Submitted:
20 February 2026
Posted:
27 February 2026
You are already at the latest version
Abstract
Aging is a fundamental biological process characterized by morphological and functional decline ultimately leading to death. Current research in aging is directed toward extending both healthspan and lifespan by elucidating the molecular and cellular mechanisms that drive aging and by developing interventions capable of delaying, preventing, or reversing age-associated physiological decline and multimorbidity. In this chapter, we take a broader view beyond the healthspan and lifespan of individuals, to consider deep issues impacting the duration and nature of our embodiment, including the nature of change, the meaning of personal persistence, and the future of humanity at multiple scales. If you don’t change, you die out (or become irrelevant); but if you change, are you still present? We argue that aging, like traumatic injury and cancer, is a fundamental challenge to an embodied mind seeking to maintain its distinct nature, differentiated from the environment. Understanding aging thus must take place within the context of a broader story of how biological individuals come to exist, how they continue to exist despite continual challenge, and how their plasticity can be leveraged for transformative change beyond mere persistence. Here, we will present our aging framework grounded in the collective intelligence of cells, then we will discuss the implication for the human- and the species-level aspects of artificial chimerism and its corollary - multiscale (non-Darwinian) evolution. We conclude with some important open questions for humanity with respect to the implications of rejuvenation and longevity technologies.
Keywords:
aging
; longevity
; cognition
; goal-directedness
; futurism
; transhumanism
; regenerative medicine
1. Introduction
Aging is a fundamental biological process characterized by morphological and functional decline ultimately leading to death (de Magalhães, 2011, Austad and Hoffman, 2018, Gladyshev, 2016). Several age-related diseases can develop with time in humans, including cardiovascular diseases, cancer, neurodegeneration, diabetes, and immune-related conditions, altering deeply the quality of life (Hayflick, 2007, Lopez-Otin et al., 2023). Beyond individual pathophysiology, the demographic shift toward an aging population imposes a substantial burden on societies, global healthcare infrastructure and economic stability (Restrepo and Rozental, 1994, Tang et al., 2022).
Current research in aging is directed toward extending both healthspan and lifespan by elucidating the molecular and cellular mechanisms that drive aging and by developing interventions capable of delaying, preventing, or reversing age-associated physiological decline and multimorbidity (de Magalhães, 2011, De Magalhães and Ocampo, 2022, Lopez-Otin et al., 2023, Onken and Driscoll, 2010). The field has witnessed a few successes, indeed for example, partial epigenetic reprogramming has shown its ability to safely reset epigenetic clocks and improve tissue function in mice without stripping cell identity (Browder et al., 2022). Partial epigenetic reprogramming has been successfully applied in non-human primate models to restore visual function, marking a significant translational milestone for cellular rejuvenation therapies (Ksander et al., 2023). Epigenetic clocks have also been developed that allow evaluation of biological age through the analysis of DNA methylation patterns that accumulate over time and regulate gene expression. These clocks estimate biological age by measuring age-related modifications to DNA, particularly the accumulation of methyl groups at CpG regions of DNA (Duan et al., 2022). However, despite hopeful results, no actual organismal rejuvenation in mammals has been demonstrated and it remains the next frontier in the field.
Might radical life extension be impossible? The only things truly impossible are forbidden by physics; perhaps entropy or other physical limitations of cellular materials cap human potential? Our perspective begins with discussion of an existence proof that shows this not to be the case: planarian flatworms(Cebrià et al., 2018), a bilaterian similar to our ancestors. Asexual planaria resist injury (regenerating entire worms from fragments) and cancer. They are also effectively immortal, having no evidence of organismal aging (individual cells senesce and die but are regenerated). They exhibit many behaviors and have a true centralized brain that can learn (Pagán, 2014, Sarnat, 1985, Jacobson, 1963), and even retain that memory through complete regeneration of the brain(Mitchell et al., 1975, Best and Rubenstein, 1962, Corning, 1966, Corning, 1967, Blackiston et al., 2015). We don’t know what will happen to the memories and personality of the inevitable human patients whose brain tissue gets partially replaced by stem cells or newly-regenerated cells for degenerative disease, but planaria are showing us that it is possible for a complex being with a nervous system to live essentially forever, in both mind and body. If there is no physical reason why it can’t be done, the rest is engineering. But the planarian teaches us more than the possibility of eternal renewal. Because it achieves this via continuous turnover of its cells, and despite a very noisy (mixoploid!) genome (reviewed in (Fields et al., 2020)), it is also telling us something important about the nature of persistence, change, and the relationship of the genetically-specified hardware to the functional physiological intelligence that supervenes on it.
In this chapter, we take a broader view here beyond healthspan and lifespan, to ask about persistence and humanity at multiple scales, and what it means to be healthy in a persistent and disease-free human body. If we can resolve morphogenetic failures, what about cognitive ones? Can we stay psychologically healthy while accumulating hundreds of years and experience? Does the long-term future of longevity mean retaining our current bodies, with all of their susceptibilities and limitations, and when does longevity shift from maintaining status quo to accommodating intentional change? What is it to be human if we can fuse with technology and/or AI and how do we remain the same through change?
This is the paradox of change: if you don’t change, you die out (or become irrelevant); if you do change, do you still exist? All of these questions must be considered within the context of the developmental and regenerative processes that build a complex body out of active components (cells) which have degrees of autonomy and their own ancient agendas (Lyon, 2015, Lyon, 2006). Indeed, our bodies are a multi-scale agential material which implements numerous competencies to build and maintain complex form despite a myriad of challenges (Hartl and Levin, 2025, Levin, 2023b, Harris, 2018, Rubin, 2007, Rubin, 2006, Rubin et al., 1996, Levin, 2019). The ubiquitous ability of living tissues to form memories, pursue setpoints, re-interpret exogenous cues and their own genetic information, and improvise solutions to novel scenarios, comprise a key target for reparative, longevity, and enhancement strategies. The reprogrammability of biology at all scales offers a powerful way to achieve complex control over system-level properties far beyond micromanaging the molecular hardware of life.
Thus, we argue that roadmaps to longevity must exploit the native biological capacity to continuously meet large-scale anatomical target morphologies – regeneration (Levin, 2024a). Moreover, because of the deep symmetries between the autopoietic processes of the mind and body that we highlight, disruptive longevity technologies will not be found purely within the province of chemistry or even physiology, but also within cybernetics and cognitive science, using novel frameworks to communicate complex goals to molecular, cell, and organ-level systems in vivo (Corlett and Fletcher, 2014, Pio-Lopez et al., 2022).
In this chapter, we present our aging framework grounded in the collective intelligence of cells and the multi-competency architecture (Pio-Lopez et al., 2025, Pio-Lopez and Levin, 2024). We discuss the exploration of the space of possible bodies and minds facilitated by anti-aging treatments and their natural extensions; control of regeneration (or more broadly, competency of navigation in anatomical and physiological spaces) is the key to radical longevity within standard human embodiments and far beyond. We also touch on the implication for the human and the species of artificial chimerism and its corollary the multiscale (non-Darwinian) evolution. We will then conclude on the important open questions for humanity of rejuvenation and longevity technologies.
2. The Ship of Theseus: Aging in the Broader Context of a Life Cycle of Embodied Minds
The phenomenon of aging, and the quest for longevity, both fundamentally rest on the question of what we are. What is it that ages and prematurely “dies”, and what does it mean to persist for longer (or, forever)? The standard story of reproduction, birth, adulthood, and death obscures some key aspects of our life history. To understand aging and death, and to develop effective management of this process, we must look more closely at our origins as a distinct being which has the possibility of persistence with an environment.
The chief lesson of developmental biology is that we are the result of a continuous process that offers no sharp distinctions between body and mind. Each of us begins as an unfertilized oocyte, and slowly and gradually, with no magic lightning bolt that separates “mere physics” from “cognition”, traverses all of the disciplines. Despite our discrete funding agencies, journals, and departments, the same material transitions gradually from being the province of chemistry, through developmental physiology, behavioral science, and eventually psychology and psychoanalysis (Figure 1A). It can also take detours into oncology and bioengineering.
What is it that we call “1 embryo”, when it is clearly composed of millions of individual cells? An “embryo” refers to successful alignment. Both physically (Gray et al., 2011) and in terms of the region of anatomical space all of the cells commit to reach together, an “embryo” is a shared, biophysically-encoded model of morphogenetic setpoints that a cellular collective will pursue (Levin, 2019, Levin, 2022). Crucially, this is not, as is often taught, merely a feed-forward process of local rules that eventually lead to an “emergent” complex outcome. Those kinds of cellular-automata-like models, describing the lowest rung of the cybernetic hierarchy, do not fit the extensive data showing the ability of cellular collectives working in novel ways to achieve specific outcomes despite perturbations and barriers (McMillen and Levin, 2024, Levin, 2023b). Instead, morphogenesis involves homeostatic, homeodynamic, and allostatic processes that have setpoints (i.e., anatomical goal states toward which cell groups continually reduce error). In other words, the body arises as the result of the behavior of a swarm intelligence in which cells cooperate, navigating the space of anatomical possibilities toward specific outcomes (at which point, large-scale transformation shifts toward maintenance).
An embryonic blastoderm can be experimentally partitioned into regions, each of which will form “an embryo” (Lutz, 1949), when the cells of an island do not feel the presence of others. What we call “an embryo” arises from a computational medium which can self-organize into some number of Individuals (Figure 1B), all of which cooperate to reach their anatomical goal states. The material can self-scale (Rahbaran et al., 2021), as is seen in monozygotic twins, triplets, etc. (not half-bodies) that form when early embryos split; there are anywhere from 0 to half a dozen or more potential beings in typical embryonic blastoderms. Thus, the question of “how many individuals are there” in a given body is not set by the genetics, but by the real-time autopoietic (Maturana and Varela, 1980) processes of morphogenesis that establish boundaries between an individual and the outside world. That boundary, or a system’s “cognitive light cone” (Levin, 2019), demarcates the region of spacetime committed to specific goal states: in other words, the boundary of cells which are minimizing error toward an anatomical specification or the spatiotemporal size of their setpoint. That directed construction process can overcome not only surgical splitting but also merging, rearranging, genomic modification, hybridization, and a plethora of remarkably disruptive interventions at many scales (discussed in detail in (Nanos and Levin, 2022, Clawson and Levin, 2023, Levin, 2023b)), in its continuous efforts to minimize discrepancies between current state and encoded setpoints.
Two important consequences for aging and longevity result. First, the process that produced a specific body – the one we wish to keep from degeneration during aging – is an active, dynamic construction. Second, development is not a unique stage, irrelevant to adulthood: it is the first example of a universal process for most biological bodies – regeneration as homeostasis and error correction.
A cellular collective is continuously maintaining alignment over its parts (Rubin, 2007, Rubin, 2006, Rubin et al., 1996). While individual cells expertly navigate physiological and metabolic state spaces (Fields and Levin, 2022), and their molecular networks navigate transcriptional spaces, each level is constantly bending the option space of their parts to achieve their scale-specific outcomes. A classical experiment is instructive here (Figure 2A): in a grafted appendage, cells distal to the join site remodel into a new structure even though there is no local damage. The molecular biology of those cells is modified by a large-scale, high-level abstract setpoint (the overall bodyplan) which no cell or molecular network can comprehend. This is the main principle on which our bodies are built, and it is seen in cognitive science as well: the flow of ions across our muscle membranes is modified by abstract, high-level cognitive goals that drive voluntary locomotion (Figure 2B). In other words, our bodies comprise a multi-scale system that transduces high-level setpoints into the activity of cells and physiological circuits. This in turn opens opportunities for biomedical interventions that learn to communicate with these intermediate levels (Figure 2C) to manipulate the system at levels higher than the current molecular biology targets on which almost everything in biomedicine is focused today.
Homeostasis is not just for restoring after massive perturbations. It occurs at every level, working against the daily wear-and-tear of the body; it takes a highly active and dynamic process just to persist (“remain in one place” in Anatomical morphospace). For example, if neural inputs are interrupted, disorganization of tissue (Takeda et al., 1996, Nagato et al., 1995) and cancer (Outzen et al., 1976, Pawlowski and Weddell, 1967, Kaminishi et al., 1997), result. The mediator of maintenance cues, enabling cell groups to determine the direction of adjustment (error-reduction activities) is not always the nervous system, but it is often accomplished by very similar bioelectric processes occurring via non-excitable cells (Bates, 2015, Harris, 2021, Levin, 2021a). In other words, everything is fundamentally regeneration (which is, fundamentally, error minimization with respect to a stored setpoint). Development is just regeneration from 1 cell. Adult maintenance is small-scale regeneration against dying and damaged cells. Even cancer is, fundamentally, a problem of cells disconnecting from the collective’s large-scale network and its organ maintenance setpoint and reversion to individual cell-level goal states (Rubin, 1985, Moore et al., 2017).
The price of being a complex, multi-scale being in a world ruled by increasing entropy at the base layer, is the eternal vigilance required to reinforce a large-scale setpoint within an unreliable medium, always in danger of degradation of both hardware and information. This has important implications beyond the plasticity of the body – it is also the basis of our cognitive flexibility (Figure 3). We exist in the NOW moment between past instances of experience, frozen as engrams within our brain and body(Levin, 2024b), and the future toward which we actively, improvisationally, interpret and edit those memories in the continuous regeneration and metamorphosis of a self- and world-model (Conway, 2005, Conway and Pleydell-Pearce, 2000). In other words, just like our bodies are a dynamic process of self-construction, maintenance, and various degrees of metamorphosis, our cognitive Self is a self-constructing story which strikes a kind of critical balance between persistence-as-is (maintenance) and adaptation toward anticipated needs (Tschantz et al., 2022, Sennesh et al., 2022, Deans, 2021, Solms, 2023). Just as body morphogenesis adapts to perturbations to navigate anatomical space, we use physiological networks to navigate the 3D space of behavior. It is very likely that our cognitive architecture, and its immense problem-solving plasticity, is an evolutionary adaptation of the same mechanisms and algorithms first used for managing the top-down control of multicellular bodies (Fields et al., 2020). The symmetry between the autopoiesis of mind and body has important implications for both, the biomedicine of augmenting regenerative repair over long time periods, and the social and philosophical questions around extremely long-lived minds.
Under radical uncertainty at the lower levels, our unified body and mind systems (Solms, 2023, Johnson et al., 2022, Ciaunica, 2024) evolved to maintain healthy large-scale structure built over the shifting sands of micro-level degradation and noise. Thus, what evolution builds are not fixed solutions to static problems, but problem-solving systems with adaptive plasticity toward future opportunities(Hartl and Levin, 2025). The morphogenetic process lying between genotype and phenotype is not merely complex or pleiotropic, it uses processes well-studied by cybernetics and behavioral sciences to accommodate novel scenarios and meet their goals or develop novel ones (Davies and Levin, 2023). The framework of a directed, homeostatic heterarchy of processes that continuously interact to implement a self- and world-model, at both cognitive and somatic levels, is essential background for thinking about longevity and evaluating the relevance of theories of aging based on noise, as well as opening the space of possible intervention targets. The symmetry and continuum between mind and body is also important, as we think about the consequences of anti-aging treatments. What evolution makes are problem-solving agents – improvising systems evolved for robustness over a fundamentally unreliable medium – which has major implications for addressing aging as a problem of cognitive decline of the somatic intelligence, just as cancer can be seen as a dissociative disorder of that intelligence (Levin, 2021b, Wolsky, 1978, Tsonis, 1987).
3. The Collective Intelligence of the Body: Implications for Aging and Rejuvenation
Classical data have shown that living bodies can be remarkably resilient to massive perturbation as well as progressive entropic challenge (Figure 4). Recent results at the intersection of the diverse intelligence and regenerative biophysics field have shown (to date, in non-human model systems) that relatively low-complexity inputs can induce the formation of whole organs, the regeneration of limbs, the repair of complex birth defects, the normalization of cancer, and the self-assembly of functional biobots (reviewed in (Levin, 2021a)). All of this has been done in the context of native genomes, not requiring cell reprogramming, genomic editing, or exogenous scaffolds, but via control of physiological networks that store and process anatomical information.
Importantly for longevity research, which is a system-wide phenomenon, the repair properties of the body extend beyond anatomical morphospace. For example, they can solve challenges spanning physiological and transcriptional spaces. When flatworms are exposed to barium, a non-specific potassium channel blocker and stressor absent from their evolutionary history, ion flows are disrupted which triggers the rapid degeneration of planarian heads. However, they quickly grow replacement head structures that are entirely resistant to barium by adapting the expression of just a handful of genes to compensate for the inability to pass K+ ions (Emmons-Bell et al., 2019). In other words, out of ~20K transcriptional effectors, the cells rapidly identified precisely which genes would solve their toxic stress and repair their body. This phenomenon reveals intrinsic problem-solving capabilities, still unmatched by our AI technologies, that enable organisms to reconfigure their genetic activity profiles when confronting unprecedented environmental conditions.
We next discuss existing theories of aging, in the context of our view of bodies as fundamentally dynamic goal-seeking systems that work to implement specific form and function in the face of a wide range of challenges. We can categorize aging theories into two groups based on the proposed ultimate cause of aging: damage-based theories (Gladyshev et al., 2021, Hayflick, 2007, Lopez-Otin et al., 2023) and programmatic ones (de Magalhães, 2011, de Magalhaes, 2023, Kirkwood and Melov, 2011).
