to enter clinical evaluation. Understanding how these interventions reshape systemic neutrophil landscapes, including LDNs, will be essential for optimizing their efficacy and safety.
Looking ahead, integrating multiomics technologies, advanced imaging, and longitudinal patient sampling will be key to unraveling the full spectrum of LDNs biology. Moreover, dissecting the interplay between LDNs, tumor associated neutrophils, and other immune compartments may reveal new therapeutic vulnerabilities and opportunities for combination strategies. As our understanding deepens, LDNs are poised to transition from emerging biomarkers to actionable targets in precision oncology, offering new avenues to improve outcomes for patients with breast, lung, and other solid tumors.
Authorship
TM and RS performed the full literature review, drafted the manuscript, and prepared the figures. BFC, DG, and IMS critically revised the manuscript, contributed to parts of the literature review, and assisted in figure preparation. MV, CF, and ARF contributed to portions of the literature review and reviewed the manuscript. AJ, SB and MVB provided guidance on the work and reviewed the manuscript. MGC supervised the work, provided overall guidance, and critically revised and validated the final manuscript. All authors read and approved the final version of the manuscript.
Acknowledgments
We would like to thank breast and lung cancer patients worldwide, whose experiences inspired this work, with special recognition to the patients at Hospital Prof. Doutor Fernando Fonseca. We also gratefully acknowledge the support of the Research Unit iNOVA4Health (UID/4462/2025) and the Associated Laboratory LS4FUTURE (LA/P/0087/2020), both financially supported by Fundação para a Ciência e Tecnologia / Ministério da Educação, Ciência e Inovação. This work was further supported by grants from Fundação para a Ciência e Tecnologia (2021.08031.BD and 2025.00568.BD).
Conflicts of Interest/ Disclosure
The authors declare that this review was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
List of abbreviations
| CEBP | CCAAT/Enhancer Binding Protein |
| CTLs | Cytotoxic T lymphocytes |
| CXCL | C-X-C motif chemokine ligand |
| CXCR | C-X-C chemokine receptor |
| ECM | Extracellular matrix |
| G-CSF | Granulocyte colony-stimulating factor |
| GFI-1 | Growth factor independent 1 |
| GMPs | Granulocyte–monocyte progenitors |
| HDNs | High-density neutrophils |
| HSCs | Hematopoietic stem cells |
| IL | Interleukin |
| JAK/STAT | Janus kinase / Signal transducer and activator of transcription |
| LDNs | Low-density neutrophils |
| MAPK | Mitogen-activated protein kinase |
| MHC | Major histocompatibility complex |
| MMPs | Matrix metalloproteinases |
| MPO | Myeloperoxidase |
| NADPH | Nicotinamide adenine dinucleotide phosphate |
| NE | Neutrophil elastase |
| NETs | Neutrophil extracellular traps |
| NETosis | NET-mediated neutrophil cell death |
| NLR | Neutrophil-to-lymphocyte ratio |
| NOD | Nucleotide-binding oligomerization domain |
| NSCLC | Non-small cell lung carcinoma |
| PAD4 | Protein-arginine deiminase 4 |
| PBMC | Peripheral blood mononuclear cell |
| PD-L1 | Programmed death-ligand 1 |
| PRRs | Pattern recognition receptors |
| PU.1 | PU box-binding protein 1 |
| RNS | Reactive nitrogen species |
| ROS | Reactive oxygen species |
| RUNX1 | Runt-related transcription factor 1 |
| TANs | Tumor-associated neutrophils |
| TGF-β | Transforming growth factor-β |
| TIME | Tumor immune microenvironment |
| TLRs | Toll-like receptors |
| TME | Tumor microenvironment |
| TNF-α | Tumor necrosis factor-α |
| TRAIL | TNF-related apoptosis-inducing ligand |
| VEGF | Vascular endothelial growth factor |
References
- Kraus, R.F.; Gruber, M.A. Neutrophils—From Bone Marrow to First-Line Defense of the Innate Immune System. Front. Immunol. 2021, 12, 767175. [Google Scholar] [CrossRef]
- Kolaczkowska, E.; Kubes, P. Neutrophil recruitment and function in health and inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef]
- Mayadas, T.N.; Cullere, X.; Lowell, C.A. The Multifaceted Functions of Neutrophils. Annu. Rev. Pathol. Mech. Dis. 2014, 9, 181–218. [Google Scholar] [CrossRef]
- Lehman, H.K.; Segal, B.H. The role of neutrophils in host defense and disease. J. Allergy Clin. Immunol. 2020, 145, 1535–1544. [Google Scholar] [CrossRef]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil Extracellular Traps Kill Bacteria. Science (80- ) 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- Fuchs, T.A.; Abed, U.; Goosmann, C.; Hurwitz, R.; Schulze, I.; Wahn, V.; Weinrauch, Y.; Brinkmann, V.; Zychlinsky, A. Novel cell death program leads to neutrophil extracellular traps. J. Cell Biol. 2007, 176, 231–241. [Google Scholar] [CrossRef]
- Malech, H.L.; DeLeo, F.R.; Quinn, M.T. The Role of Neutrophils in the Immune System: An Overview. 2014, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Carnevale, S.; Di Ceglie, I.; Grieco, G.; Rigatelli, A.; Bonavita, E.; Jaillon, S. Neutrophil diversity in inflammation and cancer. Front. Immunol. 2023, 14, 1180810. [Google Scholar] [CrossRef]
- Wu, G.; Pan, B.; Shi, H.; Yi, Y.; Zheng, X.; Ma, H.; Zhao, M.; Zhang, Z.; Cheng, L.; Huang, Y.; et al. Neutrophils’ dual role in cancer: from tumor progression to immunotherapeutic potential. Int. Immunopharmacol. 2024, 140, 112788. [Google Scholar] [CrossRef] [PubMed]
