Submitted:
10 February 2026
Posted:
10 February 2026
You are already at the latest version
Abstract
Pertussis is a highly contagious acute respiratory infection caused by Bordetella pertussis (B. pertussis), an exclusively human pathogen that remains a significant global public health concern despite widespread vaccination. Cyclical resurgences continue to occur, particularly in highly immunized populations, largely driven by waning immunity following acellular pertussis vaccination and persistent asymptomatic transmission among adolescents and adults. The pathogenesis of B. pertussis infection is mediated by a coordinated repertoire of adhesins and toxins that enable efficient colonization of the respiratory epithelium, disruption of mucociliary clearance, and profound modulation of host immune responses. Key virulence factors include pertussis toxin, adenylate cyclase toxin, and tracheal cytotoxin, which together impair innate immune defenses, alter leukocyte trafficking, suppress phagocytic function, and induce epithelial injury. While antibody-mediated immunity limits disease severity, it is insufficient to prevent colonization and onward transmission, underscoring the critical role of cell-mediated immunity. Protective clearance of infection is strongly associated with T helper 1 and T helper 17 immune responses, mucosal immunoglobulin A production, and the establishment of airway tissue-resident memory T cells—immune components that are suboptimally induced by current acellular vaccines. Advances in controlled human infection models have provided direct evidence that existing vaccines fail to prevent nasopharyngeal carriage, accelerating the development of next-generation vaccine strategies. These approaches aim to restore infection-like immune programming through mucosal delivery, enhanced innate immune activation, and broader antigenic presentation. Understanding the immune pathogenesis of B. pertussis infection provides a critical framework for redesigning vaccines capable of achieving durable, transmission-blocking immunity.
Keywords:
Bordetella pertussis
; pertussis pathogenesis
; immune evasion
; mucosal immunity
; vaccine-induced immunity
1. Introduction
Pertussis (whooping cough) is a highly contagious acute respiratory infection caused by Bordetella pertussis (B. pertussis), an exclusively human pathogen with a high secondary attack rate among susceptible contacts. Despite widespread vaccination and significant reductions in incidence compared with the pre-vaccine era, pertussis remains a global health problem with cyclical outbreaks even in highly immunized populations [1,2]. In the absence of immunization, pertussis was historically a childhood disease with high morbidity and mortality; however, waning immunity following childhood vaccination has shifted a substantial burden to older age groups [3,4]. Immunity following acellular pertussis vaccination declines relatively rapidly, with observational data showing that protection diminishes within a few years after the last dose and may be limited beyond 7–10 years, contributing to increased susceptibility among adolescents and adults [3,5,6]. These older age groups now act as an important reservoir of infection, often transmitting B. pertussis to unvaccinated or partially vaccinated infants who are at greatest risk of severe illness, hospitalization, and death [7].
The pathogenesis of B. pertussis involves a coordinated array of adhesins and toxins that facilitate airway colonization and immune evasion. Key virulence factors include pertussis toxin (PT), filamentous hemagglutinin, pertactin and other adhesins, as well as adenylate cyclase toxin, which together modulate host defenses and promote bacterial persistence [2,8]. Pertussis toxin, in particular, disrupts G protein signaling in host cells through ADP-ribosylation, subverting immune responses and contributing to disease manifestations [9]. Chemically inactivated pertussis toxin is a central component of current acellular vaccines and is critical for inducing protective antibody responses, although vaccine-induced immunity is not long-lasting [5,6]. Although antigenic changes in circulating strains such as pertactin deficiency and potential adaptation under vaccine pressure have been described, current evidence supports continued effectiveness of approved vaccines in preventing severe disease, but with an acknowledged limitation in the duration of protection [3,10].
The key steps in the pathogenesis of B. pertussis infection and the associated host immune responses are summarized in Figure 1.
Following inhalation, B. pertussis adheres to the ciliated respiratory epithelium through surface adhesins, including filamentous hemagglutinin, pertactin, and fimbriae. After colonization, the bacterium produces multiple virulence factors that disrupt host defense mechanisms. Pertussis toxin (PT) induces ADP-ribosylation of inhibitory Gi proteins, leading to dysregulated intracellular cyclic adenosine monophosphate (cAMP) signaling, impaired leukocyte trafficking, and characteristic lymphocytosis. Adenylate cyclase toxin (ACT) enters phagocytic cells via complement receptor 3 (CD11b/CD18), causing pathological cAMP accumulation and suppression of phagocytosis and oxidative burst. Tracheal cytotoxin (TCT), a peptidoglycan fragment released during bacterial cell wall turnover, selectively damages ciliated epithelial cells, resulting in impaired mucociliary clearance. Although innate immune recognition through Toll-like receptor 4 (TLR4) activates inflammatory pathways, toxin-mediated immune modulation skews adaptive responses toward weak Th1 and Th17 immunity. Antibody responses limit disease severity but are insufficient to prevent bacterial persistence and transmission.
Clinically, pertussis progresses through three characteristic stages. The catarrhal stage presents with nonspecific symptoms including rhinorrhea, sneezing, and a mild persistent cough; this phase is highly infectious and may be indistinguishable from viral upper respiratory infections, particularly in infants who often lack the classic inspiratory “whoop” [2,7]. The paroxysmal stage follows, defined by recurrent, forceful coughing paroxysms that may be accompanied by vomiting, cyanosis, or hypoxia; in unvaccinated children, an inspiratory whoop is a recognized hallmark, and this stage carries the highest risk of complications including apnea and secondary bacterial pneumonia [7]. The convalescent stage involves a gradual decline in cough severity, although paroxysmal coughing can persist for weeks or progress to a chronic pattern.
Recent epidemiological data confirm a resurgence of pertussis in many regions, with increased incidence noted among adolescents and adults—populations with waning vaccine-derived immunity and under-recognized clinical disease [4,7,11]. These older individuals are increasingly identified as key contributors to transmission, particularly to young infants too young to complete primary immunization. As a result, many countries have expanded immunization strategies to include booster doses for older children, adolescents, and adults in an effort to reduce transmission and protect vulnerable infants [7,12].
2. Bacterial Structure and Virulence Factors
Lipopolysaccharide (LPS) of B. pertussis represents a structurally atypical but immunologically active endotoxin that plays a central role in host–pathogen interactions. Although B. pertussis LPS exhibits lower endotoxic activity than classical enterobacterial LPS, it is nevertheless recognized by the host innate immune system through Toll-like receptor 4 (TLR4), initiating downstream inflammatory signaling cascades [13,14]. The lipid A moiety of B. pertussis LPS is characterized by a compact structure with reduced phosphorylation and acylation by six fatty acid residues, features that modulate its interaction with TLR4 and contribute to immune evasion while retaining immunostimulatory capacity [15]. Importantly, synthetic lipidation of detoxified pertussis toxoid has been shown to restore TLR4 agonist activity, significantly enhancing vaccine immunogenicity and protective efficacy by bridging innate and adaptive immune responses [9,16].
This finely tuned endotoxin activity enables B. pertussis to establish infection rapidly while inducing robust cytokine release, including interleukin-1β, tumor necrosis factor-α and interleukin-6, which contribute to the systemic inflammatory response characteristic of severe pertussis [17]. Simultaneously, the pathogen deploys an array of toxins—most notably pertussis toxin and adenylate cyclase toxin—to subvert host immunity. Pertussis toxin promotes marked lymphocytosis by inhibiting leukocyte trafficking, while adenylate cyclase toxin suppresses phagocytic killing through pathological elevation of intracellular cyclic AMP, collectively dampening early immune containment [18,19].
B. pertussis is a small, Gram-negative, strictly aerobic, non-motile coccobacillus with limited metabolic capabilities and a relatively small genome, reflecting its adaptation to an exclusively human host [1]. It is fastidious in vitro, requiring specialized media such as Bordet–Gengou agar or charcoal-based agar supplemented with blood and antibiotics for optimal growth. The outer membrane of B. pertussis contains numerous surface-exposed proteins and endotoxins that are essential for adhesion, immune modulation and persistence within the respiratory tract [20]. Unlike many encapsulated respiratory pathogens, B. pertussis does not possess a classical polysaccharide capsule; instead, virulence is mediated through a complex repertoire of secreted toxins and surface adhesins, including pertussis toxin, adenylate cyclase-hemolysin, pertactin, filamentous hemagglutinin (FHA), and tracheal colonization factor [18,20].
