Submitted:
10 February 2026
Posted:
12 February 2026
You are already at the latest version
Abstract
Feline immunodeficiency virus (FIV) is a lentivirus sharing significant structural and pathological similarities to human immunodeficiency virus (HIV), making it a valuable surrogate model for HIV vaccine design and development. Currently, there is no available effective vaccine could protect cats against FIV infection. This study aims to use some artificial intelligence and immunoinformatic to design a novel multi-epitope DNA vaccine targeting some conserved regions of FIV’s gag, pol, and env genes. The mapped B and T-cell epitopes across the key proteins of the FIV genomes were screened for their ability to trigger strong immune responses, while avoiding allergenic or toxic responses and were linked to the immune adjuvant PADRE. Analysis of the vaccine construct revealed a stable, soluble, and biocompatible vaccine construct with a well-folded tertiary structure capable of binding toll-like receptor 9 (TLR9) and eliciting a robust humoral and cellular immune response. These results demonstrate a promising FIV vaccine candidate with potential insight into future directions in next generation HIV vaccines. Further experimental validation is required to confirm the potential protective power of these putative vaccines in the protection of cats against FIV natural field infection.
Keywords:
feline immunodeficiency virus (FIV)
; human immunodeficiency virus (HIV)
; multi-epitope DNA vaccine
; artificial intelligence
; immunoinformatic
1. Introduction
Human immunodeficiency virus (HIV) remains a major challenge in infectious disease management, largely due to its high mutation rates, which facilitate rapid resistance to current antiretroviral therapy treatments. Resistance-associated mutations generally form in a single host, but can subsequently spread between individuals and enable the spread of therapy-resistant strains among individuals, thereby complicating effective treatment and vaccine development [1,2,3,4]. From a global perspective, this shows a growing health crisis in which antiretroviral therapy resistance is estimated to be the result of up to 10 million deaths annually by 2050 [5].
The growing resistance to current antiretroviral therapies demonstrates the need for alternative approaches to treatment, including next-generation vaccines such as a multi-epitope vaccine [6]. In HIV research, feline immunodeficiency virus (FIV) has served as a valuable surrogate model for comparison. FIV is a lentivirus which cause similar clinical syndrome in cats as those caused by HIV in humans, including progressive immune system deterioration through the decreased concentration of CD4+ T cells [7]. Both viruses share the conserved lentiviral gene products: gag, pol and env which underpin a shared genomic organization and give rise to similar structural and functional features [7,8]. Despite causing less severe clinical disease, FIV’s genetic structure, disease progression, and elicited immune response make it a highly effective model for studying HIV treatment strategies [9,10].
Continuous research on FIV has identified many antiviral factors that make it effective for studying lentiviral immunity and drug resistance [8,11,12]. In addition to this, FIV was the first lentivirus in which antiviral resistance was observed, providing significant insight into the design of HIV therapy [7]. However, effective vaccines for FIV are limited, and the immune mechanisms that protect against mutations are not yet fully understood [13]. With recent advancements in artificial intelligence (AI), there is potential to use AI-based tools to develop a targeted vaccine against FIV efficiently. This can be done by predicting the optimal epitopes, designing protein structures, and predicting the immune system’s response [6,10,14,15,16].
Traditional vaccine development using attenuated or inactivated viruses has been successful for many pathogens, but has proven to be challenging for rapidly mutating viruses [17]. Lentiviruses, such as FIV and HIV, show these challenges due to high mutation rates and integration into the host genome which limit the effectiveness of these vaccines [8]. Advances in artificial intelligence are promising in addressing limitations in traditional vaccine development, such as time and cost and enable rapid screening of datasets to identify epitopes, model structure and ligand-receptor interactions, as well as predict a host's immune response to the vaccine [18,19,20,21]. These advancements in technology offer the potential to facilitate treatment breakthroughs, especially when paired with analogous models like FIV.
The new trend in the design of the next generation vaccine is empowered by the new tools of artificial intelligence and machine learning tools. This approach showed a promising trend in the development and validation of novel vaccines against many viral diseases of humans, animals and birds [13,22,23,24,25,26,27,28]. The first step in this new vaccine pipelines is the selection of the target protein sequences per each virus that convey the protection and prove to induce both humoral and cell mediated immunity. Meanwhile, the most effective vaccine should be prepared against the currently circulating strains of the viral pathogens in the field. Monitoring of the genomic sequences of the key proteins of these viruses will ensure the designed vaccines are matching the homologous filed strains [29,30,31,32,33,34,35,36,37,38,39,40,41].
The present study aimed to identify highly conserved regions (gag, pol, and env) across known FIV genome isolates, which have analogous function to HIV in terms of viral entry, assembly, structure, and replication. The multi-epitope DNA vaccine construct demonstrates high antigenicity, structural stability, and high affinity docking with the body's innate immune defenses (TLR9), and a robust immune response. These results support this vaccine's potential to serve not only as an FIV vaccine candidate, but also as a computational model to inform HIV vaccine development.
2. Materials and Methods
2.1. Retrieval of the FIV Sequences and the Multiple Sequence Alignment (MSA)
Full amino acid sequences of 112 FIV isolates (see supplementary materials S1) were obtained from the NCBI database. Sequences were aligned using Geneious to compare regions of the genes and derive consensus sequences for pol, gag, and env.
2.2. Prediction and Mapping of Some Common T Cell Epitopes Across Some Key Target FIV Proteins
MHC I epitopes were predicted by using amino acid sequences for each gene of the FIV Petaluma strain [M25381.1] in IEDB MHC I Binding Prediction Tool to obtain 9-mers. Characterization of feline MHC I is poor, resulting in the utilization of DLA-8803401. The use of this allele was justified by comparing expressed FLA molecules (XXX) with previously used DLA molecules (XXX). The results indicated high similarity between select FLA molecules and DLA-8803401 [Acc. NM_001014767.1] and was selected for prediction of epitopes. Resulting epitopes were ranked by percentile and with results ≤2.0 being included. Results were further filtered by removing redundant core peptides.
Each gene amino acid sequence of the FIV Petaluma strain was inserted into IEDB MHC II Binding Predictions to obtain 15-mers. The use of human HLA-DR [DRB1*01:01, DRB1*04:01, DRB1*07:01, DRB1*11:01, DRB1*15:01] was justified by utilizing NCBI Blastp to compare the feline MHC class II DRB β1 extracellular domain (FLA-DRB) [Acc. U51527] to homo sapiens reference proteins. The significant sequence homology (Supplementary Table XXX) and previous use in associated studies XXX supported the use of HLA-DRB alleles for MHC II predictions of the feline epitopes. Resulting epitopes were ranked by percentile and with results ≤2.0 being included. Results were further filtered by removing redundant core peptides.
2.3. Prediction and Mapping of Some Common B Cell Epitopes of the Target Viral Proteins
Each gene of the FIV Petaluma strain was inserted into ABCpred to identify linear B-cell epitopes. Results ≥ 0.80 were retained and cross-validated utilizing BepiPred which amino acid utilizes propensity scales to suggest antibody-accessible epitopes. Surface exposure was confirmed utilizing Colabfold to provide the 3D structure of gag, pol, and env. Each of these genes was inserted into UCSF ChimeraX and epitopes exhibiting surface exposure were identified by a solvent-accessible surface area exceeding 300 Å2, indicating sufficient accessibility for immune recognition.
2.3. The Immunological and Safety Profiling of the Mapped FIV Epitopes Across the Key Proteins
T and B cell epitopes were subject to antigenicity, allergenicity, and toxicity testing using VaxiJen v2.0, AllerTOP v2.1, and ToxinPred, respectively. Epitopes were selected based on high antigenicity and a lack of allergenicity and toxicity. The criteria were only epitopes with high antigenicity scores (≥ 0.4), non-allergenic epitopes, and non-toxic epitopes.
2.4. Prediction of the Ability of Mapped T- Cell Epitopes for the Feline Cytokine Production and Cross Reactivity
The MHC II T-cell epitopes were analyzed using IL4Pred to predict the production of the cytokine IL-4. Hybrid prediction method and a SVM threshold of 0.2 were utilized within this server with positive values predicting successful induction. The potential of selected T-cell epitopes to induce IFN-γ was evaluated using the IFNepitope server. Hybrid prediction approach and a threshold of 0.2 were used to identify IFN- γ induction potential. Epitope cross reactivity with non-redundant protein sequences of Felis catus (taxid:9685) was evaluated using NCBI Blastp to assess homology with host proteins.
2.5. In Silico Construction of the FIV Multiepitope DNA Vaccine
All B and T cell epitopes were connected using linkers, along with adjuvants Feline β-defensin 1 and PADRE, to form a multi-epitope construct. Each epitope and adjuvant joined using linkers such as: KK, HE, GSGSG, and GPGPG. At the C-terminal end, a 6-His tag was incorporated into the construct.
2.5. Analysis of the Physicochemical and the Peptide Solubility Properties of the FIV Vaccine Construct
By submitting the FASTA sequence, the transmembrane regions of the protein were predicted using TMHMM-2.0’s standard TMHMM prediction. NETPHOS-3.1 was used to identify the phosphorylation sites, and the protein's hydrophilicity was evaluated using Expasy. Glycosylation sites of the protein were determined using NetNGlyc 1.0.
2.6. Prediction of the Secondary and Tertiary Structures of the Potentially Expressed Proteins
The secondary structure prediction of the peptide utilized NPS, while the tertiary structure of the protein was created using Colabfold and visualized in ChimeraX. The pre-optimized protein received a Z-score through ProSA, and further analysis of the structure was performed by MolProbity. The protein structure was optimized using GalaxyRefine and rerun through both ProSA and MolProbity [42].
2.7. Molecular Docking and Simulation
Toll-like receptor 9 was selected to engage the innate immune system and stimulate antigen presentation (Gupta). The full-length feline TLR9 molecules was obtained from the NCBI database (see supplementary S3) and subject to conversion to a peptide in Expasy, followed by tertiary structure prediction in Colabfold. The docking between the multi-epitope protein TLR9 was simulated using HDock by uploading the PDB file of the sequence with no specific restraints and global docking, with the chemical interactions being interpreted by PDBePISA.