- Fridlender, Z.G.; Sun, J.; Kim, S.; Kapoor, V.; Cheng, G.; Ling, L.; Worthen, G.S.; Albelda, S.M. Polarization of Tumor-Associated Neutrophil Phenotype by TGF-β: “N1” versus “N2” TAN. Cancer Cell 2009, 16, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Ohms, M.; Möller, S.; Laskay, T. An Attempt to Polarize Human Neutrophils Toward N1 and N2 Phenotypes in vitro. Front. Immunol. 2020, 11, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, D.P.; Correia, B.F.; Salvador, R.; de Sousa, N.; Jacinto, A.; Braga, S.; Cabral, M.G. Circulating low density neutrophils of breast cancer patients are associated with their worse prognosis due to the impairment of T cell responses. Oncotarget 2021, 12, 2388–2403. [Google Scholar] [CrossRef] [PubMed]
- Sagiv, J.Y.; Michaeli, J.; Assi, S.; Mishalian, I.; Kisos, H.; Levy, L.; Damti, P.; Lumbroso, D.; Polyansky, L.; Sionov, R.V.; et al. Phenotypic Diversity and Plasticity in Circulating Neutrophil Subpopulations in Cancer. Cell Rep. 2015, 10, 562–573. [Google Scholar] [CrossRef] [PubMed]
- Scapini, P.; Marini, O.; Tecchio, C.; Cassatella, M.A. Human neutrophils in the saga of cellular heterogeneity: insights and open questions. Immunol. Rev. 2016, 273, 48–60. [Google Scholar] [CrossRef]
- Vanhaver, C.; Aboubakar Nana, F.; Delhez, N.; Luyckx, M.; Hirsch, T.; Bayard, A.; Houbion, C.; Dauguet, N.; Brochier, A.; van der Bruggen, P.; et al. Immunosuppressive low-density neutrophils in the blood of cancer patients display a mature phenotype. Life Sci. Alliance 2024, 7, e202302332. [Google Scholar] [CrossRef]
- Shaul, M.E.; Eyal, O.; Guglietta, S.; Aloni, P.; Zlotnik, A.; Forkosh, E.; Levy, L.; Weber, L.M.; Levin, Y.; Pomerantz, A.; et al. Circulating neutrophil subsets in advanced lung cancer patients exhibit unique immune signature and relate to prognosis. FASEB J. 2020, 34, 4204–4218. [Google Scholar] [CrossRef]
- Correia, B.F.; Grosa, D.; Salvador, R.; Brites, I.; Martins, T.; Vitorino, M.; Sousa, C.X.; Cristóvão-Ferreira, S.; Braga, S.; Jacinto, A.; et al. Neutrophils matter: new clinical insights on their role in the progression of metastatic breast cancer. Breast Cancer Res. 2025, 27, 223. [Google Scholar] [CrossRef]
- Valadez-Cosmes, P.; Maitz, K.; Kindler, O.; Raftopoulou, S.; Kienzl, M.; Santiso, A.; Mihalic, Z.N.; Brcic, L.; Lindenmann, J.; Fediuk, M.; et al. Identification of Novel Low-Density Neutrophil Markers Through Unbiased High-Dimensional Flow Cytometry Screening in Non-Small Cell Lung Cancer Patients. Front Immunol 2021, 12, 703846. [Google Scholar] [CrossRef]
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global cancer statistics 2024, GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin 2022, 74, 229–263. [Google Scholar] [CrossRef]
- Cowland, J.B.; Borregaard, N. Granulopoiesis and granules of human neutrophils. Immunol. Rev. 2016, 273, 11–28. [Google Scholar] [CrossRef]
- Pillay, J.; den Braber, I.; Vrisekoop, N.; Kwast, L.M.; de Boer, R.J.; Borghans, J.A.M.; Tesselaar, K.; Koenderman, L. In vivo labeling with 2H2O reveals a human neutrophil lifespan of 5.4 days. Blood 2010, 116, 625–627. [Google Scholar] [CrossRef]
- Othman, A.; Sekheri, M.; Filep, J.G. Roles of neutrophil granule proteins in orchestrating inflammation and immunity. FEBS J. 2022, 289, 3932–3953. [Google Scholar] [CrossRef]
- Baz, A.A.; Hao, H.; Lan, S.; Li, Z.; Liu, S.; Chen, S.; Chu, Y. Neutrophil extracellular traps in bacterial infections and evasion strategies. Front Immunol 2024, 15, 1357967. [Google Scholar] [CrossRef] [PubMed]
- Sadik, C.D.; Kim, N.D.; Luster, A.D. Neutrophils cascading their way to inflammation. Trends Immunol. 2011, 32, 452–460. [Google Scholar] [CrossRef]
- Hyun, Y.-M.; Hong, C.-W. Deep insight into neutrophil trafficking in various organs. J. Leukoc. Biol. 2017, 102, 617–629. [Google Scholar] [CrossRef] [PubMed]
- Herrero-Cervera, A.; Soehnlein, O.; Kenne, E. Neutrophils in chronic inflammatory diseases. Cell Mol. Immunol. 2022, 19, 177–191. [Google Scholar] [CrossRef]
- Silvestre-Roig, C.; Braster, Q.; Ortega-Gomez, A.; Soehnlein, O. Neutrophils as regulators of cardiovascular inflammation. Nat. Rev. Cardiol. 2020, 17, 327–340. [Google Scholar] [CrossRef] [PubMed]
- Overbeeke, C.; Tak, T.; Koenderman, L. The journey of neutropoiesis: how complex landscapes in bone marrow guide continuous neutrophil lineage determination. Blood 2022, 139, 2285–2293. [Google Scholar] [CrossRef]
- Hageb, A.; Farjia, M.; Osei-Sarpong, C.; Silvestre-Roig, C. Ontogenetic drivers of neutrophil heterogeneity. Exp. Hematol. 2025, 151, 104863. [Google Scholar] [CrossRef]
- Montaldo, E.; Lusito, E.; Bianchessi, V.; Caronni, N.; Scala, S.; Basso-Ricci, L.; Cantaffa, C.; Masserdotti, A.; Barilaro, M.; Barresi, S.; et al. Cellular and transcriptional dynamics of human neutrophils at steady state and upon stress. Nat. Immunol. 2022, 23, 1470–1483. [Google Scholar] [CrossRef]
- Thind, M.K.; Uhlig, H.H.; Glogauer, M.; Palaniyar, N.; Bourdon, C.; Gwela, A.; Lancioni, C.L.; Berkley, J.A.; Bandsma, R.H.J.; Farooqui, A. A metabolic perspective of the neutrophil life cycle: new avenues in immunometabolism. Front. Immunol. 2024, 14. [Google Scholar] [CrossRef]
- Friedman, A.D. C/EBPα in normal and malignant myelopoiesis. Int. J. Hematol. 2015, 101, 330–341. [Google Scholar] [CrossRef]
- Mehta, H.M.; Corey, S.J. G-CSF, the guardian of granulopoiesis. Semin. Immunol. 2021, 54, 101515. [Google Scholar] [CrossRef]
- Rosales, C. Neutrophil: A Cell with Many Roles in Inflammation or Several Cell Types? Front Physiol 2018, 9, 113. [Google Scholar] [CrossRef] [PubMed]
- McKenna, E.; Mhaonaigh, A.U.; Wubben, R.; Dwivedi, A.; Hurley, T.; Kelly, L.A.; Stevenson, N.J.; Little, M.A.; Molloy, E.J. Neutrophils: Need for Standardized Nomenclature. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef]
- Elghetany, M.T. Surface Antigen Changes during Normal Neutrophilic Development: A Critical Review. Blood Cells, Mol Dis 2002, 28, 260–274. [Google Scholar] [CrossRef]
- Häger, M.; Cowland, J.B.; Borregaard, N. Neutrophil granules in health and disease. J. Intern. Med. 2010, 268, 25–34. [Google Scholar] [CrossRef]
