Submitted:
25 February 2026
Posted:
26 February 2026
You are already at the latest version
Abstract
Chronic stress is defined as a prolonged state of emotional disturbance and psychological strain resulting from an inability to maintain internal homeostasis. It is recognized as a significant risk factor for breast cancer, primarily through the chronic activation of the sympathetic nervous system and the hypothalamic-pituitary-adrenal axis. This neuroendocrine activation leads to elevated systemic levels of epinephrine, norepinephrine, and glucocorticoids. By binding to their respective adrenergic and glucocorticoid receptors, these hormones disrupt immune homeostasis and exacerbate oxidative stress within the tumor microenvironment. Such physiological shifts promote critical oncogenic processes, including angiogenesis and tumor cell proliferation, thereby driving the development, progression, and distant metastasis of breast cancer. Mastication, or the act of chewing, serves as a practical and effective behavioral strategy for modulating the deleterious effects of chronic psychological stress. Recent ani-mal studies have provided compelling evidence that chewing can attenuate excessive stress responses. Specifically, it has been shown to mitigate stress-induced breast cancer progression and metastasis by modulating the expression of stress hormones, their corresponding receptors, and key downstream signaling pathways. These findings suggest that the rhythmic activity of chewing may exert a protective effect against stress-related tumor exacerbation. Consequently, further clinical research is warranted to determine whether chewing interventions can serve as a viable complementary strategy alongside conventional breast cancer prevention and treatment protocols.
Keywords:
chronic psychological stress
; oxidative stress
; mastication
; glucocorticoid
; norepinephrine
; breast cancer
; hypothalamic-pituitary-adrenal axis
; sympathetic nervous system
1. Introduction
Cancer remains a leading cause of global morbidity and mortality, accounting for approximately one in six deaths. Among women, breast cancer is the most prevalent malignancy, with an estimated 2.3 million new cases and 670,000 deaths reported in 2022 [1]. Its pathogenesis is multifactorial, driven by a complex interplay of genetic and environmental factors. Established risk factors include a positive family history, BRCA1 and BRCA2 mutations, prolonged estrogen exposure, and lifestyle factors such as alcohol consumption, physical inactivity, and obesity [2]. Beyond these conventional factors, growing evidence underscores the role of chronic psychological stress in breast cancer initiation, progression, and metastasis [3,4,5]. The association between chronic stress and breast cancer was recognized as early as 200 CE, when Galen observed a higher incidence of malignancy in women with melancholia [6]. Since then, numerous studies have explored the biological mechanisms through which psychosocial factors influence oncogenesis [7,8,9]. Persistent stress, arising from life events such as occupational strain, financial hardship, chronic illness, or interpersonal conflict, induces long-term physiological alterations that extend beyond the transient responses seen in acute stress.
Psychophysiological models suggest that chronic stress facilitates breast cancer development and progression primarily through neuroendocrine and immune dysregulation. Persistent stress activates both the hypothalamic-pituitary-adrenal (HPA) axis and the sympathetic nervous system (SNS) [10,11], leading to sustained elevations in glucocorticoids (GCs), epinephrine, and norepinephrine (NE). These stress hormones impair immune surveillance and create a permissive environment for tumor growth and dissemination [12,13]. GCs modulate metabolism, immune signaling, and circadian rhythms, while the activation of β-adrenergic receptors (β-ARs) promotes angiogenesis, invasion, and metastasis. Notably, β-adrenergic blockers have been shown to slow the progression of breast cancer and improve clinical outcomes [14], highlighting the importance of this pathway.
Given its pervasive nature, chronic psychological stress represents a critical yet frequently overlooked factor in breast cancer management. Consequently, effective stress-reduction strategies may be vital for mitigating cancer progression [15]. Of particular interest, recent evidence identifies mastication, or chewing, as a beneficial stress-coping behavior [16,17,18]. Under stressful conditions, chewing appears to alleviate physiological disturbances by attenuating HPA axis and noradrenergic overactivity. However, few systematic reviews have specifically addressed the impact of chewing behavior on cancer progression in the context of chronic stress. This review aims to summarize current knowledge regarding the effects of chewing on chronic psychological stress and explore its potential as a complementary strategy for managing stress-related breast cancer through the modulation of neuroendocrine and immune pathways.
2. General Overview of Breast Cancer
Breast cancer is fundamentally characterized by the uncontrolled proliferation of malignant cells within breast tissue. Although it occurs in men, it remains the most common form of malignancy in women. Invasive ductal carcinoma and invasive lobular carcinoma are the two predominant histological subtypes, originating in the milk ducts and lobules, respectively [2]. The pathogenesis of breast cancer is a complex, multistep process that remains only partially understood. Globally, it represents a substantial health burden, accounting for approximately one-third of all female malignancies and nearly 15% of cancer-related deaths in women worldwide [1,19]. Incidence increases markedly with advancing age, particularly after puberty. Despite significant advancements in diagnostics and therapeutics, breast cancer remains a critical challenge due to its pronounced heterogeneity, variable clinical course, and intricate molecular mechanisms. This disease encompasses multiple molecular subtypes, reflecting its multifactorial etiology and diverse biological pathways [2,20].
The etiology of breast cancer involves a multifactorial interplay of genetic, hormonal, environmental, and behavioral determinants. Non-modifiable factors include increasing age, female sex, family history, and specific germline mutations, most notably high-penetrance mutations in BRCA1 or BRCA2 [2,20]. Hormonal and reproductive factors, such as prolonged estrogen exposure, early menarche, nulliparity, and late menopause, are also well-established risks. Conversely, modifiable risk factors include obesity, sedentary behavior, alcohol consumption, smoking, and exposure to ionizing radiation [19,20,21,22,23,24,25]. A comprehensive understanding of these categories is essential for informing effective prevention and management strategies.
In addition to these conventional factors, emerging evidence suggests that long-term psychological stress contributes significantly to breast cancer development [5,7,8]. Persistent activation of the HPA axis and the SNS under chronic stress leads to sustained elevations of GCs and NE [7,9]. These stress hormones bind to their respective receptors, the glucocorticoid receptor (GR) and β-AR, thereby promoting tumor initiation, growth, and metastasis via neuroendocrine and immunological pathways. Elucidating the processes through which stress-related pathways interact with genetic, hormonal, and environmental factors provides a more integrated understanding of tumor biology. Therefore, a rigorous investigation of these interrelated factors is essential for developing effective prevention strategies and identifying reliable biomarkers and novel therapeutic targets to improve patient outcomes and reduce the global burden of breast cancer.
3. Stress and Stress Responses
Stress is defined as the non-specific response of the body to any demand for change, a syndrome initially characterized by Hans Selye as being triggered by various noxious agents [26]. It encompasses physiological and psychological responses to internal or external challenges, known as stressors, that threaten homeostasis. Stressors may be physical (e.g., pathogens, radiation, and noise) or psychological (e.g., social conflict and emotional burden). In response, the body activates coordinated behavioral and physiological mechanisms to restore homeostatic balance and ensure survival.
The stress response involves multiple interconnected systems, including the HPA axis, SNS, and the neuroendocrine and immune systems, alongside neural circuits such as the locus coeruleus-norepinephrine system. These systems release mediators that act on receptors in both the brain and peripheral tissues to initiate adaptive responses [10,27,28,29]. The HPA axis serves as a central regulator of the stress response. Corticotropin-releasing hormone (CRH), secreted by the hypothalamus [28,29], stimulates the anterior pituitary gland to release adrenocorticotropic hormone (ACTH), which in turn induces the secretion of GCs from the adrenal cortex. While essential for coping with acute stress, chronic activation of the HPA axis can result in dysregulated GC secretion (Figure 1). As primary stress hormones, GCs bind to GR and play critical roles in diverse biological processes, including the regulation of immune responses, metabolism, and inflammation [30,31,32].