2.8. In Silico Prediction of the Immune Stimulation Simulation of the Designed FIV Vaccine Construct
The peptide's immune response was simulated using C-Immsim. Initial vaccination was administered on day 7 (step 21), followed by boosters at day 21 (step 63) and day 35 (step 105). The immune response was observed through each administration until day 70 (total simulation steps = 1000) [43].
2.9. In Silico Cloning of the Multiepitope FIV Vaccine Construct
The plasmid pcDNA3.1+ was obtained from Addgene, and restriction enzymes EcoRI and XhoI were used to insert the peptide using Snapgene.
2.10. Software

Table 1.
List of the immunoinformatic software utilized for predictions of various epitopes.
| Software | Reference | Software | Reference |
|---|---|---|---|
| Geneious | - | Alphafold | [44,45] |
| ABCpred | [38] | Colabfold | [46] |
| IEDB | [47] | ProSA | [48,49] |
| NCBI Blastp | - | MolProbity | [50] |
| Vaxijenv2.0 | [51] | GalaxyRefine | [52,53] |
| Allertop v2.1 | [54] | HDock | [55] |
| Toxinpred | [56] | PDBePISA | [57] |
| TMHMM-2.0 | [58] | C-Immsim | [59] |
| NETPHOS-3.1 | [60,61] | Addgene | - |
| Expasy [62] | [62] | Snapgene | - |
| NetNGlyc 1.0 | [63] | IL4Pred | [64] |
| NPS | - | IFNEpitope | [65] |
3. Results
3.1. FIV Multiple Sequence Analysis
Viral sequences were collected from the NCBI GenBank. Using Geneious, multiple sequence alignment was performed- highlighting conserved regions of the sequence that could be targeted across multiple isolates. The aligned viral sequences were used to create a consensus sequence for further evaluation and comparison to predicted epitopes of the Petaluma strain [M25381.1]. All valid predicted B and T-cell epitopes are shown in supplementary Table S2.
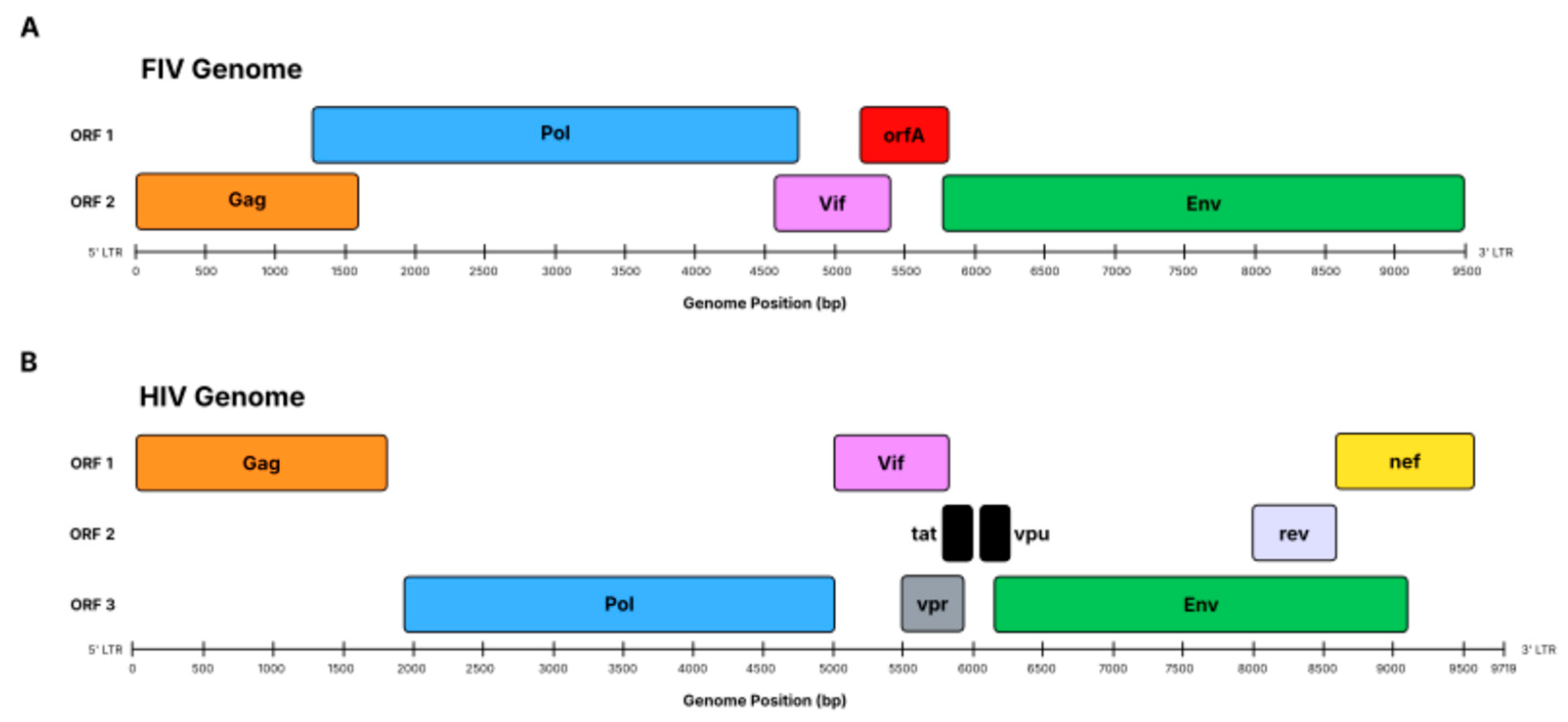
Figure 1.
Comparative genomic structure and organization of the feline immunodeficiency virus (FIV) and the human immunodeficiency virus (HIV). Both viruses share conserved gag, pol, and env genes and genes that regulate replication and immune evasion. The structural and functional similarities highlight why FIV serves as a surrogate model for HIV vaccine design. This schematic was created using Lucidspark and previous organizational models [11,66,67,68].
Figure 1.
Comparative genomic structure and organization of the feline immunodeficiency virus (FIV) and the human immunodeficiency virus (HIV). Both viruses share conserved gag, pol, and env genes and genes that regulate replication and immune evasion. The structural and functional similarities highlight why FIV serves as a surrogate model for HIV vaccine design. This schematic was created using Lucidspark and previous organizational models [11,66,67,68].
3.2. Mapping Some B-Cell Epitope Across the Key Proteins of the FIV Genomes
The gag, pol, and env genes of the FIV Petaluma strain were used to predict linear B-cell epitopes using ABCpred, followed by BepiPred for further verification of antibody accessibility. Protein conformational analysis using Colabfold and UCSF Chimera predicted surface exposed epitopes, where epitopes exhibiting surface exposure greater than 300 Å2 were considered accessible for immune recognition. The epitopes were tested for antigenicity using VaxiJen, with a threshold score of ≥0.4 indicating an epitope as a probable antigen. Allergenicity was predicted using AllerTOP v2.1 and toxicity using ToxinPred. Epitopes were screened against Felis catus proteome (taxid:9685) using NCBI BLAST and epitopes with significant homology to the host protein were excluded. Of the predicted epitopes, the ones with the highest antigenicity values, non-allergenic, non-toxic, and non-cross-reactive ones were chosen for inclusion in the vaccine construct (Table 1). All predicted antigenicity, allergenicity, and toxicity are shown in supplementary Table S2.
Table 2.
Mapping of the B-Cell Epitopes across the key proteins of the FIV genome.
| Gene | Epitope (aa) | Antigenicity | Allergenicity | Toxicity | Surface exposed | Cross- Reactivity |
|---|---|---|---|---|---|---|
| Gag | 161IQTVNGAPQYVALDPK176 | 1.266 | Non- allergen |
Non-toxin | Yes | No |
| Gag | 332VKLYLKQSLSIANANP347 | 0.847 | Non- allergen |
Non-toxin | Yes | No |
| Pol | 744GEGILDKRAEDAGYDL759 | 1.15 | Non- allergen |
Non-toxin | Yes | No |
| Pol | 513GPHQICYQVYQKEGNP528 | 0.742 | Non- allergen |
Non-toxin | Yes | No |
| Env | 298KVNISLCLTGGKMLYN313 | 0.873 | Non- allergen |
Non-toxin | Yes | No |
| Env | 523KAVEMYNIAGNWSCTS538 | 0.813 | Non- allergen |
Non-toxin | Yes | No |
3.3. Mapping and Selection of Some Cytotoxic T Lymphocyte (CD8+) Epitopes Across the Key Proteins of the FIV Genomes
The gag, pol, and env genes of the FIV Petaluma strain were used to predict cytotoxic T lymphocyte (CTL) via the IEDB MHC I-binding Prediction server. The canine allele was selected based on conserved sequence similarity with FLA molecules and previous use in a immunoinformatic study [69]. Analysis of sequence homology is shown in supplementary S3. Epitopes were ranked, with peptides receiving a percentile rank ≤2.0 being retained as well as redundant core peptides being removed. The same parameters as B-cell epitope prediction were used for the prediction of the antigenicity, allergenicity, toxicity and cross reactivity prediction of the CTL epitopes. Chosen epitopes are shown in Table 2, while all valid epitopes are shown in supplementary Table S2.
Table 3.