The bacterial cell envelope contains structurally distinctive lipids and carbohydrates that contribute to immune recognition and signaling. Host cell engagement by adenylate cyclase toxin requires interaction with complement receptor 3 (CD11b/CD18) on leukocytes, facilitating toxin translocation and pathological cAMP accumulation, a mechanism central to immune suppression during early infection [19].
3. Adhesion and Colonization Mechanisms
Successful adhesion and colonization of the respiratory epithelium represent the critical first steps in B. pertussis infection. Multiple adhesins have been identified, although their precise hierarchy and regulation during human infection remain incompletely defined. The best-characterized adhesins include filamentous hemagglutinin, pertactin, fimbrial proteins (Fim2 and Fim3), and type IV pili [20,21,22]. Among these, FHA plays a dominant role in tracheal colonization by mediating high-affinity binding to ciliated epithelial cells via integrins and sulfated glycolipids [21]. Experimental studies have demonstrated that immunization with FHA reduces bacterial burden during early colonization, underscoring its functional importance and relevance as a vaccine antigen [23].
Fimbriae contribute primarily to the initial stages of adherence and are regulated by the BvgAS two-component virulence regulatory system, which coordinates expression of multiple colonization and toxin genes in response to environmental cues [20,24]. Following attachment to the epithelial surface, type IV secretion system–associated effectors facilitate bacterial persistence and limited internalization into host cells, although B. pertussis is predominantly considered an extracellular pathogen [25].
B. pertussis is restricted to the human respiratory tract, with infection initiated through inhalation of aerosolized bacteria. Colonization of the ciliated epithelium leads to production of tracheal cytotoxin, a peptidoglycan fragment that selectively damages ciliated cells, impairing mucociliary clearance and driving the characteristic cough of pertussis [26]. This epithelial injury promotes bacterial persistence and facilitates transmission to new susceptible hosts.
Early studies demonstrated that B. pertussis binds selectively to ciliated cells of the human respiratory tract, suggesting the involvement of specific host receptors [27]. Pertactin has been shown to interact with host immunoglobulin-like domains, including IgD expressed on subsets of B cells, enhancing cellular adhesion under experimental conditions [28]. However, adhesion of viable Bordetella species to respiratory epithelial cells is mediated by multiple redundant mechanisms, and no single receptor has been identified as absolutely required for epithelial binding. Consequently, the precise adhesin–receptor interactions governing adult-to-adult and adult-to-infant transmission remain incompletely understood, highlighting a critical gap in current knowledge and an important target for future research.
4. Toxins Produced by Bordetella pertussis
The toxins produced by B. pertussis play a central role in disease pathogenesis and are largely responsible for the complex local and systemic manifestations of pertussis. Among these, pertussis toxin (PT) and adenylate cyclase–hemolysin toxin (ACT) represent the most extensively characterized and biologically significant virulence factors [18,29]. These toxins act in a coordinated manner to disrupt innate and adaptive immune responses, facilitate bacterial persistence within the respiratory tract, and promote transmission. Adenylate cyclase toxin is a bifunctional RTX family exotoxin that profoundly alters host cell signaling through dysregulation of intracellular cyclic adenosine monophosphate (cAMP). Following binding to complement receptor 3 (CD11b/CD18) on myeloid cells, the catalytic domain of ACT is translocated into the host cytosol, where it catalyzes uncontrolled cAMP production, resulting in protein kinase A activation and subsequent functional paralysis of mononuclear phagocytes [30,31]. This intoxication inhibits phagocytosis, oxidative burst, and cytokine production, thereby allowing B. pertussis to evade early immune clearance during infection [19]. Pertussis toxin is a classical AB₅ toxin that exerts systemic immunomodulatory effects by ADP-ribosylating inhibitory Gi/Go G-proteins, leading to dysregulation of G-protein–coupled receptor signaling pathways and excessive accumulation of intracellular cAMP [32]. Through these mechanisms, PT induces profound leukocytosis by impairing leukocyte extravasation from the bloodstream, alters lymphocyte trafficking, and suppresses effective immune responses, contributing to disease severity and prolonged infection [29]. In addition to PT and ACT, other secreted and surface-associated factors—including filamentous hemagglutinin (FHA), fimbriae, and tracheal cytotoxin—exert auxiliary toxic and immunomodulatory effects that enhance colonization and persistence within the respiratory epithelium [18,33]. Collectively, these virulence determinants support sustained adherence, impair mucociliary clearance, and establish a protected ecological niche for bacterial survival. Genomic studies have revealed considerable genetic plasticity within circulating B. pertussis populations. During natural infection, toxin-deficient or toxin-altered variants may arise as part of within-host adaptation, whereas such variants are not represented in vaccine formulations, which rely on stable, chemically inactivated toxin antigens [6]. This evolutionary divergence between vaccine strains and circulating isolates has raised concerns regarding antigenic mismatch and has complicated efforts toward complete eradication of pertussis, underscoring biological constraints on the development of fully avirulent yet immunogenic vaccines [34].
4.1. Pertussis Toxin (PT)
Pertussis toxin is widely regarded as the single most critical virulence determinant in the pathophysiology of pertussis and serves as a principal antigenic component of acellular pertussis vaccines [32,35]. PT is synthesized as a multimeric protein encoded by the ptx operon, which directs toxin production, post-translational modification, and secretion via the Ptl type IV secretion system [36]. PT consists of an enzymatically active S1 subunit and a pentameric B oligomer responsible for host cell binding. Following internalization, the S1 subunit catalyzes ADP-ribosylation of Giα proteins, uncoupling inhibitory G-protein signaling and leading to persistent activation of adenylate cyclase with pathological increases in intracellular cAMP [32]. These effects disrupt immune cell migration, inhibit phagocytic responses, and promote lymphocytosis—one of the hallmark laboratory features of severe pertussis [29].Genetic instability of the ptx operon has been described in laboratory-passaged B. pertussis strains, particularly in the reference strain Tohama I, where spontaneous mutations or loss of operon function may occur during repeated subculturing [37]. Such instability complicates in vitro toxin production and has historically necessitated careful strain selection for vaccine antigen manufacture. Importantly, although PT-deficient mutants may emerge experimentally or transiently during infection, circulating clinical isolates generally retain functional toxin expression, highlighting the essential role of PT in bacterial fitness and transmission [6,38].Beyond its direct pathogenic effects, PT may also contribute indirectly to bacterial survival by modulating microbial competition and host–microbe interactions within the respiratory tract, although these ancillary roles remain incompletely understood [17].
4.2. Adenylate Cyclase Toxin (ACT)
Adenylate cyclase toxin (ACT), encoded by the cyaA gene, is another major virulence factor expressed by B. pertussis, as well as B. parapertussis and B. bronchiseptica [30]. ACT is a large (~1706 amino acid) multifunctional protein comprising an N-terminal calmodulin-dependent adenylate cyclase domain and a C-terminal RTX hemolysin domain responsible for membrane interaction and translocation [31,39].
The N-terminal catalytic domain is activated upon binding host calmodulin, enabling rapid and unregulated cAMP synthesis in intoxicated cells. The large size and modular architecture of ACT facilitate efficient translocation across host cell membranes, an essential determinant of its biological potency [40]. Post-translational palmitoylation by the CyaC acyltransferase is required for full toxin activity and host cell targeting [41].
Although ACT is not essential for lethality in murine respiratory infection models, it plays a critical role during early infection by suppressing innate immune responses and enhancing bacterial colonization efficiency [19,42]. Distinct structural variants of ACT have been described across Bordetella species, with virulent B. pertussis strains typically producing a full-length, highly active toxin, whereas other species may generate truncated or alternative forms with differing biological activity [39].
4.3. Tracheal Cytotoxin (TCT)
Tracheal cytotoxin (TCT) is a key virulence factor of B. pertussis that contributes directly to epithelial damage and disruption of mucociliary clearance, a defining pathological feature of pertussis. Unlike classical protein exotoxins, TCT is a low-molecular-weight peptidoglycan fragment released during bacterial cell wall turnover and is not actively secreted through specialized secretion systems [33,43]. Chemically, TCT corresponds to a disaccharide-tetrapeptide monomer of peptidoglycan (N-acetylglucosamine–N-acetylmuramic acid–tetrapeptide) with a molecular mass of approximately 921 Da, rather than a lipid-based structure [44].
Production and accumulation of TCT are influenced by environmental conditions, including nutrient availability. Experimental data indicate that iron-limited conditions—resembling the host respiratory tract—favor increased release of peptidoglycan fragments, including TCT, whereas iron-replete environments suppress their accumulation [45]. This regulatory pattern may enhance pathogenic effects specifically within the human airway.
