Submitted:
24 February 2026
Posted:
25 February 2026
You are already at the latest version
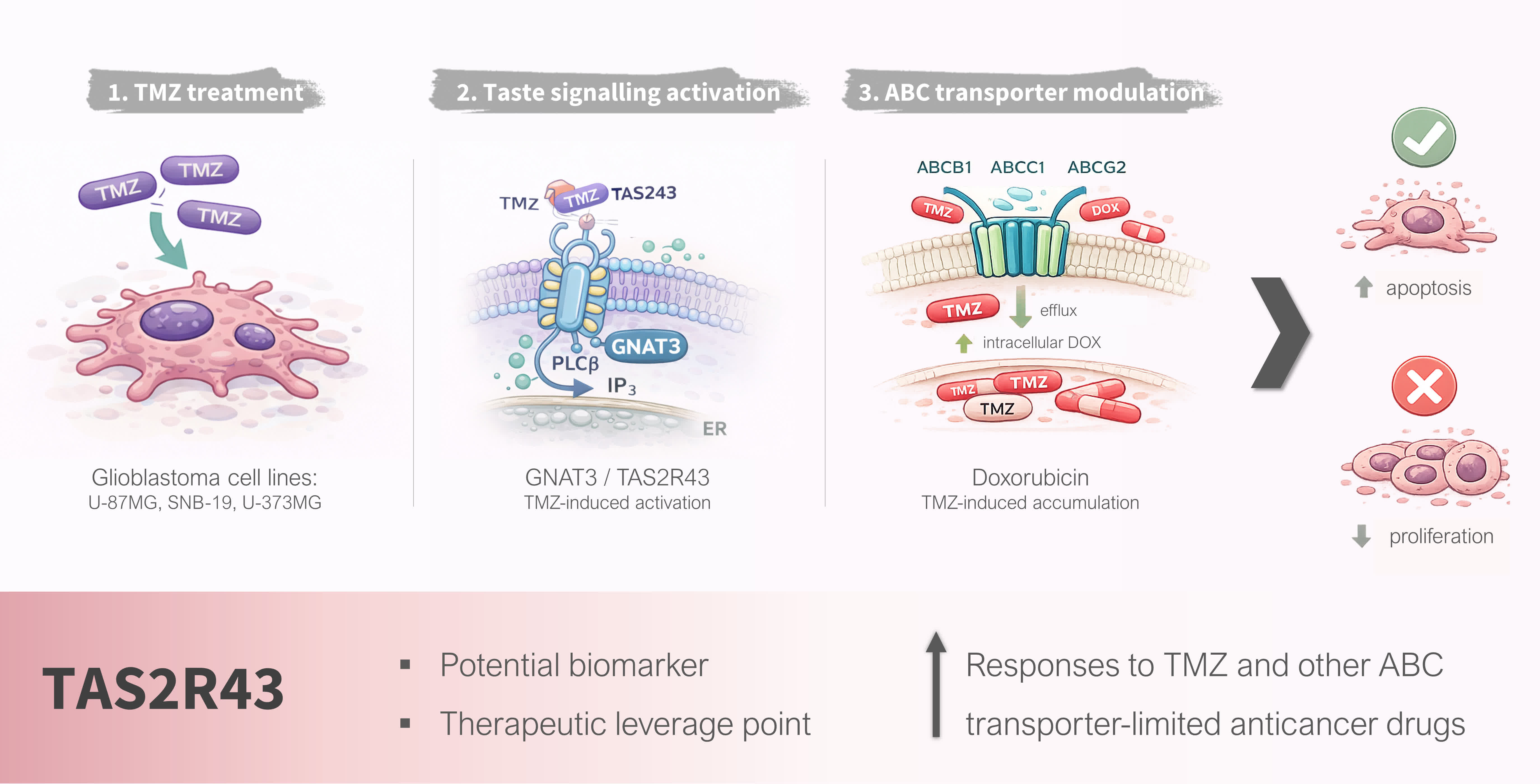
Abstract
Bitter taste receptors (TAS2Rs) are increasingly recognized as extraoral chemosensors that modulate diverse biological processes, including cancer cell behaviour and drug responsiveness. Many TAS2R ligands correspond to therapeutic compounds; however, their contribution to the response of brain tumours to chemotherapy remains unexplored. Here, we investigated whether the bitter taste signalling pathway is modulated by temozolomide (TMZ), the standard chemotherapeutic agent for glioblastoma, with an impact in treatment efficacy in glioblastoma cells. We show that TMZ elicits intracellular Ca²⁺ responses compatible with activation of GPCR signalling, and induces antiproliferative and pro-apoptotic effects in multiple human glioblastoma cell lines. Pharmacological inhibition of bitter taste receptors, as well as genetic silencing of the taste transduction G protein GNAT3, significantly attenuated TMZ-induced cytotoxicity, suggesting the involvement of bitter taste signalling in the process. In silico ligand prediction combined with receptor expression profiling identified TAS2R43 as a candidate mediator of these effects, and TAS2R43 knockdown markedly reduced TMZ-induced loss of cell viability, and apoptosis. Moreover, TMZ enhanced intracellular accumulation of the ABC transporter substrate doxorubicin, suggesting modulation of multidrug efflux mechanisms. Collectively, our findings identify TAS2R43 as a previously unrecognized modulator of glioblastoma responsiveness cells to TMZ and support a role for bitter taste signalling in shaping intracellular drug availability. These results highlight TAS2R43 as a potential biomarker and therapeutic leverage point to improve responses to TMZ and other ABC transporter-limited anticancer drugs.
Keywords:
glioblastoma
; temozolomide
; bitter taste receptors
; taste signalling
; TAS2R43
1. Introduction
Apart from sensing bitter compounds in the oral cavity, TAS2Rs have been also described in several extraoral tissues, like the gastrointestinal tract, airway epithelium, heart, testis and in the brain and brain barriers [1,2,3,4]. Experimental and clinical evidence indicates that TAS2Rs – members of the G protein-coupled receptor (GPCR) superfamily – and their ligands contribute to cancer progression and metastasis by regulating key processes such as tumorigenesis, proliferation, migration, invasion, and oncogenic signalling pathways [5,6]. However, whether TAS2R signalling influences the response of highly chemoresistant tumours such as glioblastoma to standard-of-care drugs remains unknown. Temozolomide (TMZ) is an alkylating agent widely used for the treatment of glioblastoma and other neoplasms of the central nervous system [7], known for its ability to methylate DNA, resulting in DNA damage and induction of tumour cells death by apoptosis [8]. However, its efficacy is limited because some tumour cells can repair TMZ-induced DNA damage through the action of O6-methylguanine DNA methyltransferase gene [9], relapsing of chemo- and radioresistant cancer stem-like cells and because these are highly infiltrative tumours [10,11,12]. Additionally, brain barriers, and brain cancer cells, overexpress multidrug efflux transporters which extrude several CNS-targeting medicines [13,14,15,16,17], which fail to achieve therapeutic concentrations at their target cells [18]. We recently identified 20 out of the 26 human TAS2R in glioblastoma cells that mediate signal transduction in response to a wide variety of bitter agonists in other cells [19,20,21].
Given its bitter properties, we hypothesized that TMZ might bind TAS2Rs and investigated the functional readout of this interaction. We first analysed if TMZ activates the bitter taste signalling pathway in glioblastoma cell lines and found that the presence of TAS2R43 is important for its anticancer efficacy in these cells.
2. Materials and Methods
2.1. Materials
Temozolomide (TMZ; CAS No 85622-93-1) was purchased from Cayman Chemical (#14163). A stock solution was prepared in dimethyl sulfoxide (DMSO) and freshly dissolved in Tyrode’s solution or culture medium before the experiments, where the DMSO final concentration did not exceed 1%. A vehicle control was included in all the experiments. Probenecid (CAS No 57-66-9), a known bitter taste receptors’ antagonist, was obtained from Sigma-Aldrich (#P8761), dissolved in 1 N NaOH at 0.17 M and diluted in Tyrode’s solution or culture medium. FURA-2AM (#F1221), pluronic acid F-127, Lipofectamine™ 2000 (#11668027), Opti-MEM medium (#11058-021), small interfering RNA (siRNA) targeting α-gustducin (GNAT3; #4392420; ID s51191; sequence: S-GCGAGAUGCAAGAACCGUATtt and AS-UACGGUUCUUGCAUCUCGCtc) and bitter taste receptor TAS2R43 (#AM16708; ID 202331; sequence: S-CCCUACUAUCUUUUAUGCUtt and AS-AGCAUAAAAGAUAGUAGGGtc), and scramble siRNA (#4390843) were purchased in ThermoFisher Scientific. MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] was purchased from Gerbu Biotechnik GmbH (#1006).
2.2. Cell Culture
Human malignant glioblastoma cell lines U-87MG and U-373MG were kindly provided by Dr. Joseph Costello (University of California, San Francisco), and SNB-19 was obtained from German Collection of Microorganisms and Cell Cultures. These glioblastoma cell lines have different morphology and proliferation rates: U-87MG are cluster invasion cells, whereas the most chemoresistant SNB-19 and U-373MG (similar to U-251MG) are individual invasion and expansive-growth cells, respectively [22,23].
All cell lines were grown in Dulbecco’s modified Eagle’s medium (DMEM) high glucose with stable glutamine (bioWest #L0103) supplemented with 10% (v/v) FBS and penicillin (100 IU/mL)/streptomycin (100 μg/mL), and incubated in a humidified atmosphere containing 5% CO2 at 37 ºC.