Damage-based theories hold that the cause of aging has to be found in entropy leading to the accumulated molecular damage to vital biological components—such as DNA, telomeres, mitochondria, or proteins—due to inherent repair system limitations (Gladyshev et al., 2021, Hayflick, 2007, Lopez-Otin et al., 2023). In other words, aging originates in the inevitability of rising disorder in physics. We find the DNA damage (Kirkwood, 1989, Medawar, 1952), telomere (Razgonova et al., 2020), protein damage (Bratic and Larsson, 2013), and the deleteriome (integrating genetic and cellular damages) (Gladyshev, 2016) theories in this framework for example. Current antiaging therapeutic approaches predominantly come from the perspective of damage-based theories, with a notable focus on epigenetic reprogramming techniques employing Yamanaka factors and their variants (De Magalhães and Ocampo, 2022).
Programmatic theories, in contrast, suggest that aging is genetically or evolutionary programmed, or quasi-programmed, due to ecological-based constraints and not merely due to entropic forces (de Magalhaes, 2023, Gems, 2022, Skulachev and Skulachev, 2014). In other words, aging could be avoidable by biological systems, but specific evolutionary trade-offs have actively selected for it. Under this category, we find the antagonistic pleiotropy (genes regulating growth and development cause aging later in life) (Austad and Hoffman, 2018, Bartke et al., 2013), software design flaw (aging stems from intrinsic flaws or bugs in the genetic “software” (the developmental program encoded in DNA) that continues running via epigenetic changes throughout life ) (de Magalhaes, 2023), group selected-adaptation (aging evolved to control or the death rate to stabilize population dynamics) (Mitteldorf, 2017) and the pathogen-control (aging evolved to limit the spread pathogens in a population) (Lidsky and Andino, 2020, Lidsky and Andino, 2022, Lidsky et al., 2023, Lidsky et al., 2022) theories.
We add here a third type of aging theory grounded in biological systems seen as collective intelligences (Pio-Lopez et al., 2025, Pio-Lopez and Levin, 2024), which is compatible with entropic forces and evolutionary modulation of commitment to repair, but introduces a novel cause of aging that cannot be reduced to either one: dissociative disorders of goal-seeking systems (Corlett and Fletcher, 2014, Montague et al., 2012). This theory is based on the emerging field of diverse intelligence and basal cognition (Baluška and Levin, 2016, Lyon, 2006, Lyon, 2015), which uses cybernetic and behavior science tools (Pezzulo and Levin, 2016, Pezzulo and Levin, 2015, Friston et al., 2015, Rosenblueth et al., 1943) to understand goal-seeking systems across multiple biological scales besides the conventional organism scale — upward to swarms and ecosystems, and downward to sub-organismal levels such as cells, tissues, and organs (Levin, 2022, Pezzulo and Levin, 2016, Levin, 2023c).
Examples include competencies of bacteria and biofilms (Prindle et al., 2015, Martinez-Corral et al., 2019, Ben-Jacob, 2009, Lyon, 2015), and even several different kinds of learning in systems as simple as molecular networks (Biswas et al., 2022, Biswas et al., 2021). These have major implications for understanding habituation and memory in regulatory networks (Mathews et al., 2023, Keresztes et al., 2025, Veres et al., 2024, Csermely et al., 2020), as well as cardiac memory (Chakravarthy and Ghosh, 1997, Headrick et al., 2015, Kunecki et al., 2018) and active cell perception and responses to stimuli (Bugaj et al., 2017, Mitchell and Lim, 2016, Dine et al., 2018) in a variety of cells. Various types of cells demonstrate flexible responses by utilizing molecular and/or bioelectrical signaling systems, which allow them to make contextual decisions (Koseska and Bastiaens, 2017, Biswas et al., 2021, Biswas et al., 2022, Levin, 2022, Levin, 2023a). These are especially relevant to aging and longevity because, just as in neuroscience, they remind us that crucial regulatory information can arise as a function of physiological history and be maintained at the physiological, dynamical systems level – not necessarily at the hardware level visible to omics and similar ways of characterizing molecules. In other words, as with behavior-shaping and control theory, a major target for longevity interventions may not be physical media (DNA, proteins) but persistent states, memories, and priors (in the active inference sense) held by tissues over time. Development of tools to reset theses represents an important avenue that is yet barely explored in longevity research. However, in regenerative biology for example, it has already been shown that the effects of a powerful dominant mutant in the crucial Notch pathway can, in effect, be repaired “in software” by appropriate electroceutical exposure which rescues normal form and function of the brain, face, heart, etc. (Pai and Levin, 2022, Pai et al., 2015).
Throughout the lifespan, coordinated cellular collectives must navigate toward a target morphology in morphospace. This represents anatomical homeostasis (Harris, 1987, Levin et al., 2018): the capacity to reach and maintain a characteristic anatomical state against perturbations that might push the system away from its intended configuration. This framework offers a new understanding of developmental biology: rather than viewing tissue formation solely as the automatic execution of genetic programming through biochemical pathways, it characterizes the process as one where cellular collective dynamically navigate toward their target architecture, constantly adjusting and problem-solving based on contextual cues to minimize developmental/regenerative errors as they arise. Salamanders exemplify these organisms with extraordinary tissue renewal abilities, capable of regrowing entire lost limbs with precision—the process stops automatically once the correct anatomical configuration has been achieved. Comparable healing powers exist across diverse taxa, including deer that regenerate antlers, complete with vasculature, nerve, and bone(Nieto-Diaz et al., 2012, Li, 2012), and human liver and fingertips (Illingworth, 1974). Remodeling, like outright regeneration, illustrates the key concept of cells harnessed toward a large-scale goal; for example, tadpoles whose craniofacial organs were scrambled readily reorganize their face to achieve typical adult frog morphology and then they stop (Vandenberg et al., 2012).
The key point is that a common assumption of molecular, cell, and developmental biology is holding back research in this (and other) fields: models of open-loop “emergence” (e.g., cellular automata-like models) in which there are no goals, and nothing knows anything, simply do not fit a large sector of relevant biology. Instead, cybernetic models of homeostatic processes, which reduce error of the collective relative to a setpoint memory (stored via partially known mechanisms (Levin, 2021a)), are much more applicable. This is not problematic, despite the persistent taboo on teleology in the life sciences (McShea, 2012, Teufel, 2011, Reese, 1994), because we have had rigorous tools for dealing with knowledgeable, goal-driven systems (regardless of their composition) for over 80 years. Cognitive capacities are not magic, and what’s being imported here is not mysteries but the insights of cybernetics and cognitive neuroscience (Pezzulo and Levin, 2016, Pezzulo and Levin, 2015, Rosenblueth et al., 1943). The tools used to study other kinds of collective intelligence are being used to reveal how individual cells are aligned in anatomical homeostatic processes (McMillen and Levin, 2024, Couzin, 2007). Such systems, like the “traffic jam” analogy for cancer (not due to problems in any specific car’s hardware), are susceptible to a different kind of disorder: scenarios in which the setpoint no longer motivates the group to reduce error, or the setpoint information itself becomes degraded.
One possible theory of aging then is loss of goal-directedness (Pio-Lopez et al., 2025): the progressive decline in coordinated, and goal-directed morphological behavior that occurs across multiple organizational scales, after the developmental phase. During embryogenesis, cells exhibit robust collective intelligence, navigating morphospace to achieve specific anatomical and physiological targets through coordinated multicellular problem-solving. However, this sophisticated goal-directed activity may degrade after the adult target morphology has been reached – not because of damage or specific mechanisms evolved to ensure it, but because it is common for cognitive systems to experience dysregulation once they’ve met their goal but do not have a new one to pursue (Pinquart, 2002). Recent computational modeling of regulative morphogenesis demonstrates that even in the absence of genetic damage or entropic decay, complex tissues spontaneously begin to deteriorate sometime after completing developmental objectives – this was an emergent effect, not driven by any low-level component of the in silico model.
This framework has significant implications for understanding aging: the degradation of tissue-level and organismal-level goal-directedness permits individual cells to escape hierarchical regulatory constraints and pursue autonomous objectives, as observed in oncogenesis where cells prioritize local over collective goals or as seen in the genetic atavistic behavior where cells activate different ancient genetic pathways during aging (Pio-Lopez and Levin, 2026). Therapeutically, this perspective suggests that interventions aimed at restoring coordinated tissue behavior through reactivation of dormant regenerative programs may prove more effective than approaches targeting isolated molecular defects (Dai et al., 2025, Nagai et al., 2024). In other words, in order to rejuvenate, we need to re-invigorate the importance of the setpoint, or perhaps impose a new morphological goal to the biological system (see sections below on the future of the body). It is possible that the longevity-increasing effects of exercise are due to small-scale challenges to the body structure which trigger repair, while the extremely large-scale challenges of bi-weekly fission and regeneration are what keep planaria perpetually young.
Is there evidence for increasing dissociation of subunits (cells) from a global setpoint in aging? One well-known one is the increase of cancer in old age (Pilleron et al., 2019, Thakkar et al., 2014). The other was discovered recently, and reveals an interesting dissociation of transcriptomic control at the evolutionary scale: atavistic dissociation (Pio-Lopez and Levin, 2026). The “Atavistic Genetic Expression Dissociation (AGED)” framework distinguishes aging from cancer’s reversion to ancient gene expression (Davies and Lineweaver, 2011, Lineweaver and Davies, 2021), characterizing it as the progressive fragmentation of coordinated multicellular regulation across tissues and gene networks. This dissociation appears as a diverse, tissue-dependent alterations of differential genetic expression: rather than uniform regression across all tissues to a shared ancestral state, tissues can revert along distinct evolutionary pathways (Pio-Lopez and Levin, 2026). This observation has been found for two multi-tissue aging databases covering skin, ovarian, immune, senescent and mesenchymal-senescent cells, but interestingly not for brain cells and mesenchymal stem cells. This indicates that cells drift through phylogenetic/evolutionary space, with tissues fragmenting their coordinated genetic identity and reverting along separate pathways, similarly to the loss of cellular identity found during aging (Izgi et al., 2022, Yang et al., 2023). Overall, these data may suggest that in a multi-competency architecture, the scaling of goals can shift bidirectionally: upward toward integration and higher-scale goals like anatomical homeostasis, or downward toward fragmentation, enabling cells to pursue independent goals and triggering ancestral regression (Pio-Lopez and Levin, 2026) and/or cancer (Levin, 2019).
Another specific way that anatomical homeostasis can break down is if the pattern memory degrades. What could underlie a progressive cognitive decline of the collective intelligence of the body? Work over the last 25 years shows that some aspects of large-scale target morphology are represented through stable bioelectrical patterns (Levin and Martyniuk, 2018). Described as the “physiological software of life” (Funk and Scholkmann, 2023, Funk, 2013, Burr and Northrop, 1935), these bioelectrical control systems constitute an informational framework enabling dynamic coordinated anatomical organization across scales (Pio-Lopez and Levin, 2024). This system regulates key processes like regenerative repair, developmental organization, and oncogenic suppression (Levin, 2021b, Levin, 2021a, Levin, 2023a, Levin et al., 2017, Pio-Lopez and Levin, 2024), specifically by encoding setpoints such as the layout of the vertebrate face (Vandenberg et al., 2011), the number of heads (Durant et al., 2019), and the directionality of primary body axes (Levin et al., 2002). Re-writing of these bioelectric pattern memories, with ion channel misexpression, optogenetics, or ion channel drugs, causes the cells to build the corresponding structures (reviewed in (Levin, 2021a)). Recently, the whole-body bioelectric pattern differences between mortal and immortal hydra have been characterized (Kapsetaki et al., 2025). Time-dependent disruption in such bioelectrical prepattern could cause a loss of morphostatic information, and be a downstream mechanism driving reduction of effective upkeep of tissues and organs (Pio-Lopez and Levin, 2024), causing the degradation of the anatomy, structurally and functionally.
Such degradation could be due to damage to ion channels and other aspects of the electrome. For example, evidence links ion channel malfunction to age-related decline, including senescence that induces characteristic changes of voltage potential in senescing human cells (Sediqi and Levin, 2025), stem cell exhaustion, chronic inflammation, loss of proteostasis, and altered nutrient sensing pathways (Lopez-Otin et al., 2023, Pio-Lopez and Levin, 2024). However, it could also be due to physiological dynamics in bioelectric networks (Pietak and Levin, 2017) which do not require an underlying error to manifest (this type of disorder is well known in neuroscience as cognitive disorders driven not by organic disease but by experiences, trauma, or canalized trains of depressive thought etc.). In other words, patterns can arise and change in bioelectric networks in ways that are to some extent independent of the hardware – a feature that evolution greatly exploits in the function of the nervous system but that evolved from developmental dynamics of somatic cells.
Taken together, these models suggest new targets for a radical regenerative medicine that shift the focus from repairing molecular pathways to manipulating the collective decision-making of cell swarms, to direct large-scale tissue repair and organ regeneration (Lagasse and Levin, 2023, Pio-Lopez and Levin, 2023).
4. Beyond Persistence of Standard Human Bodies: Expanding the Space of Possible Bodies and Minds
We argue that the definitive solution to longevity lies through control of native regenerative pathways. The endgame of that field is the “anatomical compiler” – a system that receives a description of the anatomy needed (for a plant, animal, biobot, organ, etc.) and produces the set of stimuli that need to be given to cells to build exactly that. It is not a 3D printer, but a communication device, translating between the goals of the biologist and those of the cellular collective. Not only aging, but birth defects, traumatic injury, and cancer will all be solved once we have a way of controlling what cells will build.
Driven by all of these pressing medical needs, the morphogenetic code will be cracked, and the anatomical compiler will eventually be built. Crucially, this means that we will be able to direct not only the repair and longevity of evolutionarily-standard organs, but any desired structure. In other words, we will no longer be locked in to the human form that the meanderings of evolution have given us. We think it entirely implausible that, given the ability to control growth and form, humans will stop at the current configuration, with all of its limitations and susceptibility to morbidity and mortality. The search for repair and life extension leads directly and inevitably to a widening of the possibilities for human (and trans-human) embodiment.
What implications will this have? We can start thinking about this using available examples in the animal kingdom. One of the most instructive is that of the caterpillar and butterfly transformation (Figure 5). First, the soft-bodied creature which crawls is remodeled to live as a hard-bodied being in a higher-dimensional (3D) space, acquiring sex organs and food preferences as well as a completely different behavioral controller needed for this new lifestyle. Second, caterpillars that are trained retain that information upon metamorphosis (Blackiston et al., 2015, Blackiston et al., 2008, Sheiman and Tiras, 1996), despite the fact that the brain is largely liquified and rebuilt. The most amazing aspect of this is not that the information is simply retained – it’s that it is somehow generalized and remapped onto an entirely new body, because the specific memories of a caterpillar (crawling behavior to eat leaves upon seeing a specific stimulus) are of no use to a butterfly (but, flying behavior to drink nectar upon that same stimulus, is relevant). Radical morphogenetic change preserves not fidelity to the past but adaptive, functional saliency toward future needs.
Will we retain memories and personalities if we augment and modify our bodies, or just replace aging neurons with progeny of naïve stem cells? It is not known, but planarian flatworms regenerate memories as well as body structures (Blackiston et al., 2015, Corning, 1967, Corning, 1966, Goldsmith et al., 1978), imprinting them on newly-developing brains, and the example of insect metamorphosis suggests that it is at least possible. In this context, one can imagine the point of view of the caterpillar, facing the singularity of its transformation, after which it can be argued to both exist and not. One can also imagine the point of view of the butterfly, inheriting memories and behavioral tendencies from a past life, and feeling perhaps at best a passing kinship with caterpillars. Whose duration should a longevity treatment prolong? The human life cycle is not as extreme; nevertheless childhood, and its attendant changes of physical and mental characteristics and priorities, show that these kinds of issues are not alien to us (one can easily map the above story to the same existential questions of passing through and puberty).
It is possible (Figure 6) that there are additional steps of metamorphosis that are open to us beyond the maturation of puberty, once we have the know-how to control morphogenesis and acquire the longevity to experience additional phases of life (indeed, it may not be necessary to artificially induce it – perhaps we have other forms which have simply never been activated because no one has lived long enough – a situation which will likely change).
The continuous plasticity of morphogenesis discussed above means that biology is extremely interoperable, with novel components that are both natural and engineered (Wang and Guo, 2016, Yum et al., 2014). This opens new avenues for expanding the space of possible bodies and minds. If we unlock regenerative medicine for rejuvenation, we can become biocyborg (Pio-Lopez, 2021). While a standard cyborg combines organic tissue with biomechatronic parts—such as prosthetic limbs, pacemakers, or neural implants—through external integration, a biocyborg achieves enhancement through biological means. The technology and the body share the same physicality; both components are fundamentally biological, making the hybridity less about hardware-software interfaces and more about redesigned living systems. The aim of longevity treatments is often stated as maintaining the anatomy, but the paradox of change reminds us that persistence need not mean stasis and stagnation.
This research naturally leads to the expansion of the space of synthetic beings and cyborgs of all sorts – a process that was begun with the first usage of wooden canes, continuing through glasses, insulin pumps, sensory augmentation and substitution devices, and eventually fully-integrated AI-powered internal prosthetics. Contemporary technologies and proof-of-concept demonstrations reveal a vast spectrum of potential biological forms or ‘life as it could be’, suggesting that life’s diversity will expand dramatically in the future (Doursat et al., 2013, Doursat and Sanchez, 2014, Kamm and Bashir, 2014, Ebrahimkhani and Levin, 2021). Such emergent biological entities will arise through the combination of naturally evolved elements across hierarchical scales—from genetic material through cellular, tissue, and organ levels—with engineered components including nanomaterials, electronic/chemical/optical interfaces, and computational algorithms (Nanos and Levin, 2022, Clawson and Levin, 2023) (see Figure 1C and Figure 2). Substitutions are possible at any level of biological complexity. In other words, we will create new forms of artificial chimerism between different kind of intelligences.
