Submitted:
25 February 2026
Posted:
26 February 2026
You are already at the latest version
Abstract
The evolution of resistance in mosquitoes to conventional pesticides such as pyrethroids presents a challenge to vector control. Thus, alternative active ingredients for pesticides to manage pyrethroid resistant populations of mosquitoes are needed. The goal of this study was to evaluate the toxic and repellent efficacies of geraniol, a plant secondary metabolite, as a potential alternative for controlling pyrethroid-resistant Aedes aegypti. We found that addition of geraniol to rearing water of 1st instar larvae caused concentration-dependent mortality within 24 h in both strains. The resistance ratio of geraniol (2.8) was modest compared to that of cypermethrin (435.3). Topical application of geraniol to adult female mosquitoes caused dose-dependent mortality in both strains within 24 h. The resistance ratio of geraniol (1.1) was minimal compared to that for cypermethrin (457). In spatial repellency assays, geraniol repelled adult females from both strains in a dose-dependent manner. The repellency resistance ratio of geraniol (2.6) was modest compared to that for pyrethrum extract (>132). Our findings suggest that geraniol has potential use as a versatile chemical tool for controlling pyrethroid-resistant populations of Ae. aegypti.
Keywords:
mosquito
; geraniol
; larvicide
; adulticide
; repellent
1. Introduction
Mosquito-borne arboviruses such as dengue, Zika, and chikungunya represent significant global health threats, collectively infecting hundreds of millions of people and causing tens of thousands of deaths annually [1,2,3]. Aedes aegypti, the yellow fever mosquito, is one of the most important vectors for transmission of these viruses to humans. Control strategies for Ae. aegypti have relied on chemical approaches that use insecticides with limited modes of action (e.g., pyrethroids), which has led to insecticide resistance in mosquito populations and challenges to the control of mosquito-borne arboviral diseases [4,5,6]. Pyrethroid resistance in mosquitoes can arise through multiple mechanisms, including target-site insensitivity (e.g., knockdown resistance, kdr, mutations in voltage-gated sodium channels) and metabolism (e.g., overexpression of detoxification enzymes, such as cytochrome P450 monooxygenases) [7,8].
In addition, overuse of some insecticides (e.g., pyrethroids), has raised concerns about threats to non-target species and environmental health. Pyrethroids act on neural signaling proteins, the voltage-gated sodium channels, which are conserved across other arthropods and vertebrates, including humans [4,9,10,11]. As a result, there is a need to develop new environmentally-friendly chemical tools for vector control.
Plant essential oils and their constituents represent a promising source of bioactive compounds for developing new pesticides. Essential oils are rich in secondary metabolites, which often play crucial roles in the chemical defenses of plants [12,13,14,15,16]. Some secondary metabolites exhibit insecticidal activity, making them potential active ingredients for developing biopesticides. Furthermore, some secondary metabolites are considered safer for human health, less harmful to non-target organisms, and more environmentally sustainable than conventional synthetic insecticides [12,14].
Geraniol is a common, abundant secondary metabolite found in essential oils of several plants, including geranium, citronella, and rose [17,18,19,20].This compound is a monoterpene alcohol with a rose-like scent that is listed by the U.S. Environmental Protection Agency (EPA) as a minimum risk pesticide [21,22,23,24]. Essential oils rich in geraniol have larvicidal and ovicidal effects against several species of mosquitoes, including Culex pipiens, Culex quinquefasciatus, Aedes albopictus, and Ae. aegypti [25,26,27,28]. Consistent with these studies, geraniol alone has been demonstrated to show larvicidal activity against multiple mosquito species, including Cx. pipiens, Cx. quinquefasciatus, Anopheles stephensi, Ae. albopictus, and Ae. aegypti [21,25,26,29,30]. Moreover, geraniol and/or essential oils enriched with geraniol have been shown to elicit adulticidal activity [28,31]. However, a limitation of these prior studies is that they were conducted primarily on insecticide-susceptible mosquito strains. Thus, the toxic efficacy of geraniol against insecticide-resistant mosquitoes is underexplored.
Prior studies have also investigated geraniol’s potential use as a mosquito repellent, which has yielded mixed results. For example, some studies have found that geraniol is a relatively strong repellent, outperforming other natural repellents such as citronella and linalool against susceptible strains of Ae. aegypti [14,24]. However, another study found geraniol to be an ineffective repellent against Ae. aegypti [32]. Intriguingly, in An. gambiae, geraniol showed similar repellent efficacy against both a pyrethroid susceptible and pyrethroid resistant strain, but weaker repellency against an organophosphate-resistant strain [4]. Thus, our understanding of the potential use of geraniol as a repellent is limited and requires further investigation, especially against insecticide resistant mosquito strains.
The goal of this study is to address the above research gaps by comparing the larvicidal, adulticidal, and repellent effects of geraniol between a pyrethroid-susceptible and a pyrethroid-resistant Aedes aegypti strain. Given that a previous computational study [33] predicted that geraniol interacts with voltage-gated sodium channels as well as certain detoxification enzymes involved in pyrethroid resistance (e.g., glutathione S-transferase, esterase) in Ae. aegypti, we hypothesize that pyrethroid-resistant Ae. aegypti will exhibit toxic resistance to geraniol. On the other hand, given that geraniol was equally effective as a repellent against pyrethroid-susceptible and pyrethroid-resistant strains of Anopheles gambiae, we hypothesize that pyrethroid-resistant Ae. aegypti will show limited repellency resistance to geraniol.
2. Materials and Methods
2.1. Aedes aegypti Colonies and Strains
Two different strains of Aedes aegypti mosquitoes were used in the present study: Liverpool (LVP-IB12, MRA-735, obtained by David W. Becnel) and Puerto Rico (PR-NR-48830, obtained by G.G. Clark & J.J. Becnel). The Puerto Rico strain is highly resistant to pyrethroids via both target site (kdr) and metabolic resistance [34,35,36]. From here, we refer to the Liverpool strain as pyrethroid-susceptible (PS) and the Puerto Rico strain as pyrethroid-resistant (PR). To rear mosquitoes, larvae from both strains were fed with 1 tablet per day of commercial fish food (Tropical Tablets, Tetramin, Blacksburg, VA, USA). Adult mosquitoes were maintained on 10% sucrose; for egg production, adult females were fed defibrinated rabbit blood (Hemostat Laboratories, Dixon, CA) via a membrane feeder (Hemotek Ltd., Blackburn, UK). All mosquito colonies were maintained in environmentally-controlled rearing chambers set at the following conditions: 28 °C, 80% relative humidity, and 12:12 light: dark cycle.