Results of the prediction of the top ranked CTL epitopes across the key FIPV proteins.
| Gene | Epitope (aa) | Restricting MHC allele | Antigenicity | Allergenicity | Toxicity | Cross- Reactivity |
|---|---|---|---|---|---|---|
| Gag | 55DLQERREKF63 | DLA-8803401 | 1.229 | Non- allergen |
Non-toxin | No |
| Gag | 138RMANVSTGR146 | DLA-8803401 | 1.108 | Non- allergen |
Non-toxin | No |
| Pol | 267SLAVHSLNF275 | DLA-8803401 | 1.053 | Non- allergen |
Non-toxin | No |
| Pol | 397RMLIDFREL405 | DLA-8803401 | 0.967 | Non- allergen |
Non-toxin | No |
| Env | 379RTQSQPGSW387 | DLA-8803401 | 1.365 | Non- allergen |
Non-toxin | No |
| Env | 512YTAFAMQEL520 | DLA-8803401 | 1.085 | Non- allergen |
Non-toxin | No |
3.4. Mapping and Selection of Some Helper T Lymphocyte (CD4+) Epitopes Across the Key Proteins of the FIV Genomes
The genes from the FIV Petaluma strain were input into the IEDB MHC II Binding Prediction server to obtain epitopes with a percentile rank of ≤2.0. MHC II alleles HLA-DR [DRB1*01:01, DRB1*04:01, DRB1*07:01, DRB1*11:01, DRB1*15:01] was justified by utilizing NCBI Blastp to compare the feline MHC class II DRB β1 extracellular domain (FLA-DRB) and results are shown in supplementary S4. Using these predicted epitope sequences, each was then tested for allergenicity, antigenicity, and toxicity. The cytokine induction potential of the MHC II T-cell epitopes were analyzed using IL4Pred and IFNEpitope. All of the selected epitopes demonstrated production of IL4 which suggests the constructs ability to induce an Th2 response [70]. Although only two epitopes were predicted to induce IFN-γ, later immune simulation demonstrated a robust potential to produce IFN-γ. Epitopes used in the construct are shown in Table 3 and full epitope results are available in supplementary Table S2.
Table 4.
Results of the prediction of the top ranked HTL epitopes across the key FIPV proteins.
| Gene | Epitope (aa) | Restricting MHC allele | Antigenicity | Allergenicity | Toxicity | IL-4 | IFN-γ | Cross- Reactivity |
|---|---|---|---|---|---|---|---|---|
| Gag | 134RWAIRMANVTTGREP148 | HLA-DRB1*01:01 | 1.082 | Non- allergen |
Non-toxin | Yes | No | No |
| Gag | 269LTQEQQAEPRFAPAR283 | HLA-DRB1*04:01 | 0.656 | Non- allergen |
Non-toxin | Yes | No | No |
| Gag | 217QLWFTAFSANLTPTD231 | HLA-DRB1*01:01 | 0.843 | Non- allergen |
Non-toxin | Yes | Yes | No |
| Pol | 312FTQNQQWIGPEEAEE326 | HLA-DRB1*07:01 | 0.613 | Non- allergen |
Non-toxin | Yes | No | No |
| Pol | 401DPDYAPYTAFTLPRK415 | HLA-DRB1*11:01 | 0.933 | Non- allergen |
Non-toxin | Yes | No | No |
| Env | 386GSWFRAISSWKQRNR400 | HLA-DRB1*15:01 | 0.459 | Non-allergen | Non-toxin | Yes | Yes | No |
3.5. Selection of the Top Ranked Epitope and the Design of the FIV-DNA Vaccine Construct
The 18 chosen epitopes were equally distributed across B, HTL, and CTL results (6 from each). The 6 chosen epitopes consisted of the most immunogenic sequences and represented each gene equally for both CTL and B-cell epitopes. Due to limited valid epitopes, only one HTL env epitope was used, with replacement by the next highest ranked pol epitope. Highly immunogenic epitopes were linked using EAAAK, GPGPG, AAY, and KK. PADRE was added to the N-terminus of the peptide as an adjuvant to stimulate a helper T-cell response and increase vaccine potency [71]. A histidine tag was then added at the C-terminus to facilitate protein purification and detection. The full construct is shown in Figure 2.
3.6. The Physicochemical Properties of the Designed FIV-DNA Vaccine Construct
The physicochemical and solubility properties of the vaccine construct were assessed to evaluate suitability for stability, expression, and immunogenic performance. Expasy Protparam results indicated the construct has a molecular weight of ~35.4 kDa (320 amino acids) and a basic pl of 9.91 indicating interaction with negatively charged immune cell membranes. Protparam also indicated a hydrophilic GRAVY score of -0.6 and a stable protein value of 29.71. Full analysis in supplementary S4. DeepTMHMM2.0 probabilities predicted that there are no transmembrane regions in the vaccine protein, with residues classified as extracellular. This supports the construct being non-membrane bound and demonstrates potential accessibility for antigen-processing [72] (Figure 3A).
NetPhos-3.1 indicated 48 phosphorylation sites distributed across residues of the construct (Figure 3B). The numerous predicted phosphorylation sites indicate that the vaccine protein may have increased immunogenicity via activation of antigen presentation by innate immune cells, increased activation of T-cells, and proliferation of B-cells [73]. The analysis by Expasy revealed that the vaccine protein is primarily hydrophilic by nature and dissolvable in water, with a hydrophilic peak of -3.1 and a hydrophobic peak of 1.5 (Figure 3C). The predicted high solubility indicates antigen dissolution and presentation to the immune system [38]. NetNGlyc 1.0 predicted four glycosylation sites that marginally exceed the threshold of 0.5; indicating the protein may not have any glycan-mediated epitope masking. (Figure 3D) [74].
3.7. Results of the Predicted Antigenicity and Allergenicity of the FIV-DNA Vaccine Construct
Utilizing VaxiJen 2.0, the antigenicity of the vaccine peptide with adjuvants was estimated at 0.7192. When the software was used solely with the peptides (without adjuvants), the antigenicity was estimated to be 0.7288. This shows that the vaccine, with or without adjuvants, is antigenic. AllerTOP 2.1 was used on the vaccine with and without adjuvants and was confirmed to be non-allergenic. The vaccine was confirmed to be non-toxic, using ToxinPred, in its complete and adjuvant-free forms as well.
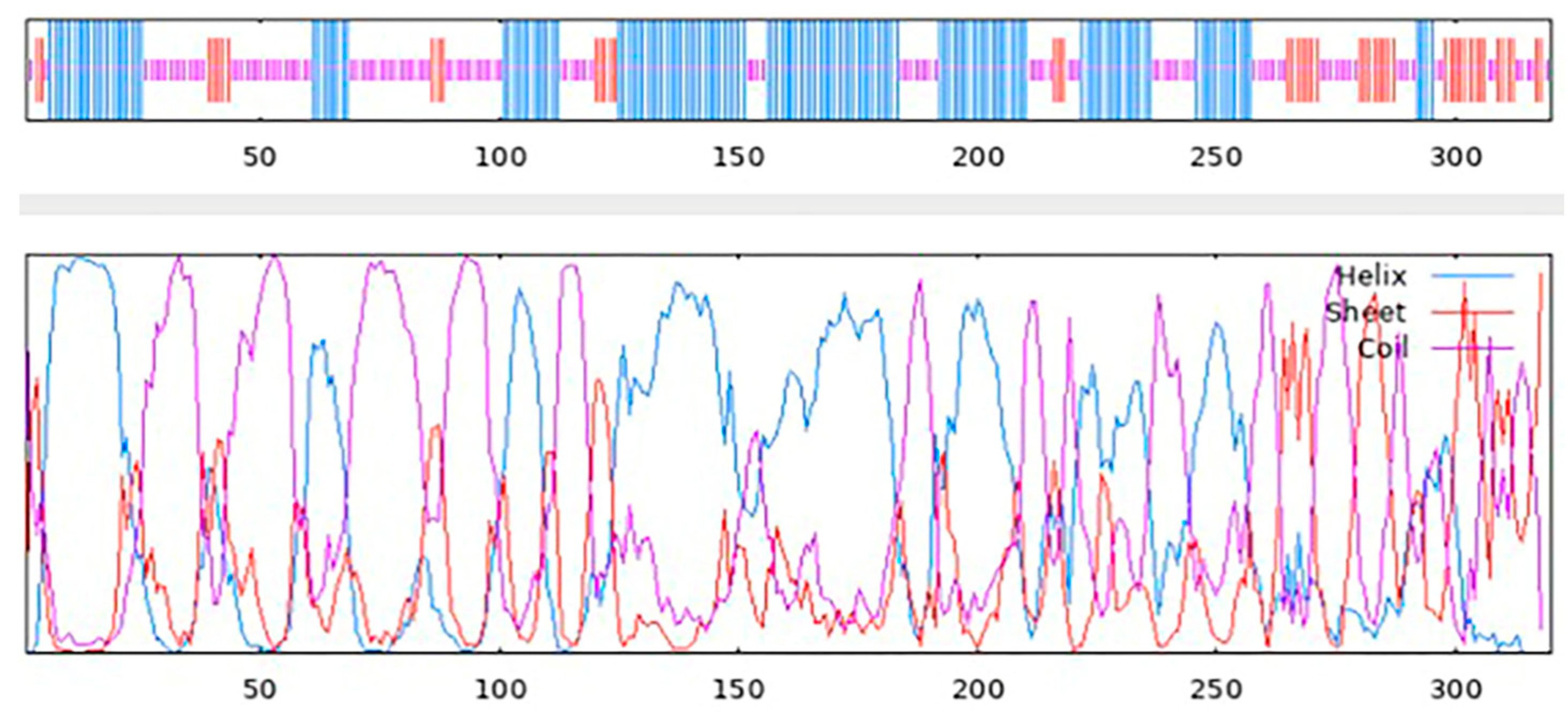
3.8. The Secondary Structure of the Multiepitope FIV-DNA Vaccine Construct
The NPS online software was used to examine the protein's secondary structure, indicating the composition to be 45.31% alpha helix, 15% extended strands, and 39.69% random coils (Figure 3). This suggests the construct is partially ordered due to the alpha helix content, but conformationally adaptable based on the coils and strands [75].
Figure 4.
Secondary structure analysis of protein, where alpha helix is represented by helix, extended strands are represented by sheet, and random coils are represented by coil.
Figure 4.
Secondary structure analysis of protein, where alpha helix is represented by helix, extended strands are represented by sheet, and random coils are represented by coil.
