Submitted:
23 February 2026
Posted:
24 February 2026
You are already at the latest version
Abstract
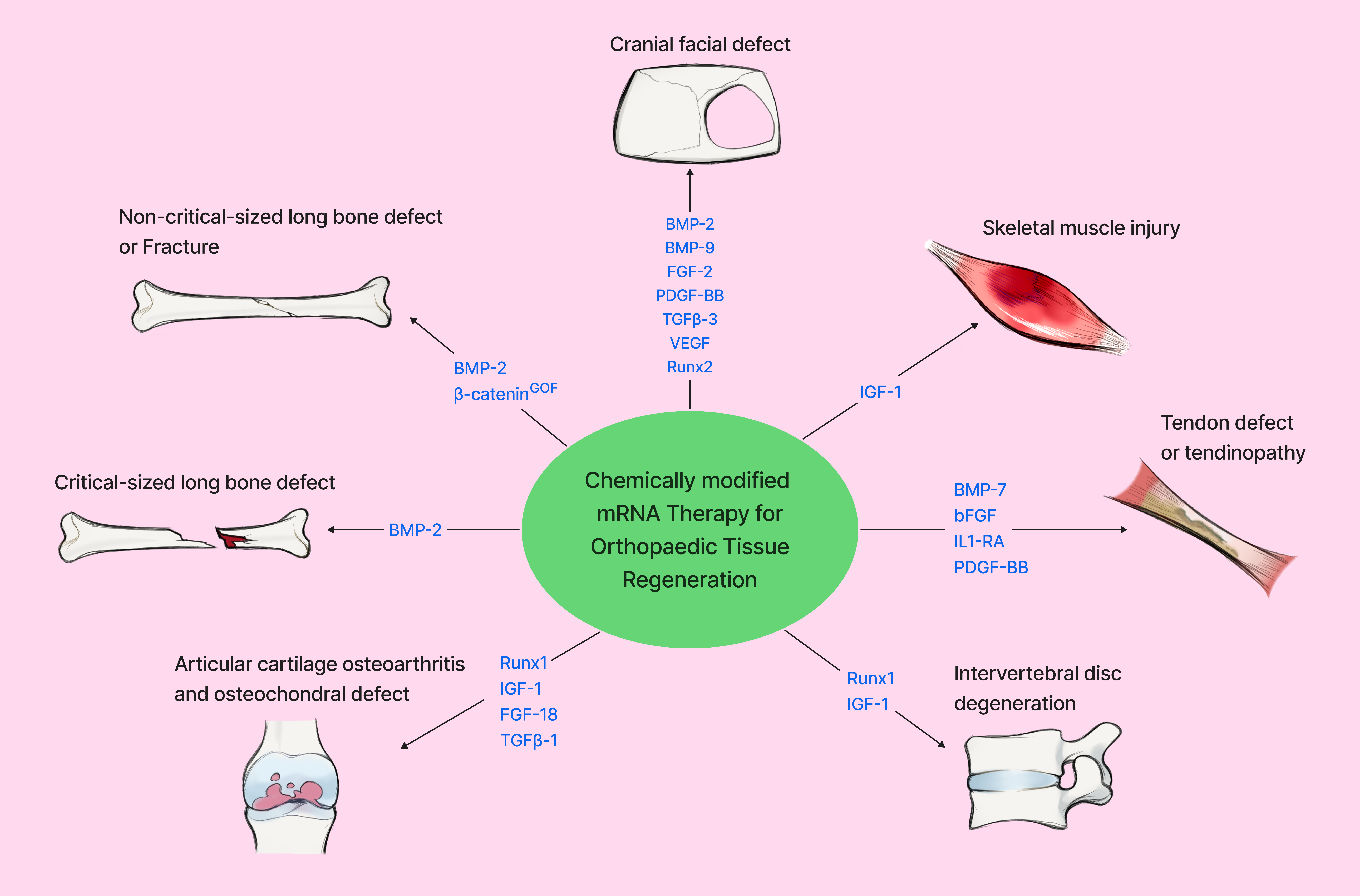
Musculoskeletal disorders and injuries are highly prevalent and encompass a broad range of conditions, including bone fractures and segmental defects, tendinopathies and tendon injury, and cartilage disorders such as osteoarthritis, cartilage defects, and intervertebral disc disease. These conditions can arise from diverse causes including trauma and injury, tumor resection, congenital abnormalities, and age-related degeneration. In the past decades, administration of chemically modified mRNA (cmRNA) encoding growth factors and transcriptional regulators has demonstrated effectiveness in repairing musculoskeletal tissues in preclinical studies. This review summarizes recent advancements in bone, tendon, cartilage, intervertebral disc, and muscle regeneration achieved through the localized delivery of protein-encoding mRNAs to express therapeutic target proteins. Delivery of cmRNA encoding growth factors such as BMP-2, BMP-9, VEGF, FGF-18, and IGF-1, or transcriptional regulators including Runx1 to various animal models has shown beneficial effects on bone, tendon, cartilage, and muscle injury repair in preclinical models. Alongside these progresses, the advantages and disadvantages of applying chemically modified mRNA for musculoskeletal tissue regeneration are also discussed. While studies show the promise of cmRNA for therapeutic applications in orthopaedic tissue regeneration, more research is required to optimize growth factors and delivery methods, as well as validate long-term safety and efficacy prior to successful translation into new therapies to benefit patients.
Keywords:
chemically modified protein encoding mRNA
; bone
; cartilage
; tendon
; skeletal muscle
; intervertebral disc
; biomaterials
1. Introduction
Disorders of the musculoskeletal system are a major public health burden and a leading cause of disability, affecting an estimated 27.7% of the population worldwide in 2014 [1]. Regeneration of musculoskeletal tissues including bone, tendon, cartilage, intervertebral disc, and skeletal muscle remains a significant clinical challenge, particularly in terms of large segmental defects, chronic degeneration, or impaired healing. Current treatment strategies such as autografts, allografts, recombinant growth factors, gene therapy, and surgical reconstruction often carry limitations such as donor-site morbidity, graft supply, variable efficacy, or off-target effects [2]. These challenges have driven increasing interest in messenger RNA (mRNA)-based therapeutics, which offer localized expression of growth factors without the integration risks associated with retroviral, lentiviral mediated gene therapy or the supraphysiological dose required for recombinant protein therapy.
The application of growth factors using gene therapy, ex vivo cell therapy, and biomaterials has proven effective for musculoskeletal tissue regeneration or repair [3,4]; however, its clinical utility has been limited by the challenges associated with recombinant protein delivery and gene therapy approaches. Currently, recombinant human bone morphogenic proteins-2 (rhBMP2)(Infusion Graft) and -7 (rhBMP7) have been approved by the FDA for therapeutic applications in bone regeneration, but have since been limited due to adverse side effects impeding clinical applications [5]. The supraphysiological dose required to accommodate for a short in-vivo half-life increases the cost of treatment and exacerbates adverse effects [6,7]. In attempts to address these limitations, gene therapy approaches employing viral and non-viral vectors for tissue regeneration have demonstrated efficacy, but are constrained by safety concerns of viral vectors and low transfection efficiencies of non-viral vectors [8,9,10,11].
Following the success of COVID-19 mRNA vaccine in controlling the COVID-19 pandemic, therapeutic mRNA has gained significant momentum as a novel strategy for therapeutic target delivery. Numerous preclinical studies have explored the feasibility of protein encoding chemically modified mRNA for musculoskeletal tissue repair. A previous review article summarized RNA therapies for musculoskeletal conditions and focused primarily on mRNA delivery vehicles [12]. This review article focuses on the applications of protein-encoding mRNA therapy for regenerating bone, tendon, cartilage, intervertebral disc and muscle.
2. Mrna for Bone Regeneration
2.1. Mrna-Based Therapeutic Strategies for Craniomaxillofacial Bone Regeneration
Large osseous defects of the craniomaxillofacial region, often caused by trauma, infection, tumor resection, or skeletal abnormalities are difficult to regenerate and remain a significant clinical challenge as they can lead to significant physical and psychological impacts [13,14]. Current clinical treatment methods include but are not limited to allografts, autografts, graft-substitutes, or the application of recombinant growth factors such as BMPs [14]. Limitations to each of these techniques necessitate novel treatment options. mRNA therapy for bone repair has the potential to overcome limitations of existing treatment strategies by enabling the localized expression of bone morphogenic proteins (BMPs) and/or other various angiogenic and osteogenic gene targets.