2.2. Chemicals
Geraniol (≥ 99%, (2E)-3,7-dimethylocta-2,6-dien-1-ol) was purchased from Thermo Fisher Scientific (Waltham, MA, USA). Cypermethrin (>96%) was obtained from Thermo Fisher Scientific (Acros Organics, Geel, Belgium). Transfluthrin (≥99%) was obtained from Thermo Fisher Scientific (Chicago, IL, USA). Pyrethrum extract (≥50% sum of pyrethrins) was purchased from Sigma-Aldrich (St. Louis, MO, USA).
2.3. Larvicidal Bioassay
Larvicidal activities of geraniol and cypermethrin were tested against 1st-instar larvae of PS and PR strains using an established bioassay [13,37]. Cypermethrin was used as a positive control/reference compound for confirming toxic resistance of larvae of the PR strain [37,38]. In brief, 5 larvae were added to wells of 24-well Falcon Multiwell plates (Becton Dickinson Labware, Franklin Lakes, NJ, USA) containing 985 µL of distilled water and 5 µl of a food suspension. The food suspension consisted of 13 mg of finely ground fish food flakes (Tetramin, Blacksburg, VA, USA) suspended in 1 mL of deionized water. After adding larvae, solvent control wells received 10 µL of 100% acetone, whereas treatment wells received 10 µL of geraniol or cypermethrin dissolved in 100% acetone at various concentrations (freshly prepared on the day of testing). The final concentrations of geraniol in the wells were 1.2, 3.7, 11, 33, or 100 ppm; the final concentrations of cypermethrin in the wells were 0.0004, 0.004, 0.04, 0.4, 4.0, or 40 ppm. The plates were held in the rearing chambers and larval mortality was assessed 24 h post-treatment. Larvae were classified as dead if they failed to respond after gently probing with a pipette tip. Each control and chemical concentration was tested in quadruplicate wells on 8 independent plates (32 wells total) for each strain.
2.4. Topical Adulticidal Bioassay
Topical adulticidal activities of geraniol and cypermethrin were tested against adult females of PS and PR strains using an established bioassay [39,40]. Cypermethrin was used as a positive control/reference compound for confirming toxic resistance of adult females of the PR strain [38]. Ten mosquitoes (3-10 days post-eclosion) were briefly immobilized on ice and 500 nL of 100% acetone (solvent control) or a treatment (geraniol or cypermethrin dissolved at various concentrations in 100% acetone) were applied to the thorax of each mosquito using a Hamilton repeating dispenser (Hamilton Company, Reno, Nevada). The doses of geraniol were 0.20, 0.55, 1.85, 5.50, and 16.5 µg/mosquito; the doses of cypermethrin were 0.00019, 0.0019, 0.019, 0.19, 1.9, and 19.2 ng/mosquito.
After treating the mosquitoes, they were transferred to small cages (32 oz. containers) and provided with access to 10% sucrose. The cages were held in the rearing chamber and toxicity was evaluated based on the percentage of mosquitoes that were dead 24 h post-treatment. Each dose of geraniol and cypermethrin was tested on 5 independent cages of 10 mosquitoes each.
2.5. Mosquito Airborne Repellency Test (MART Assay)
Spatial or airborne repellency of geraniol and pyrethrum extract were tested using a previously described method [41], with the minor modification that 12 mosquitoes were used per replicate instead of 16. Transfluthrin was initially used as a positive control/reference compound for confirming repellency resistance of adult females of the PR strain [42]. However, in preliminary experiments, we did not detect repellency of this compound against either PS or PR strain, which may have been attributed to its vapor toxicity, resulting in immobile or knocked down mosquitoes. Thus, we switched to pyrethrum extract as a positive control/reference compound for repellency [42]. Groups of twelve adult females were transferred into glass cylinders (length 12.5 cm, outer diameter 2.5 cm; TriKinetics Inc, Walthman, MA, USA) with mesh netting secured on both ends with rubber bands (the netting was temporarily removed from one end to introduce the mosquitoes). Before adding the mosquitoes, a line was drawn on each tube at the half-way point of its length. Mosquitoes were acclimated in the tubes for 30 min in a rearing chamber (28 °C, 80% relative humidity) before starting experiments.
While mosquitoes were acclimating to the tubes, fifty microliters of 100% acetone (solvent control) or a treatment (geraniol or pyrethrum extract dissolved in 100% acetone) were applied to round filter papers (diameter 2.5 cm; Cytiva, Fisher Scientific, Waltham, MA, USA). Geraniol was applied at 0.5, 1.0, 3.8, 12.0, and 38.0 µg/cm2 for experiments on PS mosquitoes, and 3.8, 12.0, 38.0, 112.0, and 336.0 µg/cm2 for experiments on PR mosquitoes. Pyrethrum extract was applied at 1.1, 3.3, 12.0, 33.0, and 127.3 µg/cm2 for experiments on PS mosquitoes, and 101.0, 254.6, 336.0, 509.2, 1,018.3, and 2,036.7 µg/cm2 for experiments on PR mosquitoes. The filter papers were placed in plastic caps from 50 mL polypropylene centrifuge tubes (VWR, Radnor, PA USA) under a fume hood for 10 min to allow acetone to evaporate.
To initiate an experiment, one end of a glass tube containing mosquitoes was fitted with a cap containing the treatment filter paper, whereas the other end was fitted with a cap containing the solvent control filter paper. In addition, to confirm that mosquitoes showed no preference towards either end of the tube, double-solvent controls were performed in which both ends were fitted with caps containing filter papers treated with 100% acetone. Fifteen minutes after fitting the caps, the number of mosquitoes on the treatment and control halves of the tubes were counted and the repellency of the treatment was calculated using Formula 1. Any mosquitoes that were knocked down (i.e., not upright, suggesting intoxication or poor health) were excluded. In the double-solvent controls and pyrethrum extract experiments, no knock down of mosquitoes was observed. In geraniol experiments, a few individuals of the PR strain were knocked down at the two highest doses. That is, in the 112.0 µg/cm2 treatment, knocked-down mosquitoes were observed in 2 of 6 replicates (1 or 2 individuals). In the 336.0 µg/cm2 treatment, a few knocked-down mosquitoes were observed in each replicate: 2 individuals in 5 of the 6 replicates and 1 knocked-down individual in 1 of the 6 replicates. The double-solvent controls for each experiment showed minimal repellency for both the PS and PR mosquitoes (Figure S1), indicating no preference of mosquitoes towards either end of the tubes in the absence of geraniol or pyrethrum extract.
Formula 1.
