Submitted:
19 February 2026
Posted:
27 February 2026
You are already at the latest version
Abstract
The gut microbiota–brain axis has emerged as a key regulator of central nervous system (CNS) development and function. Beyond its established roles in immunity and behavior, growing evidence indicates that microbial-derived metabolites influence glial biology, including oligodendrocyte development and myelination. In this review, we synthesize current findings supporting a role for microbial metabolites—particularly short-chain fatty acids (SCFAs), tryptophan-derived indoles, and secondary bile acids—in regulating oligodendrocyte precursor cell (OPC) proliferation, differentiation, and myelin formation. We examine the molecular and cellular mechanisms involved, including epigenetic regulation through histone deacetylase inhibition, G-protein–coupled receptor signaling, nuclear receptor activation, and metabolic support pathways. Experimental and clinical studies indicate that butyrate modulates oligodendrocyte lineage progression via chromatin remodeling, indole derivatives signal through the aryl hydrocarbon receptor (AhR) to attenuate neuroinflammation and promote glial homeostasis, and secondary bile acids act through receptors such as TGR5 and the vitamin D receptor to support pro-remyelinating environments. Additionally, microbial acetate contributes to acetyl-CoA pools necessary for lipid biosynthesis and epigenetic regulation within oligodendrocytes. Notably, dysbiosis and reduced availability of microbial metabolites have been consistently reported in Multiple Sclerosis (MS), a demyelinating disorder characterized by impaired remyelination, suggesting that microbiome-targeted strategies—including dietary interventions and postbiotic supplementation—may complement existing therapies. Collectively, current evidence supports the concept that microbial metabolites function as systemic modulators of CNS myelination, linking dietary and microbial inputs to oligodendrocyte biology and highlighting promising avenues for translational research in demyelinating diseases.
Keywords:
microbiota–gut–brain axis
; oligodendrocytes
; myelination
; short-chain fatty acids
; epigenetic regulation
; multiple sclerosis
; remyelination
; postbiotics
1. Introduction
The myelin sheath represents one of the most remarkable evolutionary innovations of the vertebrate nervous system. This multilayered lipid membrane, produced by oligodendrocytes in the central nervous system (CNS), enables saltatory conduction of action potentials, increasing nerve impulse velocity by up to 100-fold while reducing metabolic costs [1,2]. Beyond its insulating function, myelin provides essential metabolic support to axons through the transfer of lactate and other energy substrates, establishing a symbiotic relationship critical for neuronal survival [3,4].
1.1. The Oligodendrocyte Lineage: A Developmental Overview
Oligodendrocyte development proceeds through a well-characterized sequence of cellular stages. Oligodendrocyte precursor cells (OPCs), identified by expression of NG2, PDGFR, and Olig2, arise from ventricular zone progenitors and migrate extensively throughout the CNS [6,7]. Upon receiving appropriate environmental cues, OPCs exit the cell cycle and differentiate into pre-myelinating oligodendrocytes, characterized by elaboration of complex membrane processes. Terminal differentiation yields mature myelinating oligodendrocytes expressing myelin structural proteins including myelin basic protein (MBP), proteolipid protein (PLP), and myelin-associated glycoprotein (MAG) [8,9].
This developmental trajectory was traditionally viewed as governed primarily by intrinsic genetic programs and local signaling from the neurovascular niche, including growth factors (PDGF, FGF), axonal signals (neuregulin-1), and extracellular matrix components [10,11]. However, emerging evidence has broadened this perspective to include systemic influences originating from an unexpected source: the gut microbiota.
1.2. The Microbiota-Gut-Brain Axis: A Paradigm Shift
The human gut harbors approximately microorganisms representing over 1,000 species, collectively termed the gut microbiota [12]. This microbial community functions as a virtual endocrine organ, producing an extraordinary diversity of metabolites that enter systemic circulation and influence distant organ systems, including the brain [13,14]. The bidirectional communication network linking gut microbiota to brain function—the microbiota-gut-brain axis—has emerged as a critical determinant of neurodevelopment, behavior, and neurological disease [15,16].
The significance of the microbiota for CNS myelination was first revealed by studies in germ-free (GF) mice, which are raised in sterile isolators and lack any microbial colonization. These animals exhibit striking defects in myelin gene expression and white matter structure, particularly in the prefrontal cortex [17]. Importantly, many of these defects can be rescued by post-natal colonization with a complex microbiota, demonstrating that microbial signals are not merely correlative but causally related to myelination [18].
Recent work further demonstrates that gut microbiota composition directly influences optic nerve fiber maturation and myelination in germ-free and gnotobiotic mice [5].
1.3. Microbial Metabolites: The Molecular Messengers
Among the myriad molecules produced by gut bacteria, three classes have emerged as particularly important regulators of oligodendrocyte biology:
- 1.
- 2.
- 3.
1.4. Scope and Objectives of This Review
This comprehensive review aims to synthesize current knowledge regarding the mechanisms by which microbial metabolites regulate oligodendrocyte lineage dynamics and myelination. We will examine:
- The pathways through which metabolites access the CNS
- Epigenetic mechanisms, particularly HDAC inhibition by SCFAs
- G-protein coupled receptor (GPCR) signaling cascades
- Metabolic support of myelin lipid biosynthesis
- Clinical implications for Multiple Sclerosis and other demyelinating disorders
- Therapeutic frontiers including postbiotics and dietary interventions
2. The Microbiota-Gut-Brain Axis: Pathways of Metabolite Delivery
For microbial metabolites to influence oligodendrocyte biology, they must first traverse multiple physiological barriers to access the CNS parenchyma. Understanding these pathways is essential for appreciating how gut-derived signals reach their cellular targets.
2.1. Intestinal Absorption and Systemic Distribution
SCFAs are absorbed across the colonic epithelium through multiple mechanisms. Passive diffusion of protonated forms occurs at the acidic luminal pH, while active transport is mediated by monocarboxylate transporters (MCT1, MCT4) and the sodium-coupled transporter SMCT1 (SLC5A8) [25,26]. Once absorbed, SCFAs enter the portal circulation and are partially metabolized by the liver, with remaining fractions reaching systemic circulation at concentrations of 100-400 M for acetate and 1-10 M for propionate and butyrate [27,28].
Tryptophan metabolites and secondary bile acids similarly enter systemic circulation through specific transporters. Indole derivatives are absorbed via passive diffusion and reach plasma concentrations of 0.5-10 M [29]. Secondary bile acids utilize the apical sodium-dependent bile acid transporter (ASBT) and organic solute transporters (OST/OST) [30].
2.2. Blood-Brain Barrier Penetration
The blood-brain barrier (BBB) represents a selective interface that restricts molecular traffic between systemic circulation and the CNS. Critically, the gut microbiota itself influences BBB integrity. Germ-free mice exhibit increased BBB permeability due to reduced expression of tight junction proteins including occludin, claudin-5, and ZO-1 [31]. Colonization with SCFA-producing bacteria or direct butyrate administration restores BBB integrity, creating a regulatory feedback loop wherein the microbiota controls access of its own metabolites to the brain.
SCFAs cross the BBB primarily via MCT1, which is expressed on brain endothelial cells. Brain concentrations reach approximately 17.0 M for acetate, 2.1 M for propionate, and 2.8 M for butyrate in rodents [32]. These concentrations, while lower than systemic levels, are within the range reported to activate cellular receptors and modulate enzymatic activity in experimental systems.
Indole derivatives, being lipophilic, can cross the BBB via passive diffusion. IPA reaches brain concentrations sufficient to activate the pregnane X receptor (PXR) and provide neuroprotection [29,148]. Secondary bile acids utilize organic anion transporting polypeptides (OATPs) expressed at the BBB, though their CNS concentrations remain less well characterized [34].
2.3. Microglial and Astrocytic Intermediaries
Microbial metabolites do not always act directly on oligodendrocytes; their effects are frequently mediated through other glial populations. This indirect signaling represents a crucial amplification mechanism.
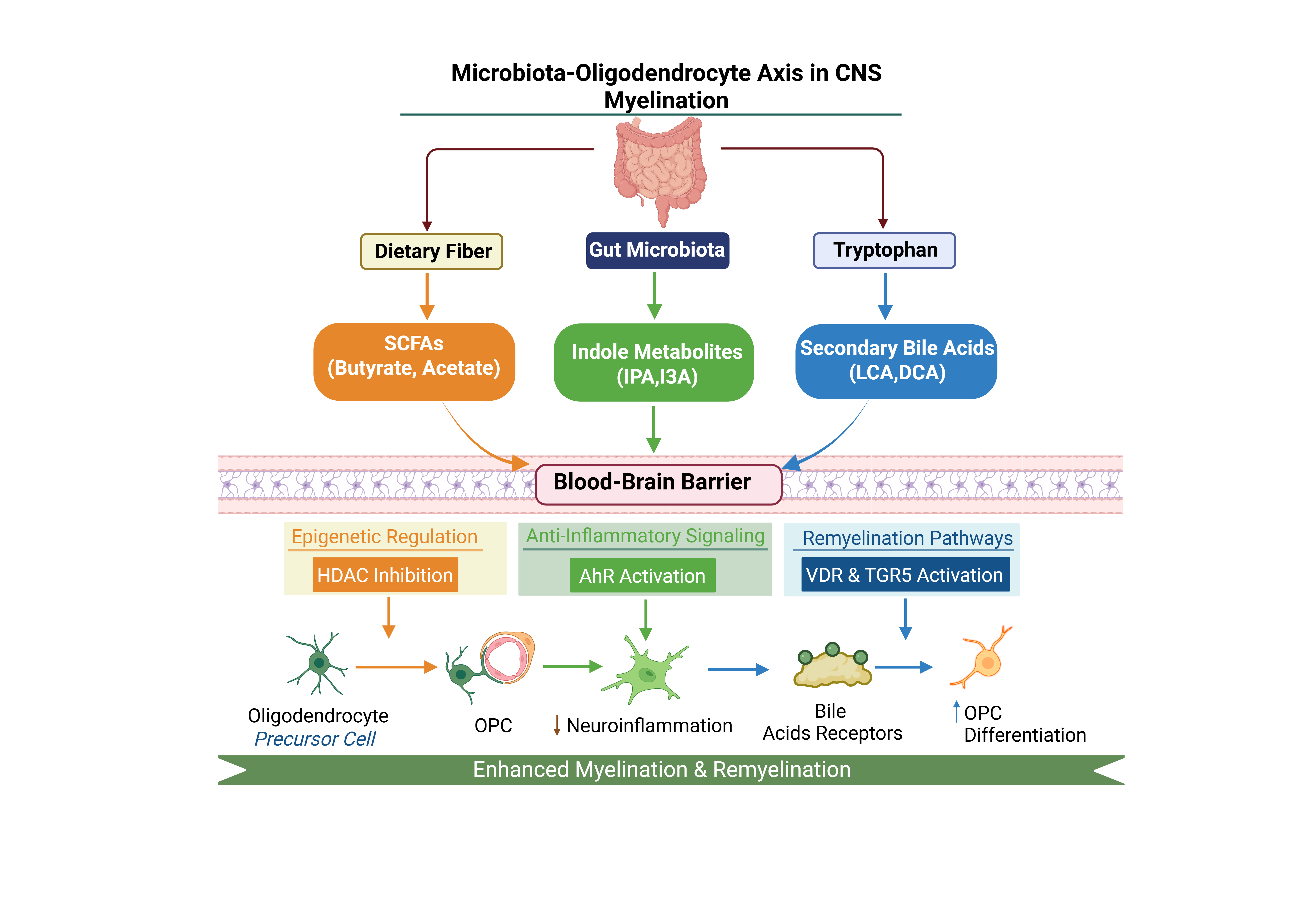
As summarized in Figure 1, microbial metabolites target multiple glial cell types through specific transport mechanisms.