2.1.1. Bmp-Encoding Mrna Delivery for Calvarial Bone Regeneration
Recombinant forms of bone morphogenic proteins, particularly BMP-2 and BMP-9, have been widely investigated for their applications in bone regeneration [15,16,17,18] . Therefore, chemically modified mRNA encoding BMPs represented some of the earliest mRNA-based attempts for bone regeneration. Among the BMP family, BMP-2 is the most effective BMP [17] and has been widely investigated in mRNA-based bone regeneration. Elangovan S et al. first investigated the effects of chemically modified ribonucleic acid (cmRNA) encoding BMP-2 delivered using polyethylenimine (PEI) for inducing osteogenic differentiation of human bone marrow mesenchymal stem cells (hBMMSCs) in vitro, followed by an in vivo evaluation of cmRNA-based therapy for the repair of a rat calvarial bone defect model in comparison with conventional plasmid DNA gene therapy. The polyplex were injected into the collagen scaffold and implanted into a 5mm diameter calvarial bone defect in rats. In vitro results demonstrated a two-fold increase in BMP-2 protein expression by transfected hBMMSCs compared to PEI-pDNA at the same dose. In vivo, μCT results revealed partial defect healing and increased bone volume to trabecular volume (BV/TV) in defects treated with PEI-cmRNA encoding BMP-2 (25µg BMP-2 cmRNA) polyplexes compared to 25µg BMP-2 pDNA polyplexes and significantly increased healing compared to empty defects. Histology analysis further validate extensive bone bridging with more mature mineralized bone tissue in the BMP-2 PEI-cmRNA group, while the pDNA group only showed minimal formation of new bone tissue in the defect margin [19].
Surisaeng T et al. demonstrated that BMP-2 cmRNA/LNP loaded on either silk fibroin (SF) or gelatin silk fibroin (G) scaffolds showed an increased quantity of mineralized bone matrix within the defect, increased percentages of calvarial defect closure, and increased BV/TV compared to empty defects. Using the SF scaffold, only the 15µg dose of BMP-2 mRNA showed a statistically significant increase in BV/TV, while using the G scaffold all BMP-2 mRNA doses (1.5, 5, and 15µg) significantly increased BV/TV. However, only partial defect closure was achieved at 15µg BMP-2 mRNA dosage using either scaffold (57 or 58% closure). This study further demonstrated that implantation of BMP-2 cmRNA transfected rat bone marrow mesenchymal stem cells (rBMSCs) or rat fibroblasts resulted in near complete defect healing compared to partial healing of the delivery of BMP-2 mRNA alone [13]. These results demonstrated that the presence of cells that continuously express BMP-2 is very important. Collectively, these studies demonstrated the efficacy of cmRNA encoding BMP-2 in promoting calvarial bone regeneration and its advantages over conventional plasmid DNA therapy.
In addition to BMP-2 mRNA, others have also investigated BMP-9 mRNA for calvarial bone defect repair. Khorsand B et al. compared the regenerative capacity of nanoplexes of polyethylenimine (PEI)-delivered BMP-9 cmRNA (10µg) and BMP-9 pDNA (10µg) incorporated into perforated collagen membranes (PCMs) in a critical sized calvarial defect model in rats. PEI-cmRNA encoding BMP-9 loaded on PCMs significantly increased mineralized bone and BV/TV relative to the scaffold only group, and a trended increase in BV/TV was observed compared to PEI-pDNA-PCM treatment group [20]. The same research team compared cmRNA encoding BMP-2 and BMP-9 complexed with PEI at 50µg dose and injected the PEI-cmRNA into collagen scaffolds and freeze dried for subsequent use for in vivo bone regeneration. It was found that both BMP-2 and BMP-9 cmRNAs regenerated significantly more bone compared to an empty defect (control) with complete defect closing. Although there was no difference in callus formation or BV/TV between BMP-9 and BMP-2 cmRNA groups, the connectivity density was twofold greater and more bone tissue was discovered in the BMP-9 cmRNA group compared to the BMP-2 cmRNA group [21]. These findings suggest that BMP-9 may be a more effective growth factor than BMP-2 for bone regeneration, when delivered in cmRNA form.
2.1.2. Cmrna Encoding Other Angiogenic and Osteogenic Genes for Craniofacial Bone Repair
While BMPs have been the most extensively investigated in terms of bone repair, other growth factors including fibroblast growth factor-2 (FGF-2), platelet-derived growth factor-BB (PDGF-BB), transforming growth factor beta-3 (TGFβ-3), runt-related transcription factor 2 (Runx2), and vascular endothelial growth factor (VEGF) are also potential gene targets for chemically modified mRNA therapy.
Co-Delivery of BMP mRNA with Additional Growth Factor mRNAs for Craniofacial Bone Defect Repair
Geng Y et al. investigated the synergistic effects of VEGF and BMP-2 cmRNA for bone repair. In vitro assay demonstrated VEGF and BMP-2 secretion by cmRNA lasted for 48 hours. Furthermore, BMP-2 mRNA transfected rat bone marrow stem cells (BMSCs) (2µg mRNA transfected 2×105 cells) demonstrated increased osteogenic potential in vitro and enhanced critical sized defect repair in rat calvarial bones when using a collagen scaffold. VEGF and BMP-2 cmRNA transfected rBMSCs synergistically enhanced bone defect repair compared to BMP-2 mRNA transfected BMSCs by simultaneously promoting osteogenesis and angiogenesis, resulting in improved bone healing and almost complete defect closure. Additionally, the regenerated bone with BMP2 and VEGF cmRNAs showed higher gene expression of osteogenic related genes and angiogenesis related genes (CD31) as demonstrated by immunohistochemistry and western blot [22].
Similarly, Tsou HK et al. explored the effects of co-application of 10µg BMP-2 and TGFβ-3 cmRNA (unknown amount) -loaded polyplex (PEG-Pasp (DET) block copolymer) nanomicelles on a gelatin sponge scaffold on calvarial defect healing in 8 weeks old male mice. At two weeks post administration, only a small bone island had formed in the BMP-2+TGFβ-3 mRNA group, while there was no bone formation in the BMP-2 mRNA group. The BMP-2 and TGFβ-3 mRNA combined group regenerated over 10-fold new bone volume (BV) at 8 weeks compared to the BMP-2 mRNA nanomedicine administration alone. However, the defects were only partially healed with bone volume/defect volume ratios around 12% in the combined group. [23] These results indicated a potential synergistic relationship between TGFβ-3 and BMP-2 in enhancing endochondral bone repair, likely due to the role of TGFβ-3 in promoting chondrogenesis.
Surisaeng T et al. evaluated the effect of BMP-2 mRNA applied alone or in combination with FGF-2 and PDGF-BB for calvarial bone defect repair. Although BMP-2 cmRNA significantly enhanced bone regeneration in the calvarial defects, neither FGF-2 nor PDGF-BB cmRNA alone promoted bone repair. Surprisingly, co-delivery of PDGF-BB cmRNA with BMP2 cmRNA suppressed the bone regenerative effects of BMP-2 cmRNA [13]. In contrast, both PDGF-BB and FGF-2 in their recombinant protein forms have previously been shown in enhancing bone regeneration [24,25]. Collectively, these findings indicate that co-delivery of selective growth factors such as VEGF or TGFβ-3 can further enhance bone formation whereas other factors such as FGF-2 or PDGF-BB may be ineffective or inhibitory.
cmRNA Encoding RUNX2 and VEGF for Craniofacial Bone Defect Repair
In addition to the co-delivery of mRNA encoding growth factors with BMP-2, the combination of other growth factors such as Runx2 and VEGF has also been investigated for promoting osteogenic differentiation and craniofacial bone defect repair. Zhang M et al. injected polyethylene glycol (PEG) and polyamino acid (Poly(N-[N′-(2-aminoethyl)-2-aminoethyl]aspartamide)) (PEG-PAsp(DET)) nanomicelles loaded with Runx2 (10µg) and VEGF mRNAs (10µg) to a mandibular defect model at 7 days after the creation of a 4mm mandible defect. Results revealed that while each mRNA individually significantly enhanced bone regeneration, their combined administration produced a synergistic effect, further promoting bone formation compared with either RUNX2 or VEGF mRNA alone, as evidenced by increased BV and BV/TV in µCT and histology analyses. However, complete defect healing was not achieved, even in the combined RUNX2 and VEGFA mRNA treatment groups [26].