2.6. Statistical Analysis
All data were analyzed using Prism 10 (GraphPad Software). For toxicity assays, the median lethal concentrations (LC50) for larvae or doses (LD50) for adults were determined by plotting percent mortalities against log transformations of the treatment tested and fitting a non-linear regression (log(agonist) vs. normalized response function). For repellency assays, the median effective doses (ED50) were calculated similarly. Statistical comparisons of the LC50, LD50, or ED50 values between the PS and PR strains were compared through extra sum-of-squares F-tests (α = 0.05).
3. Results
3.1. Geraniol and Cypermethrin Toxicity Against PS and PR Larvae
Addition of geraniol to the rearing water of first-instar Ae. aegypti larvae resulted in concentration-dependent mortality within 24 h in both the PS and PR strains (Figure 1a). The LC50 of geraniol for the PS strain (13.6 ppm, 95% CI: 11.0–16.6 ppm) was lower (p < 0.0001) than that of the PR strain (38.6 ppm, 95% Cl: 33.7–44.1 ppm), indicating a larvicidal resistance ratio of 2.8. To provide context for the larvicidal resistance ratio of geraniol, similar experiments were performed using cypermethrin. Previously, we found the resistance ratio of larvae from this same PR strain ranged from 30 to 131 for cypermethrin [37,38]. As shown in Figure 1b, the 24 h LC50 of cypermethrin for the PS strain in the present study was 0.02 ppm (95% CI: 0.01–0.04 ppm), whereas that for the PR strain was dramatically higher (p < 0.0001) at 7.40 ppm (95% CI: 1.01–10.16 ppm), indicating a resistance ratio of 435.3.
3.2. Geraniol and Cypermethrin Toxicity Against PS and PR Adult Females
Topical application of geraniol to adult female A. aegypti resulted in dose-dependent mortality within 24 h in both the PS and PR strains (Figure 2a). The LD50 for the PS strain (6.5 µg/mosquito, 95% CI: 5.1–8.3 µg/mosquito) was similar (p = 0.6) to that for the PR strain (7.1 µg/mosquito, 95% CI: 5.7–8.8 µg/mosquito), indicating an adulticidal resistance ratio of 1.1. To provide context for the adulticidal resistance ratio of geraniol, similar experiments were performed using cypermethrin. Previously, we found the resistance ratio of adult females from this same PR strain of Ae. aegypti to cypermethrin was 84 [38]. As shown in Figure 2b, the 24 h LD50 of cypermethrin for the PS strain in the present study was 7.8 × 10⁻3 ng/mosquito (95% CI: 3.42 × 10⁻3–1.76 × 10⁻2 ng/mosquito), whereas that for the PR strain was dramatically higher (p < 0.0001) at 3.55 ng/mosquito (95% CI: 2.42–5.10 × 4.81 ng/mosquito), indicating a resistance ratio of 457.
3.3. Geraniol and Pyrethrum Extract Spatial Repellency Against PS and PR Adult Females
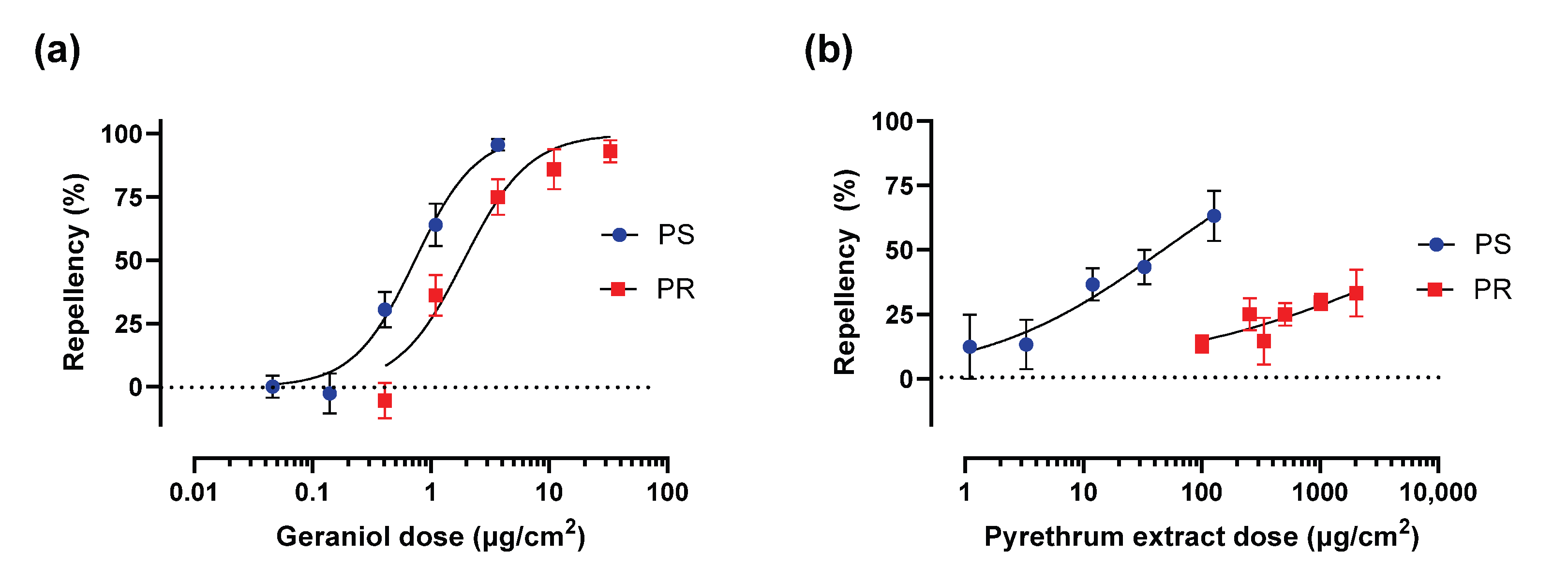
Adult female Ae. aegypti from both PS and PR strains were spatially repelled by geraniol in a dose dependent manner (Figure 3a). The ED50 for the PS strain (0.74 µg/cm2, 95% Cl: 0.58-0.95 µg/cm2) was lower (p < 0.0001) than that for the PR strain (1.90 µg/cm2, 95% CI: 1.38–2.63 µg/cm2), indicating a repellency resistance ratio of 2.6.
To provide context for the repellency resistance ratio of geraniol, similar experiments were performed using pyrethrum extract. Previously, Yang et al. (2019) found the repellency resistance ratio of adult females from this same PR strain of A. aegypti to pyrethrum extract was 7.9 relative to the pyrethroid-susceptible Orlando strain of A. aegypti. As shown in Figure 3b, the ED50 of pyrethrum extract for the PS strain was 47.36 µg/cm² (95% CI: 23.82–140.7 µg/cm²). In contrast, pyrethrum extract showed weak dose-dependent spatial repellency against the PR strain that did not exceed 30%, indicating an ED50 that is beyond detectable limits. Based on the highest dose used in the present study (2,036.7 µg/cm²) we can assume the repellency resistance ratio is at least 132. Attempts to use higher doses of pyrethrum extract to drive higher repellency responses for both the PS and PR strains were unsuccessful due to vapor toxicity of the extract.