2.3. Evaluation of the Impact of TAS2R in TMZ Cytotoxicity
2.3.1. Effects of TMZ in Intracellular Ca2+ Responses of Glioblastoma Cells
Intracellular Ca2+ mobilisation is a marker of GPCR activation, including taste receptors. Thus, we measured intracellular Ca2+ oscillations in response to TMZ stimulus by single-cell Ca2+ imaging assays, in the presence or absence of the TAS2Rs antagonist probenecid, in glioblastoma SNB-19 and U-373MG cells. Briefly, approximately 3.5x104 glioblastoma cells were seeded in μ-slide 8 well ibiTreat (Ibidi #80826) and grown until 60-70% confluency, followed by measurement of changes in intracellular Ca2+ levels after stimulation. Glioblastoma cells were loaded with 5 μM of FURA-2 AM and 0.02% pluronic acid F-127 in culture medium for 45 min. Next, cells were washed twice with Tyrode’s solution pH 7.4 [NaCl 140 mM, KCl 5 mM, MgCl2 1.0 mM, CaCl2 2.0 mM, Na-pyruvate 10 mM, Glucose 10 mM, HEPES 10 mM, NaHCO3 5.0 mM] and loaded with Tyrode’s for 30 min. After that, dose-response experiments were performed with a range of TMZ concentrations (50-200 µM), in the presence or absence of 30 min incubation with probenecid (1 mM). The μ-slide plates were placed on a Widefield Axio Observer Z1 inverted microscope (Zeiss). Stock solution of TMZ and probenecid was freshly prepared in Tyrode’s solution before the experiments. The stimulus was applied manually with a micropipette after baseline was recorded. The intracellular Ca2+ levels were evaluated by quantifying the ratio of the fluorescence emitted at 520 nm following alternate excitation at 340 nm and 380 nm, using a Lambda DG4 apparatus (Sutter Instrument) and a 520 nm bandpass filter (Zeiss) under a Fluar 40x/1.30 Oil M27 objective (Zeiss). Data were processed using the Fiji software (MediaWiki). Changes in fluorescence ratio (F=F340/F380) were measured in at least 20 cells, in three or more independent experiments. Response intensity, or intracellular Ca2+ variation (ΔF/F0), was calculated in the following way: ΔF/F0=(F-F0)/F0, where F0 corresponds to fluorescence ratio average at baseline (2 min acquisition before stimulus) and F correspond to maximum peak of fluorescence ratio evoked by the stimulus applied to the cells.
2.3.2. Evaluation of the Impact of GNAT3 Knockdown in TMZ Cytotoxicity
After Ca2+ imaging experiments, the cytotoxicity of TMZ was assessed in glioblastoma cells. Initially, a range of TMZ concentrations (50-500 μM) were used, and significant effects in cell viability were only observed at 500 μM. Thus, MTT experiments with probenecid and GNAT3 knockdown were carried out using this TMZ concentration to gain further support, that specific activation of the bitter taste signalling is elicited by TMZ. The apoptotic effect of TMZ, in the presence or absence of probenecid, was also evaluated in glioblastoma cells by Hoechst 33342 nuclei staining. Glioblastoma cells were seeded in a coverslip and the stimuli were carried out by incubating the cells for 72 h with TMZ (500 μM) or vehicle (DMSO 1%), in the presence or absence of 1 mM probenecid diluted in culture medium. After removing the medium, cells were fixed with PFA 4% for 10 min followed by incubation for 10 min with Hoechst 33342 (diluted 1:1000). After several washes, cells were mounted onto microscope slides and visualized under a confocal microscope LSM 710 (Zeiss) using a magnification of 63x (Plan-Apochromat 63x/1.4 Oil DIC M27). Apoptotic cells were distinguished from healthy or necrotic cells by the observation of condensed DNA and fragmented nuclei. The apoptotic rate was calculated using the ratio between the number of apoptotic cells and the total number of cells.
For GNAT3 knockdown with specific siRNA, cells were grown until 60% confluency or transfected for 24 h with a mixture of GNAT3 siRNA (10 nM) and Lipofectamine™ 2000 in Opti-MEM medium, following the manufacturer’s instructions. A scramble siRNA (10 nM) was also used as negative control for GNAT3 specific targeting. Then, cells were incubated for 72 h with TMZ (500 μM) or vehicle (DMSO 1%), in the presence or absence of 1 mM probenecid diluted in culture medium. Then, 100 μL culture medium was removed and 10 μL of MTT solution (5 mg/mL in PBS) was added for approximately 45 min at 37 °C in a humidified atmosphere containing 5% CO2. Untreated cells and ethanol 70% treated cells were used as negative and positive controls, respectively. Following MTT incubation, formazan crystals were dissolved in DMSO for 15 min, and absorbance was read at 570 nm in a microplate spectrophotometer xMark™ (Bio-Rad). The viability of glioblastoma cells was expressed as a percentage of the absorbance determined in the vehicle control.
2.4. Identification of the TMZ Target TAS2R on Glioblastoma Cells
Following the demonstration that the bitter taste signalling pathway was involved in the mediation of the effects of TMZ, we assessed in-silico which TAS2R could have a higher likelihood of binding to TMZ according to the webserver VirtualTaste method [24]. Using this approach, six TAS2Rs – TAS2R10, R14, R38, R43, R44 and R45 – were identified as possible target-receptors for TMZ. TAS2R38 and TAS2R43 were both detected in glioblastoma cells (U-87MG, SNB-19, and U-373MG) by RT-PCR [25]. Additionally, TAS2R43 protein detection in glioblastoma cells was assessed by immunocytochemistry, as described before [25]. Finally, the role of TAS2R43 on the response to TMZ was assessed in glioblastoma cells after TAS2R43 knockdown with specific siRNA, as described in section 2.3.2 for GNAT3.
2.5. Effect of TMZ on ABC Transporters
Since TMZ is highly lipophilic and triggers cell apoptosis through DNA methylation and damage, we explored the hypothesis that the reduced effects of TMZ seen upon TAS2R43 knockdown could be a result of reduced cellular concentrations of the drug. Thus, we tested whether TAS2R43-mediated downregulation of efflux transporters like ABCB1, ABCC1 and ABCG2 [26]. These efflux transporters are expressed in glioblastoma and have been implicated in multidrug resistance and restricted brain penetration of anticancer drugs [27,28,29,30]. Thus, TMZ-induced intracellular accumulation of DOX, well-known substrate of these efflux transporters was evaluated in U-87MG, SNB-19 and U-373MG glioblastoma cells. In these assays, DOX was chosen as a fluorescent probe, allowing functional assessment of their activity in a single experimental setup. Reference inhibitors of ABCB1 (verapamil), ABCC1 (reversan) and ABCG2 (Ko143) were used, to dissect the contribution of each transporter to TMZ-induced changes in DOX intracellular accumulation [31,32,33].
Cells were seeded in multi-well plates and maintained under standard culture conditions until use. On the assay day, cells were washed and pre-incubated for 1 hour with verapamil (5 µM), reversan (10 µM) or Ko143 (100 nM), in the presence or absence of probenecid (1 mM). After this pre-incubation period, cells were incubated for 1 h with a solution containing DOX (1 µM) and TMZ (1 µM) to allow intracellular accumulation under the different conditions. At the end of the incubation, cells were washed thrice with ice-cold PBS, and lysed with Triton X-100 1% in PBS for 30 minutes at 37 ºC, protected from light. The fluorescence of DOX in the lysates was quantified using a SpectraMax Gemini spectrofluorometer (Molecular Devices) at excitation/emission wavelengths of 480/508 nm. In each plate, wells containing only DOX/TMZ solution were included as positive controls for fluorometric detection of each compound. Data were expressed as relative intracellular accumulation compared with the corresponding vehicle-treated controls (DMSO ≤ 1%).
2.6. Data Analysis
Statistical analysis and dataset comparisons were performed using GraphPad Prism 9.3.1 (GraphPad Software). Statistical significance was determined by One-Way ANOVA followed by the software’s recommended multiple comparisons post-hoc test. Results are presented as mean ± SEM of at least three independent experiments, and data were considered statistically different for a p-value<0.05.
3. Results
3.1. TMZ Elicits Intracellular Ca2+ Responses in Glioblastoma Cell Lines
We assessed the SNB-19 and U-373MG response to TMZ (10–200 μM) using a Ca2+ imaging assay (Figure 1) in the presence or absence of probenecid, a blocker of TAS2R16, R38, and R43, to assess if any of these receptors could interact with TMZ eliciting a taste receptor typical response in these cells, the mobilization of intracellular Ca2+ [34,35]. Our results showed that TMZ was able to trigger Ca2+ mobilization in these glioblastoma cells. TMZ at 100 µM (ΔF/F0=0.220±0.039) triggered a significant increase in intracellular Ca2+ levels in SNB-19 cells (Figure 1A) in comparison to the vehicle (ΔF/F0=0.025±0.008), abolished in the presence of probenecid (ΔF/F0=0.025±0.006). U-373MG cells stimulated with 50 and 100 µM TMZ showed higher Ca2+ levels (ΔF/F0=0.437±0.137 and ΔF/F0=0.718±0.155) in comparison with vehicle control (ΔF/F0=0.052±0.007) (Figure 1B). In addition, the TMZ effects on intracellular Ca2+ levels in the presence of probenecid were reverted (ΔF/F0=0.034±0.012). Neither 10 µM nor 200 µM TMZ triggered significant Ca2+ responses in both SNB-19 and U-373MG glioblastoma cells (Figure 1).
3.2. The Bitter Taste Signalling Pathway Enhances the Anticancer Effects of TMZ
Next, we proceeded to dose-response (50-500 µM) cytotoxicity assays, and found 500 µM TMZ was the minimum concentration required to reduce significantly the viability of U-87MG (49.08%±1.92), SNB-19 (44.46%±0.97) and U-373MG (50.92%±0.45) cells (Figure 2A).
