Submitted:
26 February 2026
Posted:
27 February 2026
You are already at the latest version
Abstract
Model systems that mimic human cardiac structure and function are essential for the development of novel diagnostics and effective treatments for cardiovascular diseases. While non-human vertebrate models, from zebrafish to pig, remain vital to cardiovascular research, the translatability of findings to human patients is often limited. Therefore, animal experiments should be supplemented with human model systems, including human induced pluripotent stem cell-derived cells, 3D engineered constructs, and last but not least, native tissue preparations and isolated primary cardiomyocytes. However, while human myocardium remains the gold standard, human heart tissue – and particularly tissue from control hearts – remains scarce, and its use in research is generally restricted to settings where tissue has been excised from diseased or failing hearts. While it is in principle possible to use tissue from rejected non-failing donor hearts that cannot be transplanted, legal hurdles (for example in Germany) can restrict the use of non-transplanted donor organs in research. Given the challenges associated with accessing and using human tissue in biomedical research, an integrated strategy towards combining non-human vertebrate models, in silico models, and human tissue-derived models is recommended, enhancing the chances of successful research and development, and helping bridge the gap between preclinical and clinical research.
Keywords:
animal models
; vertebrate models
; stem cell models
; human myocardium
; donor tissue
Introduction
The development of novel, affordable, and effective treatments for cardiovascular diseases greatly relies on the use of model systems that mimic human cardiac structure and function. While human heart tissue would be the best model for investigating human cardiovascular function and pathology, its limited availability (which is subject to ethical and legal restrictions) means that non-human models play crucial roles in cardiovascular research and development. However, the extent to which cardiovascular diseases can be recapitulated ex vivo is limited, owing to complex aetiologies, the involvement of extra-cardiac influences on the heart (e.g., circulating hormones, immune cells, and nervous system effects), and additional factors such as age, sex, and comorbidities, as well as the progressive nature of structural and functional pathological remodelling (which are all in turn affected by factors such as environmental cues, lifestyle, and medical treatments). This complexity cannot easily be mimicked in vitro, and it may not be well captured in non-human vertebrate models either. As a result, the translatability of basic science findings to patients has remained limited, and the vast majority of candidate drugs that enter human testing fail, often due to a lack of efficacy or unexpected toxicity in spite of prior promising results in non-human models [1,2,3,4,5,6,7]. Combinations of models that mimic human pathophysiological mechanisms more accurately are therefore needed to improve bio-medical translation [8].
This paper is based on discussions at the 5th Translational Workshop of The German Cardiac Society (DGK) and the German Centre for Cardiovascular Research (DZHK) in Hamburg, Germany, September 2024, dedicated to Models of Human Myocardium in Medical Research. It reviews existing in vivo, ex vivo, in vitro, and in silico models, and discusses the utility and limitations of using human cells and tissue for clinically motivated research, with special consideration of the research landscape in Germany.
Non-Human Vertebrate Models of the Heart for Cardiac Research
Non-human vertebrate models allow cardiovascular researchers to perform experiments that cannot be performed on humans, while facilitating the identification of novel therapeutic and treatment approaches, before these are tested in patients [9,10]. Vertebrate models can be used to mimic complex inter-organ (e.g., metabolic, neurohormonal, or immunological) crosstalk with the heart, while allowing control of factors ranging from genetic background to ageing, comorbidities, and treatments. Here, we review some of the more commonly used vertebrate models in cardiac research (see also Table 1). The advantages and disadvantages of each model, as well as examples of research questions for each, are shown in Figure 1 [11,12,13].
Zebrafish: An Example of Non-Mammalian Model Systems
Despite being evolutionarily distant from primates, non-mammalian animal models such as zebrafish, worms [14,15], flies [16,17], and chick embryos [18,19] have been used to model several human cardiovascular disorders, including congenital heart defects and cardiomyopathies [20,21].
A unique advantage of zebrafish is that their embryos are transparent, allowing for in vivo optical measurements of cardiac structure and function. Additionally, their genome can be manipulated relatively easily, and they possess high regenerative capacities that may provide opportunities to identify novel targets for human cardiac repair, such as following ischaemic injury [21,22]. Zebrafish have a short breeding cycle and high fecundity, which permits rapid generation of models [23].
Crucially, zebrafish have hearts that share electrophysiological properties with human hearts, including conserved fundamentals of excitation–contraction coupling, similar action potential trajectories (both species have a pronounced plateau phase, for example) and comparable major inward and outward current systems, including sodium, calcium, and potassium currents [24]. However, while useful for biochemical assays such as drug screening, zebrafish hearts differ from human hearts in a host of biophysical properties, including chamber anatomy, size, and haemodynamics, meaning that findings cannot easily be translated to humans.
In conclusion, zebrafish are well-suited for live imaging, genetic and drug screening, as well as developmental and cardiac electrophysiological research. Yet, they differ substantially in size, structure, mechanical activity, and genetic make-up from humans, which can complicate translation of findings (Figure 1).
Mice: the Most Widely Used Small Mammalian Model
Mice are the most commonly used animals in biomedical research [25], due in part at least to the availability of standardised recombinatorial lines that allow a level of genetic manipulation unmatched in other vertebrate models [26]. Since the first report of a transgenic mouse [27], the cumulative global production of transgenic founders has reached the millions [28]. Other key advantages of the mouse as a model system are early sexual maturation, short gestational periods, and comparatively short lifespans, allowing for studies over multiple developmental stages of an individual, or even over generations. These features, along with their small size, comparatively low cost of maintenance, and the relative ease of surgical interventions have contributed to the widespread use of mice in cardiovascular research [25,29].
Given the wide range of ready-to-use models that are commercially available, mice are a convenient choice for experimental investigations. Murine anatomy, genetics, and physiology have been well studied, and reference values are well established. Furthermore, mouse models of a wide variety of diseases and cardiac conditions, including cardiomyopathies, heart failure, and developmental defects, have been established [11,30,31,32,33]. However, due to substantial differences in physiological cardiac function (such as heart rate, action potential shape and duration, calcium dynamics, etc; Table 1), as well as in responses to pathological stimuli and therapeutic interventions – including resistance to the development of atrial fibrillation [34] and atherosclerosis [35] – the translatability of findings in mice to human patients remains limited. In addition, one of the most commonly used mouse strains (C57BL/6J) carries a loss-of-function mutation of the gene encoding the mitochondrial transhydrogenase, which makes it resistant to a number of common aetiologies of heart failure, such as pressure-overload induced left or right heart failure, hypertrophic cardiomyopathy, and heart failure with a preserved ejection fraction, induced by high fat diet and the vasopressor N(ω)-nitro-L-arginine methyl ester (L-NAME) [36,37,38,39,40]. ‘Humanised’ mice, which are genetically modified to express functional human genes, have been proposed to extend the utility and translational potential of murine models [41]. In addition, several central repositories for genetic mouse models now contain recently created wild-derived mice that better represent human genetic background diversity (www.mmrrc.org; www.findmice.org; [42,43]).
Thus, the mouse is a valuable model for both genetic and epigenetic manipulation, enabling thorough mechanistic studies of various disease conditions and developmental disorders. Limitations include differences in size, genetics, and electro-mechanical function compared to humans (Figure 1).
Rabbits: The Intermediate Size Mammalian Model
Rabbits (commonly New Zealand white) have been dubbed ‘the largest of the small mammalian models’ in cardiovascular research [44]. While rabbit hearts are 30 to 40 times smaller than human hearts by weight, they are large enough to allow use of scaled-down instrumentation used in human interventions, including surgical and implantable devices [45]. Rabbit hearts resemble human hearts much more closely than mouse hearts in terms of cellular electrophysiology, including the shape of action potentials and arrhythmia wavelength, as well as their response to ischaemia and pharmacological interventions [45,46,47]. In contrast to mice and rats, rabbits also display a positive force–frequency relationship with calcium handling properties that are similar to humans [48,49]. Furthermore, both electrophysiological and mechanical remodelling associated with ageing and disease resemble key changes that occur in humans [50,51].
Disadvantages compared to murine models include the later sexual maturity, longer reproductive cycle, and longer lifespan of rabbits, making multi-generational and ageing studies more challenging. Rabbit housing and maintenance are also more costly. There are relatively few transgenic rabbit models and, in contrast to mice, these do not allow the easy ‘mix-and-match’ recombination that systems such as the established Cre/LoxP or Flp/FRT lines offer for mice. Regrettably, there is no central repository for genetic rabbit models, leading to duplication of efforts in generating models, and at least occasionally to the loss of transgenic model lines maintained by individual labs [52]. Encouragingly, however, genetic manipulation via point mutations and small insertions can work well, and new genetic techniques using TALEN, or CRISPR/Cas9 have simplified the generation of transgenic mammals, including rabbits [53,54].
Overall, the rabbit is a valuable model for studying cardiac electrophysiology and mechanics in health and disease. Limitations include relatively high demands for time and resources, as well as the limited availability of transgenic models (compared to mice), as well as their somewhat unusual immune system (with one variable heavy chain and an unusually complex immunoglobulin A system) (Figure 1).
Pigs: A ‘Human-Sized’ Mammalian Model
Several large vertebrate models are used in cardiovascular research, of which non-human primates have the closest phylogenetic relationship to humans. However, other large vertebrate models including goats, sheep, or pigs are also used.
Like humans, pigs are large, omnivorous, diurnal mammals, and the two species share relevant similarities in terms of anatomy, physiology, and biochemistry, with the pig’s cardiovascular system being considered a good model for humans [22]. Pigs and humans have hearts of similar size and mass, and there is overlap in terms of heart rate, cardiac output, and ejection fraction (Table 1). Accordingly, pigs have been used as models for many cardiovascular diseases, including atherosclerosis, hypercholesterolemia, hypertrophic cardiomyopathy, as well as for the exploration of cardiac regeneration and cell therapy [55,56,57,58,59]. Genetic modification is well established for the pig, including conditional gene targeting [60] and blastocyst complementation [61]. Examples of in vivo genome editing of the porcine heart have been published [62,63]. In view of the high financial and logistic burdens, generation and maintenance of genetically modified pig models is conducted by few labs worldwide [64]. Porcine-expanded potential stem cells have also been differentiated into cardiomyocytes, which may provide a useful platform for preclinical testing of cardiac therapies [65,66]. Given the similarities in size and cardiac anatomy of pigs and humans, pigs are also used for developing and testing cardiovascular devices for diagnosis and intervention [67]. In addition, research involving pigs also holds promise for xenotransplantation, including the potential to grow humanised hearts in pigs, although significant challenges remain [68,69].
Pigs are comparatively slow to reach sexual maturity (5 to 6 months) and have long gestational periods (approximately 16 weeks) and lifespans (up to 20 years), making generational and ageing studies difficult. In addition, while the possibility of using clinical equipment for research on pigs makes them a favoured model for device development and procedure testing, the associated infrastructural demands, such as for operating theatre facilities, raise the threshold for wider use.
Therefore, due to similarities between porcine and human hearts, pigs are a valuable translational model for research into pathogenesis, diagnostics, and drug- or device-based therapy. Limitations include their long reproductive cycle and lifespan, often underestimated phylogenetic distance to humans, high costs, and infrastructural demands (Figure 1).
Challenges of Using Non-Human Vertebrate Models of the Heart
Vertebrate models of the human heart are vital to cardiovascular research, particularly where experiments cannot be performed on humans. However, it is important that researchers consider interspecies differences that may affect the translatability of results, including mitochondrial function and dynamics [70], ion currents [71,72], DNA methylation and gene expression [73], as well as cellular [74] and myocardial architecture [75].
The use of living organisms as models of human disease also requires carefully balancing potential benefits for human health that can be gained against the distress, pain, and/or suffering of animals involved. Scientific research is guided by the 3R principles, referring to ‘replacement’ when possible, ‘reduction’ of the number of animals used, and ‘refinement’ of protocols, interventions, and upkeep to minimise animal discomfort while maximising scientific insight. Implicit in this is a fourth R: responsibility on the part of investigator for the experimental animal [76,77,78]. Computational modelling and simulation can contribute to refined planning of animal experiments, to data integration, and to projection from animal-based data for estimation of effects in humans [79,80].
The use of non-human vertebrate models in pharmaceutical screening suffers from several principal drawbacks. Both false-positive (when therapies appear effective in animals but fail in human trials) and false-negative observations (when therapies that would perform well in humans perform poorly in animal models and are abandoned prematurely) are common [4,13]. This explains the desire for human models, from in vitro use of patient cells to exploration of tissue and organ explants, and on to clinical data mining approaches. The aim here is to improve the accuracy of testing, and to reduce the percentage of false-negative and false-positive conclusions in pre-clinical development, before human trials are commenced. Thus, human wet and dry experimental models are needed to bridge the gap between bench and bedside more effectively.
