Submitted:
15 February 2026
Posted:
17 February 2026
You are already at the latest version
Abstract
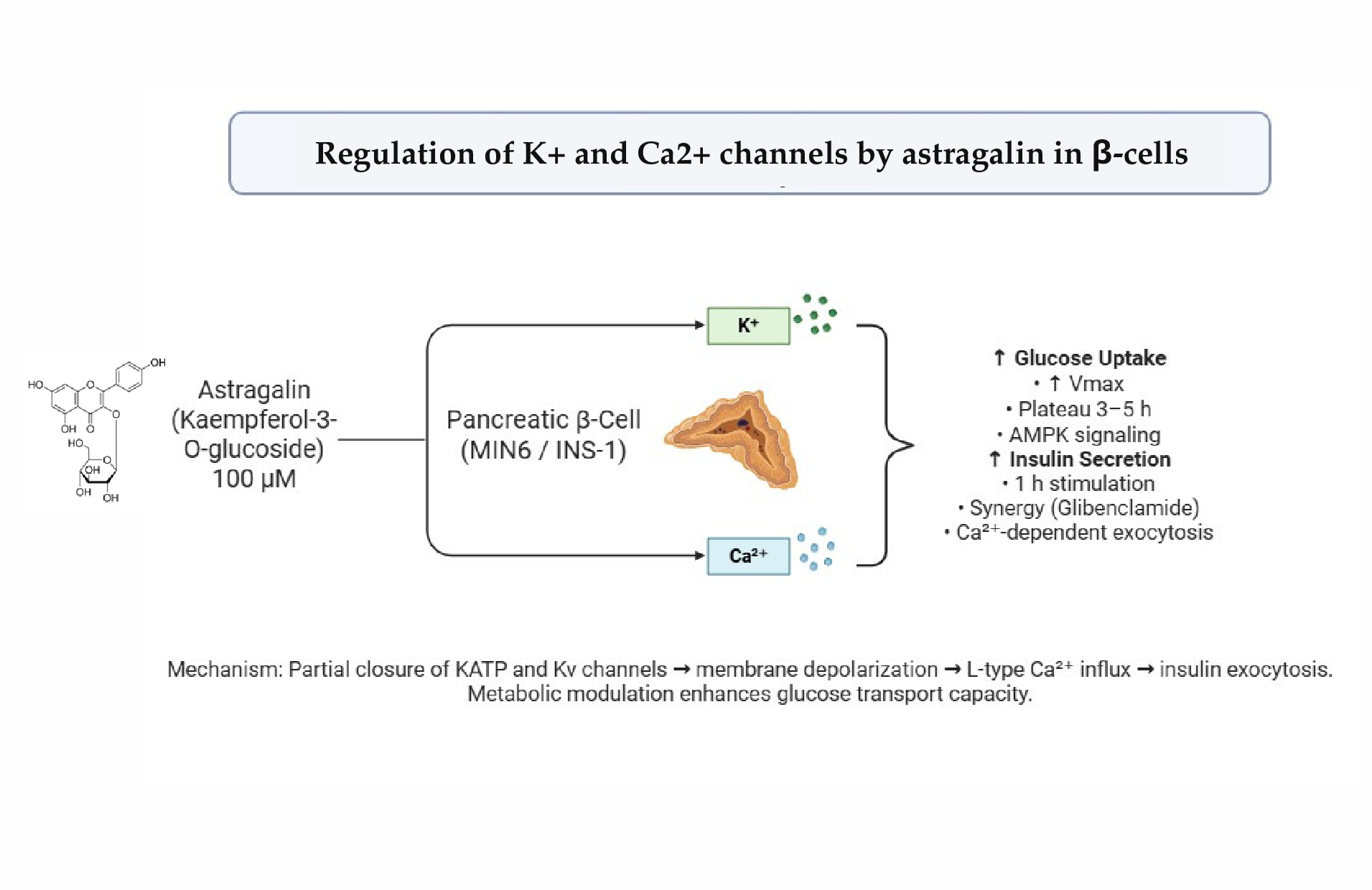
Type 2 diabetes mellitus is marked by chronic hyperglycemia and insulin resistance, leading to progressive tissue damage. Flavonoids such as astragalin have emerged as promising antidiabetic compounds. This study investigated the effects of astragalin on glucose uptake, insulin secretion and ionic mechanisms in pancreatic β-cells (MIN6 and INS-1). Glucose uptake and insulin secretion were quantified by bioluminescence and ELISA, respectively, while ionic currents were assessed by whole-cell patch clamp using selective pharmacological blockers. Astragalin progressively enhanced glucose uptake, reaching a plateau between 3 and 5 h, suggesting improved mitochondrial function and modulation of calcium- and AMPK-dependent signaling pathways. Insulin secretion was significantly stimulated after 1 h of treatment with 100 µM astragalin, involving ATP-sensitive K⁺ channels, voltage-dependent K⁺ channels, and L-type Ca²⁺ channels. Electrophysiological patch-clamp studies showed that astragalin reduces potassium channel currents, indicating partial channel closure and consequent membrane depolarization as corroborated by calcium involvement by using verapamil, an ionic environment associated with insulin exocytosis. These findings suggest that astragalin acts as a metabolic modulator and secretagogue in β-cells coupling insulin-stimulus secretion, representing a potential candidate for antidiabetic therapeutic strategies.
Keywords:
diabetes mellitus
; medicinal plants
; patch-clamp
; kinetics of glucose
; ionic channels
1. Introduction
Type 2 Diabetes Mellitus (T2DM) is a syndrome characterized by chronic hyperglycemia resulting from impaired insulin production and/or function, leading to alterations in glucose, lipid, protein, and other carbohydrate metabolism. This sustained hyperglycemia is the major cause of tissue damage associated with the disease and contributes to insulin resistance [1,2]. Medicinal plants, which form the basis of traditional medicine, are used by approximately 80% of the population in developing countries for the treatment of various diseases, including diabetes [3]. However, the efficacy, safety of phytoconstituents, and the molecular mechanisms underlying their activities remain poorly understood [4,5].
Flavonoids are secondary metabolites widely distributed throughout the plant kingdom and present in the human diet, exhibiting diverse biological activities, including antidiabetic effects [6,7,8]. Among them, astragalin (kaempferol-3-O-β-glucoside), the focus of this study, displays multiple activities described in the literature and has demonstrated antidiabetic efficacy in experimental models by regulating glucose homeostasis and exhibiting metformin-mimetic effects [9,10].