Selye described the stress response as the general adaptation syndrome, which progresses through three phases: (1) alarm, characterized by acute physiological changes; (2) resistance, during which the body attempts to adapt to the persistent stressor; and (3) exhaustion, where prolonged stress overwhelms adaptive capacity, leading to systemic dysfunction [26,33]. Recent animal studies have demonstrated that pathologically elevated GC levels and in-creased GR expression are causally associated with enhanced angiogenesis, cancer cell proliferation, accelerated tumor progression, and metastasis [34,35].
The SNS further contributes to the stress response by releasing catecholamines, specifically epinephrine and NE (Figure 1). These neurotransmitters exert context-dependent effects on the immune system [30,36]. While acute SNS activation can enhance innate immunity, prolonged activity impairs the adaptive immune responses. Chronic catecholamine exposure reduces T cell receptor expression and suppresses the function of CD4+ and CD8+ T cells, thereby diminishing immune surveillance [37,38]. Overall, prolonged stress disrupts immune homeostasis. Elevated GC levels suppress key immune functions, alter cytokine profiles, and reduce the population and cytotoxic activity of T cells and natural killer (NK) cells. This dysregulation weakens host defenses, increasing susceptibility to infection and potentially driving the progression of cancer and chronic inflammatory diseases.
Chronic stress induces oxidative stress, characterized by an imbalance between the systemic production of reactive oxygen species (ROS) and the capacity of antioxidant defenses to neutralize these intermediates or repair the resulting damage [39]. This imbalance leads to the excessive accumulation of ROS, resulting in the oxidative modification of cellular components, including lipids, proteins and DNA. Elevated ROS levels perturb redox homeostasis and disrupt normal cellular functions. Evidence suggests that chronic stress increases the production of ROS and reactive nitrogen species (RNS) through elevated GC and NE levels acting via GRs and β2-ARs. The resulting increases in ROS and RNS levels induce DNA damage and promote oncogenic transformation [39,40].
4. Chronic Stress Acts as a Risk Factor for Breast Cancer
Chronic psychological stress is increasingly recognized as a global health concern due to its impact on multiple physiological processes. Long-term stress is implicated in the pathogenesis of numerous conditions, including dementia, cardiovascular disease, gastric ulcers, obesity, diabetes, metabolic syndrome, osteoporosis, arthritis, and neurodegenerative diseases. Therefore, effective stress management is essential for preventing and treating stress-related disorders, including breast cancer.
Animal studies have confirmed that chronic stress leads to persistent activation of the HPA axis and SNS, resulting in elevated levels of GCs and NE. These stress hormones bind to their respective receptors, the GR and β-ARs, promoting tumor progression through diverse signaling pathways (Figure 2). Specifically, chronic activation of β₂-ARs has been shown to enhance angiogenesis [36,37], the epithelial-to-mesenchymal transition (EMT) [41], and metastasis via the AKT-p53 and plexin A1 pathways [10]. Preclinical data indicate that sympathetic activation promotes tumor progression, whereas parasympathetic stimulation exerts inhibitory effects [42]. Clinical studies further suggest that β-blockers may reduce cancer recurrence and metastasis [43], although large-scale trials are required to confirm their efficacy.
GCs act via GRs to regulate immune responses [30,44], inflammation [32], and metabolism [31]. Long-term exposure to cortisol decreases T cell activity and proliferation, thereby reducing the body’s capacity to mount an effective immune response [44]. Sustained SNS activation simultaneously impairs T cell responses, further dysregulating immune function. Continued exposure to catecholamines causes the downregulation of CD4+ and CD8+ T cell receptor expression, resulting in reduced activation and proliferation. Such dysregulation weakens the adaptive immune response, leading to significant clinical implications [38].
Furthermore, chronic psychological stress impairs the immune system’s ability to produce antibodies, rendering individuals more vulnerable to infections. Moreover, it serves as a potent trigger for systemic inflammation [32,45], increasing levels of pro-inflammatory cytokines, such as interleukin-1β (IL-1β), IL-6, and tumor necrosis factor-α (TNF-α) [44]. Studies suggest that long-term stress alters cytokine signaling, contributing to chronic inflammation, which is closely associated with breast cancer progression. While acute GC activity is typically anti-inflammatory, prolonged GR activation paradoxically promotes a pro-inflammatory tumor microenvironment, angiogenesis, and resistance to apoptosis [34,35,46,47]. Furthermore, sustained GC exposure stimulates estrogen biosynthesis via aromatase activation, further promoting tumor growth and correlating with a poor prognosis [48].
In addition to inflammatory changes, chronic stress disrupts immune homeostasis by impairing CD4+, CD8+, and NK cell functions, while promoting the expansion of regulatory T cells (Tregs) via the TGF-β1/Smad2/3/Foxp3 axis [48,49]. These immunosuppressive effects facilitate tumor escape and metastasis. Stress-induced activation of β-adrenergic and GR signaling also enhances the recruitment of myeloid-derived suppressor cells (MDSCs) and tumor-associated macrophages (TAMs). These TAMs secrete CXCL1 via GR and β-adrenergic activation, by binding to the CXCR2 receptor, CXCL1 facilitates the mobilization of splenic MDSCs to distant organs, such as the lungs, establishing premetastatic niches (PMNs). Additionally, TAMs secrete IL-10, matrix metalloproteinase-2 (MMP-2), MMP-9, and vascular endothelial growth factor (VEGF), further driving progression [5,50].
Furthermore, chronic stress exerts hyperglycemic and obesogenic effects, both of which are associated with increased cancer risk. Stress-induced elevation of NE and GCs can also polarize macrophages into an M2-like phenotype, which secretes prostaglandin E2, upregulates VEGF, and induces neovascularization. These M2-like TAMs facilitate cancer cell escape by secreting high levels of IL-10 and MMP-9 while impairing macrophage-mediated clearance of cancer cells [51]. Chronic stress also reshapes the lung microenvironment by inducing fibronectin accumulation, suppressing T-cell infiltration, and enhancing neutrophil recruitment. This stress response disrupts neutrophil circadian rhythms and triggers neutrophil extracellular trap (NET) formation through glucocorticoid signaling. Consequently, glucocorticoids released during chronic stress facilitate NETs, creating a niche conducive to metastasis [52].
Accumulating evidence suggests that oxidative stress is a major contributor to stress-induced oncogenesis. Chronic psychological stress increases the production of ROS and RNS by elevating GC levels and β₂-AR activation [34,35,53]. These molecules induce DNA damage and promote oncogenic transformation. Key mediators include inducible nitric oxide synthase (iNOS) and 4-hydroxynonenal (4-HNE), which facilitate angiogenesis, EMT, and metastasis [38,39,54]. Conversely, the antioxidant enzyme mitochondrial superoxide dismutase 2 (SOD2) protects cells from such damage [55]. Animal experiments have shown that chronic restraint stress significantly elevates iNOS and 4-HNE levels, while decreasing SOD2 expression in breast cancer tissue, suggesting that oxidative stress significantly enhances tumor proliferation. Taken together, chronic psychological stress promotes breast cancer progression and metastasis by dysregulating neuroendocrine, immune, and oxidative pathways. These interactions, primarily mediated by the HPA axis and SNS via GC/GR and NE/β₂-AR signaling, create a permissive environment for angiogenesis, immunosuppression, and metastatic niche formation.