4. Discussion
The present study found that geraniol elicited concentration-dependent larvicidal activity against both PS and PR Ae. aegypti. The results for PS larvae were consistent with previous studies that found concentration-dependent larvicidal activity of geraniol against pyrethroid-susceptible strains of Cx. pipiens, Cx. quinquefasciatus, An. stephensi, Ae. albopictus, and Ae. aegypti [22,25,29,30]. Notably, larvae of the PR strain possessed modest cross-resistance (2.8 resistance ratio) to the toxicity of geraniol, but this was much lower than that for cypermethrin, a pyrethroid (435.3 resistance ratio). We also found that geraniol elicited dose-dependent adulticidal activity against both PS and PR Ae. aegypti. The results for PS adults were consistent with previous studies that found dose-dependent adulticidal activity in pyrethroid-susceptible strains of Cx. pipiens, Ae. aegypti, and An. quadrimaculatus [28,31]. In contrast to the larvicidal results, we found that adult females of the PR strain did not exhibit detectable cross-resistance to geraniol (1.1. resistance ratio) while they maintained strong resistance to cypermethrin (457 resistance ratio).
The PR strain of Ae. aegypti used in the present study (Puerto Rico) possesses target site resistance via kdr mutations in voltage-gated sodium channels and metabolic resistance likely via elevated expression of numerous CYP450 mRNAs and at least one GST mRNA [43]. Approximately 50% of the total resistance in the PR strain is attributed to enhanced CYP450 activity [43]. Thus, geraniol appears to bypass most of these resistance mechanisms in larvae and fully bypass them in adult females, suggesting that geraniol has strong potential as a toxin to control pyrethroid resistant populations of Ae. aegypti.
To understand why larvae, but not adult females, of the PR strain exhibited modest cross-resistance to geraniol will require further investigation into geraniol’s mode of toxic action and how mosquitoes detoxify the compound. To date, biochemical targets of geraniol in mosquitoes remain unclear, but geraniol is known to inhibit acetylcholinesterase in beetles [44] and modulate octopaminergic signaling in neurons of cockroaches [45], suggesting neurological disruption as a potential mode of toxic action. A recent computational study suggested that geraniol may interact with voltage-gated sodium channels (VGSCs) in Ae. aegypti by binding to residues that are not implicated in kdr resistance [33]. Thus, if the toxicity of geraniol is associated with interactions with VGSCs, it may bind to the channels via a mechanism distinct from pyrethroids, which is consistent with the relatively modest cross-resistance to geraniol detected in larvae and adult females, respectively, compared to that for cypermethrin. The same computational study [33] also suggested that geraniol may interact with glutathione S-transferases (GSTs) and α-esterases in Ae. aegypti, which have both been implicated in metabolic resistance of mosquitoes to pesticides [46]. Thus, life stage-specific differences in the expression of these detoxification enzymes, as has been found in some mosquito species [42,47], could potentially lead to stage-specific differences in detoxification capacities for geraniol contributing to the different toxic resistance ratios in larvae vs. adult females. Future studies examining the impacts of synergists that inhibit CYP450s (e.g., PBO) or GSTs (e.g., diethyl maleate) on the toxicity of geraniol may help further elucidate the mechanisms of cross-resistance in larvae of the PR strain.
The present study also demonstrated that geraniol elicited dose-dependent spatial repellency against both PS and PR Ae. aegypti. The results in the PS strain were consistent with previous studies that found geraniol repelled pyrethroid susceptible strains of Ae. aegypti in both arm-in-cage and diffuser-based field assays [14,24]. However, our results conflict with a previous study in Ae. aegypti that found geraniol to be ineffective as a repellent when applied to ponies [32]. Notably, adult females of the PR strain possessed modest cross-resistance (2.6 resistance ratio) to the repellency of geraniol, but this was much lower than that for pyrethrum extract (>132 resistance ratio). Our repellency findings with geraniol are consistent with a previous study in the same PR strain of Ae. aegypti that found various degrees of repellency resistance to pyrethrum extract and pyrethroids (transfluthrin and metofluthrin), as well as other compounds, such as DEET, 2-undecanone, and IR3535, leading the investigators to propose that the pyrethroid resistance mechanisms in the PR strain offer broad resistance to a wide diversity of repellents [42]. It should be noted that the results for the PR strain of Ae. aegypti might not apply to pyrethroid-resistant strains of other mosquito species, because a pyrethroid resistant strain of An. gambiae did not exhibit cross-resistance to geraniol, whereas a carbamate-resistant strain of An. gambiae showed strong cross-resistance to geraniol [4]. Thus, the efficacies of various repellents against different insecticide resistant mosquito species should be confirmed on a species by species (and perhaps a strain by strain) basis.
Presumably, the strong repellency resistance to pyrethrum extract in the PR strain is due to kdr mutations that would weaken the interactions between natural pyrethrins and VGSCs; pyrethrins have been shown to drive the majority of the repellent activity associated with pyrethrum extract [48]. As mentioned above, although geraniol was predicted to interact with VGSCs in silico, it’s binding mechanism is hypothesized to be distinct from that of pyrethroids or pyrethrins [33]. Thus, it is more likely that the modest repellency cross-resistance to geraniol is associated with the metabolic resistance mechanisms in the PR strain, which may also explain the broad repellent cross-resistance to DEET, 2-undecanone, and IR3535, which are not known to modulate VGSCs. Future studies examining the impacts of synergists that inhibit CYP450s (e.g., PBO) or GSTs (e.g., diethyl maleate) on the repellency of geraniol and other non-pyrethrin/pyrethroid compounds may help further elucidate the mechanisms of broad cross resistance to repellents in adult females of the PR strain. Nevertheless, given the relatively weak repellency cross-resistance to geraniol in the PR strain compared to that for pyrethrum extract found in the present study, our data suggest that geraniol bypasses most of the pyrethroid resistance mechanisms in Ae. aegypti and thereby has strong potential for use as a repellent against pyrethroid resistant populations of Ae. aegypti.
