Submitted:
24 February 2026
Posted:
25 February 2026
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Following the COVID-19 pandemic in 2020, there has been growing interest in investigating the transmission routes of SARS-CoV-2 and identifying the potential reservoirs in wildlife. To date, the virus has been detected in a wide range of wildlife animals. SARS-CoV-2 has also been demonstrated the ability to infect dogs, raising concerns that other carnivores, such as free-living canids, might serve as a potential sources of infection. The aim of this study was to investigate the presence of specific IgG antibodies against SARS-CoV-2 in golden jackals (Canis aureus) and red foxes (Vulpes vulpes) in Serbia. A total of 165 individuals from ten districts including the City of Belgrade were collected and examined from the beginning of August 2024 to the end of March 2025. Of this number, 17 samples were positive (17/165), which represents 10.3% of all specimens tested. Among 83 red foxes, antibodies against SARS-CoV-2 were detected in 11 animals (13.3%), whereas reactive sera were identified in 6 of 82 jackals (7.3%). The analyses were performed using a commercial multi-species ELISA (IDVet, France), with results confirmed by an in-house ELISA, previously developed for the detection of COVID-19 in humans and adapted for canids. Our findings confirm that wild animals are indeed exposed to SARS-CoV-2 infection, and that golden jackals and red foxes may serve as competent hosts for infection, posing a threat for human health. These results represent the first detection of SARS-CoV-2 seroconversion in wild carnivores in Serbia, highlighting the need for further surveillance and investigation.
Keywords:
SARS-CoV-2
; wildlife
; serology
; red foxes
; golden jackals
; one health
; Serbia
1. Introduction
Members of the family Coronaviridae are ubiquitous worldwide and comprise a group of enveloped, positive-sense single-stranded RNA viruses. Coronaviruses (CoVs) are classified into four genera: Alphacoronavirus, Betacoronavirus, Gammacoronavirus, and Deltacoronavirus. Alphacoronaviruses and betacoronaviruses have been confirmed primarily in bats, whereas birds are recognized as the main reservoirs of gamma- and deltacoronaviruses [1]. Coronaviruses are also characterized by a high mutation rate, which contributes to their genetic diversity and adaptive potential. It has long been recognized that coronaviruses can cause respiratory and gastrointestinal diseases in domestic and wild animals. Well-known representatives include canine coronavirus (CCoV), which causes enteric disease in dogs and related species, and canine respiratory coronavirus (CRCoV), which is associated with mild to severe respiratory disorders in dogs [2,3].
In December 2019, a novel coronavirus, severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), emerged in Wuhan, China, causing coronavirus disease 2019 (COVID-19) [4,5]. The clinical presentation of COVID-19 ranges from mild symptoms such as fever, dry cough, fatigue, and loss of taste or smell to severe disease and fatal outcomes [6]. The rapid spread of the disease overwhelmed healthcare systems, caused substantial economic losses, and led to the implementation of unprecedented preventive measures worldwide, including travel and trade restrictions. Globally, COVID-19 spread at an exceptional pace and resulted in approximately 676 million confirmed human infections and about 6.9 million deaths [7].
Genetically, SARS-CoV-2 is closely related to earlier zoonotic coronaviruses responsible for the severe acute respiratory syndrome (SARS) outbreak in 2002–2003 and the Middle East respiratory syndrome (MERS) outbreak in 2012 [8,9]. However, the epidemiological characteristics of COVID-19 differ considerably from those of previous outbreaks, which were geographically limited. In contrast, COVID-19 exhibited rapid global spread, reaching pandemic proportions with high morbidity and mortality. It is widely assumed that SARS-CoV-2 originated from wildlife and spilled over to humans at a live animal market in the Chinese province of Hubei. Bats are considered the natural reservoir of the virus, while the intermediate host has not been definitively identified, although the Asian palm civet (Paradoxurus hermaphrodites) has been suggested as a potential candidate [10].
SARS-CoV-2 is a single-stranded RNA virus belonging to the genus Betacoronavirus. The viral genome encodes four main structural proteins: membrane (M), envelope (E), spike (S), and nucleocapsid (N). The N and S proteins play a key role in inducing the humoral immune response and antibody production [6]. Betacoronaviruses, including SARS-CoV-2, utilize the angiotensin-converting enzyme 2 (ACE2) receptor for host cell entry, a receptor that is present in many animal species [11]. Nevertheless, susceptibility varies among species; for example, experimentally challenged coyotes did not become infected, suggesting that they are not competent hosts for SARS-CoV-2 [12]. Since the beginning of the pandemic, several variants of concern have emerged, including Alpha, Beta, Delta, and Omicron. These variants arise from viral mutations, particularly in the spike protein, and differ in transmissibility, pathogenicity, and their impact on diagnostics and vaccine performance [13].
SARS-CoV-2 is a novel zoonotic virus with significant zoonotic and reverse zoonotic potential. It is assumed that the virus initially spilled over from wildlife to humans and subsequently spread predominantly through human-to-human transmission via close contact and respiratory droplets. However, the virus has also been confirmed in companion animals (dogs and cats), ferrets, and deer, indicating its ability to cross species barriers and cause reverse zoonotic transmission [14,15]. Situations involving close contact between humans and animals, such as pet ownership, hunting activities, and farming, are of particular concern. Notably, SARS-CoV-2 has been detected in white-tailed deer living in proximity to urban areas, suggesting human-to-deer spillover followed by deer-to-deer transmission [16]. Of special interest are minks that are highly susceptible to COVID-19 and develop severe respiratory illness with 35-55% of mortality. Outbreaks on mink farms in the Denmark further demonstrated efficient human-to-animal transmission, and viral mutations observed in mink were subsequently associated with spillback and transmission of infection back to humans, which resulted in culling of minks and closing of mink farms [17].
Numerous studies have investigated the presence of SARS-CoV-2 in domestic, wild, and zoo animals to better understand viral origin and identify potential animal reservoirs. According to reports from the World Organization for Animal Health (WOAH), since March 2020 the virus spread to 36 countries and was detected in at least 29 animal species across 775 reported events [18]. Carnivores are considered to be of a higher risk to acquire SARS-CoV-2 infection. Studies in domestic and wild carnivores indicate that infection is often asymptomatic or associated with mild and transient viremia, although mild gastrointestinal and respiratory signs have been reported, particularly in felids. SARS-CoV-2 has been confirmed in wild felids such as lions, tigers, and pumas, as well as in wild canids, including foxes and jackals. In contrast, pigs, cattle, and sheep appear to be resistant to SARS-CoV-2 infection [19,20].