Then, we evaluated the effects of TMZ on the apoptosis of glioblastoma cells in the presence or absence of probenecid, by staining the cell nuclei with Hoechst 33342 and counting the nuclei with apoptotic vesicles. As expected, TMZ induced apoptosis in these cells, but this effect was reduced to control levels in the presence of probenecid (Figure 2B). Our observations suggested that TMZ could be a TAS2Rs ligand and that the taste signalling pathway could modulate its effects or penetration in these cells. To test this hypothesis, we proceeded with knockdown experiments, of the specific guanine nucleotide binding protein that mediates taste receptor mediated signalling cascades, GNAT3 (Figure 3). TMZ alone induced a reduction in the cells’ viability of approximately 37.4% in U-87MG, 41.1% in SNB-19, and 45.9% in U-373MG in comparison with non-, mock- and siRNA scramble-transfected cells (Figure 3B). Notably, the presence of probenecid, or the silencing of GNAT3, reduced the effects of TMZ on the viability of glioblastoma cells (Figure 3B), reinforcing the results obtained in the previous experiments.
Overall, TMZ elicited Ca2+ responses in a dose-dependent manner and showed cytotoxic effects in glioblastoma cells, which were significantly reduced in the presence of probenecid or by GNAT3 silencing, highlighting that the activation of the bitter taste signalling pathway modulates TMZ effects in these cells.
3.3. TAS2R43 Mediates the Effects of TMZ in the Viability of Glioblastoma Cells
After confirming that the blockage of the bitter taste signalling pathway reduced the cytotoxic effect of TMZ in these cell lines, the next step was identifying which TAS2Rs were involved. Based on the webserver VirtualTaste predictions (Figure 4A), TMZ could bind six TAS2Rs: R38 (0.72) > R10 (0.68) > R43 (0.65) > R45 (0.62) > R44 (0.60) > R14 (0.59). However, only TAS2R38 and R43 bind probenecid [34,35], and the former was not expressed in all the cell lines analysed (Figure 4B).
Therefore, we proceeded with the analysis of glioblastoma cells viability upon TAS2R43 knockdown (Figure 5A). TMZ alone induced a reduction of approximately 41.78±2.44% in the viability of U-87MG, SNB-19 and U-373MG cells in comparison with non-, mock- and siRNA scramble-transfected cells but was unable to reduce the viability of TAS2R43-silenced cells (Figure 5B), suggesting that the reduction of cell viability induced by TMZ depends on TAS2R43 activation.
3.4. TMZ Induces Intracellular DOX Accumulation in Glioblastoma Cells
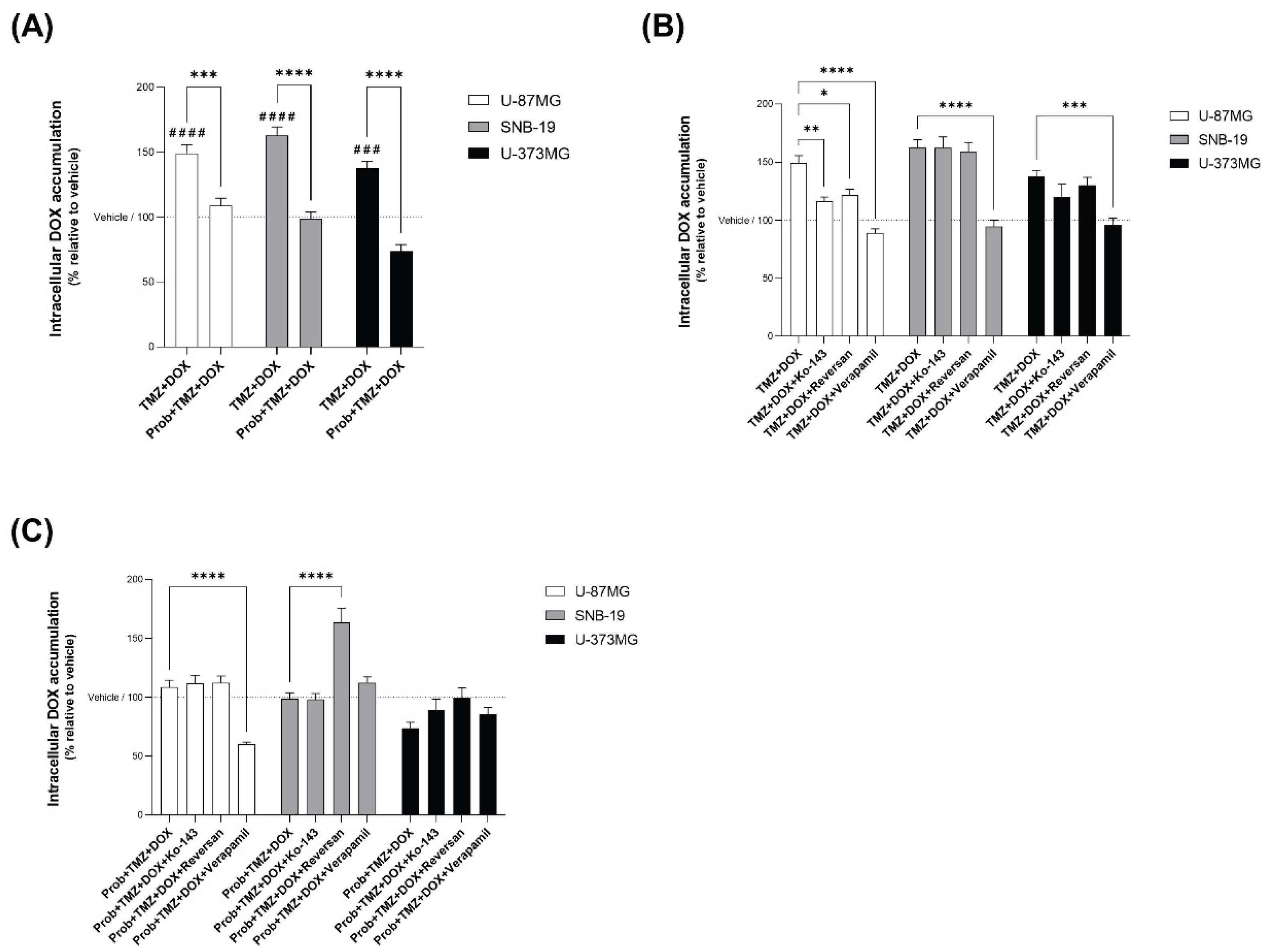
In order to evaluate the effect of TAS2R43 activation by TMZ on ABC transporter modulation, we assessed the transport of DOX, a well-known substrate of ABC transporters upon TMZ stimulation in U-87MG, SNB-19 and U-373MG glioblastoma cells. First, we showed that TMZ increased intracellular DOX accumulation by approximately 49%, 63% and 38% in U-87MG, SNB-19 and U-373MG cells, respectively, compared with vehicle, and this increase was significantly reduced in the presence of probenecid (Figure 6A).
Next, we aimed at investigating which ABC transporter influences TMZ-facilitated DOX intracellular accumulation using specific inhibitors of ABCB1, ABCC1 and ABCG2. In U-87MG cells, inhibition of ABCB1, ABCC1 and ABCG2 significantly reduced TMZ-induced DOX accumulation, with changes in the order of 20–40%, supporting the involvement of all three transporters in intracellular DOX uptake (Figure 6B). In contrast, in SNB-19 and U-373MG cells, only ABCB1 inhibition produced significant changes in TMZ-induced DOX accumulation, whereas ABCC1 inhibition caused a 30–40% change in DOX accumulation specifically in probenecid-treated SNB-19 cells (Figure 6B). Together, these findings indicate that TAS2R43 activation by TMZ promotes intracellular DOX accumulation through modulation of ABC transporters, with cell-line-specific contributions of ABCB1, ABCC1 and ABCG2.
Finally, the contribution of individual ABC transporters to TAS2R43-dependent DOX accumulation was evaluated by probenecid incubation. Overall, inhibition of ABCB1 or ABCG2 did not consistently modify DOX levels upon probenecid blockade, indicating that these transporters are not major downstream effectors in this context (Figure 6C). In contrast, in SNB-19 cells, ABCC1 inhibition led to a significant change (approx. 65%) in DOX accumulation in the presence of probenecid, suggesting that ABCC1 is a likely downstream effector of TAS2R43 in mediating TMZ-induced intracellular drug accumulation in these cells (Figure 6C).
Overall, our results suggest that TAS2R43 contributes to TMZ-induced intracellular DOX accumulation in glioblastoma cells, most likely via ABC transporter modulation.
4. Discussion
In recent years, increasing evidence has shown that several compounds, including flavonoids, alkaloids, cannabinoids, and lactones, exert anticancer effects such as anti-proliferative, pro-apoptotic, anti-angiogenic, and anti-metastatic activities in different cancers, including glioblastoma [36,37,38]. Many of these compounds are known or predicted ligands of TAS2Rs and some, such as resveratrol and epigallocatechin gallate, have been proposed as adjuvants to temozolomide (TMZ) therapy [39].
