Submitted:
17 February 2026
Posted:
25 February 2026
You are already at the latest version
Abstract
The beaded periwinkle (Cenchritis muricatus) inhabits supratidal rocky environments characterized by strong gradients in salinity, desiccation, and hydrodynamic disturbance. Preliminary observations suggested that individuals leave dry rocks more frequently when surrounded by seawater than freshwater, prompting an exploratory investigation of potential environmental cues underlying this behavior. Field-based pilot experiments in which periwinkles were placed on isolated dry rocks surrounded by either seawater or freshwater were conducted, while additional treatments varied rock height, surface char- acteristics, water depth, and salinity. Across experiments, periwinkles migrated away from rocks surrounded by seawater more frequently than those surrounded by freshwa- ter, although effect sizes varied and interactions with other factors were inconsistent. High variance and limited replication constrained statistical inference, and analyses are there- fore interpreted descriptively. Despite these limitations, results suggest that movement is not driven by immediate habitat benefits but may reflect sensitivity to salinity-associated contextual cues linked to large-scale disturbance risk. Escape behavior may be adaptive over long temporal scales associated with storm exposure, even if it appears maladaptive under experimentally constrained conditions. These findings highlight the importance of experimental scale in behavioral ecology and motivate future studies incorporating con- nected substrates, refuge gradients, and field-based validation to better resolve how su- pratidal gastropods respond to changing coastal conditions.
Keywords:
knobby periwinkle
; behaviour
; salinity
; migration
; seawater
; freshwater
; rugosity
1. Introduction
Rocky shore environments impose strong and rapidly fluctuating gradients in salinity, temperature, desiccation, and hydrodynamic stress [1,2,13,19]. Supratidal gastropods habiting these environments occupy a particularly challenging niche as they experience prolonged aerial exposure. This exposure is punctuated by episodic seawater inundation occurring during storms and extreme tides. Movement behaviour in these systems is therefore shaped by trade-offs between foraging opportunities, refuge availability, and risks associated with wave dislodgement and immersion [3,11,12].
Periwinkles (family Littorinoidea) exhibit a wide range of behavioural and physiological adaptations that allow persistence across these intertidal and supratidal zones. These include a high tolerance to desiccation, nocturnal or rain-associated foraging, mucus-mediated trail following, and the ability to assess fine-scale habitat features such as the surface texture and moisture availability [4,5,6]. In supratidal habitats, periwinkles are typically found on dry rocks above the high-tide line, where direct contact with seawater is infrequent and usually associated with storm events or wave splash [7].
The Caribbean beaded (knobby) periwinkle, Cenchritis muricatus (C. muricatus), is a semi-terrestrial species of marine origin that commonly inhabits supratidal rocky substrates [7]. Its tolerance to extreme desiccation, its reliance on damp conditions for foraging, and reduced population densities in areas of high wave exposure have been documented in previous studies [8,9,10]. Together, these findings suggest that C. muricatus experience strong selection pressures related to hydrodynamic disturbances and immersion of seawater.
Behavioural responses to perceived risks are often context-dependent and may be triggered by indirect environmental cues rather than immediate physical stress [11,12]. The presence of surrounding seawater may act as a proxy signal for elevated wave energy or flood risk, especially when combined with spatial isolation in intertidal and supratidal systems [1,9,12] Experimental investigations of such responses are complicated by scale mismatches due to small-scale laboratory and mesocosm setups which may exaggerate confinement and limit the availability of natural refuges [14].
This exploratory experimental study examines the movement behaviour of C. muricatus when placed on isolated dry rocks surrounded by either seawater or freshwater. Motivated by preliminary observations, it was tested whether the salinity of surrounding water was associated with differential likelihood of periwinkles leaving their home rock, and whether this response was modulated by rock size, orientation, surface rugosity, or water depth. The experimental design necessarily imposed spatial constraints; therefore, this study is explicitly framed as a hypothesis-generating investigation into potential cues which may influence periwinkle movement under constrained conditions [15,16].