3.9. The Tertiary Structure of the Multiepitope FIV-DNA Vaccine Construct
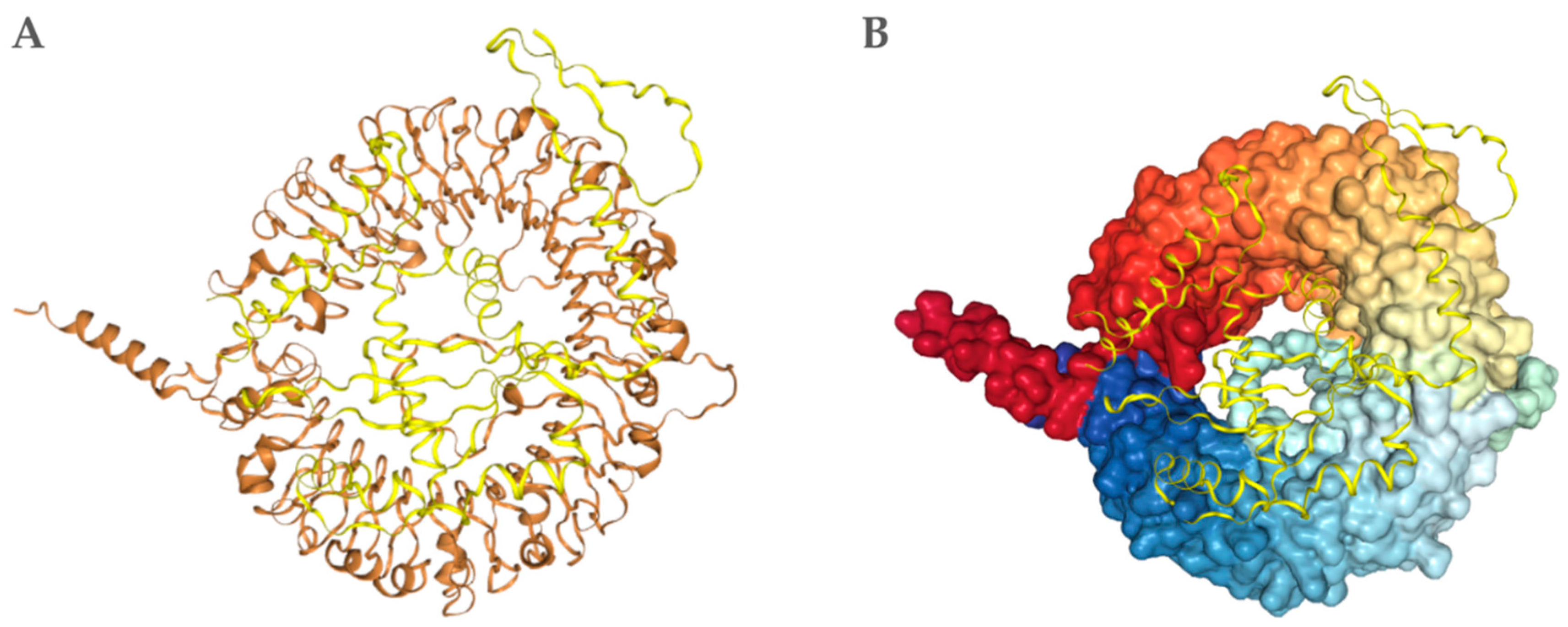
The predicted tertiary structure of the protein was created using Colabfold and the highest-scoring model returned was chosen for further visualized in ChimeraX (Figure 5A). The pre-optimized protein received a Z-score of -9.05 through ProSA (Figure 5B) and further analysis of the structure was performed by MolProbity, which analyzed the Ramachandran plots (Figure 5C) to show 66.04% of the protein had favorable geometry. The protein structure was optimized using GalaxyRefine and rerun through both ProSA and MolProbity. The resulting Z-score was -3.28, with a significantly improved 97.48% favored protein geometry and two Ramachandran outlier. The refinement substantially improved geometry and quality of the construct and enabled further analysis.
3.10. Results of the Molecular Docking with Ligand-Binding Domain of the Feline TLR9
The refined tertiary structure of the vaccine construct was validated using ProSA prior to docking analysis. The vaccine construct and the binding domain of feline toll-like receptor (TLR9) was then submitted to HDock for docking, followed by PDBePISA for post-docking analysis. HDock provided ten predicted complexes, model 1 presented with the lowest docking energy (-304.77) and the highest confidence score (0.9567), indicating a favorable binding orientation. PDBePISA predicted a total interface area of 1,933 Å2 and a ΔG of -8.7 kcal/mol upon complex formation which suggests a energy-favorable interaction between the construct and TLR9. Structural analysis showed 9 hydrogen bonds and 3 salt bridges forming between the ligand and TLR9, allowing a strong non-covalent interaction between the pair. The following are the binding residues between the construct and TLR9: Hydrogen bonds: A:GLN367–B:TYR172, A:ARG601–B:VAL211,A:ARG656–B:ALA214,A:ARG726–B:TYR217, A:GLN38–B:LYS10,A:ASN309–B:ARG142,A:SER365–B:ARG149, A:MET311–B:ARG179. Salt bridges: A:ARG338–B:GLU180.
See supplementary S5 and S6 for additional TLR9 docking and analysis information.
Figure 6.
Molecular docking visualization of the FIV-DNA vaccine construct with the feline TLR9. (A) The receptor (brown) is shown in ribbon configuration, while the vaccine construct (yellow) is shown in ribbon confirmation occupying the receptor binding domain of TLR9. (B) Surface representation of TLR9 colored by structural domains, highlighting the bonding interface with the ligand (yellow).
Figure 6.
Molecular docking visualization of the FIV-DNA vaccine construct with the feline TLR9. (A) The receptor (brown) is shown in ribbon configuration, while the vaccine construct (yellow) is shown in ribbon confirmation occupying the receptor binding domain of TLR9. (B) Surface representation of TLR9 colored by structural domains, highlighting the bonding interface with the ligand (yellow).
3.11. The Immune Stimulation Results of the Designed Multiepitope FIV-DNA Vaccine Construct
The C-ImmSim server was used to simulate how the immune system would respond after three administrations 14 days apart of the vaccine throughout the course of 70 days. The B-cell population (Figure 7A) showed a rapid increase in total population and memory B-cells following initial administration on day 7 and with subsequent boosters on day 21 and day 35. The sustained memory B-cell population and class switching between IgM and IgG suggests an effective immune response and enhanced antigen presentation. Production of antibodies, mainly IgM and IgG (Figure 6B), showed the potential to evoke long lasting immunity to the vaccine construct. Substantial activation of T-helper cells (Figure 6C) as well moderate activation of cytotoxic T-cells (Figure 6D) suggests a robust CD4+ immune response with a gradual increase in CD8+ cells over time. Activation of cytokines IL-2, IL-4, and IL-10 indicate a moderate Th2-mediated immune response (Figure 6E). Strong IFN-γ predictions indicate that the response is Th1 skewed. The activation of both cellular and humoral immunity supports the vaccine constructs potential to evoke an immune response in vivo.
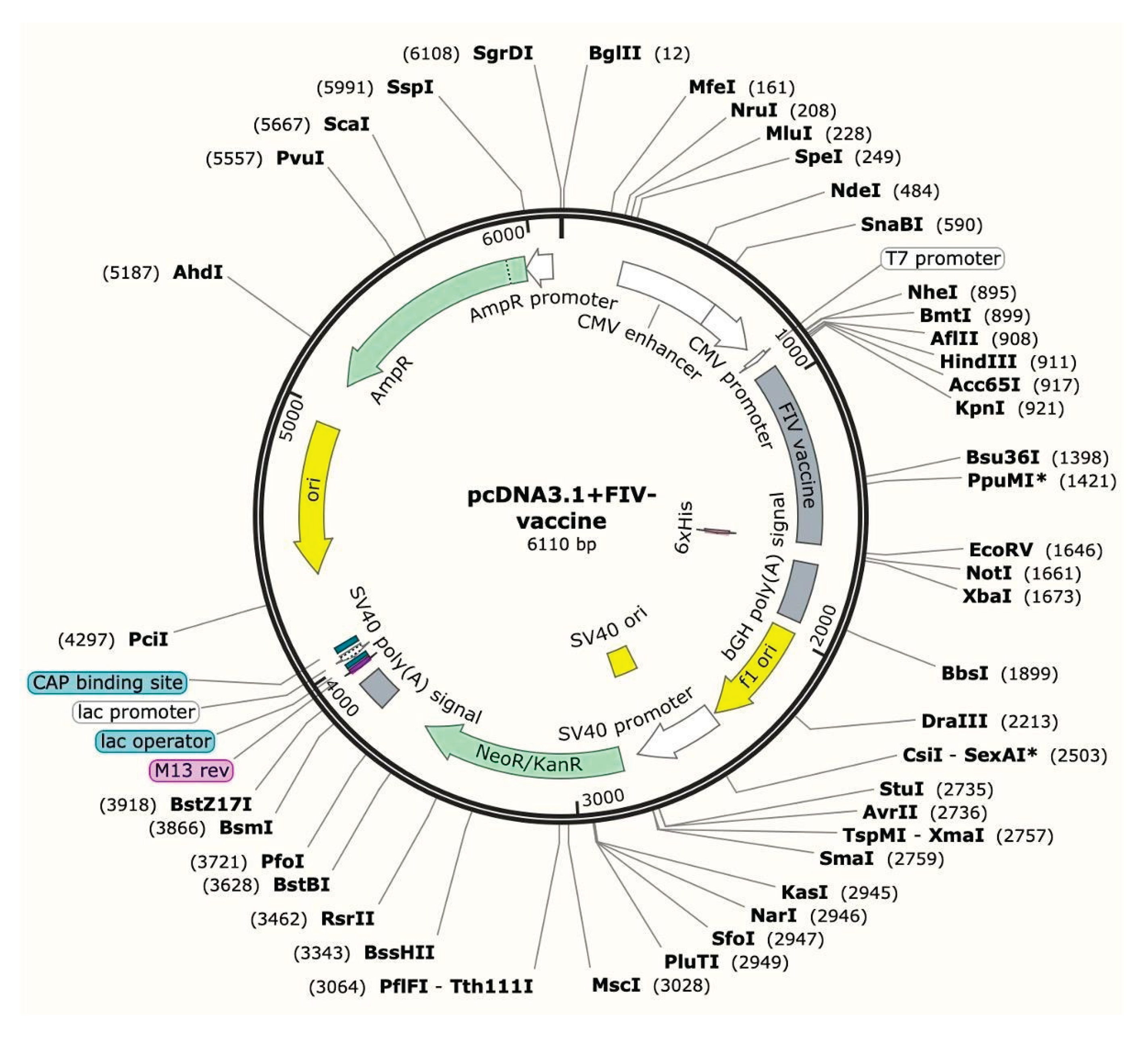
3.9. In Silico Cloning Results of the Final Recombinant Vaccine Construct
The final multi-epitope vaccine construct was cloned in silico into the mammalian expression vector pcDNA3.1(+) vector using EcoRI and Xhol restriction sites. The recombinant plasmid was designed and verified in SnapGene to ensure proper insertion into the expression vector.