In conclusion, the results from our study support the notion that geraniol is a valuable natural product for developing novel mosquito control tools to combat pyrethroid resistance. However, it should be noted that the present study evaluated only one representative pyrethroid-susceptible and pyrethroid-resistant strain of Ae. aegypti. Thus, future studies in other species and resistant strains of mosquitoes are needed to confirm the broad applicability of these findings. Nevertheless, the promising results of the present study should motivate future studies to better characterize the biochemical and physiological modes of action of geraniol as a toxicant and repellent.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Figure S1: Double solvent controls (100% acetone on each side) for MART assays. For geraniol experiments, values represent mean ± SEM based on 12 replicates per dose for PS and 6 replicates per dose for PR. For pyrethrum extract experiments, values represent mean ± SEM based on 5 replicates per dose for PS and 8 replicates per dose for PR. None of the means were significantly different from zero as determined by a one-sample t-test (if data were normally distributed) or Wilcoxon signed-rank test (if data were not normally distributed). The results indicate that in the absence of geraniol or pyrethrum extract, mosquitoes from both strains show no preference for a side of the tube.
Author Contributions
Conceptualization, P.N.F. and P.M.P.; methodology, P.N.F. and P.M.P.; validation P.N.F..; formal analysis, P.N.F. and P.M.P.; investigation, P.N.F. and P.M.P.; resources, P.N.F. and P.M.P.; data curation, P.N.F. and P.M.P.; writing—original draft preparation, P.N.F. and P.M.P.; writing—review and editing, P.N.F., P.M.P.; visualization, P.N.F. and P.M.P.; supervision, P.M.P.; project administration, P.M.P; funding acquisition, P.N.F. and P.M.P. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by funds provided by an Internal Grants Graduate Program (2025-045) of The Ohio State University (OSU) College of Food, Agricultural, and Environmental Sciences (CFAES), and by the state and federal appropriated to the OSU CFAES Wooster Campus.
Data Availability Statement
Data are contained within the article or Supplementary Material.
Acknowledgments
The authors would like to acknowledge Erick J. Martinez, Esha Shrestha, and Jafina Tubbs of OSU Entomology for their assistance in rearing the mosquitoes used in this study, and Dr. Reed Johnson of OSU Entomology for his constructive feedback on the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Benelli, G.; Mehlhorn, H. Declining Malaria, Rising of Dengue and Zika Virus: Insights for Mosquito Vector Control. Parasitol Res 2016, 115, 1747–1754. [Google Scholar] [CrossRef] [PubMed]
- Mulatier, M.; Cohuet, A.; Carrasco, D. Chapter 34: Repellents for Mosquito-Borne Disease Control: Beyond the Repellency Effect. In Sensory ecology of disease vectors; Ignell, R., Lazzari, C.R., Lorenzo, M.G., Hill, S.R., Eds.; Brill | Wageningen Academic, 2022; pp. 879–911. ISBN 978-90-8686-380-8. [Google Scholar]
- Poinsignon, A.; Fournet, F.; Ngowo, H.S.; Franco Martins Barreira, V.; Pinto, J.; Bartumeus, F.; Kaindoa, E.W.; Corbel, V. Advances in Surveillance and Control Methods for Aedes-Borne Diseases and Urban Vectors: Report of the International Conference, August 2024, Tanzania. Parasites Vectors 2025, 18, 212. [Google Scholar] [CrossRef] [PubMed]
- Deletre, E.; Martin, T.; Duménil, C.; Chandre, F. Insecticide Resistance Modifies Mosquito Response to DEET and Natural Repellents. Parasites Vectors 2019, 12, 89. [Google Scholar] [CrossRef]
- Norris, E.J.; Gross, A.D.; Bartholomay, L.C.; Coats, J.R. Plant Essential Oils Synergize Various Pyrethroid Insecticides and Antagonize Malathion in Aedes aegypti. Medical Vet Entomology 2019, 33, 453–466. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, X.; Brown, D.J.; An, M.; Xue, R.-D.; Liu, N. Insecticide Resistance: Status and Potential Mechanisms in Aedes Aegypti. Pesticide Biochemistry and Physiology 2023, 195, 105577. [Google Scholar] [CrossRef]
- Hazarika, H.; Rajan, R.K.; Pegu, P.; Das, P. Insecticide Resistance in Mosquitoes: Molecular Mechanisms, Management, and Alternatives. J Pest Sci 2025. [Google Scholar] [CrossRef]
- Mosquitoes: Biology, Pathogenicity and Management; Omkar, Ed.; Springer Nature Singapore: Singapore, 2024; ISBN 978-981-97-4162-5. [Google Scholar]
- Lin, H.-H.; Li, Z.-T.; Tzeng, H.-Y.; Chang, C.; Dai, S.-M. Correlation between Pyrethroid Knockdown Resistance and Mutation Frequency of Voltage-Gated Sodium Channel and Its Application in Aedes Aegypti Management. Pesticide Biochemistry and Physiology 2024, 198, 105710. [Google Scholar] [CrossRef]
- Mohammadi, H.; Ghassemi-Barghi, N.; Malakshah, O.; Ashari, S. Pyrethroid Exposure and Neurotoxicity: A Mechanistic Approach. Archives of Industrial Hygiene and Toxicology 2019, 70, 74–89. [Google Scholar] [CrossRef]
- Sabarwal, A.; Kumar, K.; Singh, R.P. Hazardous Effects of Chemical Pesticides on Human Health–Cancer and Other Associated Disorders. Environmental Toxicology and Pharmacology 2018, 63, 103–114. [Google Scholar] [CrossRef]