Human studies have revealed that psychological stress significantly increases salivary amylase (a sympathetic nervous system stress marker) and cortisol levels [56]. Furthermore, case-control studies demonstrate that patients with periodontitis exhibit significantly higher stress and elevated salivary cortisol levels, highlighting a potential link between psychological stress, cortisol secretion, and periodontal disease [57,58]. Compared to patients with other malignancies, those with breast cancer often experience higher levels of psychological distress arising from life-threatening diagnoses, physical alterations, treatment side effects, and social dysfunction [59]. These factors frequently lead to anxiety, depression, and insomnia. Accumulating evidence indicates that chronic psychological stress contributes to breast cancer development, progression, and metastasis through neuroendocrine and immune regulatory mechanisms [3,4]. A cohort study revealed that women with a history of chronic stress had a significantly higher prevalence of being overweight and exhibited a greater incidence of aggressive breast cancer subtype, which could potentially be linked to compromised immunosurveillance [60]. Additionally, a clinical study provided substantial evidence indicating that familial breast cancer risk acts as a chronic life stressor, associated with higher levels of self-reported distress [61].
5. Chewing Attenuates Promotion of Breast Cancer by Chronic Stress
Masticatory function plays a critical role in maintaining nutritional status, overall health, and cognitive function. Tooth loss and the resulting decline in masticatory performance not only impair daily activities but also increase the risk of dementia. Persistent masticatory dysfunction activates the SNS and the HPA axis, thereby elevating circulating NE and GC levels. These stress hormones act on their respective receptors, eliciting metabolic, physiological, and behavioral responses [16,17].
Animal studies have demonstrated that occlusal disharmony and bite-raising conditions induce excessive HPA activation and elevate GC levels [62,63]. Similarly, tooth extraction and masticatory dysfunction impair hippocampal-dependent spatial memory, indicating that chewing plays an important role in maintaining hippocampal cognitive function [64]. Early work by Vincent et al. showed that rats allowed to chew during restraint- and water-immersion stress developed fewer gastric lesions than those exposed to stress alone, suggesting that trigeminal afferent input during chewing activates mechanisms that attenuate stress responses [65]. Spatial cognitive function is primarily controlled by the hippocampus, which is highly sensitive to stress and is among the first brain regions to undergo structural and functional changes. Animal studies have further demonstrated that stress-induced elevations in corticosterone levels impair hippocampus-dependent learning and memory, whereas chewing ameliorates these stress-induced cognitive deficits [66].
Miyake et al. reported that rodents provided with wooden sticks to chew during immobilization stress exhibited attenuated suppression of spatial memory and preserved GR expression in the hippocampus [67]. Chronic stress typically downregulates hippocampal GRs, impairing the negative feedback control of the HPA axis. Chewing during immobilization stress ameliorates stress-induced impairments in N-methyl-D-aspartate (NMDA) receptor-mediated long-term potentiation (LTP), possibly through the activation of histamine H1 receptors [68].
Osteoporosis is a skeletal disorder characterized by reduced bone mass and microstructural deterioration, which increases the risk of fracture [69]. Evidence from animal and human studies indicates a link between chronic mild stress and bone loss. Chewing under chronic stress conditions prevents elevations in plasma GC and NE levels, thereby reducing osteoclast-mediated bone resorption, promoting osteoblast-mediated bone formation, and improving trabecular microarchitecture and bone strength [70].
In whole-brain analyses, Ono et al. found that glucose uptake in the hypothalamus increased significantly under stress but normalized when chewing occurred during stress [71]. Concurrent measurements of plasma corticosterone levels confirmed that chewing markedly counteracted the stress-induced elevation in corticosterone levels. A region-of-interest analysis revealed a significant reduction in glucose uptake in the paraventricular nucleus (PVN) of the hypothalamus [71]. Okada et al. reported that restraint stress increases blood pressure, heart rate, and core temperature in rats, all of which are significantly suppressed when chewing occurs during stress [72]. Similarly, Koizumi et al. showed that chewing during immobilization stress ameliorated stress-induced increases in circulating NE levels, as well as reducing the incidence of post-stress arrhythmias associated with sympathetic hyperactivity [73].
How does chewing under stress suppress the SNS and HPA axis activity? The stress-ameliorating effects of chewing likely involve integrated mechanisms between the HPA axis, SNS, and the amygdala-the emotional center responsible for evaluating sensory input and mediating behavioral responses [74]. The PVN, located upstream of the HPA axis, integrates neuroendocrine and autonomic functions to maintain homeostasis [17,75]. Expression of CRH in the PVN is a critical modulator of stress systems. CRH stimulates the release of ACTH from the pituitary gland, which in turn promotes cortisol secretion from the adrenal cortex (Figure 1). This cascade is regulated by negative feedback through steroid receptors in the pituitary gland, hypothalamus, hippocampus, and amygdala. Moreover, CRH neurons in the PVN activate the SNS via the locus coeruleus, thereby enhancing catecholamine release [76].
Chewing has been shown to reduce the number of CRH-, c-Fos-, and phosphorylated ERK1/2 (p-ERK1/2)-positive cells in the PVN, all of which increase under stress [77]. These intracellular changes precede CRH expression, suggesting that chewing modulates early neural responses to stress. Thus, the PVN appears to be a central site mediating the stress-ameliorating effects of chewing, influenced by both neural and humoral factors. Amygdala ablation studies suggest that the amygdala regulates HPA activity. The central amygdala (CeA) projects directly to the PVN [78,79], with GABA acting as the principal inhibitory neurotransmitter. Under stress, GABA efflux in the basolateral amygdala (BLA) increases and is further enhanced by chewing, suggesting that trigeminal inputs strengthen GABAergic transmission. This increased inhibitory signaling suppresses stress-induced p-ERK1/2 expression in the PVN. Consequently, the activation of BLA GABAergic neurons mediates chewing-induced suppression of stress hormone secretion via hypothalamic modulation [78,80]. Additionally, chewing-induced activation of the histamine system, transmitted via the mesencephalic trigeminal nucleus to the tuberomammillary nucleus, enhances NMDA function through H1 receptors, thereby restoring hippocampal LTP.
The hippocampus is particularly vulnerable to stress [78,79]. Oral proprioceptive inputs transmitted via the mesencephalic trigeminal nucleus activate histaminergic neurons in the tuberomammillary nucleus, which project widely throughout the brain [79]. Chewing under stress activates the histamine system, which enhances NMDA function via phospholipase C activation, restoring hippocampal LTP [77,81]. Furthermore, the amygdala modulates hippocampal function both directly and indirectly. Kim et al. demonstrated that activation of GABAergic neurons attenuates stress-induced suppression of hippocampal LTP [81,82]. Thus, the combined activation of the histaminergic and GABAergic systems likely underlies the chewing-induced recovery of hippocampal plasticity.
Stress activates the SNS and increases cardiovascular load. The periaqueductal gray (PAG), a central regulator of cardiovascular control, shows increased p-ERK1/2-positive cells under stress. This activation is normalized by chewing [83]. As the amygdala projects to the PAG, chewing may suppress stress-induced sympathetic activation and cardiovascular responses through enhanced GABAergic inhibition from the BLA [84]. Collectively, these findings indicate that chewing acts as an effective stress-coping mechanism that modulates both neuroendocrine and autonomic stress pathways. In mouse models of breast cancer, chewing during chronic stress significantly attenuated cancer progression and metastasis by reducing angiogenesis and cancer cell proliferation [34,35]. Mechanistically, chewing lowered circulating levels of GCs and NE, while downregulating GR and β₂-AR expression in breast cancer cells. It also reduces oxidative stress by decreasing iNOS and 4-HNE expression and increasing SOD2 expression.
Human studies have established that chewing significantly reduces in salivary cortisol levels [85,86]. In healthy populations, chewing serves as a practical behavior for stress mitigation by effectively decreasing salivary cortisol levels, suggesting that optimizing chewing behavior could further enhance its psychological stress-relief effects [87]. The mechanisms underlying the effects of chewing on psychological stress relief are linked to the neuronal pathways in the brain. Functional magnetic resonance imaging (fMRI) studies have demonstrated that gum chewing increases bilateral blood flow in cerebral areas, including the sensorimotor cortex and insula, suggesting enhanced neuronal activity [88,89,90]. Chewing gum has been shown to alleviate stress responses, improve working memory, and enhance cognitive performance [88]. Notably, chewing gum during stress exposure suppresses the transmission of stress-dependent information in the brain by attenuating sensory processing and neural propagation within these circuits. In particular, chewing gum appears to relieve stress by counteracting stress-induced increases in activity and interconnectivity within sensory, interoceptive, and control networks [17,71,91].