Figure 8.
In silico cloning of the multi-epitope vaccine construct into the pcDNA3.1(+) mammalian expression vector. The FIV-multiepitope DNA construct was inserted between the EcoRI and XhoI restriction sites. .
Figure 8.
In silico cloning of the multi-epitope vaccine construct into the pcDNA3.1(+) mammalian expression vector. The FIV-multiepitope DNA construct was inserted between the EcoRI and XhoI restriction sites. .
4. Discussion
This study aimed to use artificial intelligence-assisted immunoinformatics to develop a multi-epitope DNA vaccine targeting conserved regions of feline immunodeficiency virus (FIV) gag, pol, and env genes. Utilizing 112 FIV isolates, the vaccine construct was able to effectively target genes of known circulating isolates to maximize coverage of this vaccine. The developed vaccine construct showed physicochemical and structural stability, with high antigenicity and a tertiary structure suitable for downstream analysis. The construct’s capability to produce a robust humoral and cellular immune response in silico was demonstrated by immune simulation with a prime-booster vaccination strategy, yielding strong antibody production with class switching, sustained helper T-cell responses, a significant cytotoxic T-cell response, and a cytokine profile indicative of effective activation of the immune system.
The immune stimulation findings suggest that the vaccine preferentially promotes cell-mediated immunity with a strong CD8+ T-cell response associated with the robust IFN-γ response to each booster. This cellular immune profile is particularly important for combating retroviral infections, such as FIV and HIV, that are able to integrate into host genomes rapidly. As such, neutralizing antibodies will reduce viral entry and establishment of infection by early recognition of the virus, but a cytotoxic response is necessary for clearance of the rapidly infected cells and is crucial for effective immune control of retroviruses [12,76,77]. FIV and HIV vaccine literature emphasize the importance of durable viral control being correlated with the magnitude of the virus-specific CD8+ T-cell response, with IFN-γ playing a central role in promoting the cytotoxic response [4,12,78]. Further vaccine research on FIV demonstrates anti-FIV T-cell immunity is crucial for protection against FIV, and possibly HIV-1 [77].
The cytokine profile, predominant in IL-2, IL-4, and IL-10, demonstrate the induction of an effective Th2-mediated humoral immunity necessary for antibody production, maturation, and immune memory, as identified in previous multiepitope vaccine strategies [70,79]. The vaccines production of persistent memory B cells, with antibody production and IgM to IgG class switching, is a hallmark of successful vaccination strategies [70,79,80]. The inclusion of PADRE likely further supported Th2-cell activation and antigen presentation, consistent with its role in previously described multiepitope vaccines [81,82]. Together with the strong CD8+ T-cell response, these results show the induction of both humoral and cellular immunity and support the vaccine’s potential to induce an effective immune response.
The design strategy used in this study closely follows framework of previous immunoinformatic vaccines targeting viral pathogens, including feline infectious peritonitis, infectious bronchitis virus, human papillomavirus, cyprinid herpesvirus. As in these studies, the inclusion of epitopes from multiple proteins aids in broadening immune coverage and reduce the likelihood of escape [42,69,83,84]. Lentiviral and retroviral vaccine designs are often unable to maintain effectiveness due to the large amount of antigenic diversity and mutations occurring. To combat this, 18 epitopes were selected targeting the more conserved epitopes from gag and pol genes, allowing a durable immune response that is less dependent on virus strain, while the env-derived (envelope) epitopes are able to elicit antibody production despite antigenic diversity. The balance of conserved and variable gene targets has also been described in feline immunodeficiency and immunodeficiency-like virus studies [85,86].
Immunological and safety screening is another advantage of immunoinformatic-driven vaccine design. Filtering all epitopes according to antigenicity, allergenicity, toxicity, and cross-reactivity allows for early removal of possibly harmful proteins. Selection based on these requisites has been identified as a critical step in improving translational feasibility and in reducing adverse immune outcomes [51].
The structural and physicochemical analysis further supports the feasibility of this vaccine construct. The immunoinformatic utilized indicated that the construct is stable, soluble, and suitable for immune recognition, while also maintaining a well-folded refined tertiary structure. Proper folding has been shown to be critical for epitope recognition and processing by the innate immune system, a step that is critical for downstream activation of adaptive immunity and successful vaccination [42,69]. The vaccines stable and accessible structure provides a reliable foundation for evaluating its interaction with a receptor, TLR9.
The multi-epitope FIV-DNA vaccine construct demonstrated a stable and energetically favorable interaction with toll-like receptor 9 (TLR9), supported by the formation of hydrogen bonds and salt bridges. Stimulation of this receptor has been shown to play a role in innate antiviral responses in felines [87,88], therefore supporting the previously mentioned downstream activation of adaptive immunity. These results demonstrate a mechanistic basis for this vaccine inducing an immune response in felines.
Several limitations of this study should be acknowledged, many of which are common among immunoinformatic-based multi-epitope vaccine design. Expression of this vaccine in vivo cannot be fully accounted for through computational predictions, as post-translational modifications, host immune system variability, biological complexities, and many other factors contribute to the expression of a vaccine in vivo. Epitope selection, structural behavior, receptor interactions, and immune responses are thus largely host-dependent and the computational findings in this study only serve as preliminary evidence for experimental validation. Similar constraints have been noted in multiepitope vaccine studies [42,69,84].
Future studies should experimentally validate the proposed vaccine construct, beginning with in-vitro experimentation to evaluate the proteins stability, safety and initial efficacy. Further in-vivo studies can provide stronger evaluation of safety, immune response, and the vaccines effectiveness in a complex system. The vaccine’s potential would then be defined by assessing dosing strategies particularly against various FIV strain challenges. The application of immunoinformatic for vaccine research can be used to rapidly update and refine current vaccines as new isolates emerge, particularly in the case of HIV, as identifying epitopes and testing many properties can be streamlined using this approach.
Conclusions
In conclusion, this study demonstrates that an immunoinformatic driven approach to vaccine design can produce a stable and effective vaccine targeting FIV in silico. The favorable structural characteristics and strong immune response may indicate this FIV-DNA vaccine construct’s ability to produce effective immunity in-vivo. As for rapidly mutating retroviruses like HIV, this approach shows rapid in silico methods of targeting multiple regions of consistently evolving viruses and may provide an effective platform for design of future treatments.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
TM: Methodology, data analysis, writing first draft, writing final draft ; AS; Methodology, data analysis, writing first draft, writing final draft; TL; Methodology, data analysis, writing first draft, writing final draft ; MGH: Conceptualization, methodology, data analysis, funding, writing first draft, writing final draft.
Funding
This study was supported by Long Island University (LIU) (Grant No. 40262), with financing provided by Helaine Lerner and Joan Rechnitz, as part of the Heilbrunn Family College of Veterinary Medicine Research and Scholarship Fund (Research Project #1) at LIU.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable to this article.
Acknowledgments
We thank Fiona Reagan for her technical assistance.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Pennings, P.S., HIV drug resistance: problems and perspectives. Infectious Disease Reports 2013, 5(11), e5.