2.2. Mrna Therapy for the Repair of Long Bone Defects
2.2.1. Cmrna for Repair of Non-Critical Sized Long Bone Defects or Fractures
The physiological mechanism of fracture healing is a complex, innate, regenerative process. However, this process is susceptible to failure and can potentially result in unfavorable results for the patient including delayed or non-union fractures[27]. Non-union is estimated to occur in 2% of all fractures, with instances of non-union rising to as high as 20% for certain long bone shaft fracture injuries [28]. Several mRNA therapy studies on femoral or tibial fractures in animal models have aimed to address this clinical need.
Balamayor E et al. first reported that BMP-2 cmRNA expressing stem cells demonstrated enhanced osteogenic differentiation in vitro. BMP-2 cmRNA (2.5µg) loaded on Fibrin + C12-EPE/hBMP-2 cmRNA lipoids was applied to a 3mm non-critical sized rat femoral defect model, resulting in the increased formation of mineralized callus volume and regenerated bone as early as 2 weeks at a very low dose compared to the fibrin only control defect. Additionally, more mature bone was observed in the BMP-2 cmRNA group as evidenced by the formation of haversian canals and the presence of bone lining cells in the bone tissue [29]. Badieyan ZS et al., further reported that vacuum-dried cmRNA-loaded collagen sponges, termed transcript activated matrices (TAMs), exerted 100% transfection efficiency of reporter target protein and can translate target protein for 6 days. TAM loaded 2.5µg BMP-2 cmRNA applied to a 2mm drill hole non-critical rat femur defect resulted in a statistically significant increase in bone callus volume, bone area, and osteoid volume as confirmed by μCT and histomorphometric analysis [30]. These findings indicate the therapeutic potential of BMP-2 encoding mRNA in non-critical size long bone defect repair.
In addition to BMP-2 cmRNA , other osteogenic factors for mRNA therapy have also been explored in tibia fracture healing. Nelson AL et al. examined the activation of the Wnt/β-catenin pathway using β-cateninGOF cmRNA encapsulated within SM-102 lipid nanoparticles to promote endochondral ossification during fracture healing in mice. After injection of β-cateninGOF mRNA/LNP at 6 days post-fracture at various doses (10 to 45µg in 25-30µl volume) to a mouse tibia fracture site, it was found that 45µg β-cateninGOF cmRNA promoted bone fracture healing as revealed by increased bone formation and decreased cartilage composition and a higher BV/TV compared to control group. Similar effects were observed with 25ng of rWnt3A delivered at the fracture site [31].
2.2.2. Mrna Therapy for Repair of Critical Sized Segmental Long Bone Defects
Critical sized segmental bone defects remain a significant clinical challenge in terms of treatment. These defects, which can be caused by tumor resection, trauma, infection, or other diseases, currently rely on surgical treatment such as bone autografts, allografts, or free flaps[26,32]. The delivery of cmRNA has shown therapeutic potential in animal models to repair critical size segmental bone defects of the femur. De La Vega RE et al. evaluated BMP-2 cmRNA delivery versus recombinant BMP-2 protein (rhBMP-2) in a rat critical sized femoral defect model. Various doses of BMP-2 cmRNA or rhBMP-2 protein was loaded on a collagen sponge and implanted into a 5mm defect covered with a deep muscle pouch. BMP-2 cmRNA promoted dose-dependent bone regeneration, with complete defect bridging observed at 50μg but not at lower doses. The 50μg cmRNA group exhibited greater BV/TV and Tb.N at 8 weeks after surgery, faster mechanical recovery, initiated bone remodeling faster than control group, and produced superior bone quality when compared with all other groups. Similar results were observed with the application of 11µg rhBMP-2. Additionally BMP-2 delivered via cmRNA remained localized at the defect with no off-target detection in other tissues or organs and no adverse effects, whereas an ectopic callus was formed in the rhBMP-2 group at 11 µg dosage [33].
3. Mrna Therapy for Tendon Regeneration and Healing
Tendon related injuries account for 30% of musculoskeletal injury related conditions [34], yet advancements in repair techniques are necessary as surgical interventions have high rates of re-injury and attempts to reduce inflammation have variable success [34]. The use of novel therapeutics such as cmRNA presents a potential innovative approach to the healing of tendon injuries. Several studies thus far have explored the potential to achieve localized protein expression through cmRNA transfection in tendon tissues.
Groth K et al. demonstrated that injection of naked cmRNA encoding the reporter gene Luciferase in saline solution into tendon tissue explants in a variety of species including ovine, equine, bovine, rats, and porcine resulted in high expression of reporter gene (BLI) in the injection site. The authors further observed dose-dependent expression of reporter genes following injection in saline. They also proved the expression of Laz reporter gene and human BMP-7 protein in the explant tissues. They subsequently tested different solvents for injection of 50µg Luc cmRNA in bovine explants and found HEPES-buffered glucose (HBG 5 %) reached maximum expression after 24 hours among all solvents tested. Furthermore, 5% glucose in HEPES or saline reached equal Luciferase expression levels while DreamfectGold and Branched PEI did not induce Luciferase expression compared to an untreated tendon. The authors further tested the injection of naked cmRNA in the tendon of rats in vivo and found the expression of the reporter gene Luciferase (Luc) peaked in 24 hours and declined to baseline at 7 days. Finally, human BMP-7 cmRNA(100µg) injected into a ruptured rat tendon resulted in significantly high expression of hBMP-7 at 2 days and decreased collagen III expression in the regenerating tissues at day 7. Another animal tendon injury model using collagenase diluted in a fibrin and thrombin solution into superficial digital flexor tendons (SDFT) more closely resembled the clinical picture of acute tendinopathy in sheep. cmRNALuc was further tested using 5% HBG injection solution in a sheep tendinopathy model. They found target gene Luciferase expression did not increase over time in intact tendon, while dose-dependent Luciferase expression was found in the injured tendons [35]
Moreover, Herbst E et al. performed a pilot study by using 5% HEPES buffered glucose as injection solution to inject 110µg (1mg/ml cmRNA in 110µl) of basic fibroblast growth factor (bFGF) cmRNA or Luciferase cmRNA into a proximal and distal 2mm Achilles tendon defect in rats. The results showed Luciferase expression can be detected for up to 3 days post-treatment. The bFGF cmRNA group demonstrated increased tendon stiffness at 14 days compared with the Luciferase cmRNA control group, reaching levels similar to that of a contralateral healthy side tendon. No differences were observed in terms of load to failure or in the expression of COL1,2,3,4 or procollagen I compared with the Luciferase cmRNA-treated group. The bFGF cmRNA-treated group exhibited more oval-shaped nuclei than the Luciferase cmRNA control group. No side effects were detected [36]. It is noteworthy to mention that these earlier studies injected naked mRNA without any lipid nanoparticles or other delivery carriers, thereby requiring large doses of cmRNA.
