
Submitted:
18 February 2026
Posted:
27 February 2026
You are already at the latest version
Abstract
Morphological characteristics of pollen grains, such as shape, size, number of pores, and exine thickness, may vary among species, thereby enabling the reliable use of palynologi-cal data in taxonomic studies. In this context, this study aims to investigate the pollen morphology of selected seven taxa of the Betulaceae, including Alnus glutinosa, Betula pen-dula, Carpinus betulus, Carpinus orientalis, Corylus avellana, Corylus colurna and Ostrya carpinifolia. Detailed morphological measurements were carried out using Light Micros-copy (LM), and high-resolution images were obtained using Scanning Electron Microsco-py (SEM). Thirty measurements were taken under the LM for each morphological charac-ter, including polar axis length (P), equatorial diameter (E), pore length (plg), pore width (plt), and exine thickness (Ex). Interspecific differences were evaluated using one-way ANOVA, Tukey’s HSD test, and Principal Component Analysis (PCA), and a pollen iden-tification key for the investigated species was developed in this study. The first principal component (PC1) was primarily associated with pollen size (P and E), along which Carpinus betulus was clearly separated from the remaining species. The second principal component (PC2) was mainly related to pore length (plg) and particularly contributed to the separation of Alnus glutinosa from the other small-pollen species.
Keywords:
pollen morphology
; Betulaceae
; Carpinus betulus
; Alnus glutinosa
; PCA
1. Introduction
Pollen morphology is a sub-discipline of palynology that encompasses the examination of morphological characteristics of pollen grains, including their shape, size, exine structure, aperture type, and surface ornamentation. These features provide essential information for the systematic classification of plants, the clarification of taxonomic relationships, and the interpretation of evolutionary processes. Morphological differences observed among taxa enable palynological data to be used reliably in taxonomic studies [1,2,3]. Moreover, pollen morphology plays a crucial role in plant systematics by revealing phylogenetic relationships among taxa and in paleopalynological studies by facilitating the reconstruction of past plant assemblages and their spatial distributions [1,4]. In this context, the detailed investigation of pollen morphology in selected taxa of the Betulaceae constitutes one of the main objectives of the present study.
Betulaceae is represented worldwide by two subfamilies, seven genera, and approximately 205 plant species. Betula and Alnus belong to the subfamily Betuloideae, whereas Carpinus, Corylus, and Ostrya are classified within the subfamily Coryloideae [5]. Among the Betuloideae taxa examined for pollen morphology in this study, Alnus glutinosa (L.) Gaertn. has a broad distribution range extending across northwestern Africa, Europe, the Caucasus, Anatolia, and northern Iran. In Türkiye, A. glutinosa is known to occur throughout the entire Northern Anatolia region, as well as in Southern and Eastern Anatolia, where it shows regional and local distributions. The second Betuloideae species investigated, Betula pendula Roth., is widely distributed across Europe, Asia, Finland, Sweden, Norway, and Central Europe, while in Türkiye it occurs naturally mainly in Northeastern and Eastern Anatolia [5].
Among the Coryloideae taxa studied, Carpinus betulus L. has a general distribution covering Europe, Türkiye, the Caucasus, and Iran. Within Türkiye, the species is widely distributed throughout the Black Sea Region and occurs in limited and localized populations in the Amanos Mountains. Another species examined for pollen morphology, Carpinus orientalis Mill., is naturally distributed across the Balkans, Dalmatia, Italy, Sicily, Türkiye, Iran, and the Caucasus. In Türkiye, it is predominantly found in coastal areas, particularly on dry, calcareous south-facing slopes, and mainly at low elevations [5]. Among the palynologically investigated species, Corylus colurna L. is distributed in Southeastern Europe and Western Asia, while in Türkiye it occurs in regions such as the Kazdağları Mountains, Bolu, Kastamonu, Ankara–Nallıhan, Zonguldak–Yenice, and the surroundings of Rize and Trabzon [5,6]. Corylus avellana L. is distributed throughout Europe, North Africa, the Caucasus, and Western Asia, and in Türkiye it occurs both naturally and in cultivation throughout the entire Black Sea mountain range, from areas close to sea level up to elevations of approximately 1800 m [5]. With regard to the pollen morphologically studied species Ostrya carpinifolia Scop., its general distribution is centered around the Mediterranean region, while in Türkiye it is known to occur in both northern and southern parts of Anatolia [5,7].
The first study on pollen morphology of the Betulaceae was conducted by Wodehouse in 1935 [8]. This study demonstrated that pollen sizes of Betulaceae taxa range between 20 and 40 µm and that the number of pores varies from 3 to 7. In his study on the pollen morphology of the Betulaceae, Erdtman [1] examined approximately 40 species belonging to six genera. He reported that the number of pores in this family ranges from 3 to 7 and that the pollen grains exhibit an aspidote structure. He also noted that the equatorial diameters of the pollen grains range between 20 and 30 µm. Furthermore, Erdtman [1] described the presence of arc-like structures extending from pore to pore in the pollen grains of taxa belonging to the genus Alnus. Kuprianova [9], in her study on Betulaceae pollen, divided the taxa within the family into three subfamilies. According to this classification, Alnus and Betula were placed in the subfamily Betuleae, Corylus in Coryleae, and Carpinus and Ostrya in Carpineae. Praglowski [10] reported that Betulaceae pollen grains are isopolar and aspidote, with pollen sizes ranging from 15 to 40 µm, in a pollen morphological study of certain trees and shrubs in Sweden. Aytuğ et al. [11], in his investigation of pollen from various plant species in Istanbul, examined five species belonging to the Betulaceae, measuring pollen type, aperture number, polar axis, equatorial diameter and, exine thickness, and provided a concise description of their pollen morphology. Blackmore et al. [12] conducted a detailed study on the pollen morphology of Betulaceae taxa and developed a comprehensive pollen identification key for the investigated species. Similarly, Alan [13], in his study on pollen morphology of Betulaceae taxa, reported that pollen shape varies from suboblate to spheroidal and also constructed a pollen identification key for the studied species.
The pollen grains of Alnus glutinosa are generally suboblate to oblate in shape, with 3–7 pores and a psilate–granulate ornamentation, and variability in pore number has been emphasized in numerous studies. Previous investigations on pollen morphology have reported considerable variation in pollen size in A. glutinosa, with the polar axis length ranging from 16 to 29.5 µm and the equatorial diameter varying between 23 and 31 µm [10,11,12,13,14,15,16,17,18]. Studies focusing on exine thickness in this species have generally demonstrated that exine thickness ranges between 0.75 and 2.0 µm [11,12,13,16].