Human Models for Cardiac Research
Human Induced Pluripotent Stem Cells and Human Engineered Heart Tissue
Cardiomyocytes derived from human induced pluripotent stem cells (hiPSC-CM) provide a sustainable source of cells that can be maintained in cell culture for months. hiPSC can be differentiated into several cardiomyocyte subtypes, including atrial, ventricular, and pacemaker-like cells [81,82]. These cells can be used for basic research in genetics, signalling, biophysics, or regeneration. Genome manipulation, such as introducing mutations, deletions, insertions, knockouts, or knock-ins, enable the creation of disease models, investigation of the consequences of genome alterations and of genetic defect corrections, and the use of reporters and optogenetic actuators to study cell function [83,84,85,86,87].
The unique genetic relationship with their donors provides a potential for the exploration of genetic causes of cardiac disorders at a patient-specific level [88,89,90,91,92]. This has supported therapy selection for individual patients [93]and allowed re-classification of genetic variants of hitherto unknown importance [94].
One possibility to improve the functional depth and robustness of readouts, the maturity of hiPSC-CM, and the similarity to native heart muscle is to engineer 3D cardiac tissues and to incorporate other cell types such as fibroblasts, and smooth muscle, endothelial or immune cells. These 3D models are not only increasingly being used as a platform for gene and cell therapy studies, drug screening or disease modelling [90,95,96,97,98,99,100,101,102], but they are making their way into direct clinical use [103]. Combinations of multiple cell types in tissue-engineered heart muscle emerge as a critical precondition for maturing hiPSC-CM. These models allow for cell–cell interaction studies [104,105], including research in genetically mixed models to determine the contribution of specific cell types to a phenotype, or investigations into genome repair for restoration of cardiac function in disease [106]. Novel tissue constructs, such as assembloids composed of atrial, atrioventricular, and ventricular cardiomyocyte spheroids, allow for the study of complex disorders affecting heart rhythm [107,108]. Combining multiple tissue-engineered model systems furthermore opens the door to organ–organ interaction studies [109].
Key limitations of hiPSC-CM include their metabolic, electrophysiological, and structural immaturity, as the cells generally resemble embryonic or neonatal, rather than mature cardiomyocytes. Given this limitation, maturation strategies for iPSC-CM are actively being explored on multiple fronts, including long-term culture [110], use of different culture media [111], including protocols to foster metabolic maturation [110,111,112,113,114], and mechanical or electrical stimulation [115], incorporation into 3D heart organoids, or combinatorial approaches [116]. Thus far however, the most mature hiPSC-CM remain phenotypically different from adult cardiomyocytes [117]. Another challenge of working with hiPSC-CM is managing variability that occurs from clone to clone, from differentiation to differentiation, between passages, and from laboratory to laboratory – even when nominally using the same cell lines and protocols [118]. These limitations underscore the need for generally agreed-upon quality assessment routines. Stem cell journals and societies have published relevant guidelines [119] and adopted minimal standards that, for example, require the use of isogenic controls when studying the effects of genetic variants and validation of results in several cell lines, sufficient numbers of replicates, and independent differentiation batches.
Human Isolated Primary Cardiomyocytes and Non-Cardiomyocytes
Unlike hiPSC-CM, primary cardiomyocytes are directly isolated from human myocardial tissue, obtained from different regions of the heart. These cells retain core characteristics of mature cardiomyocytes, including their structural, functional, and epigenetic profile [120], while keeping patient-specific phenotypes (e.g., related to age, sex, comorbidities, medical treatments, and the progressive remodelling of cardiac diseases). These native phenotypes make findings more directly translatable. Isolated cardiomyocytes allow for a high degree of experimental control and provide a comparatively inexpensive, convenient pathophysiologically relevant model for evaluating cellular behaviour and morphology [121,122], as well as ion channel function, contraction and electrophysiological characteristics [10]. Human primary cardiomyocytes have traditionally been used both in basic and translational research, particularly in signalling studies and drug efficacy/toxicity testing [10,123,124,125]. More recently, culturing human primary cardiomyocytes without overt dedifferentiation changes has enabled genetic manipulation, which can in turn be used to study the effects of genome alterations, as proof of concept for gene therapies, to investigate the consequences of tachypacing over time, to analyse biomarker release and function, or to utilise genetically encoded biosensors to study cellular function for example [125,126,127,128,129].
However, the functional viability of isolated cells is limited to a relatively short period, typically no more than one day to one week [127,129], before they progressively lose their structural and functional integrity [120,127,130,131,132,133,134]. Cold preservation of mouse hearts and human isolated myocytes can extend the time during which they can be used in functional research without major structural, functional, or transcriptional effects [129,135]. This time window can also be extended by isolating cells on consecutive days from living cardiac tissue slices (discussed in the next section [134]), as live tissue slices can be maintained for several weeks [136]. Whether or not even longer-term cryopreservation of cells, with subsequent recovery of key live cell function, is indeed plausible remains to be elucidated [137]. Although multiple cell types (e.g., cardiomyocytes, fibroblasts, neurons, etc.) can be obtained during the isolation process, so that cell–cell interaction studies are possible in principle, cellular models cannot be used to study long-term therapeutic interventions or to replicate in vivo complexity and systemic physiological connections.
Myocardial Tissue Slices
Myocardial tissue slices are living sections of human heart tissue that are usually cut to a thickness of 250 to 350 µm and kept in culture while mimicking certain physiological conditions. They retain key aspects of the structure (e.g., matrix/cell integration), function (e.g., electrical activity and contractions), and heterocellular signalling of native tissue [122,136,138,139,140]. Remarkably, myocardial tissue slices can maintain electrophysiological behaviour that is close to that of the same tissue-block surface before cutting (as shown in slices from rabbit hearts [140]). They also maintain contractility, gene expression profiles and the response to important pharmacological or hormonal stimuli for up to 3 months [138,141,142]. Similar to hiPSC-CM and 3D models, myocardial slices can be used to explore disease mechanisms and can be matched with patient data [143]. They can also be used to evaluate or identify the therapeutic effects of drugs [144,145], and for the study of cardiac contractility modulation [146] and cardiac toxicity screening [147]. This model can also be used to separate human myocardium from the extracardiac neural innervation that may otherwise confound results in vivo, providing a tissue model with controlled conditions. Direct effects on the myocardium can thus be studied in high detail and with high fidelity. Furthermore, due to the extended viability in culture [138,148], the study of long-term effects, such as of cardiac tissue remodelling, becomes possible. The analysis of the culture medium offers another opportunity, as released metabolic products, cytokines or enzymes, as well as nutrients can be quantified [149].
Limitations of myocardial slices include the tissue damage that occurs during slicing, which directly affects near-surface cell layers. Furthermore, given the transmurally varying alignment of cardiac cells relative to the cutting plane, the functional variability from slice to slice, even if cut from the same tissue block, can be significant. Furthermore, although permitting a high level of control, e.g., over the diastolic load or the composition of the culture medium, the local biophysical (e.g., stress/strain conditions, electrical source-sink relations) and biochemical environments (e.g., nutrient accessibility, hormonal signalling, oxygen delivery) differ from in vivo conditions [149]. Slices are also disconnected from the circulation and nervous system, so that the exploration of inter-organ crosstalk, of effects of the immune system, and of response to systemic signals is restricted.
Despite these limitations, myocardial tissue slices represent one of the closest approximations to native human myocardium available in vitro. For myocardial tissue slices to realise their full translational potential, coordinated efforts towards protocol harmonisation, inter-laboratory reproducibility assessment, and consensus definition of minimal reporting standards will be essential [150].
In Silico Models
Computational models of the human heart span from single-cell to organ-in-chest levels, with a primary focus on electrophysiology and mechanical function. Mechanistic in silico models are versatile and scalable, allowing researchers to explore various physiological and pathological scenarios in a well-controlled and cost-effective way, and without the use of animal or human samples [151,152,153,154,155]. Validated models can predict emergent behaviour on different biological levels of integration, to optimise data interpretation, diagnosis and treatment. In direct iteration with wet-lab research, they can serve as the start (hypothesis-generating) and end (data interpretation) of integrated knowledge development [156], though start and end are arbitrary designations in what is probably more akin to a spiral of knowledge development [157]. In combination with artificial intelligence-based data assessment, mechanistic in silico models are becoming increasingly predictive [158,159] and are beginning to directly aid personalised cardiac treatment [153,160,161,162,163].
Regulatory acceptance of in silico data is increasing. The Comprehensive in vitro Proarrhythmia Assay (CiPA) initiative, aimed to develop a novel, mechanistic, and model-informed assessment of the pro-arrhythmic potential of new drugs, including the use of in silico computer modelling to calculate pro-arrhythmic risk scores (as well as hiSPC-CM), is an example of this development, as it involves regulatory agencies such as the FDA and EMA as stakeholders [164,165]. In April 2025, for example, the FDA announced a phasing out of mandatory animal testing for some drug types, to be replaced with computational models (among other sources of data) [166]. The same month, the National Institutes of Health announced a plan to prioritise human-based research technologies, including computational models that ‘simulate complex human systems, disease pathways, and drug interactions’ [167]. The use of in silico models to assess cardiovascular safety is therefore increasingly embedded within regulatory science and drug development [168]. As computational approaches are poised to inform high-consequence decisions, their credibility must be established within structured risk-based frameworks such as V&V40 (Verification & Validation 40) [169], and aligned with the emerging principles of Good Simulation Practices (GSP) [170]. These frameworks emphasise rigorous verification (ensuring that the computational implementation correctly represents underlying mathematical models), validation (demonstrating that model outputs are consistent with the relevant biological or clinical reality for the defined context of use), and uncertainty quantification (characterising parametric, structural, and methodological uncertainties and their impact on decision-relevant outputs).
Limitations of in silico models include the fact that their utility depends on the quality and extent of input data, both for mechanistic and probabilistic models. And while computational approaches can help translate insight across species, validation using human data remains paramount.
Availability of Human Tissue for Cardiac Research
Human cardiac tissue availability is a bottleneck for basic research. Myocardial tissue cannot be obtained by voluntary live-donation, and while biopsy material or samples from surgical interventions may be collected for certain clinically diagnostic purposes, access to control tissue from non-diseased hearts is not an option, except for post-mortem or (in some countries) non-transplanted donor hearts. The use of human heart tissue for functional and molecular research is therefore generally restricted to settings where it has been excised for medical reasons, such as atrial tissue resected for cardiac bypass surgery, ventricular tissue excised during aortic valve replacement or for ventricular assist device implantation, during septal myectomy in patients with obstructive hypertrophic cardiomyopathy or aortic stenosis, or whole hearts excised from transplant recipients. In all these scenarios, the excised tissue is from diseased hearts, so that ‘control experiments’ compare one medical condition with another (e.g., atrial tissue from patients with atrial fibrillation can be compared to tissue from patients in sinus rhythm – but both groups include patients with cardiac conditions that necessitated cardiac surgery). There is therefore an obvious unmet need for non-failing human heart tissue that can be used as a reference for control investigations. Post-mortem organ donation of non-failing heart tissue for research and teaching is possible, but the suitability of tissue for functional studies is determined by the time taken to access it, as delays of more than a few minutes pose major limitations for studies relying on metabolically competent organs and cells. A realistic source of live non-failing control myocardium would be donor hearts that, for unforeseen reasons, cannot be used for their primary medical purpose – transplantation – but this is not always possible [171].
Use of Non-Failing Human Heart Tissue for Scientific Research: Challenges and Opportunities
Access to non-failing control myocardium in the form of rejected donor hearts is problematic in Germany. While there is a shortage of organ donors in Germany and elsewhere anyway [172,173,174,175,176,177], not all available organs can be transplanted. For example, 96 of the 399 hearts (24%) that would have been available for transplantation in 2023 were not used (Figure 2). Whatever the reasons for non-transplantation, those donor organs may not be utilised for research in Germany. This means that about one in four donor hearts end up as biological waste, despite the fact that a single donor heart could provide enough human cells and control tissue to support dozens, if not hundreds of research projects utilising in vitro models, such as live tissue slices, isolated cells, or cultures. This waste occurs because German transplantation legislation does not include (and hence, does not permit) the use of donor organs for any other than medical purposes. Documentation of donor consent is focused exclusively on tissue and organ use for direct therapeutic purposes, and options in case of non-transplantation, including the potential use of donated tissue in research or education, are not included in these forms. This contrasts with the UK, for example, where the donor or the relatives/ nominated representatives are consulted, using a clearly structured consent/authorisation form. This is completed by NHS staff during consultation and it includes specific tick-box questions covering secondary (non-medical) uses, such as for research, training, or quality assurance. The overall rate of consent for research use of organs found to be unsuitable for transplantation is very high, at 93% for the UK as a whole [178]. The lack of a similar structured approach to consent for secondary uses in Germany, by contrast, means that it is difficult (if not impossible) to even approximate the willingness of patients to consent to their tissue being used for research purposes.