TCT selectively targets ciliated epithelial cells of the trachea and bronchi. It induces nitric oxide synthase expression and excessive nitric oxide production, leading to oxidative stress, ciliary dysfunction, and eventual ciliated cell extrusion [46]. These effects result in impaired mucociliary clearance and create a permissive niche for bacterial persistence, thereby facilitating prolonged infection and efficient transmission.
Unlike pertussis toxin, TCT does not appear to significantly modulate systemic anti-PT antibody responses [29]. However, immune recognition of TCT at the mucosal surface plays an important role in limiting bacterial attachment during early exposure. High levels of TCT-specific IgA and IgM have been detected in nasal secretions following exposure, suggesting that mucosal antibodies may act as a first-line barrier preventing epithelial damage and reducing effective colonization [47]. Subsequent systemic IgG and IgA responses are also generated, contributing to longer-term immune control without evidence of antigenic variation in TCT that would compromise antibody binding [48].
Importantly, primary exposure to B. pertussis may remain clinically silent, particularly in partially immune individuals, yet such asymptomatic or minimally symptomatic carriers can serve as major reservoirs for transmission within susceptible populations [49]. TCT-mediated epithelial injury is thought to facilitate bacterial shedding and environmental dissemination, reinforcing its role not only in pathogenesis but also in epidemiological spread [46,50]. Experimental models have demonstrated that neutralization of TCT—particularly at the mucosal surface—reduces epithelial damage and limits bacterial transmission, supporting the concept that targeting TCT-induced pathology could complement toxin-based vaccine strategies [29,47].
4.4. Human Carriage and Controlled Human Infection Models
In response to the growing recognition that transmission is often driven by minimally symptomatic or asymptomatic carriage, controlled human infection models (CHIMs) have been developed to safely induce nasopharyngeal colonization in healthy adults under intensive monitoring. In the PERISCOPE program, intranasal inoculation with a well-characterized clinical isolate (B1917) reproducibly established colonization without classical pertussis symptoms, enabling high-resolution study of early host-pathogen interactions and correlates of protection. Colonization and shedding were quantified by serial sampling (pernasal/nasopharyngeal swabs, nasal washes and throat swabs) combined with culture and qPCR, with eradication therapy (e.g., azithromycin) administered at a predefined timepoint to terminate carriage [51,52].
These platforms are particularly valuable for next-generation vaccines because they directly measure the outcome that current aP vaccines fail to control: acquisition of colonization and bacterial burden in the upper airway. The most advanced application to date is the BPZE1 challenge program (CHAMPION-1), in which participants received a single intranasal dose of live-attenuated BPZE1 or placebo and were subsequently challenged with virulent B. pertussis under quarantine conditions. Using repeated quantitative culture/qPCR readouts, BPZE1 reduced the proportion of volunteers with detectable colonization and lowered overall bacterial load (area-under-the-curve style endpoints) compared with placebo, supporting the concept that mucosal, infection-like priming can blunt carriage in humans [42,53].
Methodologically, CHIMs also provide an experimental sandbox to refine sampling and microbiological quantification. In addition to standard culture and DNA qPCR, viability-linked approaches (such as propidium monoazide pre-treatment coupled to qPCR) can help distinguish viable from non-viable organisms and may better approximate infectious shedding [54]. Together, these methods enable quantitative, time-resolved mapping of colonization kinetics and accelerate down-selection of vaccine platforms before large field efficacy trials.
5. Immune Response to Bordetella pertussis
Protective immunity against B. pertussis is complex and multifactorial, requiring the coordinated activation of innate, humoral, and cellular immune responses. Naturally acquired infection induces broader and more durable immunity than vaccination, particularly acellular pertussis vaccines, although neither form of immunity is lifelong [55,56]. Effective control of bacterial replication and prevention of persistent colonization depend on the integration of antibody-mediated protection with robust T-cell–dependent immune mechanisms [57].
Antibodies play a crucial role in limiting disease severity by neutralizing pertussis toxin, inhibiting bacterial adhesion, and promoting opsonophagocytic clearance. However, humoral immunity alone is insufficient to prevent infection or transmission. Experimental and clinical data demonstrate that antibody responses require appropriate innate immune signaling—particularly through pattern recognition receptors such as Toll-like receptor 4 (TLR4)—to support effective T-helper cell differentiation and immunological memory formation [17,58]. In the absence of such signals, antibody production may occur but lacks protective durability and functional efficacy.
During B. pertussis infection, the innate immune system responds rapidly to bacterial components including lipopolysaccharide, toxins, and adhesins. These factors activate epithelial cells, macrophages, dendritic cells, and neutrophils, initiating cytokine and chemokine cascades that shape downstream adaptive responses [19]. Paradoxically, B. pertussis exploits innate immune pathways to skew host responses in its favor. Pertussis toxin and adenylate cyclase toxin suppress effective phagocytic killing, impair antigen presentation, and bias T-cell polarization away from protective Th1/Th17 responses, thereby facilitating sustained bacterial survival in the respiratory tract [17,59,60].
Cell-mediated immunity, particularly Th1- and Th17-type responses, is essential for bacterial clearance. Interferon-γ–producing Th1 cells promote macrophage activation, while Th17 cells enhance neutrophil recruitment and mucosal defense [61]. The relative deficiency of these responses following acellular vaccination explains, at least in part, the failure to achieve sterilizing immunity and the persistence of asymptomatic carriage in vaccinated individuals [56,62].
6. Cellular Interactions During Infection
Direct interactions between B. pertussis and the respiratory epithelium are central to colonization and disease initiation. The bacterium preferentially binds to ciliated epithelial cells lining the upper and lower airways, where it establishes a localized extracellular niche [63]. Experimental infection models demonstrate species-specific differences in epithelial binding efficiency, with murine cells often supporting higher levels of bacterial attachment and colonization than human cells, likely due to differences in receptor availability, epithelial architecture, or microvillus organization [1].
Adhesion to epithelial surfaces is mediated by a repertoire of surface-expressed adhesins, including filamentous hemagglutinin (FHA), fimbriae (Fim2 and Fim3), and pertactin [21]. These adhesins facilitate stable attachment, resist mechanical clearance by mucociliary activity, and promote bacterial persistence. FHA plays a dominant role by interacting with integrins and sulfated glycoconjugates on epithelial and immune cells, while fimbriae contribute to early-stage attachment and colonization [64].
Beyond epithelial cells, B. pertussis interacts dynamically with immune cells such as macrophages and dendritic cells. Although the bacterium is primarily extracellular, limited internalization into host cells has been observed and may contribute to immune modulation rather than intracellular replication [50]. These interactions influence antigen presentation, cytokine secretion, and the quality of the adaptive immune response, reinforcing the importance of host cell–pathogen contact in shaping disease outcome.
7. Inflammatory Response in Pertussis
The inflammatory response to B. pertussis infection is characterized by a distinctive imbalance between robust local inflammation and relatively modest systemic manifestations. Lung inflammation is driven by activation and subsequent dysregulation of neutrophils, alveolar macrophages, and epithelial cells, which collectively orchestrate both innate and adaptive immune responses [19,40].
A hallmark of pertussis pathology is extensive leukocyte activation followed by immune cell dysfunction and death. Neutrophils and macrophages that ingest B. pertussis are exposed to adenylate cyclase toxin–mediated cAMP overload and pertussis toxin–induced signaling disruption, leading to impaired chemotaxis, defective phagolysosomal function, and eventual cell lysis [29,59]. The accumulation and subsequent destruction of these immune cells contribute to local tissue damage, airway inflammation, and impaired gas exchange, which may underlie the mild hypoxia observed during infection despite preserved ventilatory drive [65].
Emerging evidence implicates dysregulation of intracellular calcium (Ca²⁺) homeostasis as a key mechanism underlying immune cell dysfunction and death during pertussis. Failure to maintain Ca²⁺ balance disrupts actin cytoskeleton dynamics in phagocytes, impairing motility, chemotaxis, and effective immune surveillance [65]. In neutrophils with compromised phagocytic capacity, this imbalance exacerbates inflammatory injury while paradoxically limiting bacterial clearance.
8. Immune Pathogenesis of Bordetella pertussis
The immune pathogenesis of B. pertussis infection is defined by the paradoxical coexistence of intense immune activation and profound immune dysfunction. Rather than evading immune recognition entirely, B. pertussis actively manipulates innate and adaptive immune responses to favor prolonged survival in the respiratory tract, persistent inflammation, and efficient transmission [18,66]. Disease severity and failure to achieve sterilizing immunity result primarily from immune dysregulation rather than uncontrolled bacterial replication.