This development of new synthetic beings, with new Umwelts made possible by sensory-motor augmentation (Bach-y-Rita, 1990, Bach-y-Rita, 1981, Bach-y-Rita et al., 1969, Bach-y-Rita, 1967, Sampaio et al., 2001, Eagleman and Perrotta, 2022, Perrotta et al., 2021) has been probably the most developed with the field of brain-computer interfaces (BCI) and neuroprosthetic limbs (Degenhart et al., 2020, Buccelli et al., 2019). These technologies enable restoration of motor capabilities in individuals experiencing paralysis or limb loss through direct neural communication pathways. The control architectures employed in these neuroprosthetic platforms include neuromodulation implantable devices, prosthetic limbs with neural integration, neurorobotic systems, BCI control methodologies, and sensory feedback mechanisms within such interfaces (Wright et al., 2016, Lebedev, 2014). It has been shown that paralyzed patients can communicate via a BCI and neural decoding even years after the onset of the paralysis by imagining writing with a pen on a piece of paper (Willett et al., 2021). Recently, a brain-machine interface allowed a quadriplegic patient to play chess solely through imagined motor commands, establishing the clinical feasibility of motor imagery-based BCI technology (Drew, 2024). But this is not all, an important application of these neural interfaces is to redirect the neural signals towards external robotics systems. Hochberg et al. demonstrated that patients with tetraplegia successfully controlled a robotic limb through a BCI, executing complex three-dimensional reaching and grasping actions—one participant even achieved sufficient dexterity to independently drink coffee using the prosthetic arm (Hochberg et al., 2012). Interestingly, these external body parts not only extend the body but redesign the body schema. Indeed, it has been shown that monkeys integrated a third robotic arms. In a pioneering work, Nicolelis and colleagues showed that through visual feedback alone, monkeys successfully generated reaching and grasping motions in a robotic arm without moving their own limbs. The acquisition of brain-machine interface control coincided with functional reorganization across multiple cortical regions, indicating that the robot’s dynamic characteristics became integrated into the brain’s motor and sensory representational maps (Carmena et al., 2003).
Therefore, these extensions not only give new motor capabilities but also change our sensory processing. This shows that in addition of the plasticity of the body and its representation, we also have plasticity of the nervous system to new embodiments. Exploiting this property, researchers also developed sensory augmentation or substitution devices (Shull and Damian, 2015). Neuroprosthetics and sensory augmentation go the opposite path of BCI, instead of achieving neural decoding they send information to the body/brain by stimulation (electrical, magnetic or light-gated for example). More specifically, visual neuroprostheses have been developed for blind patients (Normann et al., 2009, Pio-Lopez et al., 2021). Experimental studies have reported the perception of brief light sensations, known as phosphenes, in the visual field when electrical stimulation is applied to visual pathways. Because multi-electrode stimulation can evoke spatial patterns of phosphenes, this effect is widely treated as a key physiological building block for visual neuroprostheses aimed at restoring ‘phosphenized’ visual experience in blindness (Normann et al., 2009, Pio-Lopez et al., 2021). Sensory substitution devices have also been created to replace missing sensory input by translating information from one modality (e.g., vision) into another (e.g., audition or touch), allowing users to access environmental cues through an alternate channel (Macpherson, 2018). Beyond substitution, another study shows that “new” signals can be incorporated into spatial perception, for example by providing an auditory compass cue that users learn to integrate with their existing sense of space (Nagel et al., 2005).
Crucially, this field is not limited to passive technology added to a living body; instead, high levels of autonomy and cognitive competency can exist on both sides of the hybrid. This ranges from AI-powered implants (Mohamed et al., Clawson and Levin, 2023) to chimeric living materials. For example, synthetic biology approaches have demonstrated that it is possible to biologically implement artificial-intelligence methods, including formal artificial neural networks (Nesbeth et al., 2016, Stano et al., 2018). In principle, such AI-inspired computational schemes could be encoded in biological substrates and delivered to human cells, enabling cell-based execution of predefined information-processing functions. Likewise, the creation of multicellular patient-specific biobots with healing potential (Gumuskaya et al., 2023) reveals the vast space of possibilities.
The space of new bodies and minds can also be enriched with hybrots and animats. They are hybrid systems in which dissociated neural cultures are embedded in closed-loop control architectures to operate either physical robots (hybrots) or simulated agents (animats), providing a powerful experimental setting to probe how neural networks acquire sensorimotor functions. In an influential early implementation, cortical neurons from rat brains were dissociated and grown as a two-dimensional culture on a microelectrode array that both recorded spiking activity and delivered electrical stimulation. In this preparation, distinct spatiotemporal firing patterns in the culture were mapped onto the movement commands of a virtual agent navigating a maze, while sensory feedback about the agent’s heading and proximity to obstacles was fed back to the neurons, thereby enabling the culture to gradually establish relationships between its internally generated activity and task-relevant “sensory” inputs (Demarse et al., 2001). Building on this framework, the MEART (multi-electrode array art) system coupled a similar neural culture to robotic drawing arms, using patterned stimulation derived from the current drawing and a predefined target image to shape the network’s output (Bakkum et al., 2007). More broadly, such closed-loop chimeric systems offer a tractable model for studying how neural ensembles reorganize when learning new behaviors in novel bodies and environments, without the interpretative complexity introduced by intact nervous systems, while still retaining meaningful goal-directed capabilities.
In addition to the implications of longevity-motivated research on the future of the body, we point out the reverse part of this virtuous cycle: the use of synthetic model systems to better understand and implement longevity. For example, Anthrobots – living constructs made from wild-type, adult, human patient cells (Gumuskaya et al., 2025) are actually younger (as measured by epigenetic clock) than the cells they are made of. This illustrates not only the ability to test longevity interventions on patient-specific avatars with behavior and a convenient lifespan, but also the impact of lifestyle and morphogenesis itself on markers of aging in the absence of any reprogramming factors, genomic editing, etc.
All these prosthetic/chimeric technologies illustrate the plasticity of life, from bodies to minds. They show an artificial chimerism (Clawson and Levin, 2023, Nanos and Levin, 2022, Pio-Lopez, 2021), a deeper integration of living things and technologies with important implications for our understanding of life, cognition, to be human and humanity (see Figure 2).
5. Artificial Chimerism and Multi-Scale (Non-Darwinian) Evolution
These prosthetic technologies are blurring the lines between familiar binary categories around natural organisms and machines, which we inherited from a pre-scientific time of limited capability and imagination. Indeed, the distinction between normal, pathological, and enhanced states is not well-defined, as these categories exist along a continuum rather than as discrete entities (Hofmann, 2017). Therapeutic interventions and enhancement practices overlap substantially: any efficacious medical treatment improves a patient’s functional status—an enhancement in the broad sense—although not all enhancement procedures are therapeutic in nature (Dekkers and Rikkert, 2007). Contemporary neuroprosthetic research illustrate this conceptual ambiguity. Visual neuroprostheses, initially developed for therapeutic purposes, can evolve into enhancement procedures with an ultra-violet filter for example, while vaccination can be conceptualized either as prophylactic medicine or an enhancement of immune system capacity. These examples demonstrate that normality, pathology, and enhancement represent overlapping domains with indistinct boundaries rather than mutually exclusive categories. Aging research in the same grey area by providing resistance to all age-related diseases and potentially radical longevity.
The plasticity of mind and bodies is blurring the lines between the “natural” and the “artificial”, or between “organism” and “machine” (Bongard and Levin, 2021) (Figure 7). As biological tissues are increasingly integrated with electronic components and computational capabilities, the conceptual boundary separating these concepts loses its definitional clarity. No fundamental criterion exists to demarcate these categories cleanly, and viable entities can be envisioned across the entire spectrum—from predominantly biological systems with minor computational augmentation to largely synthetic architectures incorporating small amounts of living tissue. Rather than serving as a rigorous classificatory tool to separate life from pure machinery, teleonomy is more productively understood as a continuous variable that quantifies goal-directed behavior across diverse substrates, offering a unifying framework for analyzing agency independent of material composition. The same applies to dichotomies such as “evolved” versus “designed”: these labels offer limited explanatory power regarding an agent’s cognitive traits, functional capabilities, or behavioral propensities. Contemporary research in Artificial Life for example increasingly employs evolutionary computation to generate novel designs (Bedau, 2005), while human engineering itself represents the output of biological systems shaped by natural selection, rendering the evolved-designed distinction fundamentally porous.
Therefore, longevity technologies along with multiscale chimerism will accelerate a multiscale and non-Darwinian evolution via the exploration of the space of synthetic beings. Unlike traditional evolutionary processes driven by random mutation and natural selection operating over generational timescales, this framework enables intentional reconfiguration of living systems across organizational levels—from molecular networks to whole organisms—within individual lifespans. By decoupling evolutionary change from reproductive cycles and mortality, longevity interventions create extended temporal windows for iterative modification and functional optimization of biological substrates pushing forward the exploration of the option space of possible minds and bodies. This will have several important implications for the individual and evolution.
Over long period of times, the individual will have the possibility to bioengineer his own evolution. This kind of biocyborg is already there, they are not only living in science-fiction. For example, Elisabeth Parrish is known for having accomplished gene therapies in Colombia in order to increase the size of her telomeres (Regalado, 2015). Another example is Josiah Zayner that injected himself a CRISPR solution to inactivate the gene responsible for myostatin production to see if it will allow him to build more muscle (Mehlman et al., 2023). This raises the deep question of what it is to be a species, if one day the individual can become its own species. Moreover, transformative risks extend beyond the individual level to encompass entire populations when such technological interventions or engineered genetic modifications become heritable and transmissible to subsequent generations. Indeed, astrophysics researcher claimed that the minimum viable number of one population to start a new colony (or human species) on another planet is only 98 (Marin and Beluffi, 2018).
6. Open Questions for the Future of Humanity
Mental Aging: Software, Hardware, or Both?
The question of whether mental aging is “software” (accumulated patterns, habits, schemas) or “hardware” (physical brain degradation) has profound implications. Crucially, the hardware component appears more reversible than previously thought. Brain plasticity research demonstrates that aged brains can keep substantial capacity for structural and functional change through cognitive training, environmental enrichment, and physical exercise. The aging brain develops compensatory by recruiting additional neural regions to maintain cognitive function despite degradation (Goh and Park, 2009). When older adults engage in sustained novel tasks, they can increase neural volume and alter activation patterns, showing genuine neuroplasticity persists into late life.
The cognitive component presents a more complex problem. Indeed, if biological rejuvenation restores a teenager’s neuroplasticity to an 80-year-old, would they genuinely think like a teenager? Perhaps not, or at least not immediately. Decades of habits, preferences and experience accumulated during decades may not vanish and may persist in rejuvenated hardware. However, a rejuvenated brain could theoretically rewire and regain youthful cognitive plasticity. The answer to this question, which longevity research will likely provide, will be an important example of a scientific, empirical approach that impacts on ancient philosophical questions of personal identity. However, it is fundamentally rooted in the deep question about the mind-brain (or mind-body (Ciaunica, 2024, Ciaunica et al., 2023)) relationship, which technologies like longevity treatments will bring to the forefront. While it is commonly thought that neuroscience has a good handle on how neural tissue supports minds, there are strikingly incongruous laboratory and clinical observations of mismatches between brain tissue and cognitive capacity that suggest that we are far from a full understanding of this fundamental issue (reviewed in (Kofman and Levin, 2025)).
Cognitive Limits of Extreme Longevity
Can a human mind remain coherent and sane over thousands or millions of years? We know that time can lead to neurodegeneration and dementia but what about a healthy brain/mind on these extended timelines? Would boredom, loss of meaning, or other high-level problems (perhaps not relevant to high-longevity pets?) set in, or is the capacity to identify challenges, interesting problems, and rich social relationships unlimited in humans and their progeny?
What about memory saturation – possible limits on learning capacity? It has been estimated that the human brain may can store 2.5 petabytes of information (Vazza and Feletti, 2020), the equivalent of 300 years of streaming videos. If true, and one lives 1000 years, would the memories of early centuries fade? What would be the identity of one person on such timescale if they lose all foundational memories that defined their history and personalities?
Death has also been often presented as a powerful motivational driver in life (Jacobsen, 2017). Some have argued that the human motivational system has evolved to sustain approximately 80 years lifespan, and extreme longevity may cause cognitive or spiritual exhaustion. Opponents of aging have also claimed that death is what give life meaning. However, we would argue that nothing tuned our current lifespan to maximize meaning, and if limited lives were important for meaning, would a 20- or a 10-year lifespan be desirable? We think not. Indeed, maybe the kind of wisdom, compassion, and maturity we want to see in our species needs more decades to develop than the meandering processes of evolution had given us to date. Perhaps our development requires tweaking of the lifespan just as we use dentistry, education, exercise, and assistive devices to adjust other accidents of birth that limit our potential.
Social Stratification
A society where the body does not correlate with chronological age is at risk of an important social stratification. Indeed, the body’s appearance can signal social cues, or social status. When a 25-year-old body might contain 100 years of experience, or vice versa, fundamental social coordination vanishes. The “real young” versus “faux-young” distinction might indeed matter significantly. Young adults navigating early life transitions (career uncertainty, relationship formation, identity exploration) face different psychological challenges than centenarians in young bodies who’ve already resolved these developmental tasks multiple times. Without appearance cues, intergenerational relationships become ambiguous—and all require renegotiation when you can’t assess shared life stage. These are just examples and merely speculations but a post-mortal society where rejuvenation have been achieved will inevitably challenge the way individuals interact and make society.
Species-Level Longevity and Humanity’s Future
At the species level, longevity fundamentally alters evolutionary dynamics. Human-engineered longevity creates “non-Darwinian evolution” — we actively override natural selection. This has profound implications. First, longer lives may extend the chronology of individuals but also the rhythm of civilization. Indeed, through history, society evolved with the replacement of generations. This may cause a cultural stagnation risk: if older cohorts don’t die and society does not change, they retain power and resources indefinitely, potentially blocking social innovation and adaptation (Lucke & Hall, 2005). Revolutionary ideas typically require generational replacement.
However, longevity may also lead to cumulative intelligence and knowledge that would not be lost with death. Scientists, artists, and philosophers might devote centuries to a single theme, or transfer learning between fields and achieving intellectual depth impossible within a single mortal lifespan. The same mechanism that threatens stagnation could foster unparalleled progress. If the anatomical compiler become reality, any individual may choose their own evolution redefining the notion of species. The human species could dissolve into a plurality of self-directed life forms: a society of unique species coexisting within a shared cultural network.
7. Conclusions
Bodies result from homeostatic processes in which competent subunits (cells, and the molecular networks within them) functionally align to implement high-level goal states in anatomical and other spaces. Morphogenesis also exhibits allostatic (predictive) and problem-solving competencies. By transcending the assumption that we are merely mechanical, emergent results of chemical processes which have no goals and no intelligence of their own, we gain the opportunity to apply the many tools and insights of cognitive neuroscience to the agential material of life. This will be essential, for the goal of longevity research, which likely cannot be accomplished by approaches focused entirely at the physics and chemistry levels. We envision a near future in which the advances of regenerative medicine and diverse intelligence research have gone beyond the molecular hardware of our bodies, to the software of life, and beyond that, to communicating with the multiscale agential material of which we are made to be able to specify the anatomical setpoints of morphogenetic processes, and gain total control over living form and function. Future work must take advantage of the powerful role of bioelectricity as a mechanism to scale the competencies of cells (Levin, 2019), as well as investigate additional biophysical modalities for storing setpoints (biomechanical and perhaps biophoton layers (Beloussov et al., 2007, Beloussov, 2001, Cohen and Popp, 2003, Voeikov and Beloussov, 2007, Belousov et al., 2007, Bischof, 2000)), applying the perspectives described above to the powerful model systems of aging research.
To the prevalent damage and programmatic theories of the causes of aging, we add two new components. One is the bioelectric prepatterns that encode some setpoints, and are a powerful target for future research in gerontology and its interventions. The other is a unique class of disorders that apply to goal-seeking systems. A mechanical clock can only break down at the hardware level. A dynamical computational or cognitive system can break down because of specific inputs, thoughts, persistent patterns, and physiological priors that are invisible to omics techniques that only characterize molecules. Tools must be developed to enable a kind of somatic psychiatry – to read and intervene not only in material and metabolic states but at the level of memories and decision-making patterns across scales (Mathews et al., 2023, Veres et al., 2024, Csermely et al., 2020, Piedimonte and Benedetti, 2016).
Like the Ship of Theseus, our persisting body is a self-modifying target state within the space on anatomical possibilities, maintained by the integrated alignment of cell groups guided by a collective setpoint. It is these pattern memories that must be reinforced, or improved, for true individual- and species-level longevity. Bodies receive inputs from both DNA and the environment, and because of the cognitive processes occurring at all scales (Baluska et al., 2024, Miller et al., 2023, Baluska et al., 2022b, Baluska et al., 2022a), are likely susceptible to as yet-ignored failure modes caused by physiological experiences, not organic disease (Miller et al., 2024, Pio-Lopez et al., 2025).