6. Conclusions and Future Perspectives
Modern society faces chronic psychological stress as a major public health crisis, given its pervasive effects on multiple physiological systems. Persistent activation of stress-related pathways, specifically the HPA axis and SNS, precipitates the sustained elevation of GCs and catecholamines. These hormonal shifts suppress immune surveillance, alter cytokine profiles, and augment the production of pro-inflammatory mediators and oxidative stress markers. Consequently, the delicate balance between tumor-suppressive and tumor-promoting mechanisms is disrupted, thereby facilitating the development, progression, and metastasis of breast cancer by enhancing angiogenesis, cell proliferation, and resistance to apoptosis.
Chewing is a simple, cost-effective, and practical behavior observed in both animals and humans. Accumulating evidence indicates that chewing plays a protective role in modulating stress responses. Experimental studies in rodents have demonstrated that chewing during exposure to various stressors can attenuate excessive activation of the HPA axis, normalize stress hormone levels, and uphold the equilibrium between immune and oxidative systems. Furthermore, chewing has been shown to mitigate the impacts of chronic stress on breast cancer progression and metastasis, likely by regulating stress-related signaling pathways and preserving physiological homeostasis.
Given its non-invasive and readily accessible nature, chewing represents a potentially valuable behavioral intervention to counteract the adverse effects of chronic psychological stress. Further research, including well-designed clinical trials, is warranted to validate whether chewing-based strategies can complement conventional approaches for preventing and treating breast cancer in humans.
Author Contributions
Conceptualization, K.A. and K-Y.K.; writing—original draft preparation, K.A., M.I. and K-Y.K.; writing—review and editing, K.A., S.O., K.K., A.S., K.Y., T.T. and K-Y.K. All authors have read and agreed to the published version of the manuscript.
Data Availability Statement
No new data were created or analyzed in this study.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ACTH β-AR BLA CeA CRH EMT GC GR HPA |
Adrenocorticotropic hormone β-adrenergic receptor Basolateral amygdala Central amygdala Corticotropin-releasing hormone Epithelial-mesenchymal transition Glucocorticoid Glucocorticoid receptor Hypothalamic-pituitary-adrenal |
| 4-HNE iNOS IL LTP MDSC MMP NK NE NET NMDA PAG PMN PVN RNS ROS SNS |
4-hydroxynonenal inducible nitric oxide synthase Interleukin Long-term potentiation Myeloid-derived suppressor cell Matrix metalloproteinase Natural killer Norepinephrine Neutrophil extracellular trap N-methyl-D-aspartate Periaqueductal gray Premetastatic niche Paraventricular nucleus Reactive nitrogen species Reactive oxygen species Sympathetic nervous system |
| SOD2 TAM VEGF |
Superoxide dismutase 2 Tumor-associated macrophage Vascular endothelial growth factor |
References
- Freihat, O.; Sipos, D.; Arpad Kovacs, A. Global burden and projections of breast cancer incidence and mortality to 2050: a comprehensive analysis of GLOBOCAN data. Front Public Health 2025, 13, 1622954. [Google Scholar] [CrossRef]
- Łukasiewicz, S.; Czeczelewski, M.; Forma, A.; Baj, J.; Robert Sitarz, R.; Stanisławek, A. Breast Cancer—Epidemiology, Risk Factors, Classification, Prognostic Markers, and Current Treatment Strategies—An Updated Review. Cancers 2021, 13, 4287. [Google Scholar] [CrossRef] [PubMed]
- Liu, H-M.; Ma, L.; Chunyu Li, C.; Cao, B.; Jiang, Y.; Han, L.; Xu, R.; Lin, J.; Zhang, D. The molecular mechanism of chronic stress affecting the occurrence and development of breast cancer and potential drug therapy. Transl. Oncol. 2022, 15, 101281. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, M.; Yu, K.; Xu, S.; Qiu, P.; Lyu, Z.; Zhang, X.; Xu, Y. Chronic stress-induced immune dysregulation in breast cancer: Implications of psychosocial factors. J. Transl. Int. Med. 2022, 11, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Wang, N.; Wang, S.; Zhang, J.; Yang, B.; Wang, Z. Chronic psychological stress promotes breast cancer pre-metastatic niche formation by mobilizing splenic MDSCs via TAM/CXCL1 signaling. J. Exp. Clin. Cancer Res. 2023, 42, 129. [Google Scholar] [CrossRef]
- Mravec, B.; Tibensky, M.; Horvathova, L. Stress and cancer. Part I: Mechanisms mediating the effect of stressors on cancer. J. Neuroimmunol. 2020, 346, 577311. [Google Scholar] [CrossRef]
- Chen, H.; Liu, D.; Guo, L.; Cheng, X.; Guo, N.; Shi, M. Chronic psychological stress promotes lung metastatic colonization of circulating breast cancer cells by decorating a pre-metastatic niche through activating β-adrenergic signaling. J. Pathol. 2018, 244, 49–60. [Google Scholar] [CrossRef]
- Bahri, N.; Najafi, T.F.; Shandiz, F.H.; Tohidinik, H.R.; Khajavi, A. The relation between stressful life events and breast cancer: a systematic review and meta-analysis of cohort studies. Breast Cancer Res. Treat. 2019, 176, 53–61. [Google Scholar] [CrossRef]
- Khan, A.; Song, M.; Dong, Z. Chronic stress: a fourth etiology in tumorigenesis? Mol. Cancer 2025, 24, 196. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, Y.; Li, Q. Mechanisms underlying the effects of stress on tumorigenesis and metastasis (Review). Int. J. Oncol. 2018, 53, 2332–2342. [Google Scholar] [CrossRef]
- Russell, G.; Lightman, S. The human stress response. Nat. Rev. Endocrinol. 2019, 15, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Glaser, R.; Kiecolt-Glaser, J.K. Stress-induced immune dysfunction: implications for health. Nat. Rev. Immunol. 2005, 5, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Le, M.; Bai, Y. Chronic Stress Mediates Inflammatory Cytokines Alterations and Its Role in Tumorigenesis. J. Inflamm. Res. 2025, 18, 1067–1090. [Google Scholar] [CrossRef] [PubMed]
- Caparica, R.; Bruzzone, M.; Agostinetto, E.; Angelis, C.D.; Fêde, Â.; Ceppi, M.; de Azambuja, E. Beta-blockers in early-stage breast cancer: a systematic review and meta-analysis. ESMO Open 2021, 6, 100066. [Google Scholar] [CrossRef]