Initial host–pathogen interaction occurs at the respiratory mucosa, where epithelial cells and resident macrophages recognize bacterial components such as lipopolysaccharide, peptidoglycan fragments, and adhesins via pattern-recognition receptors, particularly Toll-like receptor 4 (TLR4) [57]. This recognition induces early inflammatory signaling and cytokine production; however, B. pertussis rapidly subverts these responses through the coordinated action of pertussis toxin (PT), adenylate cyclase toxin (ACT), and tracheal cytotoxin (TCT) [17].
PT plays a central role in systemic immune dysregulation. By ADP-ribosylating inhibitory Gi proteins, PT disrupts G-protein–coupled receptor signaling, leading to aberrant intracellular cAMP accumulation in immune cells [67]. This signaling defect impairs chemokine receptor function, inhibiting leukocyte extravasation and resulting in the characteristic lymphocytosis observed in severe pertussis [68]. Consequently, immune cells are abundant in circulation but functionally excluded from sites of infection, delaying effective bacterial clearance.
ACT further amplifies immune paralysis at the local level by delivering a calmodulin-dependent adenylate cyclase directly into phagocytes. The resulting supraphysiologic cAMP concentrations suppress phagocytosis, oxidative burst, antigen presentation, and proinflammatory cytokine production in neutrophils, macrophages, and dendritic cells [19]. These effects compromise both innate bacterial killing and the priming of adaptive immune responses.
Simultaneously, TCT induces targeted destruction of ciliated epithelial cells through nitric oxide–mediated cytotoxicity, impairing mucociliary clearance and perpetuating bacterial persistence [69]. This epithelial injury sustains chronic airway inflammation while facilitating ongoing bacterial shedding and transmission.
Adaptive immunity to B. pertussis is similarly shaped by toxin-driven immune modulation. Protective clearance requires coordinated Th1 and Th17 cellular responses, which promote macrophage activation and neutrophil recruitment to the airway mucosa [60]. However, PT and ACT bias T-cell differentiation away from durable Th1/Th17 immunity, favoring suboptimal responses that permit asymptomatic carriage [70]. This phenomenon is particularly evident following acellular pertussis vaccination, which induces strong antibody responses but weak cellular immunity, explaining vaccine failure to prevent colonization and transmission despite protection from severe disease [56].
Humoral immunity remains essential for limiting pathology by neutralizing toxins and reducing disease severity, but antibodies alone are insufficient for sterilizing immunity. Effective antibody production and memory formation depend on intact innate signaling pathways, particularly TLR4-mediated activation, which are dampened during natural infection and inadequately stimulated by acellular vaccines [57,71].
9. Immune Pathogenesis as a Blueprint for Vaccine Redesign
The key immunopathogenesis insight driving “next-generation” pertussis vaccine design is that current acellular pertussis (aP) vaccines protect well against symptomatic disease but do not reliably prevent nasopharyngeal colonization and onward transmission, largely because they fail to elicit durable mucosal immunity and robust Th1/Th17 cellular responses in the respiratory tract [56,72,73]. Natural infection (and, to a greater extent, older whole-cell vaccines) induces a broader immune program that includes Th1/Th17 polarization, airway tissue-resident memory T cells (T_RM), and secretory IgA, which are repeatedly implicated as requirements for sterilizing immunity at the site of entry [47,56,72,73].
9.1. Redesign Target: Block Colonization and Transmission (Mucosal Immunity, T_RM, sIgA)
B. pertussis largely remains localized to the airways, and its toxins (PT, ACT) and inflammatory injury (including TCT-mediated epithelial damage) create a setting where systemic antibodies can limit severity but do not necessarily clear carriage [56,73]. This shifts the primary vaccine goal from “prevent disease” to “prevent infection/colonization in the nose and upper airway.” Reviews synthesizing animal and human data emphasize that mucosal Th17 responses, interleukin-17 (IL-17)-driven recruitment, and sIgA, alongside respiratory T_RM, are central correlates for durable anti-colonization immunity—precisely the compartments that aP vaccines weakly induce [47,72].
Consistent with this mechanistic goal, intranasal vaccination strategies have shown improved control of upper-airway colonization in preclinical models. Intranasal delivery of acellular antigens can induce B. pertussis–specific mucosal IgA and IL-17A and reduce bacterial burden in both upper and lower respiratory tracts in mice, demonstrating that “route” can partially correct the immunologic deficit of standard intramuscular aP vaccination [74].
9.2. Redesign Target: Restore Th1/Th17-Skewing Through Innate-Programming Adjuvants
Acellular vaccines are classically alum-adjuvanted and tend to skew toward Th2-biased responses, whereas clearance of B. pertussis and prevention of carriage are more strongly associated with Th1/Th17 immunity [56,72,73]. This has motivated redesign around pattern-recognition receptor (PRR) agonists that better mimic infection-like innate signals.
A practical proof-of-concept is that adding a TLR7 agonist to an aP formulation significantly enhances Th1/Th17 responses and protection in a mouse model, directly aligning vaccine outputs with the immune mechanisms required for bacterial control at mucosal surfaces [75]. TLR4 agonist biology is also central to modern vaccinology: monophosphoryl lipid A (MPL) is a TRIF-biased TLR4 agonist that supports potent adjuvant effects with improved tolerability relative to native lipid A, providing a mechanistic foundation for TLR4-oriented adjuvant strategies in pertussis vaccine redesign [76].
9.3. Redesign Target: Broaden Antigenic Breadth and Present Antigens in “Pathogen-Like” Formats (Omv, Biofilm/Clinical Isolate Relevance)
Another immunopathogenesis-informed limitation of current aP vaccines is their narrow antigen set and potential mismatch with evolving circulating strains, alongside suboptimal induction of mucosal cellular immunity [56,72,73]. To address this, outer membrane vesicle (OMV)–based vaccines aim to present multiple antigens in their native membrane context (including innate-stimulatory components), thereby improving breadth and functional quality of immunity.
OMV-based pertussis vaccine candidates have demonstrated long-lasting protection and cross-protection across strain genotypes in preclinical work [77]. Critically, intranasal OMV vaccination has been shown to induce mucosal IgA and Th17-mediated responses and—most importantly—prevent colonization in both lungs and nasal cavity, which maps directly onto the “stop transmission” design target derived from immune pathogenesis [78].
9.4. Redesign Target: Consider Live-Attenuated Nasal Vaccines to Recapitulate Infection-Like Immunity Safely
Because natural infection induces the most complete mucosal immune program (including airway T_RM), live-attenuated nasal vaccines are a rational extension of immune-pathogenesis principles—provided safety and genetic stability are addressed [47,72]. BPZE1 is a leading example, designed to induce both mucosal and systemic immunity. Human clinical data have now advanced substantially: BPZE1 has demonstrated immunogenicity and acceptable safety in adults in clinical studies [79], and a controlled human infection model phase 2b trial reported that a single nasal administration could prevent or substantially reduce colonization by virulent B. pertussis—a direct test of the transmission-blocking goal [80].
9.5. Structured Comparison of Next-Generation Pertussis Vaccine Platforms
To operationalize the immune-pathogenesis blueprint, Table 1 contrasts major vaccine platforms using a consistent set of immune correlates that are repeatedly linked to sterilizing immunity in the upper airway (mucosal IgA, Th1/Th17 polarization, and airway tissue-resident memory T cells), together with the most policy-relevant outcome: prevention of nasopharyngeal colonization and onward transmission.
10. Resistance to Antibiotics in Bordetella pertussis
Macrolides have long represented the cornerstone of both treatment and post-exposure prophylaxis for B. pertussis infection. Erythromycin, azithromycin, and clarithromycin are recommended as first-line agents owing to their activity against B. pertussis, favorable penetration into respiratory secretions, and demonstrated efficacy in shortening infectiousness and reducing secondary transmission [81,82]. For decades, antimicrobial resistance in B. pertussis was considered negligible; however, over the past fifteen years, the emergence and expansion of macrolide-resistant B. pertussis (MRBP) has fundamentally altered this paradigm.