Previous in vivo studies have shown that astragalin rapidly improves glucose tolerance and stimulates insulin secretion. In vitro assays in rat pancreatic islets confirmed this secretagogue effect, involving calcium influx, activation of ATP-dependent potassium channels and voltage-dependent calcium channels, as well as regulation of SERCA pump activity, protein kinase C, and protein kinase A. These mechanisms were evaluated using specific pharmacological agents targeting the high-glucose-induced insulin secretion pathway and employing ⁴⁵Ca²⁺ as a trigger in pancreatic islets [11].
In this context, astragalin was further investigated in one of the assays using electrophysiological approaches, including whole-cell recordings in the INS-1 pancreatic β-cell line, to contribute to the understanding of ionic mechanisms involved in this pathway. Therefore, the overall aim of this study was to characterize the precise targets of astragalin in signaling pathways in MIN6 cells (pancreatic β-cell line) for the regulation of insulin secretion.
2. Results
2.1. Glucose Uptake
This study demonstrated that astragalin stimulates glucose uptake in MIN6 cells, showing a progressive increase followed by a reduction in the rate of uptake between 3 and 5 h (Figure 1A). During the initial linear phase of glucose uptake (0.5–2 h), the average uptake rate was slightly higher with 50 µM astragalin (19.526 ΔLum/h) compared to 100 µM (17.036 ΔLum/h) (Figure 1B).
This initial difference may reflect minor fluctuations in the early measurements or cellular adjustments to the stimulus. As glucose uptake progressed, a plateau was observed between 4 and 5 h, indicating that the uptake rate had stabilized. During the initial uptake period (0.5–5 h), the relative Vmax was numerically higher in cells treated with 100 µM astragalin (66.195 ΔLum) compared with 50 µM (58.469 ΔLum) (Figure 1B), suggesting a concentration-dependent increase in the maximum glucose uptake capacity. The kinetic profile demonstrated an increase in uptake up to approximately 3 h, followed by stabilization.
2.2. Insulin Secretion
In the insulin secretion assay, treatment with astragalin (100 µM, 1 h) significantly increased secretion relative to the control, an effect similar to that observed with glibenclamide. Notably, the combination of astragalin and glibenclamide elicited an even greater insulin secretion than either compound alone.
Figure 2.
Effect of glibenclamide, TEA and verapamil on the stimulatory action of astragalin in insulin secretion by MIN6 cells. Incubation time = 1 h. Data were expressed as mean ± standard error of the mean (SEM), and n = 4 for each tested compound from two independent experiments. Statistically significant differences were indicated by ****p ≤ 0.0001; #p ≤ 0.05 and ###p ≤ 0.001, compared with astragalin group.
Figure 2.
Effect of glibenclamide, TEA and verapamil on the stimulatory action of astragalin in insulin secretion by MIN6 cells. Incubation time = 1 h. Data were expressed as mean ± standard error of the mean (SEM), and n = 4 for each tested compound from two independent experiments. Statistically significant differences were indicated by ****p ≤ 0.0001; #p ≤ 0.05 and ###p ≤ 0.001, compared with astragalin group.
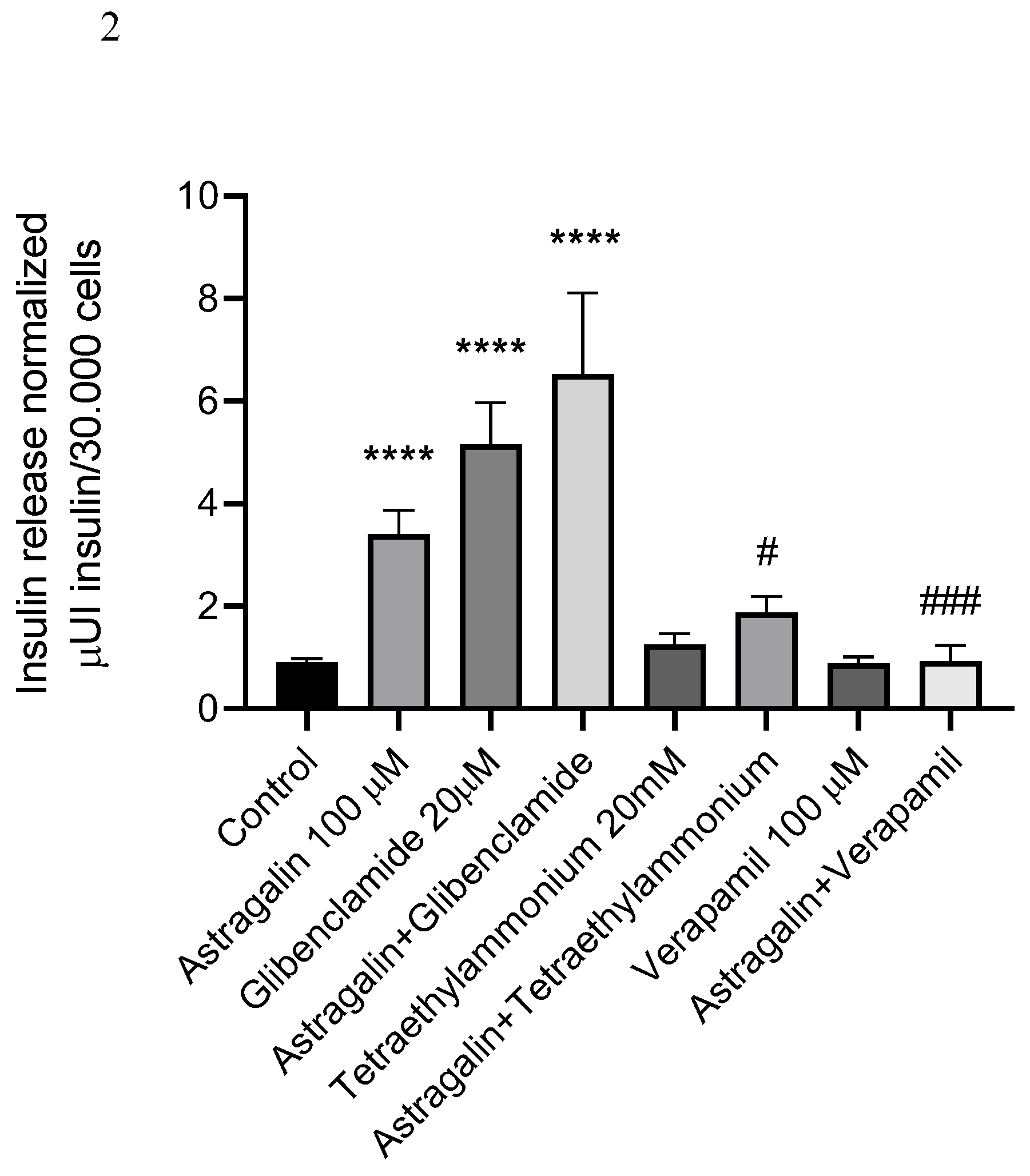
In groups treated with TEA, either alone or in combination with astragalin, a significant reduction in insulin secretion was observed compared to astragalin alone. Similarly, treatment with verapamil, either alone or combined with astragalin, reduced insulin secretion and attenuated the stimulatory effect induced by astragalin.