- Antoni, M.H.; Dhabhar, F.S. The impact of psychosocial stress and stress management on immune responses in patients with cancer. Cancer 2019, 125, 1417–1431. [Google Scholar] [CrossRef]
- Kubo, K-Y.; Iinuma, M.; Chen, H. Mastication as a Stress-Coping Behavior. Biomed. Res. Int. 2015, 2015, 876409. [Google Scholar] [CrossRef]
- Azuma, K.; Zhou, Q.; Niwa, M.; Kubo, K-Y. Association between Mastication, the Hippocampus, and the HPA Axis: A Comprehensive Review. Int. J. Mol. Sci. 2017, 18(8), 1687. [Google Scholar] [CrossRef]
- Yaman-Sözbir, Ş.; Ayaz-Alkaya, S.; Bayrak-Kahraman, B. Effect of chewing gum on stress, anxiety, depression, self-focused attention, and academic success: A randomized controlled study. Stress Health 2019, 35, 441–446. [Google Scholar] [CrossRef]
- Siegel, R.L.; Giaquinto, A.N.; Jemal, A. Cancer statistics, 2024. CA Cancer J. Clin. 2024, 74, 12–49. [Google Scholar] [CrossRef]
- Xiong, X.; Zheng, L-W.; Ding, Y.; Chen, Y-F.; Cai, Y-W.; Wang, L-P.; Huang, L.; Liu, C-C.; Shao, Z-M.; Yu, K-D. Breast cancer: Pathogenesis and treatments. Signal Transduct. Target Ther. 2025, 10, 49. [Google Scholar] [CrossRef]
- Anderson, K.N.; Schwab, R.B.; Martinez, M.E. Reproductive risk factors and breast cancer subtypes: a review of the literature. Breast Cancer Res. Treat. 2014, 144, 1–10. [Google Scholar] [CrossRef]
- Namazi, N.; Irandoost, P.; Heshmati, J.; Larijani, B; Azadbakht, L. The association between fat mass and the risk of breast cancer: A systematic review and metaanalysis. Clin. Nutr. 2019, 38, 1496–1503. [Google Scholar] [CrossRef]
- Bagnardi, V.; Rota, M.; Botteri, E.; Tramacere, I.; Islami, F.; Fedirko, V.; Scotti, L.; Jenab, M.; Turati, F.; Pasquali, E.; Pelucchi, C.; Galeone, C.; Bellocco, R.; Negri, E.; Corrao, G.; Boffetta, P.; La Vecchiaet, C. Alcohol consumption and site-specific cancer risk: a comprehensive dose-response meta-analysis. Br. J. Cancer 2015, 112, 580–593. [Google Scholar] [CrossRef]
- Gaudet, M.M.; Gapstur, S.M.; Sun, J.; Diver, W.R.; Hannan, L.M.; Thun, M.J. Active smoking and breast cancer risk: original cohort data and meta-analysis. J. Natl. Cancer Inst. 2013, 105, 515–525. [Google Scholar] [CrossRef]
- Koo, E.; Henderson, M.A.; Dwyer, M.; Skandarajah, A.R. Management and Prevention of Breast Cancer After Radiation to the Chest for Childhood, Adolescent, and Young Adulthood Malignancy. Ann. Surg. Oncol. 2015, 22, S545–S551. [Google Scholar] [CrossRef]
- Selye, H. A Syndrome produced by Diverse Nocuous Agents. Nature 1936, 138, 32. [Google Scholar] [CrossRef]
- McCarty, R.; Horwatt, K.; Konarska, M. Chronic stress and sympathetic-adrenal medullary responsiveness. Soc. Sci. Med. 1998, 26, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, M.; Stein, D.J.; Gallas-Lopes, M.; Landau, L.; de Almeida, R.M.M. Corticotropin-releasing factor receptor signaling and modulation: implications for stress response and resilience. Trends Psychiatry Psychother. 2020, 42, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Hostinar, C.E.; Sullivan, R.M.; Gunnar, M.R. Psychobiological mechanisms underlying the social buffering of the hypothalamic-pituitary-adrenocortical axis: a review of animal models and human studies across development. Psychol. Bull. 2014, 140, 256–282. [Google Scholar] [CrossRef]
- Besedovsky, H.; del Rey, A. A Glucocorticoid-Mediated Immunoregulatory Circuit Integrated at Brain Levels: Our Early Studies and a Present View. Neuroimmunomodulation 2024, 31, 230–245. [Google Scholar] [CrossRef]
- Akalestou, E.; Genser, L.; Rutter, G.A. Glucocorticoid Metabolism in Obesity and Following Weight Loss. Front Endocrinol. 2020, 11, 59. [Google Scholar] [CrossRef] [PubMed]
- Reichard, S.D.; Agathe Amouret, A.; Chiara Muzzi, C.; Sabine Vettorazzi, S.; Tuckermann, J.P.; Lühder, F.; Reichardt, H.M. The Role of Glucocorticoids in Inflammatory Diseases. Cells 2021, 10, 2921. [Google Scholar] [CrossRef] [PubMed]
- Selye, H. Forty years of stress research: principal remaining problems and misconceptions. Can. Med. Assoc. J. 1976, 115, 53–56. [Google Scholar] [PubMed]
- Zhou, Q.; Katano, M.; Zhang, J-H.; Liu, X.; Wang, K-Y.; Iinuma, M.; Kubo, K-Y.; Azuma, K. Chewing Behavior Attenuates the Tumor Progression-Enhancing Effects of Psychological Stress in a Breast Cancer Model Mouse. Brain Sci. 2021, 11, 479. [Google Scholar] [CrossRef]
- Zhang, J.H.; Wang, K-Y.; Kubo, K-Y.; Azuma, K. Chewing Behavior Attenuates Lung-Metastasis-Promoting Effects of Chronic Stress in Breast-Cancer Lung-Metastasis Model Mice. Cancers 2022, 14, 5950. [Google Scholar] [CrossRef]
- Sanders, V.M.; Kohm, A.P. Sympathetic nervous system interaction with the immune system. Int. Rev. Neurobiol. 2002, 52, 17–41. [Google Scholar]
- Lei, Y.; Liao, F.; Tian, Y.; Wang, Y.; Xia, F.; Wang, J. Investigating the crosstalk between chronic stress and immune cells: implications for enhanced cancer therapy. Front Neurosci. 2023, 17, 1321176. [Google Scholar] [CrossRef]
- Zefferino, R.; Di Gioia, S.; Conese, M. Molecular links between endocrine, nervous and immune system during chronic stress. Brain Behav. 2020, 11, e01960. [Google Scholar] [CrossRef]
- Flaherty, R.L.; Owen, M.; Fagan-Murphy, A.; Intabli, H.; Healy, D.; Patel, B.A.; Allen, M.C.; Patel, B.A.; Flint, M.S. Glucocorticoids induce production of reactive oxygen species/reactive nitrogen species and DNA damage through an iNOS mediated pathway in breast cancer. Breast Cancer Res. 2017, 19, 35. [Google Scholar] [CrossRef]
- Flaherty, R.L.; Intabli, H.; Falcinelli, M.; Bucca, G.; Hesketh, A.; Patel, B.A.; Allen, M.C.; Smith, C.P.; Flint, M.S. Stress hormone-mediated acceleration of breast cancer metastasis is halted by inhibition of nitric oxide synthase. Cancer Lett. 2019, 459, 59–71. [Google Scholar] [CrossRef]
- Silva, D.; Quintas, C.; Gonçalves, J.; Fresco, P. b2-Adrenoceptor Activation Favor Acquisition of Tumorigenic Properties in Non-Tumorigenic MCF-10A Breast Epithelial Cells. Cells 2024, 13, 262. [Google Scholar] [CrossRef]