Serological investigation plays a crucial role in detecting past exposure to SARS-CoV-2 in animal population, particularly because active infection is transient and virus is difficult to detect by molecular methods after a week whereas antibodies persist for longer periods. IgM antibodies are detectable for a limited time, followed by the development of IgG antibodies, which may be detected for up to one year, although titers decline significantly after approximately six months [21,22].
In Serbia, the first COVID-19 case was registered on 6 March 2020. According to official data from the Institute of Public Health of the Republic of Serbia “Dr Milan Jovanović Batut” and the Ministry of Health, a total of 2,459,432 confirmed COVID-19 cases and 17,606 deaths were recorded by January 2023 [23]. Data on SARS-CoV-2 infection in animals in Serbia remain scarce. Nevertheless, several studies have confirmed the presence of SARS-CoV-2 RNA and antibodies in owned dogs and cats [24,25]. Detection of SARS-CoV-2 in domestic carnivores suggests that wild carnivores may also be susceptible to infection [12].
Wild carnivores in Serbia are of particular interest as potential viral sentinels due to increasing population sizes, mostly resulting from the absence of natural predators and the implementation of oral rabies vaccination programs since 2010. According to the Statistical Yearbook of the Republic of Serbia 2023 [26], the estimated population size of red foxes is 53,999 individuals, while the golden jackal population numbers approximately 22,596. Red foxes are widely distributed throughout Serbia, whereas golden jackals are more densely populated in lowland areas, particularly in northern and central regions of the country.
The aim of this study was to assess the exposure of wild canids, specifically red foxes (Vulpes vulpes) and golden jackals (Canis aureus), to SARS-CoV-2 in Serbia and to evaluate their potential role in virus circulation at the wildlife–human interface. Blood samples from 165 free-living foxes and jackals were tested for the presence of anti-SARS-CoV-2 antibodies using a commercial ELISA and a parallel in-house ELISA assay.
2. Materials and Methods
2.1. Sample Collection and Geographic Distribution
A total of 165 blood samples from red foxes (Vulpes vulpes) and golden jackals (Canis aureus) were collected for serological testing. Of these, 83 samples originated from foxes and 82 from jackals. The samples were randomly selected based on sample quality, ensuring balanced geographical representation across all study areas. Samples from red foxes and golden jackals were collected from the beginning of August 2024 to the end of March 2025 within the framework of the annual national program for monitoring the effectiveness of oral rabies vaccination in foxes and other wild carnivores, which has been implemented in Serbia since 2011. The program is established by the Veterinary Directorate of the Ministry of Agriculture, Forestry and Water Management of the Republic of Serbia and is conducted in accordance with the Rulebook on the Program of Animal Health Protection Measures for 2024 [27]. All sampled animals were older than one year, while data on animal sex were not available. Samples were collected from ten administrative districts including the City of Belgrade, covering a total of 26 municipalities (Table 1).
Blood specimens from red foxes and golden jackals were collected directly from the heart of carcasses by trained hunters. Blood was collected into sterile vacuum blood collection tubes without anticoagulant. The samples were transported to the Virology Department of the Institute of Veterinary Medicine of Serbia (IVMS) in the portable fridges with ice packs to maintain the cold chain during transport. In the laboratory, blood samples were kept at room temperature to allow spontaneous coagulation and clot retraction. After serum separation, samples were centrifuged for 10 min at 2500 rpm. The sera were subsequently heat-inactivated at 56 °C for 30 min. Aliquots of the processed sera were transferred into sterile microtubes and stored at −20 °C until testing.
2.2. Serological Screening by Enzyme-Linked Immunosorbent Assay (ELISA)
Serological screening for specific anti-SARS-CoV-2 IgG antibodies was performed using a commercially available and validated ELISA kit, while an in-house ELISA developed at Institute for the Application of Nuclear Energy (INEP) was applied to selected samples as the comparative method.
2.2.1. Commercial ELISA
Laboratory analyses were conducted with a commercial ELISA kit “ID Screen® SARS-CoV-2 Double Antigen Multi-Species” (IDVet, Grabels, France), designed for the detection of antibodies against the nucleocapsid (N) protein of SARS-CoV-2. The assay is based on an indirect ELISA format and is suitable for use in multiple animal species. All procedures were carried out in accordance with the manufacturer’s instructions.
Briefly, serum samples were diluted 1:2 in the provided dilution buffer, and 100 µL of diluted samples, positive control, and negative control were added to microplate wells pre-coated with recombinant SARS-CoV-2 N antigen. Plates were incubated for 45 min at 37°C. After incubation, wells were washed five times with 300 µL of wash buffer to remove unbound material. Subsequently, 100 µL of horseradish peroxidase (HRP)-conjugated multispecies secondary antibody was added to each well, followed by incubation for 30 min at room temperature. Plates were then washed five times as described above. Next, 100 µL of tetramethylbenzidine (TMB) substrate solution was added to each well and, incubated for 20 min at room temperature in the dark. The enzymatic reaction was stopped by adding 100 µL of stop solution, and optical density (OD) of each sample was measured at 450 nm using a spectrophotometer Victor 3V (PerkinElmer 1420 multilabel counter).
Results were expressed as S/P percentage (S/P,%) and interpreted according to the manufacturer’s criteria. Serum samples with S/P values below 50% were considered negative, samples with S/P values between 50% and 60% were classified as doubtful, and samples with S/P values equal to or above 60% were considered positive.
2.2.2. In-House ELISA
An indirect ELISA (iELISA), originally developed for the detection of anti–SARS-CoV-2 IgG antibodies in human COVID-19 patients [28,29] and registered by the Medicines and Medical Devices Agency of Serbia (ELISA SARS-CoV-2 IgG; Certificate of registration No. 515-02-02373-20-003, dated 14 July 2020), was subsequently modified for serological testing of wild canid blood samples. The assay was adapted by replacing the original anti-human IgG secondary antibodies with anti-dog IgG, allowing the quantification of the immune response in wild canids.