10.1. Molecular Mechanisms of Macrolide Resistance
Macrolide resistance in B. pertussis is now well characterized at the molecular level and is primarily mediated by a point mutation in domain V of the 23S rRNA gene, most commonly the A2047G substitution (Escherichia coli numbering) [83]. This mutation induces a conformational alteration of the ribosomal macrolide-binding site, effectively preventing antibiotic binding and resulting in high-level resistance. Phenotypically, resistant isolates typically exhibit minimum inhibitory concentrations (MICs) exceeding 256 μg/mL for erythromycin, azithromycin, and clarithromycin, levels far above achievable serum or tissue concentrations [83,84].
Importantly, B. pertussis possesses three copies of the 23S rRNA gene, and high-level resistance generally requires mutation of all operons. This biological constraint likely explains the historical rarity of resistance and underscores why macrolide resistance emerged relatively late despite widespread antibiotic use. Once resistant clones with full operon mutation became established, however, rapid clonal expansion has been observed [38]. Notably, standardized interpretive criteria for antimicrobial susceptibility testing of B. pertussis have not been established by either CLSI or EUCAST, and confirmation of resistance relies on molecular detection of resistance mutations or demonstration of exceptionally high MICs in vitro [85].
10.2. Global Epidemiology of Macrolide-Resistant B. pertussis
China has emerged as the global epicenter of MRBP. Following the first reports of macrolide-resistant isolates in 2011, surveillance studies have demonstrated a dramatic and sustained increase in prevalence, with resistance rates exceeding 80–90% in multiple regions and approaching fixation in some northern provinces [83,86]. This extraordinary prevalence is widely attributed to intense selective pressure from macrolide overuse, combined with clonal dissemination of resistant lineages. In such settings, macrolide therapy is likely to be ineffective for both treatment and prophylaxis, substantially undermining outbreak control efforts.
Outside China, MRBP has been reported less frequently but across an expanding geographic range. In Japan, resistant isolates have been increasingly detected, including a fatal infant case confirmed to be caused by macrolide-resistant B. pertussis, underscoring the potential clinical consequences of resistance in high-risk populations [87]. In Europe, resistance remains rare but is no longer confined to isolated detections; Finland reported a resistant isolate in national surveillance in 2024, and France documented localized clusters during the same period [88]. These findings indicate that MRBP is emerging within Europe, even if prevalence remains low at present.
In the Americas, reports of MRBP have been historically sporadic; however, recent regional alerts have documented resistant isolates in several countries, including Brazil, Mexico, Peru, and the United States, suggesting wider circulation than previously appreciated [89]. Similarly, hospital-based surveillance in Hong Kong during 2023–2024 revealed a predominance of resistant strains among circulating B. pertussis isolates, raising concern that localized dominance of MRBP may develop outside mainland China under favorable conditions [90].
11. Clinical and Public Health Implications
The emergence of MRBP poses significant clinical and public health challenges. Although vaccination remains the primary tool for pertussis prevention, breakthrough infections occur due to waning immunity and incomplete protection against colonization. In this context, effective antimicrobial therapy is essential for reducing disease severity, preventing transmission, and protecting vulnerable contacts, particularly infants. Macrolide resistance eliminates the effectiveness of all first-line agents, as resistant strains demonstrate cross-resistance to erythromycin, azithromycin, and clarithromycin [83].
Trimethoprim–sulfamethoxazole (TMP-SMX) remains the only validated alternative therapy and is recommended for patients aged ≥2 months when macrolide resistance is suspected or confirmed [81]. However, TMP-SMX is contraindicated in neonates and during pregnancy, precisely the populations at highest risk for severe pertussis outcomes. Although fluoroquinolones and tetracyclines exhibit in vitro activity against B. pertussis, their use is limited by safety concerns and the absence of robust clinical efficacy data, particularly in children [85].
Beyond individual patient management, MRBP threatens the effectiveness of chemoprophylaxis strategies in households, neonatal units, and healthcare settings. Failure to eradicate carriage in index cases increases the risk of sustained transmission, potentially amplifying outbreaks even in highly vaccinated populations.
Given that MRBP is largely driven by a ribosomal target-site mutation, a complementary strategy is systematic in vitro screening of (i) newer macrolide derivatives (e.g., ketolides) with improved ribosomal interactions and (ii) adjuvant compounds that increase intracellular drug exposure (e.g., efflux pump inhibitors or membrane-permeabilizing potentiators) in combination with macrolides. Standard checkerboard assays and time–kill kinetics against contemporary MRBP and macrolide-susceptible isolates, alongside 23S rRNA genotyping, would enable quantification of synergy [fractional inhibitory concentration, (FIC) indices] and selection of candidates for in vivo validation.
In regions where MRBP prevalence is high or during outbreaks with confirmed A2047G strains, management should incorporate (a) rapid molecular resistance detection when available (e.g., 23S rRNA domain V genotyping), (b) age- and pregnancy-specific selection of the best-supported alternative agents, and (c) explicit acknowledgement of evidence gaps (especially neonates and pregnancy). Table 2 summarizes pragmatic options aligned with existing guidance and the limited MRBP-specific clinical evidence base [54,68,91,92,93,94,95,96].
Because clinical benefit from antibiotics in pertussis is greatest when given early and because antimicrobial therapy is also used to reduce infectiousness, the emergence of MRBP creates a disproportionately high-risk scenario for groups in whom (i) bacterial burden is highest (young infants), (ii) complications are more frequent (pregnancy-related exposure of neonates; immunocompromised hosts), and (iii) validated non-macrolide options are restricted by safety.
12. Implications for Surveillance, Stewardship, and Prevention
The emergence and expansion of MRBP highlight the urgent need for enhanced pertussis surveillance incorporating molecular resistance detection and genomic epidemiology. Routine sequencing of circulating isolates would enable early identification of resistant clones and facilitate tracking of their spread at national and international levels [38,88]. Treatment guidelines may require revision in regions where resistance becomes prevalent, balancing the use of alternative agents against safety considerations.
From an antimicrobial stewardship perspective, the Chinese experience serves as a cautionary example of how high antibiotic pressure can rapidly select for and entrench resistance in B. pertussis. Judicious macrolide use, combined with robust vaccination programs, is likely to be essential in preventing similar scenarios elsewhere [86]. Given increasing global mobility, international coordination in surveillance and reporting will be critical to limit dissemination of resistant strains.
13. Emerging Issues and Research Directions
A growing body of experimental and clinical evidence indicates that exposure of B. pertussis to host immune serum induces profound phenotypic and functional changes, underscoring the dynamic nature of host–pathogen interactions and revealing underexplored mechanisms of immune-mediated bacterial modulation. Serum-exposed B. pertussis organisms display altered expression of multiple surface-associated factors, including reduced hemadsorptive fimbrial activity, diminished expression of phase I virulence components, and impaired attachment to ciliated respiratory epithelial cells [89,90]. These observations suggest that humoral immune pressure not only neutralizes bacterial toxins but can directly interfere with colonization capacity, a concept with important implications for vaccine-induced protection.
Early immunological studies demonstrated that serum exposure leads to the release or downregulation of surface components involved in adhesion and immune evasion, highlighting a potential antibody-driven “phenotypic silencing” of virulence traits [89]. Among the immune-reactive components identified, the so-called leukocytosis-promoting factor—now recognized as pertussis toxin—was shown to induce lymphocyte activation and serum agglutinins capable of protecting mice against intranasal challenge [97]. Importantly, reciprocal protection models established that serum agglutinins and pertussis toxin–mediated immune effects represent independent mechanisms of protection, emphasizing the multiplicity of immune pathways involved in host defense [66].
Species-specific differences in lymphocyte responsiveness to B. pertussis antigens further highlight the complexity of immune recognition. While human, canine, and primate lymphocytes exhibit proliferative or agglutinating responses following exposure to pertussis toxin or immune serum, bovine lymphocytes do not, suggesting host-restricted immune pathways shaped by co-evolution with an exclusively human pathogen [66]. Notably, optimal lymphocyte responses in experimental systems were achieved in the presence of non-specific mitogens such as concanavalin A, pokeweed mitogen, or Bacillus Calmette–Guérin (BCG), rather than Bordetella-specific antigens alone, underscoring the importance of innate immune co-stimulation in shaping effective adaptive responses [66]. This observation directly anticipates contemporary efforts to redesign pertussis vaccines using potent innate immune adjuvants that better mimic infection-induced signaling.