2.3. Electrophysiology (Patch Clamp) Results
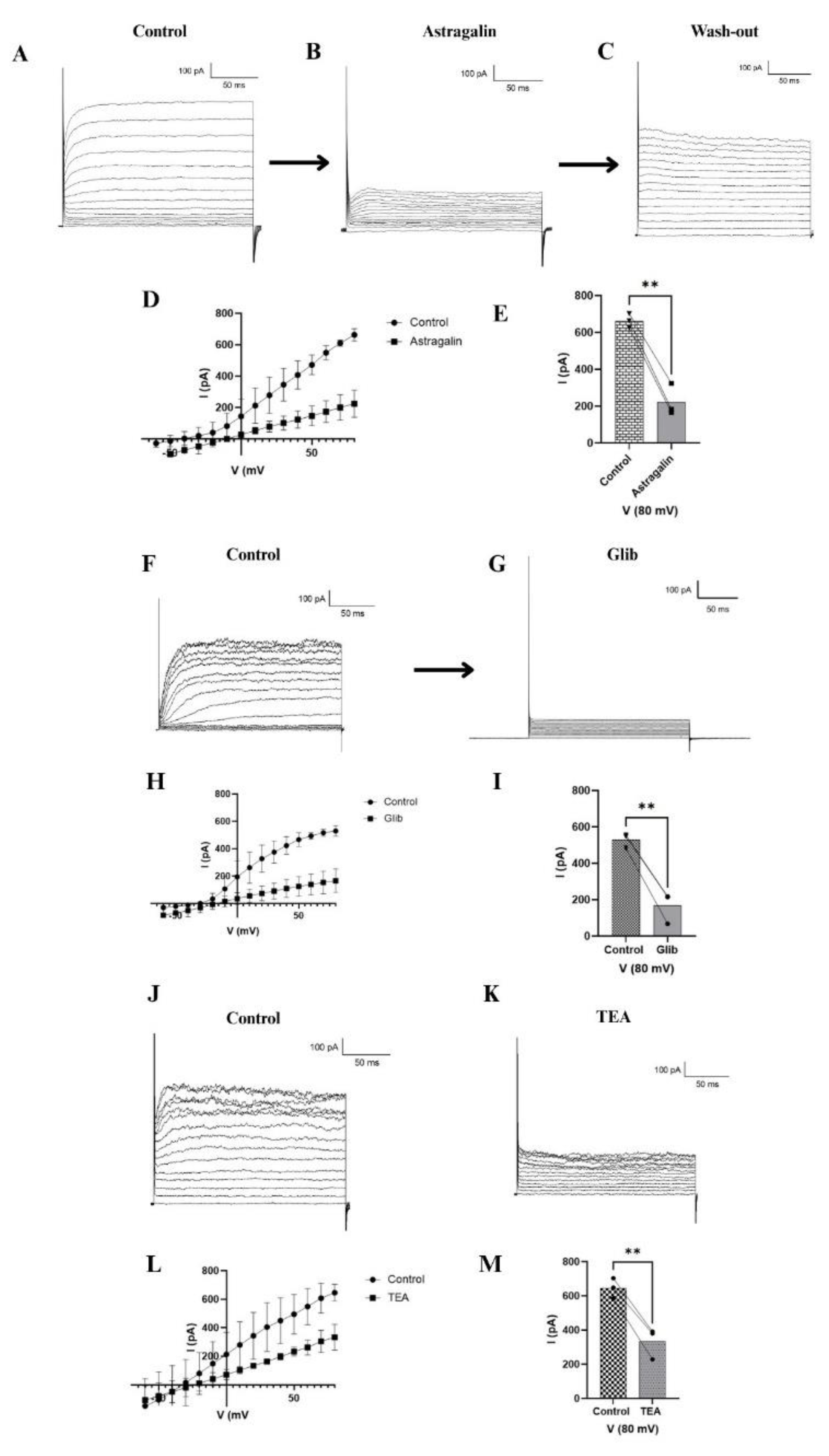
In Figure 3A, it can be observed that once stable access to the cell was achieved via membrane rupture with the micropipette, it was possible to record potassium channel activity, evidenced by an electrical current of approximately 790 pA. After 25 to 30 minutes of perfusion with 100 µM astragalin, a significant reduction in current was observed, reaching approximately 200 pA (Figure 3B). The extracellular solution was perfused again, leading to a partial recovery of the recordings toward a more open channel state (Figure 3C). To confirm the modulation of potassium channels, two specific inhibitors were tested. Glibenclamide reduced the current to approximately 100 pA (Figure 3G), while tetraethylammonium (TEA) reduced it to around 400 pA (Figure 3K). Representative control recordings for these inhibitors are shown in Figure 3F and 3J.
The current–voltage (I–V) relationships presented in Figure 3D, H, and L demonstrate that astragalin, glibenclamide, and TEA all decreased potassium channel conductance compared to the control condition. Statistical analysis performed using one-way ANOVA (n = 3) confirmed that all tested compounds significantly reduced the current (Figure 3E, I, M). Among them, astragalin and glibenclamide exhibited the most pronounced inhibitory effects, followed by TEA.
3. Discussion
The observed increase in glucose uptake suggests that astragalin enhances glucose transport capacity in pancreatic β-cells. Although the initial uptake rate was slightly higher at 50 µM, the higher Vmax observed at 100 µM indicates that astragalin increases the maximum transport capacity in a concentration-dependent manner, particularly during the saturation phase.
Rey et al. [11] reported that, at similar concentration used in this study, astragalin enhanced calcium influx in pancreatic islets and promoted insulin secretion via ATP-dependent potassium channels, L-type calcium channels, and PKA and PKC pathways. Complementarily, Sun et al. [12] showed that astragalin activated the AMPK–PGC-1α pathway in diabetic kidney injury models, stimulated mitochondrial biogenesis and reduced oxidative stress. Together, these findings suggest that the observed increase in glucose uptake in MIN6 cells may reflect both improved mitochondrial function and modulation of calcium- and AMPK-dependent signaling pathways.