2.3.1. Microglial Maturation and Function
Microglia, the resident macrophages of the CNS, are exquisitely sensitive to microbial signals. Germ-free mice exhibit immature microglia characterized by altered morphology, reduced ramification, and impaired responses to pathological stimuli [35]. SCFA supplementation rescues microglial maturation, restoring their capacity for phagocytosis and cytokine production.
Since efficient remyelination requires microglial clearance of myelin debris from demyelinated lesions, the microbial regulation of microglial function indirectly influences OPC differentiation and myelin repair [36,37]. Microglia also secrete factors that directly influence OPC behavior, including IGF-1 and activin-A, creating a complex signaling network linking microbial inputs to oligodendrocyte responses [38].
2.3.2. Astrocyte-Oligodendrocyte Crosstalk
Astrocytes represent another critical intermediary in microbiota-oligodendrocyte communication. Tryptophan-derived indoles activate the aryl hydrocarbon receptor (AhR) in astrocytes, suppressing production of pro-inflammatory cytokines including IL-6, TNF-, and IL-1 [39]. Since these cytokines inhibit OPC differentiation and can be directly toxic to oligodendrocytes, the anti-inflammatory effects of microbial indoles on astrocytes create a permissive environment for myelination.
Astrocytes also provide metabolic support to oligodendrocytes through gap junction-mediated transfer of metabolites. The panglial syncytium, formed by connexin-based coupling between astrocytes and oligodendrocytes, enables direct metabolic communication [40,41]. Microbial metabolites that influence astrocyte metabolism may therefore indirectly affect oligodendrocyte bioenergetics.
3. Short-Chain Fatty Acids: Epigenetic and Metabolic Regulation
Short-chain fatty acids represent the most extensively studied class of microbial metabolites in the context of CNS myelination. Their mechanisms of action span epigenetic regulation, G-protein coupled receptor signaling, and direct metabolic contribution.
3.1. SCFA Biosynthesis and Distribution
SCFAs are produced in the cecum and colon through bacterial fermentation of non-digestible carbohydrates, primarily dietary fiber and resistant starch. The major fermentation pathways include:
- Acetate: Produced via acetyl-CoA and the Wood-Ljungdahl pathway by diverse bacterial taxa including Bacteroides, Bifidobacterium, and Akkermansia [42].
- Propionate: Synthesized through the succinate, acrylate, and propanediol pathways, primarily by Bacteroidetes, Veillonella, and Propionibacterium [43].

3.2. Butyrate as a Histone Deacetylase Inhibitor

The most profound mechanism by which SCFAs regulate oligodendrocyte biology is through inhibition of histone deacetylases (HDACs). Butyrate acts as a non-competitive inhibitor of Class I HDACs (HDAC1, 2, 3, 8) and Class IIa HDACs (HDAC4, 5, 7, 9), with IC50 values in the low micromolar range [46,47].
3.2.1. HDAC Function in Oligodendrocyte Development
The role of HDACs in oligodendrocyte development is paradoxically complex. During the transition from OPC to mature oligodendrocyte, HDAC1 and HDAC2 are required to repress genes that maintain the progenitor state, including Sox2, Id2, Id4, and Hes5 [48]. Genetic deletion of both HDAC1 and HDAC2 in the oligodendrocyte lineage severely impairs differentiation and myelination.
However, the timing of HDAC activity is critical. Sustained HDAC activity can prevent terminal differentiation by maintaining repressive chromatin states at myelin gene promoters. The transient inhibition of HDACs by butyrate may therefore facilitate a permissive epigenetic state favoring the release of pro-myelinating genes (including Mbp, Plp1, Mag, and Mog) from epigenetic repression while allowing completion of the differentiation program [49,50].
3.2.2. Experimental Evidence for HDAC Inhibition in Myelination
Multiple lines of evidence support the pro-myelinating effects of HDAC inhibition:
- 1.
- 2.
- In vitro treatment of OPCs with butyrate or other HDAC inhibitors (valproic acid, trichostatin A) promotes expression of MBP and PLP [50].
- 3.
- Cuprizone-induced demyelination models show accelerated remyelination following butyrate supplementation [51].
- 4.
- Chromatin immunoprecipitation (ChIP) studies demonstrate increased histone H3 and H4 acetylation at myelin gene promoters following SCFA treatment [18].
3.3. G-Protein Coupled Receptor Signaling
Beyond epigenetic effects, SCFAs activate specific G-protein coupled receptors (GPCRs) that trigger intracellular signaling cascades.
3.3.1. GPR41 and GPR43
GPR41 (FFAR3) and GPR43 (FFAR2) are the primary SCFA receptors, with distinct expression patterns and ligand preferences [52,53]:
- GPR43: Highest affinity for acetate and propionate; expressed on microglia and immune cells
- GPR41: Highest affinity for propionate and butyrate; expressed on neurons and enteroendocrine cells
3.3.2. GPR109A (Hydroxycarboxylic Acid Receptor 2)
3.4. Acetate and the Acetyl-CoA Hub
Acetate, the most abundant SCFA in systemic circulation, serves as a direct metabolic substrate with implications for both lipid synthesis and epigenetic regulation.
3.4.1. Acetyl-CoA Synthetase Pathways
Upon entering cells, acetate is converted to acetyl-CoA by acetyl-CoA synthetases (AceCS1 in the cytoplasm/nucleus; AceCS2 in mitochondria) [57]. This acetyl-CoA serves multiple functions:
As illustrated in Figure 2, microbial acetate contributes to both epigenetic regulation and metabolic support of myelination through its conversion to acetyl-CoA.
3.4.2. Implications for Myelination
The metabolic link between gut-derived acetate and oligodendrocyte function is particularly significant given the extraordinary lipid demands of myelination. Myelin is approximately 70-80% lipid by dry weight, requiring massive biosynthetic capacity [61]. The flux of microbial acetate into acetyl-CoA may contribute to supporting this demand, while simultaneously providing acetyl groups for the epigenetic regulation of myelin genes.
3.5. Propionate: Immunomodulation and Beyond
Propionate, while less studied than butyrate in the CNS, exerts important immunomodulatory effects that indirectly support myelination.
3.5.1. Regulatory T Cell Induction
Propionate promotes the differentiation of regulatory T cells (Tregs) in the gut-associated lymphoid tissue through HDAC inhibition and GPR43 activation [63,64]. These Tregs can migrate to the CNS or release systemic anti-inflammatory signals that suppress Th17-mediated autoimmunity—a key pathogenic mechanism in Multiple Sclerosis [65].
Clinical trials have demonstrated that oral propionate supplementation increases circulating Treg populations in MS patients and correlates with reduced relapse rates [66]. This systemic immunomodulation creates a permissive environment for remyelination by reducing ongoing immune-mediated damage.
4. Tryptophan-Derived Metabolites and AhR Signaling
Tryptophan is an essential amino acid that serves as a precursor for multiple bioactive compounds. While host metabolism primarily directs tryptophan toward serotonin and kynurenine pathways, gut bacteria produce a distinct repertoire of indole derivatives with potent neuroactive properties [21,22,67].
4.1. Microbial Tryptophan Metabolism
Gut bacteria metabolize tryptophan through several pathways:
- 1.
- Indole pathway: Tryptophanase (TnaA) converts tryptophan to indole, which can be further modified to produce indole-3-aldehyde, indole-3-acetic acid, and indole-3-propionic acid (IPA) [68].
- 2.
- Tryptamine pathway: Decarboxylation produces tryptamine, a neuroactive amine [69].
- 3.
- Kynurenine pathway: Some bacteria possess indoleamine 2,3-dioxygenase (IDO) homologs that produce kynurenine metabolites [70].
Key bacterial genera involved in tryptophan metabolism include Lactobacillus, Clostridium, Bacteroides, and Peptostreptococcus [22].
4.2. The Aryl Hydrocarbon Receptor (AhR)
The aryl hydrocarbon receptor is a ligand-activated transcription factor belonging to the basic helix-loop-helix/Per-ARNT-Sim (bHLH-PAS) family. Originally characterized as a xenobiotic sensor, AhR is now recognized as a key mediator of microbial-host communication in the CNS [39,71].
4.2.1. Mechanism of AhR Activation
In the absence of ligand, AhR resides in the cytoplasm complexed with HSP90, XAP2, and p23. Ligand binding (e.g., by indole derivatives) triggers conformational changes, nuclear translocation, and heterodimerization with ARNT (AhR nuclear translocator). The AhR-ARNT complex binds xenobiotic response elements (XREs) in target gene promoters, modulating transcription [72].
4.2.2. AhR in Astrocytes: Suppression of Neuroinflammation
A seminal series of studies by Rothhammer and colleagues demonstrated that microbial tryptophan metabolites activate AhR in astrocytes, triggering anti-inflammatory transcriptional programs [39,73]. Specifically:
- AhR activation suppresses NF-B signaling in astrocytes
- Production of pro-inflammatory cytokines (IL-6, TNF-, CCL2) is reduced
- Astrocytes adopt a neuroprotective phenotype supporting oligodendrocyte survival
In experimental autoimmune encephalomyelitis (EAE), a mouse model of MS, dietary tryptophan supplementation or direct indole administration reduces disease severity through AhR-dependent mechanisms [39].