In contrast, pollen grains of Betula are predominantly triporate and can be distinguished from those of Corylus and Carpinus by their suboblate to spheroidal shape and well-developed poral vestibula. The surface ornamentation is described as psilate–granulate [12]. According to values reported in different studies, the polar axis length of Betula pendula pollen ranges from 16 to 35.51 µm, while the equatorial diameter varies between 18.5 and 34 µm [10,12,13,19,20,21,22,23,24,25]. Previous studies have also shown that the exine thickness of this species ranges from 0.8 to 2.52 µm [12,13,24,25].
Pollen grains of Carpinus are generally larger in size, possess three to five pores, and show morphological similarities to Betula; however, they differ in terms of the degree of poral protrusion and exine characteristics [8]. The pollen shape is suboblate to oblate, and the ornamentation is psilate–granulate. Previous studies have reported that the polar axis length of Carpinus betulus ranges between 26 and 40 µm, while the equatorial diameter varies from 26.34 to 46 µm [8,10,11,12,13,14,25,26,27]. The exine thickness of C. betulus has been reported to range between 0.7 and 1.53 µm [11,12,13,25,27].
Studies focusing on Carpinus orientalis are comparatively limited. Kedves [26] reported the pollen diameter of C. orientalis to range between 34 and 40 µm. In a subsequent study, Alan [13] reported mean values for C. orientalis of 26.6 µm for the polar axis, 28.4 µm for the equatorial diameter, and 0.87 µm for exine thickness. Akhondnezhad et al. [27], in their study on C. orientalis subsp. orientalis, reported that the polar axis length ranged from 29.23 to 37.22 µm, the equatorial diameter from 26.79 to 28.84 µm, and the exine thickness was 1.75 µm. Based on measurements conducted on C. betulus and C. orientalis, Alan [13] concluded that C. betulus exhibits larger pollen dimensions. In contrast, Praglowski [10] suggested that Carpinus orientalis displays morphological appearance and size characteristics similar to those of C. betulus.
Pollen grains of Corylus are predominantly triporate and are generally oblate to suboblate in shape, being distinguished by a characteristic triangular outline in polar view [12]. The exine ornamentation exhibits a psilate–granulate pattern. Previous palynological studies have reported that the polar axis length of Corylus avellana pollen ranges from 18 to 28 µm, while the equatorial diameter varies between 22 and 34 µm [8,10,11,12,13,14,25,26,28]. The exine thickness of this species has been reported to range between 1.09 and 1.5 µm [11,12,13,25].
Corylus colurna has been addressed in relatively fewer studies. Kedves [26] reported pollen diameters ranging from 18 to 26 µm, whereas Alan [13] recorded measurement values between 26.85 and 28.8 µm. Alan [13] further reported an exine thickness of 1.1 µm for this species.
Pollen grains of Ostrya are generally spheroidal to oblate–suboblate in shape, possess a psilate–granulate exine ornamentation, and exhibit a 3–4-porate aperture structure [12]. The ambitus of Ostrya carpinifolia is more rounded compared to other members of the Betulaceae family [10]. Studies on the pollen morphology of O. carpinifolia have reported that the polar axis length ranges between 20 and 31 µm, while the equatorial diameter varies from 19 to 28 µm [10,11,12,13,14,26]. The exine thickness of this species has been reported to range between 0.89 and 1.5 µm [11,12,13].
Although numerous studies have been conducted on pollen morphology within the Betulaceae, investigations in which the obtained measurements are tested using statistical analyses remain limited. In this context, detailed morphological measurements of pollen grains from Alnus glutinosa, Betula pendula, Carpinus betulus, Carpinus orientalis, Corylus avellana, Corylus colurna, and Ostrya carpinifolia were carried out using light microscopy, and high-resolution images were additionally obtained using scanning electron microscopy (SEM). To reveal interspecific differences, statistical evaluations were performed using one-way ANOVA (ANalysis Of Variance), Tukey’s HSD test (Tukey’s Honestly Significant Difference) and Principal Component Analysis (PCA). Furthermore, a diagnostic key highlighting species-specific differences in pollen morphology was developed.
2. Results
2.1. Palinological Results
The pollen grains of Alnus glutinosa possess a five-porate (stephanoporate) apertur type (Figure 1, Figure 2). The length of the polar axis (P) ranges from 17.53 to 24.71 µm, while the equatorial diameter (E) varies between 20.96 and 29.50 µm. The P/E ratio is 0.88, indicating a suboblate pollen shape according to the classification of Erdtman [1]. The pore length (plg) ranges from 3.05 to 4.22 µm, and the pore width (plt) from 1.27 to 2.85 µm. The exine thickness varies between 1.03 and 1.65 µm (Table 1).
The pollen grains of Betula pendula are characterized by a triporate aperture structure (Figure 1, Figure 2). The polar axis length (P) ranges from 20.64 to 24.82 µm, while the equatorial diameter (E) varies between 23.27 and 26.96 µm. The P/E ratio is 0.89, and according to Erdtman [1], the pollen shape is oblate-spheroidal. The pore length (plg) ranges from 2.58 to 3.51 µm, and the pore width (plt) from 2.03 to 3.24 µm. The exine thickness was determined to range between 0.63 and 1.16 µm (Table 1).
In Carpinus betulus, 40% of the pollen grains display a four-porate (tetraporate) aperture type, while the remaining 60% are characterized by a five-porate (stephanoporate) aperture structure (Figure 1, Figure 2). The polar axis length (P) ranges from 28.52 to 33.72 µm, while the equatorial diameter (E) varies between 34.42 and 39.67 µm. The P/E ratio is 0.86, indicating a suboblate pollen shape according to the classification of Erdtman [1]. The pore length (plg) ranges from 2.86 to 4.33 µm, and the pore width (plt) from 2.32 to 3.72 µm. The exine thickness varies between 0.84 and 1.38 µm (Table 1).
The pollen grains of Carpinus orientalis are triporate in aperture type (Figure 1, Figure 2). The polar axis length (P) ranges from 25.53 to 29.40 µm, and the equatorial diameter (E) from 30.00 to 33.91 µm. The P/E ratio is 0.88, and according to Erdtman [1], the pollen shape is classified as suboblate. The pore length (plg) ranges from 2.07 to 2.98 µm, while the pore width (plt) varies between 1.57 and 2.36 µm. The exine thickness was measured to range from 0.72 to 1.18 µm (Table 1).
The pollen grains of Corylus avellana are characterized by a triporate aperture type (Figure 1, Figure 2). The polar axis length (P) ranges from 20.71 to 23.70 µm, while the equatorial diameter (E) varies between 25.26 and 30.90 µm. The P/E ratio is 0.83, indicating a suboblate pollen shape according to the classification of Erdtman [1]. The pore length (plg) ranges from 2.08 to 2.95 µm, and the pore width (plt) from 1.82 to 2.71 µm. The exine thickness varies between 1.06 and 1.53 µm (Table 1).