- Hill, A.L.; Rosenbloom, D.I.S.; Nowak, M.A. Evolutionary dynamics of HIV at multiple spatial and temporal scales . Journal of Molecular Medicine 2012, 90(5), 543–561. [Google Scholar] [CrossRef] [PubMed]
- Pang, X. Emergence of HIV-1 drug resistance mutations among children and adolescents undergoing prolonged antiretroviral therapy in Guangxi . Journal of Global Antimicrobial Resistance 2024, 37, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Barouch, D.H. Challenges in the development of an HIV-1 vaccine . Nature 2008, 455(7213), 613–619. [Google Scholar] [CrossRef] [PubMed]
- Cedeño-Muñoz, J.S. Antibiotic resistant bacteria and antibiotic resistance genes as contaminants of emerging concern: Occurrences, impacts, mitigations and future guidelines . Science of The Total Environment 2024, 952, 175906. [Google Scholar] [CrossRef]
- Hussaini, I.M. Unveiling the arsenal against antibiotic resistance: Antibacterial peptides as broad-spectrum weapons targeting multidrug-resistant bacteria . The Microbe 2024. 5, 100169. [Google Scholar] [CrossRef]
- Asquith, C.R.M. Synthesis and comparison of substituted 1,2,3-dithiazole and 1,2,3-thiaselenazole as inhibitors of the feline immunodeficiency virus (FIV) nucleocapsid protein as a model for HIV infection . Bioorganic & Medicinal Chemistry Letters 2019, 29(14), 1765–1768. [Google Scholar] [CrossRef]
- Laitinen, T. Synthesis and evaluation of 1,2,3-dithiazole inhibitors of the nucleocapsid protein of feline immunodeficiency virus (FIV) as a model for HIV infection . Bioorganic & Medicinal Chemistry 2022, 68, 116834. [Google Scholar] [CrossRef]
- Evermann, J.F.; Kennedy, M.A. Chapter 16 - Viral Infections, in Small Animal Pediatrics; Peterson, M.E., Kutzler, M.A., Eds.; W.B. Saunders: Saint Louis, 2011; pp. 119–129. [Google Scholar]
- Weaver, J.L. 11.32 - Immunomodulation and the Risk for Neoplasia, in Comprehensive Toxicology (Third Edition); McQueen, C.A., Ed.; Elsevier: Oxford, 2018; pp. 717–731. [Google Scholar]
- Miller, C. Applications of the FIV Model to Study HIV Pathogenesis . Viruses 2018, 10(4). [Google Scholar] [CrossRef]
- Burkhard, M.J.; Dean, G.A. Transmission and immunopathogenesis of FIV in cats as a model for HIV . Curr HIV Res 2003, 1(1), 15–29. [Google Scholar] [CrossRef]
- Mary Jo, B.; Gregg, A.D. Transmission and Immunopathogenesis of FIV in Cats as a Model for HIV . Current HIV Research 2003, 1(1), 15–29. [Google Scholar] [CrossRef]
- Bhattacharya, M. A novel mutation-proof, next-generation vaccine to fight against upcoming SARS-CoV-2 variants and subvariants, designed through AI enabled approaches and tools, along with the machine learning based immune simulation: A vaccine breakthrough . International Journal of Biological Macromolecules 2023, 242, 124893. [Google Scholar] [CrossRef]
- Vaghasiya, J.; Khan, M.; Bakhda, T. Milan. A meta-analysis of AI and machine learning in project management: Optimizing vaccine development for emerging viral threats in biotechnology . International Journal of Medical Informatics 2025, 195, 105768. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, B. Chapter Five - The power of AI in viral vaccine production: A paradigm shift in efficiency and costs . In Methods in Microbiology; Srivastava, A., Mishra, V., Eds.; Academic Press, 2025; pp. 125–146. [Google Scholar]
- Jorge, S.; Dellagostin, O.A. The development of veterinary vaccines: a review of traditional methods and modern biotechnology approaches . Biotechnology Research and Innovation 2017, 1(1), 6–13. [Google Scholar] [CrossRef]
- Khalaf, W.S.; Morgan, R.N.; Elkhatib, W.F. Clinical microbiology and artificial intelligence: Different applications, challenges, and future prospects . Journal of Microbiological Methods 2025. 232-234, 107125. [Google Scholar] [CrossRef] [PubMed]
- Olawade, D.B. Leveraging artificial intelligence in vaccine development: A narrative review . Journal of Microbiological Methods 2024, 224, 106998. [Google Scholar] [CrossRef]
- Kannan, S.; Subbaram, K.; Faiyazuddin, M. Chapter 17 - Artificial intelligence in vaccine development: Significance and challenges ahead, in A Handbook of Artificial Intelligence in Drug Delivery; Philip, A., et al., Eds.; Academic Press, 2023; pp. 467–486. [Google Scholar]
- Malhotra, Y. Chapter Eight - AI in infectious disease diagnosis and vaccine development, in Methods in Microbiology; Srivastava, A., Mishra, V., Eds.; Academic Press, 2025; pp. 191–218. [Google Scholar]
- Al-Kubati, A.A.G. Immunoinformatic prediction of the pathogenicity of bovine viral diarrhea virus genotypes: implications for viral virulence determinants, designing novel diagnostic assays and vaccines development . Front Vet Sci 2023, 10, 1130147. [Google Scholar] [CrossRef] [PubMed]
- Duraisamy, N. Machine learning tools used for mapping some immunogenic epitopes within the major structural proteins of the bovine coronavirus (BCoV) and for the in silico design of the multiepitope-based vaccines . Front Vet Sci 2024, 11, 1468890. [Google Scholar] [CrossRef]
- Goud, M.D. Artificial Intelligence Driven Framework for the Design and Development of Next-Generation Avian Viral Vaccines . Microorganisms 2025, 13(10). [Google Scholar] [CrossRef]
- Hemida, M.G. The next-generation coronavirus diagnostic techniques with particular emphasis on the SARS-CoV-2 . J Med Virol 2021, 93(7), 4219–4241. [Google Scholar] [CrossRef]
- Khan, M.Y. Repurposing of Some Nucleoside Analogs Targeting Some Key Proteins of the Avian H5N1 Clade 2.3.4.4b to Combat the Circulating HPAI in Birds: An In Silico Approach . Viruses 2025, 17(7). [Google Scholar] [CrossRef]
- Khan, M.Y. Leveraging Artificial Intelligence and Gene Expression Analysis to Identify Some Potential Bovine Coronavirus (BCoV) Receptors and Host Cell Enzymes Potentially Involved in the Viral Replication and Tissue Tropism . Int J Mol Sci 2025, 26(3). [Google Scholar] [CrossRef]
- Malik, Y.S. Advances in Diagnostic Approaches for Viral Etiologies of Diarrhea: From the Lab to the Field . Front Microbiol 2019, 10, 1957. [Google Scholar] [CrossRef]
- Al-Mubarak, A.I.A. Detection of Avian Orthoavulavirus-1 genotypes VI.2.1 and VII.1.1 with neuro-viscerotropic tropism in some backyard pigeons (Columbidae) in Eastern Saudi Arabia . Front Vet Sci 2024, 11, 1352636. [Google Scholar] [CrossRef] [PubMed]
- Alturaiki, W. The pivotal roles of the host immune response in the fine-tuning the infection and the development of the vaccines for SARS-CoV-2 . Hum Vaccin Immunother 2021, 17(10), 3297–3309. [Google Scholar] [CrossRef]
- Hemida, M.G.; Al-Ghadeer, H.M. Evidence of Peste des petits Ruminants' Virus in Dromedary Camels in the Kingdom of Saudi Arabia between 2014 and 2016 . In Vet Med Int; 2019; p. 4756404. [Google Scholar]
- Hemida, M.G. Molecular characterization and phylogenetic analyses of virulent infectious bronchitis viruses isolated from chickens in Eastern Saudi Arabia . Virusdisease 2017, 28(2), 189–199. [Google Scholar] [CrossRef]
- Hemida, M.G. Prevalence and molecular characterization of some circulating strains of the peste-des-petits-ruminants virus in Saudi Arabia between 2014-2016 . PeerJ 2020. 8, p. e9035. [Google Scholar] [CrossRef]
- Hemida, M.G. Longitudinal study of Middle East Respiratory Syndrome coronavirus infection in dromedary camel herds in Saudi Arabia, 2014-2015 . Emerg Microbes Infect 2017, 6(6), p. e56. [Google Scholar] [CrossRef] [PubMed]
- Hemida, M.G. Molecular characterisation of an avian influenza (H5N8) outbreak in backyard flocks in Al Ahsa, Eastern Saudi Arabia, 2017-2018 . Vet Rec Open 2019, 6(1), p. e000362. [Google Scholar] [CrossRef] [PubMed]
- Hemida, M.G. Evidence of equine influenza A (H3N8) activity in horses from Eastern and Central Saudi Arabia: 2013-2015 . Equine Vet J 2019, 51(2), 218–221. [Google Scholar] [CrossRef]
- Hemida, M.G. Foot-and-mouth disease virus O/ME-SA/Ind 2001 lineage outbreak in vaccinated Holstein Friesian cattle in Saudi Arabia in 2016 . Vet Q 2018, 38(1), 88–98. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Raghava, G.P. Prediction of continuous B-cell epitopes in an antigen using recurrent neural network . Proteins 2006, 65(1), 40–8. [Google Scholar] [CrossRef]
- Shah, A.U.; Gauger, P.; Hemida, M.G. Isolation and molecular characterization of an enteric isolate of the genotype-Ia bovine coronavirus with notable mutations in the receptor binding domain of the spike glycoprotein . Virology 2025. 603, 110313. [Google Scholar] [CrossRef]
- Shah, A.U. Some novel field isolates belonging to lineage-1 of the genotype GI-avian infectious bronchitis virus (AIBV) show strong evidence of recombination with field/vaccinal strains . Infect Genet Evol 2025. 129, 105723. [Google Scholar] [CrossRef]
- Singh, K.; Malik, Y.S.; Hemida, M.G. Comprehensive Evolutionary and Structural Analysis of the H5N1 Clade 2.4.3.4b Influenza a Virus Based on the Sequences and Data Mining of the Hemagglutinin, Nucleoprotein and Neuraminidase Genes Across Multiple Hosts . Pathogens 2025. 14, 9. [Google Scholar] [CrossRef]
- Liu, H. Design and development of a novel multi-epitope DNA vaccine candidate against infectious bronchitis virus: an immunoinformatic approach . Archives of Microbiology 2025, 207(4), p. 84. [Google Scholar] [CrossRef]