Sturm L et al. optimized the efficiency of a cationic, hyperbranched poly(amidoamine)-based nanoparticles to deliver tdTomato mRNA to various cells including rat tendon derived stem/progenitor cells (rTDSPCs). It was found that nanoparticle (NP)-mediated mRNA delivery to tendon-derived cells was effective in a dose-dependent manner. The transfection efficiency for rTDSPC was 18.13% ± 12.07. Moderate amounts of NPs enhanced transfection efficiency, while higher doses caused cytotoxicity, evidencing the need to balance effect delivery with cell viability. Furthermore, an increase of the mRNA loading ratio (2:50, 4:50, or 6:50 w/w mRNA:NPs) had no impact on transfection efficiency[37]
More recently, Faustini B et al. aimed to improve tendon repair by targeting two pathways using injectable poly(amidoamine)-based polymers (ps-PAAQ) as nanoparticle (NP) carriers to deliver chemically modified ARCA-capped mRNAs (cmRNAs) encoding Interleukin-1 receptor antagonist (IL1RA) and Platelet-Derived Growth Factor-BB (PDGF-BB). In vitro study demonstrated that delivery of 0.5µg to 1µg of target cmRNA using the aforementioned nanoparticles efficiently induced target protein expression in tendon cells and explant tendon tissues. PDGF-BB cmRNA transfected cells exhibited enhanced proliferation and migration, as well as increased proliferation within tendon explants. IL1RA cmRNA transfected cells exhibited reduced levels of pro-inflammatory cytokines. In vivo, the authors found that delivery of 1.9µg of target cmRNA at 7 days post-surgery resulted in the most effective target protein expression and patellar tendon defect repair, compared to delivery immediately after surgery or a staged approach involving immediate delivery of IL1RA cmRNA followed by PDGF-BB cmRNA at 7 days in a pilot study. Furthermore, simultaneous delivery of 1.9 µg PDGF-BB cmRNA and 1.9 µg IL1RA cmRNA using Ps-PAAQ nanoparticles at 7 days post-surgery improved tendon repair in a rat patellar tendon window defect model (2mm diameter). The beneficial effects were mediated by a reduction in inflammatory cells (CD68), decreased expression of pro-inflammatory cytokine (COX2) at early time points post-injury, reduced levels of fibrotic markers (S100a4), and an enhancement in repair tissue structure [38]
Zhang Y et al. demonstrated that in vitro delivery of mouse IL1RAcmRNA to tendon stem cells induced IL1RA protein expression for up to 72 hours, suppressed TNFα, IL6, and iNOS expression, and restored Collagen I/III balance as well as cell migration. In vivo, a single injection of 2µg IL1RA cmRNA encapsuled within SM102 lipid nanoparticles (LNPs) in a collagenase I-induced tendinopathy model in mice reduced inflammatory markers and MMP1/13 expression and improved collagen alignment at 1week post-administration. At 4 weeks post-administration, histology results showed improved collagen organization, increased COL1, decreased COL3, and decreased COLIII/I ratio compared to the untreated group, and functional recovery was evidenced by the re-establishment of typical gait pattern as normal uninjured mice [39] . Collectively, these studies demonstrated that delivery of cmRNA encoding IL1RA, by targeting inflammation, promotes the regeneration of tendon defects and offers a potential approach for the treatment of tendinopathy. More recent studies have demonstrated that delivering cmRNA via lipid nanoparticles could more effectively induce target therapeutic protein expression with much lower dose of mRNA.
4. Mrna Therapy for the Regeneration of Articular Cartilage
4.1. Mrna for Traumatic Osteoarthritis Repair
Osteoarthritis (OA) is an increasingly prevalent cause of disability in older adults [40,41]. While conventional, non-surgical, treatment options are aimed at symptom management, approaches that are capable of regenerating or slowing the degeneration of cartilage remain limited and are still under active research [42]. In the context of mRNA-based approaches for chondrogenesis, several mRNAs encoding proteins such as runt-related transcription factor-1 (Runx1), insulin-like growth factor-1 (IGF-1), fibroblast growth factor-18 (FGF-18), and transforming growth factor beta-1 (TGFβ-1) have been investigated.
Runx1 represents a promising mRNA therapeutic target as it has a regulatory role in articular cartilage maintenance and OA development [43,44]. Overexpression of Runx1 within the knee joint of an OA mouse model has been shown to have a protective effect on articular cartilage [45]. Aini L et al. first reported using mRNA encoding Runx1 for cartilage repair. Two polyethylene glycol (PEG)-polyamino acid block copolymer-based polyplex nanomicelles were developed: PEG-PAsp (DET) and PEG-PAsp(TET). PEG-PAsp (DET) promoted strong reporter gene (Luciferase) expression at 24hours, whereas PEG-PAsp(TET) induced target reporter gene expression at 4 hours that persisted for up to 96 hours. Intra-articular injection of Runx1 mRNA (1µg in 20µl volume every 3 days for one month) via PAsp(DET) nanomicelles improved MCL- and MM-transection-induced OA, but the effects did not reach statistical significance compared to GFP mRNA. However, administration of an equivalent dose of Runx1 mRNA using PEG-PAsp(TET) nanomicelles in the same mouse OA model induced target protein expression in superficial and middle zones of the articular cartilage, significantly suppressed OA development (decreased OARSI histology score) and formation of osteophytes, and increased SOX9 and COL2 expression and cell proliferation [46]. Subsequently, Pezzotti G et al. used the same animal model to deliver Runx1 mRNA using PAsp(DET) nanomicelles and evaluated the outcomes using Raman spectroscopic analysis, which revealed cartilage regeneration at the molecular level. The mechanism of cartilage restoration was attributed to activation of remaining chondrocytes by Runx1 leading to increased hyaluronic acid synthesis and restoration of organized collagen secondary structures [47]
Treatment with IGF-1 mRNA has also shown promising results. The intra-articular injection of IGF-1 mRNA transfected adipose derived stem cells (ADSCs) (2×105) at 1 and 2 weeks post-DMM surgery prevented the degeneration of articular cartilage as proved by decreased OARSI histology score and increased COL2 and ACAN expression [48]. FGF-18 mediates proliferation and differentiation of chondrocytes and has been investigated in its recombinant form (rhFGF-18) in clinical trials for the regenerative treatment of OA [49]. Huang K et al. designed a new proprietary lipid nanoparticle TG6A with branched tails and five ester bonds. This TG6A LNP significantly increased reporter GFP mRNA expression in transfected MSCs by 9- and 41-fold compared to commercialized DLin-MC3-DMA and ALC-0315 lipid nanoparticles respectively. MSCs transfected with TG6A LNP-encapsulated circular FGF-18 mRNA exhibited enhanced chondrogenic differentiation in an in vitro 3D pellet culture differentiation model. In a rat OA model, transplantation with circular FGF18-engineered MSCs not only protected cartilage from damage, but also improved repair as evidenced by thicker cartilage layers, reduced histopathological scores, maintenance of zone structure, and higher type II collagen and extracellular matrix (ECM) deposition compared to untreated control [50]
Furthermore, Sun M et al. developed a novel articular cavity-localized lipid nanoparticle (LNP) named WG-PL14. This optimized formulation has a nearly 30-fold increase in mRNA expression as well as better articular cavity enrichment compared to commercial MC3 lipids after intra-articular injection. Treatment of anterior cruciate ligament transection induced (ACLT) OA mice with an intra-articular injection of 2µg (in 20µl/knee, once a week for 3 weeks) mRNA encoding rhFGF-18 complexed with WG-PL14 LNPs reduced OA progression. Delivery of rhFGF-18 encoding mRNA to the joint reduced osteophyte formation and increased tibial subchondral bone BV/TV,Tb.Th and bone mineral density [51]. Consequently, rhFGF-18 mRNA delivered on optimized LNPs also improved pain response, upregulated cartilage extracellular matrix (ECM)-related genes such as COL2 and ACAN, and decreased COL1, MMP13 and IL1β [51]. More recently, Kong K et al. reported that LNP/FGF18 mRNA can deliver FGF18 cmRNA deeper within cartilage than proteins. Intra-articular injection of FGF18 mRNA/LNP (6µg weekly in 20ul) or 500 ng FGF18 protein (in 20µl) for 8 weeks in the joint of DMM-induced age-related OA models improved OA symptoms via activation of the FOX3A-autophage pathway, protecting chondrocytes from degeneration and senescence [41]. Together, these findings highlight the potential of FGF-18 mRNA, delivered directly or via engineered MSCS, for the treatment of OA.
4.2. Cmrna for Osteochondral Defect Repair
In addition to using cmRNA for traumatic osteoarthritis treatment, cmRNA has also been tested in an osteochondral defect model.