Although TMZ has predicted bitterness, its potential to bind and activate specific TAS2Rs has not been addressed before. We explored the capacity of TMZ to activate the bitter taste signalling in glioblastoma cells, with functional assays with TMZ in the presence or absence of probenecid, a known TAS2R16, R38 and R43 antagonist [34,35], and by silencing the taste signalling pathway by GNAT3 knockdown. Our findings showed that TMZ reduced the cell viability, increased cell apoptosis, and elicited intracellular Ca2+ levels in U-87MG, SNB-19 and U-373MG glioblastoma cells. Interestingly, these TMZ effects were significantly reduced in the presence of probenecid and GNAT3 knockdown, suggesting that the activation of the bitter taste signalling pathway enhances somehow the anticancer effects of TMZ. The next step was the identification of the TAS2R to which TMZ binds. Based on in-silico analysis to predict the potential target-receptors for TMZ, TAS2R expression profiling in these cells, and probenecid binding, TAS2R43 emerged as the most likely TAS2R interacting with TMZ. In fact, when we silenced TAS2R43, the effects of TMZ in the glioblastoma cells analysed were also reduced similarly to what was seen with GNAT3 silencing and probenecid incubation. Thus, the expression of TAS2R43 in these cancer cells is essential for an effective action of TMZ. Interestingly, TAS2R43 is also a target of the bitter compound epigallocatechin gallate, which in turn has been widely used as an adjuvant agent in the treatment of gliomas, especially in glioblastoma, enhancing the therapeutic efficacy of TMZ [40,41,42,43].
Because TMZ induces cytotoxicity through DNA alkylation, we hypothesised that its binding to TAS2R43 could enhance its own cellular availability, by modulating ABC-mediated efflux. Efflux transporters, particularly ATP-binding cassette (ABC) transporters, remain one of the major hurdles in cancer therapy by limiting intracellular drug accumulation and treatment efficacy. This is particularly troublesome for brain tumours since anticancer drugs, such as TMZ, have also to overcome the blood brain barrier where efflux transporters are also very active [27,44]. Previous studies indicate that inhibition or genetic disruption of ABC transporters can potentiate the effects of TMZ and other anti-tumour drugs [45,46,47]. Clinical and experimental studies further associate the overexpression of multiple ABC transporters, including ABCB1, ABCC1 and ABCG2, with multidrug resistance and reduced TMZ responsiveness, indicating that combined transporter activity influences therapeutic outcome [13,28,29,48]. In addition, we previously demonstrated that another TAS2Rs, TAS2R14, regulate efflux transport across brain barriers and modulate the disposal of bitter ligands [45]. Our intracellular DOX accumulation assays showed that TMZ increased drug retention in three glioblastoma cell lines, an effect attenuated by TAS2R43 inhibition. Transporter-specific analyses indicate that TMZ-induced accumulation of DOX involves multiple ABC transporters in U-87MG cells, while in SNB-19 cells the effect is largely ABCC1-dependent, suggesting a cell-line-specific TAS2R43–ABC regulatory axis that may reflect distinct chemoresistance phenotypes. Collectively, these results support a model in which TAS2R43 contributes to shaping intracellular drug availability in TMZ-treated glioblastoma cells through modulation of ABC transport. These observations fit within previous evidence that ABC transporters at the blood–brain barrier and in glioblastoma cells can limit TMZ brain penetration and antitumor efficacy, and that pharmacological or genetic inhibition of ABCB1 and ABCG2 enhances TMZ delivery to intracranial tumours and improves therapeutic response [46,49]. Furthermore, in glioblastoma, ABCC1 overexpression has been associated with temozolomide resistance, and its downregulation restores drug sensitivity [50].
In summary, herein we demonstrated that TAS2R43 is activated by the chemotherapeutic alkylating drug TMZ, with a relevant impact on its anti-proliferative and anti-apoptotic effects. These findings also suggest that TAS2R43 could serve as a biomarker for the efficacy of TMZ and other anticancer drugs that are ABC transporter substrates. From a translational perspective, this raises the possibility that pharmacological targeting of TAS2R43, alone or in combination with ABC modulators, could be exploited to enhance the efficacy of TMZ or other ABC substrates in glioblastoma.
Author Contributions
Ana R. Costa: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Validation, Visualization, Writing-Original draft preparation. Ana C. Duarte: Formal analysis, Investigation, Methodology, Validation, Writing-Reviewing and Editing. Isabel Gonçalves: Conceptualization, Data curation, Funding acquisition, Project administration, Resources, Supervision, Validation, Writing-Reviewing and Editing. Robert Preissner: Resources, Software. José F. Cascalheira: Resources. Helena Marcelino: Resources. Cecília R. A. Santos: Conceptualization, Data curation, Funding acquisition, Project administration, Resources, Supervision, Validation, Visualization, Writing-Reviewing and Editing. All authors have read and agreed to the published version of the manuscript.
Acknowledgments
This work was supported by the Fundação para a Ciência e Tecnologia (FCT, Portugal) project grants (PTDC/BIM-ONC/7121/2014, UID/Multi/00709/2013 and UID/Multi/00709/2019), by CENTRO 2020 and Lisboa 2020 project grant (POCI-01-0145-FEDER-016822), and FEDER funds through the POCI – COMPETE 2020 – Operational Programme Competitiveness and Internationalization in Axis I – Strengthening research, technological development and innovation (POCI-01-0145-FEDER-007491). Ana R. Costa was recipient of a PhD fellowship (UI/BD/151025/2021) funded by FCT through the Portuguese state and EU budgets through the European Social Fund. Ana C. Duarte was recipient of a grant from CENTRO 2020 program through the ICON project (Interdisciplinary Challenges On Neurodegeneration; CENTRO-01-0145-FEDER-000013). We also acknowledge the support of the Portuguese Platform of Bioimaging (PPBI) [PPBI-POCI-01-0145-FEDER-022122] and the resources provided by the Fluorescence Microscopy Unit of RISE-Health, UBI.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Avau, B.; Depoortere, I. The bitter truth about bitter taste receptors: beyond sensing bitter in the oral cavity. Acta Physiol. (Oxf.) 2016, 216, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Duarte, A.C.; Santos, J.; Costa, A.R.; Ferreira, C.L.; Tomás, J.; Quintela, T.; et al. Bitter taste receptors profiling in the human blood-cerebrospinal fluid-barrier. Biochem Pharmacol. 2020, 177. [Google Scholar] [CrossRef] [PubMed]
- Li, F. Taste perception: from the tongue to the testis. Mol. Hum. Reprod. 2013, 19, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Jaggupilli, A.; Singh, N.; Upadhyaya, J.; Sikarwar, A.S.; Arakawa, M.; Dakshinamurti, S.; et al. Analysis of the expression of human bitter taste receptors in extraoral tissues. Mol. Cell Biochem. 2017, 426, 137–147. [Google Scholar] [CrossRef]
- Dorsam, R.T.; Gutkind, J.S. G-protein-coupled receptors and cancer. Nat. Rev. Cancer 2007, 7, 79–94. [Google Scholar] [CrossRef]
- O’Hayre, M.; Degese, M.S.; Gutkind, J.S. Novel insights into G protein and G protein-coupled receptor signaling in cancer. Curr. Opin. Cell Biol. 2014, 27, 126–135. [Google Scholar] [CrossRef]
- Yang, J.; Xu, Y.; Fu, Z.; Chen, J.; Fan, W.; Wu, X. Progress in research and development of temozolomide brain-targeted preparations: a review. J. Drug Target. 2023, 31, 119–133. [Google Scholar] [CrossRef]
- Strobel, H.; Baisch, T.; Fitzel, R.; Schilberg, K.; Siegelin, M.D.; Karpel-Massler, G.; et al. Temozolomide and Other Alkylating Agents in Glioblastoma Therapy. Biomedicines 2019, 7, 69. [Google Scholar] [CrossRef]
- Kaina, B.; Christmann, M.; Naumann, S.; Roos, W.P. MGMT: Key node in the battle against genotoxicity, carcinogenicity and apoptosis induced by alkylating agents. DNA Repair (AMST) 2007, 6, 1079–1099. [Google Scholar] [CrossRef]
- Bao, S.; Wu, Q.; McLendon, R.E.; Hao, Y.; Shi, Q.; Hjelmeland, A.B.; et al. Glioma stem cells promote radioresistance by preferential activation of the DNA damage response. Nature 2006, 444, 756–760. [Google Scholar] [CrossRef]
- Chen, J.; Li, Y.; Yu, T.S.; McKay, R.M.; Burns, D.K.; Kernie, S.G.; et al. A restricted cell population propagates glioblastoma growth after chemotherapy. Nature 2012, 488, 522–526. [Google Scholar] [CrossRef] [PubMed]