The pollen grains of Corylus colurna also exhibit a triporate aperture structure (Figure 1, Figure 2). The polar axis length (P) ranges from 26.01 to 29.33 µm, while the equatorial diameter (E) varies between 27.80 and 32.46 µm. The P/E ratio is 0.93 and based on Erdtman’s [1] classification, the pollen shape is oblate-spheroidal. The pore length (plg) ranges from 2.36 to 3.76 µm, and the pore width (plt) from 2.11 to 3.25 µm. The exine thickness was determined to range between 1.09 and 1.92 µm (Table 1).
The pollen grains of Ostrya carpinifolia are triporate in aperture type (Figure 1, Figure 2). The polar axis length (P) ranges from 21.09 to 23.92 µm, whereas the equatorial diameter (E) varies between 22.71 and 26.91 µm. The P/E ratio is 0.91, indicating an oblate-spheroidal pollen shape according to Erdtman [1]. The pore length (plg) ranges from 2.26 to 3.49 µm, and the pore width (plt) from 2.01 to 2.96 µm. The exine thickness was measured to range between 0.89 and 1.52 µm (Table 1).
2.2. Statistical Results
The measured pollen morphological characters (P, E, plg, plt, and Ex) of seven species belonging to the Betulaceae were statistically evaluated in this study. Descriptive statistics (mean and standard deviation) were calculated for each species and each variable in order to assess intraspecific variation (Table 2).
The results of the one-way ANOVA revealed highly significant differences among species for all measured variables (Table 3; p < 0.001). These findings indicate that pollen morphology possesses a strong discriminatory power at the species level.
Pollen size, expressed as polar axis length (P) and equatorial diameter (E), emerged as the most prominent morphological variables explaining interspecific differences. According to the Tukey HSD test, Carpinus betulus exhibited significantly higher values for both pollen length and width compared to all other species, forming a distinct group within the Betulaceae family. Carpinus orientalis and Corylus colurna clustered together with intermediate pollen dimensions, whereas Alnus glutinosa, Betula pendula, Corylus avellana, and Ostrya carpinifolia were classified as small-pollen species. No significant differences were detected in most pairwise comparisons among these four species. These results indicate that P and E are key discriminative variables for distinguishing Betulaceae species.
Analyses of pore length (plg) revealed significant differences among species (Table 3;p < 0.001). The highest plg values were recorded in Alnus glutinosa and Carpinus betulus, distinguishing these taxa from the remaining species based on pore morphology. Betula pendula and Ostrya carpinifolia exhibited intermediate pore lengths, whereas Carpinus orientalis, Corylus avellana, and Corylus colurna showed significantly lower values.
With respect to pore width (plt), the highest mean values were recorded in Carpinus betulus. Betula pendula, Corylus colurna and Ostrya carpinifolia exhibited pores of intermediate width, whereas Alnus glutinosa, Carpinus orientalis and Corylus avellana were characterized by relatively narrower pore structures. Exine thickness showed marked variation among species. Corylus colurna, and C. avellana exhibited the thickest exine within the family and formed a distinct morphological group. Alnus glutinosa and Ostrya carpinifolia were characterized by intermediate exine thickness values, while Betula pendula, Carpinus orientalis, and Carpinus betulus possessed the thinnest exine.
PCA, conducted to evaluate the combined effect of the five morphological variables, demonstrated a clear separation of species within the multivariate morphological space. The first two principal components accounted for approximately 65% of the total variance. The first principal component (PC1) was primarily associated with pollen size (P and E), along which Carpinus betulus was clearly separated from the remaining species. The second principal component (PC2) was mainly related to pore length (plg) and particularly contributed to the separation of Alnus glutinosa from the other small-pollen species (Figure 3).
In the PCA ordination, Corylus species were positioned close to each other in relation to exine thickness, whereas the small-pollen species (Alnus glutinosa, Betula pendula, Corylus avellana, and Ostrya carpinifolia) generally clustered at low PC1 values (Figure 3).
3. Discussion
All investigated taxa share similar general ornamentation patterns, appearing psilate under LM and microechinate under SEM. Consequently, species-level discrimination does not rely on exine sculpture type but primarily on pore number and quantitative morphometric parameters. The first and most decisive diagnostic criterion is pore number. Alnus glutinosa is clearly distinguished by its predominantly 5-porate pollen grains, whereas Carpinus betulus differs from all other taxa by exhibiting 4–5-porate pollen in combination with markedly larger pollen dimensions (P ≈ 31 µm; E ≈ 36 µm). The remaining taxa possess strictly 3-porate pollen grains. Within the 3-porate group, differentiation is achieved through polar axis length and exine thickness. Corylus colurna is characterized by relatively large pollen grains (P ≈ 28 µm) and the thickest exine (≈ 1.54 µm) among the studied taxa. Carpinus orientalis shows slightly smaller pollen grains (P ≈ 27 µm) and shorter pore length (≈ 2.59 µm). Species with smaller pollen grains (P < 27 µm) are further distinguished by exine thickness. Betula pendula is identified by its notably thin exine (≈ 0.86 µm), whereas Corylus avellana (≈ 1.32 µm) and Ostrya carpinifolia (≈ 1.21 µm) possess thicker exines; these latter two taxa are additionally separated by equatorial diameter values, with C. avellana exhibiting slightly larger E values (≈ 27 µm) than O. carpinifolia (≈ 25 µm).
Pollen ornamentation of all studied taxa is psilate under LM and microechinate under SEM:
| 1a. Pollen grains 5-porate ................................................................……………………….Alnus glutinosa 1b. Pollen grains 3-porate or 4–5-porate ..................................................................................................... 2 2a. Pollen grains 4–5-porate; P ≈ 31 µm; E ≈ 36 µm ......................................................... Carpinus betulus 2b. Pollen grains strictly 3-porate ………………………………………………………………................. 3 3a. Polar axis (P) > 27 µm …………………………………………………………………............................ 4 3b. Polar axis (P) < 27 µm ................................................................................................................................ 5 4a. P ≈ 28 µm; exine thickness ≈ 1.54 µm ………………………………………………….. Corylus colurna 4b. P ≈ 27 µm; pore length ≈ 2.59 µm ……………………………………………........... Carpinus orientalis 5a. Exine thickness < 1.0 µm ……………………………………………………..................... Betula pendula 5b. Exine thickness ≥ 1.0 µm ……………………………………………..................................................... 6 6a. Exine thickness ≈ 1.32 µm; E ≈ 27 µm ……………………………………………….... Corylus avellana 6b. Exine thickness ≈ 1.21 µm; E ≈ 25 µm ........................................................................ Ostrya carpinifolia |
This hierarchically structured diagnostic key is not merely descriptive but statistically supported. The present study establishes a quantitatively validated and conceptually structured framework for interpreting pollen morphological differentiation within the Betulaceae. Although previous palynological investigations [1,8,10,12] provided detailed descriptive accounts of pollen size ranges, aperture configurations, and ornamentation patterns, they largely relied on qualitative comparisons. In contrast, the integration of precise morphometric measurements with rigorous univariate (ANOVA, Tukey HSD) and multivariate (PCA) statistical analyses demonstrate that interspecific variation is hierarchically organized and statistically robust.