4.2.3. AhR in Microglia
AhR signaling in microglia modulates their activation state and phagocytic capacity. Microbial metabolites acting through microglial AhR suppress production of inflammatory mediators while maintaining the capacity for myelin debris clearance [73]. This balance is critical for efficient remyelination.
4.3. Indole-3-Propionic Acid: A Potent Neuroprotectant
Indole-3-propionic acid (IPA) is produced exclusively by gut bacteria, primarily Clostridium sporogenes, and is one of the most potent naturally occurring antioxidants [29,74].
4.3.1. Antioxidant Properties
IPA scavenges reactive oxygen species (ROS) with efficiency comparable to melatonin. Given that oligodendrocytes are exceptionally vulnerable to oxidative stress due to their high iron content and low antioxidant enzyme levels, the neuroprotective effects of IPA may be particularly relevant to myelin maintenance [75].
4.3.2. Pregnane X Receptor Activation
Beyond its antioxidant function, IPA activates the pregnane X receptor (PXR), a nuclear receptor that regulates xenobiotic metabolism and intestinal barrier function [148]. PXR activation enhances expression of tight junction proteins, potentially contributing to BBB integrity.
4.4. Clinical Relevance: Tryptophan Metabolism in MS
Multiple Sclerosis patients exhibit altered tryptophan metabolism characterized by:
These observations suggest that restoration of microbial tryptophan metabolism could represent a therapeutic strategy for promoting remyelination and suppressing neuroinflammation.
5. Secondary Bile Acids: Emerging Regulators of Remyelination
Secondary bile acids, produced by microbial transformation of host-derived primary bile acids, have recently emerged as potent signaling molecules with implications for oligodendrocyte biology.
5.1. Bile Acid Metabolism and the Microbiota
Primary bile acids (cholic acid and chenodeoxycholic acid) are synthesized in the liver from cholesterol and secreted into the intestine to facilitate lipid absorption. A fraction escapes enterohepatic recirculation and reaches the colon, where bacteria perform various modifications [23]:
- Deconjugation: Bile salt hydrolases remove glycine/taurine conjugates
- 7-dehydroxylation: Produces lithocholic acid (LCA) from chenodeoxycholic acid and deoxycholic acid (DCA) from cholic acid
- Epimerization and oxidation: Generate additional bile acid species
5.2. Bile Acid Receptors in the CNS
Secondary bile acids activate multiple receptors expressed in the brain:
5.2.1. TGR5 (GPBAR1)
TGR5 is a G-protein coupled receptor activated by LCA and DCA. In the CNS, TGR5 is expressed on microglia, astrocytes, and neurons [82]. TGR5 activation:
- Suppresses microglial inflammatory responses via cAMP elevation
- Reduces production of TNF- and IL-1
- Promotes neuroprotective microglial phenotypes
5.2.2. Vitamin D Receptor (VDR)
The vitamin D receptor, a nuclear hormone receptor, is activated by both 1,25-dihydroxyvitamin D3 and secondary bile acids, particularly LCA [83]. VDR is expressed in oligodendrocytes and OPCs, and its activation:
- Promotes OPC differentiation into mature oligodendrocytes
- Enhances expression of myelin genes
- Provides neuroprotection against inflammatory damage
5.3. Bile Acids and Remyelination
Emerging evidence suggests that secondary bile acids can promote remyelination:
- 1.
- Tauroursodeoxycholic acid (TUDCA) administration in EAE models reduces demyelination and enhances OPC differentiation [86].
- 2.
- Ursodeoxycholic acid (UDCA) is being evaluated in clinical trials for progressive MS based on its neuroprotective and immunomodulatory properties [87].
- 3.
- MS patients show altered bile acid profiles with reduced secondary bile acids, suggesting dysbiosis-driven deficiency [88].
6. Metabolic Support of Axonal Integrity and Bioenergetics
The relationship between oligodendrocytes and axons extends far beyond simple insulation. Oligodendrocytes are active metabolic partners that provide trophic support essential for axonal survival and function.
6.1. The Monocarboxylate Transporter Shuttle
Myelinated axons depend on oligodendrocyte-derived metabolic substrates for their energy needs. The monocarboxylate transporter (MCT) shuttle is the primary mechanism for this metabolic coupling [3,4]:
- 1.
- Oligodendrocytes take up glucose via GLUT1 or utilize glycogen stores
- 2.
- Glycolytic metabolism produces pyruvate, which is converted to lactate
- 3.
- MCT1 on the oligodendrocyte membrane exports lactate into the periaxonal space
- 4.
- Axonal MCT2 imports lactate for mitochondrial oxidation
Disruption of this shuttle—through MCT1 knockout or oligodendrocyte damage—leads to axonal degeneration even in the absence of demyelination [4].
6.2. Microbial Contribution to the Metabolic Axis
Microbial metabolites contribute to this bioenergetic relationship through multiple mechanisms:
6.2.1. Direct Fuel Provision
SCFAs, particularly acetate, can be directly oxidized by oligodendrocytes to generate ATP. Acetate enters the TCA cycle via acetyl-CoA, providing an alternative carbon source to glucose [89]. This may be particularly relevant during periods of high metabolic demand or when glucose availability is limited.
6.2.2. Support of Lipid Synthesis
The extraordinary lipid demands of myelination require substantial metabolic resources. Acetyl-CoA derived from microbial acetate contributes to:
- Cholesterol biosynthesis (the mevalonate pathway)
- Fatty acid synthesis (via malonyl-CoA)
- Sphingolipid production (ceramide and galactocerebroside)
Studies in germ-free mice show reduced expression of cholesterol biosynthesis genes in the prefrontal cortex, suggesting that microbial metabolites support myelin lipid production [17].
6.3. Mitochondrial Function and Oxidative Stress
Myelination is metabolically demanding and generates significant reactive oxygen species (ROS). Oligodendrocytes are particularly vulnerable to oxidative damage due to:
- High iron content (required for myelin synthesis enzymes)
- Low levels of antioxidant enzymes (catalase, superoxide dismutase)
- Abundant polyunsaturated fatty acids in myelin membranes
Recent evidence indicates that mitochondrial metabolic programming is not merely supportive but instructive for oligodendrocyte maturation. During the transition from pre-oligodendrocytes to mature oligodendrocytes, mitochondrial respiration increases alongside enhanced pyruvate dehydrogenase (Pdh) activity, promoting acetyl-CoA production and facilitating terminal differentiation and remyelination [155]. These findings suggest that intact mitochondrial flux is essential for effective oligodendrocyte lineage progression.
Microbial metabolites provide several layers of antioxidant protection:
By maintaining mitochondrial health and cellular redox balance, microbial metabolites ensure that OPCs retain the bioenergetic capacity required for the membrane expansion and protein synthesis of myelination.
7. Clinical Implications: Multiple Sclerosis and Demyelinating Disorders
Multiple Sclerosis (MS) affects approximately 2.8 million people worldwide and is characterized by inflammatory demyelination, neurodegeneration, and accumulating disability [91]. The recognition of microbiota-oligodendrocyte interactions has profound implications for understanding MS pathogenesis and developing new therapies.
7.1. Dysbiosis in Multiple Sclerosis
Multiple independent studies have documented characteristic alterations in the gut microbiome of MS patients:
Table 2.
Gut Microbiome Alterations in Multiple Sclerosis.
| Bacterial Taxon | Change in MS | Metabolic Impact |
|---|---|---|
| Faecalibacterium prausnitzii | Reduced butyrate | |
| Prevotella species | ↓ | Reduced propionate |
| Butyricimonas | ↓ | Reduced butyrate |
| Lactobacillus species | ↓ | Reduced indoles |
| Clostridium clusters IV/XIVa | ↓ | Reduced SCFAs/bile acids |
| Methanobrevibacter | ↑ | Altered fermentation |
| Akkermansia muciniphila | Variable | Context-dependent |
The consistent finding of reduced SCFA-producing bacteria suggests that MS patients experience a systemic deficiency in metabolites required for optimal oligodendrocyte function.
7.2. The Differentiation Block in Chronic Lesions
In chronic MS lesions, OPCs are often present in the lesion vicinity but fail to differentiate into mature myelinating oligodendrocytes—a phenomenon termed the “differentiation block” [92,93,94]. This failure of endogenous repair represents a major therapeutic challenge and has been attributed to multiple factors:
The deficiency of microbial-derived HDAC inhibitors and AhR ligands in MS patients may directly contribute to this differentiation block by failing to provide the epigenetic “priming” signals required for OPC maturation. Supporting this hypothesis, circulating SCFA levels negatively correlate with MS disease activity and disability progression [103,104].
The integrated actions of these metabolites, summarized in Figure 3, provide both epigenetic priming and anti-inflammatory signaling that collectively support remyelination.
7.3. Modulation of Neuroinflammation
Beyond direct effects on oligodendrocytes, microbial metabolites regulate myelination indirectly through modulation of CNS inflammation.
7.3.1. The Treg-Th17 Balance
The balance between regulatory T cells (Tregs) and pro-inflammatory Th17 cells is critical in MS pathogenesis. Th17 cells produce IL-17 and GM-CSF, which promote inflammatory demyelination, while Tregs suppress autoimmune responses [105,106].
Microbial SCFAs, particularly propionate, promote Treg differentiation through multiple mechanisms:
- 1.
- 2.
- GPR43 activation: SCFA signaling through GPR43 on T cells promotes Treg differentiation [63].
- 3.
Clinical evidence supports this mechanism: oral propionate supplementation in MS patients significantly increased circulating Treg frequency and function, correlating with reduced annualized relapse rate [66].
7.3.2. Microglial Polarization
Microglia can adopt pro-inflammatory (M1-like) or anti-inflammatory/pro-regenerative (M2-like) phenotypes, though this classification is increasingly recognized as oversimplified [110]. Similarly, astrocyte activation in demyelinating disease does not conform to a strict binary paradigm but instead represents a spectrum of context-dependent reactive states balancing protective and pathological functions [171]. Microbial metabolites promote microglial states conducive to remyelination:
Since activated microglia can directly damage oligodendrocytes through release of reactive oxygen species and inflammatory cytokines, microbial modulation of microglial states is critical for creating a permissive environment for remyelination.