2. Materials and Methods
The Study Site
This study took place at the Discovery Bay Marine Lab (DBML) located in the University of the West Indies, Jamaica spanning over two weeks from December 28th, 2023, to January 10th, 2024. Periwinkles (Figure 2) from Discovery Bay (Figure 1), Jamaica’s costal rocks near the sea, and freshwater or brackish water pools. Several locations near the DBML facilities, with each collection site about 5 m apart. Each periwinkle was collected from rocks close to the sea to ensure similar prior habitat experience among the experimental individuals. Two sites with similar characteristics were chosen: near the marine boardwalk pier and the boating dock. The rocks were taken from rock rubble deposits along the shoreline from the same or close locations in approximation to the original periwinkles
Figure 1.
Map of Discovery Bay Marine Lab where specimen collection occurred.

Figure 2.
The beaded periwinkle (C. muricatus) shells. From Bailey-Matthews National Shell Museum (https://www.shellmuseum.org/post/shell-of-the-week-the-beaded-periwinkle). Photos by James F. Kelly. Licence – none required.
Figure 2.
The beaded periwinkle (C. muricatus) shells. From Bailey-Matthews National Shell Museum (https://www.shellmuseum.org/post/shell-of-the-week-the-beaded-periwinkle). Photos by James F. Kelly. Licence – none required.

The Study Species and Experimental Rocks
The beaded periwinkle (C. muricatus; Figure 2) was the most abundant species among the study sites, and individuals often presented in groups on rocks near the splash zone. The number of periwinkles varied between experiments. A preliminary experiment was completed in a lab setup to obtain initial observations on periwinkle movement in seawater and freshwater environments.
The Experiments
Four experiments were conducted (details in Appendix A). In all but one experiment, rocks were placed in sea and freshwater tanks and then periwinkles were added to the dry areas of the rock (Figure 3). After 24 hours, periwinkles that left the rock were counted.
We interpreted the departure from the rock as a response to conditions a periwinkle perceived as unsatisfactory when the departure required travelling submerged in seawater. Staying on the rock indicated satisfactory conditions. Other treatments varied among experiments using rocks of different sizes, textures, or orientations of the main surface (horizontal or vertical).
Between January 8th to 10th, the terrain and rock size experiments were refined by only altering one variable. The rock was left the same size, shape and orientation; however, the only variable that was altered was the terrain of the rock. This ensures that the results were not due to chance. The experiments were then replicated without this alteration of every single variable, which aims to further validate how rock terrain influences the be-haviour of periwinkles where we could measure our results more accurately.
3. Results
Periwinkles left their home rock more often when it sat in seawater than in freshwater islands’ (Figure 4A, Table 1). However, this effect differed among the experiments (Figure 4B), where two experiments showed little or no difference between the salinity treatments. Interestingly, when water was about 60 cm deep, periwinkle migration did not differ between the fresh and seawater (Figure 5). The rock size also revealed no difference. This experiment highlights that the salinity effect depends on some variables) associated with the water depth, possibly the extent of water level movement or direct small wave action the experimental rock experienced. Such factors might explain why periwinkles do not err into the seawater in this one treatment but chose to do so preferentially in the lab. Overall, the salinity effect interacted with other treatments (Table 1), but those treatments suggested a range of diverse impacts on periwinkle migration without much consistency. Unsurprisingly, this interaction may arise from differences in the importance of the examined factors or, by chance, playing a significant role in the experiments with low replication.
The considerable differences between the effects of individual experiments suggest factors with a more significant role in affecting periwinkle responses to environmental influences (Figure 4). Regardless of rock properties, more periwinkles migrate through seawater than freshwater. Water depth affected experimental periwinkles differently, although we do not have statistical support for the response we observed. In any case, the departure of the water depth effect from the prevailing pattern of other responses should not be a surprise because deep seawater is likely to correlate with a lethal migration outcome.