Pollen size (polar axis and equatorial diameter) emerged as the primary axis of morphological differentiation. The exceptionally high ANOVA F-values for P (274.74; p < 0.001) and E (283.16; p < 0.001) indicate that interspecific variance substantially exceeds intraspecific variability, quantitatively confirming earlier qualitative observations that pollen size represents a key taxonomic character within the family [1,12]. The distinct separation of Carpinus betulus as a large-pollen taxon aligns with previous reports [10,13], while Alnus glutinosa, Betula pendula, Corylus avellana, and Ostrya carpinifolia formed a small-pollen group. Carpinus orientalis and Corylus colurna occupied intermediate morphometric positions. This structured size-based organization reflects genus-level differentiation patterns between Betuloideae and Coryloideae [9,12]. PCA independently confirmed this hierarchy, with pollen size dominating PC1 and accounting for the largest proportion of total variance.
Aperture morphology contributed to secondary differentiation among taxa. The pre-dominance of stephanoporate pollen in Alnus glutinosa confirms its generic distinctiveness [1], whereas the triporate configuration observed in Coryloideae taxa supports established generic boundaries [12]. Significant interspecific differences in pore length (plg; F = 42.02, p < 0.001) and pore width (plt; F = 45.59, p < 0.001) demonstrate that aperture dimensions enhance taxonomic resolution. PCA further indicated that plg contributed primarily to PC2, suggesting that aperture traits function as fine-scale discriminators rather than primary structuring variables.
Exine thickness also showed significant interspecific variation (F = 67.91, p < 0.001). Although not sufficient alone for complete species separation, its integration with pollen size and aperture traits enhances diagnostic reliability and supports a multi-character evaluation approach.
Crucially, the concordance between ANOVA, Tukey HSD, and PCA demonstrates that pollen morphology in the Betulaceae is not only descriptively distinct but statistically robust. By transforming classical descriptive palynology into a quantitatively testable and hierarchically structured morphometric framework, this study introduces a reproducible model for species-level discrimination. This statistically supported diagnostic hierarchy reduces interpretative subjectivity—historically complicated by morphological overlap among closely related taxa—and strengthens the application of pollen morphology in systematic taxonomy and Quaternary paleoenvironmental reconstruction. Furthermore, the proposed framework offers a transferable analytical template for other wind-pollinated woody plant families.
4. Materials and Methods
Pollen grains of Alnus glutinosa, Betula pendula, Carpinus betulus, Carpinus orientalis, Corylus avellana, Corylus colurna, and Ostrya carpinifolia, which constitute the research material, were collected from the Atatürk Arboretum by selecting mature male inflorescences during the pollination period (Figure 4).
The collected specimens were transferred to the Palynology Laboratory of the Faculty of Forestry, Istanbul University–Cerrahpaşa, where pollen slides were prepared using the method described by Wodehouse [8]. Measurements and microphotographs of pollen morphological characters were obtained using a Leica DM750 light microscope. The terminology proposed by Punt et al. [2], Hesse et al. [29], Beug [30], and Karlıoğlu Kılıç et al. [3] was used for the identification of pollen grains. SEM observations were carried out at the SEM Laboratory of the Faculty of Science, Istanbul University. Prior to SEM imaging, pollen grains were sputter-coated with platinum for 45 seconds using an EmiTech SC7620 sputter coater. The coated specimens were examined and photographed with a JEOL NeoScope JCM-5000 benchtop SEM. Thirty measurements were taken under the light microscope for each morphological character, including Polar axis length (P), Equatorial diameter (E), pore length (plg), pore width (plt), and exine thickness (Ex).
All statistical analyses were performed to identify differences in pollen morphology among species. First, descriptive statistics (mean and standard deviation) were calculated for each species and each variable to assess intraspecific variation. One-way ANOVA was applied separately to each morphological variable to compare mean values among species. Tukey’s HSD post hoc test was conducted to determine pairwise differences among species for variables showing statistically significant differences in the ANOVA results. PCA was performed to evaluate the multivariate morphological structure and to reveal the combined contribution of all measured variables to species discrimination. All statistical analyses were carried out using the Python programming language [31]. Descriptive statistics were calculated using the pandas package [32] while one-way ANOVA and Tukey HSD tests were performed using the SciPy [33] and statsmodels [34] packages. PCA was conducted using the scikit-learn library [35]. Figures were generated using matplotlib [36].
5. Conclusions
This study demonstrates that pollen morphological differentiation within the Betulaceae is hierarchically structured and statistically robust rather than merely descriptive. By integrating detailed LM–SEM observations with ANOVA, Tukey HSD and PCA, we identify pollen size as the dominant axis of interspecific variation, with aperture traits and exine thickness providing secondary and complementary resolution. The concordance between univariate and multivariate analyses confirms that interspecific variance significantly exceeds intraspecific variability, establishing a reproducible morphometric basis for species-level discrimination. Beyond validating classical palynological descriptions, this work introduces a quantitatively supported diagnostic hierarchy that reduces subjectivity and enhances taxonomic precision. The proposed framework offers a transferable model for other wind-pollinated woody taxa and strengthens the application of pollen morphology in both systematic botany and Quaternary paleoenvironmental research.
Author Contributions
Conceptualization, H.C., R.Y.D. and N.K.K.; methodology, N.K.K., R.Y.D. and H.C.; investigation, H.C., R.Y.D. and N.K.K.; resources, H.C., R.Y.D., and N.K.K.; data curation, R.Y.D, N.K.K.; writing—original draft preparation, H.C., N.K.K., R.Y.D. and G.G.; writing—review and editing, H.C., N.K.K, R.Y.D. and G.G.; project administration, H.C., R.Y.D and N.K.K.; funding acquisition, H.C. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Scientific Research Projects Coordination Unit of Istanbul University (Project No. FBA-2020-36100).
Data Availability Statement
All data are contained in the article.