7.4. Gut-Brain Axis in MS: Clinical Evidence
Several lines of clinical evidence support the relevance of the gut-brain axis in MS:

8. Experimental Models: Evidence from Germ-Free and Intervention Studies
The most compelling mechanistic evidence for microbial regulation of myelination comes from controlled experimental models that manipulate the microbiome or its metabolites. Recent morphometric analyses of optic nerves from germ-free (GF), gnotobiotic (OMM12), and conventionally colonized mice revealed significant alterations in axon diameter, fiber density, and g-ratio in young adult animals, supporting a direct role of gut microbiota in CNS myelination [5].
8.1. Germ-Free Mouse Studies
Germ-free (GF) mice, raised in sterile isolators without any microbial colonization, have proven invaluable for understanding microbiome-brain interactions.
8.1.1. Myelination Defects in GF Mice
GF mice exhibit multiple abnormalities in CNS myelination:
- 1.
- 2.
- Transcriptomic changes: RNA-seq reveals downregulation of cholesterol biosynthesis genes (Hmgcr, Fdft1, Sqle) and myelin structural genes [17]
- 3.
- Altered white matter volume: MRI studies show reduced white matter in prefrontal regions [115]
- 4.
- OPC differentiation defects: Reduced numbers of mature CC1+ oligodendrocytes in corpus callosum [18]
8.1.2. Reversibility and Critical Windows
Critically, many myelination defects in GF mice can be rescued by microbial colonization, but timing matters:
- Early colonization (weaning): Nearly complete rescue of myelin gene expression and white matter structure [17]
- Adult colonization: Partial rescue, suggesting a critical developmental window [18]
- SCFA supplementation alone: Substantial rescue even without live bacteria, demonstrating metabolite sufficiency [35]
These findings suggest that while the developing brain is most sensitive to microbial cues, some plasticity persists into adulthood—an encouraging observation for therapeutic applications.
8.2. Antibiotic Perturbation Studies
Antibiotic treatment provides a complementary approach to studying microbiome-myelination interactions in conventionally raised animals.
8.2.1. Effects of Antibiotic-Induced Dysbiosis
Broad-spectrum antibiotic treatment in adult mice produces:
8.2.2. Recovery Following Antibiotic Cessation
Upon antibiotic withdrawal, the gut microbiome gradually recovers, and microbial metabolite levels normalize over 2-4 weeks. Importantly, myelination markers also recover, demonstrating the dynamic responsiveness of oligodendrocytes to microbial signals [18]. This reversibility has important implications for understanding how transient dysbiosis (e.g., from illness or dietary changes) might temporarily impact CNS myelination.
8.3. Demyelination-Remyelination Models
Several experimental models allow direct assessment of microbial effects on remyelination following demyelinating injury.
8.3.1. Cuprizone Model
Cuprizone (bis-cyclohexanone-oxaldihydrazone) is a copper chelator that, when administered in the diet, causes selective oligodendrocyte death and demyelination, particularly in the corpus callosum [121]. Upon cuprizone withdrawal, spontaneous remyelination occurs over 2-4 weeks.
Studies using this model have demonstrated:
8.3.2. Experimental Autoimmune Encephalomyelitis (EAE)
EAE, induced by immunization with myelin antigens or adoptive transfer of myelin-reactive T cells, models the inflammatory demyelination of MS [122].
Microbiome interventions in EAE have shown:
Table 3.
Summary of Experimental Evidence Linking Microbiota to Myelination.
| Model System | Manipulation | Myelination Outcome | Key References |
|---|---|---|---|
| Germ-free mice | Absence of microbiota | Hypomyelination (PFC) | [17,18] |
| Antibiotic-treated | Broad-spectrum ABx | Reduced myelin genes | [18,35] |
| Cuprizone + GF | Demyelination in GF | Delayed remyelination | [119] |
| Cuprizone + butyrate | SCFA supplementation | Accelerated repair | [51] |
| EAE + GF | Autoimmune model | Reduced severity | [123,124] |
| EAE + propionate | SCFA supplementation | Reduced severity; ↑ Tregs | [65,66] |
| EAE + probiotics | Live bacteria | Reduced severity | [126,169] |
8.4. Colonization Studies with Defined Communities
Moving beyond GF versus conventionalized comparisons, researchers have begun using defined microbial communities to dissect specific bacterial contributions:
- Altered Schaedler Flora (ASF): An 8-member defined community that partially rescues GF phenotypes [128]
- Synthetic communities: Custom-designed consortia enriched in SCFA producers show enhanced myelination support [129]
- Monocolonization: Single-strain colonization identifies specific bacterial contributions [130]
These approaches are revealing that not all bacteria contribute equally to myelination support, with butyrate-producing Clostridia species appearing particularly important [131].
9. Therapeutic Frontiers: From Probiotics to Precision Postbiotics
The recognition of microbial metabolites as systemic regulators of myelination has catalyzed development of novel therapeutic strategies targeting the gut-brain axis.
9.1. Dietary Interventions
9.1.1. High-Fiber Diets
Dietary fiber provides the substrate for microbial SCFA production. Increasing fiber intake represents a simple, low-risk intervention with potential CNS benefits:
9.1.2. Ketogenic and Low-Carbohydrate Diets
Paradoxically, ketogenic diets—which are low in fiber—may also benefit myelination through alternative mechanisms:
9.1.3. Mediterranean Diet
The Mediterranean diet, rich in fiber, polyphenols, and omega-3 fatty acids, promotes a diverse, SCFA-producing microbiome:
9.2. Probiotic and Synbiotic Approaches
9.2.1. Traditional Probiotics
Administration of live beneficial bacteria aims to restore healthy microbiome composition:
Clinical trials of probiotics in MS have shown modest benefits on inflammatory markers and quality of life, though effects on myelination per se remain to be demonstrated [143].
9.2.2. Next-Generation Probiotics
Emerging approaches focus on bacteria specifically selected for their metabolite-producing capacity:
9.2.3. Synbiotics
Synbiotic formulations combine probiotics with prebiotic substrates to enhance bacterial survival and metabolite production:
9.3. Postbiotic Supplementation

9.3.1. SCFA Supplementation
Direct administration of SCFAs offers precise control over metabolite exposure:
- Oral butyrate: Sodium butyrate or tributyrin (a butyrate prodrug) can be administered orally; shows efficacy in demyelination models [51]
- Oral propionate: Clinical trials in MS show increased Tregs and reduced relapse rates [66]
- Challenges: Rapid metabolism limits systemic exposure; enteric coating or prodrug formulations improve bioavailability [28]
9.3.2. Indole Derivatives
Tryptophan metabolites represent another postbiotic avenue:
9.3.3. Bile Acid Therapeutics
Secondary bile acids and their derivatives are entering clinical development:
9.4. Fecal Microbiota Transplantation
Fecal microbiota transplantation (FMT), involving transfer of stool from healthy donors to patients, represents the most comprehensive microbiome intervention:
- Rationale: Restores entire microbial ecosystem including metabolite-producing capacity
- MS case reports: Anecdotal reports of improvement following FMT [152]
- Clinical trials: Phase I/II trials in MS are underway (ClinicalTrials.gov identifiers NCT03594487, NCT04203017)
- Challenges: Standardization, safety (infection risk), and donor selection remain obstacles [153]
9.5. Personalized Approaches and Biomarkers
The substantial inter-individual variation in gut microbiome composition suggests that personalized approaches may be necessary:
10. Critical Discussion and Future Perspectives
While the evidence linking microbial metabolites to oligodendrocyte biology is compelling, several critical questions remain to be addressed.
These findings are reinforced by recent ultrastructural studies of optic nerve fibers demonstrating hypermyelination and altered axon dimensions in germ-free and simplified microbiota models [5].
Beyond multiple sclerosis, converging evidence across neurodegenerative disorders underscores the central regulatory role of glial populations—including astrocytes, microglia, and oligodendrocytes—in shaping disease trajectories and therapeutic responsiveness [172].
10.1. Translational Challenges
10.1.1. Species Differences
Most mechanistic studies have been conducted in rodents, which differ from humans in:
Human studies measuring CNS metabolite concentrations (e.g., via magnetic resonance spectroscopy) and correlating them with myelin imaging metrics are needed.
10.1.2. Causality vs. Correlation
While dysbiosis is consistently observed in MS, establishing causality is challenging:
Longitudinal studies following at-risk individuals (e.g., those with clinically isolated syndrome) could help establish temporal relationships.
10.2. Mechanistic Gaps
10.2.1. Direct vs. Indirect Effects
The relative contribution of direct metabolite effects on oligodendrocytes versus indirect effects through microglia, astrocytes, and immune cells remains unclear. Cell-specific conditional knockouts of metabolite receptors could address this question.
10.2.2. Metabolite Synergies
Metabolites do not act in isolation. The combinatorial effects of SCFAs, indoles, and bile acids likely create a “metabolic milieu” whose properties exceed the sum of individual components. Systems biology approaches integrating multi-omics data are needed to capture these interactions [162].
10.2.3. Timing and Critical Windows
The observation that early-life colonization is more effective than adult colonization at rescuing GF myelination defects suggests critical developmental windows [17]. Key questions include:
- 1.
- Can these windows be “reopened” in adults through pharmacological or environmental interventions?
- 2.
- Are there distinct windows for initial myelination versus remyelination after injury?
- 3.
- How do early-life antibiotic exposures affect long-term myelination capacity?
10.3. Future Directions
10.3.1. Advanced Imaging
Integration of gut microbiome profiling with advanced neuroimaging could reveal brain-microbiome correlations:
10.3.2. Single-Cell Approaches
Single-cell RNA sequencing of the oligodendrocyte lineage following microbiome perturbations could reveal:
10.3.3. Organoid Systems
Human brain organoids offer new opportunities for studying microbiome-oligodendrocyte interactions:
10.3.4. Clinical Trial Design
Future clinical trials of microbiome-targeted interventions for MS should incorporate:
- Stratification by baseline microbiome composition
- Metabolomic endpoints (circulating SCFA, indole levels)
- Myelin-specific imaging outcomes
- Long-term follow-up for disability progression
11. Conclusions
The recognition that microbial metabolites serve as systemic regulators of oligodendrocyte lineage dynamics represents a paradigm shift in our understanding of CNS myelination. Through HDAC inhibition, GPCR signaling, AhR activation, and direct metabolic support, gut-derived molecules—particularly SCFAs, tryptophan derivatives, and secondary bile acids—coordinate the complex transcriptional and metabolic programs required for myelin synthesis and maintenance.