Additional confusion arises around the legal ‘ownership’ of an organ. Once it has been assigned to a recipient by Eurotransplant, for example, ‘ownership’ is formally transferred to the recipient. But – there is no guidance on whose consent (the donor’s or recipient’s) would be required for non-medical use if an organ is eventually not transplanted. Due to this legal ambiguity, donor organs which cannot be transplanted are currently discarded (Figure 2).
This situation is part of the patchwork of regulations relating to organ donation across Europe, where the use of donor tissue is generally determined on a national level (Figure 3), and which may add an additional layer of legal complexity around questions of ownership and consent, as organs are regularly shared between countries during the process of donor–recipient matching. This situation contrasts with the USA, where the use of human tissue in research is governed by federal, state, and local laws, regulations, and policies, including the Department of Health and Human Service’s ‘Common Rule’ and the Health Insurance Portability and Accountability Act [179,180]. In practice, human heart tissue can be obtained for scientific research via tissue banks and networks like the National Cancer Institute-funded Cooperative Human Tissue Network (https://chtn.cancer.gov/), Texas Children’s Hospital’s Heart Center Tissue Bank (https://www.texaschildrens.org/research/find-laboratory/heart-center-tissue-bank), and the International Institute for the Advancement of Medicine (https://iiam.org/researchers/human-tissue-for-research/). These provide non-transplantable organs and tissues (including cardiac tissue) for use in medical research on a national scale.
A key question raised in this context is whether or not it is ethically acceptable (e.g., consistent with donor intentions) for tissue that cannot be used for the originally intended purpose (clinical application) to be used for secondary objectives such as medical research, training, or quality control. It may be assumed that organ donors are altruistic, as they intend their donation to benefit others. Whether this includes the indirect benefits of scientific research into cardiac diseases cannot be assessed retrospectively. However, the high proportion of donors (or their families) who consent to their tissues being used for research purposes when given the option (93% in the UK, 89% in the Netherlands, for example [178,181]) suggests that donor intentions may be broader than simply supporting direct medical use. This is also evident from the very high proportion of patients who are willing to donate (diseased) tissue for use in scientific research, once it has been excised for medical reasons [182,183]. This means that, in view of the uniquely relevant roles of control tissue for research, an alternative question should be posed – namely whether it is ethically acceptable for tissue that cannot be transplanted to be simply discarded, instead of being used in secondary applications.
The legislative situation in Germany regarding organ donation seems unlikely to improve soon [184,185,186,187], as is evident from the response of the German government [188] to a formal parliamentary enquiry on this very topic [189] in 2023. Since the use of organs or tissue, originally intended for transplantation, for research, teaching, or quality control is unaddressed (i.e., neither permitted nor prohibited per se) in Germany, it would seem to be prudent to consider changing the informed consent process, in collaboration with organisations conducting it for organ donation, perhaps to a model similar to that of the UK. This may involve the creation of shared standard operating procedures to ensure that organs which cannot be used for transplantation do not go to waste, as well as the establishment of a coordinated national (or European) procurement network to handle, process, and share tissue for research, to prevent the current waste of uniquely valuable control tissue.
Conclusions
There are multiple model systems for human cardiovascular research, including animal tissue, iPSC-derived cells, primary human tissue and cells, and computational models. All these models have inherent benefits and limitations, related to how well they represent specific aspects of human cardiac structure and function, as well as reproducibility, amenability to conducting controlled investigations, cost, and access, to name but a few. While pronounced inter-species differences can be bridged (at least in part) using theoretical models, human heart tissue remains the gold standard for clinically relevant cardiac research. The scarcity of human heart tissue, in particular from non-failing control hearts, is compounded by the fact that donor tissue that cannot be used for its intended primary medical purpose is not yet systematically available for academic research, training, and quality control, with divergent regulations in individual European countries (Figure 3). The UK provides a good example of how consent for primary and secondary uses of organs can be effectively obtained, with very high rates of consent. Conversely, legal ambiguities, such as in Germany, can hinder the use of precious donor tissue for the advancement of medical insight. This calls, on the one hand, for careful combination of multiple model systems leveraging their respective strengths, and on the other hand, for a broader discussion with all stakeholders about the handling of donated tissue for medical progress in the present (i.e., treatment) and future (i.e., research).
Author Contributions
EARZ and PK wrote the first and the final draft of the manuscript. All authors reviewed the manuscript and approved the final version.
Funding
This paper is based on discussions at the 5th DGK/DZHK Translational Workshop ‘Models of human myocardium in functional medical research: opportunities and challenges’ held in Hamburg, Germany, 25 September 2024, organised by the Commission for Experimental Cardiovascular Medicine of the DGK. The workshop was financially supported by DGK, Novartis AG, und Boehringer Ingelheim International GmbH.
Data Availability Statement
Data sharing not applicable to this article, as no datasets were generated or analysed.
Acknowledgments
This position paper was written on behalf of the German Cardiac Society (Deutsche Gesellschaft für Kardiologie, DGK) and the German Centre for Cardiovascular Research (Deutsches Zentrum für Herz-Kreislauf-Forschung e.V., DZHK). The content of the manuscript reflects author consensus; it is not an official declaration of the views of either DZHK or DGK, and the authors do not speak on behalf of either organisation. Writing and editing support for the manuscript was provided by Dr James O’Reilly.
Conflicts of Interest Statement
KSB: speakers’ honoraria from Novartis. CM: speakers/consulting honoraria from Astra Zeneca, Boehringer Ingelheim, Bristol Myers Squibb, Cytokinetics, Lilly, Novo Nordisk. RK: founder, equity holder and advisor Sequantrix GmbH. BKP: advisor to HeartbeatBio GmbH. TS: equity holder in InVitroSys GmbH. SS: speakers/consulting honoraria from Astra Zeneca, Novartis, Berlin-Chemie, Daiichi Sankyo, Bristol Myers Squibb, Pfizer, Boehringer Ingelheim, Lilly. WHZ: founder of, equity holder in, and advisor to Repairon GmbH and myriamed GmbH. All other authors have no conflicts to declare.
References
- Morehouse, LA; Sugarman, ED; Bourassa, PA; Sand, TM; Zimetti, F; Gao, F; et al. Inhibition of CETP activity by torcetrapib reduces susceptibility to diet-induced atherosclerosis in New Zealand White rabbits. J Lipid Res 2007, 48, 1263–72. [Google Scholar] [CrossRef]
- Barter, PJ; Caulfield, M; Eriksson, M; Grundy, SM; Kastelein, JJP; Komajda, M; et al. Effects of Torcetrapib in Patients at High Risk for Coronary Events. New England Journal of Medicine 2007, 357, 2109–22. [Google Scholar] [CrossRef]
- Harrison, RK. Phase II and phase III failures: 2013–2015. Nat Rev Drug Discov 2016, 15, 817–8. [Google Scholar] [CrossRef]
- Perry, CJ; Lawrence, AJ. Hurdles in Basic Science Translation. Front Pharmacol 2017, 8, 478. [Google Scholar] [CrossRef]
- Austin, CP. Opportunities and challenges in translational science. Clin Transl Sci 2021, 14, 1629–47. [Google Scholar] [CrossRef] [PubMed]
- Sun, D; Gao, W; Hu, H; Zhou, S. Why 90% of clinical drug development fails and how to improve it? Acta Pharm Sin B 2022, 12, 3049–62. [Google Scholar] [CrossRef] [PubMed]
- Ineichen, B V.; Furrer, E; Grüninger, SL; Zürrer, WE; Macleod, MR. Analysis of animal-to-human translation shows that only 5% of animal-tested therapeutic interventions obtain regulatory approval for human applications. PLoS Biol 2024, 22, e3002667. [Google Scholar] [CrossRef] [PubMed]
- Thomas, A; Desai, P; Takahashi, N. Translational research: A patient-centered approach to bridge the valley of death. Cancer Cell 2022, 40, 565–8. [Google Scholar] [CrossRef]
- Giacomotto, J; Ségalat, L. High-throughput screening and small animal models, where are we? Br J Pharmacol 2010, 160, 204–16. [Google Scholar] [CrossRef]
- Odening, KE; Gomez, A-M; Dobrev, D; Fabritz, L; Heinzel, FR; Mangoni, ME; et al. ESC working group on cardiac cellular electrophysiology position paper: relevance, opportunities, and limitations of experimental models for cardiac electrophysiology research. EP Europace 2021, 23, 1795–814. [Google Scholar] [CrossRef]
- Zaragoza, C; Gomez-Guerrero, C; Martin-Ventura, JL; Blanco-Colio, L; Lavin, B; Mallavia, B; et al. Animal models of cardiovascular diseases. J Biomed Biotechnol 2011, 2011, 497841. [Google Scholar] [CrossRef] [PubMed]
- Tsang, HG; Rashdan, NA; Whitelaw, CBA; Corcoran, BM; Summers, KM; MacRae, VE. Large animal models of cardiovascular disease. Cell Biochem Funct 2016, 34, 113–32. [Google Scholar] [CrossRef] [PubMed]
- van der Velden, J; Asselbergs, FW; Bakkers, J; Batkai, S; Bertrand, L; Bezzina, CR; et al. Animal models and animal-free innovations for cardiovascular research: current status and routes to be explored. Consensus document of the ESC Working Group on Myocardial Function and the ESC Working Group on Cellular Biology of the Heart. Cardiovasc Res 2022, 118, 3016–51. [Google Scholar] [CrossRef] [PubMed]
- Ray, AK; Priya, A; Malik, MdZ; Thanaraj, TA; Singh, AK; Mago, P; et al. A bioinformatics approach to elucidate conserved genes and pathways in C. elegans as an animal model for cardiovascular research. Sci Rep 2024, 14, 7471. [Google Scholar] [CrossRef]
- Benian, GM; Epstein, HF. Caenorhabditis elegans Muscle. Circ Res 2011, 109, 1082–95. [Google Scholar] [CrossRef]
- Paternostro, G; Vignola, C; Bartsch, D-U; Omens, JH; McCulloch, AD; Reed, JC. Age-Associated Cardiac Dysfunction in Drosophila melanogaster. Circ Res 2001, 88, 1053–8. [Google Scholar] [CrossRef]
- Feala, JD; Omens, JH; Paternostro, G; McCulloch, AD. Discovering Regulators of the Drosophila Cardiac Hypoxia Response Using Automated Phenotyping Technology. Ann N Y Acad Sci 2008, 1123, 169–77. [Google Scholar] [CrossRef]
- Wittig, JG; Münsterberg, A. The chicken as a model organism to study heart development. Cold Spring Harb Perspect Biol 2020, 12, 1–17. [Google Scholar] [CrossRef]
- Rees, J; Winkler, A; Huettemeister, J; Stengel, L; Spangler, P; Ramesh, G; et al. Rostafuroxin ameliorates cardiac glycoside-induced cardiomyocyte electrolyte imbalances and arrhythmia in ovo. Am J Physiol Heart Circ Physiol 2026. [Google Scholar] [CrossRef]