TGFβ-1 mRNA plays an important role in cartilage homeostasis and has also been investigated for use in cartilage defect repair [52]. Fontana G et al. developed mRNA delivery vehicles using mineral coated microparticle (MCM) and fluoride MCM (FMCM) and mimicked a clinical setting using bone marrow aspirate concentrate (BMAC) as ex vivo mRNA carrier. After complexing TGFβ-1 mRNA (at an unspecified dose) with FMCMs, the mRNA was used to transfect BMAC. The transfected BMAC was then combined with autologous peripheral blood to form a clot, which was implanted into a 2.7mm condyle osteochondral defect created in a rabbit knee. Rabbits treated with the BMAC-TGFβ-1 mRNA regenerated cartilage with increased type II collagen and glycosaminoglycan deposition, as well as reduced COL1 formation; however, no improvement in Odriscoll’s histology scores was observed [53]. Collectively, these studies demonstrate that mRNA-based delivery of chondrogenic factors represents a promising approach to the protection and restoration of osteoarthritic cartilage and osteochondral defect repair.
5. Mrna for Intervertebral Disc Regeneration
Intervertebral disc degenerative disease (IVDD) affects millions of people worldwide and is a major factor contributing to low back and neck pain and associated disability. Current treatments mainly focus on symptom management using surgical approaches, yet no treatments address disc regeneration, as the underlying mechanisms leading to IVDD are still elusive.
Lin C et al. used PAsp (DET) to form nanomicelles complexed with Runx1-mRNA, which were then injected (4µg in 6µl) into a needle puncture induced disc injury model in the rat coccygeal 4-5 disc. They found that injection of nanomicelle-Runx1 mRNA complexes reduced inflammation and increased intradiscal Runx1 expression compared to naked Runx1 mRNA injection, which in turn reduced disc height loss, prevented fibrous tissue formation, increased disc hydration content as revealed by MRI, and enhanced nucleus pulposus ECM production of COL2 and ACAN at 2 and 4 weeks post-injury [54,55]. Antony J S et al. designed a new IGF-1 mRNA construct using neurotrophin-3 (NTF3) signal peptide, pro-human IGF-1 domain, and a full length IGF-1 mRNA coding sequence (Cpd.3) to enhance IGF-1 secretion. This IGF-1 mRNA construct secreted higher levels of mature IGF-1 protein than IGF-1 natural signal peptide sequence (Cpd.1) following transfection of multiple cell lines. Starting at one month after the induction of a lumbar spine disc degeneration model using needle puncture and disc denucleation (disc herniation model), injection of 20µg of Cpd.3 IGF-1 mRNA in 20µl were performed on days 30, 37, 44 and 51 using citrated saline buffer. At 90 days following the injection of Cad.3 IGF-1 mRNA, the disc height index and histology scores were significantly higher than saline-injected discs, although still inferior to uninjured controls [56]. The success of these studies using mRNA therapy in ameliorating IVDD progression demonstrated potential therapeutic benefits of delivery of Runx1 and IGF-1 mRNAs.
6. Mrna Therapy for Skeletal Muscle Injury and Repair
Antony J S et al. designed a new IGF-1 mRNA construct using brain derived growth factor (BNDF) signal peptide, pro-domain of human IGF-1, and a full length IGF-1 mRNA coding sequence (Cpd.2) to enhance IGF-1 secretion. This modified IGF-1 mRNA construct secreted 3.1- to 6.1-fold higher levels of mature IGF-1 protein than IGF-1 natural signal peptide sequence (Cpd.1) after transfection of multiple cell lines. The concentration dependent EC50 was equivalent to clinically approved recombinant protein INCRELEX [56]. Injection of Cpd.2 IGF-1 mRNA (1µg) to a notexin-induced myotoxic mouse muscle injury achieved functional muscle recovery equivalent to that of 10µg Cad.1 IGF-1-mRNA. A dose dependent injection revealed Cpd.2 IGF-1 mRNA achieved peak effects at a 3µg dose (bell-shaped effect), whereas Cpd.1 IGF-1 mRNA did not reach a bell-shaped effect even at a 30µg dosage. The healing potency of Cpd.2 IGF-1 mRNA was found to be 15-fold higher than Cpd.1 IGF-1 mRNA in a skeletal muscle injury model [56]. Furthermore, a single injection of Cpd.2 IGF-1 mRNA in a rat punch TA muscle injury model produced IGF-1 protein levels of 18pg/mg protein within the 4mm injection site, reaching the therapeutic range and activating the down-stream signaling pathway (pAKT), with effects spreading up to 4-8mm from the injection site. Human IGF-1 expression was sustained for 3 days, with an approximate 20-hour half-life, and activated the down-stream IGF-IR signaling pathway. Furthermore, injection of 3µg Cpd.2 IGF-1 mRNA significantly increased the expression of early myogenic differentiation markers PAX7 and MYH3, as well as MyoD1, MYH5, and MyoG expression within 72hrs in the 4-8 mm injection site. The late myogenic markers MYH8 and MYH4 did not change within 72 hours. This supports the therapeutic effects of Cpd.2 IGF-1 mRNA in this more severe muscle injury model [56].
7. Advantages and Disadvantages of Protein Encoding Mrna Therapy for Musculoskeletal Tissue Repair
Following the success of the COVID-19 mRNA vaccines in controlling the COVID-19 pandemic and the Nobel Prize recognition for the development of modified mRNA technology, protein encoding mRNA therapeutic gained momentum over the past decade. Therapeutic mRNA applications in the treatment of orthopaedic injuries and degenerative diseases have progressed significantly and showed promising preclinical results. It is recognized that the use of mRNA as a vaccine is advantageous in its applications as mRNA vaccines leverage muscle as a protein-producing organ to release the target antigen into the blood stream, thereby stimulating the immune system to produce cellular and humoral immunity. The immune system does not require large amounts of the target protein or sustained protein release to be effective. Another advantage is that therapeutic mRNA can be prepared rapidly, without the need for lengthy protein purification and refolding processes. Well-developed lipid nanoparticle delivery systems and modified uridine or cytidine further reduce mRNA incurred immune response.
However, application of mRNA therapeutics to musculoskeletal tissue regeneration is not without shortcoming. One limitation is most of the studies using mRNA therapeutics to repair bone presented better BV/TV and very few presented the total regenerated bone volume. In terms of bone tissue engineering or regeneration, the regenerated bone volume is of higher importance, meaning that complete defect healing is more important than partial healing with a high BV/TV. For the treatment of bone defects, most studies use substantially higher doses of mRNA compared with protein. For example, in treating a rat segmental bone defect model, 50µg BMP-2 mRNA was required to achieve complete healing, compared to 11µg BMP-2 protein, with the additional requirement to cover the mRNA/LNP using host muscle tissues [33]. In a mouse tibia fracture model, 45µg β-catenin mRNA was used and produced effects comparable to using 25ng Wnt3A protein [31]. The requirement for such high mRNA doses may be due to the lack of host cells within the bone defect after injury, resulting in a limitation of host cell transfection and subsequent degradation of large amounts of unincorporated mRNA. In contrast, protein administration with sustained released biomaterials only required a very small dose of BMP-2 (as little as 1µg) to achieve complete bridging of a critical size femoral defect [57] or calvarial bone defect [17] . Secondly, once mRNA is applied in the defect, it is not certain which cells it will transfect. For example, if mRNA/LNP transfects inflammatory cells, they will not undergo osteogenesis. It is also hard to determine the exact quantity of the target protein produced. In contrast, protein delivery can elicit immediate functional effects and even recruit host reparative cells to the defect area. Most studies that track target protein expression use reporter genes, not therapeutic genes. On the other hand, mRNA transfected stem cells (ex vivo therapy) appeared more effective, likely due to the delivery cells themselves playing an important role in the regenerative process. In this aspect, compared to retroviral or lentiviral vector ex vivo gene therapy, mRNA transfected cells do not offer extended expression of target protein, but rather transit expression only (maximum 6 days in vitro or in vivo).
However, mRNA therapy appeared more effective in tendon, articular cartilage, skeletal muscle, and intervertebral disc repair than in bone defect repair and did not require a higher dose nor a lipid nanoparticle delivery system, compared to its application in bone defect repair. This is likely due to the tendon, articular cartilage, skeletal muscle, and disc injury sites having target cells for mRNA to transfect, resulting in therapeutic target protein expression in the injury site.