One possible example is the existential question facing an ageless humanity as much as it faces living bodies that finish embryogenesis: when you have completed your goal, what do you do next? If this turns out to be a fundamental problem for morphogenesis, treatments may need to reinforce the goal state; or perhaps, the only way to persist is to change: re-motivate the regenerative homeodynamic systems with a new set of anatomical and functional goal states. That approach could also address the same question on a human level: when you have completed your goals as one type of being, maybe there are new vistas facing you as a different one. Perhaps like the caterpillar, we can live much longer, but not in the exact same form factor and mindset.
Ultimately, longevity should mean more than “live longer exactly the way you are”. It should be an opportunity to grow, to have principled, rational control over the way we change, and to make use of as long a runway as might be needed to reach our individual and species-level potential in wisdom, compassion, and capability. It is statistically improbable that this just happens to be doable in the lifespan that Darwinian evolution, which aims for “good enough to replicate”, not for any of our values, has given us. It also seems highly unlikely that a visitor to Earth in 50 or 100 years will find precisely what he would see now: the same ancient bodyplan, with its susceptibility to a wide range of viruses, bacteria, and degenerative diseases, and a limit on IQ and healthspan that was set by the activity of indifferent cosmic rays hitting DNA for millions of years.
Future humans will think back on us and marvel with incredulity that we had to live our whole lives, short as they were, in whatever body we happened to have been handed by a history of mutation and gestation. The good news, we estimate, is that given advances in relevant conceptual and empirical tools, this restrictive time period is almost over. There is a lot of work to do to improve on these ancient limitations, and we will certainly make mistakes. We think however that we can, and ethically must, do better than the cosmic rays in providing everyone with the opportunity to mold their body as well as their mind, to reach whatever potential they value. The roadmap of regenerative medicine leads not only to radical longevity, but through it, to freedom of embodiment, and a massive increase in intentional agency over not just how long we live, but how.
References
- Austad, S. N., and J. M. Hoffman. 2018. Is Antagonistic Pleiotropy Ubiquitous In Aging Biology? Evol Med Public Health 2018: 287–294. [Google Scholar] [CrossRef] [PubMed]
- Bach-Y-Rita, P. 1967. Sensory Plasticity. Applications To A Vision Substitution System. Acta Neurol Scand 43: 417–26. [Google Scholar] [CrossRef]
- Bach-Y-Rita, P. 1981. Brain Plasticity As A Basis Of The Development Of Rehabilitation Procedures For Hemiplegia. Scand J Rehabil Med 13: 73–83. [Google Scholar] [PubMed]
- Bach-Y-Rita, P. 1990. Brain Plasticity As A Basis For Recovery Of Function In Humans. Neuropsychologia 28: 547–54. [Google Scholar] [CrossRef] [PubMed]
- Bach-Y-Rita, P., C. C. Collins, F. A. Saunders, B. White, and L. Scadden. 1969. Vision Substitution By Tactile Image Projection. Nature 221: 963–4. [Google Scholar] [CrossRef]
- Bakkum, D. J., P. M. Gamblen, G. Ben-Ary, Z. C. Chao, and S. M. Potter. 2007. Meart: The Semi-Living Artist. Frontiers In Neurorobotics 1: 5. [Google Scholar] [CrossRef]
- Baluška, F., and M. Levin. 2016. On Having No Head: Cognition Throughout Biological Systems. Front Psychol 7: 902. [Google Scholar] [CrossRef]
- Baluska, F., W. B. Miller, Jr., and A. S. Reber. 2024. Sentient Cells As Basic Units Of Tissues, Organs And Organismal Physiology. J Physiol 602: 2491–2501. [Google Scholar] [CrossRef]
- Baluska, F., W. B. Miller, and A. S. Reber. 2022a. Cellular And Evolutionary Perspectives On Organismal Cognition: From Unicellular To Multicellular Organisms. Biological Journal Of The Linnean Society. [Google Scholar]
- Baluska, F., A. S. Reber, and W. B. Miller, Jr. 2022b. Cellular Sentience As The Primary Source Of Biological Order And Evolution. Biosystems 218: 104694. [Google Scholar] [CrossRef]
- Bartke, A., L. Y. Sun, and V. Longo. 2013. Somatotropic Signaling: Trade-Offs Between Growth, Reproductive Development, And Longevity. Physiol Rev 93: 571–98. [Google Scholar] [CrossRef]
- Bates, E. 2015. Ion Channels In Development And Cancer. Annu Rev Cell Dev Biol 31: 231–47. [Google Scholar] [CrossRef]
- Bedau, M. A. 2005. Artificial Life: More Than Just Building And Studying Computational Systems. Artif Life 11: 1–3. [Google Scholar] [CrossRef]
- Belousov, L. V., V. L. Voeikov, and V. S. Martynyuk. 2007. Biophotonics And Coherent Systems In Biology, New York, Ny. Springer. [Google Scholar]
- Beloussov, L. V. 2001. Morphogenetic Fields: Outlining The Alternatives And Enlarging The Context [English]. Rivista Di Biologia - Biology Forum 94: 219–35. [Google Scholar] [PubMed]
- Beloussov, L. V., V. L. Voeikov, and V. S. Martynyuk, eds. 2007. Biophotonics And Coherent Systems In Biology [English]. Springer: New York, Ny, Usa. [Google Scholar]
- Ben-Jacob, E. 2009. Learning From Bacteria About Natural Information Processing. Annals Of The New York Academy Of Sciences 1178: 78–90. [Google Scholar] [CrossRef] [PubMed]
- Best, J., and I. Rubenstein. 1962. Maze Leaning And Associated Behavior In Planaria. Journal Of Comparative & Physiological Psychology, 560–566. [Google Scholar]
- Bischof, M. 2000. Edited by L. V. Beloussov, F. A. Popp, V. Voeikov and R. Van Wijk. Field Concepts And The Emergence Of A Holistic Biophysics. In Biophotonics And Coherent Systems. Moscow. [Google Scholar]
- Biswas, S., W. Clawson, and M. Levin. 2022. Learning In Transcriptional Network Models: Computational Discovery Of Pathway-Level Memory And Effective Interventions. Int J Mol Sci 24. [Google Scholar] [CrossRef]
- Biswas, S., S. Manicka, E. Hoel, and M. Levin. 2021. Gene Regulatory Networks Exhibit Several Kinds Of Memory: Quantification Of Memory In Biological And Random Transcriptional Networks. Iscience 24: 102131. [Google Scholar] [CrossRef]
- Blackiston, D., T. Shomrat, and M. Levin. 2015. The Stability Of Memories During Brain Remodeling: A Perspective. Communicative & Integrative Biology 8: E1073424. [Google Scholar] [CrossRef]
- Blackiston, D. J., and M. Levin. 2013. Ectopic Eyes Outside The Head In Xenopus Tadpoles Provide Sensory Data For Light-Mediated Learning. The Journal Of Experimental Biology 216: 1031–40. [Google Scholar] [CrossRef]
- Blackiston, D. J., E. Silva Casey, and M. R. Weiss. 2008. Retention Of Memory Through Metamorphosis: Can A Moth Remember What It Learned As A Caterpillar? Plos One 3: E1736. [Google Scholar] [CrossRef]
- Bongard, J., and M. Levin. 2021. Living Things Are Not (20th Century) Machines: Updating Mechanism Metaphors In Light Of The Modern Science Of Machine Behavior. Frontiers In Ecology And Evolution 9. [Google Scholar] [CrossRef]
- Bratic, A., and N. G. Larsson. 2013. The Role Of Mitochondria In Aging. J Clin Invest 123: 951–7. [Google Scholar] [CrossRef] [PubMed]
- Browder, K. C., P. Reddy, M. Yamamoto, A. Haghani, I. G. Guillen, S. Sahu, C. Wang, Y. Luque, J. Prieto, and L. Shi. 2022. In Vivo Partial Reprogramming Alters Age-Associated Molecular Changes During Physiological Aging In Mice. Nature Aging 2: 243–253. [Google Scholar] [CrossRef] [PubMed]
- Buccelli, S., Y. Bornat, I. Colombi, M. Ambroise, L. Martines, V. Pasquale, M. Bisio, J. Tessadori, P. Nowak, F. Grassia, A. Averna, M. Tedesco, P. Bonifazi, F. Difato, P. Massobrio, T. Levi, and M. Chiappalone. 2019. A Neuromorphic Prosthesis To Restore Communication In Neuronal Networks. Iscience 19: 402–414. [Google Scholar] [CrossRef] [PubMed]
- Bugaj, L. J., G. P. O'donoghue, and W. A. Lim. 2017. Interrogating Cellular Perception And Decision Making With Optogenetic Tools. J Cell Biol 216: 25–28. [Google Scholar] [CrossRef]
- Burr, H. S., and F. S. C. Northrop. 1935. The Electro-Dynamic Theory Of Life. Quarterly Review Of Biology 10: 322–333. [Google Scholar] [CrossRef]
- Carmena, J. M., M. A. Lebedev, R. E. Crist, J. E. O'doherty, D. M. Santucci, D. F. Dimitrov, P. G. Patil, C. S. Henriquez, and M. A. Nicolelis. 2003. Learning To Control A Brain-Machine Interface For Reaching And Grasping By Primates. Plos Biol 1: E42. [Google Scholar] [CrossRef]
- Cebrià, F., T. Adell, and E. Saló. 2018. Rebuilding A Planarian: From Early Signaling To Final Shape. Int J Dev Biol 62: 537–550. [Google Scholar] [CrossRef]
- Chakravarthy, S. V., and J. Ghosh. 1997. On Hebbian-Like Adaptation In Heart Muscle: A Proposal For 'Cardiac Memory'. Biol Cybern 76: 207–15. [Google Scholar] [CrossRef]
- Ciaunica, A. 2024. The Forgotten Body: The Emergence Of Conscious Experiences In Early Life. Trends Cogn Sci 28: 967–968. [Google Scholar] [CrossRef]
- Ciaunica, A., E. V. Shmeleva, and M. Levin. 2023. The Brain Is Not Mental! Coupling Neuronal And Immune Cellular Processing In Human Organisms. Front Integr Neurosci 17: 1057622. [Google Scholar] [CrossRef] [PubMed]
- Clawson, W. P., and M. Levin. 2023. Endless Forms Most Beautiful 2.0: Teleonomy And The Bioengineering Of Chimaeric And Synthetic Organisms. Biological Journal Of The Linnean Society 139: 457–486. [Google Scholar] [CrossRef]
- Cohen, S., and F. A. Popp. 2003. Biophoton Emission Of The Human Body. Indian Journal Of Experimental Biology 41: 440–5. [Google Scholar] [CrossRef] [PubMed]
- Conway, M. A. 2005. Memory And The Self. Journal Of Memory And Language 53: 594–628. [Google Scholar] [CrossRef]
- Conway, M. A., and C. W. Pleydell-Pearce. 2000. The Construction Of Autobiographical Memories In The Self-Memory System. Psychol Rev 107: 261–88. [Google Scholar] [CrossRef]
- Corlett, P. R., and P. C. Fletcher. 2014. Computational Psychiatry: A Rosetta Stone Linking The Brain To Mental Illness. Lancet Psychiatry 1: 399–402. [Google Scholar] [CrossRef]
- Corning, W. C. 1966. Retention Of A Position Discrimination After Regeneration In Planarians. Psychanomic Science 5: 17–18. [Google Scholar] [CrossRef]
- Corning, W. C. 1967. Regeneration And Retention Of Acquired Information. Nasa. [Google Scholar]
- Couzin, I. 2007. Collective Minds. Nature 445: 715. [Google Scholar] [CrossRef]
- Csermely, P., N. Kunsic, P. Mendik, M. Kerestely, T. Farago, D. V. Veres, and P. Tompa. 2020. Learning Of Signaling Networks: Molecular Mechanisms. Trends Biochem Sci 45: 284–294. [Google Scholar] [CrossRef]
- Dai, X., X. Li, A. Tyshkovskiy, C. Zuckerman, N. Cheng, P. Lin, D. Paris, S. Qureshi, L. Kruglyak, X. Mao, J. Nandakumar, V. N. Gladyshev, S. Pletcher, J. Sobota, and L. Guo. 2025. Regeneration Leads To Global Tissue Rejuvenation In Aging Sexual Planarians. Nat Aging 5: 780–798. [Google Scholar] [CrossRef]
- Davies, J., and M. Levin. 2023. Synthetic Morphology With Agential Materials. Nature Reviews Bioengineering 1: 46–59. [Google Scholar] [CrossRef]
- Davies, P. C., and C. H. Lineweaver. 2011. Cancer Tumors As Metazoa 1.0: Tapping Genes Of Ancient Ancestors. Phys Biol 8: 015001. [Google Scholar] [CrossRef] [PubMed]
- De Magalhaes, J. P. 2023. Ageing As A Software Design Flaw. Genome Biol 24: 51. [Google Scholar] [CrossRef] [PubMed]
- De Magalhães, J. P. 2011. The Biology Of Ageing: A Primer. An Introduction To Gerontology 21: 1. [Google Scholar]
- De Magalhães, J. P., and A. Ocampo. 2022. Cellular Reprogramming And The Rise Of Rejuvenation Biotech. Trends In Biotechnology 40: 639–642. [Google Scholar] [CrossRef] [PubMed]
- Deans, C. 2021. Biological Prescience: The Role Of Anticipation In Organismal Processes. Front Physiol 12: 672457. [Google Scholar] [CrossRef] [PubMed]
- Degenhart, A. D., W. E. Bishop, E. R. Oby, E. C. Tyler-Kabara, S. M. Chase, A. P. Batista, and B. M. Yu. 2020. Stabilization Of A Brain-Computer Interface Via The Alignment Of Low-Dimensional Spaces Of Neural Activity. Nat Biomed Eng 4: 672–685. [Google Scholar] [CrossRef]
- Dekkers, W., and M. O. Rikkert. 2007. Memory Enhancing Drugs And Alzheimer’s Disease: Enhancing The Self Or Preventing The Loss Of It? Medicine, Health Care And Philosophy 10: 141–151. [Google Scholar] [CrossRef]
- Demarse, T. B., D. A. Wagenaar, A. W. Blau, and S. M. Potter. 2001. The Neurally Controlled Animat: Biological Brains Acting With Simulated Bodies. Auton Robots 11: 305–310. [Google Scholar] [CrossRef]
- Dine, E., A. A. Gil, G. Uribe, C. P. Brangwynne, and J. E. Toettcher. 2018. Protein Phase Separation Provides Long-Term Memory Of Transient Spatial Stimuli. Cell Syst 6: 655–663 E5. [Google Scholar] [CrossRef]
- Doursat, R., and C. Sanchez. 2014. Growing Fine-Grained Multicellular Robots. Soft Robotics 1: 110–121. [Google Scholar] [CrossRef]
- Doursat, R., H. Sayama, and O. Michel. 2013. A Review Of Morphogenetic Engineering. Natural Computing 12: 517–535. [Google Scholar] [CrossRef]
- Drew, L. 2024. Elon Musk's Neuralink Brain Chip: What Scientists Think Of First Human Trial. Nature. [Google Scholar]
- Duan, R., Q. Fu, Y. Sun, and Q. Li. 2022. Epigenetic Clock: A Promising Biomarker And Practical Tool In Aging. Ageing Research Reviews 81: 101743. [Google Scholar] [CrossRef] [PubMed]
- Durant, F., J. Bischof, C. Fields, J. Morokuma, J. Lapalme, A. Hoi, and M. Levin. 2019. The Role Of Early Bioelectric Signals In The Regeneration Of Planarian Anterior/Posterior Polarity. Biophys J 116: 948–961. [Google Scholar] [CrossRef] [PubMed]
- Eagleman, D. M., and M. V. Perrotta. 2022. The Future Of Sensory Substitution, Addition, And Expansion Via Haptic Devices. Front Hum Neurosci 16: 1055546. [Google Scholar] [CrossRef]
- Ebrahimkhani, M. R., and M. Levin. 2021. Synthetic Living Machines: A New Window On Life. Iscience 24: 102505. [Google Scholar] [CrossRef]
- Emmons-Bell, M., F. Durant, A. Tung, A. Pietak, K. Miller, A. Kane, C. J. Martyniuk, D. Davidian, J. Morokuma, and M. Levin. 2019. Regenerative Adaptation To Electrochemical Perturbation In Planaria: A Molecular Analysis Of Physiological Plasticity. Iscience 22: 147–165. [Google Scholar] [CrossRef]
- Fankhauser, G. 1945. The Effects Of Changes In Chromosome Number On Amphibian Development. The Quarterly Review Of Biology 20: 20–78. [Google Scholar]
- Farinella-Ferruzza, N. 1956. The Transformation Of A Tail Into A Limb After Xenoplastic Transformation. Experientia 15: 304–305. [Google Scholar] [CrossRef]
- Fields, C., J. Bischof, and M. Levin. 2020. Morphological Coordination: A Common Ancestral Function Unifying Neural And Non-Neural Signaling. Physiology 35: 16–30. [Google Scholar] [CrossRef]
- Fields, C., and M. Levin. 2022. Competency In Navigating Arbitrary Spaces As An Invariant For Analyzing Cognition In Diverse Embodiments. In Entropy (Basel). p. 24. [Google Scholar]
- Friston, K., M. Levin, B. Sengupta, and G. Pezzulo. 2015. Knowing One's Place: A Free-Energy Approach To Pattern Regulation. J R Soc Interface 12. [Google Scholar] [CrossRef]
- Funk, R. 2013. Ion Gradients In Tissue And Organ Biology. In Biological Systems. [Google Scholar]
- Funk, R. H. W., and F. Scholkmann. 2023. The Significance Of Bioelectricity On All Levels Of Organization Of An Organism. Part 1: From The Subcellular Level To Cells. Prog Biophys Mol Biol 177: 185–201. [Google Scholar] [CrossRef] [PubMed]
- Gems, D. 2022. The Hyperfunction Theory: An Emerging Paradigm For The Biology Of Aging. Ageing Res Rev 74: 101557. [Google Scholar] [CrossRef] [PubMed]
- Gladyshev, V. N. 2016. Aging: Progressive Decline In Fitness Due To The Rising Deleteriome Adjusted By Genetic, Environmental, And Stochastic Processes. Aging Cell 15: 594–602. [Google Scholar] [CrossRef] [PubMed]
- Gladyshev, V. N., S. B. Kritchevsky, S. G. Clarke, A. M. Cuervo, O. Fiehn, J. P. De Magalhaes, T. Mau, M. Maes, R. Moritz, L. J. Niedernhofer, E. Van Schaftingen, G. J. Tranah, K. Walsh, Y. Yura, B. Zhang, and S. R. Cummings. 2021. Molecular Damage In Aging. Nat Aging 1: 1096–1106. [Google Scholar] [CrossRef]
- Goh, J. O., and D. C. Park. 2009. Neuroplasticity And Cognitive Aging: The Scaffolding Theory Of Aging And Cognition. Restor Neurol Neurosci 27: 391–403. [Google Scholar] [CrossRef]