- Eramo, A.; Ricci-Vitiani, L.; Zeuner, A.; Pallini, R.; Lotti, F.; Sette, G.; et al. Chemotherapy resistance of glioblastoma stem cells. Cell Death Differ. 2006, 13, 1238–1241. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Zepeda, D.; Taghi, M.; Scherrmann, J.M.; Decleves, X.; Menet, M.C. ABC Transporters at the Blood-Brain Interfaces, Their Study Models, and Drug Delivery Implications in Gliomas. Pharmaceutics 2019, 12. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.K.; Singh, P.; Ali, V.; Verma, M. Role of membrane-embedded drug efflux ABC transporters in the cancer chemotherapy. Oncol. Rev. 2020, 14, 144–151. [Google Scholar] [CrossRef]
- Hermann, D.M.; Kilic, E.; Spudich, A.; Krämer, S.D.; Wunderli-Allenspach, H.; Bassetti, C.L. Role of drug efflux carriers in the healthy and diseased brain. Ann. Neurol. 2006, 60, 489–498. [Google Scholar] [CrossRef]
- Qosa, H.; Miller, D.S.; Pasinelli, P.; Trotti, D. Regulation of ABC efflux transporters at blood-brain barrier in health and neurological disorders. Brain Res. 2015, 1628, 298–316. [Google Scholar] [CrossRef]
- Saunders, N.R.; Habgood, M.D.; Møllgård, K.; Dziegielewska, K.M. The biological significance of brain barrier mechanisms: Help or hindrance in drug delivery to the central nervous system? F1000Res 2016, 5. [Google Scholar] [CrossRef]
- Pitcher, M.R.; Quevedo, J. Tools for studying drug transport and metabolism in the brain. Expert. Opin. Drug Metab. Toxicol. 2016, 12, 161–168. [Google Scholar] [CrossRef]
- Adler, E.; Hoon, M.A.; Mueller, K.L.; Chandrashekar, J.; Ryba, N.J.P.; Zuker, C.S. A novel family of mammalian taste receptors. Cell 2000, 100, 693–702. [Google Scholar] [CrossRef]
- Chandrashekar, J.; Mueller, K.L.; Hoon, M.A.; Adler, E.; Feng, L.; Guo, W.; et al. T2Rs function as bitter taste receptors. Cell 2000, 100, 703–711. [Google Scholar] [CrossRef]
- Lang, T.; Di Pizio, A.; Risso, D.; Drayna, D.; Behrens, M. Activation Profile of TAS2R2, the 26th Human Bitter Taste Receptor. Mol Nutr Food Res. 2023, 67. [Google Scholar] [CrossRef] [PubMed]
- Diao, W.; Tong, X.; Yang, C.; Zhang, F.; Bao, C.; Chen, H.; et al. Behaviors of Glioblastoma Cells in in Vitro Microenvironments. Sci. Rep. 2019, 9, 85. [Google Scholar] [CrossRef] [PubMed]
- Memme, S.; Sukhorukov, V.L.; Höring, M.; Westerling, K.; Fiedler, V.; Katzer, A.; et al. Cell surface area and membrane folding in glioblastoma cell lines differing in PTEN and p53 status. PLoS One 2014, 9. [Google Scholar]
- Fritz, F.; Preissner, R.; Banerjee, P. VirtualTaste: a web server for the prediction of organoleptic properties of chemical compounds. Nucleic Acids Res. 2021, 49, W679–W684. [Google Scholar] [CrossRef]
- Costa, A.R.; Duarte, A.C.; Gonçalves, I.; Ferreira, C.L.; Cavaco, J.E.; Ferrer, I.; et al. Taste receptors’ profiling in glioblastoma. bioRxiv 2025, 2025.06.24.661264. [Google Scholar] [CrossRef]
- Thorn, C.F.; Oshiro, C.; Marsh, S.; Hernandez-Boussard, T.; McLeod, H.; Klein, T.E.; et al. Doxorubicin pathways: pharmacodynamics and adverse effects. Pharmacogenet Genom. 2011, 21, 440–446. [Google Scholar] [CrossRef]
- Amawi, H.; Hammad, A.M.; Hall, F.S.; Hussein, N.; Rataan, A.O.; Mrayyan, A.; et al. Revisiting strategies to target ABC transporter-mediated drug resistance in CNS cancer. Cancer Biol. Med. 2025, 22. [Google Scholar] [CrossRef]
- De Faria, G.P.; De Oliveira, J.A.; De Oliveira, J.G.P.; Romano, S.D.O.; Neto, V.M.; Maia, R.C. Differences in the expression pattern of P-glycoprotein and MRP1 in low-grade and high-grade gliomas. Cancer Invest. 2008, 26, 883–889. [Google Scholar] [CrossRef]
- Roy, L.O.; Lemelin, M.; Blanchette, M.; Poirier, M.B.; Aldakhil, S.; Fortin, D. Expression of ABCB1, ABCC1 and 3 and ABCG2 in glioblastoma and their relevance in relation to clinical survival surrogates. J. Neurooncol. 2022, 160, 601–609. [Google Scholar] [CrossRef]
- Schulz, J.A.; Hartz, A.M.S.; Bauer, B. ABCB1 and ABCG2 Regulation at the Blood-Brain Barrier: Potential New Targets to Improve Brain Drug Delivery. Pharmacol. Rev. 2023, 75, 815–853. [Google Scholar] [CrossRef]
- Engle, K.; Kumar, G. Cancer multidrug-resistance reversal by ABCB1 inhibition: A recent update. Eur. J. Med. Chem. 2022, 239. [Google Scholar] [CrossRef]
- Poku, V.O.; Iram, S.H. A critical review on modulators of Multidrug Resistance Protein 1 in cancer cells. PeerJ 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- Toyoda, Y.; Takada, T.; Suzuki, H. Inhibitors of Human ABCG2: From Technical Background to Recent Updates With Clinical Implications. Front Pharmacol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Greene, T.A.; Alarcon, S.; Thomas, A.; Berdougo, E.; Doranz, B.J.; Breslin, P.A.S.; et al. Probenecid inhibits the human bitter taste receptor TAS2R16 and suppresses bitter perception of salicin. PLoS One 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Wölfle, U.; Haarhaus, B.; Kersten, A.; Fiebich, B.; Hug, M.J.; Schempp, C.M. Salicin from Willow Bark can Modulate Neurite Outgrowth in Human Neuroblastoma SH-SY5Y Cells. Phytother. Res. 2015, 29, 1494–1500. [Google Scholar] [CrossRef]
- Anwar, S.; Malik, J.A.; Ahmed, S.; Kameshwar, V.A.; Alanazi, J.; Alamri, A.; et al. Can Natural Products Targeting EMT Serve as the Future Anticancer Therapeutics? Molecules 2022, 27. [Google Scholar] [CrossRef]
- Costa, A.R.; Duarte, A.C.; Costa-Brito, A.R.; Gonçalves, I.; Santos, C.R.A. Bitter taste signaling in cancer. Life Sci. 2023, 315. [Google Scholar] [CrossRef]
- Luís, Â.; Marcelino, H.; Rosa, C.; Domingues, F.; Pereira, L.; Cascalheira, J.F. The effects of cannabinoids on glioblastoma growth: A systematic review with meta-analysis of animal model studies. Eur. J. Pharmacol. 2020, 876. [Google Scholar] [CrossRef]
- Banerjee, P.; Preissner, R. BitterSweetForest: A Random Forest Based Binary Classifier to Predict Bitterness and Sweetness of Chemical Compounds. Front Chem. 2018, 6. [Google Scholar] [CrossRef]
- Chen, T.C.; Wang, W.; Golden, E.B.; Thomas, S.; Sivakumar, W.; Hofman, F.M.; et al. Green tea epigallocatechin gallate enhances therapeutic efficacy of temozolomide in orthotopic mouse glioblastoma models. Cancer Lett. 2011, 302, 100–108. [Google Scholar] [CrossRef]
- Grube, S.; Ewald, C.; Kögler, C.; Lawson McLean, A.; Kalff, R.; Walter, J. Achievable Central Nervous System Concentrations of the Green Tea Catechin EGCG Induce Stress in Glioblastoma Cells in Vitro. Nutr. Cancer 2018, 70, 1145–1158. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.R.; You, C.G.; Zhang, N.; Sheng, H.S.; Zheng, X.S. Epigallocatechin Gallate Preferentially Inhibits O6-Methylguanine DNA-Methyltransferase Expression in Glioblastoma Cells Rather than in Nontumor Glial Cells. Nutr. Cancer 2018, 70, 1339–1347. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, S.X.; Ma, J.W.; Li, H.Y.; Ye, J.C.; Xie, S.M.; et al. EGCG inhibits properties of glioma stem-like cells and synergizes with temozolomide through downregulation of P-glycoprotein inhibition. J. Neurooncol. 2015, 121, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Oh, J.H.; Zhang, W.; Rathi, S.; Le, J.; Talele, S.; et al. How Much is Enough? Impact of Efflux Transporters on Drug delivery Leading to Efficacy in the Treatment of Brain Tumors. Pharm. Res. 2023, 40, 2731–2746. [Google Scholar] [CrossRef]
- Duarte, A.C.; Rosado, T.; Costa, A.R.; Santos, J.; Gallardo, E.; Quintela, T.; et al. The bitter taste receptor TAS2R14 regulates resveratrol transport across the human blood-cerebrospinal fluid barrier. Biochem Pharmacol. 2020, 177. [Google Scholar] [CrossRef]
- Lin, F.; De Gooijer, M.C.; Roig, E.M.; Buil, L.C.M.; Christner, S.M.; Beumer, J.H.; et al. ABCB1, ABCG2, and PTEN determine the response of glioblastoma to temozolomide and ABT-888 therapy. Clin. Cancer Res. 2014, 20, 2703–2713. [Google Scholar] [CrossRef]
- Munoz, J.L.; Walker, N.D.; Scotto, K.W.; Rameshwar, P. Temozolomide competes for P-glycoprotein and contributes to chemoresistance in glioblastoma cells. Cancer Lett. 2015, 367, 69–75. [Google Scholar] [CrossRef]
- Wu, H.; Gao, W.; Chen, P.; Wei, Y.; Zhao, H.; Wang, F. Research progress of drug resistance mechanism of temozolomide in the treatment of glioblastoma. Heliyon 2024, 10. [Google Scholar] [CrossRef]
- de Gooijer, M.C.; de Vries, N.A.; Buckle, T.; Buil, L.C.M.; Beijnen, J.H.; Boogerd, W.; etal. Improved Brain Penetration and Antitumor Efficacy of Temozolomide by Inhibition of ABCB1 and ABCG2. Neoplasia 2018, 20, 710–720. [Google Scholar] [CrossRef]
- Li, Y.; Liu, Y.; Ren, J.; Deng, S.; Yi, G.; Guo, M.; et al. miR-1268a regulates ABCC1 expression to mediate temozolomide resistance in glioblastoma. J. Neurooncol. 2018, 138, 499–508. [Google Scholar] [CrossRef]
Figure 1.
Calcium dose-response curves of SNB-19 and U-373MG glioblastoma cells to temozolomide. Different concentrations of temozolomide (TMZ; 10-200 μM), in the presence (●) or absence (○) of 1 mM probenecid, a known TAS2R inhibitor. Dot line: Ca2+ levels measured in cells with vehicle only (DMSO ≤ 0.2%). Results are presented as mean ± SEM. Statistical analysis was performed by two-way ANOVA followed by Tukey’s multiple comparisons test. [N≥3 independent experiments; §versus vehicle; #versus probenecid+TMZ].