- Sun, C.; Shen, Y.; Wang, F.; Lu, T.; Zhang, J. Sympathetic nervous system in tumor progression and metabolic regulation: mechanisms and clinical potential. J. Translational. Med. 2025, 23, 836. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, R.; de Lourdes Pereira, M.; Oliveira, M. Beta-Blockers and Cancer: Where Are We? Pharmaceuticals. 2020, 13, 105. [Google Scholar] [CrossRef] [PubMed]
- Alotiby, A. Immunology of Stress: A Review Article. J. Clin. Med. 2024, 13, 6394. [Google Scholar] [CrossRef] [PubMed]
- Slavich, G.M.; Irwin, M.R. From stress to inflammation and major depressive disorder: a social signal transduction theory of depression. Psychol. Bull. 2014, 140, 774–815. [Google Scholar] [CrossRef]
- Kuebler, U.; Zuccarella-Hackl, C.; Arpagaus, A.; Wolf, J.M.; Farahmand, F.; von Känel, R.; Ehlert, U.; Wirtz, P.H. Stress-induced modulation of NF-κB activation, inflammation-associated gene expression, and cytokine levels in blood of healthy men. Brain Behav. Immun. 2015, 46, 87–95. [Google Scholar] [CrossRef]
- Vilasco, M.; Communal, L.; Mourra, N.; Courtin, A.; Forgez, P.; Gompel, A. Glucocorticoid receptor and breast cancer. Breast Cancer Res. Treat. 2011, 130, 1–10. [Google Scholar] [CrossRef]
- Krolick, K.N.; Shi, H. Estrogenic Action in Stress-Induced Neuroendocrine Regulation of Energy Homeostasis. Cells 2022, 11, 879. [Google Scholar] [CrossRef]
- Zhang, X.; Ma, N.; Jin, C.; Cao, X. Role of gut microbiota and immune response in breast cancer progression. Biomolecules and Biomedicine 2026, 26, 100–114. [Google Scholar] [CrossRef]
- Zhang, L.; Pan, J.; Wang, M.; Yang, J.; Zhu, S.; Li, L.; Hu, X.; Wang, Z.; Pang, L.; Li, P.; Jia, Fang; Ren, G.; Zhang, Y.; Xu, D.; Qiu, F.; Huang, J. Chronic Stress-Induced and Tumor Derived SP1+ Exosomes Polarizing IL-1β+ Neutrophils to Increase Lung Metastasis of Breast Cancer. Adv. Sci. 2025, 12, 2310266. [Google Scholar] [CrossRef]
- Momčilović, S.; Milošević, M.; Kočović, D.M.; Marković, D.; Zdravković, D.; Petrinović, S.V. Macrophages at the Crossroads of Chronic Stress and Cancer. Int. J. Mol. Sci. 2025, 26, 6838. [Google Scholar] [CrossRef]
- He, X-Y.; Gao, Y.; Ng, D.; Michalopoulou, E.; George, S.; Adrover, J.M.; Sun, L.; Albrengues, J.; Daßler-Plenker, J.; Han, X.; Wan, L.; Wu, X.S.; Shui, L.S.; Huang, Y-H.; Liu, B.; Su, C.; Spector, D.L.; Vakoc, C.R.; Aelst, L.V.; Egeblad, M. Chronic stress increases metastasis via neutrophil-mediated changes to the microenvironment. Cancer Cell 2024, 42, 474–486.e12. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Deng, Z.; Lei, C.; Ding, X.; Li, J.; Wang, C. The Role of Oxidative Stress in Tumorigenesis and Progression. Cells 2024, 13, 441. [Google Scholar] [CrossRef] [PubMed]
- Li, Y-P.; Tian, F-G.; Shi, P-C.; Guo, L-Y.; Wu, H-M.; Chen, R-Q.; Xue, J-M. 4-Hydroxynonenal promotes growth and angiogenesis of breast cancer cells through HIF-1α stabilization. Asian Pac. J. Cancer Prev. 2014, 15, 10151–10156. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Vallur, P.G.; Phaëton, R.; Mythreye, K.; Hempel, N. Insights into the Dichotomous Regulation of SOD2 in Cancer. Antioxidants 2017, 6, 86. [Google Scholar] [CrossRef]
- Takai, N.; Yamaguchi, M.; Aragaki, T.; Eto, K.; Uchihashi, K.; Nishikawa, Y. Effect of psychological stress on the salivary cortisol and amylase levels in healthy young adults. Arch. Oral. Biol. 2004, 49, 9633–968. [Google Scholar] [CrossRef]
- Hingorjo, M.R.; Owais, M.; Siddiqui, S.U.; Nazar, S.; Ali, Y.S. The impact of psychological stress on salivary cortisol levels in periodontitis patients: a case-control study. BMC Oral Health 2025, 21, 25:276. [Google Scholar] [CrossRef]
- Lee, Y-H.; Suk, C.; Shin, S-I.; Hong, J-Y. Salivary cortisol, dehydroepiandrosterone, and chromogranin A levels in patients with gingivitis and periodontitis and a novel biomarker for psychological stress. Front Endocrinol. 2023, 14, 1147739. [Google Scholar] [CrossRef]
- Dinapoli, L.; Colloca, G.; Di Capua, B.; Valentini, V. Psychological Aspects to Consider in Breast Cancer Diagnosis and Treatment. Curr. Oncol. Rep. 2021, 23, 38. [Google Scholar] [CrossRef]
- Cormanique, T.F.; de Almeida, L.E.; Rech, C.A.; Rech, D.; Herrera, A.C.; Panis, C. Chronic psychological stress and its impact on the development of aggressive breast cancer. Einstein 2015, 13, 352–356. [Google Scholar] [CrossRef]
- Gold, S.M.; Zakowski, S.G.; Valdimarsdottir, H.B.; Bovbjerg, D.H. Stronger endocrine responses after brief psychological stress in women at familial risk of breast cancer. Psychoneuroendocrinology 2003, 28, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Kubo, K-Y.; Yamada, Y.; Iinuma, M.; Iwaku, F.; Tamura, Y.; Watanabe, K.; Nakamura, H.; Onozuka, M. Occlusal disharmony induces spatial memory impairment and hippocampal neuron degeneration via stress in SAMP8 mice. Neurosci. Lett. 2007, 414, 188–191. [Google Scholar] [CrossRef] [PubMed]
- Miyake, H.; Mori, D.; Katayama, T.; Fujiwara, S.; Sato, Y.; Azuma, K.; Kubo, K-Y. Novel stress increases hypothalamic-pituitary-adrenal activity in mice with a raised bite. Arch. Oral Biol. 2016, 68, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Kubo, K-Y.; Kojo, A.; Yamamoto, T.; Onozuka, M. The bite-raised condition in aged SAMP8 mice induces dendritic spine changes in the hippocampal region. Neurosci. Lett. 2008, 441, 141–144. [Google Scholar] [CrossRef]
- Vincent, G.P.; Paré, W.P.; Prenatt, J.E.; Glavin, G.B. Aggression, body temperature, and stress ulcer. Physiol. Behav. 1984, 32, 265–268. [Google Scholar] [CrossRef]
- Kubo, K-Y.; Sasaguri, K.; Ono, Y.; Yamamoto, T.; Takahashi, T.; Watanabe, K.; Karasawa, N.; Onozuka, M. Chewing under restraint stress inhibits the stress-induced suppression of cell birth in the dentate gyrus of aged SAMP8 mice. Neurosci. Lett. 2009, 466, 109–113. [Google Scholar] [CrossRef]
- Miyake, S.; Yoshikawa, G.; Yamada, K.; Sasaguri, K-I.; Yamamoto, T.; Onozuka, M.; Sato, S. Chewing ameliorates stress-induced suppression of spatial memory by increasing glucocorticoid receptor expression in the hippocampus. Brain Res. 2012, 1446, 34–39. [Google Scholar] [CrossRef]
- Ono, Y.; Kataoka, T.; Miyake, S.; Sasaguri, K.; Sato, S.; Onozuka, M. Chewing rescues stress-suppressed hippocampal long-term potentiation via activation of histamine H1 receptor. Neurosci. Res. 2009, 64, 385–390. [Google Scholar] [CrossRef]