MIN6 cells exhibit rapid glucose uptake and high GLUT2 expression, making this system highly sensitive to changes in glucose transport dynamics [13]. Studies using 2-NBDG, a fluorescent glucose analog, confirm GLUT2 predominance, facilitating the investigation of mechanisms modulated by compounds such as astragalin [14].
The kinetic pattern (an increase up to approximately 3 h followed by stabilization) suggests transient mechanisms regulating glucose transport, reinforcing the biological relevance of astragalin.
The secretion of insulin in pancreatic β-cells, such as MIN6 cells, depends on a precise coupling between glucose metabolism, membrane electrical activity, and ionic fluxes, particularly K⁺ and Ca²⁺. K+-ATP channels constitute the central link between metabolic status and membrane potential, whereas voltage-dependent K⁺ channels (Kv) and L-type Ca²⁺ channels (VDCC) refine the duration of the action potential and the pattern of Ca²⁺ entry, ultimately determining insulin granule exocytosis [15,16,17,18]. In this context, our findings indicate that astragalin acts as a potent secretagogue, integrating into multiple stages of the stimulus–secretion coupling cascade.
The effects of astragalin and glibenclamide (a classical sulfonylurea that blocks K+-ATP channels and triggers membrane depolarization followed by the opening of L-VDCC) [16,18], indicate that astragalin does not simply compete for the same binding site as glibenclamide on the K+-ATP channel but likely acts through complementary mechanisms, such as modulating additional ionic channels or components of the exocytotic machinery, in alignment with the multifaceted role of flavonoids in the electrophysiological modulation of excitable cells [19].
The involvement of Kv channels in the action of astragalin is supported by the reduction in insulin secretion observed in the presence of TEA. Considering that TEA is a classical blocker of Kv channels, the results suggest that the secretagogue effect of astragalin depends, at least in part, on the functional integrity of Kv channels. Previous studies have demonstrated that inhibition of Kv2.1 prolongs depolarization, increases Ca²⁺ influx, and enhances insulin secretion [20]. Moreover, recent evidence indicates that Kv channels such as KCNH6 can directly regulate exocytosis through interactions with secretory machinery proteins, including Munc18-1, independently of K⁺ conductance [21].
The result obtained with verapamil further reinforce the critical role of Ca²⁺ in the action of astragalin. Because verapamil blocks L-VDCC, the findings indicate that the secretagogue action of astragalin is dependent on Ca²⁺ entry through these channels. This interpretation is supported by studies demonstrating that CaV1.2 channels are strongly coupled to insulin secretion and that alterations in their expression impair the secretory response [22,23].
In addition, the literature highlights that chronic inhibition of K+-ATP channels, either due to loss-of-function mutations or prolonged exposure to sulfonylureas, can reduce Ca²⁺ sensitivity and lead to β-cell exhaustion [17,24]. In this context, the fact that astragalin modulates multiple pathways, promoting depolarization via K+-ATP channels (in synergy with glibenclamide), prolonging depolarizing events and influencing exocytosis through Kv channels, and amplifying Ca²⁺ influx through L-type channels, suggests a potentially advantageous pharmacological profile, combining a strong secretagogue effect with distributed action across the stimulus–secretion cascade.
The results of electrophysiology indicate that astragalin exerts its effects through the modulation of potassium channels. The partial recovery of the current after washout further supports a direct and reversible modulatory effect on these channels. Both ATP-sensitive and voltage-dependent potassium channels are involved in the insulin secretion process in pancreatic β-cells. Closure of ATP-sensitive potassium channels leads to membrane depolarization, which in turn triggers the opening of calcium channels and subsequent insulin release [25]. Additionally, inhibition of voltage-dependent potassium channels prevents membrane repolarization, prolonging the depolarized state of the cell and thereby sustaining insulin secretion [26]. These findings are consistent with the study by Trezza et al. [27], who demonstrated, using patch-clamp experiments in rat arterial myocytes, that quercetin, the aglycone of astragalin, is also capable of inhibiting K+-ATP channels.
Taken together, these findings provide evidence that astragalin modulates potassium channel activity and directly influences glucose homeostasis in pancreatic β-cells, highlighting its potential as a metabolic modulator and a promising candidate for future studies aimed at developing novel β-cell–targeted antidiabetic therapies.
4. Materials and Methods
4.1. Chemical Compounds
Astragalin, glibenclamide and verapamil were purchased from Sigma-Aldrich (St. Louis, MO, USA), while tetraethylammonium (TEA) was obtained from Alfa Aesar (Ward Hill, MA, USA). Stock solutions of glibenclamide and astragalin were prepared in dimethyl sulfoxide (DMSO), whereas TEA and verapamil were diluted in ultrapure water.
4.2. Cell Culture
MIN6 cells, pancreatic tumor cells derived from Mus musculus, were obtained from the Rio de Janeiro Cell Bank (BCRJ, code 0263). Cells were cultured in low-glucose Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 2 mM L-glutamine, 10% fetal bovine serum (FBS), 100 U/mL penicillin, and 100 μg/mL streptomycin, under standard conditions (37°C, 5% CO₂). Cultures were maintained according to BCRJ guidelines (2022), with passages performed at ~80% confluence. For subsequent experiments, cells in the exponential growth phase (log) were seeded at 50–80% confluence [28].
INS-1 cells are a rat pancreatic β-cell line derived from an insulinoma, widely used as an in vitro model to study insulin secretion, ion channel activity, and β-cell metabolism. (Hohmeier et al., 2000). Cells were obtained from Merck S.A (Polvilho, São Paulo, Brazil) and were cultured in RPMI 1640 medium supplemented with 10% heat-inactivated fetal bovine serum, 1 mM sodium pyruvate, 10 mM HEPES, 50 µM β-mercaptoethanol, and antibiotics (100 U/mL penicillin and 100 µg/mL streptomycin), maintained at 37 °C in a humidified atmosphere with 5% CO₂. This model provides a robust and reproducible system for investigating pancreatic β-cell physiology and pharmacology.
4.3. Glucose Uptake Assay
The glucose uptake assay was performed in MIN6 cells using the bioluminescent Glucose Uptake-Glo™ Assay (Promega, São Paulo, Brazil), based on the detection of 2-deoxyglucose-6-phosphate (2DG6P). Cells were pre-incubated overnight at 37 °C in low-glucose medium, washed twice with PBS, and treated with astragalin (50 or 100 µM), saline solution (negative control), or 2DG6P (positive control). After 30 min, cells were washed again, incubated with 2-deoxyglucose (2DG) for 10 min at 37 °C, and subsequently treated with Stop Buffer, Neutralization Buffer, and the detection reagent. The reaction was incubated for 30 min at 25 °C, followed by luminescence measurement in a microplate reader after an additional 5 h. Results were expressed as fmol/cell/min (n = 3 for each group), according to the manufacturer’s instructions [5]. Statistical analysis was carried out using one-way ANOVA for multiple comparisons. Statistical significance was considered at p ≤ 0.05.