Acknowledgments
The authors would like to thank the Scientific Research Projects Coordination Unit of Istanbul University for their support.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| µm | Micrometre |
| SEM | Scanning Electron Microscopy |
| ANOVA | ANalysis Of VAriance |
| Tukey’s HSD | Tukey’s Honestly Significant Difference |
| PCA | Principal Component Analysis |
| P | Polar axis length |
| E | Equatorial diameter |
| plg | Pore length |
| plt | Pore width |
| Ex | Exine thickness |
| LM | Light Microscopy |
References
- Erdtman, G. Pollen Morphology and Plant Taxonomy; Almqvist & Wiksell: Angiosperms, 1952. [Google Scholar]
- Punt, W.; Hoen, P.P.; Blackmore, S.; Nilsson, S.; Le Thomas, A. Glossary of pollen and spore terminology. Rev. Palaeobot. Palynol. 2007, 143, 1–81. [Google Scholar] [CrossRef]
- Karlıoğlu Kılıç, N.; Yılmaz Dağdeviren, R.; Caner, H.; Akkemik, Ü. Türkiye’de kullanılmakta olan palinoloji ve polen terimleri üzerine bir değerlendirme ve öneriler. Avrasya Terim Dergisi 2020, 8, 98–108. [Google Scholar] [CrossRef]
- Moore, P. D.; Webb, J. A. An Illustrated Guide to Pollen Analysis; Hodder and Stoughton: London, UK, 1978. [Google Scholar]
- Türkiye’nin Bütün Ağaçları ve Çalıları; Akkemik, Ü., Ed.; Türkiye İş Bankası Kültür Yayınları: Türkiye, 2023. [Google Scholar]
- Yaltırık, F.; Efe, A. Dendroloji Ders Kitabı; Çantay Kitabevi: İstanbul, Türkiye, 2000. [Google Scholar]
- Kayacık, H. Orman ve Park Ağaçlarının Özel Sistematiği: Angiospermae (Kapalı Tohumlular); İstanbul Üniversitesi Orman Fakültesi Yayınları: İstanbul, Türkiye, 1975. [Google Scholar]
- Wodehouse, R. P. Pollen Grains; McGraw-Hill Book Company: New York, 1935. [Google Scholar]
- Kuprianova, L. A. On a hitherto undescribed family belonging to the Amentiferae. Taxon 1963, 12, 12–13. [Google Scholar] [CrossRef]
- Praglowski, J. R. Notes on the pollen morphology of Swedish trees and shrubs. Grana Palynologica 1962, 3, 45–76. [Google Scholar] [CrossRef]
- Aytuğ, B. İstanbul Çevresi Bitkilerinin Polen Atlası; İstanbul Üniversitesi Orman Fakültesi Yayınları: İstanbul, Türkiye, 1971. [Google Scholar]
- Blackmore, S.; Steinmann, J.A.J.; Hoen, P. P.; Punt, W. Betulaceae and Corylaceae. Rev. Palaeobot. Palynol. 2003, 123, 71–98. [Google Scholar] [CrossRef]
- Alan, Ş. Türkiye Corylaceae ve Betulaceae Familyaları Polenlerinin Biyokimyasal ve Morfolojik Özellikleri. In Doktora Tezi; Ankara Üniversitesi: Ankara, 2010. [Google Scholar]
- Erdtman, G. An Introduction to Pollen Analysis; The Cnronica Botanica Company: Leyden, The Netherlands, 1943. [Google Scholar]
- Leopold, E.B.; Birkebak, J.; Reinink-Smith, L.; Jayachandar, A.P.; Narváez, P.; Zaborac-Reed, S. Pollen morphology of the three subgenera of Alnus. Palynology 2012, 36, 131–151. [Google Scholar] [CrossRef]
- Shayanmehr, F.; Jalali, S.H.; Hosseinzadeh Colagar, A.; Yousefzadeh, H.; Zare, H. Pollen morphology of the genus Alnus Mill. In Hyrcanian Forests, North of Iran. Appl. Environ. Res. 2014, 13, 819–831. [Google Scholar] [CrossRef]
- Kizilpinar Temizer, İ.; Turkmen, Z. Pollen morphology of A. glutinosa (L.) Gaertner subsp. glutinosa (Betulaceae). Kastamonu Üniversitesi Orman Fakültesi Dergisi 2016, 16, 423–426. [Google Scholar] [CrossRef]
- Sahli, A.; Kassout, J.; Boselli, V.A.; Ennouni, H.; Chakkour, S.; Kadaoui, K.; Houssni, M.; Ater, M. Pollen variability of Alnus glutinosa (L.) Gaerth. (Betulaceae) from southern range edge populations in Northern Morocco. Int. J. Plant Biol. 2023, 14, 797–810. [Google Scholar] [CrossRef]
- Kedves, M.; Pardutz, A. Scanning electron microscopy of some selected recent Amentiflorae pollen II. Acta Biol. Szeged. 1983, 29, 67–76. [Google Scholar]
- Mäkelä, E. M. Size distinctions between Betula pollen types-a review. Grana 1996, 35, 248–256. [Google Scholar] [CrossRef]
- Krizo, M.; Slobodnik, B. Changes in the size of pollen grains and structure of the tapetal layer in Betula pendula (Roth) during the maturation of pollen. Bull. Polish Acad. Sci. Biol. Sci 1997, 45, 2–4. [Google Scholar]
- Tetiana, S.; Kateryna, G.; Jan, B.; Radovan, O. Morphological characteristics and antioxidant activity of pollen silver birch (Betula pendula Ehrh.). Pharmacogn. Commun. 2014, 4, 25–34. [Google Scholar] [CrossRef]
- Bıçakçı, A.; Saatçioğlu, G.; Tosunoğlu, A. Allerjenik Betula (huş ağacı) polenlerinin Türkiye’deki dağılımları. Astım Allerji İmmünoloji 2017, 15, 142–153. [Google Scholar] [CrossRef]
- Payamnoor, V.; Sattarian, A.; Kordalivand, A.; Mohammadi, J. Pollen morphology and trichome types of Betula spp. in the Hyrcanian forests of northern Iran. Journal of Forestry Research 2019, 30, 1191–1198. [Google Scholar] [CrossRef]
- Acar Şahin, A.; Pınar, N.M. Pollen morphology of selected allergenıc specıes at beşevler 10. year campus, Ankara Unıversıty, Turkey. Communications Faculty of Sciences University of Ankara Series C Biology 2020, 29, 318–337. [Google Scholar]
- Kedves, M. Scanning electron microscopy of some selected recent Amentiflorae pollens I. Acta Bot. Acad. Sci. Hung. 1979, 25, 75–82. [Google Scholar]
- Akhondnezhad, S.; Nejadsattari, T.; Sattarian, A.; Asri, Y.; Bagheriieh, N.M. Pollen morphology of the genus Carpinus L. (Corylaceae) in Iran. Iran. J. Bot. 2011, 17, 233–237. [Google Scholar] [CrossRef]
- Nikolaieva, N.; Brindza, J.; Garkava, K.; Ostrovsky, R. Pollen features of hazelnut (Corylus avellana L.) from different habitats. Modern Phytomorphology 2014, 6, 53–58. [Google Scholar] [CrossRef]
- Hesse, M.; Halbritter, H.; Zetter, R.; Weber, M.; Buchner, R.; Frosch-Radivo, A.; Ulrich, S. Pollen Terminology an Illustrated Handbook, 1st ed.; Springer: Vienna, Austria, 2009. [Google Scholar]
- Beug, H.J. Leitfaden der Pollenbestimmung für Mitteleuropa und angrenzende Gebiete; Verlag Dr. Friedrich Pfeil: München, 2015. [Google Scholar]
- Python Software Foundation. Python (Version 3.10). 2023. Available online: https://www.python.org.