Figure 1.
Calcium dose-response curves of SNB-19 and U-373MG glioblastoma cells to temozolomide. Different concentrations of temozolomide (TMZ; 10-200 μM), in the presence (●) or absence (○) of 1 mM probenecid, a known TAS2R inhibitor. Dot line: Ca2+ levels measured in cells with vehicle only (DMSO ≤ 0.2%). Results are presented as mean ± SEM. Statistical analysis was performed by two-way ANOVA followed by Tukey’s multiple comparisons test. [N≥3 independent experiments; §versus vehicle; #versus probenecid+TMZ].
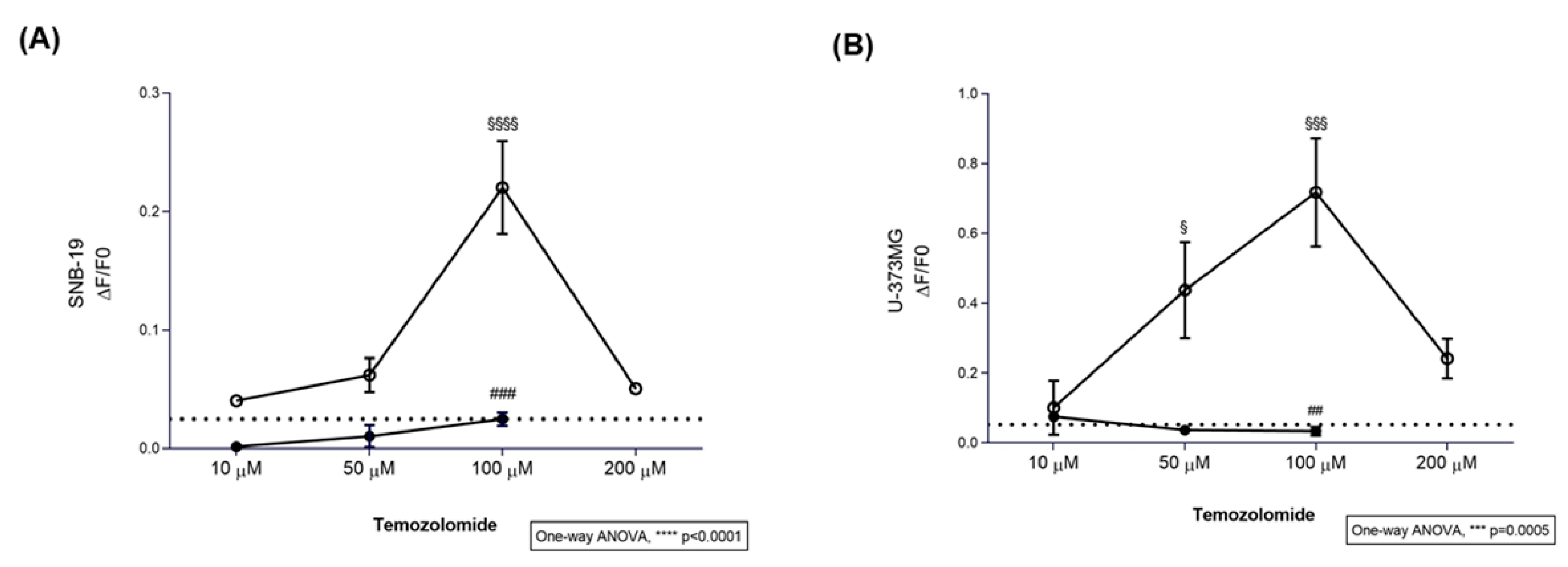
Figure 2.
Cytotoxic effect of different concentrations of temozolomide. (A) Dose-response cytotoxicity assay of glioblastoma cells exposed to different concentrations of temozolomide (TMZ; 50-500 μM) for 72 hours was assessed in U-87MG, SNB-19 and U-373MG glioblastoma cells by MTT assay. Results are presented as mean ± SEM. Statistical analysis was performed by one-way ANOVA followed by Tukey’s multiple comparisons test. [N≥3 independent experiments; **p<0.01, ***p<0.001 and ****p<0.0001]. (B) The apoptotic rate of U-87MG, SNB-19 and U-373MG glioblastoma cells incubated with 500 µM TMZ for 72h, in the presence or absence of 1 mM probenecid, was carried out by Hoechst 33342 staining (data not shown). Bar graphs represent mean ± SEM. Statistical analysis was performed by one-way ANOVA followed by Tukey’s multiple comparisons test. [N≥3 independent experiments; **p<0.01, ***p<0.001 and ****p<0.0001].
Figure 2.
Cytotoxic effect of different concentrations of temozolomide. (A) Dose-response cytotoxicity assay of glioblastoma cells exposed to different concentrations of temozolomide (TMZ; 50-500 μM) for 72 hours was assessed in U-87MG, SNB-19 and U-373MG glioblastoma cells by MTT assay. Results are presented as mean ± SEM. Statistical analysis was performed by one-way ANOVA followed by Tukey’s multiple comparisons test. [N≥3 independent experiments; **p<0.01, ***p<0.001 and ****p<0.0001]. (B) The apoptotic rate of U-87MG, SNB-19 and U-373MG glioblastoma cells incubated with 500 µM TMZ for 72h, in the presence or absence of 1 mM probenecid, was carried out by Hoechst 33342 staining (data not shown). Bar graphs represent mean ± SEM. Statistical analysis was performed by one-way ANOVA followed by Tukey’s multiple comparisons test. [N≥3 independent experiments; **p<0.01, ***p<0.001 and ****p<0.0001].
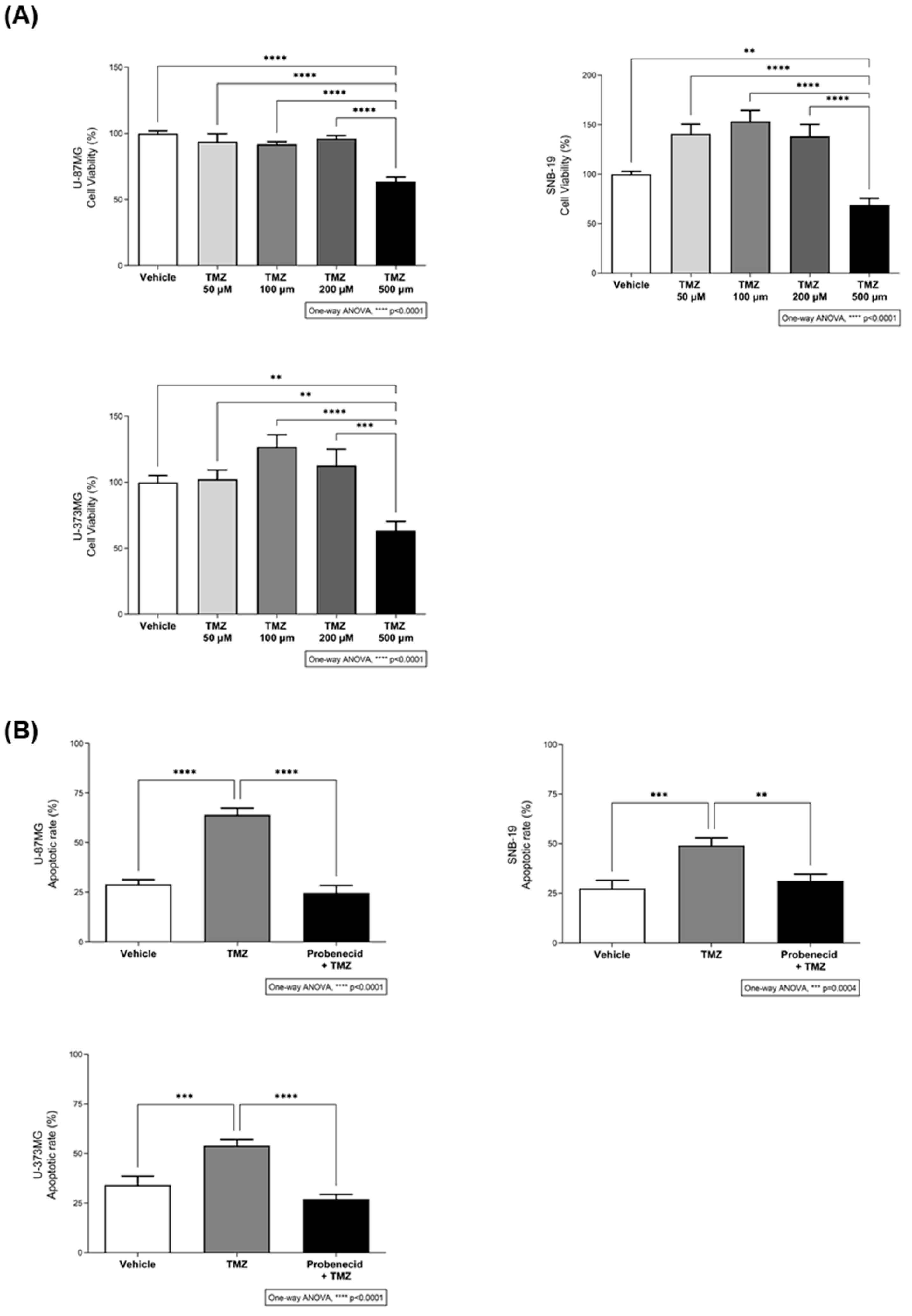
Figure 3.