- MacRae, CA; Peterson, RT. Zebrafish as tools for drug discovery. Nat Rev Drug Discov 2015, 14, 721–31. [Google Scholar] [CrossRef]
- Asnani, A; Peterson, RT. The zebrafish as a tool to identify novel therapies for human cardiovascular disease. Dis Model Mech 2014, 7, 763–7. [Google Scholar] [CrossRef] [PubMed]
- Cesarovic, N; Lipski, M; Falk, V; Emmert, MY. Animals in cardiovascular research. Eur Heart J 2020, 41, 200–3. [Google Scholar] [CrossRef] [PubMed]
- Yang, D; Jian, Z; Tang, C; Chen, Z; Zhou, Z; Zheng, L; et al. Zebrafish Congenital Heart Disease Models: Opportunities and Challenges. Int J Mol Sci 2024, 25, 5943. [Google Scholar] [CrossRef] [PubMed]
- Vornanen, M; Hassinen, M. Zebrafish heart as a model for human cardiac electrophysiology. Channels 2016, 10, 101–10. [Google Scholar] [CrossRef]
- Hickman, DL; Johnson, J; Vemulapalli, TH; Crisler, JR; Shepherd, R. Commonly Used Animal Models. In Principles of Animal Research for Graduate and Undergraduate Students; Elsevier Inc., 2017; pp. 117–75. [Google Scholar] [CrossRef]
- Gurumurthy, CB; Kent Lloyd, KC. Generating mouse models for biomedical research: Technological advances. Dis Model Mech 2019, 12, dmm029462. [Google Scholar] [CrossRef]
- Gordon, JW; Scangos, GA; Plotkin, DJ; Barbosa, JA; Ruddle, FH. Genetic transformation of mouse embryos by microinjection of purified DNA. Proceedings of the National Academy of Sciences 1980, 77, 7380–4. [Google Scholar] [CrossRef]
- Hanahan, D; Wagner, EF; Palmiter, RD. The origins of oncomice: a history of the first transgenic mice genetically engineered to develop cancer. Genes Dev 2007, 21, 2258–70. [Google Scholar] [CrossRef]
- Justice, MJ; Dhillon, P. Using the mouse to model human disease: increasing validity and reproducibility. Dis Model Mech 2016, 9, 101–3. [Google Scholar] [CrossRef]
- Chowdhury, R; Ashraf, H; Melanson, M; Tanada, Y; Nguyen, M; Silberbach, M; et al. Mouse Model of Human Congenital Heart Disease: Progressive Atrioventricular Block Induced by a Heterozygous Nkx2-5 Homeodomain Missense Mutation. Circ Arrhythm Electrophysiol 2015, 8, 1255–64. [Google Scholar] [CrossRef]
- Lindsey, ML; Brunt, KR; Kirk, JA; Kleinbongard, P; Calvert, JW; de Castro Brás, LE; et al. Guidelines for in vivo mouse models of myocardial infarction. American Journal of Physiology-Heart and Circulatory Physiology 2021, 321, H1056–73. [Google Scholar] [CrossRef]
- Rao, KS; Kameswaran, V; Bruneau, BG. Modeling congenital heart disease: lessons from mice, hPSC-based models, and organoids. Genes Dev 2022, 36, 652–63. [Google Scholar] [CrossRef]
- Salerno, N; Scalise, M; Marino, F; Filardo, A; Chiefalo, A; Panuccio, G; et al. A Mouse Model of Dilated Cardiomyopathy Produced by Isoproterenol Acute Exposure Followed by 5-Fluorouracil Administration. J Cardiovasc Dev Dis 2023, 10, 225. [Google Scholar] [CrossRef] [PubMed]
- Fu, F; Pietropaolo, M; Cui, L; Pandit, S; Li, W; Tarnavski, O; et al. Lack of authentic atrial fibrillation in commonly used murine atrial fibrillation models. PLoS One 2022, 17, e0256512. [Google Scholar] [CrossRef] [PubMed]
- Emini Veseli, B; Perrotta, P; De Meyer, GRA; Roth, L; Van der Donckt, C; Martinet, W; et al. Animal models of atherosclerosis. Eur J Pharmacol 2017, 816, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Nickel, AG; von Hardenberg, A; Hohl, M; Löffler, JR; Kohlhaas, M; Becker, J; et al. Reversal of Mitochondrial Transhydrogenase Causes Oxidative Stress in Heart Failure. Cell Metab 2015, 22, 472–84. [Google Scholar] [CrossRef]
- Schiattarella, GG; Altamirano, F; Tong, D; French, KM; Villalobos, E; Kim, SY; et al. Nitrosative stress drives heart failure with preserved ejection fraction. Nature 2019, 568, 351–6. [Google Scholar] [CrossRef]
- Müller, M; Bischof, C; Kapries, T; Wollnitza, S; Liechty, C; Geißen, S; et al. Right Heart Failure in Mice Upon Pressure Overload Is Promoted by Mitochondrial Oxidative Stress. JACC Basic Transl Sci 2022, 7, 658–77. [Google Scholar] [CrossRef]
- Kohlhaas, M; Sequeira, V; Parikh, S; Dietl, A; Richter, O; Bay, J; et al. Mitochondrial reactive oxygen species cause arrhythmias in hypertrophic cardiomyopathy. BioRxiv 2024, 2024, 10.02.616214. [Google Scholar] [CrossRef]
- Pepin, ME; Konrad, PJM; Nazir, S; Bazgir, F; Maack, C; Nickel, A; et al. Mitochondrial NNT Promotes Diastolic Dysfunction in Cardiometabolic HFpEF. Circ Res 2025, 136, 1564–78. [Google Scholar] [CrossRef]
- Rosshart, SP; Herz, J; Vassallo, BG; Hunter, A; Wall, MK; Badger, JH; et al. Laboratory mice born to wild mice have natural microbiota and model human immune responses. Science (1979) 2019, 365. [Google Scholar] [CrossRef]
- Eppig, JT; Motenko, H; Richardson, JE; Richards-Smith, B; Smith, CL. The International Mouse Strain Resource (IMSR): cataloging worldwide mouse and ES cell line resources. Mammalian Genome 2015, 26, 448–55. [Google Scholar] [CrossRef] [PubMed]
- Donahue, LR; Hrabe de Angelis, M; Hagn, M; Franklin, C; Lloyd, KCK; Magnuson, T; et al. Centralized mouse repositories. Mammalian Genome 2012, 23, 559–71. [Google Scholar] [CrossRef] [PubMed]
- Odening, KE; Kohl, P. Follow the white rabbit: Experimental and computational models of the rabbit heart provide insights into cardiac (patho-) physiology. Prog Biophys Mol Biol 2016, 121, 75–6. [Google Scholar] [CrossRef] [PubMed]
- Pogwizd, SM; Bers, DM. Rabbit models of heart disease. Drug Discov Today Dis Models 2008, 5, 185–93. [Google Scholar] [CrossRef] [PubMed]
- Panfilov, A V. Is heart size a factor in ventricular fibrillation? Or how close are rabbit and human hearts? Heart Rhythm 2006, 3, 862–4. [Google Scholar] [CrossRef]
- Odening, KE; Baczko, I; Brunner, M. Animals in cardiovascular research: Important role of rabbit models in cardiac electrophysiology. Eur Heart J 2020, 41, 2036. [Google Scholar] [CrossRef]
- Endoh, M. Force–frequency relationship in intact mammalian ventricular myocardium: physiological and pathophysiological relevance. Eur J Pharmacol 2004, 500, 73–86. [Google Scholar] [CrossRef]
- Milani-Nejad, N; Janssen, PML. Small and large animal models in cardiac contraction research: Advantages and disadvantages. Pharmacol Ther 2014, 141, 235–49. [Google Scholar] [CrossRef]
- Alpert, NR; Hasenfuss, G; Mulieri, LA; Blanchard, EM; Leavitt, BJ; Ittleman, F. The reorganization of the human and rabbit heart in response to haemodynamic overload. Eur Heart J 1992, 13, 9–16. [Google Scholar] [CrossRef]
- Cooper, LL; Odening, KE; Hwang, M-S; Chaves, L; Schofield, L; Taylor, CA; et al. Electromechanical and structural alterations in the aging rabbit heart and aorta. American Journal of Physiology-Heart and Circulatory Physiology 2012, 302, H1625–35. [Google Scholar] [CrossRef]
- Hornyik, T; Rieder, M; Castiglione, A; Major, P; Baczko, I; Brunner, M; et al. Transgenic rabbit models for cardiac disease research. Br J Pharmacol 2022, 179, 938–57. [Google Scholar] [CrossRef]
- Liu, Z; Chen, M; Chen, S; Deng, J; Song, Y; Lai, L; et al. Highly efficient RNA-guided base editing in rabbit. Nat Commun 2018, 9, 2717. [Google Scholar] [CrossRef] [PubMed]
- Yang, D; Xu, J; Chen, YE. Generation of Rabbit Models by Gene Editing Nucleases. Methods in Molecular Biology 2019, 1874, 327–45. [Google Scholar] [CrossRef]
- Davis, BT; Wang, XJ; Rohret, JA; Struzynski, JT; Merricks, EP; Bellinger, DA; et al. Targeted disruption of LDLR causes hypercholesterolemia and atherosclerosis in Yucatan miniature pigs. PLoS One 2014, 9, e93457. [Google Scholar] [CrossRef] [PubMed]
- Huang, L; Hua, Z; Xiao, H; Cheng, Y; Xu, K; Gao, Q; et al. CRISPR/Cas9-mediated ApoE-/-and LDLR-/-double gene knockout in pigs elevates serum LDL-C and TC levels. Oncotarget 2017, 8, 37751–60. [Google Scholar] [CrossRef] [PubMed]
- Montag, J; Petersen, B; Flögel, AK; Becker, E; Lucas-Hahn, A; Cost, GJ; et al. Successful knock-in of Hypertrophic Cardiomyopathy-mutation R723G into the MYH7 gene mimics HCM pathology in pigs. Sci Rep 2018, 8, 4786. [Google Scholar] [CrossRef]
- Sridharan, D; Pracha, N; Rana, SJ; Ahmed, S; Dewani, AJ; Alvi, SB; et al. Preclinical Large Animal Porcine Models for Cardiac Regeneration and Its Clinical Translation: Role of hiPSC-Derived Cardiomyocytes. Cells 2023, 12, 1090. [Google Scholar] [CrossRef]
- Liu, S; Li, K; Florencio, LW; Tang, L; Heallen, TR; Leach, JP; et al. Gene therapy knockdown of Hippo signaling induces cardiomyocyte renewal in pigs after myocardial infarction. Sci Transl Med 2021, 13, eabd6892. [Google Scholar] [CrossRef]
- Fischer, K; Schnieke, A. How genome editing changed the world of large animal research. Front Genome Ed 2023, 5, 1272687. [Google Scholar] [CrossRef]
- Barlabé, P; Aranguren, XL; Coppiello, G. Blastocyst complementation: current progress and future directions in xenogeneic organogenesis. Stem Cell Res Ther 2025, 16, 321. [Google Scholar] [CrossRef]
- Moretti, A; Fonteyne, L; Giesert, F; Hoppmann, P; Meier, AB; Bozoglu, T; et al. Somatic gene editing ameliorates skeletal and cardiac muscle failure in pig and human models of Duchenne muscular dystrophy. Nat Med 2020, 26, 207–14. [Google Scholar] [CrossRef] [PubMed]
- Rieblinger, B; Sid, H; Duda, D; Bozoglu, T; Klinger, R; Schlickenrieder, A; et al. Cas9-expressing chickens and pigs as resources for genome editing in livestock. Proceedings of the National Academy of Sciences 2021, 118, e2022562118. [Google Scholar] [CrossRef] [PubMed]
- Prather, RS; Lorson, M; Ross, JW; Whyte, JJ; Walters, E. Genetically engineered pig models for human diseases. Annu Rev Anim Biosci 2013, 1, 203–19. [Google Scholar] [CrossRef] [PubMed]
- Gao, X; Nowak-Imialek, M; Chen, X; Chen, D; Herrmann, D; Ruan, D; et al. Establishment of porcine and human expanded potential stem cells. Nat Cell Biol 2019, 21, 687–99. [Google Scholar] [CrossRef]
- Rawat, H; Kornherr, J; Zawada, D; Bakhshiyeva, S; Kupatt, C; Laugwitz, KL; et al. Recapitulating porcine cardiac development in vitro: from expanded potential stem cell to embryo culture models. Front Cell Dev Biol 2023, 11, 1111684. [Google Scholar] [CrossRef]
- Miller, JR; Henn, MC; Lancaster, TS; Lawrance, CP; Schuessler, RB; Shepard, M; et al. Pulmonary Valve Replacement With Small Intestine Submucosa-Extracellular Matrix in a Porcine Model. World J Pediatr Congenit Heart Surg 2016, 7, 475–83. [Google Scholar] [CrossRef]
- Cooper, DKC; Cozzi, E. Clinical Pig Heart Xenotransplantation—Where Do We Go From Here? Transplant International 2024, 37, 12592. [Google Scholar] [CrossRef]
- Garry, DJ; Garry, MG; Nakauchi, H; Masaki, H; Sachs, DH; Weiner, JI; et al. Allogeneic, Xenogeneic, and Exogenic Hearts for Transplantation. Methodist Debakey Cardiovasc J 2025, 21, 92–9. [Google Scholar] [CrossRef]
- Alibrandi, L; Lionetti, V. Interspecies differences in mitochondria: Implications for cardiac and vascular translational research. Vascul Pharmacol 2025, 159, 107476. [Google Scholar] [CrossRef]
- Horváth, B; Hézső, T; Szentandrássy, N; Kistamás, K; Árpádffy-Lovas, T; Varga, R; et al. Late sodium current in human, canine and guinea pig ventricular myocardium. J Mol Cell Cardiol 2020, 139, 14–23. [Google Scholar] [CrossRef]
- Linz, KW; Meyer, R. Profile and kinetics of L-type calcium current during the cardiac ventricular action potential compared in guinea-pigs, rats and rabbits. Pflügers Archiv 2000, 439, 588–99. [Google Scholar] [CrossRef] [PubMed]