The in-house ELISA procedure applied in this study is described below:
High-binding 96-well polystyrene microplates were coated overnight at 4 °C with 100 µL per well of recombinant SARS-CoV-2 nucleocapsid (N) protein (ACRO, USA; CAT# NUN-C5227) at a final concentration of 1 µg per well in carbonate–bicarbonate coating buffer (pH 9.0). Plates were washed twice with phosphate-buffered saline (PBS, pH 7.5), stabilized with 200 µL of 1% sucrose per well for 15 min at room temperature, air-dried overnight, and stored at 4 °C until use.
Serum samples were diluted 1:100 in PBST supplemented with 1% bovine serum albumin (BSA), and 100 µL of each diluted sample was added per well. Plates were incubated for 1 h at 37 °C and washed five times with PBST. Horseradish peroxidase–conjugated rabbit anti-dog IgG secondary antibody (CAT No. ab112828, Abcam, UK), diluted 1:1000 in PBST with 1% BSA, was added at 100 µL per well and incubated for 1 h at 37 °C, followed by washing.
Color development was achieved by adding 50 µL of TMB substrate solution A followed by 50 µL of TMB substrate solution B per well, incubating for 20 min at 37 °C, and stopping the reaction with 100 µL of sulfuric acid (H₂SO₄). Optical density (OD) was measured at 450 nm using a microplate reader (Victor 3V, PerkinElmer, USA).
Positive and negative control sera from red foxes and golden jackals were selected based on their results obtained with the commercially ELISA kit and pooled accordingly. Blank wells containing 100 µL PBST with 1% BSA were included. The cut-off value was defined as the mean OD of negative sera plus three standard deviations. Samples with OD values above the cut-off were considered positive.
2.3. Statistical Analyses
Differences in SARS-CoV-2 seroprevalence among species and regions were analyzed by Pearson’s chi-squared test, with 95% confidence intervals (CI). Ninety-five percent confidence intervals (95% CI) were calculated to assess the precision of seroprevalence estimates across species and regions. A p-value < 0.05 was considered statistically significant. Statistical analyses were performed using Microsoft Excel 2007 (Microsoft Corp., Redmond, WA, USA) and SPSS Statistics version 19.0 (IBM Corp., Armonk, NY, USA).
Agreement between the commercial ELISA and the adapted assay was evaluated using Cohen’s kappa (κ) coefficient [30]. The strength of agreement was interpreted according to commonly accepted criteria (poor: κ < 0.20; fair: 0.21–0.40; moderate: 0.41–0.60; substantial: 0.61–0.80; almost perfect: > 0.80).
For data visualisation was used QGIS Development Team (2026). _QGIS Geographic Information System. Open Source Geospatial Foundation Project. Available at: https://qgis.org.
3. Results
Serological screening conducted by commercial ELISA revealed that 17 (10.3%, 95% CI: 5.6–14.9) of 165 tested red foxes and golden jackals were seropositive for SARS-CoV-2. Among 83 red foxes, antibodies against SARS-CoV-2 were detected in 11 animals (13.3%, 95% CI: 6.8–22.5), whereas reactive sera were identified in 6 of 82 jackals (7.3%, 95% CI: 2.7–15). Despite the higher antibody detection rate in red foxes, no statistically significant difference between species was observed (p = 0.21).
Samples were collected across 10 regions, including the City of Belgrade, but seropositive animals were found in only four districts. The highest seroprevalence was observed in the Južnobanatski District, with 11 positive animals (35.5%, 95% CI: 19.2–54.6), including 8 red foxes and 3 jackals. In the Mačva District, 3 animals had anti-SARS-CoV-2 antibodies (15.8%, 95% CI: 3.4–39.6; 2 foxes, 1 jackal), followed by the Kolubara District with 2 positive (13.3%, 95% CI: 1.7–40.5; 1 fox, 1 jackal) and the Braničevo District with a single positive jackal (3.6%, 95% CI: 0.1–18.3) (Figure 1). Differences in seroprevalence among regions were statistically significant (p < 0.001).
Notably, Južnobanatski District accounted for 72.7% of all seropositive red foxes and 50% of jackals with detectable antibodies, highlighting this district as the main hotspot of SARS-CoV-2 exposure. Overall, the highest seroprevalence was observed in the northern part of Serbia, in Vojvodina region, indicating both species-specific and regional patterns of exposure among wild canids.
All samples were additionally tested with the adapted ELISA. Antibodies against SARS-CoV-2 were detected in 12 of 165 animals (7.3%; 95% CI: 3.8–12.4%), including 8 of 83 red foxes (9.6%; 95% CI: 4.3–18.1%) and 4 of 82 golden jackals (4.9%; 95% CI: 1.3–12.0%). The adapted ELISA showed substantial agreement with the commercial assay (Cohen’s κ = 0.66), suggesting comparable performance between the two methods under the conditions of this study.
4. Discussion
Surveys describing cases of SARS-CoV-2 infection in animals have so far been widely reported in pets, particularly domestic cats and dogs, across North America, Europe, Asia, and South America [31,32]. However, data on SARS-CoV-2 exposure in wild canids remain scarce, despite the fact that the wildlife species are well-known reservoirs of viral pathogens.
In the present study, we assessed SARS-CoV-2 exposure in free-ranging wild canids, specifically red foxes (Vulpes vulpes) and golden jackals (Canis aureus) in Serbia. Our results revealed that 17 out of 165 tested animals were seropositive, corresponding to an overall seroprevalence of 10.3%. Comparable studies conducted in other European countries have demonstrated heterogeneous results. In Croatia, ELISA reactivity was detected in 2.9% of red foxes and 4.6% of tested golden jackals [20]. In Switzerland, serological exposure to SARS-CoV-2 was confirmed in free-ranging red foxes, with an estimated seroprevalence of 3.1% [33]. In another Swiss study, only one red fox was found SARS-CoV-2 positive by qRT-PCR among 246 samples collected from 153 animals housed in zoos [34]. Conversely, a study from Poland did not identify any positive red foxes in either molecular or serological investigations involving 292 hunted animals [35]. We assume that published findings vary significantly depending on the species studied, methodological approaches, sample sizes, and SARS-CoV-2 variants circulating during the pandemic, which was consistent with previous observations [31,36,37].
Furthermore, data from investigations involving other wild canids are similarly limited. Goldberg et al. (2024) [38], during surveillance of wildlife communities across Virginia and Washington D.C., USA, detected SARS-CoV-2 antibodies in 36.4% of tested raccoons. In contrast, several studies failed to confirm SARS-CoV-2 infection in wolves, either in free-ranging populations or zoo-housed animals [39]. Similarly, no SARS-CoV-2-positive coyotes or eastern wolves were detected by RT-PCR in surveillance studies conducted in Ontario and Québec, Canada [3,40].