Other observations, while unsupported formally, inspire some possibilities. For ex-ample, the response to rugosity was almost entirely explainable by chance, as were responses to salinity in the rugosity experiment. However, the interaction between the two factors reduced the role of chance five times. This may matter because one of the assumptions was that rugosity should protect periwinkles from the impact of waves in case of water’s edge proximity.
4. Discussion
4.1. Salinity as a Contextual Movement Cue Under Constrained Conditions
Across experiments, periwinkle individuals were more likely to leave isolated rocks when surrounded by seawater compared to freshwater, although the strength of this pattern varied among treatments. This movement occurred despite the seawater representing a physiologically and ecologically hostile medium for the species [8,10]. Rather than indicating adaptive preferences for exposure to seawater, the behaviour can be more plausibly interpreted as a response mechanism to contextual cues associated with elevated environmental risk.
In natural supratidal environments, seawater contact typically coincides with storm surge, extreme tides, or high wave energy conditions which can be associated with an increased risk of dislodgement and mortality [1,9,12]. Therefore, the presence of surrounding seawater may function as this proxy signal for such large-scale disturbances which may elicit escape responses even when immediate experimental context offers no clear refuge advantage.
4.2. Constraints Imposed by Experimental Isolation
A critical limitation of the present study is that experimental rocks were fully isolated by water, restricting the availability of alternative refuges and potentially biasing movement outcomes. Under such conditions, movement may reflect stress-induced displacement rather than active selection of habitats [14].
4.3. Modulating Effects of Rock Features and Water Depth
Rock size, orientation, and surface rugosity did not produce consistent effects on movement behaviour across experiments. While some of these interactions reduced the apparent role of random variation, limited replication and high variance preclude strong inference regarding the influence of these factors. Behavioural variability is common in gastropods and may reflect both physiological constraints and individual-level differences [16,18]. Salinity tolerance and stress responses documented across taxa nevertheless suggest that environmental salinity can interact with physiological limits to shape behavioural outcomes [17].
4.4. Scale Mismatch and Behavioural Interpretation
The variability observed in this study highlights a broader issue in behavioural ecology: small-scale experiments may elicit responses that are adaptive only when interpreted at larger temporal or spatial scales [14,15]. In the case of C. muricatus, escape behaviour triggered by seawater exposure may be beneficial over evolutionary timescales associated with rare but severe disturbance events [2], even if it appears maladaptive in simplified experimental settings.
5. Conclusions
This exploratory study suggests that C. muricatus exhibit differential movement responses when surrounded by seawater compared to freshwater under experimentally constrained conditions. While statistical and design limitation prevent definitive conclusions about adaptive function, the results are consistent with the hypothesis that the presence of seawater acts as a contextual risk cue which can be linked to large-scale disturbances.
Understanding how these supratidal organisms interpret and respond to environmental cues is increasingly important in the context of climate change, as rising sea levels, intensifying storm events, and altered salinity regimes are expected to increase the frequency and magnitude of seawater exposure in costal habitats. Behavioural sensitivity to such cues may therefore play a critical role in shaping species persistence, distribution, and vulnerability across rocky shore systems.
Beyond species-specific insights, this study highlights broader methodological challenges in marine behavioural ecology, particularly in the influence of spatial scale and experimental confinement on observed responses. Small-scale manipulations may elicit behaviours that are adaptive only when interpreted across larger ecological contexts, emphasizing the need for experimental designs that incorporate connected substrates, refuge gradients, and field-based validation.
Future research should build on these findings by examining movement behaviour across continuous habitat mosaics, integrating physiological stress measurements, and evaluating responses under natural disturbance regimes. Such approaches will help clarify whether salinity-associated cues function as early warning signals of environmental risk and will improve our ability to predict organismal responses to changing coastal conditions.
Author Contributions
Conceptualization, AH. And JK.; methodology, AH.; formal analysis, JK.; writing – draft preparation, AH.; writing – reviewing and editing, AH, JK and DM.; visualization, AH, JK.; supervision, JK. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is directly available from both authors.