- Zhang, F.; Xia, Y.; Su, J.; Quan, F.; Zhou, H.; Li, Q.; Feng, Q.; Lin, C.; Wang, D.; Jiang, Z. Neutrophil diversity and function in health and disease. Signal Transduct. Target. Ther. 2024, 9, 343. [Google Scholar] [CrossRef]
- Silvestre-Roig, C.; Hidalgo, A.; Soehnlein, O. Neutrophil heterogeneity: implications for homeostasis and pathogenesis. Blood 2016, 127, 2173–2181. [Google Scholar] [CrossRef] [PubMed]
- Tsioumpekou, M.; Krijgsman, D.; Leusen, J.H.W.; Olofsen, P.A. The Role of Cytokines in Neutrophil Development, Tissue Homing, Function and Plasticity in Health and Disease. Cells 2023, 12, 1981. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, W.; Yang, F.; Xu, Y.; Feng, C.; Zhao, Y. The regulatory roles of neutrophils in adaptive immunity. Cell Commun. Signal 2019, 17, 147. [Google Scholar] [CrossRef]
- Mantovani, A.; Cassatella, M.A.; Costantini, C.; Jaillon, S. Neutrophils in the activation and regulation of innate and adaptive immunity. Nat. Rev. Immunol. 2011, 11, 519–531. [Google Scholar] [CrossRef]
- Shafqat, A.; Khan, J.A.; Alkachem, A.Y.; Sabur, H.; Alkattan, K.; Yaqinuddin, A.; Sing, G.K. How Neutrophils Shape the Immune Response: Reassessing Their Multifaceted Role in Health and Disease. Int. J. Mol. Sci. 2023, 24, 17583. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef]
- Uribe-Querol, E.; Rosales, C. Neutrophils in Cancer: Two Sides of the Same Coin. J. Immunol. Res. 2015, 2015, 1–21. [Google Scholar] [CrossRef]
- Garcia-Flores, L.A.; Dawid De Vera, M.T.; Pilo, J.; Rego, A.; Gomez-Casado, G.; Arranz-Salas, I.; Hierro Martín, I.; Alcaide, J.; Torres, E.; Ortega-Gomez, A.; et al. Increased neutrophil counts are associated with poor overall survival in patients with colorectal cancer: a five-year retrospective analysis. Front. Immunol. 2024, 15, 1415804. [Google Scholar] [CrossRef]
- Ma, J.; Kuzman, J.; Ray, A.; Lawson, B.O.; Khong, B.; Xuan, S.; Hahn, A.W.; Khong, H.T. Neutrophil-to-lymphocyte Ratio (NLR) as a predictor for recurrence in patients with stage III melanoma. Sci. Rep. 2018, 8, 4044. [Google Scholar] [CrossRef]
- Dirican, A.; Kucukzeybek, B.B.; Alacacioglu, A.; Kucukzeybek, Y.; Erten, C.; Varol, U.; Somali, I.; Demir, L.; Bayoglu, I.V.; Yildiz, Y.; et al. Do the derived neutrophil to lymphocyte ratio and the neutrophil to lymphocyte ratio predict prognosis in breast cancer? Int. J. Clin. Oncol. 2015, 20, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Shen, G.; Zhou, X.; Li, J. Therapeutic potential of tumor-associated neutrophils: dual role and phenotypic plasticity. Signal Transduct. Target. Ther. 2025, 10, 178. [Google Scholar] [CrossRef] [PubMed]
- Grecian, R.; Whyte, M.K.B.; Walmsley, S.R. The role of neutrophils in cancer. Br. Med. Bull. 2018, 128, 5–14. [Google Scholar] [CrossRef]
- SenGupta, S.; Hein, L.E.; Parent, C.A. The Recruitment of Neutrophils to the Tumor Microenvironment Is Regulated by Multiple Mediators. Front. Immunol. 2021, 12, 734188. [Google Scholar] [CrossRef]
- Shaul, M.E.; Eyal, O.; Guglietta, S.; Aloni, P.; Zlotnik, A.; Forkosh, E.; Levy, L.; Weber, L.M.; Levin, Y.; Pomerantz, A.; et al. Circulating neutrophil subsets in advanced lung cancer patients exhibit unique immune signature and relate to prognosis. FASEB J. 2020, 34, 4204–4218. [Google Scholar] [CrossRef]
- Arasanz, H.; Bocanegra, A.I.; Morilla, I.; Fernández-Irigoyen, J.; Martínez-Aguillo, M.; Teijeira, L.; Garnica, M.; Blanco, E.; Chocarro, L.; Ausin, K.; et al. Circulating Low Density Neutrophils Are Associated with Resistance to First Line Anti-PD1/PDL1 Immunotherapy in Non-Small Cell Lung Cancer. Cancers 2022, 14, 3846. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, R.; Lo Russo, G.; Signorelli, D.; Proto, C.; Prelaj, A.; Galli, G.; De Toma, A.; Viscardi, G.; Lobefaro, R.; Brambilla, M.; et al. Circulating and tumor-associated neutrophil subtypes discriminate hyperprogressive disease (HPD) from conventional progression (PD) upon immune checkpoint inhibitors (ICI) in advanced non-small cell lung cancer (NSCLC) patients (pts) and in vivo models. J. Clin. Oncol. 2020, 38, 9547–9547. [Google Scholar] [CrossRef]
- Arrazubi, V.; Goñi, S.; González-Borja, I.; Hernandez-Garcia, I.; Arasanz, H.; Pérez-Sanz, J.; Bocanegra, A.I.; Kochan, G.; Escors, D.; Ruiz de Azúa, Y.; et al. Circulating low density neutrophils are associated with resistance to anti-PD1 immunotherapy in squamous head and neck cancer. Head. Neck 2023, 45, 3075–3085. [Google Scholar] [CrossRef]
- Wang, Y.; Yao, R.; Zhang, D.; Chen, R.; Ren, Z.; Zhang, L. Circulating Neutrophils Predict Poor Survival for HCC and Promote HCC Progression Through p53 and STAT3 Signaling Pathway. J. Cancer 2020, 11, 3736–3744. [Google Scholar] [CrossRef]
- Fernandes, P.C.; Garcia, C.B.; Micheli, D.C.; Cunha, F.Q.; Murta, E.F.C.; Tavares-Murta, B.M. Circulating neutrophils may play a role in the host response in cervical cancer. Int. J. Gynecol. Cancer 2007, 17, 1068–1074. [Google Scholar] [CrossRef] [PubMed]
- Cerezo-Wallis, D.; Ballesteros, I. Neutrophils in cancer, a love–hate affair. FEBS J. 2022, 289, 3692–3703. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Ikenaga, N.; Nakata, K.; Higashijima, N.; Zhong, P.; Kubo, A.; Wu, C.; Tsutsumi, C.; Shimada, Y.; Hayashi, M.; et al. Tumor-associated neutrophils upregulate Nectin2 expression, creating the immunosuppressive microenvironment in pancreatic ductal adenocarcinoma. J. Exp. Clin. Cancer Res. 2024, 43, 258. [Google Scholar] [CrossRef]
- Wang, Z.; Hu, H.; Bao, Y.; Pang, L.; Yang, C. Neutrophils in cancer: from immune defense to tumor promotion. Cancer Biol. Med. 2025, 1–20. [Google Scholar] [CrossRef]