- Erb, M. Plant Defenses against Herbivory: Closing the Fitness Gap. Trends in Plant Science 2018, 23, 187–194. [Google Scholar] [CrossRef]
- Inocente, E.A.; Nguyen, B.; Manwill, P.K.; Benatrehina, A.; Kweka, E.; Wu, S.; Cheng, X.; Rakotondraibe, L.H.; Piermarini, P.M. Insecticidal and Antifeedant Activities of Malagasy Medicinal Plant (Cinnamosma Sp.) Extracts and Drimane-Type Sesquiterpenes against Aedes Aegypti Mosquitoes. Insects 2019, 10, 373. [Google Scholar] [CrossRef]
- Lopez, A.D.; Whyms, S.; Luker, H.A.; Galvan, C.J.; Holguin, F.O.; Hansen, I.A. Repellency of Essential Oils and Plant-Derived Compounds Against Aedes Aegypti Mosquitoes. Insects 2025, 16, 51. [Google Scholar] [CrossRef]
- Mahanta, S.; Khanikor, B. Mosquitocidal Activity of Twenty-Eight Plant Essential Oils and Their Binary Mixtures against Culex Quinquefasciatus, (Diptera: Culicidae). Heliyon 2021, 7, e06128. [Google Scholar] [CrossRef]
- Martin, J.S.; Martin, M.M.; Bernays, E.A. Failure of Tannic Acid to Inhibit Digestion or Reduce Digestibility of Plant Protein in Gut Fluids of Insect Herbivores: Implications for Theories of Plant Defense. J Chem Ecol 1987, 13, 605–621. [Google Scholar] [CrossRef]
- Dobreva, A.; Nedeltcheva-Antonova, D.; Nenov, N.; Getchovska, K.; Antonov, L. Subcritical Extracts from Major Species of Oil-Bearing Roses—A Comparative Chemical Profiling. Molecules 2021, 26, 4991. [Google Scholar] [CrossRef]
- Antonova-Nedeltcheva, D.; Dobreva, A.; Gechovska, K.; Antonov, L. Volatile Compounds Profiling of Fresh R. Alba L. Blossom by Headspace—Solid Phase Microextraction and Gas Chromatography. Molecules 2025, 30, 3102. [Google Scholar] [CrossRef] [PubMed]
- Akçura, S.; Çakmakçi, R.; Ürüşan, Z. Changes in the Essential Oil Content and Composition of Pelargonium Graveolens l’hér with Different Drying Methods. Grasas aceites 2023, 74, e497. [Google Scholar] [CrossRef]
- Itawani; Hayati, R.; Munawar, A. Physicochemical Analysis Using Gas Chromatogrphy Mass Spectrophotometer (GCMS) on the Quality of Citronella Oil (Cymbopogon Nardus L.). IOP Conf. Ser.: Earth Environ. Sci. 2024, 1297, 012011. [Google Scholar] [CrossRef]
- Chuaycharoensuk, T.; Manguin, S.; Duvallet, G.; Chareonviriyaphap, T. Assessment of Geraniol-Incorporated Polymers to Control Aedes Albopictus (Diptera: Culicidae). Parasite 2012, 19, 427–432. [Google Scholar] [CrossRef]
- Huong, L.T.; Dai, D.N.; Thin, D.B.; Hung, N.H.; Thinh, B.B. Essential Oils of Distichochlamys Benenica: Chemical Constituents, Mosquito Larvicidal and Antimicrobial Activities. Natural Product Communications 2023, 18, 1934578X231193541. [Google Scholar] [CrossRef]
- Müller, G.C.; Junnila, A.; Kravchenko, V.D.; Revay, E.E.; Butler, J.; Schlein, Y. Indoor Protection Against Mosquito and Sand Fly Bites: A Comparison Between Citronella, Linalool, and Geraniol Candles. Journal of the American Mosquito Control Association 2008, 24, 150–153. [Google Scholar] [CrossRef] [PubMed]
- Müller, G.C.; Junnila, A.; Butler, J.; Kravchenko, V.D.; Revay, E.E.; Weiss, R.W.; Schlein, Y. Efficacy of the Botanical Repellents Geraniol, Linalool, and Citronella against Mosquitoes. Journal of Vector Ecology 2009, 34, 2–8. [Google Scholar] [CrossRef]
- Liu, X.C.; Dong, H.W.; Zhou, L.; Du, S.S.; Liu, Z.L. Essential Oil Composition and Larvicidal Activity of Toddalia Asiatica Roots against the Mosquito Aedes Albopictus (Diptera: Culicidae). Parasitol Res 2013, 112, 1197–1203. [Google Scholar] [CrossRef]
- Benelli, G.; Pavela, R.; Giordani, C.; Casettari, L.; Curzi, G.; Cappellacci, L.; Petrelli, R.; Maggi, F. Acute and Sub-Lethal Toxicity of Eight Essential Oils of Commercial Interest against the Filariasis Mosquito Culex Quinquefasciatus and the Housefly Musca Domestica. Industrial Crops and Products 2018, 112, 668–680. [Google Scholar] [CrossRef]
- Moungthipmalai, T.; Puwanard, C.; Aungtikun, J.; Sittichok, S.; Soonwera, M. Ovicidal Toxicity of Plant Essential Oils and Their Major Constituents against Two Mosquito Vectors and Their Non-Target Aquatic Predators. Sci Rep 2023, 13, 2119. [Google Scholar] [CrossRef] [PubMed]
- Tabari, M.A.; Youssefi, M.R.; Esfandiari, A.; Benelli, G. Toxicity of β-Citronellol, Geraniol and Linalool from Pelargonium Roseum Essential Oil against the West Nile and Filariasis Vector Culex Pipiens (Diptera: Culicidae). Research in Veterinary Science 2017, 114, 36–40. [Google Scholar] [CrossRef]
- Dehghankar, M.; Maleki-Ravasan, N.; Tahghighi, A.; Karimian, F.; Karami, M. Bioactivities of Rose-Scented Geranium Nanoemulsions against the Larvae of Anopheles Stephensi and Their Gut Bacteria. PLoS ONE 2021, 16, e0246470. [Google Scholar] [CrossRef]
- Elghonemy, M.M.; Ayoob, F.; Abdel-Ghany, H.S.M.; Kassem, A.M.; El-Gendy, A.E.G.; Abdel-Shafy, S.; Abd-ElGawad, A.M.; Imagawa, H.; Elshamy, A.I. Larvicidal Effects of Cymbopogon Commutatus Essential Oil and Its Based Nanoformulations on Culex Pipiens: Chemical Profiling and Physicochemical Characterization. Chemistry & Biodiversity 2025, 22, e202403276. [Google Scholar] [CrossRef]
- Kaufman, P.E.; Mann, R.S.; Butler, J.F. Evaluation of Semiochemical Toxicity to Aedes Aegypti, Ae. Albopictus and Anopheles Quadrimaculatus (Diptera: Culicidae). Pest Management Science 2010, 66, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Reeves, W.K.; Miller, M.M. Aqueous 2% Geraniol as a Mosquito Repellent Failed against Aedes Aegypti on Ponies. Journal of the American Mosquito Control Association 2010, 26, 340–341. [Google Scholar] [CrossRef]