The fox and jackal samples included in this study originated from ten districts, including the City of Belgrade, while positive samples were detected in only four districts (Južnobanatski District, Mačva District, Kolubara District, and Braničevo District). The observed distribution of seropositive animals reflects the epidemiological situation regarding SARS-CoV-2 in the human population in Serbia. Following several waves of COVID-19 between March 6, 2020 and 2022, a period of reduced infection rates was recorded. However, starting from May 2024, SARS-CoV-2 showed moderate fluctuations throughout the season, with a noticeable maximum (103.1 cases per 100,000 population) at the end of September 2024 [41]. Since animal samples in our study were collected from summer 2024 to spring 2025, we assumed that there is a positive relationship between the proximity of urban areas and seropositivity in wild canids, and these results coincide with the highest peak in the post-COVID-19 season in 2024 in Serbia. Similar associations between urbanization and seropositivity in wildlife have been reported previously [38].
An exceptionally high incidence rate of SARS-CoV-2 in humans was recorded in the Južnobanatski District (2646.55 per 100,000 inhabitants) which is consistent with our findings in wild canids from the same region. Significantly, 72.7% of seropositive foxes and 50% of seropositive jackals originated from the same district. Our data are in agreement with earlier studies indicating that animals living in close proximity to humans may face an increased risk of exposure [32]. Jackals and foxes are known to approach human habitations, farms, and zoos and likely mix with domestic animals and humans [35,42]. Foxes and jackals are known to transmit numerous pathogens, many of which have zoonotic potential [43]. Red foxes live and move in small groups while jackals generally live in pairs and have territorial behavior. Jackals are omnivores and scavengers. Both species feed on rabbits, skunks, rodents, and are in close contact with other potential sources and reservoirs of SARS-CoV-2 [12,20].
In the present study, red foxes showed a higher seroprevalence than golden jackals, with antibodies detected in 11 of 83 foxes (13.3%) and 6 of 82 jackals (7.3%). This is in line with findings reported by Porter et al. (2022) [12], who showed that experimentally infected foxes developed mild clinical signs and were capable of viral shedding, whereas infected coyotes remained asymptomatic. Such results highlight the importance of further investigations in red foxes, which may represent a potential wildlife host. Additionally, it has been that red foxes are more susceptible to infection because they possess the ACE2 receptor, which facilitates SARS-CoV-2 binding and entry [36,44].
Serological assays are important diagnostic tools for SARS-CoV-2 surveillance, particularly in wildlife studies. Although RT-PCR remains the reference method for confirming active SARS-CoV-2 infection, its application in wildlife surveillance presents several practical limitations. The persistence of viral RNA is transient, the success of detection is highly dependent on the timing of sampling, and the quality of the sample is often inadequate. For these reasons, serological approaches are often more suitable for population-based surveillance and retrospective assessment of viral circulation [21].
Among serological techniques, ELISA assays offer several practical advantages. They are technically less demanding, cost-effective, suitable for high-throughput screening, and do not require biosafety level-3 (BSL-3) facilities, unlike virus neutralization tests, which are considered the gold standard but are time-consuming and require trained personnel [21,45].
Many commercial ELISA assays for SARS-CoV-2 are based on the nucleocapsid (N) protein. The N protein is highly immunogenic, abundantly expressed during infection, and elicits a strong antibody response, making it a suitable target for serological detection [46,47]. In the present study, we used a commercial multi-species ELISA from Innovative Diagnostics (IDVet), France based on the N protein. This assay has been widely applied across a broad range of animal species, including cats, dogs, rabbits, bats, wild boars, ferrets, goats, lions, cattle, and horses [48,49,50,51,52]. This test has shown high diagnostic performance, with reported sensitivity and specificity of 99.1%, as indicated by the manufacturer and supported by previous evaluations [49,51].
All samples were additionally tested using an adapted in-house ELISA. The lower number of seropositive results obtained with the adapted assay (12/165; 7.3%) compared to the commercial ELISA (17/165; 10.3%) can be explained by methodological differences between the assays. Variations in antigen presentation, assay design, and cut-off determination can influence the classification of samples, particularly those with low antibody levels [53]. Commercial assays are typically validated using large panels of well-characterized samples. In contrast, in-house assays are often developed on smaller sample sets, which may result in reduced sensitivity. Despite these differences, the substantial agreement observed between assays (Cohen’s κ = 0.66) indicates that the two approaches were broadly comparable. Similar levels of concordance between commercial and in-house serological assays have been reported previously [54].
As far as we know, this study provides the first evidence of SARS-CoV-2 exposure in wild canids in Serbia. Certain limitations should also be considered. The geographic distribution of sampled areas was not fully balanced, and the number of collected samples varied among districts. A more uniform sampling strategy would allow a clearer interpretation of the spatial distribution of SARS-CoV-2 exposure.
Finally, further optimization and validation of serological assays for wildlife samples remain essential. As emphasized by Jemeršić et al. (2021) [20], particular attention should be given to sampling methodology, species tested, and sample quality. Wildlife samples are frequently affected by hemolysis and variable storage conditions, which can affect the accuracy of results.
Conclusions
In conclusion, this study provides evidence of SARS-CoV-2 exposure in wild canids, red foxes and golden jackals, in Serbia, highlighting that wildlife species living close to human environments may be affected by emerging pathogens. Continued surveillance, including a broader range of species and longitudinal monitoring, is essential. Within a One Health framework, integrated monitoring of pathogen circulation at the human–animal–environment interface remains key for understanding of viral dynamics, potential spillover risks and future zoonotic threats.
Author Contributions
Conceptualization, D.L. and J.M.Z.; methodology, B.K, N.K. and M.G.; investigation, J.M.Z. and M.K.; software, B.M; data curation, N.J. and B.M.; writing—original draft preparation D.L., A.A. and B.K.; writing—review and editing, M.G., N.J. and N.K.; supervision D.L. and B.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Science, Technological Development and Innovation of Republic of Serbia by the Contract of implementation and funding of research work for 2026, Contract No: 451-03-33/2026-03/ 200019 and 451-03-33/2026-03/ 200030.