4.4. Insulin Secretion
The insulin concentration was measured using an enzyme-linked immunosorbent assay (ELISA) kit for insulin, based on the sandwich principle. In this method, microtiter plate wells are coated with a monoclonal antibody specific to a unique antigenic site on the insulin molecule. For this purpose, 30.000 MIN-6 cells were added to the insulin ELISA kit plate according to the datasheet instructions. The cells were then incubated for 1 h in Krebs-Hepes buffer containing 5 mM glucose, in the presence and absence of astragalin with activators/inhibitors of ion channels of interest in insulin secretion. Subsequently, a test sample aliquot containing endogenous insulin was incubated in the coated well, where binding occurred with an enzyme conjugate consisting of a biotin-conjugated anti-insulin antibody. After incubation, the unbound conjugate was removed by washing. During the second incubation step, the streptavidin-peroxidase enzyme complex binds to the biotin-anti-insulin antibody. The amount of complex formed is directly proportional to the insulin concentration present in the sample. Upon addition to the substrate solution, the intensity of the developed color reflects the insulin concentration in the sample. Data were expressed as insulin content: μUI insulin/30.000 cells [29]. Statistical analysis was carried out using one-way ANOVA for multiple comparisons. Statistical significance was considered at p ≤ 0.05.
4.5. Patch Clamp Technique
Whole-cell patch clamp electrophysiological recordings were performed on INS-1 cells (rat insulinoma pancreatic β-cells) using a MultiClamp 700B amplifier (Molecular Devices Corp., Berkeley, CA) connected to a Digidata 1440A digitizer and controlled by the pCLAMP 10 software (Molecular Devices Corp., Sunnyvale, CA). Cells were cultured on coverslips pre-treated with poly-L-lysine to enhance adhesion and visualized under an Olympus IX81 inverted microscope integrated into the patch clamp setup. The extracellular solution used during recordings contained: 115 mM NaCl, 5 mM KCl, 1 mM MgCl₂, 1 mM CaCl₂, 3 mM glucose, and 10 mM HEPES (280 mOsm/kg H₂O, pH 7.4). The internal pipette solution, optimized for potassium channel recordings, consisted of: 127 mM KCl, 1 mM CaCl₂, 1 mM MgCl₂, 10 mM EGTA, 10 mM HEPES, and 3 mM ATP (290 mOsm/kg H₂O, pH 7.4). Patch pipettes were fabricated from borosilicate glass capillaries (Sutter, 1.5 × 0.86 mm × 10 cm) using a Micropipette Puller P-1000 (Sutter Instrument Company, Novato, CA). Membrane currents were recorded in response to voltage steps ranging from –60 mV to +80 mV in 10 mV increments. Pipette resistance ranged between 6 and 8 MΩ. Pharmacological assays were conducted by perfusing the cells with glibenclamide (20 nM), TEA (20 mM), verapamil (100 µM) or astragalin (100 µM) [29]. Data analysis was performed using Clampfit 11.4.1 (Molecular Devices Corp., Sunnyvale, CA) and Origin 2025 (OriginLab Corporation, One Roundhouse Plaza, Northampton, MA, USA). Experiments were conducted with n = 4, and statistical analysis was carried out using one-way ANOVA for multiple comparisons. Statistical significance was considered at p ≤ 0.05.
5. Conclusions
In summary, the results of this study demonstrate that astragalin enhances both glucose uptake and insulin secretion in pancreatic β-cells, involving multiple ionic channels and calcium- and AMPK-dependent signaling pathways. The modulation of ATP-sensitive K⁺ channels, voltage-dependent K⁺ channels, and L-type Ca²⁺ channels highlight the multifaceted role of this flavonoid in stimulus–secretion coupling. These findings underscore the potential of astragalin as a metabolic modulator and secretagogue, supporting its relevance as a candidate for novel therapeutic strategies in the management of Type 2 Diabetes Mellitus.
Author Contributions
Conceptualization, F.R.M.B.S., P.M.S.; methodology, P.M.S., B.A.Z., A.L.R.M., P.W.Z.B., K.C., C.R., F.C.C.; software, B.A.Z. and G.B.P.; validation, F.R.M.B.S. and A.L.A.D.; formal analysis, F.R.M.B.S. and A.L.A.D.; investigation, P.M.S., B.A.Z, A.L.R.M., F.C.C., C.R.; resources, F.R.M.B.S.; data curation, P.M.S.; B.A.Z. and A.L.A.D.; writing—original draft preparation, F.R.M.B.S., P.M.S. and B.A.Z; writing—review and editing, F.R.M.B.S., P.M.S., A.L.A.D and B.A.Z.; visualization, P.M.S., G.B.P. and B.A.Z; supervision, F.R.M.B.S. and A.L.A.D; project administration, F.R.M.B.S.; funding acquisition, F.R.M.B.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received FAPESC Grant 2024TR000070 and the support CAPES/PROAP/PPG-BQA.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author(s).