- Obeagu, E.I.; Ezeala, C.C. Neutrophils as key regulators of tumor microenvironment in breast cancer: a focus on N1 and N2 polarization. Ann. Med. Surg. 2025, 87, 3509–3522. [Google Scholar] [CrossRef]
- Toghraie, F.S.; Bayat, M.; Hosseini, M.S.; Ramezani, A. Tumor-infiltrating myeloid cells; mechanisms, functional significance, and targeting in cancer therapy. Cell Oncol. 2025, 48, 559–590. [Google Scholar] [CrossRef]
- Shaul, M.E.; Levy, L.; Sun, J.; Mishalian, I.; Singhal, S.; Kapoor, V.; Horng, W.; Fridlender, G.; Albelda, S.M.; Fridlender, Z.G. Tumor-associated neutrophils display a distinct N1 profile following TGFβ modulation: A transcriptomics analysis of pro- vs. antitumor TANs. Oncoimmunology 2016, 5, e1232221. [Google Scholar] [CrossRef]
- Fang, Q.; Stehr, A.M.; Naschberger, E.; Knopf, J.; Herrmann, M.; Stürzl, M. No NETs no TIME: Crosstalk between neutrophil extracellular traps and the tumor immune microenvironment. Front. Immunol. 2022, 13, 1075260. [Google Scholar] [CrossRef] [PubMed]
- Cools-Lartigue, J.; Spicer, J.; McDonald, B.; Gowing, S.; Chow, S.; Giannias, B.; Bourdeau, F.; Kubes, P.; Ferri, L. Neutrophil extracellular traps sequester circulating tumor cells and promote metastasis. J. Clin. Invest. 2013, 123, 3446–3458. [Google Scholar] [CrossRef]
- Koenderman, L.; Vrisekoop, N. Neutrophils in cancer: from biology to therapy. Cell Mol. Immunol. 2024, 22, 4–23. [Google Scholar] [CrossRef]
- Howard, R.; Kanetsky, P.A.; Egan, K.M. Exploring the prognostic value of the neutrophil-to-lymphocyte ratio in cancer. Sci. Rep. 2019, 9, 19673. [Google Scholar] [CrossRef]
- Heshmat-Ghahdarijani, K.; Sarmadi, V.; Heidari, A.; Falahati Marvasti, A.; Neshat, S.; Raeisi, S. The neutrophil-to-lymphocyte ratio as a new prognostic factor in cancers: a narrative review. Front. Oncol. 2023, 13, 1228076. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.-H.; Tuo, M.; Ye, C.; Wu, X.-F.; Wang, H.-H.; Ren, W.-Z.; Liu, G.; Xiang, T. Prognostic value of neutrophil-to-lymphocyte ratio in gastric cancer patients undergoing neoadjuvant chemotherapy: A systematic review and meta-analysis. World J. Gastrointest. Oncol. 2024, 16, 4477–4488. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Wang, J.; Wang, X.; Gu, L.; Pei, H.; Kuai, S.; Zhang, Y.; Shang, Z. Prognostic value of the neutrophil to lymphocyte ratio in lung cancer: A meta-analysis. Clinics 2015, 70, 524–530. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, X.; Xu, R.; Luo, G.; Wang, X. Neutrophil-to-lymphocyte ratio as a prognostic factor in patients with castration-resistant prostate cancer treated with docetaxel-based chemotherapy: a meta-analysis. BMC Urol. 2025, 25, 17. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Liang, Y.; Tang, X.; Qu, H. Decrease in Blood Neutrophil-to-Lymphocyte Ratio Indicates Better Survival After Neoadjuvant Chemotherapy in Patients With Advanced Gastric Cancer. Front. Surg. 2021, 8, 745748. [Google Scholar] [CrossRef]
- Liu, S.; Wu, W.; Du, Y.; Yin, H.; Chen, Q.; Yu, W.; Wang, W.; Yu, J.; Liu, L.; Lou, W.; et al. The evolution and heterogeneity of neutrophils in cancers: origins, subsets, functions, orchestrations and clinical applications. Mol. Cancer 2023, 22, 148. [Google Scholar] [CrossRef]
- Hacbarth, E.; Kajdacsy-Balla, A. Low density neutrophils in patients with systemic lupus erythematosus, rheumatoid arthritis, and acute rheumatic fever. Arthritis Rheum. 1986, 29, 1334–1342. [Google Scholar] [CrossRef] [PubMed]
- Rankin, A.N.; Hendrix, S.V.; Naik, S.K.; Stallings, C.L. Exploring the Role of Low-Density Neutrophils During Mycobacterium tuberculosis Infection. Front. Cell Infect. Microbiol. 2022, 12. [Google Scholar] [CrossRef]
- Jaillon, S.; Ponzetta, A.; Di Mitri, D.; Santoni, A.; Bonecchi, R.; Mantovani, A. Neutrophil diversity and plasticity in tumour progression and therapy. Nat. Rev. Cancer 2020, 20, 485–503. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Hu, Y.; Gu, F.; Liang, J.; Zeng, Y.; Hong, X.; Zhang, K.; Liu, L. Phenotypic and clinical characterization of low density neutrophils in patients with advanced lung adenocarcinoma. Oncotarget 2017, 8, 90969–90978. [Google Scholar] [CrossRef]
- Hsu, B.E.; Tabariès, S.; Johnson, R.M.; Andrzejewski, S.; Senecal, J.; Lehuédé, C.; Annis, M.G.; Ma, E.H.; Völs, S.; Ramsay, L.; et al. Immature Low-Density Neutrophils Exhibit Metabolic Flexibility that Facilitates Breast Cancer Liver Metastasis. Cell Rep. 2019, 27, 3902–3915.e6. [Google Scholar] [CrossRef]
- Futoh, Y.; Kumagai, Y.; Miyato, H.; Ozawa, H.; Kanamaru, R.; Sadatomo, A.; Ohnishi, Y.; Koinuma, K.; Horie, H.; Yamaguchi, H.; et al. Peripheral low-density granulocytes after colorectal cancer surgery in predicting recurrence. BJS Open 2023, 7. [Google Scholar] [CrossRef]
- Krieg, C.; Shaul, M.; Guglietta, S.; Weber, L.; Robinson, M.; Fridlender, Z. Circulating Low Density Neutrophils In Advanced Lung Cancer Patients Exhibit Unique Immune Signatures and affect prognosis. J. Immunol. 2020, 204, 164.11–164.11. [Google Scholar] [CrossRef]
- Fu, Y.; Wen, Z.; Fan, J. Interaction of low-density neutrophils with other immune cells in the mechanism of inflammation. Mol. Med. 2025, 31, 133. [Google Scholar] [CrossRef]
- Wang, X.; Qiu, L.; Li, Z.; Wang, X.-Y.; Yi, H. Understanding the Multifaceted Role of Neutrophils in Cancer and Autoimmune Diseases. Front Immunol 2018, 9, 2456. [Google Scholar] [CrossRef]
- Hassani, M.; Hellebrekers, P.; Chen, N.; van Aalst, C.; Bongers, S.; Hietbrink, F.; Koenderman, L.; Vrisekoop, N. On the origin of low-density neutrophils. J. Leukoc. Biol. 2020, 107, 809–818. [Google Scholar] [CrossRef]
- Ng, M.S.F.; Kwok, I.; Tan, L.; Shi, C.; Cerezo-Wallis, D.; Tan, Y.; Leong, K.; Calvo, G.F.; Yang, K.; Zhang, Y.; et al. Deterministic reprogramming of neutrophils within tumors. Science (80- ) 2024, 383. [Google Scholar] [CrossRef]