This integrated view of the gut-brain axis in myelination has profound implications for understanding demyelinating diseases such as Multiple Sclerosis. The characteristic dysbiosis observed in MS patients, with depletion of SCFA-producing bacteria and altered tryptophan metabolism, may directly contribute to the failure of endogenous remyelination that characterizes progressive disease.
Therapeutically, the gut-brain axis opens multiple intervention points. Dietary modifications, probiotics, and direct postbiotic supplementation all show promise for promoting remyelination and suppressing neuroinflammation. The ongoing clinical trials of propionate, UDCA, and FMT in MS patients will provide critical evidence for the translational potential of these approaches.
As we move toward precision medicine, integration of microbiome profiling with neuroimaging and clinical outcomes will enable personalized therapeutic strategies. The next decade promises to reveal the full therapeutic potential of harnessing the gut microbiota for CNS regeneration.

Abbreviations
| AhR | Aryl Hydrocarbon Receptor |
| BBB | Blood-Brain Barrier |
| CNS | Central Nervous System |
| DCA | Deoxycholic Acid |
| EAE | Experimental Autoimmune Encephalomyelitis |
| FMT | Fecal Microbiota Transplantation |
| GF | Germ-Free |
| GPCR | G-Protein Coupled Receptor |
| HDAC | Histone Deacetylase |
| IPA | Indole-3-Propionic Acid |
| LCA | Lithocholic Acid |
| MBP | Myelin Basic Protein |
| MCT | Monocarboxylate Transporter |
| MS | Multiple Sclerosis |
| OL | Oligodendrocyte |
| OPC | Oligodendrocyte Precursor Cell |
| PLP | Proteolipid Protein |
| SCFA | Short-Chain Fatty Acid |
| TGR5 | Takeda G-Protein Receptor 5 |
| Treg | Regulatory T Cell |
| VDR | Vitamin D Receptor |
References
- Nave, K.A. Myelination and support of axonal integrity by glia. Nature 2010, 468(7321), 244–252. [Google Scholar] [CrossRef]
- Simons, M.; Nave, K.A. Oligodendrocytes: Myelination and axonal support. Cold Spring Harbor Perspectives in Biology 2015, 8(1), a020479. [Google Scholar] [CrossRef]
- Fünfschilling, U.; et al. Glycolytic oligodendrocytes maintain myelin and long-term axonal integrity. Nature 2012, 485(7399), 517–521. [Google Scholar] [CrossRef]
- Lee, Y.; et al. Oligodendroglia metabolically support axons and contribute to neurodegeneration. Nature 2012, 487(7408), 443–448. [Google Scholar] [CrossRef]
- Ronchi, G.; Pellegrino, D. Gut microbiota regulates optic nerve fiber myelination. Frontiers in Cell and Developmental Biology 2025, 13, 1526855. [Google Scholar] [CrossRef]
- Kessaris, N.; et al. Competing waves of oligodendrocytes in the forebrain and postnatal elimination of an embryonic lineage. Nature Neuroscience 2006, 9(2), 173–179. [Google Scholar] [CrossRef]
- Richardson, W.D.; et al. Oligodendrocyte wars. Nature Reviews Neuroscience 2006, 7(1), 11–18. [Google Scholar] [CrossRef]
- Emery, B. Regulation of oligodendrocyte differentiation and myelination. Science 2010, 330(6005), 779–782. [Google Scholar] [CrossRef] [PubMed]
- Zuchero, J.B.; et al. CNS myelin wrapping is driven by actin disassembly. Developmental Cell 2015, 34(2), 152–167. [Google Scholar] [CrossRef] [PubMed]
- Fancy, S.P.; et al. Myelin regeneration: A recapitulation of development? Annual Review of Neuroscience 2011, 34, 21–43. [Google Scholar] [CrossRef] [PubMed]
- Mitew, S.; et al. Mechanisms regulating the development of oligodendrocytes and central nervous system myelin. Neuroscience 2014, 276, 29–47. [Google Scholar] [CrossRef] [PubMed]
- Sender, R.; et al. Revised estimates for the number of human and bacteria cells in the body. PLoS Biology 2016, 14(8), e1002533. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; et al. The microbiota-gut-brain axis. Physiological Reviews 2019, 99(4), 1877–2013. [Google Scholar] [CrossRef] [PubMed]
- Sharon, G.; et al. The central nervous system and the gut microbiome. Cell 2016, 167(4), 915–932. [Google Scholar] [CrossRef]
- Mayer, E.A.; et al. Gut microbes and the brain: Paradigm shift in neuroscience. Journal of Neuroscience 2015, 35(41), 13857–13867. [Google Scholar] [CrossRef]
- Dinan, T.G.; Cryan, J.F. Gut instincts: Microbiota as a key regulator of brain development, ageing and neurodegeneration. Journal of Physiology 2017, 595(2), 489–503. [Google Scholar] [CrossRef]
- Hoban, A.E.; et al. Regulation of prefrontal cortex myelination by the microbiota. Translational Psychiatry 2016, 6(4), e774. [Google Scholar] [CrossRef]
- Gacias, M.; et al. Microbiota-driven transcriptional changes in prefrontal cortex override genetic differences in social behavior. eLife 2016, 5, e13442. [Google Scholar] [CrossRef]
- Cummings, J.H.; et al. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987, 28(10), 1221–1227. [Google Scholar] [CrossRef]
- Dalile, B.; et al. The role of short-chain fatty acids in microbiota-gut-brain communication. Nature Reviews Gastroenterology & Hepatology 2019, 16(8), 461–478. [Google Scholar] [CrossRef]
- Agus, A.; et al. Gut microbiota regulation of tryptophan metabolism in health and disease. Cell Host & Microbe 2018, 23(6), 716–724. [Google Scholar] [CrossRef] [PubMed]
- Roager, H.M.; Licht, T.R. Microbial tryptophan catabolites in health and disease. Nature Communications 2018, 9(1), 3294. [Google Scholar] [CrossRef] [PubMed]
- Wahlström, A.; et al. Intestinal crosstalk between bile acids and microbiota and its impact on host metabolism. Cell Metabolism 2016, 24(1), 41–50. [Google Scholar] [CrossRef] [PubMed]
- MahmoudianDehkordi, S.; et al. Altered bile acid profile associates with cognitive impairment in Alzheimer’s disease - An emerging role for gut microbiome. Alzheimer’s & Dementia 2019, 15(1), 76–92. [Google Scholar] [CrossRef]
- Sivaprakasam, S.; et al. Short-chain fatty acid transporters: Role in colonic homeostasis. Comprehensive Physiology 2017, 8(1), 299–314. [Google Scholar] [CrossRef]
- Borthakur, A.; et al. Regulation of monocarboxylate transporter 1 (MCT1) promoter by butyrate in human intestinal epithelial cells. American Journal of Physiology-Gastrointestinal and Liver Physiology 2008, 294(1), G241–G247. [Google Scholar] [CrossRef]
- Bloemen, J.G.; et al. Short chain fatty acids exchange across the gut and liver in humans measured at surgery. Clinical Nutrition 2009, 28(6), 657–661. [Google Scholar] [CrossRef]
- Boets, E.; et al. Systemic availability and metabolism of colonic-derived short-chain fatty acids in healthy subjects. American Journal of Physiology-Gastrointestinal and Liver Physiology 2017, 312(1), G37–G47. [Google Scholar] [CrossRef]
- Wikoff, W.R.; et al. Metabolomics analysis reveals large effects of gut microflora on mammalian blood metabolites. Proceedings of the National Academy of Sciences 2009, 106(10), 3698–3703. [Google Scholar] [CrossRef]
- Dawson, P.A.; et al. Bile acid transporters. Journal of Lipid Research 2009, 50(12), 2340–2357. [Google Scholar] [CrossRef]
- Braniste, V.; et al. The gut microbiota influences blood-brain barrier permeability in mice. Science Translational Medicine 2014, 6(263), 263ra158. [Google Scholar] [CrossRef] [PubMed]
- MacFabe, D.F. Short-chain fatty acid fermentation products of the gut microbiome: Implications in autism spectrum disorders. Microbial Ecology in Health and Disease 2012, 23, 19260. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, M.; et al. Symbiotic bacterial metabolites regulate gastrointestinal barrier function via the xenobiotic sensor PXR and Toll-like receptor 4. Immunity 2014, 41(2), 296–310. [Google Scholar] [CrossRef] [PubMed]
- Mertens, K.L.; et al. Bile acid signaling pathways from the enterohepatic circulation to the central nervous system. Frontiers in Neuroscience 2017, 11, 617. [Google Scholar] [CrossRef]
- Erny, D.; et al. Host microbiota constantly control maturation and function of microglia in the CNS. Nature Neuroscience 2015, 18(7), 965–977. [Google Scholar] [CrossRef]
- Lampron, A.; et al. Inefficient clearance of myelin debris by microglia impairs remyelinating processes. Journal of Experimental Medicine 2015, 212(4), 481–495. [Google Scholar] [CrossRef]
- Lloyd, A.F.; et al. Central nervous system regeneration is driven by microglia necroptosis and repopulation. Nature Neuroscience 2019, 22(7), 1046–1052. [Google Scholar] [CrossRef]
- Miron, V.E.; et al. M2 microglia and macrophages drive oligodendrocyte differentiation during CNS remyelination. Nature Neuroscience 2013, 16(9), 1211–1218. [Google Scholar] [CrossRef]
- Rothhammer, V.; et al. Type I interferons and microbial metabolites of tryptophan modulate astrocyte activity and CNS inflammation via the AhR. Nature Medicine 2016, 22(6), 586–597. [Google Scholar] [CrossRef]
- Orthmann-Murphy, J.L.; et al. Gap junctions couple astrocytes and oligodendrocytes. Journal of Molecular Neuroscience 2011, 35(1), 101–116. [Google Scholar] [CrossRef]
- Tress, O.; et al. Panglial gap junctional communication is essential for maintenance of myelin in the CNS. Journal of Neuroscience 2012, 32(22), 7499–7518. [Google Scholar] [CrossRef] [PubMed]
- Louis, P.; et al. The gut microbiota, bacterial metabolites and colorectal cancer. Nature Reviews Microbiology 2014, 12(10), 661–672. [Google Scholar] [CrossRef] [PubMed]
- Reichardt, N.; et al. Phylogenetic distribution of three pathways for propionate production within the human gut microbiota. The ISME Journal 2014, 8(6), 1323–1335. [Google Scholar] [CrossRef] [PubMed]