- Goldsmith, C. M., H. R. Hepburn, and D. Mitchell. 1978. Retention Of An Associative Learning-Task After Metamorphosis In Locusta-Migratoria-Migratorioides. Journal Of Insect Physiology 24: 737–741. [Google Scholar] [CrossRef]
- Gray, R. S., I. Roszko, and L. Solnica-Krezel. 2011. Planar Cell Polarity: Coordinating Morphogenetic Cell Behaviors With Embryonic Polarity. Dev Cell 21: 120–33. [Google Scholar] [CrossRef]
- Gumuskaya, G., N. Davey, P. Srivastava, A. Bender, L. Pio-Lopez, D. Hazel, and M. Levin. 2025. The Morphological, Behavioral, And Transcriptomic Life Cycle Of Anthrobots. Adv Sci (Weinh), E2409330. [Google Scholar] [CrossRef]
- Gumuskaya, G., P. Srivastava, B. G. Cooper, H. Lesser, B. Semegran, S. Garnier, and M. Levin. 2023. Motile Living Biobots Self-Construct From Adult Human Somatic Progenitor Seed Cells. Adv Sci (Weinh), E2303575. [Google Scholar] [CrossRef]
- Harris, A. K. 1987. Cell Motility And The Problem Of Anatomical Homeostasis. J Cell Sci Suppl 8: 121–40. [Google Scholar] [CrossRef] [PubMed]
- Harris, A. K. 2018. The Need For A Concept Of Shape Homeostasis. Biosystems 173: 65–72. [Google Scholar] [CrossRef] [PubMed]
- Harris, M. P. 2021. Bioelectric Signaling As A Unique Regulator Of Development And Regeneration. Development 148. [Google Scholar] [CrossRef] [PubMed]
- Hartl, B., and M. Levin. 2025. What Does Evolution Make? Learning In Living Lineages And Machines. Trends Genet 41: 480–496. [Google Scholar] [CrossRef]
- Hayflick, L. 2007. Biological Aging Is No Longer An Unsolved Problem. Ann N Y Acad Sci 1100: 1–13. [Google Scholar] [CrossRef]
- Headrick, J. P., L. E. See Hoe, E. F. Du Toit, and J. N. Peart. 2015. Opioid Receptors And Cardioprotection - 'Opioidergic Conditioning' Of The Heart. Br J Pharmacol 172: 2026–50. [Google Scholar] [CrossRef]
- Hochberg, L. R., D. Bacher, B. Jarosiewicz, N. Y. Masse, J. D. Simeral, J. Vogel, S. Haddadin, J. Liu, S. S. Cash, P. Van Der Smagt, and J. P. Donoghue. 2012. Reach And Grasp By People With Tetraplegia Using A Neurally Controlled Robotic Arm. Nature 485: 372–5. [Google Scholar] [CrossRef]
- Hofmann, B. 2017. Limits To Human Enhancement: Nature, Disease, Therapy Or Betterment? Bmc Med Ethics 18: 56. [Google Scholar] [CrossRef]
- Holtfreter, J. 1955. Transformation Of A Tail Into A Limb Or Gill-Like Structures. Journal Of Experimental Zoology 129: 623–648. [Google Scholar] [CrossRef]
- Illingworth, C. M. 1974. Trapped Fingers And Amputated Finger Tips In Children. J Pediatr Surg 9: 853–58. [Google Scholar] [CrossRef]
- Izgi, H., D. Han, U. Isildak, S. Huang, E. Kocabiyik, P. Khaitovich, M. Somel, and H. M. Donertas. 2022. Inter-Tissue Convergence Of Gene Expression During Ageing Suggests Age-Related Loss Of Tissue And Cellular Identity. Elife 11: E68048. [Google Scholar] [CrossRef]
- Jacobsen, M. H. 2017. Postmortal Society: Towards A Sociology Of Immortality. Taylor & Francis. [Google Scholar]
- Jacobson, A. L. 1963. Learning In Flatworms And Annelids. Psychological Bulletin 60: 74–94. [Google Scholar] [CrossRef]
- Johnson, S. G. B., A. Bilovich, and D. Tuckett. 2022. Conviction Narrative Theory: A Theory Of Choice Under Radical Uncertainty. Behav Brain Sci 46: E82. [Google Scholar] [CrossRef] [PubMed]
- Kaminishi, M., N. Shimizu, S. Shimoyama, H. Yamaguchi, E. Tsuji, F. Aoki, S. Nomura, A. Yoshikawa, S. Kuramoto, T. Oohara, K. Inada, and M. Tatematsu. 1997. Denervation Promotes The Development Of Cancer-Related Lesions In The Gastric Remnant. J Clin Gastroenterol 25 Suppl 1: S129–34. [Google Scholar] [CrossRef] [PubMed]
- Kamm, R. D., and R. Bashir. 2014. Creating Living Cellular Machines. Annals Of Biomedical Engineering 42: 445–59. [Google Scholar] [CrossRef] [PubMed]
- Kapsetaki, S. E., A. Pimkina, P. Mcmillen, P. Tayyebi, P. Erickson, and M. Levin. 2025. The Bioelectrics Of Immortality And Mortality In Cold-Sensitive Hydra Oligactis. Bioelectricity 7: 166–179. [Google Scholar] [CrossRef]
- Keresztes, D., M. Kerestely, L. Szarka, B. M. Kovacs, K. Schulc, D. V. Veres, and P. Csermely. 2025. Cancer Drug Resistance As Learning Of Signaling Networks. Biomed Pharmacother 183: 117880. [Google Scholar] [CrossRef]
- Kirkwood, T. B. 1989. Dna, Mutations And Aging. Mutat Res 219: 1–7. [Google Scholar] [CrossRef]
- Kirkwood, T. B., and S. Melov. 2011. On The Programmed/Non-Programmed Nature Of Ageing Within The Life History. Curr Biol 21: R701–7. [Google Scholar] [CrossRef]
- Kofman, K., and M. Levin. 2025. Robustness Of The Mind-Body Interface: Case Studies Of Unconventional Information Flow In The Multiscale Living Architecture. In Mind And Brain. In Press. [Google Scholar]
- Koseska, A., and P. I. Bastiaens. 2017. Cell Signaling As A Cognitive Process. Embo J 36: 568–582. [Google Scholar] [CrossRef]
- Ksander, B., M. Shah, D. Krasniqi, M. S. Gregory-Ksander, S. Rosenzweig-Lipson, K. Broniowska, M. Wathier, J. Mannick, J. Cermak, and M. Karg. 2023. Epigenetic Reprogramming-A Novel Gene Therapy That Restores Vision Loss In A Nonhuman Primate Model Of Naion. Investigative Ophthalmology & Visual Science 64: 474–474. [Google Scholar]
- Kunecki, M., T. Roleder, J. Biernat, P. Kukla, L. Tomkiewicz-Pajak, M. A. Deja, P. Podolec, K. S. Golba, and W. Plazak. 2018. Opioidergic Conditioning Of The Human Heart Muscle In Nitric Oxide-Dependent Mechanism. Adv Clin Exp Med 27: 1069–1073. [Google Scholar] [CrossRef] [PubMed]
- Lagasse, E., and M. Levin. 2023. Future Medicine: From Molecular Pathways To The Collective Intelligence Of The Body. Trends Mol Med.
- Lebedev, M. 2014. Brain-Machine Interfaces: An Overview. Translational Neuroscience 5: 99–110. [Google Scholar] [CrossRef]
- Levin, M. 2019. The Computational Boundary Of A “Self”: Developmental Bioelectricity Drives Multicellularity And Scale-Free Cognition. Frontiers In Psychology 10: 2688. [Google Scholar] [CrossRef]
- Levin, M. 2021a. Bioelectric Signaling: Reprogrammable Circuits Underlying Embryogenesis, Regeneration, And Cancer. Cell 184: 1971–1989. [Google Scholar] [CrossRef]
- Levin, M. 2021b. Bioelectrical Approaches To Cancer As A Problem Of The Scaling Of The Cellular Self. Prog Biophys Mol Biol 165: 102–113. [Google Scholar] [CrossRef]
- Levin, M. 2022. Technological Approach To Mind Everywhere: An Experimentally-Grounded Framework For Understanding Diverse Bodies And Minds. Frontiers In Systems Neuroscience 16: 768201. [Google Scholar] [CrossRef]
- Levin, M. 2023a. Bioelectric Networks: The Cognitive Glue Enabling Evolutionary Scaling From Physiology To Mind. In Anim Cogn. [Google Scholar]
- Levin, M. 2023b. Edited by P. A. Corning, S. A. Kauffman, D. Noble, J. A. Shapiro, R. I. Vane-Wright and A. Pross. Collective Intelligence Of Morphogenesis As A Teleonomic Process. In Evolution “On Purpose”: Teleonomy In Living Systems. Cambridge: Mit Press. [Google Scholar]
- Levin, M. 2023c. Darwin's Agential Materials: Evolutionary Implications Of Multiscale Competency In Developmental Biology. Cell Mol Life Sci 80: 142. [Google Scholar] [CrossRef]
- Levin, M. 2024a. The Multiscale Wisdom Of The Body: Collective Intelligence As A Tractable Interface For Next-Generation Biomedicine. Bioessays, E202400196. [Google Scholar] [CrossRef]
- Levin, M. 2024b. Self-Improvising Memory: A Perspective On Memories As Agential, Dynamically Reinterpreting Cognitive Glue. In Entropy (Basel). p. 26. [Google Scholar]
- Levin, M., and C. J. Martyniuk. 2018. The Bioelectric Code: An Ancient Computational Medium For Dynamic Control Of Growth And Form. Biosystems 164: 76–93. [Google Scholar] [CrossRef]
- Levin, M., G. Pezzulo, and J. M. Finkelstein. 2017. Endogenous Bioelectric Signaling Networks: Exploiting Voltage Gradients For Control Of Growth And Form. Annu Rev Biomed Eng 19: 353–387. [Google Scholar] [CrossRef]
- Levin, M., A. M. Pietak, and J. Bischof. 2018. Planarian Regeneration As A Model Of Anatomical Homeostasis: Recent Progress In Biophysical And Computational Approaches. Semin Cell Dev Biol 87: 125–144. [Google Scholar] [CrossRef]
- Levin, M., T. Thorlin, K. R. Robinson, T. Nogi, and M. Mercola. 2002. Asymmetries In H+/K+-Atpase And Cell Membrane Potentials Comprise A Very Early Step In Left-Right Patterning. Cell 111: 77–89. [Google Scholar] [CrossRef]
- Li, C. 2012. Deer Antler Regeneration: A Stem Cell-Based Epimorphic Process. Birth Defects Research. Part C, Embryo Today: Reviews 96: 51–62. [Google Scholar] [CrossRef] [PubMed]
- Lidsky, P. V., and R. Andino. 2020. Epidemics As An Adaptive Driving Force Determining Lifespan Setpoints. Proc Natl Acad Sci U S A 117: 17937–17948. [Google Scholar] [CrossRef] [PubMed]
- Lidsky, P. V., and R. Andino. 2022. Could Aging Evolve As A Pathogen Control Strategy? Trends Ecol Evol 37: 1046–1057. [Google Scholar] [CrossRef] [PubMed]
- Lidsky, P. V., J. Yuan, and R. Andino. 2023. Reconsidering Life History Theory Amid Infectious Diseases. Trends Ecol Evol 38: 699–700. [Google Scholar] [CrossRef]
- Lidsky, P. V., J. Yuan, J. M. Rulison, and R. Andino-Pavlovsky. 2022. Is Aging An Inevitable Characteristic Of Organic Life Or An Evolutionary Adaptation? Biochemistry (Mosc) 87: 1413–1445. [Google Scholar] [CrossRef]
- Lineweaver, C. H., and P. C. W. Davies. 2021. Comparison Of The Atavistic Model Of Cancer To Somatic Mutation Theory: Phylostratigraphic Analyses Support The Atavistic Model. In The Physics Of Cancer: Research Advances. World Scientific. [Google Scholar]
- Loftus, E. F. 2005. Planting Misinformation In The Human Mind: A 30-Year Investigation Of The Malleability Of Memory. Learn Mem 12: 361–6. [Google Scholar] [CrossRef]
- Lopez-Otin, C., M. A. Blasco, L. Partridge, M. Serrano, and G. Kroemer. 2023. Hallmarks Of Aging: An Expanding Universe. Cell 186: 243–278. [Google Scholar] [CrossRef]
- Lutz, H. 1949. Sur La Production Experimentale De La Polyembryonie Et De La Monstruosite Double Chez Les Oiseaux. Archs Anat. Microsc. Morph. Exp. 38: 79–144. [Google Scholar]
- Lyon, P. 2006. The Biogenic Approach To Cognition. Cogn Process 7: 11–29. [Google Scholar] [CrossRef] [PubMed]
- Lyon, P. 2015. The Cognitive Cell: Bacterial Behavior Reconsidered. Front Microbiol 6: 264. [Google Scholar] [CrossRef]
- Macpherson, F. 2018. Sensory Substitution And Augmentation: An Introduction. [Google Scholar]
- Marin, F., and C. Beluffi. 2018. Computing The Minimal Crew For A Multi-Generational Space Travel Towards Proxima Centauri B. Arxiv Preprint Arxiv, 1806.03856. [Google Scholar]
- Martinez-Corral, R., J. Liu, A. Prindle, G. M. Suel, and J. Garcia-Ojalvo. 2019. Metabolic Basis Of Brain-Like Electrical Signalling In Bacterial Communities. Philos Trans R Soc Lond B Biol Sci 374: 20180382. [Google Scholar] [CrossRef]
- Mathews, J., A. J. Chang, L. Devlin, and M. Levin. 2023. Cellular Signaling Pathways As Plastic, Proto-Cognitive Systems: Implications For Biomedicine. Patterns (N Y) 4: 100737. [Google Scholar] [CrossRef]
- Maturana, H. R., and F. J. Varela. 1980. Autopoiesis And Cognition The Realization Of The Living. In Boston Studies In The Philosophy Of Science,. Dordrecht: Springer Netherlands. [Google Scholar]
- Mcmillen, P., and M. Levin. 2024. Collective Intelligence: A Unifying Concept For Integrating Biology Across Scales And Substrates. Commun Biol 7: 378. [Google Scholar] [CrossRef]
- Mcshea, D. W. 2012. Upper-Directed Systems: A New Approach To Teleology In Biology. Biology & Philosophy 27: 663–684. [Google Scholar] [CrossRef]
- Medawar, P. B. 1952. An Unsolved Problem Of Biology. [Google Scholar]
- Mehlman, M. J., R. A. Conlon, and A. Pearlman. 2023. Governing Nonconventional Genetic Experimentation. Journal Of Law And The Biosciences 10: Lsad003. [Google Scholar] [CrossRef]
- Miller, W. B., Jr., F. Baluska, and A. S. Reber. 2023. A Revised Central Dogma For The 21st Century:All Biology Is Cognitive Information Processing. Prog Biophys Mol Biol 182: 34–48. [Google Scholar] [CrossRef]
- Miller, W. B., Jr., F. Baluska, A. S. Reber, and P. Slijepcevic. 2024. Why Death And Aging ? All Memories Are Imperfect. Prog Biophys Mol Biol 187: 21–35. [Google Scholar] [CrossRef]
- Mitchell, A., and W. Lim. 2016. Cellular Perception And Misperception: Internal Models For Decision-Making Shaped By Evolutionary Experience. Bioessays 38: 845–9. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, S. R., J. M. Beaton, and R. J. Bradley. 1975. Biochemical Transfer Of Acquired Information. International Review Of Neurobiology 17: 61–83. [Google Scholar] [PubMed]
- Mitteldorf, J. 2017. Aging Is A Group-Selected Adaptation: Theory, Evidence, And Medical Implications. Crc Press. [Google Scholar]
- Mohamed, N., M. S. Solanki, H. D. Praveena, A. Princy, S. Das, and D. Verma. 2023. Artificial Intelligence Integrated Biomedical Implants System Developments In Healthcare. vol. Ieee, pp. 588–591. [Google Scholar]
- Montague, P. R., R. J. Dolan, K. J. Friston, and P. Dayan. 2012. Computational Psychiatry. Trends Cogn Sci 16: 72–80. [Google Scholar] [CrossRef] [PubMed]
- Moore, D., S. I. Walker, and M. Levin. 2017. Cancer As A Disorder Of Patterning Information: Computational And Biophysical Perspectives On The Cancer Problem. Convergent Science Physical Oncology 3: 043001. [Google Scholar] [CrossRef]
- Nagai, H., Y. Adachi, T. Nakasugi, E. Takigawa, J. Ui, T. Makino, M. Miura, and Y. I. Nakajima. 2024. Highly Regenerative Species-Specific Genes Improve Age-Associated Features In The Adult Drosophila Midgut. Bmc Biol 22: 157. [Google Scholar] [CrossRef]
- Nagato, T., K. Matsumoto, H. Tanioka, J. Kodama, and H. Toh. 1995. Effect Of Denervation On Morphogenesis Of The Rat Fungiform Papilla. Acta Anat (Basel) 153: 301–9. [Google Scholar] [CrossRef]
- Nagel, S. K., C. Carl, T. Kringe, R. Martin, and P. Konig. 2005. Beyond Sensory Substitution--Learning The Sixth Sense. J Neural Eng 2: R13–26. [Google Scholar] [CrossRef]
- Nanos, V., and M. Levin. 2022. Multi-Scale Chimerism: An Experimental Window On The Algorithms Of Anatomical Control. Cells Dev 169: 203764. [Google Scholar] [CrossRef]
- Nesbeth, D. N., A. Zaikin, Y. Saka, M. C. Romano, C. V. Giuraniuc, O. Kanakov, and T. Laptyeva. 2016. Synthetic Biology Routes To Bio-Artificial Intelligence. Essays Biochem 60: 381–391. [Google Scholar]
- Nieto-Diaz, M., D. W. Pita-Thomas, T. Munoz-Galdeano, C. Martinez-Maza, R. Navarro-Ruiz, D. Reigada, M. Yunta, M. J. Caballero-Lopez, M. Nieto-Sampedro, and R. Martinez-Maza. 2012. Deer Antler Innervation And Regeneration. Frontiers In Bioscience 17: 1389–401. [Google Scholar] [CrossRef]
- Nogi, T., and M. Levin. 2005. Characterization Of Innexin Gene Expression And Functional Roles Of Gap-Junctional Communication In Planarian Regeneration. Dev Biol 287: 314–35. [Google Scholar] [CrossRef]
- Normann, R. A., B. Greger, P. House, S. F. Romero, F. Pelayo, and E. Fernandez. 2009. Toward The Development Of A Cortically Based Visual Neuroprosthesis. J Neural Eng 6: 035001. [Google Scholar] [CrossRef] [PubMed]
- Onken, B., and M. Driscoll. 2010. Metformin Induces A Dietary Restriction-Like State And The Oxidative Stress Response To Extend C. Elegans Healthspan Via Ampk, Lkb1, And Skn-1. Plos One 5: E8758. [Google Scholar] [CrossRef] [PubMed]
- Otgaar, H., O. Dodier, M. Garry, M. L. Howe, E. F. Loftus, S. J. Lynn, I. Mangiulli, R. J. Mcnally, and L. Patihis. 2023. Oversimplifications And Misrepresentations In The Repressed Memory Debate: A Reply To Ross. J Child Sex Abus 32: 116–126. [Google Scholar] [CrossRef] [PubMed]
- Outzen, H. C., R. P. Custer, and R. T. Prehn. 1976. Influence Of Regenerative Capacity And Innervation On Oncogenesis In The Adult Frog (Rana Pipiens). J Natl Cancer Inst 57: 79–84. [Google Scholar] [CrossRef]
- Pagán, O. R. 2014. The First Brain: The Neuroscience Of Planarians. [Google Scholar]
- Pai, V. P., J. M. Lemire, J. F. Pare, G. Lin, Y. Chen, and M. Levin. 2015. Endogenous Gradients Of Resting Potential Instructively Pattern Embryonic Neural Tissue Via Notch Signaling And Regulation Of Proliferation. The Journal Of Neuroscience 35: 4366–85. [Google Scholar] [CrossRef]
- Pai, V. P., and M. Levin. 2022. Hcn2 Channel-Induced Rescue Of Brain, Eye, Heart And Gut Teratogenesis Caused By Nicotine, Ethanol And Aberrant Notch Signalling. Wound Repair Regen.