- Setlur, A.S.; K, C.; Pandey, S.; Sarkar, M.; Niranjan, V. Comprehensive Molecular Interaction Studies to Construe the Repellent/Kill Activity of Geraniol During Binding Event Against Aedes Aegypti Proteins. Mol Biotechnol 2023, 65, 726–740. [Google Scholar] [CrossRef]
- Du, Y.; Nomura, Y.; Zhorov, B.; Dong, K. Sodium Channel Mutations and Pyrethroid Resistance in Aedes Aegypti. Insects 2016, 7, 60. [Google Scholar] [CrossRef] [PubMed]
- Naw, H.; Võ, T.C.; Lê, H.G.; Kang, J.-M.; Mya, Y.Y.; Myint, M.K.; Kim, T.-S.; Shin, H.-J.; Na, B.-K. Knockdown Resistance Mutations in the Voltage-Gated Sodium Channel of Aedes Aegypti (Diptera: Culicidae) in Myanmar. Insects 2022, 13, 322. [Google Scholar] [CrossRef]
- Ponce-García, G.; Del Río-Galvan, S.; Barrera, R.; Saavedra-Rodriguez, K.; Villanueva-Segura, K.; Felix, G.; Amador, M.; Flores, A.E. Knockdown Resistance Mutations in Aedes Aegypti (Diptera: Culicidae) From Puerto Rico. J Med Entomol 2016, 53, 1410–1414. [Google Scholar] [CrossRef]
- Martínez Rodríguez, E.J.; Phelan, P.L.; Canas, L.; Acosta, N.; Rakotondraibe, H.L.; Piermarini, P.M. Larvicidal Activity of Hemp Extracts and Cannabidiol against the Yellow Fever Mosquito Aedes Aegypti. Insects 2024, 15, 517. [Google Scholar] [CrossRef] [PubMed]
- Inocente, E.A.; Shaya, M.; Acosta, N.; Rakotondraibe, L.H.; Piermarini, P.M. A Natural Agonist of Mosquito TRPA1 from the Medicinal Plant Cinnamosma Fragrans That Is Toxic, Antifeedant, and Repellent to the Yellow Fever Mosquito Aedes Aegypti. PLoS Negl Trop Dis 2018, 12, e0006265. [Google Scholar] [CrossRef]
- Swale, D.R.; Engers, D.W.; Bollinger, S.R.; Gross, A.; Inocente, E.A.; Days, E.; Kanga, F.; Johnson, R.M.; Yang, L.; Bloomquist, J.R.; et al. An Insecticide Resistance-Breaking Mosquitocide Targeting Inward Rectifier Potassium Channels in Vectors of Zika Virus and Malaria. Sci Rep 2016, 6, 36954. [Google Scholar] [CrossRef]
- Calkins, T.L.; Piermarini, P.M. Pharmacological and Genetic Evidence for Gap Junctions as Potential New Insecticide Targets in the Yellow Fever Mosquito, Aedes Aegypti. PLoS ONE 2015, 10, e0137084. [Google Scholar] [CrossRef]
- Jiang, S.; Yang, L.; Bloomquist, J.R. High-throughput Screening Method for Evaluating Spatial Repellency and Vapour Toxicity to Mosquitoes. Medical Vet Entomology 2019, 33, 388–396. [Google Scholar] [CrossRef]
- Yang, L.; Norris, E.J.; Jiang, S.; Bernier, U.R.; Linthicum, K.J.; Bloomquist, J.R. Reduced Effectiveness of Repellents in a Pyrethroid-resistant Strain of Aedes Aegypti (Diptera: Culicidae) and Its Correlation with Olfactory Sensitivity. Pest Management Science 2020, 76, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Estep, A.S.; Sanscrainte, N.D.; Waits, C.M.; Louton, J.E.; Becnel, J.J. Resistance Status and Resistance Mechanisms in a Strain of Aedes Aegypti (Diptera: Culicidae) From Puerto Rico. Journal of Medical Entomology 2017, 54, 1643–1648. [Google Scholar] [CrossRef] [PubMed]
- López, M.D.; Pascual-Villalobos, M.J. Mode of Inhibition of Acetylcholinesterase by Monoterpenoids and Implications for Pest Control. Industrial Crops and Products 2010, 31, 284–288. [Google Scholar] [CrossRef]
- Jankowska, M.; Rogalska, J.; Wyszkowska, J.; Stankiewicz, M. Molecular Targets for Components of Essential Oils in the Insect Nervous System—A Review. Molecules 2017, 23, 34. [Google Scholar] [CrossRef] [PubMed]
- Liu, N. Insecticide Resistance in Mosquitoes: Impact, Mechanisms, and Research Directions. Annu. Rev. Entomol. 2015, 60, 537–559. [Google Scholar] [CrossRef]
- Bharadwaj, N.; Sharma, R.; Subramanian, M.; Ragini, G.; Nagarajan, S.A.; Rahi, M. Omics Approaches in Understanding Insecticide Resistance in Mosquito Vectors. IJMS 2025, 26, 1854. [Google Scholar] [CrossRef]
- Liu, F.; Wang, Q.; Xu, P.; Andreazza, F.; Valbon, W.R.; Bandason, E.; Chen, M.; Yan, R.; Feng, B.; Smith, L.B.; et al. A Dual-Target Molecular Mechanism of Pyrethrum Repellency against Mosquitoes. Nat Commun 2021, 12, 2553. [Google Scholar] [CrossRef]
Figure 1.
Concentration-mortality (24 h) curves for (a) geraniol and (b) cypermethrin against 1st instar larvae of pyrethroid-susceptible (PS, blue) and pyrethroid-resistant (PR, red) Ae. aegypti. For geraniol, values are means ± SEM based on 8 replicates per concentration for the PS strain and 5 replicates per concentration for the PR strain. For cypermethrin, values are means ± SEM based on 5 replicates per concentration for both the PS and PR strains. No mortality was observed in solvent controls.
Figure 1.
Concentration-mortality (24 h) curves for (a) geraniol and (b) cypermethrin against 1st instar larvae of pyrethroid-susceptible (PS, blue) and pyrethroid-resistant (PR, red) Ae. aegypti. For geraniol, values are means ± SEM based on 8 replicates per concentration for the PS strain and 5 replicates per concentration for the PR strain. For cypermethrin, values are means ± SEM based on 5 replicates per concentration for both the PS and PR strains. No mortality was observed in solvent controls.