Institutional Review Board Statement
Ethical review and approval were waived for this study, as all samples originated from archived serum collections obtained within the annual national oral rabies vaccination monitoring program implemented by the Veterinary Directorate of the Republic of Serbia (as part of the Program of Animal Health Protection Measures, Official Gazette of the Republic of Serbia, No. 21/2024), conducted in accordance with national legislation. The study did not involve the sacrifice of animals, and no animals were captured or handled specifically for research purposes; therefore, ethical approval was not required.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Naqvi, A.A.T.; Fatima, K.; Mohammad, T.; Fatima, U.; Singh, I.K.; Singh, A.; Atif, S.M.; Hariprasad, G.; Hasan, G.M.; Hassan, M.I. Insights into SARS-CoV-2 genome, structure, evolution, pathogenesis and therapies: Structural genomics approach. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165878. [Google Scholar] [CrossRef] [PubMed]
- Decaro, N.; Buonavoglia, C. An update on canine coronaviruses: Viral evolution and pathobiology. Vet. Microbiol. 2008, 132, 221–234. [Google Scholar] [CrossRef]
- Whittaker, G.; Stout, A. Coronaviruses in wild canids: A review of the literature. Qeios 2022. [Google Scholar] [CrossRef]
- Coronaviridae Study Group of the International Committee on Taxonomy of Viruses. The species Severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [CrossRef]
- Harcourt, J.; Tamin, A.; Lu, X.; Kamili, S.; Sakthivel, S.K.; Murray, J.; et al. Severe acute respiratory syndrome coronavirus 2 from a patient with coronavirus disease, United States. Emerg. Infect. Dis. 2020, 26, 1266–1273. [Google Scholar] [CrossRef]
- Zhou, F.; Yu, T.; Du, R.; Fan, G.; Liu, Y.; Liu, Z.; et al. Clinical course and risk factors for mortality of adult in patients with COVID-19 in Wuhan, China: A retrospective cohort study. Lancet 2020, 395, 1054–1062. [Google Scholar] [CrossRef]
- World Health Organization. Coronavirus disease (COVID-2019) situation reports. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019/situation-reports/situation-reports-archive (accessed on 4 January 2026).
- LeDuc, J.W.; Barry, M.A. SARS, the first pandemic of the 21st century. Emerg. Infect. Dis. 2004, 10. [Google Scholar] [CrossRef]
- Zaki, A.M.; van Boheemen, S.; Bestebroer, T.M.; Osterhaus, A.D.; Fouchier, R.A. Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia. N. Engl. J. Med. 2012, 367, 1814–1820. [Google Scholar] [CrossRef]
- Lytras, S.; et al. The animal origin of SARS-CoV-2. Science 2021, 373, 968–970. [Google Scholar] [CrossRef] [PubMed]
- Fischhoff, I.R.; Castellanos, A.A.; Rodrigues, J.P.G.L.M.; Varsani, A.; Han, B.A. Predicting the zoonotic capacity of mammals to transmit SARS-CoV-2. Proc. R. Soc. B 2021, 288, 20211651. [Google Scholar] [CrossRef] [PubMed]
- Porter, S.M.; Hartwig, A.E.; Bielefeldt-Ohmann, H.; Bosco-Lauth, A.M.; Root, J.J. Susceptibility of wild canids to SARS-CoV-2. Emerg. Infect. Dis. 2022, 28, 1852–1855. [Google Scholar] [CrossRef]
- Andre, M.; Lau, L.S.; Pokharel, M.D.; Ramelow, J.; Owens, F.; Souchak, J.; Akkaoui, J.; Ales, E.; Brown, H.; Shil, R.; Nazaire, V.; Manevski, M.; Paul, N.P.; Esteban-Lopez, M.; Ceyhan, Y.; El-Hage, N. From Alpha to Omicron: How different variants of concern of the SARS-coronavirus-2 impacted the world. Biology 2023, 12, 1267. [Google Scholar] [CrossRef]
- Decaro, N.; Balboni, A.; Bertolotti, L.; Martino, P.A.; Mazzei, M.; Mira, F.; Pagnini, U. SARS-CoV-2 infection in dogs and cats: Facts and speculations. Front. Vet. Sci. 2021, 8, 619207. [Google Scholar] [CrossRef] [PubMed]
- Munir, K.; Ashraf, S.; Munir, I.; Khalid, H.; Muneer, M.A.; Mukhtar, N.; Amin, S.; Ashraf, S.; Imran, M.A.; Chaudhry, U.; et al. Zoonotic and reverse zoonotic events of SARS-CoV-2 and their impact on global health. Emerg. Microbes Infect. 2020, 9, 2222–2235. [Google Scholar] [CrossRef]
- Pickering, B.; Lung, O.; Maguire, F.; Kruczkiewicz, P.; Kotwa, J.D.; Buchanan, T.; Gagnier, M.; Guthrie, J.L.; Jardine, C.M.; Marchand-Austin, A.; Massé, A.; McClinchey, H.; Nirmalarajah, K.; Aftanas, P.; Blais-Savoie, J.; Chee, H.Y.; Chien, E.; Yim, W.; Banete, A.; Griffin, B.D.; Yip, L.; Goolia, M.; Suderman, M.; Pinette, M.; Smith, G.; Sullivan, D.; Rudar, J.; Vernygora, O.; Adey, E.; Nebroski, M.; Goyette, G.; Finzi, A.; Laroche, G.; Ariana, A.; Vahkal, B.; Côté, M.; McGeer, A.J.; Nituch, L.; Mubareka, S.; Bowman, J. Divergent SARS-CoV-2 variant emerges in white-tailed deer with deer-to-human transmission. Nat. Microbiol. 2022, 7, 2011–2024. [Google Scholar] [CrossRef]
- Larsen, H.D.; Fonager, J.; Lomholt, F.K.; Dalby, T.; Benedetti, G.; Kristensen, B.; Urth, T.R.; Rasmussen, M.; Lassaunière, R.; Rasmussen, T.B.; Strandbygaard, B.; Lohse, L.; Chaine, M.; Møller, K.L.; Berthelsen, A.N.; Nørgaard, S.K.; Sönksen, U.W.; Boklund, A.E.; Hammer, A.S.; Belsham, G.J.; Krause, T.G.; Mortensen, S.; Bøtner, A.; Fomsgaard, A.; Mølbak, K. Preliminary report of an outbreak of SARS-CoV-2 in mink and mink farmers associated with community spread, Denmark, June to November 2020. Euro Surveill. 2021, 26, 2100009. [Google Scholar] [CrossRef]
- World Organisation for Animal Health (WOAH). Cases of SARS-CoV-2 infection in animals reported to WOAH since March 2020. Available online: https://www.woah.org/en/disease/sars-cov-2/#ui-id-2 (accessed on 21 February 2026).