- McKinney, W. Data structures for statistical computing in Python. SciPy 2010, 445, 51–56. [Google Scholar] [CrossRef]
- Virtanen, P.; Gommers, R.; Oliphant, T. E.; Haberland, M.; Reddy, T.; Cournapeau, D.; Van Mulbregt, P. SciPy 1.0: fundamental algorithms for scientific computing in Python. Nature methods 2020, 17, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Seabold, S.; Perktold, J. Statsmodels: econometric and statistical modeling with python. SciPy 2010, 7, 92–96. [Google Scholar] [CrossRef]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Duchesnay, É. Scikit-learn: Machine learning in Python. Journal of Machine Learning Research 2011, 12, 2825–2830. Available online: https://inria.hal.science/hal-00650905v2.
- Hunter, J. D. Matplotlib: A 2D graphics environment. Computing in Science & Engineering 2007, 9, 90–95. [Google Scholar] [CrossRef]
Figure 1.
Polar and equatorial views of pollen grains under LM. 1-2; Polar view of Alnus glutinosa pollen grains, 3; equatorial view of Alnus glutinosa pollen grains, 4-5; polar view of Betula pendula pollen grains, 6; equatorial view of Betula pendula pollen grains, 7-8; polar view of Carpinus betulus pollen grains, 9; equatorial view of Carpinus betulus pollen grains, 10-11; polar view of Carpinus orientalis pollen grains, 12; equatorial view of Carpinus orientalis pollen grains, 13-14; polar view of Corylus avellana pollen grains, 15; equatorial view of Corylus avellana pollen grains, 16-17; polar view of Corylus colurna pollen grains, 18; equatorial view of Corylus colurna pollen grains, 19-20; polar view of Ostrya carpinifolia pollen grains, 21; equatorial view of Ostrya carpinifolia pollen grains.
Figure 1.
Polar and equatorial views of pollen grains under LM. 1-2; Polar view of Alnus glutinosa pollen grains, 3; equatorial view of Alnus glutinosa pollen grains, 4-5; polar view of Betula pendula pollen grains, 6; equatorial view of Betula pendula pollen grains, 7-8; polar view of Carpinus betulus pollen grains, 9; equatorial view of Carpinus betulus pollen grains, 10-11; polar view of Carpinus orientalis pollen grains, 12; equatorial view of Carpinus orientalis pollen grains, 13-14; polar view of Corylus avellana pollen grains, 15; equatorial view of Corylus avellana pollen grains, 16-17; polar view of Corylus colurna pollen grains, 18; equatorial view of Corylus colurna pollen grains, 19-20; polar view of Ostrya carpinifolia pollen grains, 21; equatorial view of Ostrya carpinifolia pollen grains.
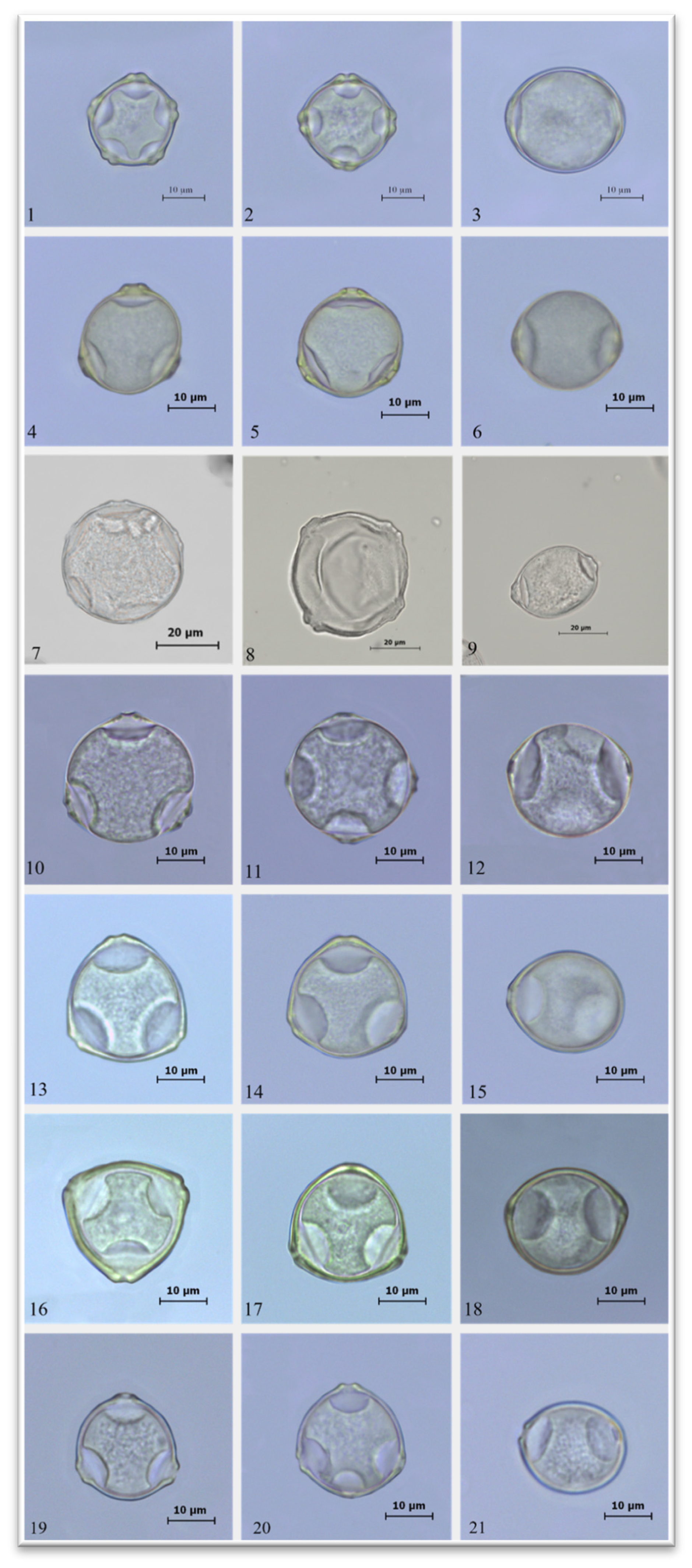
Figure 2.