- Pai, AA; Bell, JT; Marioni, JC; Pritchard, JK; Gilad, Y. A Genome-Wide Study of DNA Methylation Patterns and Gene Expression Levels in Multiple Human and Chimpanzee Tissues. PLoS Genet 2011, 7, e1001316. [Google Scholar] [CrossRef] [PubMed]
- Greiner, J; Sonak, F; Jones, WD; Madl, J; Ryeng, KA; Efimov, IR; et al. Architecture of the cardiac transverse-axial tubular system across different mammalian species. BioRxiv 2026, 2026.01, 10.698376. [Google Scholar] [CrossRef]
- Mulbjerg, H; Ringgaard, S; Agger, P. Diffusion tensor imaging reveals myocardial architectural differences between porcine and primate hearts with potential implications for cardiac xenotransplantation. Sci Rep 2025, 15, 28696. [Google Scholar] [CrossRef]
- Lee, KH; Lee, DW; Kang, BC. The ‘R’ principles in laboratory animal experiments. Lab Anim Res 2020, 36, 45. [Google Scholar] [CrossRef]
- Tannenbaum, J; Bennett, BT. Russell and Burch’s 3Rs then and now: the need for clarity in definition and purpose. J Am Assoc Lab Anim Sci 2015, 54, 120–32. [Google Scholar]
- Hubrecht, RC; Carter, E. The 3Rs and humane experimental technique: Implementing change. Animals 2019, 9, 754. [Google Scholar] [CrossRef]
- Viceconti, M; Emili, L; Afshari, P; Courcelles, E; Curreli, C; Famaey, N; et al. Possible Contexts of Use for In Silico Trials Methodologies: A Consensus-Based Review. IEEE J Biomed Health Inform 2021, 25, 3977–82. [Google Scholar] [CrossRef]
- Morotti, S; Liu, C; Hegyi, B; Ni, H; Fogli Iseppe, A; Wang, L; et al. Quantitative cross-species translators of cardiac myocyte electrophysiology: Model training, experimental validation, and applications. Sci Adv 2021, 7, eabg0927. [Google Scholar] [CrossRef]
- Cyganek, L; Tiburcy, M; Sekeres, K; Gerstenberg, K; Bohnenberger, H; Lenz, C; et al. Deep phenotyping of human induced pluripotent stem cell-derived atrial and ventricular cardiomyocytes. JCI Insight 2018, 3, e99941. [Google Scholar] [CrossRef]
- Protze, SI; Liu, J; Nussinovitch, U; Ohana, L; Backx, PH; Gepstein, L; et al. Sinoatrial node cardiomyocytes derived from human pluripotent cells function as a biological pacemaker. Nat Biotechnol 2017, 35, 56–68. [Google Scholar] [CrossRef] [PubMed]
- Wang, G; McCain, ML; Yang, L; He, A; Pasqualini, FS; Agarwal, A; et al. Modeling the mitochondrial cardiomyopathy of Barth syndrome with induced pluripotent stem cell and heart-on-chip technologies. Nat Med 2014, 20, 616–23. [Google Scholar] [CrossRef] [PubMed]
- Kime, C; Mandegar, MA; Srivastava, D; Yamanaka, S; Conklin, BR; Rand, TA. Efficient CRISPR/Cas9-Based Genome Engineering in Human Pluripotent Stem Cells. Curr Protoc Hum Genet 2016, 88, 21.4.1–21.4.23. [Google Scholar] [CrossRef] [PubMed]
- Bengel, P; Dybkova, N; Tirilomis, P; Ahmad, S; Hartmann, N; A. Mohamed, B; et al. Detrimental proarrhythmogenic interaction of Ca2+/calmodulin-dependent protein kinase II and NaV1.8 in heart failure. Nat Commun 2021, 12, 6586. [Google Scholar] [CrossRef]
- Stüdemann, T; Rössinger, J; Manthey, C; Geertz, B; Srikantharajah, R; von Bibra, C; et al. Contractile Force of Transplanted Cardiomyocytes Actively Supports Heart Function After Injury. Circulation 2022, 146, 1159–69. [Google Scholar] [CrossRef]
- Haertter, D; Hauke, L; Driehorst, T; Nishi, K; Zimmermann, W-H; Schmidt, CF. Sarcomere dynamic instability and stochastic heterogeneity drive robust cardiomyocyte contraction. BioRxiv 2024, 2024, 05.28.596183. [Google Scholar] [CrossRef]
- Moretti, A; Bellin, M; Welling, A; Jung, CB; Lam, JT; Bott-Flügel, L; et al. Patient-Specific Induced Pluripotent Stem-Cell Models for Long-QT Syndrome. New England Journal of Medicine 2010, 363, 1397–409. [Google Scholar] [CrossRef]
- Itzhaki, I; Maizels, L; Huber, I; Zwi-Dantsis, L; Caspi, O; Winterstern, A; et al. Modelling the long QT syndrome with induced pluripotent stem cells. Nature 2011, 471, 225–9. [Google Scholar] [CrossRef]
- Streckfuss-Bömeke, K; Tiburcy, M; Fomin, A; Luo, X; Li, W; Fischer, C; et al. Severe DCM phenotype of patient harboring RBM20 mutation S635A can be modeled by patient-specific induced pluripotent stem cell-derived cardiomyocytes. J Mol Cell Cardiol 2017, 113, 9–21. [Google Scholar] [CrossRef]
- Cuello, F; Knaust, AE; Saleem, U; Loos, M; Raabe, J; Mosqueira, D; et al. Impairment of the ER/mitochondria compartment in human cardiomyocytes with PLN p.Arg14del mutation. EMBO Mol Med 2021, 13. [Google Scholar] [CrossRef]
- Wu, X; Swanson, K; Yildirim, Z; Liu, W; Liao, R; Wu, JC. Clinical trials in-a-dish for cardiovascular medicine. Eur Heart J 2024, 45, 4275–90. [Google Scholar] [CrossRef] [PubMed]
- Prondzynski, M; Lemoine, MD; Zech, AT; Horváth, A; Di Mauro, V; Koivumäki, JT; et al. Disease modeling of a mutation in α-actinin 2 guides clinical therapy in hypertrophic cardiomyopathy. EMBO Mol Med 2019, 11, e11115. [Google Scholar] [CrossRef]
- Ma, N; Zhang, JZ; Itzhaki, I; Zhang, SL; Chen, H; Haddad, F; et al. Determining the Pathogenicity of a Genomic Variant of Uncertain Significance Using CRISPR/Cas9 and Human-Induced Pluripotent Stem Cells. Circulation 2018, 138, 2666–81. [Google Scholar] [CrossRef] [PubMed]
- Hinson, JT; Chopra, A; Nafissi, N; Polacheck, WJ; Benson, CC; Swist, S; et al. Titin mutations in iPS cells define sarcomere insufficiency as a cause of dilated cardiomyopathy. Science (1979) 2015, 349, 982–6. [Google Scholar] [CrossRef] [PubMed]
- Mannhardt, I; Breckwoldt, K; Letuffe-Brenière, D; Schaaf, S; Schulz, H; Neuber, C; et al. Human Engineered Heart Tissue: Analysis of Contractile Force. Stem Cell Reports 2016, 7, 29–42. [Google Scholar] [CrossRef]
- Kyrychenko, V; Kyrychenko, S; Tiburcy, M; Shelton, JM; Long, C; Schneider, JW; et al. Functional correction of dystrophin actin binding domain mutations by genome editing. JCI Insight 2017, 2, e95918. [Google Scholar] [CrossRef]
- Borchert, T; Hübscher, D; Guessoum, CI; Lam, T-DD; Ghadri, JR; Schellinger, IN; et al. Catecholamine-Dependent β-Adrenergic Signaling in a Pluripotent Stem Cell Model of Takotsubo Cardiomyopathy. J Am Coll Cardiol 2017, 70, 975–91. [Google Scholar] [CrossRef]
- Saleem, U; van Meer, BJ; Katili, PA; Mohd Yusof, NAN; Mannhardt, I; Garcia, AK; et al. Blinded, Multicenter Evaluation of Drug-induced Changes in Contractility Using Human-induced Pluripotent Stem Cell-derived Cardiomyocytes. Toxicological Sciences 2020, 176, 103–23. [Google Scholar] [CrossRef]
- Fomin, A; Gärtner, A; Cyganek, L; Tiburcy, M; Tuleta, I; Wellers, L; et al. Truncated titin proteins and titin haploinsufficiency are targets for functional recovery in human cardiomyopathy due to TTN mutations. Sci Transl Med 2021, 13, eabd3079. [Google Scholar] [CrossRef]
- Kyriakopoulou, E; Versteeg, D; de Ruiter, H; Perini, I; Seibertz, F; Döring, Y; et al. Therapeutic efficacy of AAV-mediated restoration of PKP2 in arrhythmogenic cardiomyopathy. Nature Cardiovascular Research 2023, 2, 1262–76. [Google Scholar] [CrossRef]
- Greer-Short, A; Greenwood, A; Leon, EC; Qureshi, TN; von Kraut, K; Wong, J; et al. AAV9-mediated MYBPC3 gene therapy with optimized expression cassette enhances cardiac function and survival in MYBPC3 cardiomyopathy models. Nat Commun 2025, 16, 2196. [Google Scholar] [CrossRef] [PubMed]
- Jebran, AF; Seidler, T; Tiburcy, M; Daskalaki, M; Kutschka, I; Fujita, B; et al. Engineered heart muscle allografts for heart repair in primates and humans. Nature 2025, 639, 503–11. [Google Scholar] [CrossRef] [PubMed]
- Tiburcy, M; Hudson, JE; Balfanz, P; Schlick, S; Meyer, T; Liao, MLC; et al. Defined engineered human myocardium with advanced maturation for applications in heart failure modeling and repair. Circulation 2017, 135, 1832–47. [Google Scholar] [CrossRef] [PubMed]
- Landau, S; Zhao, Y; Hamidzada, H; Kent, GM; Okhovatian, S; Lu, RXZ; et al. Primitive macrophages enable long-term vascularization of human heart-on-a-chip platforms. Cell Stem Cell 2024, 31, 1222–1238.e10. [Google Scholar] [CrossRef]
- Long, C; Li, H; Tiburcy, M; Rodriguez-Caycedo, C; Kyrychenko, V; Zhou, H; et al. Correction of diverse muscular dystrophy mutations in human engineered heart muscle by single-site genome editing. Sci Adv 2018, 4, eaap9004. [Google Scholar] [CrossRef]
- Mallapaty, S. Mini hearts, lungs and livers made in lab now grow their own blood vessels. Nature 2025, 643, 892–892. [Google Scholar] [CrossRef]
- Li, J; Wiesinger, A; Fokkert, L; Bakker, P; de Vries, DK; Tijsen, AJ; et al. Modeling the atrioventricular conduction axis using human pluripotent stem cell-derived cardiac assembloids. Cell Stem Cell 2024, 31, 1667–1684.e6. [Google Scholar] [CrossRef]
- Schneider, LV; Guobin, B; Methi, A; Jensen, O; Schmoll, KA; Setya, MG; et al. Bioengineering of a human innervated cardiac muscle model. BioRxiv 2023, 08.18.552653. [Google Scholar] [CrossRef]
- Emanuelli, G; Zoccarato, A; Reumiller, CM; Papadopoulos, A; Chong, M; Rebs, S; et al. A roadmap for the characterization of energy metabolism in human cardiomyocytes derived from induced pluripotent stem cells. J Mol Cell Cardiol 2022, 164, 136–47. [Google Scholar] [CrossRef]
- Feyen, DAM; McKeithan, WL; Bruyneel, AAN; Spiering, S; Hörmann, L; Ulmer, B; et al. Metabolic Maturation Media Improve Physiological Function of Human iPSC-Derived Cardiomyocytes. Cell Rep 2020, 32, 107925. [Google Scholar] [CrossRef]
- Li, W; Luo, X; Strano, A; Arun, S; Gamm, O; Poetsch, MS; et al. Comprehensive promotion of iPSC-CM maturation by integrating metabolic medium with nanopatterning and electrostimulation. Nat Commun 2025, 16, 2785. [Google Scholar] [CrossRef] [PubMed]
- Wickramasinghe, NM; Sachs, D; Shewale, B; Gonzalez, DM; Dhanan-Krishnan, P; Torre, D; et al. PPARdelta activation induces metabolic and contractile maturation of human pluripotent stem cell-derived cardiomyocytes. Cell Stem Cell 2022, 29, 559–576.e7. [Google Scholar] [CrossRef] [PubMed]
- Rebs, S; Eberl, H; Wagensohner, N; Dybkova, N; Unsöld, JK; Dudek, J; et al. Enhanced iPSC Cardiomyocyte Maturation via Combined 3D-Culture and Metabolic Cues. BioRxiv 2025, 2025, 10.29.684362. [Google Scholar] [CrossRef]
- Hirt, MN; Boeddinghaus, J; Mitchell, A; Schaaf, S; Börnchen, C; Müller, C; et al. Functional improvement and maturation of rat and human engineered heart tissue by chronic electrical stimulation. J Mol Cell Cardiol 2014, 74, 151–61. [Google Scholar] [CrossRef]
- Tan, SH; Ye, L. Maturation of Pluripotent Stem Cell-Derived Cardiomyocytes: a Critical Step for Drug Development and Cell Therapy. J Cardiovasc Transl Res 2018, 11, 375–92. [Google Scholar] [CrossRef]
- Yang, H; Yang, Y; Kiskin, FN; Shen, M; Zhang, JZ. Recent advances in regulating the proliferation or maturation of human-induced pluripotent stem cell-derived cardiomyocytes. Stem Cell Res Ther 2023, 14, 228. [Google Scholar] [CrossRef]
- Thomas, D; Cunningham, NJ; Shenoy, S; Wu, JC. Human-induced pluripotent stem cells in cardiovascular research: current approaches in cardiac differentiation, maturation strategies, and scalable production. Cardiovasc Res 2022, 118, 20–36. [Google Scholar] [CrossRef]
- Selfa Aspiroz, L; Mennecozzi, M; Batlle, L; Corneo, B; Healy, L; Kotter, M; et al. Promoting the adoption of best practices and standards to enhance quality and reproducibility of stem cell research. Stem Cell Reports 2025, 20, 102531. [Google Scholar] [CrossRef]
- Zhou, B; Shi, X; Tang, X; Zhao, Q; Wang, L; Yao, F; et al. Functional isolation, culture and cryopreservation of adult human primary cardiomyocytes. Signal Transduct Target Ther 2022, 7, 254. [Google Scholar] [CrossRef]