- Azuma, K.; Adachi, Y.; Hayashi, H.; Kubo, K-Y. Chronic Psychological Stress as a Risk Factor of Osteoporosis. J. UOEH 2015, 37, 245–253. [Google Scholar] [CrossRef]
- Furuzawa, M.; Chen, H.; Fujiwara, S.; Yamada, K.; Kubo, K-Y. Chewing ameliorates chronic mild stress-induced bone loss in senescence-accelerated mouse (SAMP8), a murine model of senile osteoporosis. Exp. Gerontol. 2014, 55, 12–18. [Google Scholar] [CrossRef]
- Ono, Y.; Lin, S-C.; Tzen, K-Y.; Chen, H-H.; Yang, P-F.; Lai, W-S.; Chen, J-H.; Onozuka, M.; Yen, C-T. Active coping with stress suppresses glucose metabolism in the rat hypothalamus. Stress 2012, 15, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Okada, S.; Hori, N.; Kimoto, K.; Onozuka, M.; Sato, S.; Sasaguri, K. Effects of biting on elevation of blood pressure and other physiological responses to stress in rats: biting may reduce allostatic load. Brain Res. 2007, 1185, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Koizumi, S.; Minamisawa, S.; Sasaguri, K.; Onozuka, M.; Sato, S.; Ono, Y. Chewing reduces sympathetic nervous response to stress and prevents poststress arrhythmias in rats. Am. J. Physiol. 2011, 301, H1551–H1558. [Google Scholar] [CrossRef] [PubMed]
- Pessoa, L. A Network Model of the Emotional Brain. Trends Cogn. Sci. 2017, 21, 357–371. [Google Scholar] [CrossRef]
- McEwen, B.S. The neurobiology of stress: from serendipity to clinical relevance. Brain Res. 2000, 886, 172–189. [Google Scholar] [CrossRef]
- Binder, E.B; Nemeroff, C.B. The CRF system, stress, depression and anxiety-insights from human genetic studies. Mol. Psychiatry 2010, 15, 574–588. [Google Scholar] [CrossRef]
- Sasaguri, K.; Kikuchi, M.; Hori, N.; Yuyama, N.; Onozuka, M.; Sato, S. Suppression of stress immobilization-induced phosphorylation of ERK 1/2 by biting in the rat hypothalamic paraventricular nucleus. Neurosci. Lett. 2005, 383, 160–164. [Google Scholar] [CrossRef]
- Van de Kar, L.D.; Piechowski, R.A.; Rittenhouse, P.A.; Gray, T.S. Amygdaloid lesions: differential effect on conditioned stress and immobilization-induced increases in corticosterone and renin secretion. Neuroendocrinology 1991, 54, 89–95. [Google Scholar] [CrossRef]
- Chen, H.; Iinuma, M.; Onozuka, M.; Kubo, K-Y. Chewing Maintains Hippocampus-Dependent Cognitive Function. Int. J. Med, Sci. 2015, 12, 502–509. [Google Scholar] [CrossRef]
- Prewitt, C.M.; Herman, J.P. Hypothalamo-Pituitary-Adrenocortical Regulation Following Lesions of the Central Nucleus of the Amygdala. Stress 1997, 1, 263–280. [Google Scholar] [CrossRef]
- Reznikov, L.R.; Reagan, L.P.; Fadel, J.R. Effects of acute and repeated restraint stress on GABA efflux in the rat basolateral and central amygdala. Brain Res. 2009, 1256, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.J.; Koo, J.W.; Lee, H.J.; Han, J.S. Amygdalar inactivation blocks stress-induced impairments in hippocampal long-term potentiation and spatial memory. J. Neurosci. 2005, 25, 1532–1539. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Narimatsu, Y.; Ono, Y.; Sasaguri, K-I.; Onozuka, M.; Kawata, T.; Yamamoto, T. Chewing suppresses the stress-induced increase in the number of pERK-immunoreactive cells in the periaqueductal grey. Neurosci. Lett. 2015, 599, 43–48. [Google Scholar] [CrossRef] [PubMed]
- de Abreu, A.R.; Abreu, A.R.; Santos, L.T.; de Souza, A.A.; da Silva, L.G., Jr.; Chianca, D.A., Jr.; de Menezes, C.R. Amygdalar neuronal activity mediates the cardiovascular responses evoked from the dorsolateral periaqueductal gray in conscious rats. Neuroscience 2015, 284, 737–750. [Google Scholar] [CrossRef]
- Tasaka, A.; Takeuchi, K.; Sasaki, H.; Yoshii, T.; Soeda, R.; Ueda, T.; Sakurai, K. Influence of chewing time on salivary stress markers. J. Prosthodont. Res. 2014, 58, 48–54. [Google Scholar] [CrossRef]
- Tahara, Y.; Sakurai, K.; Ando, T. Influence of chewing and clenching on salivary cortisol levels as an indicator of stress. J. Prosthodont. 2007, 16, 129–135. [Google Scholar] [CrossRef]
- Tasaka, A.; Kikuchi, M.; Nakanishi, K.; Ueda, T.; Yamashita, S.; Sakurai, K. Psychological stress-relieving effects of chewing - Relationship between masticatory function-related factors and stress-relieving effects. J. Prosthodont. 2018, 62, 50–55. [Google Scholar] [CrossRef]
- Onozuka, M.; Fujita, M.; Watanabe, K.; Hirano, Y.; Niwa, M.; Nishiyama, K.; Saito, S. Mapping brain region activity during chewing: a functional magnetic resonance imaging study. J. Dent. Res. 2002, 81, 743–746. [Google Scholar] [CrossRef]
- Hirano, Y.; Onozuka, M. Chewing and attention: a positive effect on sustained attention. Biomed. Res. Int. 2015, 2015, 367026. [Google Scholar] [CrossRef]
- Hirano, Y.; Obata, T.; Kashikura, K.; Nonaka, H.; Tachibana, A.; Ikehira, H.; Onozuka, M. Effects of chewing in working memory processing. Neurosci. Lett. 2008, 436, 189–192. [Google Scholar] [CrossRef]
- Yu, H.; Chen, X.; Liu, J.; Zhou, X. Gum chewing inhibits the sensory processing and the propagation of stress-related information in a brain network. PLoS ONE 2013, 8, e57111. [Google Scholar] [CrossRef]
Figure 1.
Illustration of the stress response. CRH secreted by the hypothalamus, stimulates the pituitary to release ACTH, which in turn induces the secretion GC from the adrenal cortex. GCs exert negative feedback control on the secretion of both CRH and ACTH. In addition, the hypothalamus activates the SNS and the adrenal medulla, leading to the secretion of epinephrine and norepinephrine. ACTH: adrenocorticotropic hormone; CRH: corticotropin-releasing hormone; GC: glucocorticoid; SNS: sympathetic nervous system.
Figure 1.
Illustration of the stress response. CRH secreted by the hypothalamus, stimulates the pituitary to release ACTH, which in turn induces the secretion GC from the adrenal cortex. GCs exert negative feedback control on the secretion of both CRH and ACTH. In addition, the hypothalamus activates the SNS and the adrenal medulla, leading to the secretion of epinephrine and norepinephrine. ACTH: adrenocorticotropic hormone; CRH: corticotropin-releasing hormone; GC: glucocorticoid; SNS: sympathetic nervous system.