Polar and equatorial views of pollen grains under SEM. 1-2; Polar view of Alnus glutinosa pollen grains, 3; equatorial view of Alnus glutinosa pollen grains, 4-5; polar view of Betula pendula pollen grains, 6; equatorial view of Betula pendula pollen grains, 7-8; polar view of Carpinus betulus pollen grains, 9; equatorial view of Carpinus betulus pollen grains, 10-11; polar view of Carpinus orientalis pollen grains, 12; equatorial view of Carpinus orientalis pollen grains, 13-14; polar view of Corylus avellana pollen grains, 15; equatorial view of Corylus avellana pollen grains, 16-17; polar view of Corylus colurna pollen grains, 18; equatorial view of Corylus colurna pollen grains, 19-20; polar view of Ostrya carpinifolia pollen grains, 21; equatorial view of Ostrya carpinifolia pollen grains.
Figure 2.
Polar and equatorial views of pollen grains under SEM. 1-2; Polar view of Alnus glutinosa pollen grains, 3; equatorial view of Alnus glutinosa pollen grains, 4-5; polar view of Betula pendula pollen grains, 6; equatorial view of Betula pendula pollen grains, 7-8; polar view of Carpinus betulus pollen grains, 9; equatorial view of Carpinus betulus pollen grains, 10-11; polar view of Carpinus orientalis pollen grains, 12; equatorial view of Carpinus orientalis pollen grains, 13-14; polar view of Corylus avellana pollen grains, 15; equatorial view of Corylus avellana pollen grains, 16-17; polar view of Corylus colurna pollen grains, 18; equatorial view of Corylus colurna pollen grains, 19-20; polar view of Ostrya carpinifolia pollen grains, 21; equatorial view of Ostrya carpinifolia pollen grains.
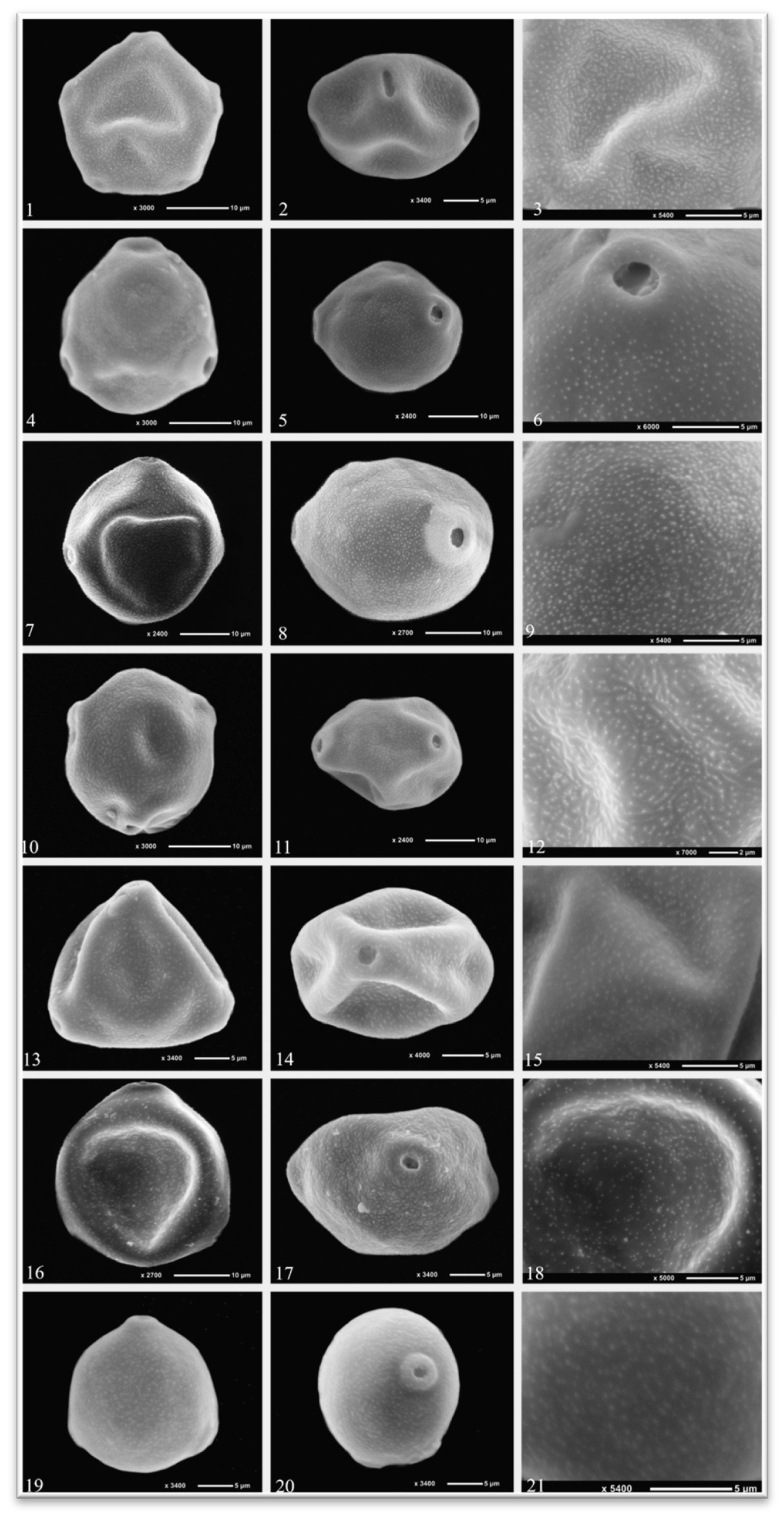
Figure 3.
The results indicate that pollen morphological characters within the Betulaceae exhibit clear and statistically significant differences among species. When pollen size, pore characteristics, and exine thickness are evaluated together, the species can be reliably distinguished based on their pollen morphology.
Figure 3.
The results indicate that pollen morphological characters within the Betulaceae exhibit clear and statistically significant differences among species. When pollen size, pore characteristics, and exine thickness are evaluated together, the species can be reliably distinguished based on their pollen morphology.