8. Perspective
The application of mRNA therapeutics in musculoskeletal regeneration represents a promising and rapidly evolving field. Despite much progress in the applications of protein encoding-mRNA for orthopaedic tissue regeneration, gaps remain in the existing literature. First, direct comparative studies are needed between mRNA targets, dosages, and delivery platforms (e.g., lipid nanoparticles, scaffolds, stem cell carriers) to optimize effects for repair of various tissues. Current studies lack standardization of outcome measures, complicating the ability to compare across studies and establish clinical relevance. Second, regarding combinatorial growth factor therapies, the synergistic and inhibitory relationships observed between BMP-2 and various growth factors [13,22,23] warrant further investigation of additional combinations to optimize therapeutic efficacy. The contradictory results might be related to the delivery nanoparticle and timing of administration rather than the growth factor’s role in bone defect healing. Third, despite tendon, articular cartilage, and intervertebral disc repair using cmRNA being less developed than for bone repair, it appears that only lower doses are required when using lipid nanoparticles for mRNA delivery of target therapeutic genes and is therefore more promising in terms of cost, potential side effects, and clinical application. Identification of therapeutic mRNA targets specific for tendon, articular cartilage, and other musculoskeletal tissues requires further investigation. BMPs have been shown effective in cartilage repair either using gene therapy or biomaterial mediated-delivery [4,58,59,60], but have not been studied using mRNA delivery for articular cartilage repair. Furthermore, even fewer studies have investigated mRNA therapy for muscle injury repair. No study has been conducted for meniscus or ligament repair likely due to difficulties in creating injury models and the delivery of target mRNA. Finally, while the short-term safety and efficacy have been evaluated in pre-clinical models, additional research is needed to assess long term effects, off-target expression, immune responses, and challenges before translation to treatment of human patients. Future studies should focus on optimizing mRNA constructs and delivery systems to maximize efficiency and minimize off target effects and cytotoxicity. Investigations into synergistic effects of growth factor combinations, particularly in tendon and cartilage repair, are warranted and may further enhance regenerative outcomes. Additionally, long-term preclinical and translational studies are needed to address safety, efficacy, and potential clinical applications.
9. Conclusion
In summary, protein encoding mRNA-based therapeutics offer a novel approach to musculoskeletal tissue regeneration that can overcome some limitations of protein and gene therapies. Current available preclinical literatures have shown efficacy of protein encoding mRNA in the regeneration or repair of craniomaxillofacial and long bone defects, tendon defects or injuries, OA and IVDD, and skeletal muscle injuries using various functional target genes, but no clinical trial has been initiated. However, comprehensive studies addressing the choice of growth factors, doses, and delivery strategies are still needed. Additionally, further information regarding long-term safety, efficacy, and translational feasibility are necessary to fully understand the therapeutic potential of mRNA therapy for musculoskeletal regeneration before clinical translation of this research.
Author Contributions
Conceptualization, X.G. J.H. and B.F.; Writing—Original Draft Preparation, B.F and X.G.; Writing—Review and Editing, X.G and J.H; Validation, X.G. and J.H.; Supervision, X.G., J.H.; Project Administration J.H, X.G; Funding Acquisition J.H. All authors have read and agreed to the published version of the manuscript.
Funding
This project was funded by philanthropic gifts from the Crown Genomic study (Steadman Philippon Research Institute code 3036).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data
sharing is not applicable.
Acknowledgments
We appreciate Ms. Xiang Xiao, B.A from The Glassel School of Art, Museum of Fine Arts, Houston, for making the Graphical abstract using Figma for this manuscript.
Conflicts: of Interest
All authors declare no conflicts of interest.
Declaration of using AI for generating manuscript
We did not use AI assistance to write this paper.
References
- Palazzo, C.; Ravaud, J. F.; Papelard, A.; Ravaud, P.; Poiraudeau, S. The burden of musculoskeletal conditions. PLoS One 2014, 9((3)), e90633. [Google Scholar] [CrossRef]
- Ehlen, Q. T.; Costello, J. P., 2nd; Mirsky, N. A.; Slavin, B. V.; Parra, M.; Ptashnik, A.; Nayak, V. V.; Coelho, P. G.; Witek, L. Treatment of Bone Defects and Nonunion via Novel Delivery Mechanisms, Growth Factors, and Stem Cells: A Review. ACS Biomater Sci Eng 2024, 10((12)), 7314–7336. [Google Scholar] [CrossRef]
- Gao, X.; Ruzbarsky, J. J.; Layne, J. E.; Xiao, X.; Huard, J. Stem Cells and Bone Tissue Engineering. Life (Basel) 2024, 14((3)). [Google Scholar] [CrossRef]
- Deng, Z.; Li, Y.; Gao, X.; Lei, G.; Huard, J. Bone morphogenetic proteins for articular cartilage regeneration. Osteoarthr. Cartil. 2018, 26, 1153–1161. [Google Scholar] [CrossRef]
- Gillman, C.E.; Jayasuriya, A.C. FDA-approved bone grafts and bone graft substitute devices in bone regeneration. Mater. Sci. Eng. C 2021, 130, 112466–112466. [Google Scholar] [CrossRef]
- Hsu, W.K.; Wang, J.C. The use of bone morphogenetic protein in spine fusion. Spine J. 2008, 8, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Watson-Levings, R.S.; Palmer, G.D.; Levings, P.P.; Dacanay, E.A.; Evans, C.H.; Ghivizzani, S.C. Gene Therapy in Orthopaedics: Progress and Challenges in Pre-Clinical Development and Translation. Front. Bioeng. Biotechnol. 2022, 10, 901317. [Google Scholar] [CrossRef] [PubMed]
- Tierney, E. G.; Duffy, G. P.; Cryan, S. A.; Curtin, C. M.; O'Brien, F. J. Non-viral gene-activated matrices: next generation constructs for bone repair. Organogenesis 2013, 9((1)), 22–8. [Google Scholar] [CrossRef]
- De la Vega, R.E.; Atasoy-Zeybek, A.; Panos, J.A.; Van Griensven, M.; Evans, C.H.; Balmayor, E.R. Gene therapy for bone healing: lessons learned and new approaches. Transl. Res. 2021, 236, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Chu, X.; Xiong, Y.; Lu, L.; Wang, Y.; Wang, J.; Zeng, R.; Hu, L.; Yan, C.; Zhao, Z.; Lin, S.; et al. Research progress of gene therapy combined with tissue engineering to promote bone regeneration. APL Bioeng. 2024, 8, 031502. [Google Scholar] [CrossRef]