Acknowledgments
The authors gratefully acknowledge the support of Discovery Bay Marine Lab for providing access to facilities, equipment, and logistical assistance essential in the completion of this study. We thank the laboratory staff for their technical support and guidance during field experiments.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| DBML | Discovery Bay Marine Lab |
Appendix A
Experiment 1. This experiment aimed to determine whether the availability of horizontal space influenced the periwinkle decision to stay or move from the rock. The tested premise was that resources (algae and organic debris resources are most likely available on horizontal rather than vertical surfaces). This experiment used 28 periwinkles and four rocks (seven periwinkles per rock). We arranged two elongated rocks with the largest surface being vertical and two rocks where it was horizontal, placed in both a seawater and freshwater tank for observation to simulate the periwinkle’s natural habitat where each treatment used two rocks (Figure A1). The horizontal rock had a total surface area of 806cm2, yielding 161.2cm? per periwinkle to provide a rough estimate of how much space the periwinkles could potentially cover. The rocks were set up in tanks, one with seawater and one with fresh water, both oriented in different directions (Figure A1). In the seawater tank, a pump created a constant seawater flow to keep results consistent. The freshwater tank also had a constant flow of freshwater. To further prove if the periwinkles had a preference, the experiment was rerun multiple times on Jan 4 to minimize overall error.
Figure A1.
Experiment 1: Rock Orientation. Wet-lab setup to assess the influence of rock position on periwinkle movement. n = 28 (n=7 per rock). A wet lab setup in the seawater environment testing a horizontal rock compared to a vertical rock (A). The freshwater environment setup compares a horizontal rock to a vertical rock (B). Water levels were maintained around 1cm.
Figure A1.
Experiment 1: Rock Orientation. Wet-lab setup to assess the influence of rock position on periwinkle movement. n = 28 (n=7 per rock). A wet lab setup in the seawater environment testing a horizontal rock compared to a vertical rock (A). The freshwater environment setup compares a horizontal rock to a vertical rock (B). Water levels were maintained around 1cm.

Experiment 2. This experiment examined whether the periwinkles responded to rock size. Preference differences for rock size likely imply that the periwinkles could assess the resource availability or the risk of drowning before deciding to leave the rock. For example, they might exhaust resources quickly and must move to find food, or they might perceive the proximity of water as a danger indicator (drowning, fish attack?). To guard against differences in periwinkle density on rocks of different sizes, we added snail individuals in different numbers to maintain one periwinkle for each 45-55cm2. Rocks of three different sizes received 61 periwinkles, with 40 placed on the large rock, 15 on the medium-sized rock, and 6 on the small rock. Each periwinkle was initially placed 1-2 cm apart from its nearest neighbor. The tanks set up was the same as in Experiment 1 (Figure A2). This experiment was repeated over the next couple of days.
Figure A2.
Experiment 2: Rock size. Wet-lab setup to assess the impact of a small, medium, and large rock on periwinkle migration. (A) In the seawater tank, the large rock had a surface area of 50.14 cm2 per periwinkle (n=28). The medium rock had a surface area of 53.25 cm2 per periwinkle (n=20). The small rock had a surface area of 48.00 cm2 (n=5). (B) In the freshwater tank, the large rock had a surface area of 49.95 cm2 per periwinkle (n=40). The medium rock had a surface area of 45.80 cm2 per periwinkle (n=15). The small rock had a surface area of 48.75 cm2 per periwinkle (n=6). Water levels were maintained around 1cm.
Figure A2.
Experiment 2: Rock size. Wet-lab setup to assess the impact of a small, medium, and large rock on periwinkle migration. (A) In the seawater tank, the large rock had a surface area of 50.14 cm2 per periwinkle (n=28). The medium rock had a surface area of 53.25 cm2 per periwinkle (n=20). The small rock had a surface area of 48.00 cm2 (n=5). (B) In the freshwater tank, the large rock had a surface area of 49.95 cm2 per periwinkle (n=40). The medium rock had a surface area of 45.80 cm2 per periwinkle (n=15). The small rock had a surface area of 48.75 cm2 per periwinkle (n=6). Water levels were maintained around 1cm.