- Sher, H. Employing computational tools to design a multi-epitope vaccine targeting human immunodeficiency virus-1 (HIV-1) . BMC Genomics 2023, 24(1), 276. [Google Scholar] [CrossRef] [PubMed]
- Fleming, J. AlphaFold Protein Structure Database and 3D-Beacons: New Data and Capabilities . Journal of Molecular Biology 2025, 437(15), p. 168967. [Google Scholar] [CrossRef]
- Jumper, J. Highly accurate protein structure prediction with AlphaFold . Nature 2021, 596(7873), 583–589. [Google Scholar] [CrossRef]
- Mirdita, M. ColabFold: making protein folding accessible to all . Nature Methods 2022, 19(6), 679–682. [Google Scholar] [CrossRef]
- Vita, R. The Immune Epitope Database (IEDB): 2018 update . Nucleic Acids Res 2019, 47(D1), D339–d343. [Google Scholar] [CrossRef]
- Wiederstein, M.; Sippl, M.J. ProSA-web: interactive web service for the recognition of errors in three-dimensional structures of proteins . Nucleic Acids Res 2007, 35(Web Server issue), W407–10. [Google Scholar] [CrossRef]
- Sippl, M.J. Recognition of errors in three-dimensional structures of proteins . Proteins 1993, 17(4), 355–62. [Google Scholar] [CrossRef]
- Chen, V.B. MolProbity: all-atom structure validation for macromolecular crystallography . Acta Crystallogr D Biol Crystallogr 2010, 66 Pt 1, 12–21. [Google Scholar] [CrossRef]
- Doytchinova, I.A.; Flower, D.R. VaxiJen: a server for prediction of protective antigens, tumour antigens and subunit vaccines . BMC Bioinformatics 2007, 8, 4. [Google Scholar] [CrossRef]
- Heo, L.; Park, H.; Seok, C. GalaxyRefine: Protein structure refinement driven by side-chain repacking . Nucleic Acids Res 2013, 41(Web Server issue), W384–8. [Google Scholar] [CrossRef]
- Lee, G.R.; Heo, L.; Seok, C. Effective protein model structure refinement by loop modeling and overall relaxation . Proteins 2016, 84 Suppl 1, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Dimitrov, I. AllerTOP v.2--a server for in silico prediction of allergens . J Mol Model 2014, 20(6), p. 2278. [Google Scholar] [CrossRef]
- Yan, Y. The HDOCK server for integrated protein–protein docking . Nature Protocols 2020, 15(5), 1829–1852. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S. In silico approach for predicting toxicity of peptides and proteins . PLoS One 2013, 8(9), e73957. [Google Scholar] [CrossRef] [PubMed]
- Krissinel, E.; Henrick, K. Inference of Macromolecular Assemblies from Crystalline State . Journal of Molecular Biology 2007, 372(3), 774–797. [Google Scholar] [CrossRef]
- Hallgren, J. DeepTMHMM predicts alpha and beta transmembrane proteins using deep neural networks. 2022.04.08.487609. bioRxiv 2022. [Google Scholar]
- Rapin, N. Computational Immunology Meets Bioinformatics: The Use of Prediction Tools for Molecular Binding in the Simulation of the Immune System . PLOS ONE 2010, 5(4), e9862. [Google Scholar] [CrossRef]
- Blom, N. Prediction of post-translational glycosylation and phosphorylation of proteins from the amino acid sequence . Proteomics 2004, 4(6), 1633–49. [Google Scholar] [CrossRef] [PubMed]
- Blom, N.; Gammeltoft, S.; Brunak, S. Sequence and structure-based prediction of eukaryotic protein phosphorylation sites . J Mol Biol 1999, 294(5), 1351–62. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, M.R. Protein identification and analysis tools in the ExPASy server . Methods Mol Biol 1999, 112, 531–52. [Google Scholar]
- Gupta, R.; Brunak, S. Prediction of glycosylation across the human proteome and the correlation to protein function . In Pac Symp Biocomput; 2002; pp. 310–22. [Google Scholar]
- Dhanda, S.K. Prediction of IL4 inducing peptides . Clin Dev Immunol 2013, 263952. [Google Scholar] [CrossRef] [PubMed]
- Dhanda, S.K.; Vir, P.; Raghava, G.P. Designing of interferon-gamma inducing MHC class-II binders . Biol Direct 2013, 8, 30. [Google Scholar] [CrossRef]
- Kenyon, J.C.; Lever, A.M. The molecular biology of feline immunodeficiency virus (FIV) . Viruses 2011, 3(11), 2192–213. [Google Scholar] [CrossRef]
- González, S.A.; Affranchino, J.L. Properties and Functions of Feline Immunodeficiency Virus Gag Domains in Virion Assembly and Budding . Viruses 2018, 10. [Google Scholar] [CrossRef]
- Cervera, L. Production of HIV-1-based virus-like particles for vaccination: achievements and limits . Appl Microbiol Biotechnol 2019, 103(18), 7367–7384. [Google Scholar] [CrossRef]
- Chawla, M. Immunoinformatics-aided rational design of a multi-epitope vaccine targeting feline infectious peritonitis virus . Front Vet Sci 2023, 10, 1280273. [Google Scholar] [CrossRef]
- Romagnani, S. Th1/Th2 cells . Inflamm Bowel Dis 1999, 5(4), 285–94. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J. Development of high potency universal DR-restricted helper epitopes by modification of high affinity DR-blocking peptides . Immunity 1994, 1(9), 751–61. [Google Scholar] [CrossRef] [PubMed]
- Krogh, A. Predicting transmembrane protein topology with a hidden Markov model: application to complete genomes . J Mol Biol 2001, 305(3), 567–80. [Google Scholar] [CrossRef]
- Zhao, T. Vaccine adjuvants: mechanisms and platforms . Signal Transduction and Targeted Therapy 2023, 8(1), 283. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Zhou, X.; Wang, X. Glycosylation: mechanisms, biological functions and clinical implications . Signal Transduction and Targeted Therapy 2024, 9(1), p. 194. [Google Scholar] [CrossRef]
- Dunker, A.K. Intrinsically disordered protein . J Mol Graph Model 2001, 19(1), 26–59. [Google Scholar] [CrossRef]
- Barouch, D.H. Challenges in the development of an HIV-1 vaccine . Nature 2008, 455(7213), 613–9. [Google Scholar] [CrossRef]
- Aranyos, A.M. An initial examination of the potential role of T-cell immunity in protection against feline immunodeficiency virus (FIV) infection . Vaccine 2016, 34(12), 1480–8. [Google Scholar] [CrossRef]
- Collins, D.R.; Gaiha, G.D.; Walker, B.D. CD8(+) T cells in HIV control, cure and prevention . Nat Rev Immunol 2020, 20(8), 471–482. [Google Scholar] [CrossRef]
- Murphy, B.G. The Late Asymptomatic and Terminal Immunodeficiency Phases in Experimentally FIV-Infected Cats-A Long-Term Study . Viruses 2023, 15(8). [Google Scholar] [CrossRef]
- Plotkin, S.A. Correlates of protection induced by vaccination . Clin Vaccine Immunol 2010, 17(7), 1055–65. [Google Scholar] [CrossRef]
- del Guercio, M.F. Potent immunogenic short linear peptide constructs composed of B cell epitopes and Pan DR T helper epitopes (PADRE) for antibody responses in vivo . Vaccine 1997, 15(4), 441–8. [Google Scholar] [CrossRef]
- Alexander, J. The optimization of helper T lymphocyte (HTL) function in vaccine development . Immunol Res 1998, 18(2), 79–92. [Google Scholar] [CrossRef]
- Rani, N.A. Development of multi epitope subunit vaccines against emerging carp viruses Cyprinid herpesvirus 1 and 3 using immunoinformatics approach . Scientific Reports 2024, 14(1), p. 11783. [Google Scholar] [CrossRef]
- Rizarullah. Designing a Novel Multiepitope Vaccine from the Human Papilloma Virus E1 and E2 Proteins for Indonesia with Immunoinformatics and Molecular Dynamics Approaches . ACS Omega 2024, 9(14), 16547–16562. [Google Scholar] [CrossRef]
- Rani, N.A. Development of multi epitope subunit vaccines against emerging carp viruses Cyprinid herpesvirus 1 and 3 using immunoinformatics approach . Sci Rep 2024, 14(1), p. 11783. [Google Scholar] [CrossRef] [PubMed]
- Uhl, E.W. FIV vaccine development and its importance to veterinary and human medicine: a review FIV vaccine 2002 update and review . Vet Immunol Immunopathol 2002, 90(3-4), 113–32. [Google Scholar] [CrossRef] [PubMed]
- Robert-Tissot, C. Stimulation with a class A CpG oligonucleotide enhances resistance to infection with feline viruses from five different families . Vet Res 2012, 43(1), p. 60. [Google Scholar] [CrossRef] [PubMed]
- Wheat, W. Activation of upper respiratory tract mucosal innate immune responses in cats by liposomal toll-like receptor ligand complexes delivered topically . J Vet Intern Med 2019, 33(2), 838–845. [Google Scholar] [CrossRef] [PubMed]
Figure 2.
Schematic representation of the FIV-multi-epitope vaccine construct. The vaccine construct is arranged from C-terminus (left) to N-terminus (right) and contains immunostimulatory adjuvant PADRE, along with sequential B and T-cell epitopes from the conserved regions of pol, gag, and env genes of FIV. The epitopes are linked using rigid linkers (EAAAK) and flexible linkers (GPGPG, AAY, and KK) to allow for proper tertiary folding of the construct. A histidine tag was added to the N-terminus of the construct to facilitate protein purification.
Figure 2.
Schematic representation of the FIV-multi-epitope vaccine construct. The vaccine construct is arranged from C-terminus (left) to N-terminus (right) and contains immunostimulatory adjuvant PADRE, along with sequential B and T-cell epitopes from the conserved regions of pol, gag, and env genes of FIV. The epitopes are linked using rigid linkers (EAAAK) and flexible linkers (GPGPG, AAY, and KK) to allow for proper tertiary folding of the construct. A histidine tag was added to the N-terminus of the construct to facilitate protein purification.
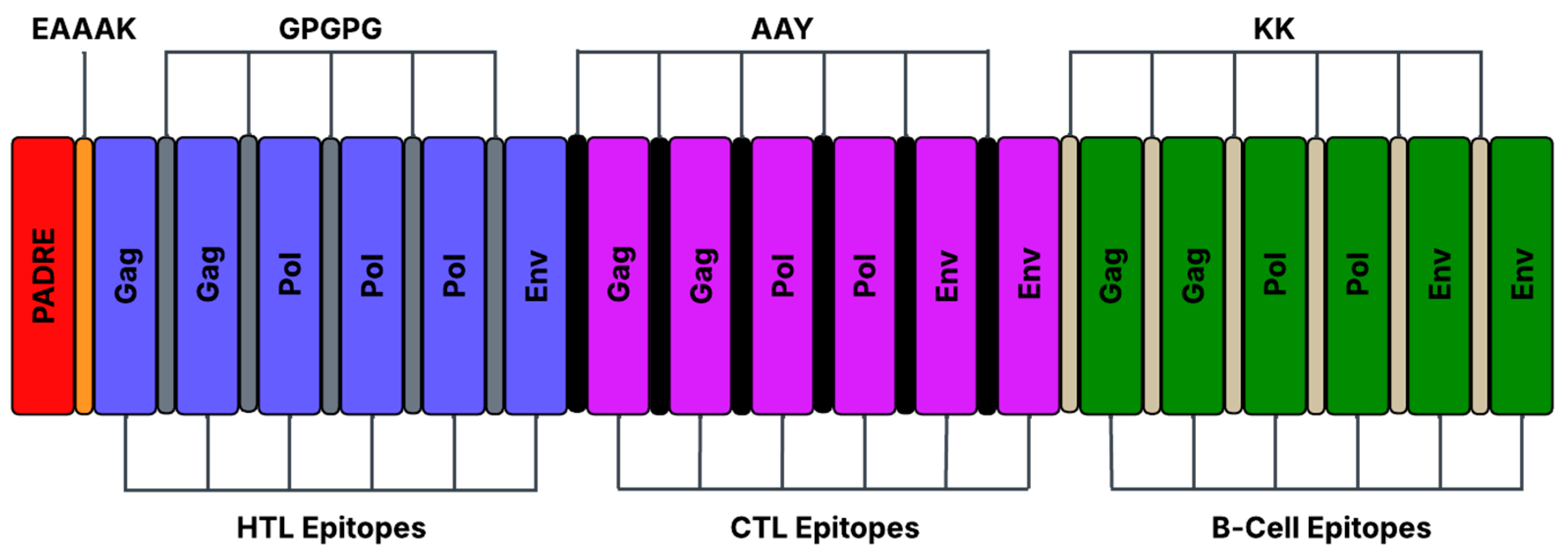
Figure 3.