- Ge, H.; Shi, Z.; Liu, C.; Lu, J.; Yao, Y.; Cheng, B. Viral and non-viral vectors for gene therapy in the treatment of bone-related disorders: molecular insights and clinical perspectives. Mol. Asp. Med. 2025, 105, 101400. [Google Scholar] [CrossRef]
- Mancino, C.; Franke, M.; Greco, A.; Sontam, T.; Mcculloch, P.; Corbo, C.; Taraballi, F. RNA therapies for musculoskeletal conditions. J. Control. Release 2024, 377, 756–766. [Google Scholar] [CrossRef] [PubMed]
- Surisaeng, T.; Wisitrasameewong, W.; Champaiboon, C.; Sa-Ard-Iam, N.; Chanamuangkon, T.; Thongnuek, P.; Tam, Y.K.; Muramatsu, H.; Weissman, D.; Pardi, N.; et al. BMP-2 mRNA-transfected BMSCs promote superior calvarial bone regeneration. Sci. Rep. 2025, 15, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Dimitriou, R.; Jones, E.; McGonagle, D.; Giannoudis, P.V. Bone regeneration: current concepts and future directions. BMC Med. 2011, 9, 66–66. [Google Scholar] [CrossRef] [PubMed]
- Jagadale, S.; Damle, M.; Joshi, M.G. Bone Tissue Engineering: From Biomaterials to Clinical Trials. Adv Exp Med Biol 2025, 1479, 73–115. [Google Scholar]
- Santos, W.; Benito-Ramal, E.; Toledano-Serrabona, J.; Sánchez-Garcés, M. Efficacy of bone morphogenetic protein in comparison with autogenous bone in regeneration of ameloblastoma bone defects. A systematic review. Med. Oral Patol. Oral Y Cirugia Bucal 2024, 29, e782–e790. [Google Scholar] [CrossRef]
- Gao, X.; Hwang, M.P.; Wright, N.; Lu, A.; Ruzbarsky, J.J.; Huard, M.; Cheng, H.; Mullen, M.; Ravuri, S.; Wang, B.; et al. The use of heparin/polycation coacervate sustain release system to compare the bone regenerative potentials of 5 BMPs using a critical sized calvarial bone defect model. Biomaterials 2022, 288, 121708–121708. [Google Scholar] [CrossRef]
- Schoonraad, S.A.; Jaimes, A.A.; Singh, A.J.; Croland, K.J.; Bryant, S.J. Osteogenic effects of covalently tethered rhBMP-2 and rhBMP-9 in an MMP-sensitive PEG hydrogel nanocomposite. Acta Biomater. 2023, 170, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Elangovan, S.; Khorsand, B.; Do, A.-V.; Hong, L.; Dewerth, A.; Kormann, M.; Ross, R.D.; Sumner, D.R.; Allamargot, C.; Salem, A.K. Chemically modified RNA activated matrices enhance bone regeneration. J. Control. Release 2015, 218, 22–28. [Google Scholar] [CrossRef]
- Khorsand, B.; Elangovan, S.; Hong, L.; Kormann, M.S.D.; Salem, A.K. A bioactive collagen membrane that enhances bone regeneration. J. Biomed. Mater. Res. Part B: Appl. Biomater. 2018, 107, 1824–1832. [Google Scholar] [CrossRef]
- Khorsand, B.; Elangovan, S.; Hong, L.; Dewerth, A.; Kormann, M.S.D.; Salem, A.K. A Comparative Study of the Bone Regenerative Effect of Chemically Modified RNA Encoding BMP-2 or BMP-9. AAPS J. 2017, 19, 438–446. [Google Scholar] [CrossRef]
- Geng, Y.; Duan, H.; Xu, L.; Witman, N.; Yan, B.; Yu, Z.; Wang, H.; Tan, Y.; Lin, L.; Li, D.; et al. BMP-2 and VEGF-A modRNAs in collagen scaffold synergistically drive bone repair through osteogenic and angiogenic pathways. Commun. Biol. 2021, 4, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Tsou, H.-K.; Wu, C.-H.; Chan, L.Y.; Kataoka, K.; Itokazu, N.; Tsuzuki, M.; Hu, H.; Zhuo, G.-Y.; Itaka, K.; Lin, C.-Y. Administration of mRNA-Nanomedicine-Augmented Calvarial Defect Healing via Endochondral Ossification. Pharmaceutics 2023, 15, 1965. [Google Scholar] [CrossRef] [PubMed]
- Friedlaender, G.E.; Lin, S.; Solchaga, L.A.; Snel, L.B.; Lynch, S.E. The Role of Recombinant Human Platelet-derived Growth Factor-BB (rhPDGF-BB) in Orthopaedic Bone Repair and Regeneration. Curr. Pharm. Des. 2013, 19, 3384–3390. [Google Scholar] [CrossRef] [PubMed]
- Jia, T.; Jacquet, T.; Dalonneau, F.; Coudert, P.; Vaganay, E.; Exbrayat-Heritier, C.; Vollaire, J.; Josserand, V.; Ruggiero, F.; Coll, J. L.; Eymin, B. FGF-2 promotes angiogenesis through a SRSF1/SRSF3/SRPK1-dependent axis that controls VEGFR1 splicing in endothelial cells. BMC Biol 2021, 19((1)), 173. [Google Scholar] [CrossRef]
- Zhang, M.; Fukushima, Y.; Nozaki, K.; Nakanishi, H.; Deng, J.; Wakabayashi, N.; Itaka, K. Enhancement of bone regeneration by coadministration of angiogenic and osteogenic factors using messenger RNA. Inflamm. Regen. 2023, 43, 1–16. [Google Scholar] [CrossRef]
- Marsell, R.; Einhorn, T. A. The biology of fracture healing. Injury 2011, 42((6)), 551–5. [Google Scholar] [CrossRef]
- Nicholson, J.; Makaram, N.; Simpson, A.; Keating, J. Fracture nonunion in long bones: A literature review of risk factors and surgical management. Injury 2021, 52, S3–S11. [Google Scholar] [CrossRef]
- Balmayor, E.R.; Geiger, J.P.; Aneja, M.K.; Berezhanskyy, T.; Utzinger, M.; Mykhaylyk, O.; Rudolph, C.; Plank, C. Chemically modified RNA induces osteogenesis of stem cells and human tissue explants as well as accelerates bone healing in rats. Biomaterials 2016, 87, 131–146. [Google Scholar] [CrossRef]
- Badieyan, Z.S.; Berezhanskyy, T.; Utzinger, M.; Aneja, M.K.; Emrich, D.; Erben, R.; Schüler, C.; Altpeter, P.; Ferizi, M.; Hasenpusch, G.; et al. Transcript-activated collagen matrix as sustained mRNA delivery system for bone regeneration. J. Control. Release 2016, 239, 137–148. [Google Scholar] [CrossRef]
- Nelson, A.L.; Mancino, C.; Gao, X.; Choe, J.A.; Chubb, L.; Williams, K.; Czachor, M.; Marcucio, R.; Taraballi, F.; Cooke, J.P.; et al. β-catenin mRNA encapsulated in SM-102 lipid nanoparticles enhances bone formation in a murine tibia fracture repair model. Bioact. Mater. 2024, 39, 273–286. [Google Scholar] [CrossRef]
- Dumic-Cule, I.; Pecina, M.; Jelic, M.; Jankolija, M.; Popek, I.; Grgurevic, L.; Vukicevic, S. Biological aspects of segmental bone defects management. Int. Orthop. 2015, 39, 1005–1011. [Google Scholar] [CrossRef]
- De La Vega, R.E.; van Griensven, M.; Zhang, W.; Coenen, M.J.; Nagelli, C.V.; Panos, J.A.; Silva, C.J.P.; Geiger, J.; Plank, C.; Evans, C.H.; et al. Efficient healing of large osseous segmental defects using optimized chemically modified messenger RNA encoding BMP-2. Sci. Adv. 2022, 8, eabl6242. [Google Scholar] [CrossRef] [PubMed]
- Andarawis-Puri, N.; Flatow, E.L.; Soslowsky, L.J. Tendon basic science: Development, repair, regeneration, and healing. J. Orthop. Res. 2015, 33, 780–784. [Google Scholar] [CrossRef] [PubMed]
- Groth, K.; Berezhanskyy, T.; Aneja, M.; Geiger, J.; Schweizer, M.; Maucksch, L.; Pasewald, T.; Brill, T.; Tigani, B.; Weber, E.; et al. Tendon healing induced by chemically modified mRNAs. 2017, 33, 294–307. [Google Scholar] [CrossRef]
- Herbst, E.; Imhoff, F.B.; Foehr, P.; Milz, S.; Plank, C.; Rudolph, C.; Hasenpusch, G.; Geiger, J.P.; Aneja, M.K.; Groth, K.; et al. Chemically Modified Messenger RNA: Modified RNA Application for Treatment of Achilles Tendon Defects. Tissue Eng. Part A 2019, 25, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Sturm, L.; Schwemberger, B.; Menzel, U.; Hackel, S.; Albers, C. E.; Plank, C.; Rip, J.; Alini, M.; Traweger, A.; Grad, S.; Basoli, V. In Vitro Evaluation of a Nanoparticle-Based mRNA Delivery System for Cells in the Joint. Biomedicines 2021, 9((7)). [Google Scholar] [CrossRef]