Experiment 3. The experiment examined the effect of rock rugosity (bumps and ridges) on periwinkle movement. The two rocks, one horizontally cracked and creviced and one with a smooth surface, received 10 periwinkles each and were placed in the tanks (Figure A3). The design was the same between the sea and freshwater tanks. The rocks were left undisturbed for 24. This experiment would reveal whether shelter availability affected periwinkles’ preference to stay on a rock even if surrounded by water. If yes, then the type of surrounding water might be less important than the attributes of the rock.
Figure A3.
Experiment 3: Rock terrain. Wet-lab setup to assess the effect of rock surface rugosity on periwinkle behaviour. A flat rock (n=10) and a cracked rock (10) in seawater (A). A flat rock (n=10) and a cracked rock (10) in freshwater (B). Water levels were maintained around 1cm.
Figure A3.
Experiment 3: Rock terrain. Wet-lab setup to assess the effect of rock surface rugosity on periwinkle behaviour. A flat rock (n=10) and a cracked rock (10) in seawater (A). A flat rock (n=10) and a cracked rock (10) in freshwater (B). Water levels were maintained around 1cm.

Experiment 4. The experiment tested if the depth of rocks in seawater affected periwinkles, aiming to observe that the periwinkles would still leave the rock regardless of depth as an escape mechanism. We placed a 60 cm long rock in the sea about 3 m from the shore, where most was submerged, leaving 11 cm of rock above the water surface, with ten periwinkles placed at the top (Figure A4). This experiment could only run once over 24 hours due to logistical constraints. A camera captured a time-lapse video of the periwinkle movement throughout the 24 hours.
Figure A4.
Experiment 4: Rock depth. Outdoor experimental setup to assess the tidal threshold that periwinkles can withstand. A 60cm long coral rock was placed with 11cm exposed (n=10). A time-lapse camera attached to a pole driven into the sea sediments recorded periwinkle movements.
Figure A4.
Experiment 4: Rock depth. Outdoor experimental setup to assess the tidal threshold that periwinkles can withstand. A 60cm long coral rock was placed with 11cm exposed (n=10). A time-lapse camera attached to a pole driven into the sea sediments recorded periwinkle movements.

References
- Denny, M. W. (1987). Life in the maelstrom: The biomechanics of wave-swept rocky shores. Trends in Ecology & Evolution, 2(3), 61–66. [CrossRef]
- Menge, B. A., & Sutherland, J. P. (1987). Community Regulation: Variation in Disturbance, Competition, and Predation in Relation to Environmental Stress and Recruitment. In Source: The American Naturalist (Vol. 130, Issue 5).
- Denny, M. W., & Paine, R. T. (1998). Celestial Mechanics, Sea-Level Changes, and Intertidal Ecology. In Reference: Biol. Bull (Vol. 194). [CrossRef]
- Davies, M. S., & Beckwith, P. (1999). Role of mucus trails and trail-following in the behaviour and nutrition of the periwinkle Littorina littorea. In Source: Marine Ecology Progress Series (Vol. 179). [CrossRef]
- Chapman, M. G. (2000). A comparative study of differences among species and patches of habitat on movements of three species of intertidal gastropods. In Journal of Experimental Marine Biology and Ecology (Vol. 244). [CrossRef]
- Little, C., Williams, G. A., & Trowbridge, C. D. (2009). The Biology of Rocky Shores. Oxford University PressOxford. [CrossRef]
- Thompson, D. The Online Guide to the Animals of Trinidad and Tobago: Cenchritis muricatus (Beaded Periwinkle). University of the West Indies, 2015. Available online: https://www.uwi.edu.