- Giese, M.A.; Hind, L.E.; Huttenlocher, A. Neutrophil plasticity in the tumor microenvironment. Blood 2019, 133, 2159–2167. [Google Scholar] [CrossRef]
- Brandau, S.; Moses, K.; Lang, S. The kinship of neutrophils and granulocytic myeloid-derived suppressor cells in cancer: Cousins, siblings or twins? Semin. Cancer Biol. 2013, 23, 171–182. [Google Scholar] [CrossRef]
- Curi, R.; Levada-Pires, A.C.; Silva, E.B.D.; Poma, S.O.; Zambonatto, R.F.; Domenech, P.; Almeida, M.M.; Gritte, R.B.; Souza-Siqueira, T.; Gorjão, R.; et al. The Critical Role of Cell Metabolism for Essential Neutrophil Functions. Cell Physiol. Biochem. 2020, 54, 629–647. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Saxena, S.; Goel, P.; Prajapati, D.R.; Wang, C.; Singh, R.K. Breast Cancer Cell–Neutrophil Interactions Enhance Neutrophil Survival and Pro-Tumorigenic Activities. Cancers 2020, 12, 2884. [Google Scholar] [CrossRef]
- Ettel, P.; Weichhart, T. Not just sugar: metabolic control of neutrophil development and effector functions. J. Leukoc. Biol. 2024, 116, 487–510. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Lu, M.; Shi, J.; Gong, Z.; Hua, L.; Li, Q.; Lim, B.; Zhang, X.H.-F.; Chen, X.; Li, S.; et al. Lung mesenchymal cells elicit lipid storage in neutrophils that fuel breast cancer lung metastasis. Nat. Immunol. 2020, 21, 1444–1455. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Shi, J.; Shen, J.; Fan, X. Metabolic reprogramming of neutrophils in the tumor microenvironment: Emerging therapeutic targets. Cancer Lett. 2025, 612, 217466. [Google Scholar] [CrossRef]
- Rice, C.M.; Davies, L.C.; Subleski, J.J.; Maio, N.; Gonzalez-Cotto, M.; Andrews, C.; Patel, N.L.; Palmieri, E.M.; Weiss, J.M.; Lee, J.-M.; et al. Tumour-elicited neutrophils engage mitochondrial metabolism to circumvent nutrient limitations and maintain immune suppression. Nat. Commun. 2018, 9, 5099. [Google Scholar] [CrossRef] [PubMed]
- Bodac, A.; Meylan, E. Neutrophil metabolism in the cancer context. Semin. Immunol. 2021, 57, 101583. [Google Scholar] [CrossRef]
- Arpinati, L.; Kaisar-Iluz, N.; Shaul, M.E.; Groth, C.; Umansky, V.; Fridlender, Z.G. Tumor-Derived Factors Differentially Affect the Recruitment and Plasticity of Neutrophils. Cancers 2021, 13, 5082. [Google Scholar] [CrossRef]
- Wang, X.; Qu, Y.; Xu, Q.; Jiang, Z.; Wang, H.; Lin, B.; Cao, Z.; Pan, Y.; Li, S.; Hu, Y.; et al. NQO1 Triggers Neutrophil Recruitment and NET Formation to Drive Lung Metastasis of Invasive Breast Cancer. Cancer Res. 2024, 84, 3538–3555. [Google Scholar] [CrossRef]
- Lecot, P.; Ardin, M.; Dussurgey, S.; Alcazer, V.; Moudombi, L.; Pereira Abrantes, M.; Hubert, M.; Swalduz, A.; Hernandez-Vargas, H.; Viari, A.; et al. Gene signature of circulating platelet-bound neutrophils is associated with poor prognosis in cancer patients. Int. J. Cancer 2022, 151, 138–152. [Google Scholar] [CrossRef]
- Tay, S.H.; Zharkova, O.; Lee, H.Y.; Toh, M.M.X.; Libau, E.A.; Celhar, T.; Narayanan, S.; Ahl, P.J.; Ong, W.Y.; Joseph, C.; et al. Platelet TLR7 is essential for the formation of platelet–neutrophil complexes and low-density neutrophils in lupus nephritis. Rheumatology 2024, 63, 551–562. [Google Scholar] [CrossRef] [PubMed]
- Hsu, B.E.; Roy, J.; Mouhanna, J.; Rayes, R.F.; Ramsay, L.; Tabariès, S.; Annis, M.G.; Watson, I.R.; Spicer, J.D.; Costantino, S.; et al. C3a elicits unique migratory responses in immature low-density neutrophils. Oncogene 2020, 39, 2612–2623. [Google Scholar] [CrossRef] [PubMed]
- Mezquita, L.; Preeshagul, I.; Auclin, E.; Saravia, D.; Hendriks, L.; Rizvi, H.; Park, W.; Nadal, E.; Martin-Romano, P.; Ruffinelli, J.C.; et al. Predicting immunotherapy outcomes under therapy in patients with advanced NSCLC using dNLR and its early dynamics. Eur. J. Cancer 2021, 151, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Carrión-Barberà, I.; Lood, C. Performance of the neutrophil-to-lymphocyte ratio as a prognostic tool for survival in solid cancers. Front. Oncol. 2025, 15, 1616477. [Google Scholar] [CrossRef]
- Jadoon, S.K.; Soomro, R.; Ahsan, M.N.; Ijaz Khan, R.M.; Iqbal, S.; Yasmin, F.; Najeeb, H.; Saleem, N.; Cho, N.; Resham Shaikh, T.G.; et al. Association of neutrophil-to-lymphocyte ratio with clinical, pathological, radiological, laboratory features and disease outcomes of invasive breast cancer patients: A retrospective observational cohort study. Medicine 2023, 102, e33811. [Google Scholar] [CrossRef] [PubMed]
- Obeagu, E.I.; Obeagu, G.U. Exploring neutrophil functionality in breast cancer progression: A review. Medicine 2024, 103, e37654. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Shen, Y.; Huang, H.; Pan, S.; Jiang, J.; Chen, W.; Zhang, T.; Zhang, C.; Ni, C. A Rosetta Stone for Breast Cancer: Prognostic Value and Dynamic Regulation of Neutrophil in Tumor Microenvironment. Front. Immunol. 2020, 11, 1779. [Google Scholar] [CrossRef]
- Chen, J.; Deng, Q.; Pan, Y.; He, B.; Ying, H.; Sun, H.; Liu, X.; Wang, S. Prognostic value of neutrophil-to-lymphocyte ratio in breast cancer. FEBS Open Bio 2015, 5, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Garner, H.; Martinovic, M.; Liu, N.Q.; Bakker, N.A.M.; Velilla, I.Q.; Hau, C.-S.; Vrijland, K.; Kaldenbach, D.; Kok, M.; de Wit, E.; et al. Understanding and reversing mammary tumor-driven reprogramming of myelopoiesis to reduce metastatic spread. Cancer Cell 2025, 43, 1279–1295.e9. [Google Scholar] [CrossRef]
- Gianni, C.; Palleschi, M.; Schepisi, G.; Casadei, C.; Bleve, S.; Merloni, F.; Sirico, M.; Sarti, S.; Cecconetto, L.; Di Menna, G.; et al. Circulating inflammatory cells in patients with metastatic breast cancer: Implications for treatment. Front. Oncol. 2022, 12, 882896. [Google Scholar] [CrossRef]