The effect of temozolomide on the viability of glioblastoma cells requires the activation of the bitter taste signalling pathway. (A) Immunofluorescence analysis of α-gustducin (GNAT3) expression after siRNA transfection in SNB-19 glioblastoma cells. Protein levels of siRNA GNAT3-transfected cells are decreased in comparison with non-, mock-, and siRNA scramble-transfected cells. The quantification of GNAT3 fluorescence intensity (green) was performed in different regions of interest of confocal microscopy images obtained from three independent experiments. Nuclei were stained with Hoechst 33342 (not shown). Scale bar: 10 µm. Bar graphs represent mean ± SEM. Statistical analysis was performed by one-way ANOVA followed by Tukey’s multiple comparisons test. [N=3 independent experiments; ***p<0.001]. (B) Effects of TMZ in the viability of U-87MG, SNB-19 and U-373MG glioblastoma cells transfected or mock-transfected for 24 h with GNAT3 or a scramble siRNA, and incubated with 500 µM TMZ for 72 h, in the presence or absence of 1 mM probenecid. Bar graphs represent mean ± SEM. Statistical analysis was performed by one-way ANOVA followed by Tukey’s multiple comparisons test. [N≥3 independent experiments; *p<0.05, **p<0.01 and ****p<0.0001; §versus non-transfected; #versus non-transfected+TMZ].
Figure 3.
The effect of temozolomide on the viability of glioblastoma cells requires the activation of the bitter taste signalling pathway. (A) Immunofluorescence analysis of α-gustducin (GNAT3) expression after siRNA transfection in SNB-19 glioblastoma cells. Protein levels of siRNA GNAT3-transfected cells are decreased in comparison with non-, mock-, and siRNA scramble-transfected cells. The quantification of GNAT3 fluorescence intensity (green) was performed in different regions of interest of confocal microscopy images obtained from three independent experiments. Nuclei were stained with Hoechst 33342 (not shown). Scale bar: 10 µm. Bar graphs represent mean ± SEM. Statistical analysis was performed by one-way ANOVA followed by Tukey’s multiple comparisons test. [N=3 independent experiments; ***p<0.001]. (B) Effects of TMZ in the viability of U-87MG, SNB-19 and U-373MG glioblastoma cells transfected or mock-transfected for 24 h with GNAT3 or a scramble siRNA, and incubated with 500 µM TMZ for 72 h, in the presence or absence of 1 mM probenecid. Bar graphs represent mean ± SEM. Statistical analysis was performed by one-way ANOVA followed by Tukey’s multiple comparisons test. [N≥3 independent experiments; *p<0.05, **p<0.01 and ****p<0.0001; §versus non-transfected; #versus non-transfected+TMZ].
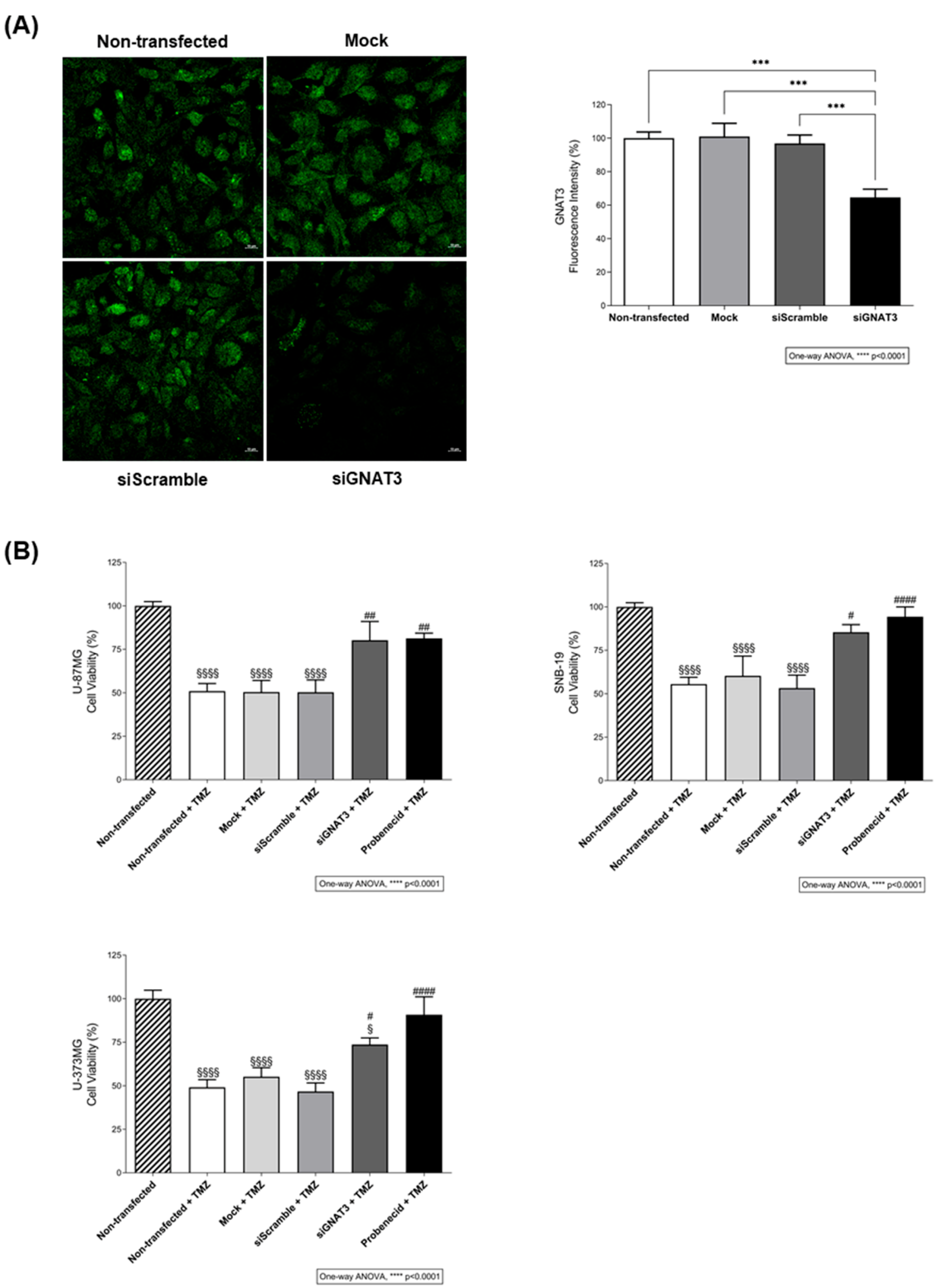
Figure 4.
TMZ target prediction. (A) TMZ is predicted to bind TAS2R38 > R10 > R43 > R45 > R44 > R14 according to the webserver VirtualTaste algorithm. Results are presented as probability. (B) mRNA expression profile of TAS2R38 and R43 in glioblastoma cell lines (U-87MG, SNB-19, U-373MG). Only TAS2R43 mRNA and protein were detected in all cell lines. The identities of the amplified products were confirmed by Sanger sequencing. MW: molecular weight (base pair); C-: negative control.
Figure 4.
TMZ target prediction. (A) TMZ is predicted to bind TAS2R38 > R10 > R43 > R45 > R44 > R14 according to the webserver VirtualTaste algorithm. Results are presented as probability. (B) mRNA expression profile of TAS2R38 and R43 in glioblastoma cell lines (U-87MG, SNB-19, U-373MG). Only TAS2R43 mRNA and protein were detected in all cell lines. The identities of the amplified products were confirmed by Sanger sequencing. MW: molecular weight (base pair); C-: negative control.
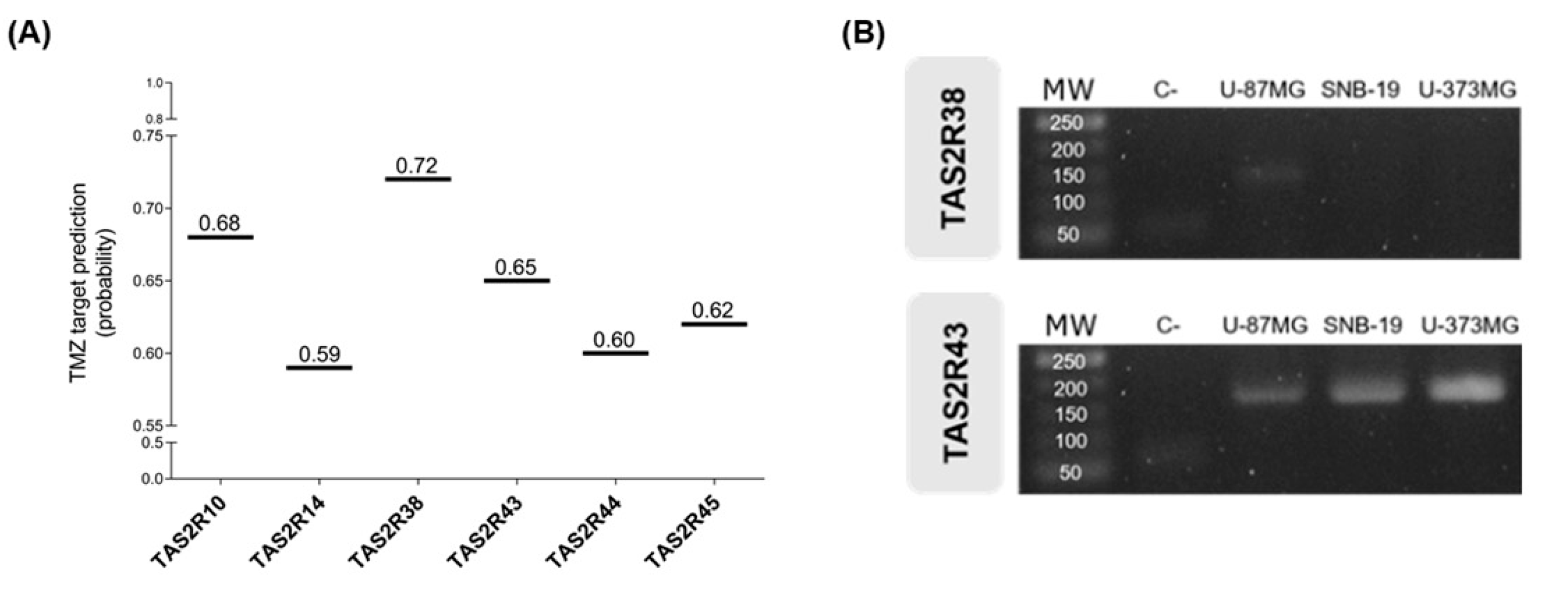
Figure 5.