- Emson, R. H., Morritt, D., Andrews, E. B., & Young, C. M. (2002). Life on a hot dry beach: Behavioural, physiological, and ultrastructural adaptations of the littorinid gastropod Cenchritis (Tectarius) muricatus. Marine Biology, 140(4), 723–732. [CrossRef]
- Piovia-Scott, J. (2009). Effects of disturbance on Cenchritis muricatus (beaded periwinkle) populations on small islands in the Bahamas. In BULLETIN OF MARINE SCIENCE (Vol. 84, Issue 3).
- Rosewater, J. (1963). Resistance to desiccation in dormancy by Tectarius muricatus. Nautilus, 76, 111.
- Lima And, S. L., & Dill, L. M. (n.d.). Behavioral decisions made under the risk of predation: a review and prospectus.
- Brown, J. S., Laundre, J. W., & Gurung, M. (1999). The Ecology of Fear: Optimal Foraging, Game Theory, and Trophic Interactions. Journal of Mammalogy, 80(2), 385–399. [CrossRef]
- Helmuth, B., Broitman, B. R., Blanchette, C. A., Gilman, S., Halpin, P., Harley, C. D. G., O’Donnell, M. J., Hofmann, G. E., Menge, B., & Strickland, D. (2006). Mosaic patterns of thermal stress in the rocky intertidal zone: Implications for climate change. Ecological Monographs, 76(4), 461–479. [CrossRef]
- Underwood, A. J., & Chapman, M. G. (2003). Power, precaution, Type II error and sampling design in assessment of environmental impacts. Journal of Experimental Marine Biology and Ecology, 296(1), 49–70. [CrossRef]
- Resetarits, W. J. (2005). Habitat selection behaviour links local and regional scales in aquatic systems. Ecology Letters, 8(5), 480–486. [CrossRef]
- Moisez, E., & Seuront, L. (2020). Deciphering the known unknowns in the behavioural ecology of the intertidal gastropod Littorina littorea. Journal of Experimental Marine Biology and Ecology, 524. [CrossRef]
- Marković, M., Šoštarić, J., Kojić, A., Popović, B., Bubalo, A., Bošnjak, D., & Stanisavljević, A. (2022). Zinnia (Zinnia elegans L.) and Periwinkle (Catharanthus roseus (L.) G. Don) Responses to Salinity Stress. Water (Switzerland), 14(7). [CrossRef]
- Chapperon, C., & Seuront, L. (2011). Variability in the motion behaviour of intertidal gastropods: ecological and evolutionary perspectives. Journal of the Marine Biological Association of the United Kingdom, 91(1), 237–244. [CrossRef]
- Underwood, A. J. (2000). Experimental ecology of rocky intertidal habitats: what are we learning? In Journal of Experimental Marine Biology and Ecology (Vol. 250). [CrossRef]
Figure 3.
Wet-lab experimental setup – rocks with periwinkles. Freshwater (A). Seawater (B).

Figure 4.
Mean periwinkle numbers moved over all the combined experiments (A). Different treatments (individual experiments) were significantly different, but not individually (B).
Figure 4.
Mean periwinkle numbers moved over all the combined experiments (A). Different treatments (individual experiments) were significantly different, but not individually (B).