Acknowledgments
P.M.S. and B.A.Z. thank to FAPESC-Pos-doc fellowship nº 735/2024 and Master fellowship nº 61/2024; and F.R.M.B.S. thank to CNPq nº 305891/2023-1 by Productivity grant.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| AMPK | AMP-Activated Protein Kinase |
| ANOVA | Analysis of Variance |
| ATP | Adenosine Triphosphate |
| CaV1.2 | Voltage-Gated Calcium Channel Subtype 1.2 |
| DMEM | Dulbecco’s Modified Eagle Medium |
| DMSO | Dimethyl Sulfoxide |
| ELISA | Enzyme-Linked Immunosorbent Assay |
| FBS | Fetal Bovine Serum |
| GLUT2 | Glucose Transporter Type 2 |
| K⁺-ATP | ATP-Sensitive Potassium Channels |
| Kv | Voltage-Gated Potassium Channels |
| PKA | Protein Kinase A |
| PKC | Protein Kinase C |
| RPMI | Roswell Park Memorial Institute Medium |
| SERCA | Sarco/Endoplasmic Reticulum Ca²⁺-ATPase |
| T2DM | Type 2 Diabetes Mellitus |
| TEA | Tetraethylammonium |
| VDCC | Voltage-Dependent Calcium Channels |
| 2DG | 2-Deoxyglucose |
| 2DG6P | 2-Deoxyglucose-6-Phosphate |
| 2-NBDG | 2-(N-(7-Nitrobenz-2-oxa-1,3-diazol-4-yl)Amino)-2-Deoxyglucose |
References
- Punthakee, Z.; Goldenberg, R.; Katz, P. Definition, Classification and Diagnosis of Diabetes, Prediabetes and Metabolic Syndrome. Canadian Journal of Diabetes 2018, 42, S10–S15. [Google Scholar] [CrossRef]
- Martín, M.Á.; Ramos, S. Dietary Flavonoids and Insulin Signaling in Diabetes and Obesity. Cells 2021, 10, 1474. [Google Scholar] [CrossRef]
- Ranilla, L.G.; Kwon, Y.-I.; Apostolidis, E.; Shetty, K. Phenolic Compounds, Antioxidant Activity and in Vitro Inhibitory Potential against Key Enzymes Relevant for Hyperglycemia and Hypertension of Commonly Used Medicinal Plants, Herbs and Spices in Latin America. Bioresource Technology 2010, 101, 4676–4689. [Google Scholar] [CrossRef]
- Farzaei, F.; Morovati, M.R.; Farjadmand, F.; Farzaei, M.H. A Mechanistic Review on Medicinal Plants Used for Diabetes Mellitus in Traditional Persian Medicine. J Evid Based Complementary Altern Med 2017, 22, 944–955. [Google Scholar] [CrossRef]
- Behl, T.; Gupta, A.; Albratty, M.; Najmi, A.; Meraya, A.M.; Alhazmi, H.A.; Anwer, M.K.; Bhatia, S.; Bungau, S.G. Alkaloidal Phytoconstituents for Diabetes Management: Exploring the Unrevealed Potential. Molecules 2022, 27. [Google Scholar] [CrossRef]
- Cazarolli, L.; Zanatta, L.; Alberton, E.; Reis Bonorino Figueiredo, M.; Folador, P.; Damazio, R.; Pizzolatti, M.; Mena Barreto Silva, F. Flavonoids: Cellular and Molecular Mechanism of Action in Glucose Homeostasis. MRMC 2008, 8, 1032–1038. [Google Scholar] [CrossRef]
- Aragon Novoa, D.M.; Silva, F.R.M.B. The Role of Natural Products on Diabetes Mellitus Treatment; 2024. [Google Scholar] [CrossRef]
- Rey, D.; Fernandes, T.A.; Sulis, P.M.; Gonçalves, R.; Sepúlveda R, M.; Silva Frederico, M.J.; Aragon, M.; Ospina, L.F.; Costa, G.M.; Silva, F.R.M.B. Cellular Target of Isoquercetin from Passiflora Ligularis Juss for Glucose Uptake in Rat Soleus Muscle. Chemico-Biological Interactions 2020, 330, 109198. [Google Scholar] [CrossRef]
- Riaz, A.; Rasul, A.; Hussain, G.; Zahoor, M.K.; Jabeen, F.; Subhani, Z.; Younis, T.; Ali, M.; Sarfraz, I.; Selamoglu, Z. Astragalin: A Bioactive Phytochemical with Potential Therapeutic Activities. Adv Pharmacol Sci 2018, 2018, 9794625. [Google Scholar] [CrossRef]
- Li, Q.; Yang, Z.; Lu, H.; Liu, F.; Zhou, D.; Zou, Y. Astragalin Exerted Hypoglycemic Effect by Both Inhibiting α-Glucosidase and Modulating AMPK Signaling Pathway. Nutrients 2025, 17. [Google Scholar] [CrossRef]