- Oh, JG; Kho, C; Hajjar, RJ; Ishikawa, K. Experimental models of cardiac physiology and pathology. Heart Fail Rev 2019, 24, 601–15. [Google Scholar] [CrossRef]
- Pitoulis, FG; Watson, SA; Perbellini, F; Terracciano, CM. Myocardial slices come to age: an intermediate complexity in vitro cardiac model for translational research. Cardiovasc Res 2020, 116, 1275–87. [Google Scholar] [CrossRef]
- Molina, CE; Abu-Taha, IH; Wang, Q; Roselló-Díez, E; Kamler, M; Nattel, S; et al. Profibrotic, electrical, and calcium-handling remodeling of the atria in heart failure patients with and without atrial fibrillation. Front Physiol 2018, 9, 1383. [Google Scholar] [CrossRef]
- Ahmad, S; Tirilomis, P; Pabel, S; Dybkova, N; Hartmann, N; Molina, CE; et al. The functional consequences of sodium channel NaV1.8 in human left ventricular hypertrophy. ESC Heart Fail 2019, 6, 154–63. [Google Scholar] [CrossRef] [PubMed]
- Grammatika Pavlidou, N; Dobrev, S; Beneke, K; Reinhardt, F; Pecha, S; Jacquet, E; et al. Phosphodiesterase 8 governs cAMP/PKA-dependent reduction of L-type calcium current in human atrial fibrillation: a novel arrhythmogenic mechanism. Eur Heart J 2023, 44, 2483–94. [Google Scholar] [CrossRef] [PubMed]
- Berisha, F; Götz, KR; Wegener, JW; Brandenburg, S; Subramanian, H; Molina, CE; et al. CAMP Imaging at Ryanodine Receptors Reveals β2-Adrenoceptor Driven Arrhythmias. Circ Res 2021, 129, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Beneke, K; Molina, CE. Live Cell Imaging of Cyclic Nucleotides in Human Cardiomyocytes. Methods Mol Biol 2022, 2483, 195–204. [Google Scholar] [CrossRef]
- Pabel, S; Knierim, M; Stehle, T; Alebrand, F; Paulus, M; Sieme, M; et al. Effects of Atrial Fibrillation on the Human Ventricle. Circ Res 2022, 130, 994–1010. [Google Scholar] [CrossRef]
- Aceituno, C; Revuelta, D; Jiménez-Sábado, V; Ginel, A; Molina, CE; Hove-Madsen, L. Impact of Overnight Storage of Human Atrial Myocytes on Intracellular Calcium Homeostasis and Electrophysiological Utility. Biomolecules 2024, 14, 1415. [Google Scholar] [CrossRef]
- Mitcheson, JS; Hancox, JC; Levi, AJ. Action potentials, ion channel currents and transverse tubule density in adult rabbit ventricular myocytes maintained for 6 days in cell culture. Pflugers Arch 1996, 431, 814–27. [Google Scholar] [CrossRef]
- Banyasz, T; Lozinskiy, I; Payne, CE; Edelmann, S; Norton, B; Chen, B; et al. Transformation of adult rat cardiac myocytes in primary culture. Exp Physiol 2008, 93, 370–82. [Google Scholar] [CrossRef]
- Guo, GR; Chen, L; Rao, M; Chen, K; Song, JP; Hu, SS. A modified method for isolation of human cardiomyocytes to model cardiac diseases. J Transl Med 2018, 16, 288. [Google Scholar] [CrossRef] [PubMed]
- Seidel, T; Fiegle, DJ; Baur, TJ; Ritzer, A; Nay, S; Heim, C; et al. Glucocorticoids preserve the t-tubular system in ventricular cardiomyocytes by upregulation of autophagic flux. Basic Res Cardiol 2019, 114. [Google Scholar] [CrossRef] [PubMed]
- Greiner, J; Schiatti, T; Kaltenbacher, W; Dente, M; Semenjakin, A; Kok, T; et al. Consecutive-Day Ventricular and Atrial Cardiomyocyte Isolations from the Same Heart: Shifting the Cost–Benefit Balance of Cardiac Primary Cell Research. Cells 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Pfeilschifter, B; Martinez-Vilchez, A; Iqbal, Z; Potue, P; Fiegle, DJ; Morhenn, K; et al. Cold storage of mouse hearts prior to cardiomyocyte isolation preserves electromechanical function, microstructure, and gene expression for 24 h. Basic Res Cardiol 2025, 120, 1055–74. [Google Scholar] [CrossRef]
- Brandenburger, M; Wenzel, J; Bogdan, R; Richardt, D; Nguemo, F; Reppel, M; et al. Organotypic slice culture from human adult ventricular myocardium. Cardiovasc Res 2012, 93, 50–9. [Google Scholar] [CrossRef]
- Wang, Z; Tong, C; Xu, M; Feng, S; Dong, M; Rao, R; et al. Optimized methods for the functional cryopreservation of adult human primary cardiomyocytes. Cryobiology 2026, 122, 105583. [Google Scholar] [CrossRef]
- Fischer, C; Milting, H; Fein, E; Reiser, E; Lu, K; Seidel, T; et al. Long-term functional and structural preservation of precision-cut human myocardium under continuous electromechanical stimulation in vitro. Nat Commun 2019, 10, 117. [Google Scholar] [CrossRef]
- Watson, SA; Duff, J; Bardi, I; Zabielska, M; Atanur, SS; Jabbour, RJ; et al. Biomimetic electromechanical stimulation to maintain adult myocardial slices in vitro. Nat Commun 2019, 10, 2168. [Google Scholar] [CrossRef]
- Wang, K; Terrar, D; Gavaghan, DJ; Mu-u-min, R; Kohl, P; Bollensdorff, C. Living cardiac tissue slices: An organotypic pseudo two-dimensional model for cardiac biophysics research. Prog Biophys Mol Biol 2014, 115, 314–27. [Google Scholar] [CrossRef]
- Klumm, MJ; Heim, C; Fiegle, DJ; Weyand, M; Volk, T; Seidel, T. Long-Term Cultivation of Human Atrial Myocardium. Front Physiol 2022, 13, 839139. [Google Scholar] [CrossRef]
- van der Geest, JSA; de Boer, TP; Terracciano, CM; Thum, T; Dendorfer, A; Doevendans, PA; et al. Living myocardial slices: walking the path towards standardization. Cardiovasc Res 2025, 121, 1011–23. [Google Scholar] [CrossRef]
- Abu-Khousa, M; Fiegle, DJ; Sommer, ST; Minabari, G; Milting, H; Heim, C; et al. The Degree of t-System Remodeling Predicts Negative Force-Frequency Relationship and Prolonged Relaxation Time in Failing Human Myocardium. Front Physiol 2020, 11, 182. [Google Scholar] [CrossRef]
- Amesz, JH; Langmuur, SJJ; Zhang, L; Manintveld, OC; Schinkel, AFL; de Jong, PL; et al. Biomechanical response of ultrathin slices of hypertrophic cardiomyopathy tissue to myosin modulator mavacamten. Biomedicine and Pharmacotherapy 2024, 170, 116036. [Google Scholar] [CrossRef] [PubMed]
- Krammer, T; Baier, MJ; Hegner, P; Zschiedrich, T; Lukas, D; Wolf, M; et al. Cardioprotective effects of semaglutide on isolated human ventricular myocardium. Eur J Heart Fail 2025, 27, 1315–25. [Google Scholar] [CrossRef] [PubMed]
- Bierhuizen, MFA; Amesz, JH; Langmuur, SJJ; Lam, B; Knops, P; Veen, KM; et al. Acute Biomechanical Effects of Cardiac Contractility Modulation in Living Myocardial Slices from End-Stage Heart Failure Patients. Bioengineering 2025, 12, 174. [Google Scholar] [CrossRef] [PubMed]
- Shi, R; Reichardt, M; Fiegle, DJ; Küpfer, LK; Czajka, T; Sun, Z; et al. Contractility measurements for cardiotoxicity screening with ventricular myocardial slices of pigs. Cardiovasc Res 2023, 119, 2469–81. [Google Scholar] [CrossRef]
- Habeler, W; Peschanski, M; Monville, C. Organotypic heart slices for cell transplantation and physiological studies. Organogenesis 2009, 5, 62–6. [Google Scholar] [CrossRef]
- Baron, V; Sommer, ST; Fiegle, DJ; Pfeuffer, A-KM; Peyronnet, R; Volk, T; et al. Effects of electro-mechanical uncouplers, hormonal stimulation and pacing rate on the stability and function of cultured rabbit myocardial slices. Front Bioeng Biotechnol 2024, 12, 1363538. [Google Scholar] [CrossRef]
- Quinn, TA; Granite, S; Allessie, MA; Antzelevitch, C; Bollensdorff, C; Bub, G; et al. Minimum Information about a Cardiac Electrophysiology Experiment (MICEE): Standardised reporting for model reproducibility, interoperability, and data sharing. Prog Biophys Mol Biol 2011, 107, 4–10. [Google Scholar] [CrossRef]
- Niederer, SA; Lumens, J; Trayanova, NA. Computational models in cardiology. Nat Rev Cardiol 2019, 16, 100–11. [Google Scholar] [CrossRef]
- Trayanova, NA; Lyon, A; Shade, J; Heijman, J. Computational modeling of cardiac electrophysiology and arrhythmogenesis: toward clinical translation. Physiol Rev 2024, 104, 1265–333. [Google Scholar] [CrossRef]
- Lai, C; Yin, M; Kholmovski, EG; Sani, MM; Gilotra, NA; Chrispin, J; et al. Predicting Sudden Cardiac Death in Patients With Sarcoidosis Using a Multimodal Artificial Intelligence Model. JACC Clin Electrophysiol 2025, S2405-500X(25)00829-1. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, C; Sakata, K; Ali, SY; Loeffler, S; Prakosa, A; Tice, B; et al. Arrhythmogenic substrates in atrial fibrillation and the role of ablation lesions: A longitudinal biatrial digital twin study. Cardiovasc Res 2026. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K; Magtibay, K; Trayanova, N; Vigmond, E. A model of β-adrenergic stimulation in human ventricular cells for tissue-scale simulations of sympathetically modulated tachycardias. J Physiol 2026. [Google Scholar] [CrossRef] [PubMed]
- Loewe, A; Hunter, PJ; Kohl, P. Computational modelling of biological systems now and then: revisiting tools and visions from the beginning of the century. Philosophical Transactions of the Royal Society A: Mathematical, Physical and Engineering Sciences 2025, 383, 20230384. [Google Scholar] [CrossRef]
- Quinn, TA; Kohl, P. Combining wet and dry research: experience with model development for cardiac mechano-electric structure-function studies. Cardiovasc Res 2013, 97, 601–11. [Google Scholar] [CrossRef]
- Roney, CH; Sim, I; Yu, J; Beach, M; Mehta, A; Alonso Solis-Lemus, J; et al. Predicting Atrial Fibrillation Recurrence by Combining Population Data and Virtual Cohorts of Patient-Specific Left Atrial Models. Circ Arrhythm Electrophysiol 2022, 15, e010253. [Google Scholar] [CrossRef]
- Corral-Acero, J; Margara, F; Marciniak, M; Rodero, C; Loncaric, F; Feng, Y; et al. The ‘Digital Twin’ to enable the vision of precision cardiology. Eur Heart J 2020, 41, 4556–64. [Google Scholar] [CrossRef]
- Shade, JK; Prakosa, A; Popescu, DM; Yu, R; Okada, DR; Chrispin, J; et al. Predicting risk of sudden cardiac death in patients with cardiac sarcoidosis using multimodality imaging and personalized heart modeling in a multivariable classifier. Sci Adv 2021, 7, eabi8020. [Google Scholar] [CrossRef]
- Sakata, K; Bradley, RP; Prakosa, A; Yamamoto, CAP; Ali, SY; Loeffler, S; et al. Assessing the arrhythmogenic propensity of fibrotic substrate using digital twins to inform a mechanisms-based atrial fibrillation ablation strategy. Nature Cardiovascular Research 2024, 3, 857–68. [Google Scholar] [CrossRef]
- Sakata, K; Yamamoto, CAP; Ali, SY; Loeffler, S; Prakosa, A; Tice, BM; et al. Assessment of persistent atrial fibrillation arrhythmogenesis in the right atrium using digital twins. Heart Rhythm 2026, 23, e123–32. [Google Scholar] [CrossRef]
- Waight, MC; Prakosa, A; Li, AC; Truong, A; Bunce, N; Marciniak, A; et al. Heart Digital Twins Predict Features of Invasive Reentrant Circuits and Ablation Lesions in Scar-Dependent Ventricular Tachycardia. Circ Arrhythm Electrophysiol 2025, 18, e013660. [Google Scholar] [CrossRef]
- Vicente, J; Zusterzeel, R; Johannesen, L; Mason, J; Sager, P; Patel, V; et al. Mechanistic Model-Informed Proarrhythmic Risk Assessment of Drugs: Review of the “CiPA” Initiative and Design of a Prospective Clinical Validation Study. Clin Pharmacol Ther 2018, 103, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Li, Z; Mirams, GR; Yoshinaga, T; Ridder, BJ; Han, X; Chen, JE; et al. General Principles for the Validation of Proarrhythmia Risk Prediction Models: An Extension of the CiPA In Silico Strategy. Clin Pharmacol Ther 2020, 107, 102–11. [Google Scholar] [CrossRef] [PubMed]
- Food and Drug Administration. FDA Announces Plan to Phase Out Animal Testing Requirement for Monoclonal Antibodies and Other Drugs 2025. Available online: https://www.fda.gov/news-events/press-announcements/fda-announces-plan-phase-out-animal-testing-requirement-monoclonal-antibodies-and-other-drugs (accessed on 10 February 2026).