- Faustini, B.; Lettner, T.; Wagner, A.; Tempfer, H.; Cesur, N.P.; Lehner, C.; Brouwer, C.; Roelofs, K.; Mykhailyk, O.; Plank, C.; et al. Improved tendon repair with optimized chemically modified mRNAs: Combined delivery of Pdgf-BB and IL-1Ra using injectable nanoparticles. Acta Biomater. 2025, 195, 451–466. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, X.; Li, H.; Zhang, R.; Zhang, T.; Juma, T.; Zhou, Y.; Guo, Q.; Zhao, H.; Cao, Y. Anti-inflammatory therapy for tendinopathy using Il1rn mRNA encapsulated in SM102 lipid nanoparticles. Front. Bioeng. Biotechnol. 2025, 13, 1641236. [Google Scholar] [CrossRef]
- Hunter, D. J.; Bierma-Zeinstra, S. Osteoarthritis. Lancet 2019, 393((10182)), 1745–1759. [Google Scholar] [CrossRef]
- Kong, K.; Li, B.; Chang, Y.; Zhao, C.; Qiao, H.; Jin, M.; Wu, X.; Fan, W.; Wang, L.; Qi, Y.; et al. Delivery of FGF18 using mRNA-LNP protects the cartilage against degeneration via alleviating chondrocyte senescence. J. Nanobiotechnology 2025, 23, 1–21. [Google Scholar] [CrossRef]
- Zhang, W.; Ouyang, H.; Dass, C.R.; Xu, J. Current research on pharmacologic and regenerative therapies for osteoarthritis. Bone Res. 2016, 4, 15040. [Google Scholar] [CrossRef]
- Yano, F.; Ohba, S.; Murahashi, Y.; Tanaka, S.; Saito, T.; Chung, U.-I. Runx1 contributes to articular cartilage maintenance by enhancement of cartilage matrix production and suppression of hypertrophic differentiation. Sci. Rep. 2019, 9, 7666. [Google Scholar] [CrossRef]
- LeBlanc, K.T.; Walcott, M.E.; Gaur, T.; O'COnnell, S.L.; Basil, K.; Tadiri, C.P.; Mason-Savas, A.; Silva, J.A.; van Wijnen, A.J.; Stein, J.L.; et al. Runx1 Activities in Superficial Zone Chondrocytes, Osteoarthritic Chondrocyte Clones and Response to Mechanical Loading. J. Cell. Physiol. 2014, 230, 440–448. [Google Scholar] [CrossRef]
- Zhang, Y.; Zuo, T.; McVicar, A.; Yang, H.-L.; Li, Y.-P.; Chen, W. Runx1 is a key regulator of articular cartilage homeostasis by orchestrating YAP, TGFβ, and Wnt signaling in articular cartilage formation and osteoarthritis. Bone Res. 2022, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Aini, H.; Itaka, K.; Fujisawa, A.; Uchida, H.; Uchida, S.; Fukushima, S.; Kataoka, K.; Saito, T.; Chung, U.-I.; Ohba, S. Messenger RNA delivery of a cartilage-anabolic transcription factor as a disease-modifying strategy for osteoarthritis treatment. Sci. Rep. 2016, 6, srep18743. [Google Scholar] [CrossRef]
- Pezzotti, G.; Zhu, W.; Terai, Y.; Marin, E.; Boschetto, F.; Kawamoto, K.; Itaka, K. Raman spectroscopic insight into osteoarthritic cartilage regeneration by mRNA therapeutics encoding cartilage-anabolic transcription factor Runx1. Mater. Today Bio 2022, 13, 100210. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Peng, Z.; Xu, Y.; Sheng, Z.; Liu, Y.; Liao, Y.; Wang, Y.; Wen, Y.; Yi, J.; Xie, C.; et al. Engineered adipose-derived stem cells with IGF-1-modified mRNA ameliorates osteoarthritis development. Stem Cell Res. Ther. 2022, 13, 1–15. [Google Scholar] [CrossRef]
- Song, Z.; Li, Y.; Shang, C.; Shang, G.; Kou, H.; Li, J.; Chen, S.; Liu, H. Sprifermin: Effects on Cartilage Homeostasis and Therapeutic Prospects in Cartilage-Related Diseases. Front. Cell Dev. Biol. 2021, 9, 786546. [Google Scholar] [CrossRef]
- Huang, K.; Liu, X.; Qin, H.; Li, Y.; Zhu, J.; Yin, B.; Zheng, Q.; Zuo, C.; Cao, H.; Tong, Z.; et al. FGF18 encoding circular mRNA-LNP based on glycerolipid engineering of mesenchymal stem cells for efficient amelioration of osteoarthritis. Biomater. Sci. 2024, 12, 4427–4439. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Ma, B.; Pan, Z.; Zhao, Y.; Tian, L.; Fan, Y.; Kong, W.; Wang, J.; Xu, B.; Ao, Y.; et al. Targeted Therapy of Osteoarthritis via Intra-Articular Delivery of Lipid-Nanoparticle-Encapsulated Recombinant Human FGF18 mRNA. Adv. Heal. Mater. 2024, 13, e2400804. [Google Scholar] [CrossRef]
- Plaas, A.; Velasco, J.; Gorski, D.; Li, J.; Cole, A.; Christopherson, K.; Sandy, J. The relationship between fibrogenic TGFβ1 signaling in the joint and cartilage degradation in post-injury osteoarthritis. Osteoarthr. Cartil. 2011, 19, 1081–1090. [Google Scholar] [CrossRef]
- Fontana, G.; Nemke, B.; Lu, Y.; Chamberlain, C.; Lee, J.-S.; Choe, J.A.; Jiao, H.; Nelson, M.; Amitrano, M.; Li, W.-J.; et al. Local delivery of TGF-β1-mRNA decreases fibrosis in osteochondral defects. Bioact. Mater. 2025, 45, 509–519. [Google Scholar] [CrossRef]
- Lin, C.-Y.; Crowley, S.T.; Uchida, S.; Komaki, Y.; Kataoka, K.; Itaka, K. Treatment of Intervertebral Disk Disease by the Administration of mRNA Encoding a Cartilage-Anabolic Transcription Factor. Mol. Ther. - Nucleic Acids 2019, 16, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-C.; Tsou, H.-K.; Chang, H.-H.; Chan, L.Y.; Zhuo, G.-Y.; Maeda, T.; Lin, C.-Y. Runx1 Messenger RNA Delivered by Polyplex Nanomicelles Alleviate Spinal Disc Hydration Loss in a Rat Disc Degeneration Model. Int. J. Mol. Sci. 2022, 23, 565. [Google Scholar] [CrossRef]
- Antony, J.S.; Birrer, P.; Bohnert, C.; Zimmerli, S.; Hillmann, P.; Schaffhauser, H.; Hoeflich, C.; Hoeflich, A.; Khairallah, R.; Satoh, A.T.; et al. Local application of engineered insulin-like growth factor I mRNA demonstrates regenerative therapeutic potential in vivo. Mol. Ther. - Nucleic Acids 2023, 34, 102055. [Google Scholar] [CrossRef]
- Lee, S.S.; Huang, B.J.; Kaltz, S.R.; Sur, S.; Newcomb, C.J.; Stock, S.R.; Shah, R.N.; Stupp, S.I. Bone regeneration with low dose BMP-2 amplified by biomimetic supramolecular nanofibers within collagen scaffolds. Biomaterials 2013, 34, 452–459. [Google Scholar] [CrossRef]
- Matsumoto, T.; Cooper, G.M.; Gharaibeh, B.; Meszaros, L.B.; Li, G.; Usas, A.; Fu, F.H.; Huard, J. Cartilage repair in a rat model of osteoarthritis through intraarticular transplantation of muscle-derived stem cells expressing bone morphogenetic protein 4 and soluble flt-1. Arthritis Rheum. 2009, 60, 1390–1405. [Google Scholar] [CrossRef]
- Gao, X.; Cheng, H.; Awada, H.; Tang, Y.; Amra, S.; Lu, A.; Sun, X.; Lv, G.; Huard, C.; Wang, B.; et al. A comparison of BMP2 delivery by coacervate and gene therapy for promoting human muscle-derived stem cell-mediated articular cartilage repair. Stem Cell Res. Ther. 2019, 10, 1–13. [Google Scholar] [CrossRef]
- Gao, X.; Wright, N.; Huard, M.; Tan, J.; Ruzbarsky, J.; Lu, A.; Chubb, L.; Tuan, R.; Philippon, M.J.; Wang, Y.; et al. Comparison of 5 BMPs for their chondrogenic potentials and microfracture-mediated cartilage repair using heparin/PEAD coacervate sustained release polymer. Bioact. Mater. 2025, 52, 588–603. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