- Rey, D.; Miranda Sulis, P.; Alves Fernandes, T.; Gonçalves, R.; Silva Frederico, M.J.; Costa, G.M.; Aragon, M.; Ospina, L.F.; Mena Barreto Silva, F.R. Astragalin Augments Basal Calcium Influx and Insulin Secretion in Rat Pancreatic Islets. Cell Calcium 2019, 80, 56–62. [Google Scholar] [CrossRef]
- Sun, M.; Ye, H.; Zheng, C.; Jin, Z.; Yuan, Y.; Weng, H. Astragalin Ameliorates Renal Injury in Diabetic Mice by Modulating Mitochondrial Quality Control via AMPK-Dependent PGC1α Pathway. Acta Pharmacol Sin 2023, 44, 1676–1686. [Google Scholar] [CrossRef]
- Niu, B.; Ma, L.; Yao, L.; Zhang, Y.; Su, H. HCV Affects KATP Channels through GnT-IVa-Mediated N-Glycosylation of GLUT2 on the Surface of Pancreatic β-Cells Leading to Impaired Insulin Secretion. Endocrine 2024, 84, 427–440. [Google Scholar] [CrossRef]
- Samario-Román, J.; Velasco, M.; Larqué, C.; Cárdenas-Vázquez, R.; Ortiz-Huidobro, R.I.; Hiriart, M. NGF Effects Promote the Maturation of Rat Pancreatic Beta Cells by Regulating GLUT2 Levels and Distribution, and Glucokinase Activity. PLOS ONE 2024, 19, e0303934. [Google Scholar] [CrossRef]
- Rorsman, P.; Ashcroft, F.M. Pancreatic β-Cell Electrical Activity and Insulin Secretion: Of Mice and Men. Physiological Reviews | American Physiological Society 2017, 98. [Google Scholar] [CrossRef]
- Ashcroft, F.M. KATP Channels and the Metabolic Regulation of Insulin Secretion in Health and Disease: The 2022 Banting Medal for Scientific Achievement Award Lecture. Diabetes 2023, 72, 693–702. [Google Scholar] [CrossRef]
- ElSheikh, A.; Shyng, S.-L. KATP Channel Mutations in Congenital Hyperinsulinism: Progress and Challenges towards Mechanism-Based Therapies. Front. Endocrinol. 2023, 14, 1161117. [Google Scholar] [CrossRef]
- Seino, S. Cell Signalling in Insulin Secretion: The Molecular Targets of ATP, cAMP and Sulfonylurea. Diabetologia 2012, 55, 2096–2108. [Google Scholar] [CrossRef]
- Velasco, M.; Díaz-García, C.M.; Larqué, C.; Hiriart, M. Modulation of Ionic Channels and Insulin Secretion by Drugs and Hormones in Pancreatic Beta Cells. Molecular Pharmacology 2016, 90, 341–357. [Google Scholar] [CrossRef]
- MacDonald, P.E.; Sewing, S.; Wang, J.; Joseph, J.W.; Smukler, S.R.; Sakellaropoulos, G.; Wang, J.; Saleh, M.C.; Chan, C.B.; Tsushima, R.G.; et al. Inhibition of Kv2.1 Voltage-Dependent K+Channels in Pancreatic β-Cells Enhances Glucose-Dependent Insulin Secretion*. Journal of Biological Chemistry 2002, 277, 44938–44945. [Google Scholar] [CrossRef]
- Yang, X.; Yang, Y. The Crucial Role of Potassium Ion Channels in Diabetes Mellitus and Its Complications: A Review. Channels 2025, 19, 2531949. [Google Scholar] [CrossRef]
- Tanaka, Y.; Farkhondeh, A.; Yang, W.; Ueno, H.; Noda, M.; Hirokawa, N. Kinesin-1 Mediates Proper ER Folding of the CaV1.2 Channel and Maintains Mouse Glucose Homeostasis. EMBO Rep 2024, 25, 4777–4802. [Google Scholar] [CrossRef]
- Nitert, M.D.; Nagorny, C.L.F.; Wendt, A.; Eliasson, L.; Mulder, H. CaV1.2 Rather than CaV1.3 Is Coupled to Glucose-Stimulated Insulin Secretion in INS-1 832/13 Cells. 2008. [Google Scholar] [CrossRef]
- York, N.W.; Yan, Z.; Osipovich, A.B.; Tate, A.; Patel, S.; Piston, D.W.; Magnuson, M.A.; Remedi, M.S.; Nichols, C.G. Loss of β-Cell KATP Reduces Ca2+ Sensitivity of Insulin Secretion and Trpm5 Expression. Diabetes 2024, 74, 376–383. [Google Scholar] [CrossRef]