At the mucosal level, the interaction between B. pertussis and the respiratory epithelium remains a critical research focus. The bacterium exhibits a strong tropism for ciliated epithelial cells, and early colonization rapidly impairs ciliary function, as demonstrated in tracheal organ culture models [98]. Within hours of inoculation, B. pertussis expresses hemadsorptive fimbriae, closely associates with specialized epithelial surface structures, and releases cytotoxic factors—including tracheal cytotoxin—that damage both ciliated and non-ciliated cells [99]. These processes collectively undermine mucociliary clearance, establishing a permissive environment for persistent infection and transmission.
Human observational studies provide compelling evidence for the protective role of mucosal antibodies. In infants with pertussis, higher concentrations of B. pertussis–specific IgA in nasal secretions are inversely correlated with disease severity, supporting a central role for mucosal IgA in modulating clinical outcomes [100]. In animal models, both IgA and IgG have been shown to confer protection; however, they act at different stages of infection. Secretory IgA primarily interferes with bacterial attachment and epithelial colonization, whereas IgG acts earlier by neutralizing bacteria and toxins before stable adherence to the respiratory tract occurs [101]. These distinctions have profound implications for vaccine design, as current acellular vaccines induce strong systemic IgG responses but limited mucosal IgA, thereby protecting against severe disease without reliably preventing colonization or transmission.
Additional evidence suggests that cellular immunity contributes to control of B. pertussis beyond antibody-mediated mechanisms. The proliferation of intracellular or closely associated bacteria can be inhibited by lymphoid cells derived from fetal or immunoglobulin-depleted neonatal lymph nodes, indicating that non-antibody cellular mechanisms may restrict bacterial persistence [102]. Among B. pertussis products, leukoagglutinin activity—historically attributed to pertussis toxin—has been identified as the primary factor capable of interfering with the bactericidal activity of normal human serum, leading to flocculation of a subset of the bacterial population in vivo [29]. This selective interaction suggests population heterogeneity during infection and raises important questions regarding bacterial subpopulations that may preferentially evade immune clearance.
14. Future research priorities
Collectively, these findings highlight several critical directions for future research. First, there is a need to better characterize how antibody pressure modulates B. pertussis phenotype and virulence gene expression during infection and following vaccination. Second, the differential roles of mucosal IgA, systemic IgG, and cellular immunity in preventing disease versus blocking transmission require further elucidation in both human studies and refined animal models. Third, understanding species-specific immune responses may inform the development of more predictive experimental systems for vaccine evaluation. Finally, integrating these immunological insights into vaccine design—particularly strategies that enhance mucosal immunity and infection-like innate signaling—remains a central challenge for achieving durable, transmission-blocking protection against pertussis.
Author Contributions
Conceptualization, X.X. and Y.Y.; methodology, X.X.; software, X.X.; validation, X.X., Y.Y. and Z.Z.; formal analysis, X.X.; investigation, X.X.; resources, X.X.; data curation, X.X.; writing—original draft preparation, X.X.; writing—review and editing, X.X.; visualization, X.X.; supervision, X.X.; project administration, X.X.; funding acquisition, Y.Y. All authors have read and agreed to the published version of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Melvin JA, Scheller EV, Miller JF, Cotter PA. B. pertussispathogenesis: current and future challenges. Nat Rev Microbiol. 2014;12(4):274-88. [CrossRef]
- Kapil P, Merkel TJ. Pertussis vaccines and protective immunity. Curr Opin Immunol. 2019;59:72-78. [CrossRef]
- Schwartz KL, Kwong JC, Deeks SL, Campitelli MA, Jamieson FB, Marchand-Austin A, Stukel TA, Rosella L, Daneman N, Bolotin S, Drews SJ, Rilkoff H, Crowcroft NS. Effectiveness of pertussis vaccination and duration of immunity. CMAJ. 2016;188(16):E399-E406. [CrossRef]
- Seppälä E, Bråthen Kristoffersen A, Bøås H, Frimann Vestrheim D, Greve-Isdahl M, Freiesleben De Blasio B, Steens A. Pertussis epidemiology including direct and indirect effects of the childhood pertussis booster vaccinations, Norway, 1998-2019. Vaccine. 2022;40(23):3142-3149. [CrossRef]
- Burdin N, Handy LK, Plotkin SA. What Is Wrong with Pertussis Vaccine Immunity? The Problem of Waning Effectiveness of Pertussis Vaccines. Cold Spring Harb Perspect Biol. 2017;9(12):a029454. [CrossRef]
- Mooi FR, Van Der Maas NA, De Melker HE. Pertussis resurgence: waning immunity and pathogen adaptation - two sides of the same coin. Epidemiol Infect. 2014;142(4):685-94. [CrossRef]
- Kardos P, Correia de Sousa J, Heininger U, Konstantopoulos A, MacIntyre CR, Middleton D, Nolan T, Papi A, Rendon A, Rizzo A, Sampson K, Sette A, Sobczyk E, Tan T, Weil-Olivier C, Weinberger B, Wilkinson T, Wirsing von König CH. Understanding the impact of adult pertussis and current approaches to vaccination: A narrative review and expert panel recommendations. Hum Vaccin Immunother. 2024;20(1):2324547. [CrossRef]
- Hodder SL, Mortimer EA Jr. Epidemiology of pertussis and reactions to pertussis vaccine. Epidemiol Rev. 1992;14:243-67. [CrossRef]
- Gregg KA, Merkel TJ. Pertussis Toxin: A Key Component in Pertussis Vaccines? Toxins (Basel). 2019 Sep 21;11(10):557. [CrossRef]
- Duda-Madej A, Łabaz J, Topola E, Bazan H, Viscardi S. Pertussis-A Re-Emerging Threat Despite Immunization: An Analysis of Vaccine Effectiveness and Antibiotic Resistance. Int J Mol Sci. 2025;26(19):9607. [CrossRef]
- Sheng Y, Ma S, Zhou Q, Xu J. Pertussis resurgence: epidemiological trends, pathogenic mechanisms, and preventive strategies. Front Immunol. 2025;16:1618883. [CrossRef]
- Macina D, Evans KE. B. pertussisin School-Age Children, Adolescents and Adults: A Systematic Review of Epidemiology and Mortality in Europe. Infect Dis Ther. 2021;10(4):2071-2118. [CrossRef]
- Zughaier SM, Shafer WM, Stephens DS. Antimicrobial peptides and endotoxin inhibit cytokine and nitric oxide release but amplify respiratory burst response in human and murine macrophages. Cell Microbiol. 2005;7(9):1251-62. [CrossRef]
- Marr N, Hajjar AM, Shah NR, Novikov A, Yam CS, Caroff M, Fernandez RC. Substitution of the B. pertussislipid A phosphate groups with glucosamine is required for robust NF-kappaB activation and release of proinflammatory cytokines in cells expressing human but not murine Toll-like receptor 4-MD-2-CD14. Infect Immun. 2010;78(5):2060-9. [CrossRef]
- Caroff M, Deprun C, Richards JC, Karibian D. Structural characterization of the lipid A of B. pertussis1414 endotoxin. J Bacteriol. 1994;176(16):5156-9. [CrossRef]
- Maeshima N, Evans-Atkinson T, Hajjar AM, Fernandez RC. B. pertussisLipid A Recognition by Toll-like Receptor 4 and MD-2 Is Dependent on Distinct Charged and Uncharged Interfaces. J Biol Chem. 2015;290(21):13440-53. [CrossRef]
- Carbonetti NH. Bordetella pertussis: new concepts in pathogenesis and treatment. Curr Opin Infect Dis. 2016;29(3):287-94. [CrossRef]
- Mattoo S, Cherry JD. Molecular pathogenesis, epidemiology, and clinical manifestations of respiratory infections due to B. pertussisand other Bordetella subspecies. Clin Microbiol Rev. 2005;18(2):326-82. [CrossRef]
- Rossi Paccani S, Benagiano M, Capitani N, Zornetta I, Ladant D, Montecucco C, D’Elios MM, Baldari CT. The adenylate cyclase toxins of Bacillus anthracis and B. pertussispromote Th2 cell development by shaping T cell antigen receptor signaling. PLoS Pathog. 2009;5(3):e1000325. [CrossRef]
- Locht C. Molecular aspects of B. pertussispathogenesis. Int Microbiol. 1999;2(3):137-44.