- National Institutes of Health. NIH to prioritize human-based research technologies 2025. Available online: https://www.nih.gov/news-events/news-releases/nih-prioritize-human-based-research-technologies (accessed on 10 February 2026).
- Ridder, BJ; Leishman, DJ; Bridgland-Taylor, M; Samieegohar, M; Han, X; Wu, WW; et al. A systematic strategy for estimating hERG block potency and its implications in a new cardiac safety paradigm. Toxicol Appl Pharmacol 2020, 394, 114961. [Google Scholar] [CrossRef] [PubMed]
- Assessing the credibility of computational modeling through verification and validation: application to medical devices; American Society of Mechanical Engineers, 2018.
- Toward Good Simulation Practice; Viceconti, M, Emili, L, Eds.; Springer Nature Switzerland: Cham, 2024. [Google Scholar] [CrossRef]
- Iaizzo, PA. The Visible Heart® project and free-access website “Atlas of Human Cardiac Anatomy”. Europace 2016, 18, iv163–72. [Google Scholar] [CrossRef]
- Bunnik, EM. Ethics of allocation of donor organs. Curr Opin Organ Transplant 2023, 28, 192–6. [Google Scholar] [CrossRef]
- Pick, F; Krug, N. Was Deutschland von anderen Ländern lernen kann. Spiegel. 2024. Available online: https://www.spiegel.de/gesundheit/organspende-register-was-deutschland-von-anderen-laendern-lernen-kann-a-d1eecb0f-27d0-4bc8-9d58-2c22bacf469a (accessed on 22 October 2024).
- Eurotransplant. Deutschland 2024. Available online: https://www.eurotransplant.org/region/deutschland/ (accessed on 9 October 2024).
- Eurotransplant Statistics Report Library. Heart donation, deceased donors in 2023, by country, by allocation phase. n.d. [Google Scholar]
- IRODaT. 2024. Available online: https://www.irodat.org/?p=database&c=DE#data (accessed on 9 October 2024).
- Statista. Number of postmortem organ donors in Germany from 1998 to 2023 2024. Available online: https://www.statista.com/statistics/1384771/postmortem-organ-donors-germany/ (accessed on 9 October 2024).
- Mumford, L. NHS BLOOD & TRANSPLANT RESEARCH, INNOVATION AND NOVEL TECHNOLOGIES ADVISORY GROUP RESEARCH CONSENT/AUTHORISATION RATES. 2022.
- Bledsoe, MJ; Grizzle, WE. Use of human specimens in research: the evolving United States regulatory, policy, and scientific landscape. Diagn Histopathol 2013, 19, 322–30. [Google Scholar] [CrossRef]
- Grizzle, WE. Issues in the Use of Human Tissues to Support Precision Medicine. J Health Care Poor Underserved 2019, 30, 66–78. [Google Scholar] [CrossRef]
- Lutomski, JE; Manders, P. From opt-out to opt-in consent for secondary use of medical data and residual biomaterial: An evaluation using the RE-AIM framework. PLoS One 2024, 19, e0299430. [Google Scholar] [CrossRef]
- Fitzpatrick, PE; McKenzie, KD; Beasley, A; Sheehan, JD. Patients attending tertiary referral urology clinics: Willingness to participate in tissue banking. BJU Int 2009, 104, 209–13. [Google Scholar] [CrossRef]
- Jack, AL; Womack, C. Why surgical patients do not donate tissue for commercial research: review of records. BMJ 2003, 327, 262. [Google Scholar] [CrossRef]
- Rehsmann, J. A revealing scandal: The German transplant scandal between structural failures, moralizing rules, and ambivalent manipulations. Journal of Liver Transplantation 2023, 11, 100168. [Google Scholar] [CrossRef]
- Deutscher Bundestag. Organspenden: Mehrheit für die Entscheidungslösung 2020. Available online: https://www.bundestag.de/dokumente/textarchiv/2020/kw03-de-transplantationsgesetz-674682 (accessed on 9 October 2024).
- Deutsche Welle. German lawmakers reject “opt-out” organ donor bill 2020. Available online: https://www.dw.com/en/german-parliament-explicit-consent-still-necessary-from-organ-donors/a-52022245 (accessed on 9 October 2024).
- Kappler, HE; Kohl, P. Wissenschaftliche Nutzung von nicht transplantiertem Humangewebe. CardioNews 2023. Available online: https://app.cardionews.de/Politik-Gesellschaft/Wissenschaftliche-Nutzung-von-nicht-transplantiertem-Humangewebe-442919.html (accessed on 23 February 2026).
- Deutscher Bundestag 20. Wahlperiode. Antwort der Bundesregierung auf die Kleine Anfrage der Fraktion der CDU/CSU – Drucksache 20/7480 – Nutzung postmortaler Spender-Organe zu Forschungszwecken. Drucksache 20/7731 2023. Available online: https://dserver.bundestag.de/btd/20/077/2007731.pdf (accessed on 23 February 2026).
- Deutscher Bundestag 20. Wahlperiode. Kleine Anfrage der Fraktion der CDU/CSU Nutzung postmortaler Spender-Organe zu Forschungszwecken. Drucksache 20/7480 2023. Available online: https://dserver.bundestag.de/btd/20/074/2007480.pdf (accessed on 23 February 2026).
- Fedlex. Federal Act of 30 September 2011 on Research involving Human Beings, SR 810. 30 2011. Available online: https://www.fedlex.admin.ch/eli/cc/2013/617/en (accessed on 6 January 2026).
- Federal Public Health Food Chain Safety and Environment. Organ donation and donation of body to science. Federal Public Health Food Chain Safety and Environment 2016. Available online: https://www.health.belgium.be/en/organ-donation-and-donation-body-science (accessed on 20 December 2025).
- Ministry of Health Welfare and Sport Donor Register. Scientific Research 2025. Available online: https://english.donorregister.nl/recording-your-choise/what-choices-do-you-have/scientific-research (accessed on 9 January 2026).
- Ustaw, Dziennik. Ustawa z dnia 1 lipca 2005 r. o pobieraniu, przechowywaniu i przeszczepianiu komórek, tkanek i narządów [Act of 1 July 2005 on the Collection, Storage, and Transplantation of Cells, Tissues, and Organs.] Dz. U. 2005 No. 169 item 141 n.d. Available online: https://isap.sejm.gov.pl/isap.nsf/DocDetails.xsp?id=WDU20051691411 (accessed on 9 January 2026).
- Retsinformation.dk. Vejledning om samtykke til transplantation fra afdøde personer og til transplantationsrelateret forskning [Guidance on consent for transplantation from deceased persons and for transplantation-related research], VEJ nr 10009 of 05/12/2019. RetsinformationDk n.d. Retsinformation.dk. Available online: https://www.retsinformation.dk/eli/retsinfo/2019/10099 (accessed on 9 January 2026).
- Agence de la biomédicine. Baromètre d’opinion 2024 2024. Available online: https://back.agence-biomedecine.fr/uploads/Dossier_de_presse_complet_en_PDF_6598766daa.pdf (accessed on 8 January 2026).
- Rechtsinformationssystem des Bundes. Organtransplantationsgesetz [Federal Act on the Transplantation of Human Organs], §2, Abs. 2. n.d. Available online: https://www.ris.bka.gv.at/GeltendeFassung.wxe?Abfrage=Bundesnormen&Gesetzesnummer=20008119 (accessed on 9 January 2026).
- NHS Blood and Transplant. Understanding consent for organ donation. n.d. Available online: https://www.organdonation.nhs.uk/about-organ-donation/consent/ (accessed on 23 February 2026).
Figure 1.
Advantages and disadvantages of various vertebrate models used in cardiovascular research [11,12,13].