- Louis, P.; Flint, H.J. Diversity, metabolism and microbial ecology of butyrate-producing bacteria from the human large intestine. FEMS Microbiology Letters 2010, 294(1), 1–8. [Google Scholar] [CrossRef]
- Vital, M.; et al. Revealing the bacterial butyrate synthesis pathways by analyzing (meta) genomic data. mBio 2014, 5(2), e00889-14. [Google Scholar] [CrossRef]
- Davie, J.R. Inhibition of histone deacetylase activity by butyrate. The Journal of Nutrition 2003, 133(7), 2485S–2493S. [Google Scholar] [CrossRef]
- Waldecker, M.; et al. Inhibition of histone-deacetylase activity by short-chain fatty acids and some polyphenol metabolites formed in the colon. The Journal of Nutritional Biochemistry 2008, 19(9), 587–593. [Google Scholar] [CrossRef]
- Shen, S.; et al. Age-dependent epigenetic control of differentiation inhibitors is critical for remyelination efficiency. Nature Neuroscience 2008, 11(9), 1024–1034. [Google Scholar] [CrossRef]
- Conway, G.D.; et al. Histone deacetylase activity is required for human oligodendrocyte progenitor differentiation. Glia 2012, 60(12), 1944–1953. [Google Scholar] [CrossRef]
- Shen, S.; et al. Histone modifications affect timing of oligodendrocyte progenitor differentiation in the developing rat brain. Journal of Cell Biology 2005, 169(4), 577–589. [Google Scholar] [CrossRef]
- Chen, T.; et al. Butyrate suppresses demyelination and enhances remyelination. Journal of Neuroinflammation 2019, 16(1), 165. [Google Scholar] [CrossRef]
- Brown, A.J.; et al. The orphan G protein-coupled receptors GPR41 and GPR43 are activated by propionate and other short chain carboxylic acids. Journal of Biological Chemistry 2003, 278(13), 11312–11319. [Google Scholar] [CrossRef] [PubMed]
- Le Poul, E.; et al. Functional characterization of human receptors for short chain fatty acids and their role in polymorphonuclear cell activation. Journal of Biological Chemistry 2003, 278(28), 25481–25489. [Google Scholar] [CrossRef] [PubMed]
- Kimura, I.; et al. Short-chain fatty acids and ketones directly regulate sympathetic nervous system via G protein-coupled receptor 41 (GPR41). Proceedings of the National Academy of Sciences 2011, 108(19), 8030–8035. [Google Scholar] [CrossRef] [PubMed]
- Thangaraju, M.; et al. GPR109A is a G-protein–coupled receptor for the bacterial fermentation product butyrate and functions as a tumor suppressor in colon. Cancer Research 2009, 69(7), 2826–2832. [Google Scholar] [CrossRef]
- Fu, S.P.; et al. Anti-inflammatory effects of BHBA in both in vivo and in vitro Parkinson’s disease models are mediated by GPR109A-dependent mechanisms. Journal of Neuroinflammation 2015, 12(1), 9. [Google Scholar] [CrossRef]
- Mews, P.; et al. Acetyl-CoA synthetase regulates histone acetylation and hippocampal memory. Nature 2017, 546(7658), 381–386. [Google Scholar] [CrossRef]
- Camargo, N.; et al. Oligodendroglial myelination requires astrocyte-derived lipids. PLoS Biology 2017, 15(5), e1002605. [Google Scholar] [CrossRef]
- Wellen, K.E.; et al. ATP-citrate lyase links cellular metabolism to histone acetylation. Science 2009, 324(5930), 1076–1080. [Google Scholar] [CrossRef]
- Pietrocola, F.; et al. Acetyl coenzyme A: A central metabolite and second messenger. Cell Metabolism 2015, 21(6), 805–821. [Google Scholar] [CrossRef]
- Saher, G.; et al. Cholesterol in myelin biogenesis and hypomyelinating disorders. Biochimica et Biophysica Acta 2011, 1811(12), 698–703. [Google Scholar] [CrossRef]
- Mews, P.; et al. Alcohol metabolism contributes to brain histone acetylation. Nature 2019, 574(7780), 717–721. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.M.; et al. The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis. Science 2013, 341(6145), 569–573. [Google Scholar] [CrossRef] [PubMed]
- Arpaia, N.; et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 2013, 504(7480), 451–455. [Google Scholar] [CrossRef] [PubMed]
- Haghikia, A.; et al. Dietary fatty acids directly impact central nervous system autoimmunity via the small intestine. Immunity 2015, 43(4), 817–829. [Google Scholar] [CrossRef]
- Duscha, A.; et al. Propionic acid shapes the multiple sclerosis disease course by an immunomodulatory mechanism. Cell 2020, 180(6), 1067–1080. [Google Scholar] [CrossRef]
- Gao, K.; et al. Tryptophan metabolism: A link between the gut microbiota and brain. Advances in Nutrition 2020, 11(3), 709–723. [Google Scholar] [CrossRef]
- Zelante, T.; et al. Tryptophan catabolites from microbiota engage aryl hydrocarbon receptor and balance mucosal reactivity via interleukin-22. Immunity 2013, 39(2), 372–385. [Google Scholar] [CrossRef]
- Williams, B.B.; et al. Discovery and characterization of gut microbiota decarboxylases that can produce the neurotransmitter tryptamine. Cell Host & Microbe 2014, 16(4), 495–503. [Google Scholar] [CrossRef]
- Gao, J.; et al. Impact of the gut microbiota on intestinal immunity mediated by tryptophan metabolism. Frontiers in Cellular and Infection Microbiology 2018, 8, 13. [Google Scholar] [CrossRef]
- Quintana, F.J.; Sherr, D.H. Aryl hydrocarbon receptor control of adaptive immunity. Pharmacological Reviews 2013, 65(4), 1148–1161. [Google Scholar] [CrossRef] [PubMed]
- Denison, M.S.; Nagy, S.R. Activation of the aryl hydrocarbon receptor by structurally diverse exogenous and endogenous chemicals. Annual Review of Pharmacology and Toxicology 2003, 43(1), 309–334. [Google Scholar] [CrossRef] [PubMed]
- Rothhammer, V.; et al. Microglial control of astrocytes in response to microbial metabolites. Nature 2018, 557(7707), 724–728. [Google Scholar] [CrossRef] [PubMed]
- Dodd, D.; et al. A gut bacterial pathway metabolizes aromatic amino acids into nine circulating metabolites. Nature 2017, 551(7682), 648–652. [Google Scholar] [CrossRef]
- Thorburne, S.K.; Bhagat, S. Low glutathione and high iron govern the susceptibility of oligodendroglial precursors to oxidative stress. Journal of Neurochemistry 1996, 67(3), 1014–1022. [Google Scholar] [CrossRef]
- Lim, C.K.; et al. Kynurenine pathway metabolomics predicts and provides mechanistic insight into multiple sclerosis progression. Scientific Reports 2017, 7(1), 41473. [Google Scholar] [CrossRef]
- Hartai, Z.; et al. Kynurenine metabolism in multiple sclerosis. Acta Neurologica Scandinavica 2005, 112(2), 93–96. [Google Scholar] [CrossRef]
- Chen, J.; et al. Multiple sclerosis patients have a distinct gut microbiota compared to healthy controls. Scientific Reports 2016, 6(1), 28484. [Google Scholar] [CrossRef]
- Jangi, S.; et al. Alterations of the human gut microbiome in multiple sclerosis. Nature Communications 2016, 7(1), 12015. [Google Scholar] [CrossRef]
- Ridlon, J.M.; et al. Bile salt biotransformations by human intestinal bacteria. Journal of Lipid Research 2006, 47(2), 241–259. [Google Scholar] [CrossRef]
- Gérard, P. Metabolism of cholesterol and bile acids by the gut microbiota. Pathogens 2013, 3(1), 14–24. [Google Scholar] [CrossRef] [PubMed]
- Keitel, V.; et al. The G-protein coupled bile salt receptor TGR5 is expressed in liver sinusoidal endothelial cells. Hepatology 2019, 70(3), 982–997. [Google Scholar] [CrossRef] [PubMed]
- Makishima, M.; et al. Vitamin D receptor as an intestinal bile acid sensor. Science 2002, 296(5571), 1313–1316. [Google Scholar] [CrossRef] [PubMed]
- Munger, K.L.; et al. Serum 25-hydroxyvitamin D levels and risk of multiple sclerosis. JAMA 2006, 296(23), 2832–2838. [Google Scholar] [CrossRef]
- Mokry, L.E.; et al. Vitamin D and risk of multiple sclerosis: A Mendelian randomization study. PLoS Medicine 2015, 12(8), e1001866. [Google Scholar] [CrossRef]
- Yanguas-Casás, N.; et al. TUDCA: An agonist of the bile acid receptor GPBAR1/TGR5 with anti-inflammatory effects in microglial cells. Journal of Cellular Physiology 2017, 232(8), 2231–2245. [Google Scholar] [CrossRef]
- Parry, G.J.; et al. Safety, tolerability, and cerebrospinal fluid penetration of ursodeoxycholic acid in patients with amyotrophic lateral sclerosis. Clinical Neuropharmacology 2010, 33(1), 17–21. [Google Scholar] [CrossRef]
- Bhargava, P.; et al. Bile acid metabolism is altered in multiple sclerosis and supplementation ameliorates neuroinflammation. Journal of Clinical Investigation 2020, 130(7), 3467–3482. [Google Scholar] [CrossRef]
- Moschen, A.R.; et al. Dietary factors: Major regulators of the gut’s microbiota. Gut & Liver 2012, 6(4), 411–416. [Google Scholar] [CrossRef]
- Dong, W.; et al. Sodium butyrate activates NRF2 to ameliorate diabetic nephropathy possibly via inhibition of HDAC. Journal of Endocrinology 2014, 223(1), 27–36. [Google Scholar] [CrossRef]
- Walton, C.; et al. Rising prevalence of multiple sclerosis worldwide: Insights from the Atlas of MS, third edition. Multiple Sclerosis Journal 2020, 26(14), 1816–1821. [Google Scholar] [CrossRef] [PubMed]
- Kuhlmann, T.; et al. Differentiation block of oligodendroglial progenitor cells as a cause for remyelination failure in chronic multiple sclerosis. Brain 2008, 131(7), 1749–1758. [Google Scholar] [CrossRef] [PubMed]
- Franklin, R.J. Why does remyelination fail in multiple sclerosis? Nature Reviews Neuroscience 2002, 3(9), 705–714. [Google Scholar] [CrossRef] [PubMed]