- Bakker, N.A.M.; Garner, H.; van Dyk, E.; Champanhet, E.; Klaver, C.; Duijst, M.; Voorwerk, L.; Nederlof, I.; Voorthuis, R.; Liefaard, M.C.; et al. Triple-negative breast cancer modifies the systemic immune landscape and alters neutrophil functionality. npj Breast Cancer 2025, 11, 5. [Google Scholar] [CrossRef]
- Ramessur, A.; Ambasager, B.; Valle Aramburu, I.; Peakman, F.; Gleason, K.; Lehmann, C.; Papayannopoulos, V.; Coombes, R.C.; Malanchi, I. Circulating neutrophils from patients with early breast cancer have distinct subtype-dependent phenotypes. Breast Cancer Res. 2023, 25, 125. [Google Scholar] [CrossRef]
- Unanua, N.C.; Fontecha, O.; Sanchez, L.T.; Martínez-Aguillo, M.; Ruiz, I.M.; Labiano, I.; Huerta-Hernández, A.E.; Lecumberri, A.; Caseda, I.; Maqueda, M.A.; et al. 6P Circulating low-density neutrophils (LDNs) are associated with resistance to immunotherapy as frontline treatment for non-small cell lung cancer (NSCLC): Updated results and proteomic characterization. Ann. Oncol. 2024, 35, S217. [Google Scholar] [CrossRef]
- Ohashi, K.; Nishito, Y.; Fukuda, H.; Sadahiro, R.; Yoshida, Y.; Watanabe, S.; Motoi, N.; Sonobe, Y.; Mizuno, H.; Tsunoda, H.; et al. Neutrophil-to-lymphocyte ratio is a prognostic factor reflecting immune condition of tumor microenvironment in squamous cell lung cancer. Sci. Rep. 2024, 14, 429. [Google Scholar] [CrossRef]
- Zhang, C.; Huo, Y.; Shang, X.; Zhang, T.; Tang, N.; Wang, H. Tumor mutational burden adjusted by neutrophil-to-lymphocyte ratio serves as a potential biomarker for atezolizumab-treated patients with extensive stage small cell lung cancer. Respir. Res. 2024, 25, 253. [Google Scholar] [CrossRef]
- Mariean, C.R.; Tiucă, O.M.; Mariean, A.; Szekely, T.-B.; Niculescu, R.; Sabau, A.H.; Al-Akel, C.F.; Cotoi, O.S. The Impact of the Histologic Types of Lung Cancer on CBC-Derived Inflammatory Markers—Current Knowledge and Future Perspectives. J. Clin. Med. 2025, 14, 3038. [Google Scholar] [CrossRef]
- Zhou, J.; Liu, H.; Jiang, S.; Wang, W. Role of tumor-associated neutrophils in lung cancer (Review). Oncol. Lett. 2022, 25, 2. [Google Scholar] [CrossRef]
- Yamazaki, N.; Kiyohara, Y.; Uhara, H.; Iizuka, H.; Uehara, J.; Otsuka, F.; Fujisawa, Y.; Takenouchi, T.; Isei, T.; Iwatsuki, K.; et al. Cytokine biomarkers to predict antitumor responses to nivolumab suggested in a phase 2 study for advanced melanoma. Cancer Sci. 2017, 108, 1022–1031. [Google Scholar] [CrossRef] [PubMed]
- Laino, A.S.; Woods, D.; Vassallo, M.; Qian, X.; Tang, H.; Wind-Rotolo, M.; Weber, J. Serum interleukin-6 and C-reactive protein are associated with survival in melanoma patients receiving immune checkpoint inhibition. J. Immunother. Cancer 2020, 8, e000842. [Google Scholar] [CrossRef] [PubMed]
- Tazzyman, S.; Barry, S.T.; Ashton, S.; Wood, P.; Blakey, D.; Lewis, C.E.; Murdoch, C. Inhibition of neutrophil infiltration into A549 lung tumors in vitro and in vivo using a CXCR2-specific antagonist is associated with reduced tumor growth. Int. J. Cancer 2011, 129, 847–858. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, M.H.; Rayes, R.F.; Cools-Lartigue, J.; Spicer, J.D. Neutrophil extracellular traps in cancer. Nat. Rev. Cancer 2025. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Y.; He, Y.; Liang, L. Dysregulated arginine metabolism is associated with pro-tumor neutrophil polarization in liver cancer. Front Immunol 2025, 16, 1673665. [Google Scholar] [CrossRef]
- Lynch, C.; Pitroda, S.P.; Weichselbaum, R.R. Radiotherapy, immunity, and immune checkpoint inhibitors. Lancet Oncol. 2024, 25, e352–e362. [Google Scholar] [CrossRef]
- Principe, D.R.; Kamath, S.D.; Korc, M.; Munshi, H.G. The immune modifying effects of chemotherapy and advances in chemo-immunotherapy. Pharmacol. Ther. 2022, 236, 108111. [Google Scholar] [CrossRef]
- Ning, X.; Wang, W.-M.; Jin, H.-Z. Low-Density Granulocytes in Immune-Mediated Inflammatory Diseases. J. Immunol. Res. 2022, 2022, 1–11. [Google Scholar] [CrossRef]
- Carmona-Rivera, C.; Khaznadar, S.S.; Shwin, K.W.; Irizarry-Caro, J.A.; O’Neil, L.J.; Liu, Y.; Jacobson, K.A.; Ombrello, A.K.; Stone, D.L.; Tsai, W.L.; et al. Deficiency of adenosine deaminase 2 triggers adenosine-mediated NETosis and TNF production in patients with DADA2. Blood 2019, 134, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.W.; Houghton, A.M. Targeting neutrophils for cancer therapy. Nat. Rev. Drug Discov. 2025, 24, 666–684. [Google Scholar] [CrossRef]
- Eruslanov, E.; Nefedova, Y.; Gabrilovich, D.I. The heterogeneity of neutrophils in cancer and its implication for therapeutic targeting. Nat. Immunol. 2025, 26, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Chen, X.; Lu, Y.; Sun, N.; Weisgerber, K.E.; Xu, M.; Bai, R.-Y. Neutrophil Dynamics in Response to Cancer Therapies. Cancers 2025, 17, 2593. [Google Scholar] [CrossRef]
- Koh, C.-H.; Bhoo-Pathy, N.; Ng, K.-L.; Jabir, R.S.; Tan, G.-H.; See, M.-H.; Jamaris, S.; Taib, N.A. Utility of pre-treatment neutrophil–lymphocyte ratio and platelet–lymphocyte ratio as prognostic factors in breast cancer. Br. J. Cancer 2015, 113, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Wu, J.; Jia, H.; Yang, Y.; Zhang, X.; Chen, K.; Su, F. The Peripheral Blood Neutrophil-To-Lymphocyte Ratio Is Superior to the Lymphocyte-To-Monocyte Ratio for Predicting the Long-Term Survival of Triple-Negative Breast Cancer Patients. PLoS One 2015, 10, e0143061. [Google Scholar] [CrossRef]
- Kim, I.S.; Gao, Y.; Welte, T.; Wang, H.; Liu, J.; Janghorban, M.; Sheng, K.; Niu, Y.; Goldstein, A.; Zhao, N.; et al. Immuno-subtyping of breast cancer reveals distinct myeloid cell profiles and immunotherapy resistance mechanisms. Nat. Cell Biol. 2019, 21, 1113–1126. [Google Scholar] [CrossRef]
Figure 1.