Results of physicochemical property analysis of the FIV-DNA vaccine construct. A Transmembrane prediction results; B Predicted phosphorylation sites; C Hydrophilicity prediction; D Predicted glycosylation sites.
Figure 3.
Results of physicochemical property analysis of the FIV-DNA vaccine construct. A Transmembrane prediction results; B Predicted phosphorylation sites; C Hydrophilicity prediction; D Predicted glycosylation sites.
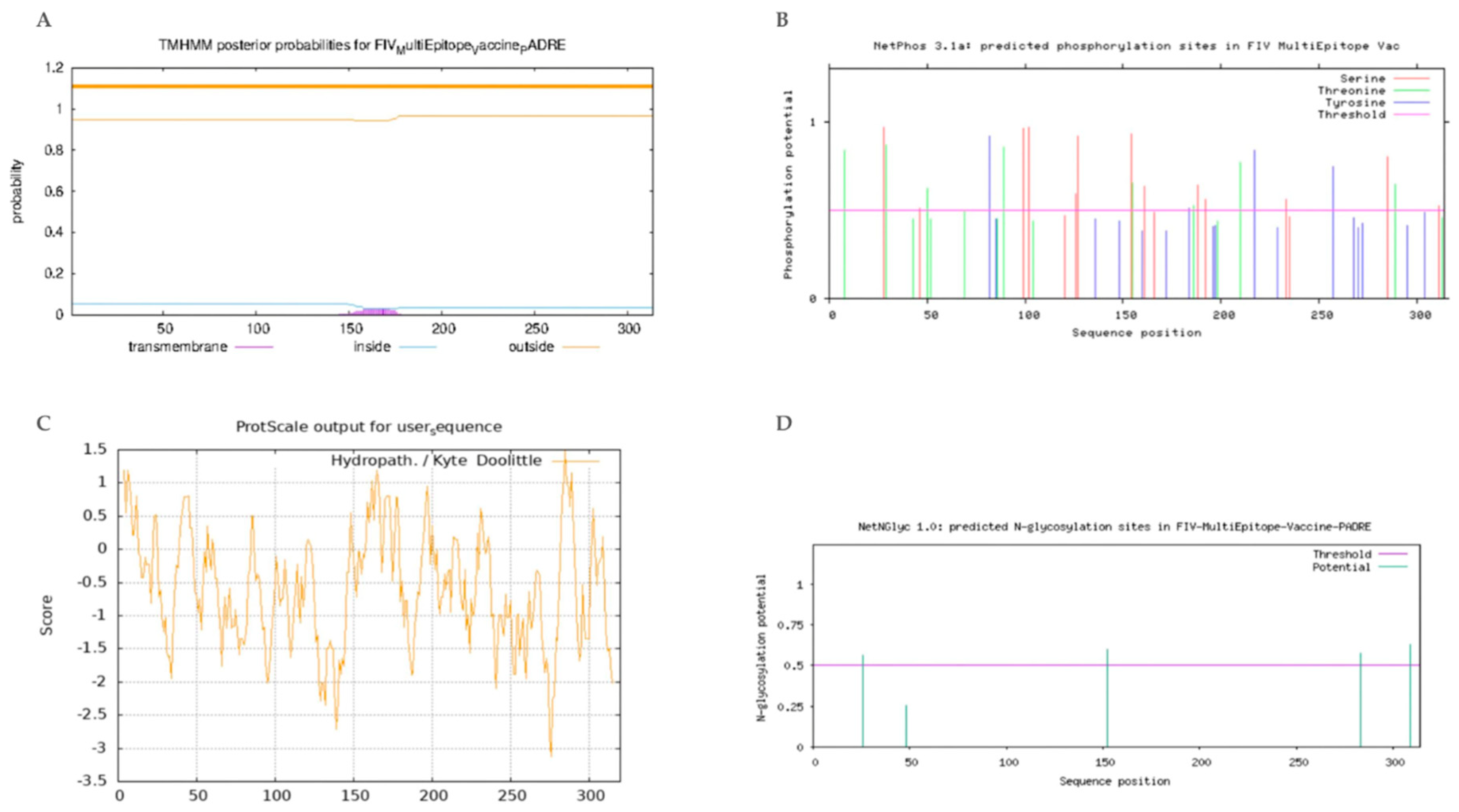
Figure 5.
Results of the tertiary structure and analysis of the protein structure. A. pre-optimized protein structure; B Pre-optimized Z-score residue analysis; C Pre-optimized Ramachandran plot; D Post-optimization protein structure; E Post-optimization Z-score residue analysis; F Post-optimization Ramachandran plot.
Figure 5.
Results of the tertiary structure and analysis of the protein structure. A. pre-optimized protein structure; B Pre-optimized Z-score residue analysis; C Pre-optimized Ramachandran plot; D Post-optimization protein structure; E Post-optimization Z-score residue analysis; F Post-optimization Ramachandran plot.
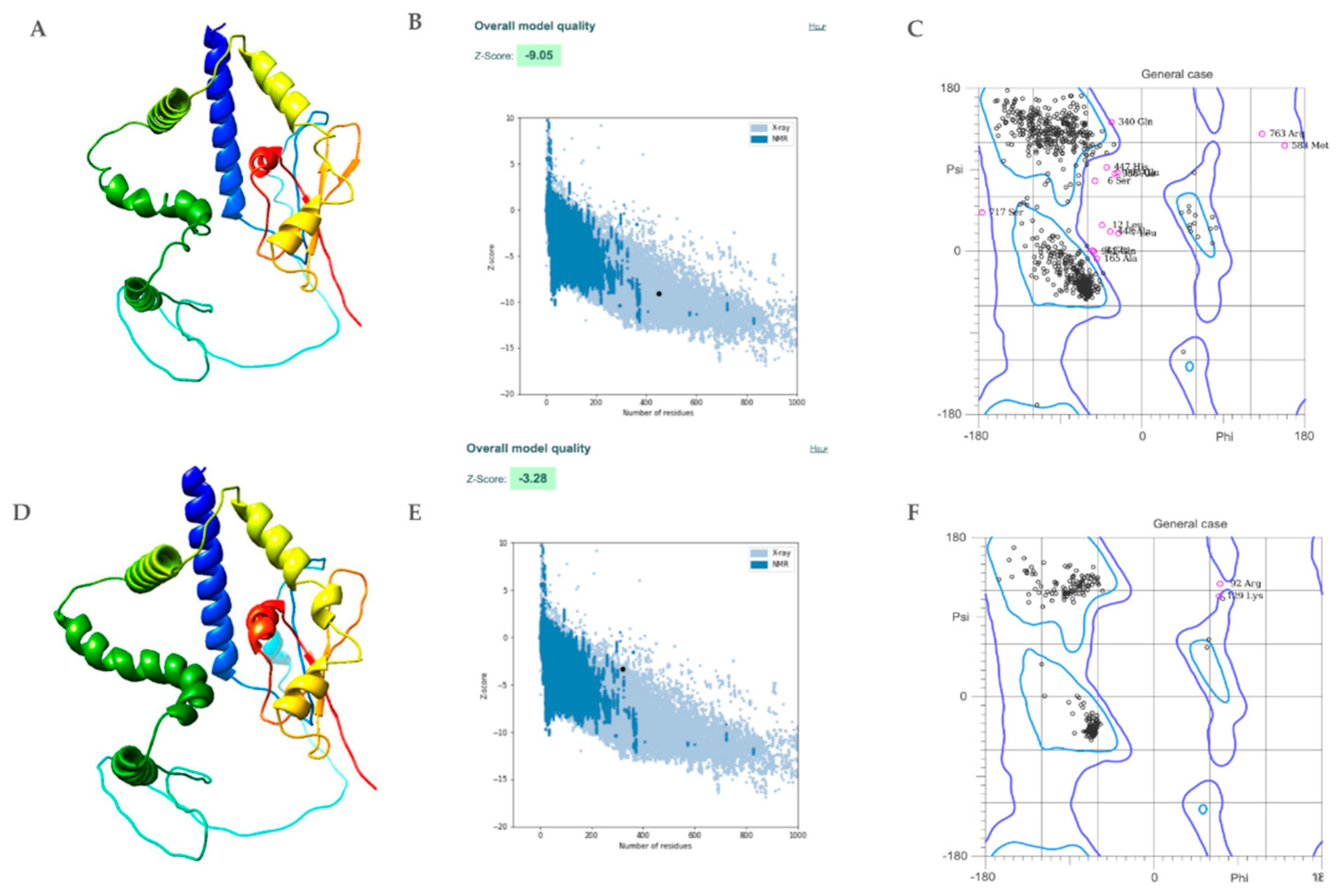
Figure 7.
Immune stimulation results of the vaccine construct using C-Immsim. (A) B-cell proliferation following repeated doses. (B) Antibody titters (IgM, IgG) following exposure to antigen. (C) Helper and memory T-cell production. (D) Cytotoxic T-cell activation. (E) Cytokine profile showing mainly increased IFN-γ and IL-2.
Figure 7.
Immune stimulation results of the vaccine construct using C-Immsim. (A) B-cell proliferation following repeated doses. (B) Antibody titters (IgM, IgG) following exposure to antigen. (C) Helper and memory T-cell production. (D) Cytotoxic T-cell activation. (E) Cytokine profile showing mainly increased IFN-γ and IL-2.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