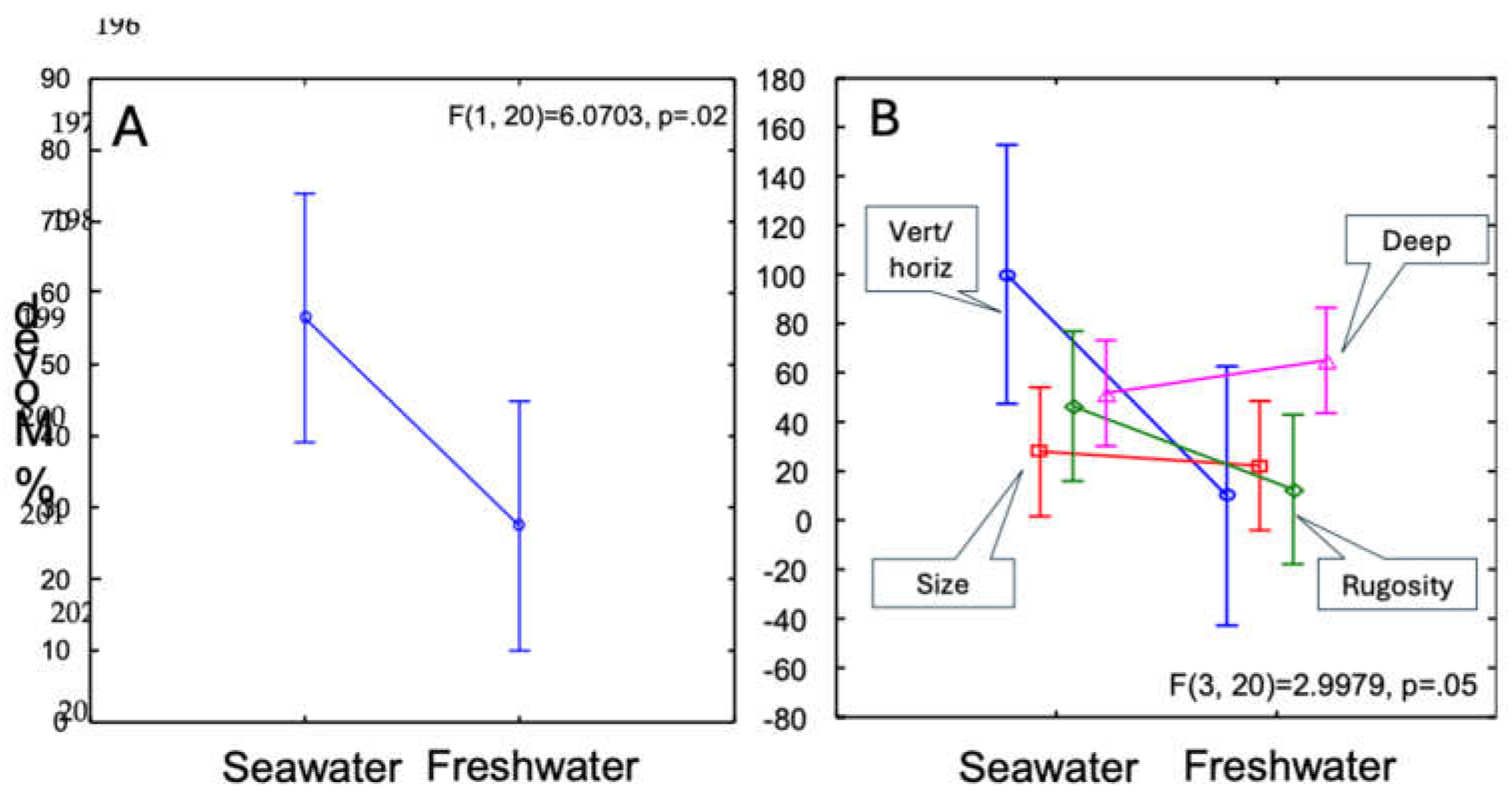
Figure 5.
Water salinity effect on periwinkle movement across the experiments (x-axis).