- Neutrophil maturation and phenotypic marker expression. Neutrophils arise from HSCs and GMPs expressing CD34, CD117, and CD33, and progress through promyelocyte and myelocyte stages marked by CD66b, CD15, CD33, and CD49d expression. Transition to metamyelocytes and band cells is accompanied by nuclear remodeling, acquisition of CD16, and loss of CD49d, defining the shift from the mitotic to the post-mitotic pool. Mature neutrophils display a segmented nucleus, high granule content, and elevated CD16 and CD10 expression.
Figure 1.
- Neutrophil maturation and phenotypic marker expression. Neutrophils arise from HSCs and GMPs expressing CD34, CD117, and CD33, and progress through promyelocyte and myelocyte stages marked by CD66b, CD15, CD33, and CD49d expression. Transition to metamyelocytes and band cells is accompanied by nuclear remodeling, acquisition of CD16, and loss of CD49d, defining the shift from the mitotic to the post-mitotic pool. Mature neutrophils display a segmented nucleus, high granule content, and elevated CD16 and CD10 expression.

Figure 2.
- Neutrophil functional heterogeneity and plasticity in cancer. TANs exhibit pronounced phenotypic and functional plasticity within the TME, ranging from antitumoral N1-like states to protumoral N2-like states. N1 neutrophils exert cytotoxic and immunostimulatory functions that support antitumor immunity and restrain early metastatic dissemination. Conversely, N2 neutrophils facilitate tumor progression by promoting angiogenesis, extracellular matrix remodeling, immunosuppression, and metastasis, including through the formation of NETs.
Figure 2.
- Neutrophil functional heterogeneity and plasticity in cancer. TANs exhibit pronounced phenotypic and functional plasticity within the TME, ranging from antitumoral N1-like states to protumoral N2-like states. N1 neutrophils exert cytotoxic and immunostimulatory functions that support antitumor immunity and restrain early metastatic dissemination. Conversely, N2 neutrophils facilitate tumor progression by promoting angiogenesis, extracellular matrix remodeling, immunosuppression, and metastasis, including through the formation of NETs.

Figure 3.
- Circulating neutrophil subsets and phenotypic diversity in cancer. Density gradient centrifugation distinguishes two major subsets of circulating neutrophils: HDNs, which sediment in the granulocytic fraction under homeostatic conditions, and LDNs, which emerge within the PBMC layer in pathological settings, including cancer.
Figure 3.
- Circulating neutrophil subsets and phenotypic diversity in cancer. Density gradient centrifugation distinguishes two major subsets of circulating neutrophils: HDNs, which sediment in the granulocytic fraction under homeostatic conditions, and LDNs, which emerge within the PBMC layer in pathological settings, including cancer.

Figure 4.
- Crosstalk between LDNs and immune cells in inflammation and cancer. LDNs display functional plasticity, engaging with platelets, macrophages, T cells, and B cells to modulate inflammation and tumor progression. High PSGL-1 expression drives LDNs–platelet aggregation, promoting NETs release and creating pro-thrombotic, tumor-permissive, pre-metastatic niches. LDNs communicate bidirectionally with macrophages, amplifying inflammation via NETs-mediated inflammasome activation or evading phagocytosis through inhibitory “don’t-eat-me” signals. While LDNs generally suppress T cell responses through metabolic, oxidative, and checkpoint-dependent mechanisms, thereby promoting an immunosuppressive TME, certain context-and maturation-dependent subsets can instead enhance T cell activation. Emerging evidence also indicates that LDNs modulate B cell activity.
Figure 4.
- Crosstalk between LDNs and immune cells in inflammation and cancer. LDNs display functional plasticity, engaging with platelets, macrophages, T cells, and B cells to modulate inflammation and tumor progression. High PSGL-1 expression drives LDNs–platelet aggregation, promoting NETs release and creating pro-thrombotic, tumor-permissive, pre-metastatic niches. LDNs communicate bidirectionally with macrophages, amplifying inflammation via NETs-mediated inflammasome activation or evading phagocytosis through inhibitory “don’t-eat-me” signals. While LDNs generally suppress T cell responses through metabolic, oxidative, and checkpoint-dependent mechanisms, thereby promoting an immunosuppressive TME, certain context-and maturation-dependent subsets can instead enhance T cell activation. Emerging evidence also indicates that LDNs modulate B cell activity.

Figure 5.
- LDNs are emerging as critical mediators of tumor progression, immunosuppression, and therapy resistance in breast and lung cancer. Elevated LDNs levels associate with aggressive disease, poor clinical outcomes, and reduced responses to chemotherapy and immune checkpoint blockade, particularly in TNBC and NSCLC. Their inflammatory, NETotic, and T cell-suppressive programs position LDNs as both prognostic biomarkers and actionable therapeutic targets across solid tumors.
Figure 5.
- LDNs are emerging as critical mediators of tumor progression, immunosuppression, and therapy resistance in breast and lung cancer. Elevated LDNs levels associate with aggressive disease, poor clinical outcomes, and reduced responses to chemotherapy and immune checkpoint blockade, particularly in TNBC and NSCLC. Their inflammatory, NETotic, and T cell-suppressive programs position LDNs as both prognostic biomarkers and actionable therapeutic targets across solid tumors.


Table 1.
– LDNs in Breast and Lung Cancer. Table 1 provides an overview of key prospective, observational, and mechanistic studies examining LDNs and dysfunctional neutrophil subsets in breast and lung cancer. Collectively, these studies identify LDNs as markers of poor prognosis and therapeutic resistance and highlight their emergence as products of tumor driven myeloid reprogramming.