- Pawlowski, A., and G. Weddell. 1967. Induction Of Tumours In Denervated Skin. Nature 213: 1234–1236. [Google Scholar] [CrossRef]
- Perrotta, M. V., T. Asgeirsdottir, and D. M. Eagleman. 2021. Deciphering Sounds Through Patterns Of Vibration On The Skin. Neuroscience 458: 77–86. [Google Scholar] [CrossRef]
- Pezzulo, G., and M. Levin. 2015. Re-Membering The Body: Applications Of Computational Neuroscience To The Top-Down Control Of Regeneration Of Limbs And Other Complex Organs. Integr Biol (Camb) 7: 1487–517. [Google Scholar] [CrossRef]
- Pezzulo, G., and M. Levin. 2016. Top-Down Models In Biology: Explanation And Control Of Complex Living Systems Above The Molecular Level. J R Soc Interface 13. [Google Scholar] [CrossRef] [PubMed]
- Piedimonte, A., and F. Benedetti. 2016. Words And Drugs: Same Mechanisms Of Action? Journal Of Contemporary Psychotherapy 46: 159–166. [Google Scholar] [CrossRef]
- Pietak, A., and M. Levin. 2017. Bioelectric Gene And Reaction Networks: Computational Modelling Of Genetic, Biochemical And Bioelectrical Dynamics In Pattern Regulation. J R Soc Interface 14. [Google Scholar] [CrossRef] [PubMed]
- Pilleron, S., D. Sarfati, M. Janssen-Heijnen, J. Vignat, J. Ferlay, F. Bray, and I. Soerjomataram. 2019. Global Cancer Incidence In Older Adults, 2012 And 2035: A Population-Based Study. Int J Cancer 144: 49–58. [Google Scholar] [CrossRef]
- Pinquart, M. 2002. Creating And Maintaining Purpose In Life In Old Age: A Meta-Analysis. Ageing International 27: 90–114. [Google Scholar] [CrossRef]
- Pio-Lopez, L. 2021. The Rise Of The Biocyborg: Synthetic Biology, Artificial Chimerism And Human Enhancement. New Genetics And Society 40: 599–619. [Google Scholar] [CrossRef]
- Pio-Lopez, L., B. Hartl, and M. Levin. 2025. Aging As A Loss Of Goal-Directedness: An Evolutionary Simulation And Analysis Unifying Regeneration With Anatomical Rejuvenation. Adv Sci (Weinh) 12: E09872. [Google Scholar] [CrossRef]
- Pio-Lopez, L., F. Kuchling, A. Tung, G. Pezzulo, and M. Levin. 2022. Active Inference, Morphogenesis, And Computational Psychiatry. Front Comput Neurosci 16: 988977. [Google Scholar] [CrossRef]
- Pio-Lopez, L., and M. Levin. 2023. Morphoceuticals: Perspectives For Discovery Of Drugs Targeting Anatomical Control Mechanisms In Regenerative Medicine, Cancer And Aging. Drug Discov Today 28: 103585. [Google Scholar] [CrossRef]
- Pio-Lopez, L., and M. Levin. 2024. Aging As A Loss Of Morphostatic Information: A Developmental Bioelectricity Perspective. Ageing Res Rev 97: 102310. [Google Scholar] [CrossRef]
- Pio-Lopez, L., and M. Levin. 2026. Atavistic Genetic Expression Dissociation (Aged) During Aging: Meta-Phylostratigraphic Evidence Of Cellular And Tissue-Level Phylogenetic Dissociation. Aging Cell 25: E70305. [Google Scholar] [CrossRef]
- Pio-Lopez, L., R. Poulkouras, and D. Depannemaecker. 2021. Visual Cortical Prosthesis: An Electrical Perspective. J Med Eng Technol 45: 394–407. [Google Scholar] [CrossRef] [PubMed]
- Prindle, A., J. Liu, M. Asally, S. Ly, J. Garcia-Ojalvo, and G. M. Suel. 2015. Ion Channels Enable Electrical Communication In Bacterial Communities. Nature 527: 59–63. [Google Scholar] [CrossRef] [PubMed]
- Rahbaran, M., E. Razeghian, M. S. Maashi, A. T. Jalil, G. Widjaja, L. Thangavelu, M. Y. Kuznetsova, P. Nasirmoghadas, F. Heidari, F. Marofi, and M. Jarahian. 2021. Cloning And Embryo Splitting In Mammalians: Brief History, Methods, And Achievements. Stem Cells Int, 2347506. [Google Scholar] [CrossRef] [PubMed]
- Razgonova, M. P., A. M. Zakharenko, K. S. Golokhvast, M. Thanasoula, E. Sarandi, K. Nikolouzakis, P. Fragkiadaki, D. Tsoukalas, D. A. Spandidos, and A. Tsatsakis. 2020. Telomerase And Telomeres In Aging Theory And Chronographic Aging Theory (Review). Mol Med Rep 22: 1679–1694. [Google Scholar] [CrossRef]
- Reese, H. W. 1994. Teleology And Teleonomy In Behavior Analysis. Behav Anal 17: 75–91. [Google Scholar] [CrossRef]
- Regalado, A. 2015. A Tale Of Do-It-Yourself Gene Therapy. Mit Technology Review 11. Technologyreview. Com/2015/10/14/165802/A-Taleof-Do-It-Yourself-Gene-Therapy/Erişim Tarihi. Available online: Https://Www.
- Restrepo, H. E., and M. Rozental. 1994. The Social Impact Of Aging Populations: Some Major Issues. Soc Sci Med 39: 1323–38. [Google Scholar] [CrossRef]
- Rosenblueth, A., N. Wiener, and J. Bigelow. 1943. Behavior, Purpose, And Teleology. Philosophy Of Science 10: 18–24. [Google Scholar] [CrossRef]
- Rubin, H. 1985. Cancer As A Dynamic Developmental Disorder. Cancer Res 45: 2935–42. [Google Scholar]
- Rubin, H. 2006. What Keeps Cells In Tissues Behaving Normally In The Face Of Myriad Mutations? Bioessays 28: 515–24. [Google Scholar] [CrossRef]
- Rubin, H. 2007. Ordered Heterogeneity And Its Decline In Cancer And Aging. Advances In Cancer Research 98: 117–47. [Google Scholar] [PubMed]
- Rubin, H., M. Chow, and A. Yao. 1996. Cellular Aging, Destabilization, And Cancer. Proc Natl Acad Sci U S A 93: 1825–30. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, E., S. Maris, and P. Bach-Y-Rita. 2001. Brain Plasticity: 'Visual' Acuity Of Blind Persons Via The Tongue. Brain Res 908: 204–7. [Google Scholar] [CrossRef]
- Sarnat, H. B., and M.G. And Netsky. 1985. The Brain Of The Planarian As The Ancestor Of The Human Brain. Canadian Journal Of Neurological Sciences 12: 296–302. [Google Scholar] [CrossRef] [PubMed]
- Sediqi, H., and M. Levin. 2025. Bioelectric Characterization Of Senescing Human Keratinocytes. Iscience 28: 113275. [Google Scholar] [CrossRef]
- Sennesh, E., J. Theriault, D. Brooks, J. W. Van De Meent, L. F. Barrett, and K. S. Quigley. 2022. Interoception As Modeling, Allostasis As Control. Biol Psychol 167: 108242. [Google Scholar] [CrossRef]
- Shao, X., C. Chen, E. F. Loftus, G. Xue, and B. Zhu. 2022. Dynamic Changes In Neural Representations Underlie The Repetition Effect On False Memory. Neuroimage 259: 119442. [Google Scholar] [CrossRef]
- Sheiman, I. M., and K. L. Tiras. 1996. Edited by C. I. Abramson, Z. P. Shuranova and Y. M. Burmistrov. Memory And Morphogenesis In Planaria And Beetle. In Russian Contributions To Invertebrate Behavior. Westport, Ct: Praeger. [Google Scholar]
- Shull, P. B., and D. D. Damian. 2015. Haptic Wearables As Sensory Replacement, Sensory Augmentation And Trainer - A Review. J Neuroeng Rehabil 12: 59. [Google Scholar] [CrossRef]
- Skulachev, M. V., and V. P. Skulachev. 2014. New Data On Programmed Aging—Slow Phenoptosis. Biochemistry (Moscow) 79: 977–993. [Google Scholar] [CrossRef]
- Solms, M. 2023. The Statistical Mechanics Of Felt Uncertainty Under Active Inference. Behav Brain Sci 46: E108. [Google Scholar] [CrossRef]
- Stano, P., Y. Kuruma, and L. Damiano. 2018. Synthetic Biology And (Embodied) Artificial Intelligence: Opportunities And Challenges. Adaptive Behavior 26: 41–44. [Google Scholar] [CrossRef]
- Takeda, M., Y. Suzuki, N. Obara, and Y. Nagai. 1996. Apoptosis In Mouse Taste Buds After Denervation. Cell Tissue Res 286: 55–62. [Google Scholar] [CrossRef] [PubMed]
- Tang, B., Z. Li, S. Hu, and J. Xiong. 2022. Economic Implications Of Health Care Burden For Elderly Population. Inquiry 59: 469580221121511. [Google Scholar] [CrossRef] [PubMed]
- Teufel, T. 2011. Wholes That Cause Their Parts: Organic Self-Reproduction And The Reality Of Biological Teleology. Studies In History And Philosophy Of Biological And Biomedical Sciences 42: 252–60. [Google Scholar] [CrossRef] [PubMed]
- Thakkar, J. P., B. J. Mccarthy, and J. L. Villano. 2014. Age-Specific Cancer Incidence Rates Increase Through The Oldest Age Groups. Am J Med Sci 348: 65–70. [Google Scholar] [CrossRef]
- Tschantz, A., L. Barca, D. Maisto, C. L. Buckley, A. K. Seth, and G. Pezzulo. 2022. Simulating Homeostatic, Allostatic And Goal-Directed Forms Of Interoceptive Control Using Active Inference. Biol Psychol 169: 108266. [Google Scholar] [CrossRef]
- Tsonis, P. A. 1987. Embryogenesis And Carcinogenesis: Order And Disorder. Anticancer Res 7: 617–23. [Google Scholar]
- Vandenberg, L. N., D. S. Adams, and M. Levin. 2012. Normalized Shape And Location Of Perturbed Craniofacial Structures In The Xenopus Tadpole Reveal An Innate Ability To Achieve Correct Morphology. Developmental Dynamics 241: 863–78. [Google Scholar] [CrossRef]
- Vandenberg, L. N., R. D. Morrie, and D. S. Adams. 2011. V-Atpase-Dependent Ectodermal Voltage And Ph Regionalization Are Required For Craniofacial Morphogenesis. Dev Dyn 240: 1889–904. [Google Scholar] [CrossRef]
- Vazza, F., and A. Feletti. 2020. The Quantitative Comparison Between The Neuronal Network And The Cosmic Web. Frontiers In Physics 8: 525731. [Google Scholar] [CrossRef]
- Veres, T., M. Kerestely, B. M. Kovacs, D. Keresztes, K. Schulc, E. Seitz, Z. Vassy, D. V. Veres, and P. Csermely. 2024. Cellular Forgetting, Desensitisation, Stress And Ageing In Signalling Networks. When Do Cells Refuse To Learn More? Cell Mol Life Sci 81: 97. [Google Scholar] [CrossRef] [PubMed]
- Voeikov, V. L., and L. V. Beloussov. 2007. Edited by L. V. Beloussov, V. L. Voeikov and V. S. Martynyuk. From Mitogenetic Rays To Biophotons [English]. In Biophotonics And Coherent Systems In Biology. New York: Springer. [Google Scholar]
- Wang, Y., and L. Guo. 2016. Nanomaterial-Enabled Neural Stimulation. Front Neurosci 10: 69. [Google Scholar] [CrossRef] [PubMed]
- Willett, F. R., D. T. Avansino, L. R. Hochberg, J. M. Henderson, and K. V. Shenoy. 2021. High-Performance Brain-To-Text Communication Via Handwriting. Nature 593: 249–254. [Google Scholar] [CrossRef]
- Wolsky, A. 1978. Regeneration And Cancer. Growth 42: 425–6. [Google Scholar]
- Wright, J., V. G. Macefield, A. Van Schaik, and J. C. Tapson. 2016. A Review Of Control Strategies In Closed-Loop Neuroprosthetic Systems. Front Neurosci 10: 312. [Google Scholar] [CrossRef]
- Yang, J. H., M. Hayano, P. T. Griffin, J. A. Amorim, M. S. Bonkowski, J. K. Apostolides, E. L. Salfati, M. Blanchette, E. M. Munding, M. Bhakta, Y. C. Chew, W. Guo, X. Yang, S. Maybury-Lewis, X. Tian, J. M. Ross, G. Coppotelli, M. V. Meer, R. Rogers-Hammond, D. L. Vera, Y. R. Lu, J. W. Pippin, M. L. Creswell, Z. Dou, C. Xu, S. J. Mitchell, A. Das, B. L. O'connell, S. Thakur, A. E. Kane, Q. Su, Y. Mohri, E. K. Nishimura, L. Schaevitz, N. Garg, A. M. Balta, M. A. Rego, M. Gregory-Ksander, T. C. Jakobs, L. Zhong, H. Wakimoto, J. El Andari, D. Grimm, R. Mostoslavsky, A. J. Wagers, K. Tsubota, S. J. Bonasera, C. M. Palmeira, J. G. Seidman, C. E. Seidman, N. S. Wolf, J. A. Kreiling, J. M. Sedivy, G. F. Murphy, R. E. Green, B. A. Garcia, S. L. Berger, P. Oberdoerffer, S. J. Shankland, V. N. Gladyshev, B. R. Ksander, A. R. Pfenning, L. A. Rajman, and D. A. Sinclair. 2023. Loss Of Epigenetic Information As A Cause Of Mammalian Aging. Cell 186: 305–326 E27. [Google Scholar] [CrossRef]
- Yum, K., S. G. Hong, K. E. Healy, and L. P. Lee. 2014. Physiologically Relevant Organs On Chips. Biotechnol J 9: 16–27. [Google Scholar] [CrossRef]
Figure 1.