The anti-proliferative effects of temozolomide in glioblastoma cells depend on the activation of TAS2R43. (A) Immunofluorescence analysis of TAS2R43 expression after siRNA transfection in SNB-19 glioblastoma cells. Protein levels of siRNA TAS2R43-transfected cells are decreased in comparison with non-, mock-, and siRNA scramble-transfected cells. The quantification of TAS2R43 fluorescence intensity (green) was performed in different regions of interest of confocal microscopy images obtained from three independent experiments. Nuclei were stained with Hoechst 33342 (not shown). Scale bar: 10 µm. Bar graphs represent mean ± SEM. Statistical analysis was performed by one-way ANOVA followed by Tukey’s multiple comparisons test. [N=3 independent experiments; **p<0.01]. (B) Effects of TMZ in the viability of U-87MG, SNB-19, and U-373MG glioblastoma cells transfected or mock-transfected for 24 h with TAS2R43 or a scramble siRNA and incubated with 500 µM TMZ for 72 h. Bar graphs represent mean ± SEM. Statistical analysis was performed by one-way ANOVA followed by Tukey’s multiple comparisons test. [N=3 independent experiments; *p<0.05, **p<0.01, ***p<0.001 and ****p<0.0001; §versus non-transfected; #versus non-transfected+TMZ].
Figure 5.
The anti-proliferative effects of temozolomide in glioblastoma cells depend on the activation of TAS2R43. (A) Immunofluorescence analysis of TAS2R43 expression after siRNA transfection in SNB-19 glioblastoma cells. Protein levels of siRNA TAS2R43-transfected cells are decreased in comparison with non-, mock-, and siRNA scramble-transfected cells. The quantification of TAS2R43 fluorescence intensity (green) was performed in different regions of interest of confocal microscopy images obtained from three independent experiments. Nuclei were stained with Hoechst 33342 (not shown). Scale bar: 10 µm. Bar graphs represent mean ± SEM. Statistical analysis was performed by one-way ANOVA followed by Tukey’s multiple comparisons test. [N=3 independent experiments; **p<0.01]. (B) Effects of TMZ in the viability of U-87MG, SNB-19, and U-373MG glioblastoma cells transfected or mock-transfected for 24 h with TAS2R43 or a scramble siRNA and incubated with 500 µM TMZ for 72 h. Bar graphs represent mean ± SEM. Statistical analysis was performed by one-way ANOVA followed by Tukey’s multiple comparisons test. [N=3 independent experiments; *p<0.05, **p<0.01, ***p<0.001 and ****p<0.0001; §versus non-transfected; #versus non-transfected+TMZ].
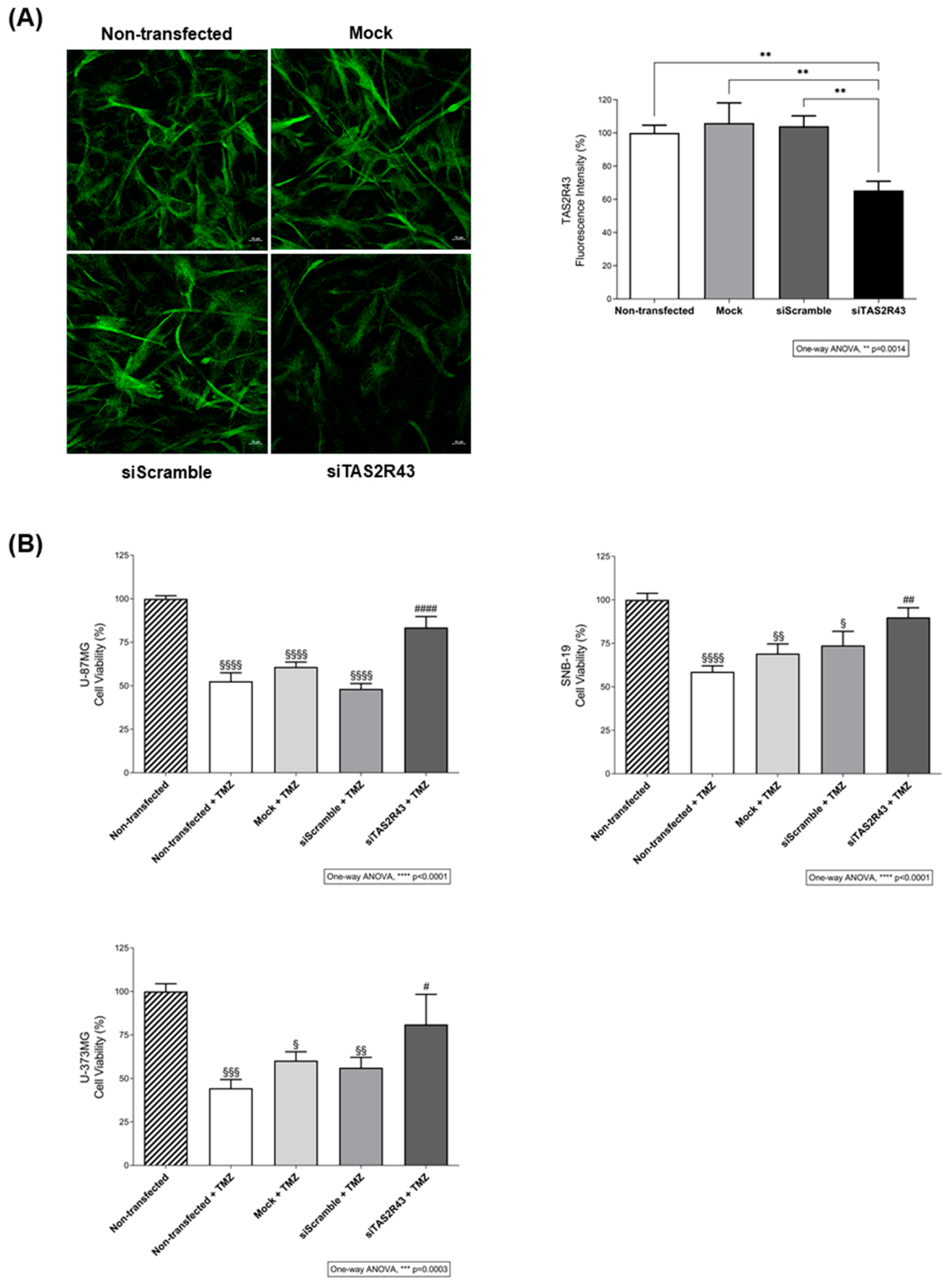
Figure 6.
TAS2R43 contributes to TMZ-induced intracellular DOX accumulation in glioblastoma cells, possibly via ABC transporter modulation. (A) Intracellular doxorubicin (DOX; 1 µM) accumulation in U-87MG, SNB-19 and U-373MG glioblastoma cells incubated with TMZ (1 µM) for 1 h, in the presence or absence of probenecid (1 mM), compared with vehicle (DMSO ≤ 1%). (B) Effects of ABCB1, ABCC1 and ABCG2 inhibition on TMZ-facilitated DOX accumulation in U-87MG, SNB-19 and U-373MG cells. Cells were pre-incubated for 1 h with verapamil (5 µM), reversan (10 µM) or Ko143 (100 nM), followed by 1 h incubation with a solution containing DOX (1 µM) and TMZ (1 µM). (C) Effects of ABCB1, ABCC1 and ABCG2 inhibition on DOX accumulation upon TAS2R43 inhibition with probenecid. Bar graphs represent mean ± SEM. Statistical analysis was performed by one-way ANOVA followed by Tukey’s multiple comparisons test. [N≥3 independent experiments; *p<0.05, **p<0.01, ***p<0.001 and ****p<0.0001; #versus vehicle].
Figure 6.
TAS2R43 contributes to TMZ-induced intracellular DOX accumulation in glioblastoma cells, possibly via ABC transporter modulation. (A) Intracellular doxorubicin (DOX; 1 µM) accumulation in U-87MG, SNB-19 and U-373MG glioblastoma cells incubated with TMZ (1 µM) for 1 h, in the presence or absence of probenecid (1 mM), compared with vehicle (DMSO ≤ 1%). (B) Effects of ABCB1, ABCC1 and ABCG2 inhibition on TMZ-facilitated DOX accumulation in U-87MG, SNB-19 and U-373MG cells. Cells were pre-incubated for 1 h with verapamil (5 µM), reversan (10 µM) or Ko143 (100 nM), followed by 1 h incubation with a solution containing DOX (1 µM) and TMZ (1 µM). (C) Effects of ABCB1, ABCC1 and ABCG2 inhibition on DOX accumulation upon TAS2R43 inhibition with probenecid. Bar graphs represent mean ± SEM. Statistical analysis was performed by one-way ANOVA followed by Tukey’s multiple comparisons test. [N≥3 independent experiments; *p<0.05, **p<0.01, ***p<0.001 and ****p<0.0001; #versus vehicle].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