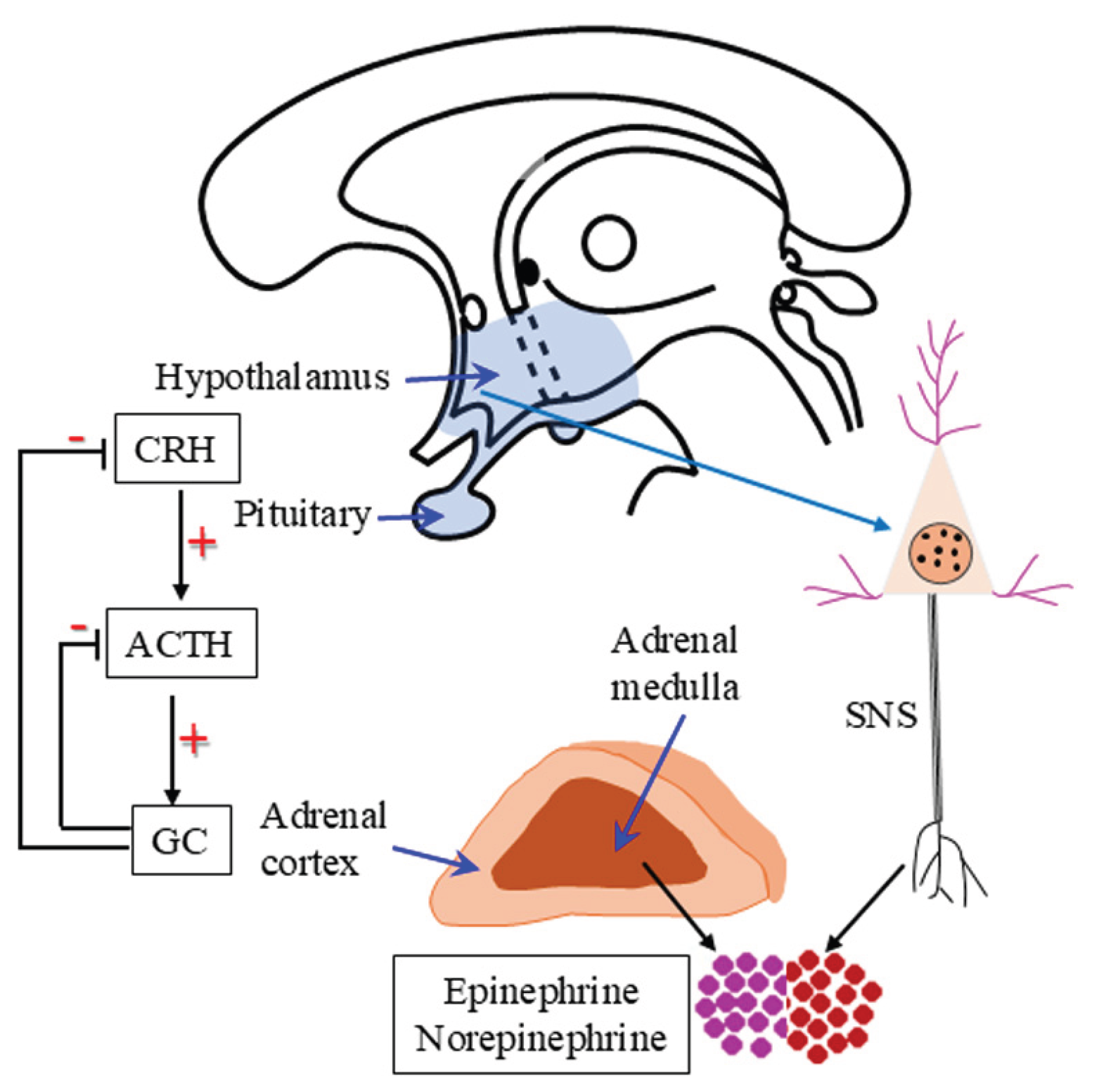
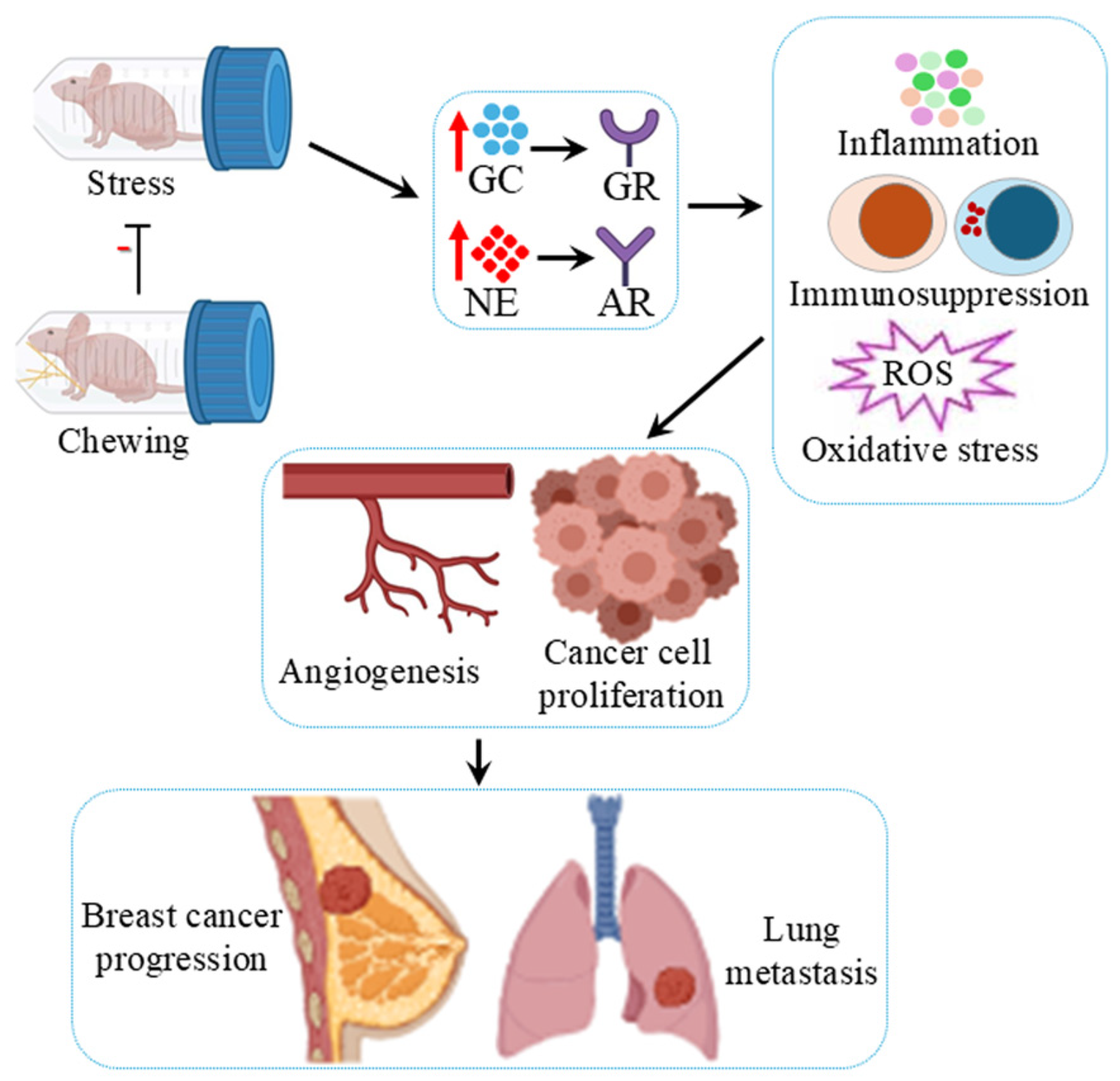
Figure 2.
A simplified diagram illustrates the relationship among chewing, chronic stress, and stress-related breast cancer. Chronic stress leads to elevated levels of GC and NE, through binding to their respective receptors, the GR and AR, disrupt immune homeostasis, trigger inflammation, and increase oxidative stress. These alterations promote angiogenesis, cancer cell proliferation, and ultimately the development, progression and metastasis of breast cancer. In contrast, chewing attenuates stress responses and mitigates the promotive effects of chronic stress on breast cancer progression, and lung metastasis. AR; adrenergic receptor; GC; glucocorticoid; GR; glucocorticoid receptor; NE; norepinephrine.
Figure 2.
A simplified diagram illustrates the relationship among chewing, chronic stress, and stress-related breast cancer. Chronic stress leads to elevated levels of GC and NE, through binding to their respective receptors, the GR and AR, disrupt immune homeostasis, trigger inflammation, and increase oxidative stress. These alterations promote angiogenesis, cancer cell proliferation, and ultimately the development, progression and metastasis of breast cancer. In contrast, chewing attenuates stress responses and mitigates the promotive effects of chronic stress on breast cancer progression, and lung metastasis. AR; adrenergic receptor; GC; glucocorticoid; GR; glucocorticoid receptor; NE; norepinephrine.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