Table 1.
– LDNs in Breast and Lung Cancer. Table 1 provides an overview of key prospective, observational, and mechanistic studies examining LDNs and dysfunctional neutrophil subsets in breast and lung cancer. Collectively, these studies identify LDNs as markers of poor prognosis and therapeutic resistance and highlight their emergence as products of tumor driven myeloid reprogramming.
| Cancer Type | Authors, Journal; DOI | Design | Population | Material, Timing | Key Findings |
|---|---|---|---|---|---|
| Breast cancer | Saraiva et al., Oncotarget 2021; [12] doi:10.18632/oncotarget.28135 | Prospective observational | 48 non-metastatic, 12 metastatic;7 healthy donors | Peripheral blood; baseline; fresh biopsies and surgical specimens | LDNs↑ → poor prognosis; ↓CT response; T cell suppression |
| Metastatic breast cancer | Correia et al., Breast Cancer Res. 2025; [17] doi:10.1186/s13058-025-02201-8 | Prospective observational | Patients with metastatic breast cancer | peripheral blood; tissue biopsies 2023-2025 | Neutrophils are associated with tumor progression, increased metastasis, and worse prognosis |
| Breast cancer | Koh et al., British Journal of Cancer 2015; [126] doi:10.1038/bjc.2015.183 | Prospective observational | 1435 patients (stage I-III) | Peripheral blood pre-treatment | High NLR (quintile 5) → worse OS; independent prognostic factor; 5-year survival 51.1% vs 76.4% |
| Breast cancer (TNBC focus) | Jia et al., PLoS One 2015; [127] doi:10.1371/journal.pone.0143061 |
Retrospective cohort | 1570 operable breast cancer patients | Peripheral blood baseline | NLR superior to LMR for prognosis; high NLR → poor DFS/OS in TNBC (HR=2.58 for DFS, HR=3.05 for OS) |
| Breast cancer | Hsu et al., Cell Reports 2019; [78] doi:10.1016/j.celrep.2019.05.091 | Mechanistic (preclinical) | Mouse models of breast cancer liver metastasis | Blood and tissue samples from tumor-bearing mice | iLDNs exhibit metabolic flexibility; promote liver metastasis via NETosis; G-CSF mobilizes iLDNs; HDNs inhibit metastasis |
| Breast cancer | Sagiv et al., Cell Reports 2015; [13] doi:10.1016/j.celrep.2014.12.039 |
Mechanistic (preclinical+clinical samples) | Mouse models + cancer patients | Peripheral blood; density gradient separation | LDNs heterogeneous (immature MDSCs + mature cells); TGF-β-dependent conversion from HDNs; immunosuppressive properties |
| Breast cancer (TNBC) | Kim et al., Nature Cell Biology 2019; [128] doi:10.1038/s41556-019-0373-7 |
Mechanistic (preclinical+clinical datasets) | Multiple murine TNBC models + clinical datasets | Tumor tissue and blood | Neutrophil-enriched subtype resistant to ICI; systemic accumulation of immunosuppressive neutrophils; MES-to-NES conversion mediates acquired ICI resistance |
| Metastatic breast cancer | Bakker et al., NPJ Breast Cancer 2025; [107] doi:10.1038/s41523-025-00721-2 |
Prospective translational | TNBC patients | Peripheral blood; functional assays | Neutrophils showed increased migration, granule protein expression, ROS production, and NETs formation, consistent with pro-tumor LDNs dysfunction |
| Breast cancer | Garner et al., Cancer Cell 2025; [105] doi:10.1016/j.ccell.2025.04.007 |
Preclinical + translational | Mice + human samples | Bone marrow + peripheral blood | Tumor-driven myelopoiesis → dysfunctional LDNs-like and promotes metastasis. Anti-IL-1β reverses these changes and reduces metastatic spread |
| NSCLC | Arasanz et al., Cancers 2022; [53] doi:10.3390/cancers14163846 | Prospective observational | Advanced NSCLC patients (first-line anti-PD1/PDL1) | Peripheral blood; baseline and during treatment | Elevated baseline LDNs predict primary resistance to ICI monotherapy; LDNs mediate humoral immunosuppression |
| Advanced lung cancer | Shaul et al., FASEB Journal 2020; [52] doi:10.1096/fj.201902467R |
Prospective observational + mechanistic |
64 advanced, 35 early-stage patients, 15 healthy, 13 COPD | Peripheral blood; CyTOF analysis | LDNs highly enriched in advanced disease (median 7.0%); elevated LDNs (>10%) → poor prognosis; unique immune signatures (CD66b⁺/CD10⁻/CXCR4⁺/PDL1⁺ subset) |
|
Lung adenocarcinoma |
Liu et al., Oncotarget 2017; [77] doi:10.18632/oncotarget.18771 |
Prospective observational | 52 advanced lung adenocarcinoma patients, 13 healthy controls | Peripheral blood; flow cytometry | LDNs/HDNs ratio ↑ in patients; ratio correlates with disease progression; positive correlation with CD8⁺ T cells; decreased with treatment |
|
NSCLC |
Mezquita et al., European Journal of Cancer 2021; [99] doi:10.1016/j.ejca.2021.03.011 |
Multicenter prospective | NSCLC patients: 1485 ICI; 173 CT | Peripheral blood; baseline and cycle 2 | Persistently high NLR → poor survival (OS=5 months); immature neutrophils correlate with poor PFS/OS; 12-week death rate 49% |
| NSCLC | Valadez-Cosmes et al., Frontiers in Immunology 2021; [18] doi:10.3389/fimmu.2021.703846 |
Mechanistic (clinical samples) | 26 NSCLC patients, 14 healthy | Peripheral blood; high-dimensional flow cytometry screening | NSCLC: LDNs highly enriched (median 20.4%); novel markers identified: CD36↑, CD41↑, CD61↑, CD226↑; 12 markers downregulated, 41 upregulated |
CD: cluster of differentiation; COPD: chronic obstructive pulmonary disease; CT: chemotherapy; CyTOF: cytometry by time of flight; DFS: disease-free survival; G-CSF: granulocyte colony-stimulating factor; HDNs: high-density neutrophils; HR: hazard ratio; ICI: immune checkpoint inhibitor; iLDNs: immature low-density neutrophils; IL-1β: interleukin-1 beta; LDNs: low-density neutrophils; LMR: lymphocyte-to-monocyte ratio; MDSCs: myeloid-derived suppressor cells; MES: mesenchymal subtype; NES: neutrophil-enriched subtype; NETs: neutrophil extracellular trap; NLR: neutrophil-to-lymphocyte ratio; NSCLC: non-small cell lung cancer; OS: overall survival; PD-1: programmed cell death protein 1; PD-L1: programmed death-ligand 1; PFS: progression-free survival; ROS: reactive oxygen species; TGF-β: transforming growth factor beta; TNBC: triple-negative breast cancer.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