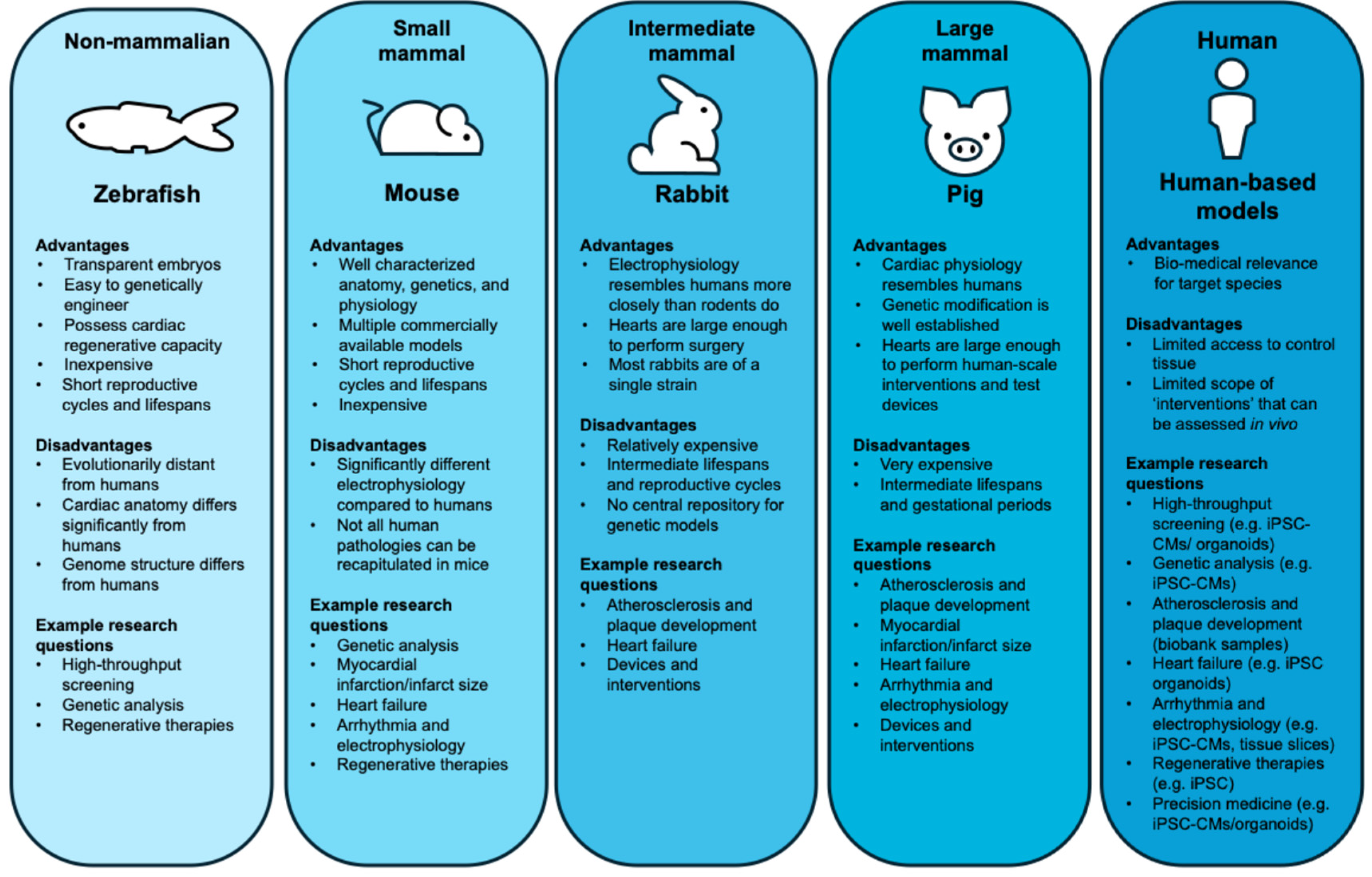
Figure 2.
Why non-failing hearts are not used as control tissue in scientific research in Germany. a Eurotransplant Statistics Report Library 2024 [175].
Figure 2.
Why non-failing hearts are not used as control tissue in scientific research in Germany. a Eurotransplant Statistics Report Library 2024 [175].
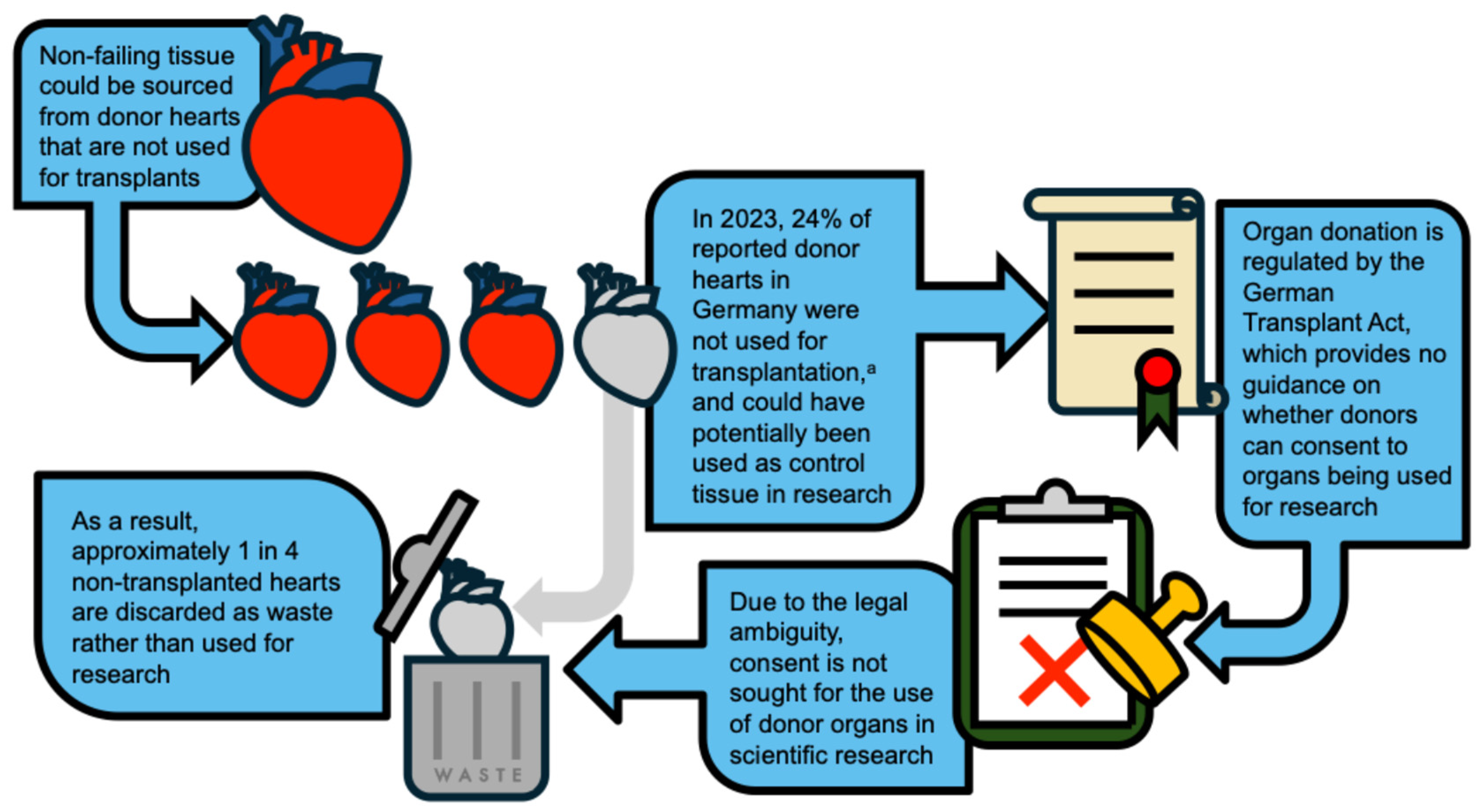
Figure 3.
Laws and guidelines relating to the use of non-transplanted donor hearts for research in selected European countries [190,191,192,193,194,195,196,197].
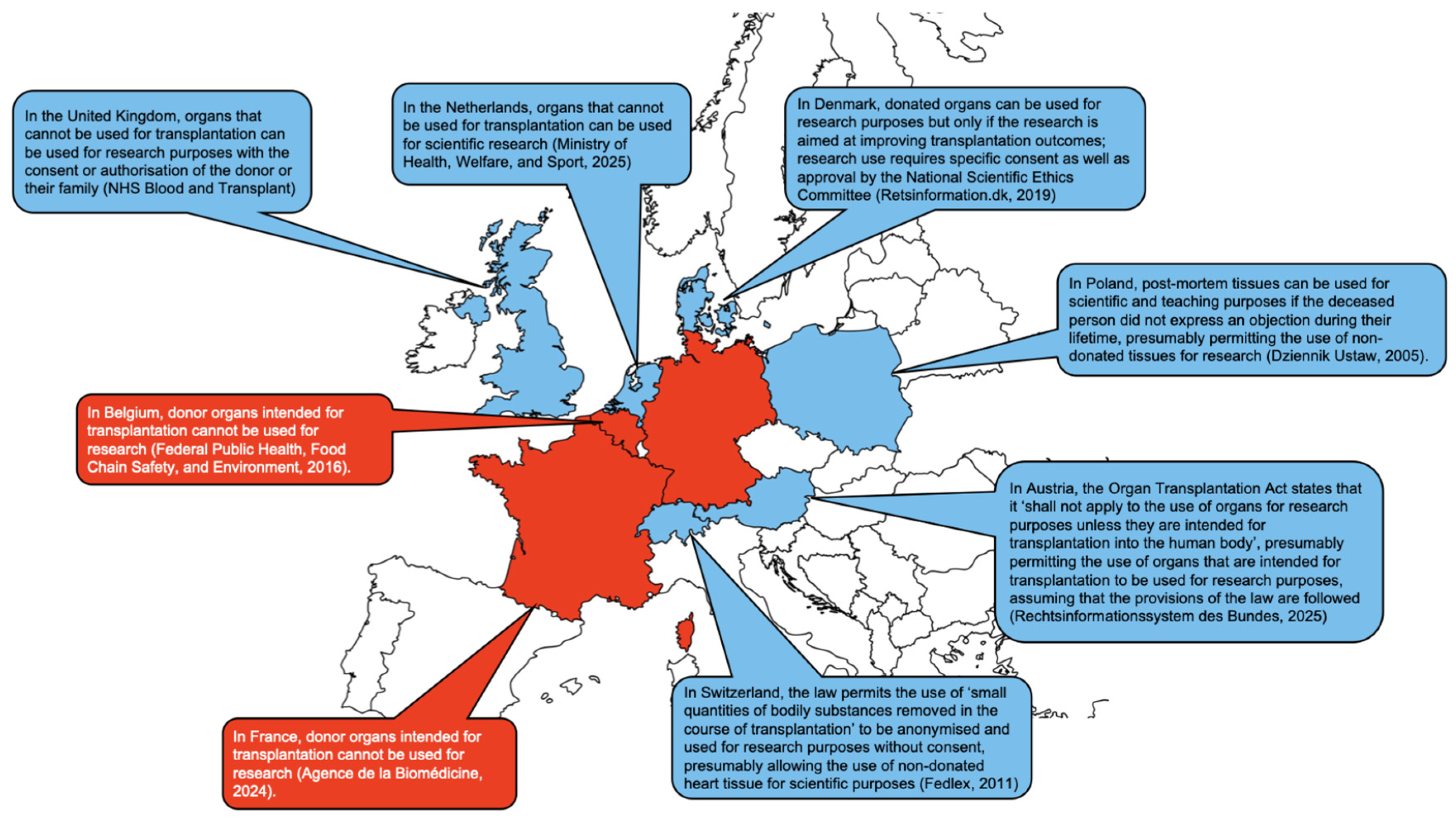

Table 1.
Key parameters of selected vertebrate models commonly used in cardiovascular research (other commonly used models include invertebrates, chick embryos, rats, sheep, goats, non-human primates, and historically also cats and dogs).
Table 1.
Key parameters of selected vertebrate models commonly used in cardiovascular research (other commonly used models include invertebrates, chick embryos, rats, sheep, goats, non-human primates, and historically also cats and dogs).
| Parameter | Non-mammalian, e.g., zebrafish |
Small mammal, e.g., mouse |
Intermediate mammal, e.g., rabbit |
Large mammal, e.g., pig |
Human |
| Body weight | 0.5 g | 20–45 g | 1-5 kg | 40–90 kg | 50–100 kg |
| Heart weight | 0.5 mg | 0.15–0.2 g | 5–10 g | 145–450 g | 150–480 g |
| Resting heart rate | 110–170 bpm | 450–800 bpm | 140-280 bpm | 70–120 bpm | 60–100 bpm |
| Cardiac output | 0.09–0.172 mL/min | 13–15 mL/min | 260-420 mL/min | 2,720–3,620 mL/min | 3,340–9,790 mL/min |
| Ejection fraction | 40–50% | 65–90% | 55–65% | 50–55% | 55–60% |
| Ventricular action potential duration | 150–250 ms | 80–180 ms | 150–280 ms | 180–300 ms | 200–400 ms |
| Conduction velocity | 4–10 mm/s | 350–600 mm/s | 300-500 mm/s | 500–800 mm/s | 500–900 mm/s |
| Reentry space factora | 0.42 | 0.34 | 0.69 | 1.43 | 1.5 |
| Approximate cost per animal | €2.00–2.50/tank | €4–7 | €250–600 | €80–400+ | - |
| Husbandry notes | Swarm fish, density 4–5 animals per litre | Individually ventilated or standard cages | Larger pens | Large facilities or catheterisation laboratories, low volume but translational | - |
| Anaesthesia, and effects on cardiovascular endpoints | Tricaine: bradycardia, decreased heart rate and contractility, effects dose-dependent | Isoflurane and opioids: decreased heart rate and blood pressure | Volatile or injectable anaesthetics: altered haemodynamics, effects on heart rhythm | Isoflurane and similar: more frequent arrhythmias, infarct variability, effects on blood pressure effects | - |
| Typical n per group in cardiovascular studies | 30–100+ embryos | 8–15 | 6–10 | 6–12 | - |
a Calculated as maximum ventricular circumference divided by (conduction velocity × action potential duration). Colour indicates good (green) or satisfactory (yellow) match to human cardiac parameters.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