- Relman DA, Domenighini M, Tuomanen E, Rappuoli R, Falkow S. Filamentous hemagglutinin of Bordetella pertussis: nucleotide sequence and crucial role in adherence. Proc Natl Acad Sci U S A. 1989;86(8):2637-41. [CrossRef]
- Geuijen CA, Willems RJ, Bongaerts M, Top J, Gielen H, Mooi FR. Role of the B. pertussisminor fimbrial subunit, FimD, in colonization of the mouse respiratory tract. Infect Immun. 1997;65(10):4222-8. [CrossRef]
- Cherry JD, Gornbein J, Heininger U, Stehr K. A search for serologic correlates of immunity to B. pertussiscough illnesses. Vaccine. 1998;16(20):1901-6. [CrossRef]
- Chen Q, Stibitz S. The BvgASR virulence regulon of Bordetella pertussis. Curr Opin Microbiol. 2019;47:74-81. [CrossRef]
- Burns DL. Secretion of Pertussis Toxin from Bordetella pertussis. Toxins (Basel). 2021;13(8):574. [CrossRef]
- Cundell DR, Kanthakumar K, Taylor GW, Goldman WE, Flak T, Cole PJ, Wilson R. Effect of tracheal cytotoxin from B. pertussison human neutrophil function in vitro. Infect Immun. 1994;62(2):639-43. [CrossRef]
- Tuomanen EI, Hendley JO. Adherence of B. pertussisto human respiratory epithelial cells. J Infect Dis. 1983;148(1):125-30. [CrossRef]
- Leininger E, Roberts M, Kenimer JG, Charles IG, Fairweather N, Novotny P, Brennan MJ. Pertactin, an Arg-Gly-Asp-containing B. pertussissurface protein that promotes adherence of mammalian cells. Proc Natl Acad Sci U S A. 1991;88(2):345-9. [CrossRef]
- Carbonetti NH. Contribution of pertussis toxin to the pathogenesis of pertussis disease. Pathog Dis. 2015;73(8):ftv073. [CrossRef]
- Hewlett EL, Kim KJ, Lee SJ, Gray MC. Adenylate cyclase toxin from Bordetella pertussis: current concepts and problems in the study of toxin functions. Int J Med Microbiol. 2000;290(4-5):333-5. [CrossRef]
- Guiso N. Bordetella Adenylate Cyclase-Hemolysin Toxins. Toxins (Basel). 2017;9(9):277. [CrossRef]
- Graf R, Codina J, Birnbaumer L. Peptide inhibitors of ADP-ribosylation by pertussis toxin are substrates with affinities comparable to those of the trimeric GTP-binding proteins. Mol Pharmacol. 199242(5):760-4.
- Heiss LN, Lancaster JR Jr, Corbett JA, Goldman WE. Epithelial autotoxicity of nitric oxide: role in the respiratory cytopathology of pertussis. Proc Natl Acad Sci U S A. 1994;91(1):267-70. [CrossRef]
- Eberhardt CS, Siegrist CA. What Is Wrong with Pertussis Vaccine Immunity? Inducing and Recalling Vaccine-Specific Immunity. Cold Spring Harb Perspect Biol. 2017;9(12):a029629. [CrossRef]
- Guiso N, Koenig CHWV. Pertussis Vaccination: The Challenges Ahead. Semin Respir Crit Care Med. 2025;46(1):14-19. [CrossRef]
- Ricci S, Rappuoli R, Scarlato V. The pertussis toxin liberation genes of B. pertussisare transcriptionally linked to the pertussis toxin operon. Infect Immun. 1996;64(4):1458-60. [CrossRef]
- Lam C, Octavia S, Ricafort L, Sintchenko V, Gilbert GL, Wood N, McIntyre P, Marshall H, Guiso N, Keil AD, Lawrence A, Robson J, Hogg G, Lan R. Rapid increase in pertactin-deficient B. pertussisisolates, Australia. Emerg Infect Dis. 2014;20(4):626-33. [CrossRef]
- Bart MJ, Harris SR, Advani A, Arakawa Y, Bottero D, Bouchez V, Cassiday PK, Chiang CS, Dalby T, Fry NK, Gaillard ME, van Gent M, Guiso N, Hallander HO, Harvill ET, He Q, van der Heide HG, Heuvelman K, Hozbor DF, Kamachi K, Karataev GI, Lan R, Lutyńska A, Maharjan RP, Mertsola J, Miyamura T, Octavia S, Preston A, Quail MA, Sintchenko V, Stefanelli P, Tondella ML, Tsang RS, Xu Y, Yao SM, Zhang S, Parkhill J, Mooi FR. Global population structure and evolution of B. pertussisand their relationship with vaccination. mBio. 2014;5(2):e01074. [CrossRef]
- Park J, Zhang Y, Buboltz AM, Zhang X, Schuster SC, Ahuja U, Liu M, Miller JF, Sebaihia M, Bentley SD, Parkhill J, Harvill ET. Comparative genomics of the classical Bordetella subspecies: the evolution and exchange of virulence-associated diversity amongst closely related pathogens. BMC Genomics. 2012;13:545. [CrossRef]
- Confer DL, Eaton JW. Phagocyte impotence caused by an invasive bacterial adenylate cyclase. Science. 1982;217(4563):948-50. [CrossRef]
- Bouchez V, Douché T, Dazas M, Delaplane S, Matondo M, Chamot-Rooke J, Guiso N. Characterization of Post-Translational Modifications and Cytotoxic Properties of the Adenylate-Cyclase Hemolysin Produced by Various B. pertussisand Bordetella parapertussis Isolates. Toxins (Basel). 2017;9(10):304. [CrossRef]
- Kroes MM, Miranda-Bedate A, Jacobi RHJ, van Woudenbergh E, den Hartog G, van Putten JPM, de Wit J, Pinelli E. Bordetella pertussis-infected innate immune cells drive the anti-pertussis response of human airway epithelium. Sci Rep. 2022;12(1):3622. [CrossRef]
- Goldman WE, Klapper DG, Baseman JB. Detection, isolation, and analysis of a released B. pertussisproduct toxic to cultured tracheal cells. Infect Immun. 1982;36(2):782-94. [CrossRef]
- Rosenthal RS, Nogami W, Cookson BT, Goldman WE, Folkening WJ. Major fragment of soluble peptidoglycan released from growing B. pertussisis tracheal cytotoxin. Infect Immun. 1987;55(9):2117-20. [CrossRef]
- Cookson BT, Tyler AN, Goldman WE. Primary structure of the peptidoglycan-derived tracheal cytotoxin of Bordetella pertussis. Biochemistry. 1989;28(4):1744-9. [CrossRef]
- Heiss LN, Flak TA, Lancaster JR Jr, McDaniel ML, Goldman WE. Nitric oxide mediates B. pertussistracheal cytotoxin damage to the respiratory epithelium. Infect Agents Dis. 1993 Aug;2(4):173-7.