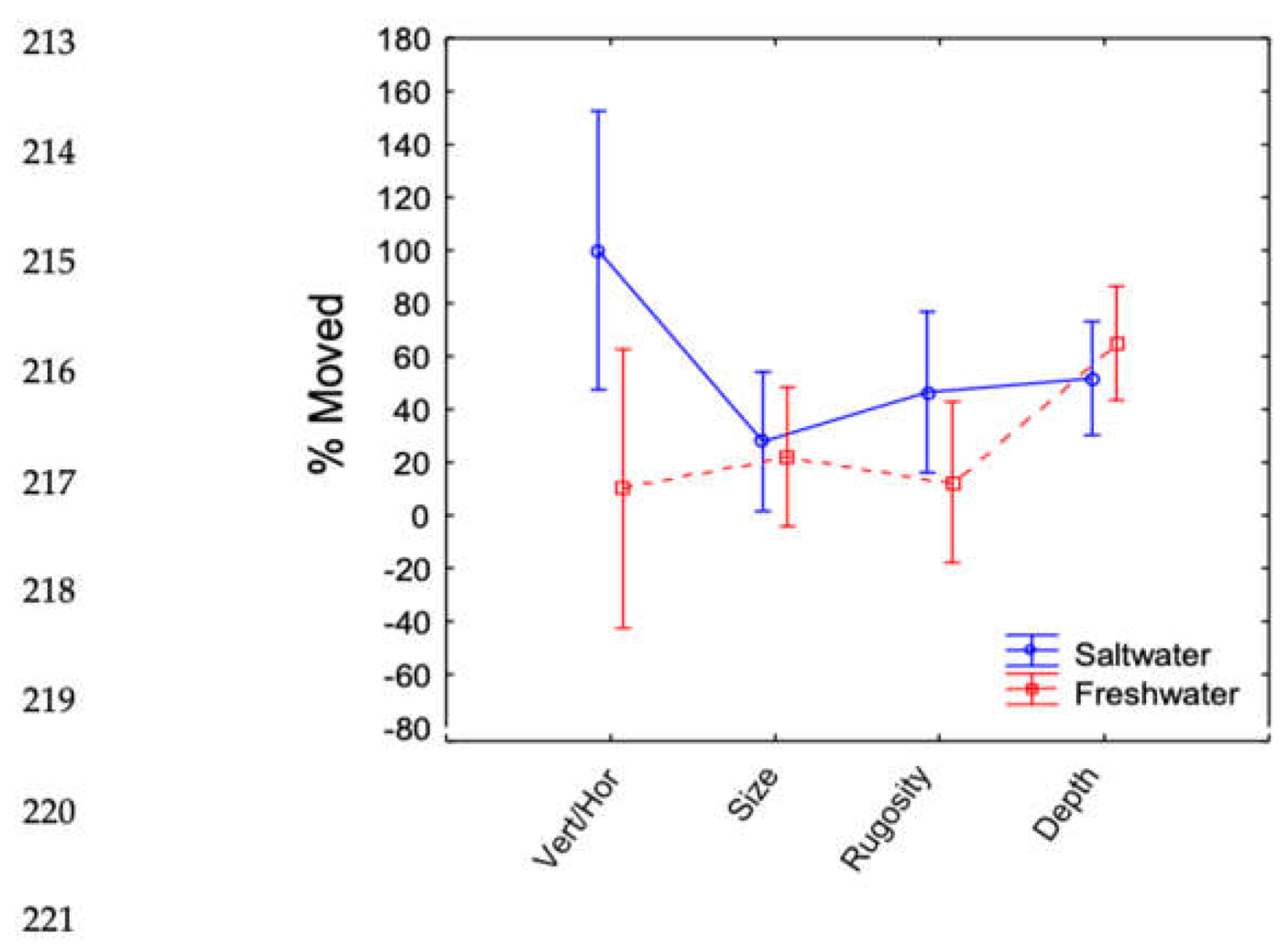

Table 1.
Factorial ANOVA results on periwinkle migration response to seawater and freshwater rock island environments across the four experiments. Note that the interactions between the Experiment and Salinity were significant at an alpha-level of 0.1.
Table 1.
Factorial ANOVA results on periwinkle migration response to seawater and freshwater rock island environments across the four experiments. Note that the interactions between the Experiment and Salinity were significant at an alpha-level of 0.1.
| SS | Degr. of Freedom | MS | F | P-value | |
| Intercept | 32184.04 | 1 | 32184.04 | 50.53258 | 0.000001 |
| Experiment | 6786.20 | 3 | 2262.07 | 3.55170 | 0.0329 |
| Fresh/Salt | 3866.14 | 1 | 3866.14 | 6.070 | 0.0229 |
| Experiment*Fresh/Salt | 5728.09 | 3 | 1909.36 | 2.998 | 0.0549 |
| Error | 12737.94 | 20 | 636.90 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