- Shi, J.; Wen, Z.; Zhong, G.; Yang, H.; Wang, C.; Huang, B.; et al. Susceptibility of ferrets, cats, dogs, and other domesticated animals to SARS-coronavirus 2. Science 2020, 368, 1016–1020. [Google Scholar] [CrossRef]
- Jemeršić, L.; Lojkić, I.; Krešić, N.; Keros, T.; Zelenika, T.A.; Jurinović, L.; Skok, D.; Bata, I.; Boras, J.; Habrun, B.; Brnić, D. Investigating the presence of SARS-CoV-2 in free-living and captive animals. Pathogens 2021, 10, 635. [Google Scholar] [CrossRef]
- Miller, M.R.; Braun, E.; Ip, H.S.; Tyson, G.H. Domestic and wild animal samples and diagnostic testing for SARS-CoV-2. Vet. Q. 2023, 43, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hamer, S.A.; Pauvolid-Corrêa, A.; Zecca, I.B.; Davila, E.; Auckland, L.D.; Roundy, C.M.; Tang, W.; Torchetti, M.K.; Killian, M.L.; Jenkins-Moore, M.; et al. SARS-CoV-2 infections and viral isolations among serially tested cats and dogs in households with infected owners in Texas, USA. Viruses 2021, 13, 938. [Google Scholar] [CrossRef] [PubMed]
- Institute of Public Health of the Republic of Serbia “Dr Milan Jovanović Batut”. Health Statistical Yearbook of the Republic of Serbia 2023. 2024. Available online: https://www.batut.org.rs/download/aktuelno/04012023.pdf (accessed on 10 February 2026).
- Stanojević, S.; Radojičić, S.; Mišić, D.; Srejić, D.; Vasiljević, D.V.; Prokić, K.; Ilić, N. Frequency of SARS-CoV-2 infection in dogs and cats: Results of a retrospective serological survey in Šumadija District, Serbia. Prev. Vet. Med. 2022, 208, 105755. [Google Scholar] [CrossRef]
- Petrović, T.; Videnović, D.; Savić, S.; Vidanović, D.; Gajdov, V.; Lazić, G.; Samojlović, M.; Lupulović, D.; Lazić, S. Study on presence of SARS-CoV-2 in pet dogs and cats – Results from Serbia. In Proceedings of the 14th Annual Meeting EPIZONE, Barcelona, Spain, 18–20 May 2022. [Google Scholar]
- Statistical Office of the Republic of Serbia. Statistical Yearbook of the Republic of Serbia 2023. Available online: https://publikacije.stat.gov.rs/G2023/pdf/G20232056.pdf (accessed on 12 December 2025).
- Official Gazette of the Republic of Serbia. No. 21/2024. Available online: http://demo.paragraf.rs/demo/combined/Old/t/t2024_03/SG_021_2024_005.htm (accessed on 21 February 2026).
- Baralić, M.; Laušević, M.; Ćujić, D.; Bontić, A.; Pavlović, J.; Brković, V.; Kezić, A.; Mihajlovski, K.; Hadži Tanović, L.; Assi Milošević, I.; Lukić, J.; Gnjatović, M.; Todorović, A.; Stojanović, N.M.; Jovanović, D.; Radović, M. The importance of natural and acquired immunity to SARS-CoV-2 infection in patients on peritoneal dialysis. Vaccines 2024, 12, 135. [Google Scholar] [CrossRef]
- Kotur-Stevuljević, J.; Roksandić Milenković, M.; Vekić, J.; Dimić, N.; Dimić, D.; Ćujić, D.; Guzonjić, A.; Gnjatović, M.; Todorović, A.; Bogavac-Stanojević, N. Leukocyte telomere dynamic change in patients with mild to moderate COVID-19 during three weeks of follow-up: Relation with therapy. J. Med. Biochem. 2026, 45, 1–11. [Google Scholar] [CrossRef]
- Cohen, J. A coefficient of agreement for nominal scales. Educ. Psychol. Meas. 1960, 20, 37–46. [Google Scholar] [CrossRef]
- EFSA AHAW Panel (EFSA Panel on Animal Health and Animal Welfare); Nielsen, S.S.; Alvarez, J.; Bicout, D.J.; Calistri, P.; Canali, E.; Drewe, J.A.; Garin-Bastuji, B.; Gonzales Rojas, J.L.; Gortázar, C.; Herskin, M.; Michel, V.; Miranda Chueca, M.A.; Padalino, B.; Pasquali, P.; Roberts, H.C.; Spoolder, H.; Velarde, A.; Viltrop, A.; Winckler, C.; Adlhoch, C.; Aznar, I.; Baldinelli, F.; Boklund, A.; Broglia, A.; Gerhards, N.; Mur, L.; Nannapaneni, P.; Stahl, K. SARS-CoV-2 in animals: Susceptibility of animal species, risk for animal and public health, monitoring, prevention and control. EFSA J. 2023, 21, 7822. [Google Scholar] [CrossRef]
- Abay, Z.; Sadikaliyeva, S.; Nurpeisova, A.; Jekebekov, K.; Shorayeva, K.; Yespembetov, B.; Nurabayev, S.; Kerimbayev, A.; Khairullin, B.; Yoo, H.; et al. Breaking the Barrier: SARS-CoV-2 Infections in Wild and Companion Animals and Their Implications for Public Health. Viruses 2024, 16, 956. [Google Scholar] [CrossRef]
- Kuhn, J.; Marti, I.; Ryser-Degiorgis, M.-P.; Wernike, K.; Jones, S.; Tyson, G.; Delalay, G.; Scherrer, P.; Borel, S.; Hosie, M.J.; et al. Investigations on the Potential Role of Free-Ranging Wildlife as a Reservoir of SARS-CoV-2 in Switzerland. Viruses 2024, 16, 1407. [Google Scholar] [CrossRef] [PubMed]
- Chan, T.; Ginders, J.; Kuhlmeier, E.; Meli, M.L.; Bönzli, E.; Meili, T.; Hüttl, J.; Hatt, J.-M.; Hindenlang Clerc, K.; Kipar, A.; et al. Detection of SARS-CoV-2 RNA in a Zoo-Kept Red Fox (Vulpes vulpes). Viruses 2024, 16, 521. [Google Scholar] [CrossRef] [PubMed]
- Goll, A.; Krupińska, M.; Nowicka, J.; Baranowicz, K.; Rabalski, L.; Lass, A.; Gorska, A.; Sironen, T.; Kant, R.; Grzybek, M. Wild Red Foxes (Vulpes vulpes) Do Not Participate in SARS-CoV-2 Circulation in Poland. One Health 2024, 19, 100845. [Google Scholar] [CrossRef]