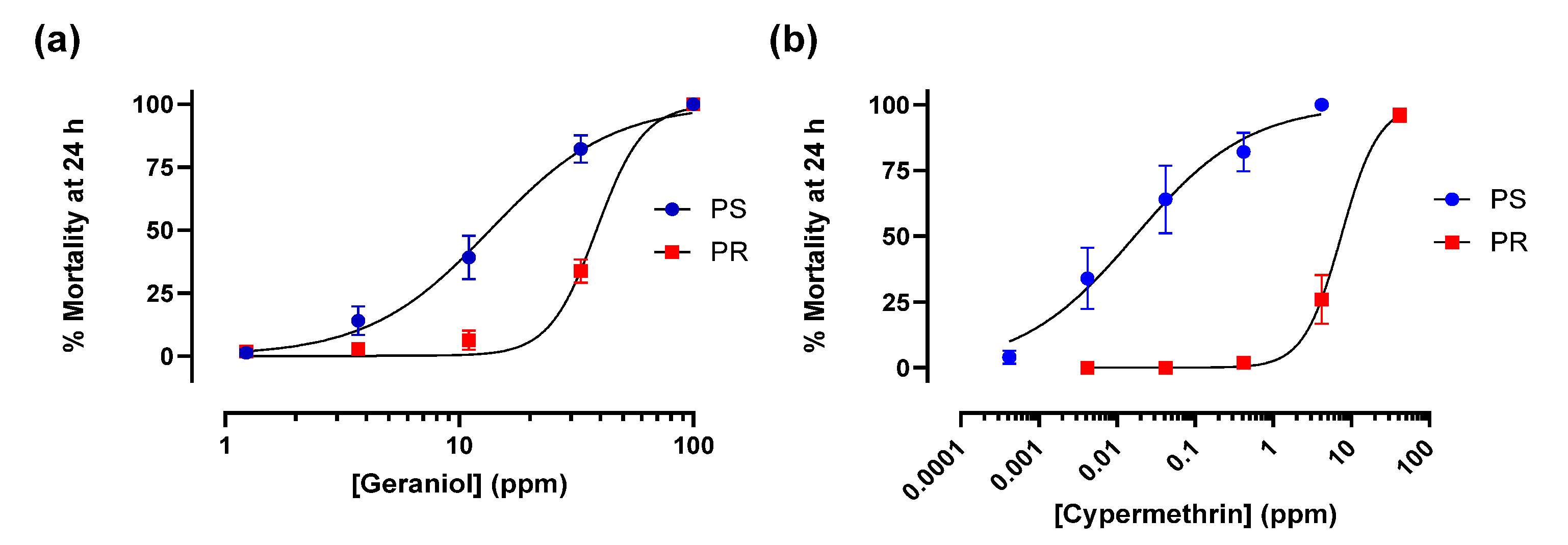
Figure 2.
Dose-mortality curves for 24 h adulticidal activity of (a) geraniol and (b) cypermethrin against adult females of pyrethroid-susceptible (PS, blue) and pyrethroid-resistant (PR, red) Ae. aegypti. Values are means ± SEM based on 5 replicates per dose for both geraniol and cypermethrin for both strains. No mortality was observed in solvent controls.
Figure 2.
Dose-mortality curves for 24 h adulticidal activity of (a) geraniol and (b) cypermethrin against adult females of pyrethroid-susceptible (PS, blue) and pyrethroid-resistant (PR, red) Ae. aegypti. Values are means ± SEM based on 5 replicates per dose for both geraniol and cypermethrin for both strains. No mortality was observed in solvent controls.
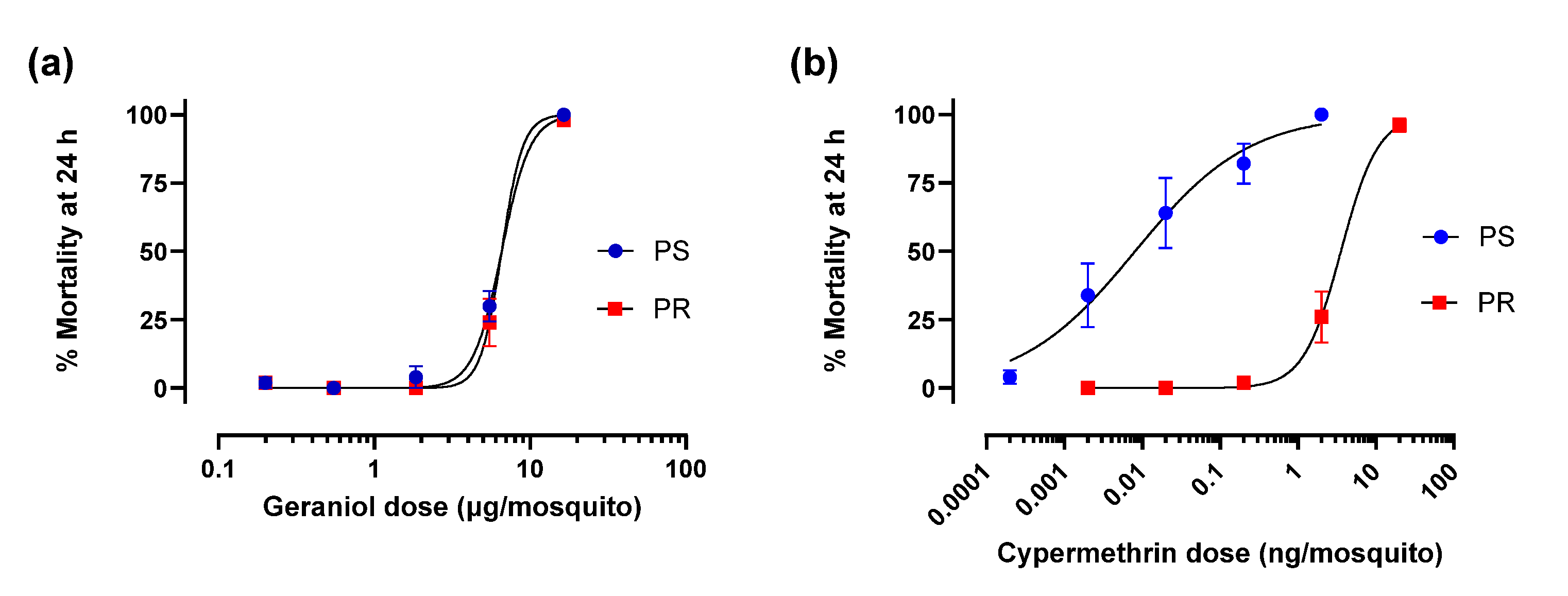
Figure 3.
Dose-repellency (15 min) curves for (a) geraniol and (b) pyrethrum extract against adult females of pyrethroid-susceptible (PS, blue) and pyrethroid-resistant (PR, red) Ae. aegypti. For geraniol, values represent mean ± SEM based on 6-12 replicates per dose for PS and 6 replicates per dose for PR. For pyrethrum extract, values represent mean ± SEM based on 5 replicates per dose for PS and 5-8 replicates per dose for PR. Figure S1 shows the respective double-solvent controls for the geraniol and pyrethrum extract experiments.
Figure 3.
Dose-repellency (15 min) curves for (a) geraniol and (b) pyrethrum extract against adult females of pyrethroid-susceptible (PS, blue) and pyrethroid-resistant (PR, red) Ae. aegypti. For geraniol, values represent mean ± SEM based on 6-12 replicates per dose for PS and 6 replicates per dose for PR. For pyrethrum extract, values represent mean ± SEM based on 5 replicates per dose for PS and 5-8 replicates per dose for PR. Figure S1 shows the respective double-solvent controls for the geraniol and pyrethrum extract experiments.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