The origin story of embodied Selves. (A) Our smooth, continuous transformation, from the chemicals in a quiescent oocyte to that of a complex meta-cognitive human embedded in a social milieu, ranges across numerous disciplines. These disciplines are not truly binary (reflecting no discontinuities of substrate) but represent convenient names for sets of tools (from physical rewiring to cybernetics, behavioral science, psychoanalysis, etc.) that can be used to interact with a system. (B) Developing as “an embryo” requires a demarcation process in which a group of cells aligns toward a specific outcome in anatomical space, separating itself from the outside world via a boundary of states it will actively manage with respect to setpoints in physiological, transcriptional, and morpho-space. A single blastoderm can be partitioned into a number of different self-organizing islands, showing that the number of individuals within a living medium is variable. This process of self-construction, held together by a target morphology self-model, is relevant throughout our life history long after development, guiding maintenance, cancer suppression, and the future possibilities of subsequent transformation.
Figure 1.
The origin story of embodied Selves. (A) Our smooth, continuous transformation, from the chemicals in a quiescent oocyte to that of a complex meta-cognitive human embedded in a social milieu, ranges across numerous disciplines. These disciplines are not truly binary (reflecting no discontinuities of substrate) but represent convenient names for sets of tools (from physical rewiring to cybernetics, behavioral science, psychoanalysis, etc.) that can be used to interact with a system. (B) Developing as “an embryo” requires a demarcation process in which a group of cells aligns toward a specific outcome in anatomical space, separating itself from the outside world via a boundary of states it will actively manage with respect to setpoints in physiological, transcriptional, and morpho-space. A single blastoderm can be partitioned into a number of different self-organizing islands, showing that the number of individuals within a living medium is variable. This process of self-construction, held together by a target morphology self-model, is relevant throughout our life history long after development, guiding maintenance, cancer suppression, and the future possibilities of subsequent transformation.
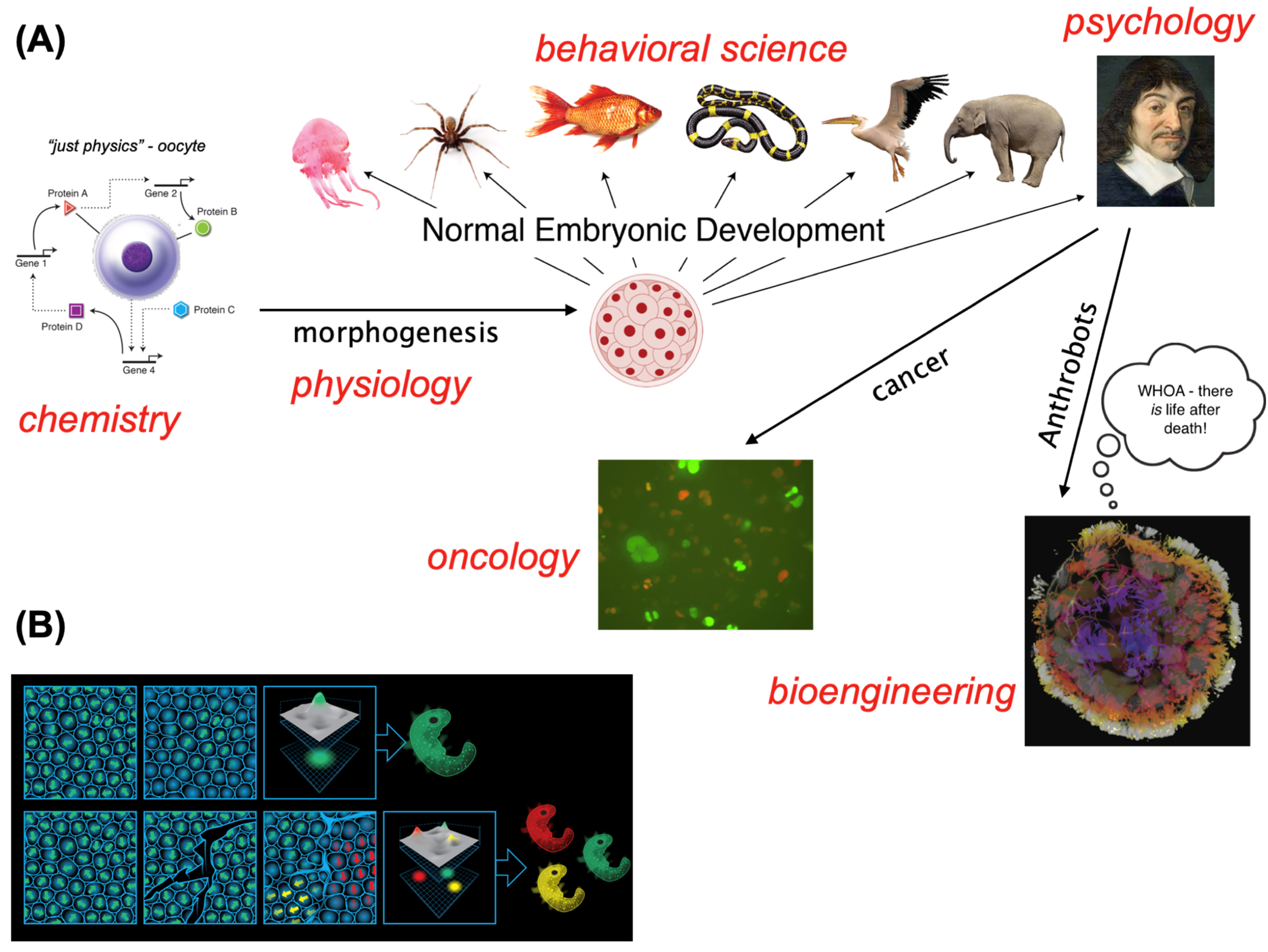
Figure 2.
Multi-scale control of living matter. (A) When a tail is grafted into the flank of an amphibian, it metamorphoses into a limb – the structure more appropriate to that location (Farinella-Ferruzza, 1956, Holtfreter, 1955). The cells at the tip of the tail (marked in red) turn into fingers even though there is no local damage or injury. They do it because the error-correcting process at the organism level propagates influence down to the cell- and molecular-level processes, which know nothing of large-scale target morphology but end up taking actions which serve its endpoint. (B) Similarly in voluntary motion, the chemistry of ions at muscle cell membranes effectively changes in response to high-level abstract goals in financial, social, and other spaces because the body architecture is an effective transducer of influence across scales. (C) The increasing understanding of the physiological layers linking chemistry to cognition is giving rise to technologies that allow communication and control of complex states well beyond the molecular level (Levin, 2024a). Panel C by Jeremy Guay of Peregrine Creative.
Figure 2.
Multi-scale control of living matter. (A) When a tail is grafted into the flank of an amphibian, it metamorphoses into a limb – the structure more appropriate to that location (Farinella-Ferruzza, 1956, Holtfreter, 1955). The cells at the tip of the tail (marked in red) turn into fingers even though there is no local damage or injury. They do it because the error-correcting process at the organism level propagates influence down to the cell- and molecular-level processes, which know nothing of large-scale target morphology but end up taking actions which serve its endpoint. (B) Similarly in voluntary motion, the chemistry of ions at muscle cell membranes effectively changes in response to high-level abstract goals in financial, social, and other spaces because the body architecture is an effective transducer of influence across scales. (C) The increasing understanding of the physiological layers linking chemistry to cognition is giving rise to technologies that allow communication and control of complex states well beyond the molecular level (Levin, 2024a). Panel C by Jeremy Guay of Peregrine Creative.
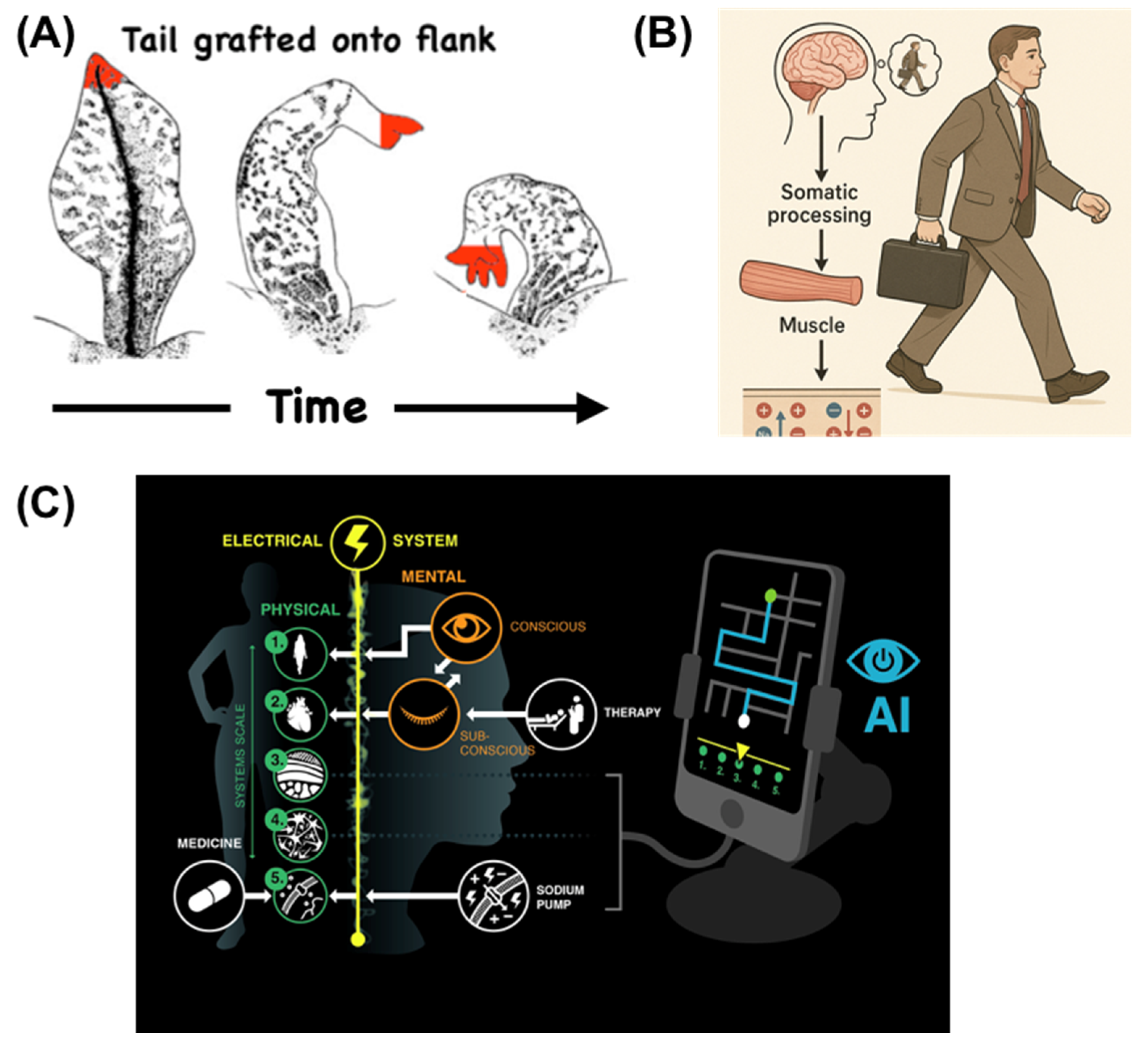
Figure 3.
A bowtie architecture and genetic/behavioral memory bottlenecks that force improvisation. (A) An embodied mind persisting across time is comprised of a set of “Selflets” – snapshots of time during which the agent has no access to the past. Instead, we have access to compressed biophysical encodings of past experiences into engrams. Inevitably, these memories must be actively and continuously interpreted. Because the process of encoding loses information (discards details and correlations), the reconstruction process is creative (Otgaar et al., 2023, Shao et al., 2022, Loftus, 2005) – confabulatory, improvisational, seeking to maintain saliency and adaptive power for the future in terms of large-scale goals and motivations, not accuracy with respect to low-level details of the past (Levin, 2024b). (B) The origin of this system, allowing immense plasticity, is the ancient need for morphogenesis to interpret genomic information actively, not literally, because both genomes and environment change unpredictably. Each embryo does not receive the actual form and function of their ancestors, but a parts list of molecular hardware that is then used to construct a coherent organism under current circumstances. This is why subtle physiological prompts (Nogi and Levin, 2005) can induce perfectly viable planarian flatworms with 2-heads (C) and why tadpoles whose only eye was induced to form on their tails (D) can see out of those eyes without new rounds of selection and adaptation to the new sensory-motor architecture (Blackiston and Levin, 2013). Panels A,B by Jeremy Guay of Peregrine Creative.
Figure 3.
A bowtie architecture and genetic/behavioral memory bottlenecks that force improvisation. (A) An embodied mind persisting across time is comprised of a set of “Selflets” – snapshots of time during which the agent has no access to the past. Instead, we have access to compressed biophysical encodings of past experiences into engrams. Inevitably, these memories must be actively and continuously interpreted. Because the process of encoding loses information (discards details and correlations), the reconstruction process is creative (Otgaar et al., 2023, Shao et al., 2022, Loftus, 2005) – confabulatory, improvisational, seeking to maintain saliency and adaptive power for the future in terms of large-scale goals and motivations, not accuracy with respect to low-level details of the past (Levin, 2024b). (B) The origin of this system, allowing immense plasticity, is the ancient need for morphogenesis to interpret genomic information actively, not literally, because both genomes and environment change unpredictably. Each embryo does not receive the actual form and function of their ancestors, but a parts list of molecular hardware that is then used to construct a coherent organism under current circumstances. This is why subtle physiological prompts (Nogi and Levin, 2005) can induce perfectly viable planarian flatworms with 2-heads (C) and why tadpoles whose only eye was induced to form on their tails (D) can see out of those eyes without new rounds of selection and adaptation to the new sensory-motor architecture (Blackiston and Levin, 2013). Panels A,B by Jeremy Guay of Peregrine Creative.
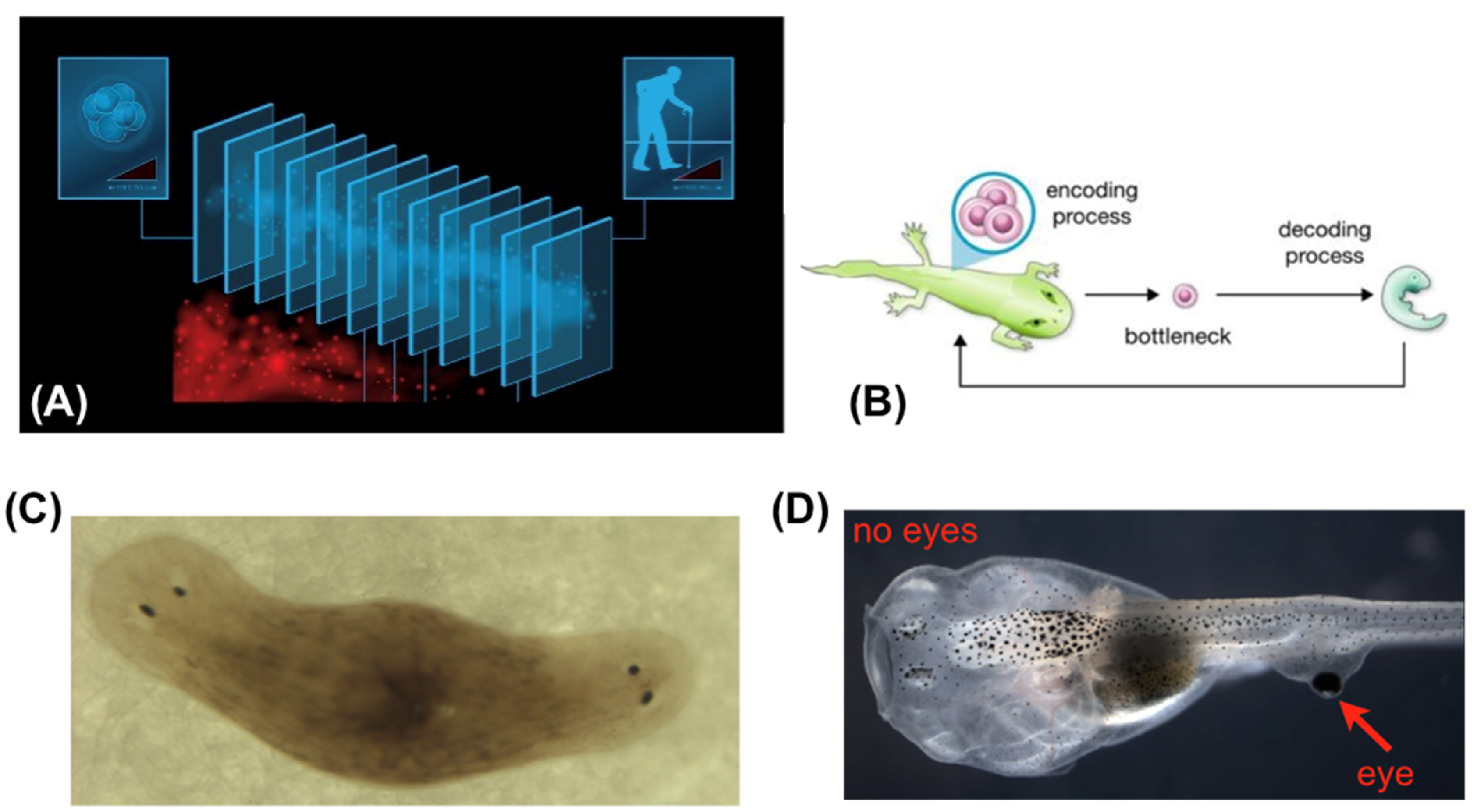
Figure 4.