- Proks, P.; Lippiat, J.D. Membrane Ion Channels and Diabetes. Current Pharmaceutical Design 2006, 12, 485–501. [Google Scholar] [CrossRef]
- Thompson, B.; Satin, L.S. Beta-Cell Ion Channels and Their Role in Regulating Insulin Secretion. Comprehensive Physiology 2021, 11, 1–21. [Google Scholar] [CrossRef]
- Trezza, A.; Cicaloni, V.; Porciatti, P.; Langella, A.; Fusi, F.; Saponara, S.; Spiga, O. From in Silico to in Vitro: A Trip to Reveal Flavonoid Binding on the Rattus Norvegicus Kir6.1 ATP-Sensitive Inward Rectifier Potassium Channel. PeerJ 2018, 6, e4680. [Google Scholar] [CrossRef]
- Nakashima, K.; Kanda, Y.; Hirokawa, Y.; Kawasaki, F.; Matsuki, M.; Kaku, K. MIN6 Is Not a Pure Beta Cell Line but a Mixed Cell Line with Other Pancreatic Endocrine Hormones. Endocr J 2009, 56, 45–53. [Google Scholar] [CrossRef]
- Mendes, C.P.; Postal, B.G.; Oliveira, G.T.C.; Castro, A.J.G.; Frederico, M.J.S.; Moraes, A.L.L.; Neuenfeldt, P.D.; Nunes, R.J.; Menegaz, D.; Silva, F.R.M.B. Insulin Stimulus-secretion Coupling Is Triggered by a Novel Thiazolidinedione/Sulfonylurea Hybrid in Rat Pancreatic Islets. Journal Cellular Physiology 2019, 234, 509–520. [Google Scholar] [CrossRef]
Figure 1.
Effect of astragalin on glucose uptake in MIN6 cells. (A) Kinetics of glucose uptake over time (0.5–5 h) at two concentrations of astragalin (50 µM and 100 µM). (B) Glucose uptake plateau (relative Vmax), defined as the average uptake between 4 and 5 h, showing the effect of astragalin concentration on the maximum glucose transport capacity. Data were expressed as mean ± standard error of the mean (SEM), and n = 3 for each tested compound. Statistically significant differences were indicated by *p ≤ 0.05 and **p ≤ 0.01.
Figure 1.
Effect of astragalin on glucose uptake in MIN6 cells. (A) Kinetics of glucose uptake over time (0.5–5 h) at two concentrations of astragalin (50 µM and 100 µM). (B) Glucose uptake plateau (relative Vmax), defined as the average uptake between 4 and 5 h, showing the effect of astragalin concentration on the maximum glucose transport capacity. Data were expressed as mean ± standard error of the mean (SEM), and n = 3 for each tested compound. Statistically significant differences were indicated by *p ≤ 0.05 and **p ≤ 0.01.
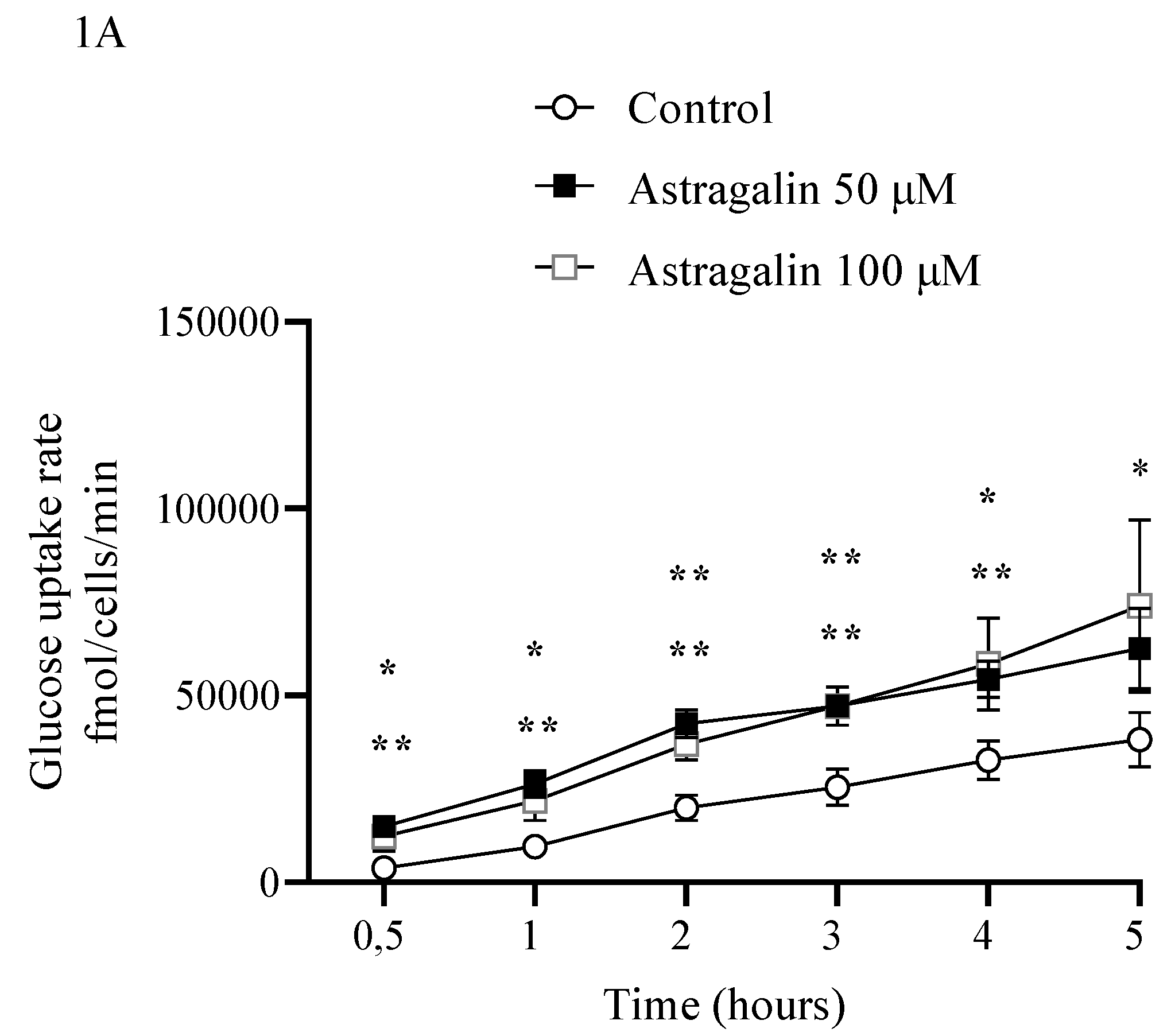
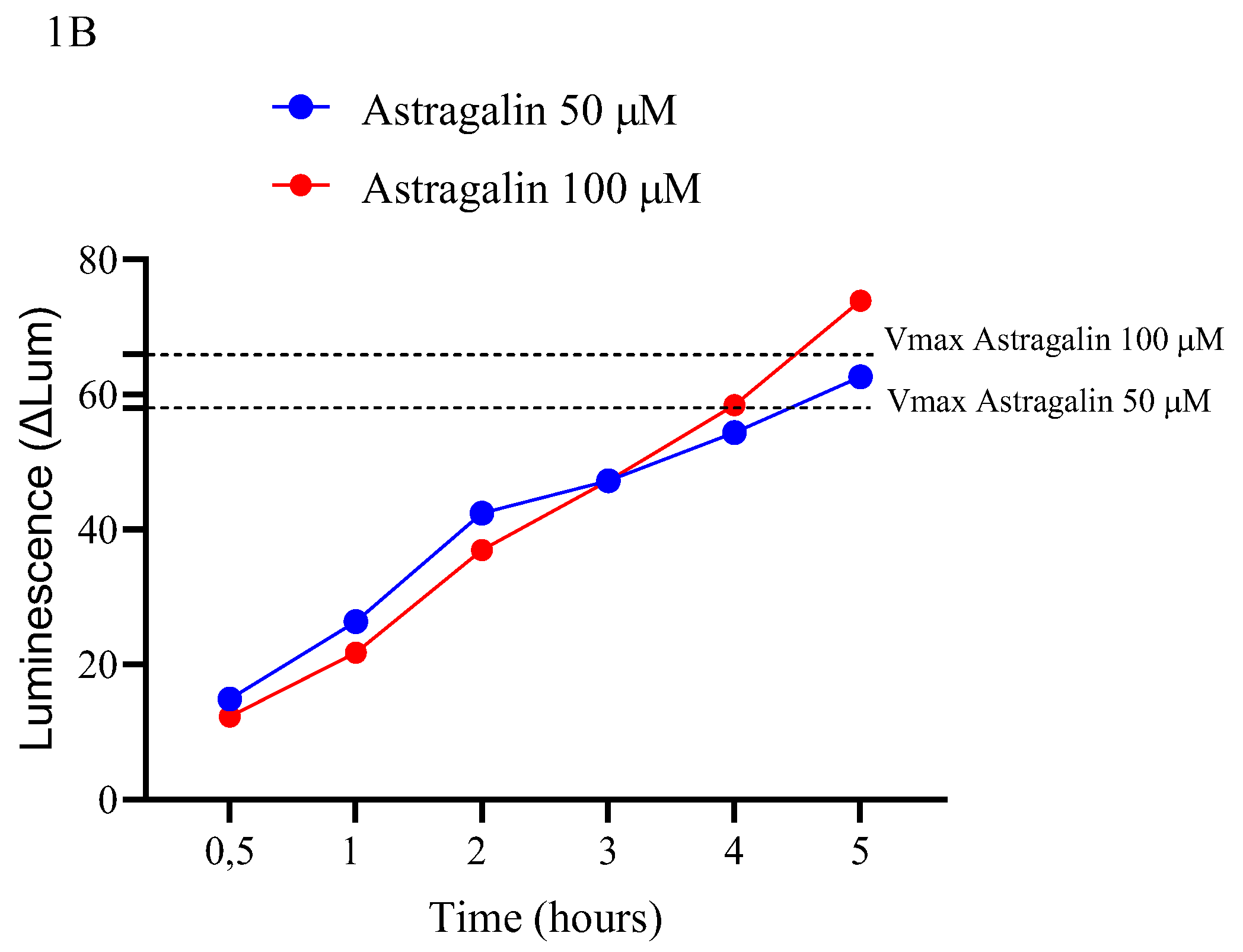
Figure 3.
Results from electrophysiological assays using INS-1 cells. Control currents are shown in Figure A, followed by the effects of astragalin on ionic currents (Figure B), glibenclamide (Figure G), and TEA (Figure K). The representative current–voltage (I–V) relationship is presented in Figure D, H, L. Statistical analysis is shown in Figure E, I, M with data expressed as mean ± standard error of the mean (SEM), and n = 3 for each tested compound. Statistically significant differences were indicated by *p ≤ 0.05 and **p ≤ 0.01.
Figure 3.
Results from electrophysiological assays using INS-1 cells. Control currents are shown in Figure A, followed by the effects of astragalin on ionic currents (Figure B), glibenclamide (Figure G), and TEA (Figure K). The representative current–voltage (I–V) relationship is presented in Figure D, H, L. Statistical analysis is shown in Figure E, I, M with data expressed as mean ± standard error of the mean (SEM), and n = 3 for each tested compound. Statistically significant differences were indicated by *p ≤ 0.05 and **p ≤ 0.01.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