- Chang, A.; et al. Premyelinating oligodendrocytes in chronic lesions of multiple sclerosis. New England Journal of Medicine 2002, 346(3), 165–173. [Google Scholar] [CrossRef] [PubMed]
- Syed, Y.A.; et al. Inhibition of CNS remyelination by the presence of semaphorin 3A. Journal of Neuroscience 2011, 31(10), 3719–3728. [Google Scholar] [CrossRef]
- Fancy, S.P.; et al. Dysregulation of the Wnt pathway inhibits timely myelination and remyelination in the mammalian CNS. Genes & Development 2009, 23(13), 1571–1585. [Google Scholar] [CrossRef]
- Moyon, S.; et al. Demyelination causes adult CNS progenitors to revert to an immature state and express immune cues that support their migration. Journal of Neuroscience 2017, 37(5), 1197–1210. [Google Scholar] [CrossRef]
- Koreman, E.; et al. Chromatin remodeling and epigenetic regulation of oligodendrocyte myelination and myelin repair. Molecular and Cellular Neuroscience 2018, 87, 18–26. [Google Scholar] [CrossRef]
- Rone, M.B.; et al. Oligodendrogliopathy in multiple sclerosis: Low glycolytic metabolic rate promotes oligodendrocyte survival. Journal of Neuroscience 2016, 36(17), 4698–4707. [Google Scholar] [CrossRef]
- Schirmer, L.; et al. Neuronal vulnerability and multilineage diversity in multiple sclerosis. Nature 2019, 573(7772), 75–82. [Google Scholar] [CrossRef]
- Absinta, M.; et al. A lymphocyte–microglia–astrocyte axis in chronic active multiple sclerosis. Nature 2021, 597(7878), 709–714. [Google Scholar] [CrossRef] [PubMed]
- Lassmann, H. Multiple sclerosis pathology. Cold Spring Harbor Perspectives in Medicine 2018, 8(3), a028936. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; et al. Gut dysbiosis and lack of short chain fatty acids in a Chinese cohort of patients with multiple sclerosis. Neurochemistry International 2019, 129, 104468. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; et al. Short-chain fatty acids induce both effector and regulatory T cells by suppression of histone deacetylases. Mucosal Immunology 2015, 8(1), 80–93. [Google Scholar] [CrossRef]
- Korn, T.; et al. IL-17 and Th17 cells. Annual Review of Immunology 2009, 27, 485–517. [Google Scholar] [CrossRef]
- Kleinewietfeld, M.; et al. Sodium chloride drives autoimmune disease by the induction of pathogenic TH17 cells. Nature 2013, 496(7446), 518–522. [Google Scholar] [CrossRef]
- Furusawa, Y.; et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504(7480), 446–450. [Google Scholar] [CrossRef]
- Nastasi, C.; et al. The effect of short-chain fatty acids on human monocyte-derived dendritic cells. Scientific Reports 2015, 5(1), 16148. [Google Scholar] [CrossRef]
- Kaisar, M.M.; et al. Butyrate conditions human dendritic cells to prime type 1 regulatory T cells via both histone deacetylase inhibition and G protein-coupled receptor 109A signaling. Frontiers in Immunology 2017, 8, 1429. [Google Scholar] [CrossRef]
- Ransohoff, R.M. A polarizing question: Do M1 and M2 microglia exist? Nature Neuroscience 2016, 19(8), 987–991. [Google Scholar] [CrossRef]
- Wenzel, T.J.; et al. Short-chain fatty acids (SCFAs) alone or in combination regulate select immune functions of microglia-like cells. Molecular and Cellular Neuroscience 2020, 105, 103493. [Google Scholar] [CrossRef] [PubMed]
- Berer, K.; et al. Gut microbiota from multiple sclerosis patients enables spontaneous autoimmune encephalomyelitis in mice. Proceedings of the National Academy of Sciences 2017, 114(40), 10719–10724. [Google Scholar] [CrossRef]
- Cekanaviciute, E.; et al. Gut bacteria from multiple sclerosis patients modulate human T cells and exacerbate symptoms in mouse models. Proceedings of the National Academy of Sciences 2017, 114(40), 10713–10718. [Google Scholar] [CrossRef] [PubMed]
- Tremlett, H.; et al. Gut microbiota in early pediatric multiple sclerosis: A case-control study. European Journal of Neurology 2016, 23(8), 1308–1321. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; et al. Microbiota influence the development of the brain and behaviors in C57BL/6J mice. PLoS ONE 2018, 13(8), e0201829. [Google Scholar] [CrossRef]
- Thion, M.S.; et al. Microbiome influences prenatal and adult microglia in a sex-specific manner. Cell 2018, 172(3), 500–516. [Google Scholar] [CrossRef]
- Desbonnet, L.; et al. Gut microbiota depletion from early adolescence in mice: Implications for brain and behaviour. Brain, Behavior, and Immunity 2015, 48, 165–173. [Google Scholar] [CrossRef]
- Leclercq, S.; et al. Low-dose penicillin in early life induces long-term changes in murine gut microbiota, brain cytokines and behavior. Nature Communications 2017, 8(1), 15062. [Google Scholar] [CrossRef]
- McMurran, C.E.; et al. The microbiota regulates murine inflammatory responses to toxin-induced CNS demyelination but has minimal impact on remyelination. Proceedings of the National Academy of Sciences 2019, 116(50), 25311–25321. [Google Scholar] [CrossRef]
- Langley, M.R.; et al. High fat diet consumption results in mitochondrial dysfunction, oxidative stress, and oligodendrocyte loss in the central nervous system. Biochimica et Biophysica Acta-Molecular Basis of Disease 2020, 1866(3), 165630. [Google Scholar] [CrossRef]
- Matsushima, G.K.; Morell, P. The neurotoxicant, cuprizone, as a model to study demyelination and remyelination in the central nervous system. Brain Pathology 2001, 11(1), 107–116. [Google Scholar] [CrossRef]
- Constantinescu, C.S.; et al. Experimental autoimmune encephalomyelitis (EAE) as a model for multiple sclerosis (MS). British Journal of Pharmacology 2011, 164(4), 1079–1106. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.K.; et al. Proinflammatory T-cell responses to gut microbiota promote experimental autoimmune encephalomyelitis. Proceedings of the National Academy of Sciences 2011, 108 (Suppl 1), 4615–4622. [Google Scholar] [CrossRef] [PubMed]
- Berer, K.; et al. Commensal microbiota and myelin autoantigen cooperate to trigger autoimmune demyelination. Nature 2011, 479(7374), 538–541. [Google Scholar] [CrossRef] [PubMed]
- Ochoa-Repáraz, J.; et al. A polysaccharide from the human commensal Bacteroides fragilis protects against CNS demyelinating disease. Mucosal Immunology 2010, 3(5), 487–495. [Google Scholar] [CrossRef]
- Mangalam, A.; et al. Human gut-derived commensal bacteria suppress CNS inflammatory and demyelinating disease. Cell Reports 2017, 20(6), 1269–1277. [Google Scholar] [CrossRef]
- Mizuno, M.; et al. The dual role of short fatty acid chains in the pathogenesis of autoimmune disease models. PLoS ONE 2017, 12(2), e0173032. [Google Scholar] [CrossRef]
- Dewhirst, F.E.; et al. Phylogeny of the defined murine microbiota: Altered Schaedler flora. Applied and Environmental Microbiology 1999, 65(8), 3287–3292. [Google Scholar] [CrossRef]
- Faith, J.J.; et al. Identifying gut microbe-host phenotype relationships using combinatorial communities in gnotobiotic mice. Science Translational Medicine 2014, 6(220), 220ra11. [Google Scholar] [CrossRef]
- Tan, J.; et al. The role of short-chain fatty acids in health and disease. Advances in Immunology 2016, 121, 91–119. [Google Scholar] [CrossRef]
- Atarashi, K.; et al. Treg induction by a rationally selected mixture of Clostridia strains from the human microbiota. Nature 2013, 500(7461), 232–236. [Google Scholar] [CrossRef] [PubMed]
- So, D.; et al. Dietary fiber intervention on gut microbiota composition in healthy adults: A systematic review and meta-analysis. The American Journal of Clinical Nutrition 2018, 107(6), 965–983. [Google Scholar] [CrossRef] [PubMed]
- Baxter, N.T.; et al. Dynamics of human gut microbiota and short-chain fatty acids in response to dietary interventions with three fermentable fibers. mBio 2019, 10(1), e02566-18. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, K.C.; et al. Diet quality is associated with disability and symptom severity in multiple sclerosis. Neurology 2018, 90(1), e1–e11. [Google Scholar] [CrossRef]
- Kim, D.Y.; et al. Inflammation-mediated memory dysfunction and effects of a ketogenic diet in a murine model of multiple sclerosis. PLoS ONE 2012, 7(5), e35476. [Google Scholar] [CrossRef]
- Storoni, M.; Plant, G.T. The therapeutic potential of the ketogenic diet in treating progressive multiple sclerosis. Multiple Sclerosis International 2015, 2015, 681289. [Google Scholar] [CrossRef]
- Brenton, J.N.; et al. Phase II study of ketogenic diets in relapsing multiple sclerosis: Safety, tolerability and potential clinical benefits. Journal of Neurology, Neurosurgery & Psychiatry 2019, 93(6), 637–644. [Google Scholar] [CrossRef]
- Swidsinski, A.; et al. Reduced mass and diversity of the colonic microbiome in patients with multiple sclerosis and their improvement with ketogenic diet. Frontiers in Microbiology 2017, 8, 1141. [Google Scholar] [CrossRef]
- Katz Sand, I.; et al. The role of diet in multiple sclerosis: Mechanistic connections and current evidence. Current Nutrition Reports 2018, 7(3), 150–160. [Google Scholar] [CrossRef]
- De Filippis, F.; et al. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut 2016, 65(11), 1812–1821. [Google Scholar] [CrossRef]
- Tankou, S.K.; et al. A probiotic modulates the microbiome and immunity in multiple sclerosis. Annals of Neurology 2018, 83(6), 1147–1161. [Google Scholar] [CrossRef] [PubMed]