The body as a Ship of Theseus, beset by storms both small and great. (A) Due to the constant turnover of molecules and cells in the body, persistence requires constant active maintenance, guiding new cells into a large-scale structure that resists degeneration, cancer, and aging over some time period. Personal identity, as mental content changes over one’s life, and one’s changing body, persist as processes, not objects, by virtue of this active process. The actual Ship of Theseus is not the boards, but the large-scale plan of the ship in the mind of the repair workers; similarly, the structure of the body is not in the hardware encoded by the DNA but the pattern memories stored by physiological networks of cells that enable the collective to control molecular pathways in accordance to a body-scale anatomical specification. Those are the processes that can be targeted for transformative regenerative applications, including longevity. Examples of large-scale challenges include amputation of limbs (which regenerate in amphibians regardless of the location of amputation and stop precisely when a correct limb has been produced) (B), and duplications of the genome, which produce polyploid newts with enormous cells but kidney tubules sized correctly due to the cells’ ability to adapt to an unexpected change in their hardware and either use fewer cells, or single giant cells that wrap around themselves to leave a lumen(Fankhauser, 1945). Images by Jeremy Guay of Peregrine Creative.
Figure 4.
The body as a Ship of Theseus, beset by storms both small and great. (A) Due to the constant turnover of molecules and cells in the body, persistence requires constant active maintenance, guiding new cells into a large-scale structure that resists degeneration, cancer, and aging over some time period. Personal identity, as mental content changes over one’s life, and one’s changing body, persist as processes, not objects, by virtue of this active process. The actual Ship of Theseus is not the boards, but the large-scale plan of the ship in the mind of the repair workers; similarly, the structure of the body is not in the hardware encoded by the DNA but the pattern memories stored by physiological networks of cells that enable the collective to control molecular pathways in accordance to a body-scale anatomical specification. Those are the processes that can be targeted for transformative regenerative applications, including longevity. Examples of large-scale challenges include amputation of limbs (which regenerate in amphibians regardless of the location of amputation and stop precisely when a correct limb has been produced) (B), and duplications of the genome, which produce polyploid newts with enormous cells but kidney tubules sized correctly due to the cells’ ability to adapt to an unexpected change in their hardware and either use fewer cells, or single giant cells that wrap around themselves to leave a lumen(Fankhauser, 1945). Images by Jeremy Guay of Peregrine Creative.
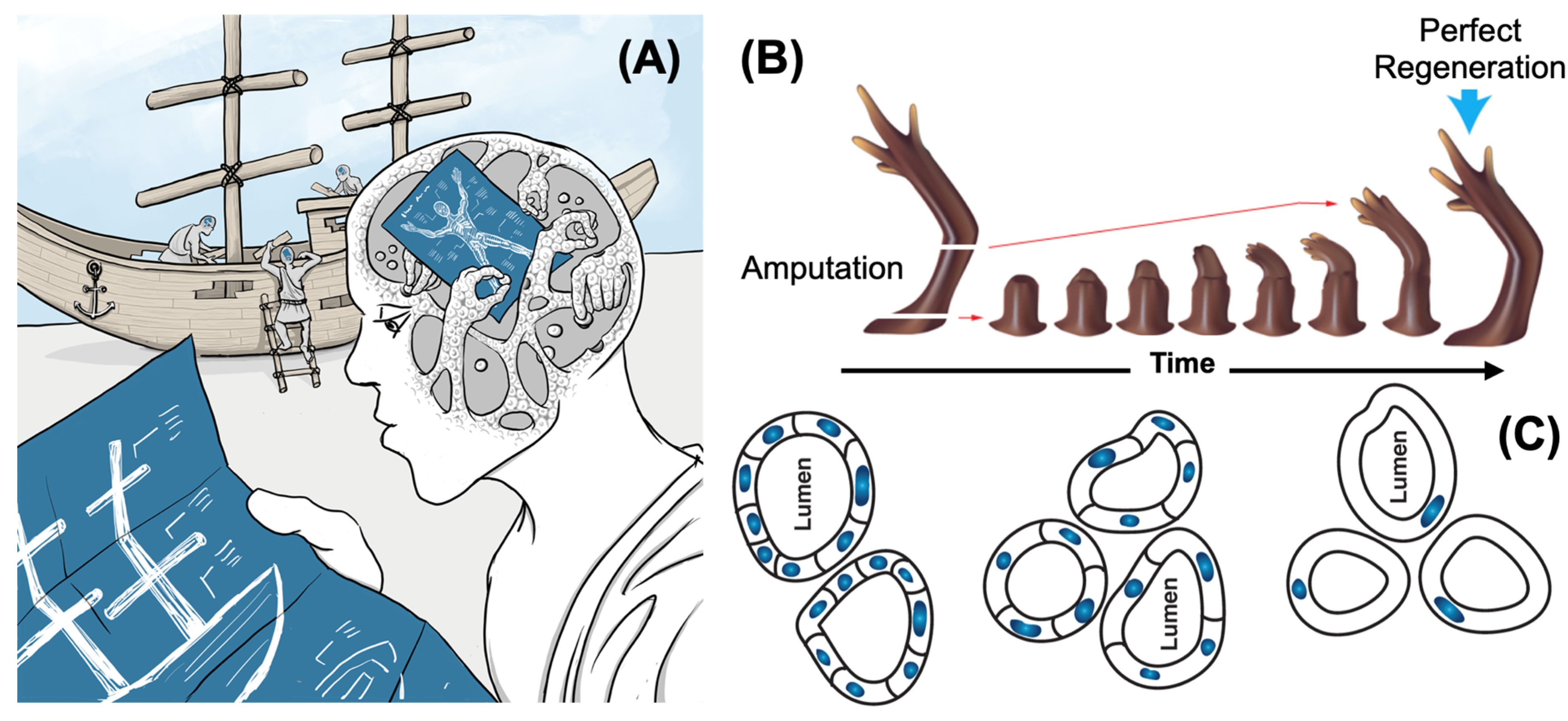
Figure 5.
metamorphosis as a singularity. Caterpillars largely deconstruct their brain, building a new one suitable for an entirely new body, but memories not only persist but are remapped onto a new embodiment and lifestyle. Image by Jeremy Guay of Peregrine Creative.
Figure 5.
metamorphosis as a singularity. Caterpillars largely deconstruct their brain, building a new one suitable for an entirely new body, but memories not only persist but are remapped onto a new embodiment and lifestyle. Image by Jeremy Guay of Peregrine Creative.
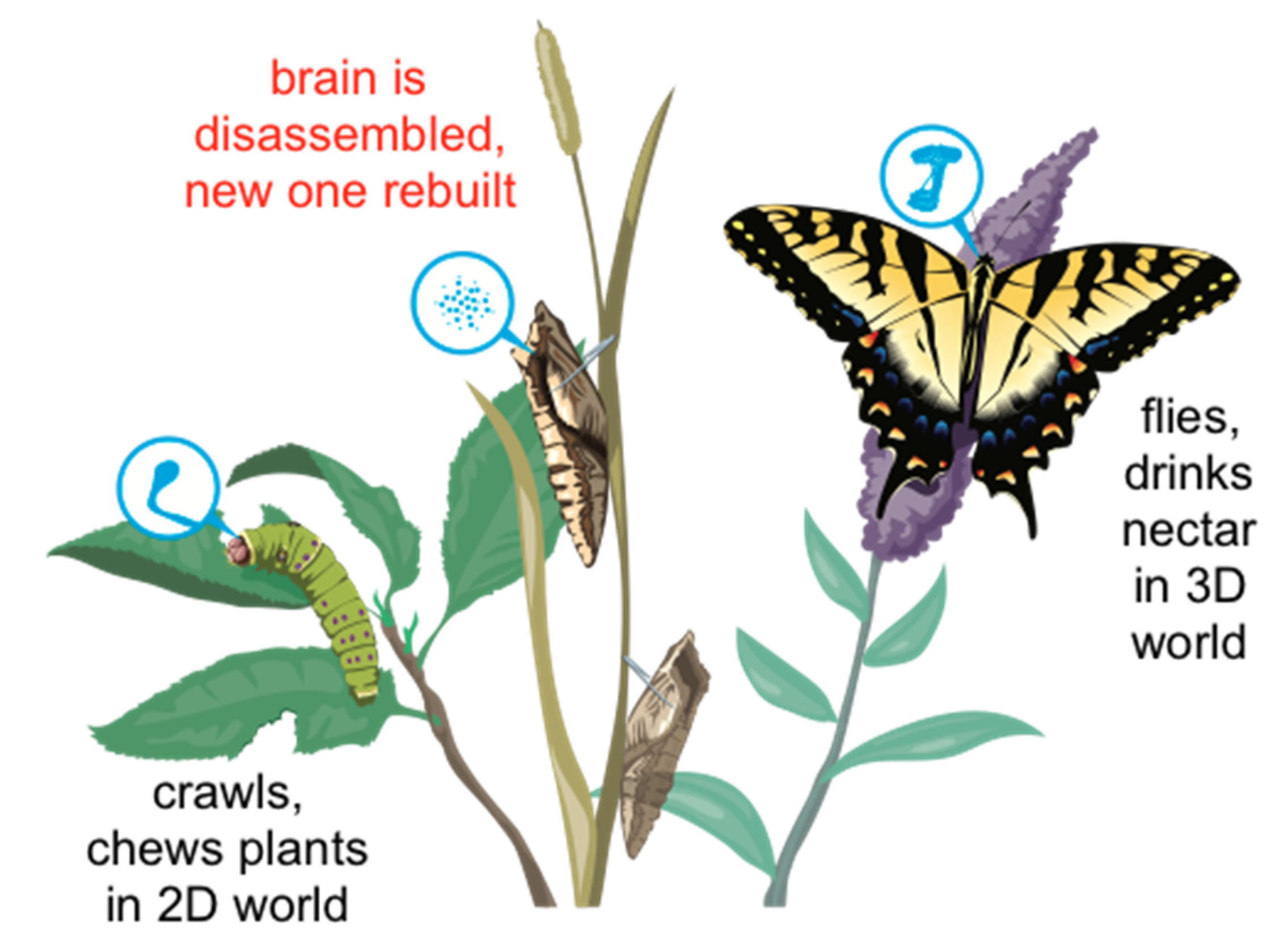
Figure 6.
Human embodiment. The “human” (modern adult form, represented by the yellow glow) that is most commonly dealt with in questions of philosophy of mind, law, etc. stands at the continuum reaching back to a single cell (vertical axis), on both the developmental and evolutionary timescales. However, it is also the center of another continuum, of technological and biological change (horizontal axis). All of these make clear that identifying what a “human” is, or should be, is not trivial, which has major implications of what exactly we are trying to prolong in longevity interventions. What is absolutely crucial is that all of these variants form a smooth continuum of intermediate forms, making it likely impossible to define meaningful crisp categories that can be used to delimit ethical and social categories for “humans vs. machines”. Image by Jeremy Guay of Peregrine Creative.
Figure 6.
Human embodiment. The “human” (modern adult form, represented by the yellow glow) that is most commonly dealt with in questions of philosophy of mind, law, etc. stands at the continuum reaching back to a single cell (vertical axis), on both the developmental and evolutionary timescales. However, it is also the center of another continuum, of technological and biological change (horizontal axis). All of these make clear that identifying what a “human” is, or should be, is not trivial, which has major implications of what exactly we are trying to prolong in longevity interventions. What is absolutely crucial is that all of these variants form a smooth continuum of intermediate forms, making it likely impossible to define meaningful crisp categories that can be used to delimit ethical and social categories for “humans vs. machines”. Image by Jeremy Guay of Peregrine Creative.
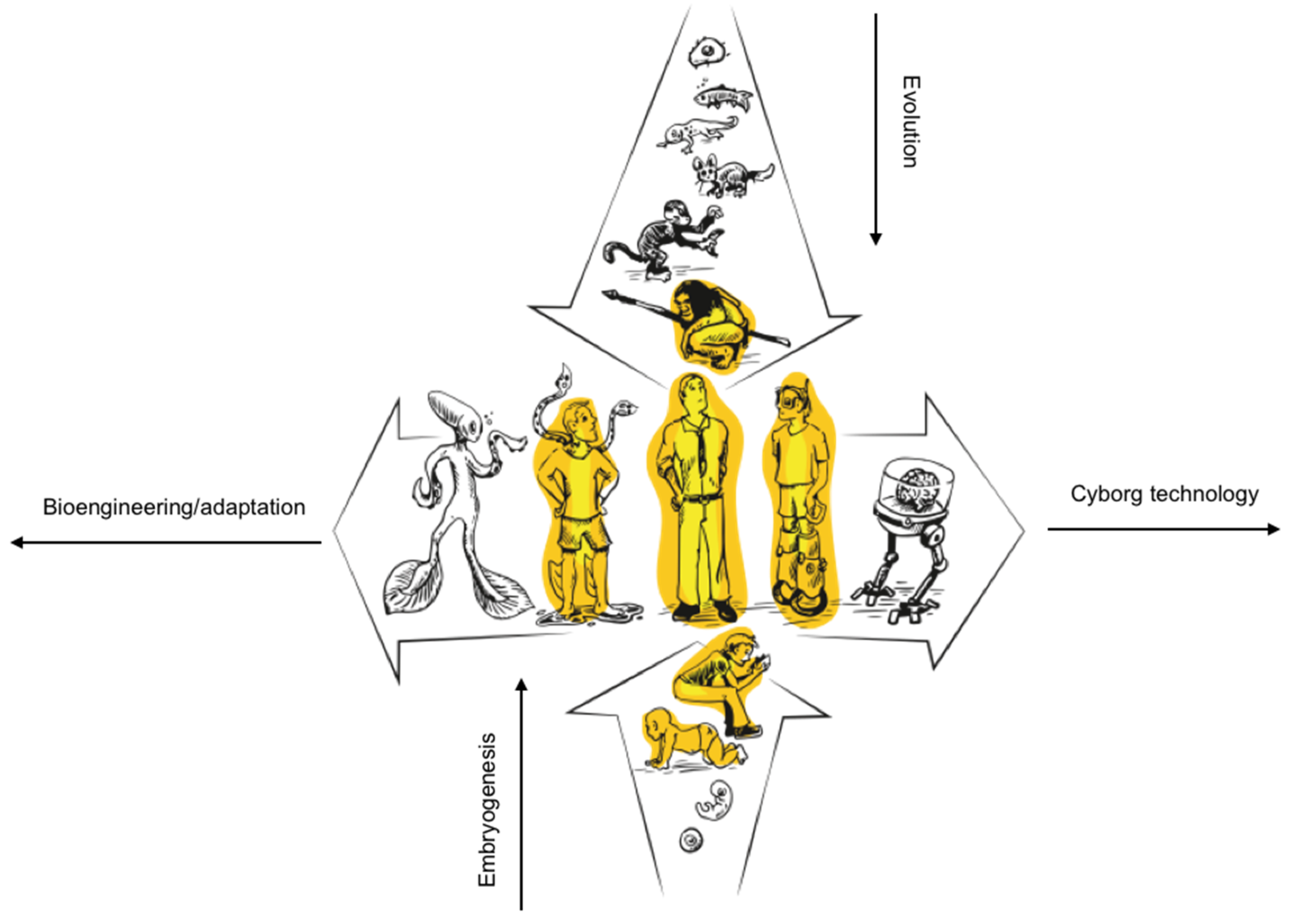
Figure 7.
Multi-scale chimerism. Bioengineering technologies now permit systematic replacement of components at multiple scales, with substituted elements varying along independent axes of cognitive sophistication and developmental origin (evolved versus engineered). The application of evolutionary algorithms in machine design, combined with advances in synthetic morphogenesis, has rendered obsolete traditional categorical distinctions between naturally evolved biological organisms and artificially constructed systems exhibiting goal-directed behavior. Image by Jeremy Guay of Peregrine Creative.
Figure 7.
Multi-scale chimerism. Bioengineering technologies now permit systematic replacement of components at multiple scales, with substituted elements varying along independent axes of cognitive sophistication and developmental origin (evolved versus engineered). The application of evolutionary algorithms in machine design, combined with advances in synthetic morphogenesis, has rendered obsolete traditional categorical distinctions between naturally evolved biological organisms and artificially constructed systems exhibiting goal-directed behavior. Image by Jeremy Guay of Peregrine Creative.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