- Delahay, R.J.; de la Fuente, J.; Smith, G.C.; Sharun, K.; Snary, E.L.; Flores Girón, L.; Nziza, J.; Fooks, A.R.; Brookes, S.M.; Lean, F.Z.X.; et al. Assessing the Risks of SARS-CoV-2 in Wildlife. One Health Outlook 2021, 3, 7. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.Y.; Huang, J.S.; Gong, Q.L.; Sun, J.M.; Li, Y.J.; Liu, B.; Zhang, Y.M.; Shi, C.W.; Yang, G.L.; Yang, W.T.; Wang, C.F. SARS-CoV-2 Prevalence in Wildlife 2020–2022: A Worldwide Systematic Review and Meta-Analysis. Microbes Infect. 2024, 26, 105350. [Google Scholar] [CrossRef]
- Goldberg, A.R.; Langwig, K.E.; Brown, K.L.; Marano, J.M.; Rai, P.; King, K.M.; Sharp, A.K.; Ceci, A.; Kailing, C.D.; Kailing, M.J.; et al. Widespread Exposure to SARS-CoV-2 in Wildlife Communities. Nat. Commun. 2024, 15, 6210. [Google Scholar] [CrossRef]
- Tinto, B.; Revel, J.; Virolle, L.; Chenet, B.; Reboul Salze, F.; Ortega, A.; Beltrame, M.; Simonin, Y. Monitoring SARS-CoV-2 Seroprevalence in Domestic and Exotic Animals in Southern France. Trop. Med. Infect. Dis. 2023, 8, 426. [Google Scholar] [CrossRef]
- Greenhorn, J.E.; Kotwa, J.D.; Bowman, J.; Bruce, L.; Buchanan, T.; Buck, P.A.; Davy, C.M.; Dibernardo, A.; Flockhart, L.; Gagnier, M.; et al. SARS-CoV-2 Wildlife Surveillance in Ontario and Québec. Can. Commun. Dis. Rep. 2022, 48, 243–251. [Google Scholar] [CrossRef]
- Ristić, M.; et al. Sentinel Surveillance of Acute Respiratory Infections and Distribution of SARS-CoV-2, Influenza A and B, and Respiratory Syncytial Virus in the Post-COVID-19 Period. Vojnosanit. Pregl. 2025, 82, 490–503. [Google Scholar] [CrossRef]
- Campbell, S.J.; Ashley, W.; Gil-Fernandez, M.; Newsome, T.M.; Di Giallonardo, F.; Ortiz-Baez, A.S.; Mahar, J.E.; Towerton, A.L.; Gillings, M.; Holmes, E.C.; Carthey, A.J.R.; Geoghegan, J.L. Red Fox Viromes in Urban and Rural Landscapes. Virus Evol. 2020, 6, veaa065. [Google Scholar] [CrossRef] [PubMed]
- Davidson, W.R.; Appel, M.J.; Doster, G.L.; Baker, O.E.; Brown, J.F. Diseases and Parasites of Red Foxes, Gray Foxes, and Coyotes from Commercial Sources Selling to Fox-Chasing Enclosures. J. Wildl. Dis. 1992, 28, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Luan, J.; Lu, Y.; Jin, X.; Zhang, L. Spike Protein Recognition of Mammalian ACE2 Predicts the Host Range and an Optimized ACE2 for SARS-CoV-2 Infection. Biochem. Biophys. Res. Commun. 2020, 526, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Tindih Sheltone, H.; Obara, I.; Alessandro, M.; et al. First Detection of SARS-CoV-2-like Antibodies in Domestic and Wild Animals in Kenya. Authorea 2022, May 25.
- Liu, W.; Liu, L.; Kou, G.; Zheng, Y.; Ding, Y.; Ni, W.; Wang, Q.; Tan, L.; Wu, W.; Tang, S.; Xiong, Z. Evaluation of Nucleocapsid and Spike Protein-Based Enzyme-Linked Immunosorbent Assays for Detecting Antibodies against SARS-CoV-2. J. Clin. Microbiol. 2020, 58, e00461-20. [Google Scholar] [CrossRef]
- Kontou, P.I.; Braliou, G.G.; Dimou, N.L.; Nikolopoulos, G.; Bagos, P.G. Antibody Tests in Detecting SARS-CoV-2 Infection: A Meta-Analysis. Diagnostics 2020, 10, 319. [Google Scholar] [CrossRef]
- Barua, S.; Hoque, M.; Adekanmbi, F.; Kelly, P.; Jenkins-Moore, M.; Torchetti, M.K.; Chenoweth, K.; Wood, T.; Wang, C.M. Antibodies to SARS-CoV-2 in Dogs and Cats, USA. Emerg. Microbes Infect. 2021, 10, 1669–1674. [Google Scholar] [CrossRef] [PubMed]
- Dias, J.M.; Costa, M.O.; de Souza, A.O.; Faria, M.L.; Golcman, D.H.; Selim, M.B.; Medici, E.P.; Haga, G.S.I.; Romaldini, A.H.C.N.; Okuda, L.H. Serological Assay for SARS-CoV-2 in Domestic and Wild Animals in Middle and Southeast Regions in Brazil. Fortune J. Health Sci. 2024, 7, 299–306. [Google Scholar]
- Pomorska Mól, M.; Turlewicz Podbielska, H.; Gogulski, M.; Ruszkowski, J.R.; Kubiak, M.; Kuriga, A.; Barket, P.; Postrzech, M. A Cross-Sectional Retrospective Study of SARS-CoV-2 Seroprevalence in Domestic Cats, Dogs and Rabbits in Poland. BMC Vet. Res. 2021, 17, 322. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Bastit, L.; Marfil, S.; Pradenas, E.; Blanco, J.; Vergara-Alert, J.; Segalés, J. Comparison of Three Commercial ELISA Kits for Detection of Antibodies Against SARS-CoV-2 in Serum Samples from Different Animal Species. Viruses 2025, 17, 716. [Google Scholar] [CrossRef] [PubMed]
- Udom, K.; Jairak, W.; Chamsai, E.; Charoenkul, K.; Boonyapisitsopa, S.; Bunpapong, N.; Techakriengkrai, N.; Amonsin, A. Serological Survey of Antibodies Against SARS-CoV-2 in Dogs and Cats, Thailand. Transbound. Emerg. Dis. 2022, 69, 2140–2147. [Google Scholar] [CrossRef]
- Whitman, J.D.; Hiatt, J.; Mowery, C.T.; Shy, B.R.; Yu, R.; Yamamoto, T.N.; Rathore, U.; Goldgof, G.M.; Whitty, C.; Woo, J.M.; et al. Test Performance Evaluation of SARS-CoV-2 Serological Assays. medRxiv [Preprint] 2020, 2020.04.25.20074856. Update in: Nat. Biotechnol. 2020, 38, 1174–1183. https://doi.org/10.1038/s41587-020-0659-0. [CrossRef]
- Gobena, D.; Gudina, E.K.; Degfie, T.T.; et al. Diagnostic Performance Evaluation of Elecsys Anti-SARS-CoV-2 Assay Against RT-PCR for SARS-CoV-2 Detection and Surveillance in Ethiopian Referral Hospitals: Cross-Sectional Study. Sci. Rep. 2026, 16, 5207. [Google Scholar] [CrossRef]
Figure 1.