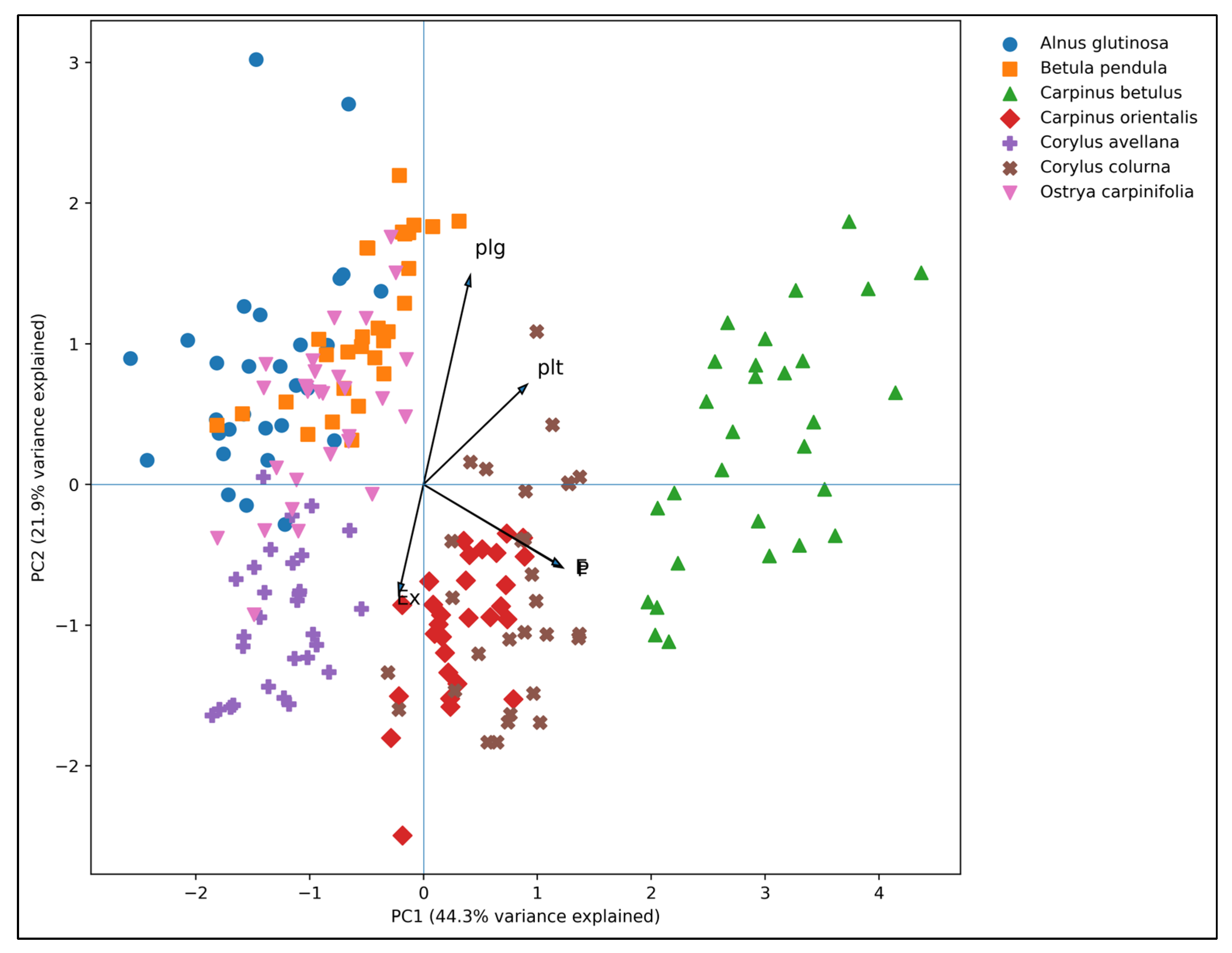
Figure 4.
Location map of the study area.

Table 1.
Measurement of pollen morphological characters.
| Species | P (µm) |
E (µm) |
P/E | Plg (µm) | Plt (µm) | Ex (µm) | Number of pores | Ornamentation | |
|---|---|---|---|---|---|---|---|---|---|
| LM | SEM | ||||||||
|
Alnus glutinosa |
17.53-24.71 | 20.96-29.50 | 0.88 | 3.05-4.22 | 1.27-2.85 | 1.03-1.65 | 5 (Stephanoporate) |
Psilate | Microechinate |
|
Betula pendula |
20.64-24.82 | 23.27-26.96 | 0.89 | 2.58-3.51 | 2.03-3.24 | 0.63-1.16 | 3 (Triporate) |
Psilate | Microechinate |
| Carpinus betulus | 28.52-33.72 | 34.42-39.67 | 0.86 | 2.86-4.33 | 2.32-3.72 | 0.84-1.38 | 4-5 (Tetraporate, Stephanoporate) |
Psilate | Microechinate |
| Carpinus orientalis | 25.53-29.40 | 30-33.91 | 0.88 | 2.07-2.98 | 1.57-2.36 | 0.72-1.18 | 3 (Triporate) |
Psilate | Microechinate |
| Corylus avellana | 20.71-23.70 | 25.26-30.90 | 0.83 | 2.08-2.95 | 1.82-2.71 | 1.06-1.53 | 3 (Triporate) |
Psilate | Microechinate |
|
Corylus colurna |
26.01-29.33 | 27.8-32.46 | 0.93 | 2.36-3.76 | 2.11-3.25 | 1.09-1.92 | 3 (Triporate) |
Psilate | Microechinate |
| Ostrya carpinifolia | 21.09-23.92 | 22.71-26.91 | 0.91 | 2.26-3.49 | 2.01-2.96 | 0.89-1.52 | 3 (Triporate) |
Psilate | Microechinate |
Table 2.
Means and standard deviations of the species.
| Species | P mean |
sd |
E mean |
sd |
plg mean |
sd |
plt mean |
sd |
Ex mean |
sd |
|---|---|---|---|---|---|---|---|---|---|---|
| Alnus glutinosa | 21.77 | 1.92 | 24.71 | 2.13 | 3.50 | 0.36 | 2.06 | 0.41 | 1.27 | 0.15 |
| Betula pendula | 22.64 | 1.07 | 25.34 | 0.83 | 3.06 | 0.26 | 2.76 | 0.30 | 0.86 | 0.15 |
| Carpinus betulus | 31.35 | 1.63 | 36.54 | 1.55 | 3.47 | 0.47 | 3.17 | 0.43 | 1.04 | 0.13 |
| Carpinus orientalis | 27.62 | 0.99 | 31.55 | 1.00 | 2.59 | 0.24 | 2.10 | 0.22 | 0.94 | 0.12 |
| Corylus avellana | 22.63 | 0.80 | 27.03 | 1.46 | 2.48 | 0.24 | 2.21 | 0.26 | 1.32 | 0.15 |
| Corylus colurna | 28.16 | 1.00 | 30.35 | 1.29 | 2.99 | 0.36 | 2.76 | 0.34 | 1.54 | 0.21 |
| Ostrya carpinifolia | 22.62 | 0.78 | 25.00 | 1.37 | 3.05 | 0.30 | 2.56 | 0.32 | 1.21 | 0.17 |
Table 3.
ANOVA results.
| Variable | ||
|---|---|---|
| P | 274.74 | p < 0.001 |
| E | 283.16 | p < 0.001 |
| plg | 42.02 | p < 0.001 |
| plt | 45.59 | p < 0.001 |
| Ex | 67.91 | p < 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