- Kouchaki, E.; et al. Clinical and metabolic response to probiotic supplementation in patients with multiple sclerosis: A randomized, double-blind, placebo-controlled trial. Clinical Nutrition 2017, 36(5), 1245–1249. [Google Scholar] [CrossRef] [PubMed]
- Salami, M.; et al. How probiotic bacteria influence the motor and mental behaviors as well as immunological and oxidative biomarkers in multiple sclerosis? A double blind clinical trial. Journal of Functional Foods 2019, 52, 8–13. [Google Scholar] [CrossRef]
- Mimee, M.; et al. Programming a human commensal bacterium, Bacteroides thetaiotaomicron, to sense and respond to stimuli in the murine gut microbiota. Cell Systems 2015, 1(1), 62–71. [Google Scholar] [CrossRef]
- Charbonneau, M.R.; et al. Developing a new class of engineered live bacterial therapeutics to treat human diseases. Nature Communications 2020, 11(1), 1738. [Google Scholar] [CrossRef]
- Depommier, C.; et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: A proof-of-concept exploratory study. Nature Medicine 2019, 25(7), 1096–1103. [Google Scholar] [CrossRef]
- Zmora, N.; et al. Personalized gut mucosal colonization resistance to empiric probiotics is associated with unique host and microbiome features. Cell 2018, 174(6), 1388–1405. [Google Scholar] [CrossRef]
- Venkatesh, M.; et al. Symbiotic bacterial metabolites regulate gastrointestinal barrier function via the xenobiotic sensor PXR and Toll-like receptor 4. Immunity 2014, 41(2), 296–310. [Google Scholar] [CrossRef]
- Konopelski, P.; et al. Indole-3-propionic acid, a tryptophan-derived bacterial metabolite, reduces weight gain in rats. Nutrients 2022, 14(7), 1507. [Google Scholar] [CrossRef]
- Busbee, P.B.; et al. Use of natural AhR ligands as potential therapeutic modalities against inflammatory disorders. Nutrition Reviews 2013, 71(6), 353–369. [Google Scholar] [CrossRef]
- Elia, A.E.; et al. Tauroursodeoxycholic acid in the treatment of patients with amyotrophic lateral sclerosis. European Journal of Neurology 2016, 23(1), 45–52. [Google Scholar] [CrossRef] [PubMed]
- Makkawi, S.; et al. Fecal microbiota transplantation associated with 10 years of stability in a patient with SPMS. Neurology-Neuroimmunology Neuroinflammation 2018, 5(4), e459. [Google Scholar] [CrossRef] [PubMed]
- Allegretti, J.R.; et al. The evolution of the use of faecal microbiota transplantation and emerging therapeutic indications. The Lancet 2019, 394(10196), 420–431. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M.; et al. Mapping human microbiome drug metabolism by gut bacteria and their genes. Nature 2019, 570(7762), 462–467. [Google Scholar] [CrossRef]
- Sajad, M.; et al. Pyruvate Dehydrogenase-Dependent Metabolic Programming Affects the Oligodendrocyte Maturation and Remyelination. Molecular Neurobiology 2024, 61, 397–410. [Google Scholar]
- Xiao, L.; et al. A catalog of the mouse gut metagenome. Nature Biotechnology 2015, 33(10), 1103–1108. [Google Scholar] [CrossRef]
- Ventura-Antunes, L.; et al. Different scaling of white matter volume, cortical connectivity, and gyrification across rodent and primate brains. Frontiers in Neuroanatomy 2013, 7, 3. [Google Scholar] [CrossRef]
- Miller, D.J.; et al. Prolonged myelination in human neocortical evolution. Proceedings of the National Academy of Sciences 2012, 109(41), 16480–16485. [Google Scholar] [CrossRef]
- Storm-Larsen, C.; et al. Gut microbiota composition during a 12-week intervention with delayed-release dimethyl fumarate in multiple sclerosis. Multiple Sclerosis and Related Disorders 2020, 39, 101898. [Google Scholar] [CrossRef]
- Cantarel, B.L.; et al. Gut microbiota in multiple sclerosis: Possible influence of immunomodulators. Journal of Investigative Medicine 2015, 63(5), 729–734. [Google Scholar] [CrossRef]
- Wunsch, M.; et al. The enteric nervous system is a potential autoimmune target in multiple sclerosis. Acta Neuropathologica 2022, 144(5), 871–888. [Google Scholar] [CrossRef]
- Shoaie, S.; et al. Quantifying diet-induced metabolic changes of the human gut microbiome. Cell Metabolism 2015, 22(2), 320–331. [Google Scholar] [CrossRef] [PubMed]
- MacKay, A.L.; Laule, C. Magnetic resonance of myelin water: An in vivo marker for myelin. Brain Plasticity 2016, 2(1), 71–91. [Google Scholar] [CrossRef] [PubMed]
- Alexander, A.L.; et al. Diffusion tensor imaging of the brain. Neurotherapeutics 2007, 4(3), 316–329. [Google Scholar] [CrossRef] [PubMed]
- Marques, S.; et al. Oligodendrocyte heterogeneity in the mouse juvenile and adult central nervous system. Science 2016, 352(6291), 1326–1329. [Google Scholar] [CrossRef]
- Jäkel, S.; et al. Altered human oligodendrocyte heterogeneity in multiple sclerosis. Nature 2019, 566(7745), 543–547. [Google Scholar] [CrossRef]
- Madhavan, M.; et al. Induction of myelinating oligodendrocytes in human cortical spheroids. Nature Methods 2018, 15(9), 700–706. [Google Scholar] [CrossRef]
- Marton, R.M.; et al. Differentiation and maturation of oligodendrocytes in human three-dimensional neural cultures. Nature Neuroscience 2019, 22(3), 484–491. [Google Scholar] [CrossRef]
- Lavasani, S.; et al. A novel probiotic mixture exerts a therapeutic effect on experimental autoimmune encephalomyelitis mediated by IL-10 producing regulatory T cells. PLoS ONE 2010, 5(2), e9009. [Google Scholar] [CrossRef]
- Stacho, R.; Zucha, D.; Kirdajova, D.; Valihrach, L. Applications of Spatial Transcriptomics in Ischemic Stroke Research. American Journal of Pathology 2026. [Google Scholar] [CrossRef]
- Abulaban, A.A.; Al-Kuraishy, H.M.; et al. The Janus Face of Astrocytes in Multiple Sclerosis: Balancing Protection and Pathology. Brain Research Bulletin 2025, 226, 111356. [Google Scholar] [CrossRef]
- Jubair, H. Glial Cells in Dementia: From Cellular Dysfunction to Therapeutic Frontiers. Cell Transplantation 2026, 35, 09636897251414216. [Google Scholar] [CrossRef]
- Deng, X.; Zhao, M.; et al. New insights into acute ischemic stroke from the perspective of spatial omics. Theranostics 2025, 15(15), 7902–7924. [Google Scholar] [CrossRef]
Figure 1.
Transport mechanisms and cellular targets of microbial metabolites in the CNS. Butyrate, acetate, indoles, and bile acids cross the blood-brain barrier via specific transporters (MCT1, OATP) or passive diffusion. Once in the CNS parenchyma, these metabolites act on multiple glial populations: microglia, astrocytes, and oligodendrocyte precursor cells (OPCs)/oligodendrocytes (OLs). This multi-cellular targeting enables coordinated regulation of myelination through direct and indirect signaling pathways.
Figure 1.
Transport mechanisms and cellular targets of microbial metabolites in the CNS. Butyrate, acetate, indoles, and bile acids cross the blood-brain barrier via specific transporters (MCT1, OATP) or passive diffusion. Once in the CNS parenchyma, these metabolites act on multiple glial populations: microglia, astrocytes, and oligodendrocyte precursor cells (OPCs)/oligodendrocytes (OLs). This multi-cellular targeting enables coordinated regulation of myelination through direct and indirect signaling pathways.

Figure 2.
Metabolic fate of microbial acetate in oligodendrocytes. Acetate is converted to acetyl-CoA by acetyl-CoA synthetases (AceCS1/2). This central metabolite serves three key functions: (1) entry into the TCA cycle for ATP production; (2) provision of acetyl groups for histone acetylation via histone acetyltransferases (HATs), regulating gene expression; and (3) serving as a precursor for lipid biosynthesis, including myelin membrane components. This multi-faceted role positions acetate as a critical link between microbial metabolism and oligodendrocyte function.
Figure 2.
Metabolic fate of microbial acetate in oligodendrocytes. Acetate is converted to acetyl-CoA by acetyl-CoA synthetases (AceCS1/2). This central metabolite serves three key functions: (1) entry into the TCA cycle for ATP production; (2) provision of acetyl groups for histone acetylation via histone acetyltransferases (HATs), regulating gene expression; and (3) serving as a precursor for lipid biosynthesis, including myelin membrane components. This multi-faceted role positions acetate as a critical link between microbial metabolism and oligodendrocyte function.

Figure 3.
Integrated mechanisms by which microbial metabolites promote myelination. SCFAs (particularly butyrate) inhibit histone deacetylases (HDACs), leading to epigenetic priming of OPCs for differentiation. Indoles activate the aryl hydrocarbon receptor (AhR), suppressing neuroinflammation. Bile acids signal through receptors such as VDR and TGR5, promoting anti-inflammatory and pro-differentiation pathways. Together, these coordinated actions enhance OPC differentiation, reduce inflammation, and promote both developmental myelination and remyelination after injury.
Figure 3.
Integrated mechanisms by which microbial metabolites promote myelination. SCFAs (particularly butyrate) inhibit histone deacetylases (HDACs), leading to epigenetic priming of OPCs for differentiation. Indoles activate the aryl hydrocarbon receptor (AhR), suppressing neuroinflammation. Bile acids signal through receptors such as VDR and TGR5, promoting anti-inflammatory and pro-differentiation pathways. Together, these coordinated actions enhance OPC differentiation, reduce inflammation, and promote both developmental myelination and remyelination after injury.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