Spatial distribution of tested samples and seropositive wild canids in Serbian districts. Inset: geographic position of Serbia within Europe.
Figure 1.
Spatial distribution of tested samples and seropositive wild canids in Serbian districts. Inset: geographic position of Serbia within Europe.
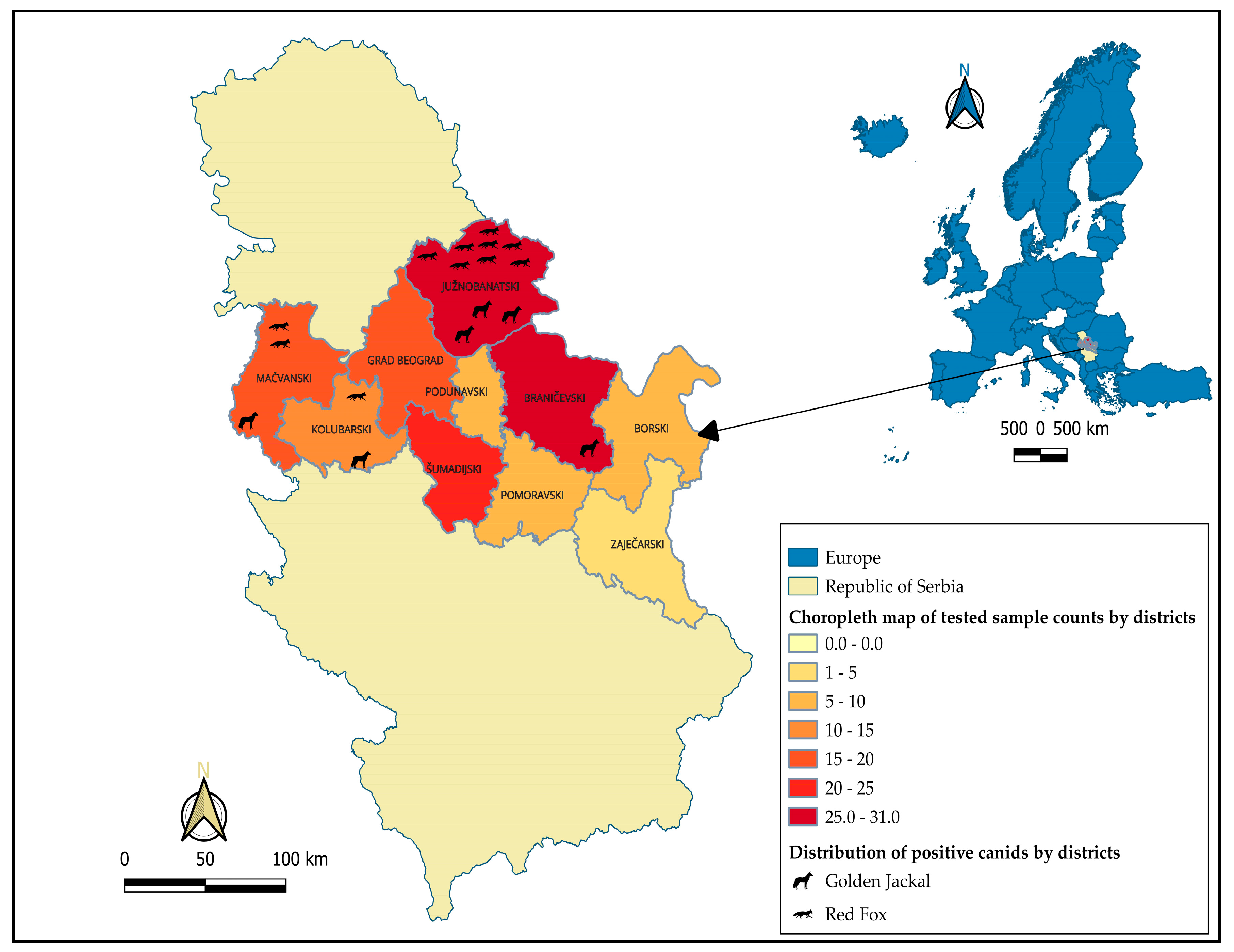

Table 1.
Number of blood samples collected by species and districts in Serbia.
| District | Species | Total | |
|---|---|---|---|
| Red fox | Golden jackal | ||
| Borski | 6 | 4 | 10 |
| Braničevski | 7 | 21 | 28 |
| Južnobanatski | 16 | 15 | 31 |
| Kolubarski | 13 | 2 | 15 |
| Mačvanski | 16 | 3 | 19 |
| Podunavski | 0 | 7 | 7 |
| Pomoravski | 1 | 5 | 6 |
| Šumadijski | 16 | 9 | 25 |
| Zaječarski | 1 | 3 | 4 |
| City of Belgrade | 7 | 13 | 20 |
| Total | 83 | 82 | 165 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