- Solans L, Locht C. The Role of Mucosal Immunity in Pertussis. Front Immunol. 2019 Jan 14;9:3068. [CrossRef]
- Smith AM, Guzmán CA, Walker MJ. The virulence factors of Bordetella pertussis: a matter of control. FEMS Microbiol Rev. 2001 May;25(3):309-33. [CrossRef]
- Wendelboe AM, Njamkepo E, Bourillon A, Floret DD, Gaudelus J, Gerber M, Grimprel E, Greenberg D, Halperin S, Liese J, Muñoz-Rivas F, Teyssou R, Guiso N, Van Rie A; Infant Pertussis Study Group. Transmission of B. pertussisto young infants. Pediatr Infect Dis J. 2007 Apr;26(4):293-9. [CrossRef]
- Cherry JD. The epidemiology of pertussis: a comparison of the epidemiology of the disease pertussis with the epidemiology of B. pertussisinfection. Pediatrics. 2005;115(5):1422-7. [CrossRef]
- Bartkus JM, Juni BA, Ehresmann K, Miller CA, Sanden GN, Cassiday PK, Saubolle M, Lee B, Long J, Harrison AR Jr, Besser JM. Identification of a mutation associated with erythromycin resistance in Bordetella pertussis: implications for surveillance of antimicrobial resistance. J Clin Microbiol. 2003 Mar;41(3):1167-72. [CrossRef]
- Wang Z, Han R, Liu Y, Du Q, Liu J, Ma C, Li H, He Q, Yan Y. Direct Detection of Erythromycin-Resistant B. pertussisin Clinical Specimens by PCR. J Clin Microbiol. 2015 Nov;53(11):3418-22. [CrossRef]
- Petráčková D, Farman MR, Amman F, Linhartová I, Dienstbier A, Kumar D, Držmíšek J, Hofacker I, Rodriguez ME, Večerek B. Transcriptional profiling of human macrophages during infection with Bordetella pertussis. RNA Biol. 2020 May;17(5):731-742. [CrossRef]
- Zhang Q, Li M, Wang L, Xin T, He Q. High-resolution melting analysis for the detection of two erythromycin-resistant B. pertussisstrains carried by healthy schoolchildren in China. Clin Microbiol Infect. 2013;19(6):E260-2. [CrossRef]
- Wendelboe AM, Van Rie A, Salmaso S, Englund JA. Duration of immunity against pertussis after natural infection or vaccination. Pediatr Infect Dis J. 2005;24(5 Suppl):S58-61. [CrossRef]
- Warfel JM, Edwards KM. Pertussis vaccines and the challenge of inducing durable immunity. Curr Opin Immunol. 2015;35:48-54. [CrossRef]
- Higgins SC, Jarnicki AG, Lavelle EC, Mills KH. TLR4 mediates vaccine-induced protective cellular immunity to Bordetella pertussis: role of IL-17-producing T cells. J Immunol. 2006;177(11):7980-9. [CrossRef]
- Fedele G, Nasso M, Spensieri F, Palazzo R, Frasca L, Watanabe M, Ausiello CM. Lipopolysaccharides from B. pertussisand Bordetella parapertussis differently modulate human dendritic cell functions resulting in divergent prevalence of Th17-polarized responses. J Immunol. 2008;181(1):208-16. [CrossRef]
- Higgs R, Higgins SC, Ross PJ, Mills KH. Immunity to the respiratory pathogen Bordetella pertussis. Mucosal Immunol. 2012;5(5):485-500. [CrossRef]
- Ross PJ, Sutton CE, Higgins S, Allen AC, Walsh K, Misiak A, Lavelle EC, McLoughlin RM, Mills KH. Relative contribution of Th1 and Th17 cells in adaptive immunity to Bordetella pertussis: towards the rational design of an improved acellular pertussis vaccine. PLoS Pathog. 2013;9(4):e1003264. [CrossRef]
- Warfel JM, Zimmerman LI, Merkel TJ. Acellular pertussis vaccines protect against disease but fail to prevent infection and transmission in a nonhuman primate model. Proc Natl Acad Sci U S A. 2014;111(2):787-92. [CrossRef]
- Rodríguez ME, Hellwig SM, Pérez Vidakovics ML, Berbers GA, van de Winkel JG. B. pertussisattachment to respiratory epithelial cells can be impaired by fimbriae-specific antibodies. FEMS Immunol Med Microbiol. 2006;46(1):39-47. [CrossRef]
- Guiso N, Capiau C, Carletti G, Poolman J, Hauser P. Intranasal murine model of B. pertussisinfection. I. Prediction of protection in human infants by acellular vaccines. Vaccine. 1999;17(19):2366-76. [CrossRef]
- Belcher T, Dubois V, Rivera-Millot A, Locht C, Jacob-Dubuisson F. Pathogenicity and virulence of B. pertussis and its adaptation to its strictly human host. Virulence. 2021;12(1):2608-2632. [CrossRef]
- Weiss AA, Patton AK, Millen SH, Chang SJ, Ward JI, Bernstein DI. Acellular pertussis vaccines and complement killing of Bordetella pertussis. Infect Immun. 2004;72(12):7346-51. [CrossRef]
- Mills KH. Immunity to Bordetella pertussis. Microbes Infect. 2001 Jul;3(8):655-77. [CrossRef]
- von Olleschik-Elbheim L, el Bayâ A, Schmidt MA. ADP-ribosylation of alpha-Gi proteins by pertussis toxin. Positional dissection of acceptor sites using membrane anchored synthetic peptides. Methods Mol Biol. 2000;145:203-17. [CrossRef]
- Carbonetti NH. Pertussis leukocytosis: mechanisms, clinical relevance and treatment. Pathog Dis. 2016;74(7):ftw087. [CrossRef]
- Luker KE, Tyler AN, Marshall GR, Goldman WE. Tracheal cytotoxin structural requirements for respiratory epithelial damage in pertussis. Mol Microbiol. 1995;16(4):733-43. [CrossRef]
- Allen AC, Wilk MM, Misiak A, Borkner L, Murphy D, Mills KHG. Sustained protective immunity against B. pertussisnasal colonization by intranasal immunization with a vaccine-adjuvant combination that induces IL-17-secreting TRM cells. Mucosal Immunol. 2018;11(6):1763-1776. [CrossRef]
- Cherry JD. Why do pertussis vaccines fail? Pediatrics. 2012;129(5):968-70. [CrossRef]
- Ni Chasaide C, Mills KHG. Next-Generation Pertussis Vaccines Based on the Induction of Protective T Cells in the Respiratory Tract. Vaccines (Basel). 2020;8(4):621. [CrossRef]
- Kapil P, Merkel TJ. Pertussis vaccines and protective immunity. Curr Opin Immunol. 2019;59:72-78. [CrossRef]
- Boehm DT, Wolf MA, Hall JM, Wong TY, Sen-Kilic E, Basinger HD, Dziadowicz SA, de la Paz Gutierrez M, Blackwood CB, Bradford SD, Begley KA, Witt WT, Varney ME, Barbier M, Damron FH. Intranasal acellular pertussis vaccine provides mucosal immunity and protects mice from Bordetella pertussis. NPJ Vaccines. 2019;4:40. [CrossRef]
- Misiak A, Leuzzi R, Allen AC, Galletti B, Baudner BC, D’Oro U, O’Hagan DT, Pizza M, Seubert A, Mills KHG. Addition of a TLR7 agonist to an acellular pertussis vaccine enhances Th1 and Th17 responses and protective immunity in a mouse model. Vaccine. 2017;35(39):5256-5263. [CrossRef]
- Mata-Haro V, Cekic C, Martin M, Chilton PM, Casella CR, Mitchell TC. The vaccine adjuvant monophosphoryl lipid A as a TRIF-biased agonist of TLR4. Science. 2007;316(5831):1628-1632. [CrossRef]
- Gaillard ME, Bottero D, Errea A, Ormazabal M, Zurita ME, Moreno G, Rumbo M, Castuma C, Bartel E, Flores D, van der Ley P, van der Ark A, Hozbor DF. Acellular pertussis vaccine based on outer membrane vesicles capable of conferring both long-lasting immunity and protection against different strain genotypes. Vaccine. 2014;32(8):931-937. [CrossRef]
- Raeven RHM, Rockx-Brouwer D, Kanojia G, van der Maas L, Bindels THE, Ten Have R, van Riet E, Metz B, Kersten GFA. Intranasal immunization with outer membrane vesicle pertussis vaccine confers broad protection through mucosal IgA and Th17 responses. Sci Rep. 2020;10(1):7396. [CrossRef]
- Keech C, Miller VE, Rizzardi B, Hoyle C, Pryor MJ, Ferrand J, Solovay K, Thalen M, Noviello S, Goldstein P, Gorringe A, Cavell B, He Q, Barkoff AM, Rubin K, Locht C. Immunogenicity and safety of BPZE1, an intranasal live attenuated pertussis vaccine, versus tetanus-diphtheria-acellular pertussis vaccine: a randomised, double-blind, phase 2b trial. Lancet. 2023;401(10379):843-855. [CrossRef]
- Gbesemete D, Ramasamy MN, Ibrahim M, Hill AR, Raud L, Ferreira DM, Guy J, Dale AP, Laver JR, Coutinho T, Faust SN, Reed TAN, Babbage G, Weissfeld L, Lang W, Locht C, Samal V, Goldstein P, Solovay K, Rubin K, Noviello S, Read RC. Efficacy, immunogenicity, and safety of the live attenuated nasal pertussis vaccine, BPZE1, in the UK: a randomised, placebo-controlled, phase 2b trial using a controlled human infection model with virulent Bordetella pertussis. Lancet Microbe. 2025;6(12):101211. [CrossRef]
- Cherry JD. Pertussis in young infants throughout the world. Clin Infect Dis. 2016 Dec 1;63(Suppl 4):S119-S122. [CrossRef]
- Altunaiji S, Kukuruzovic R, Curtis N, Massie J. Antibiotics for whooping cough (pertussis). Cochrane Database Syst Rev. 2007;2007(3):CD004404. [CrossRef]
- Li L, Deng J, Ma X, Zhou K, Meng Q, Yuan L, Shi W, Wang Q, Li Y, Yao K. High prevalence of macrolide-resistant B. pertussis and ptxP1 genotype, mainland China, 2014-2016. Emerg Infect Dis. 2019;25(12):2205-2214. [CrossRef]
- Wang Z, Li Y, Hou T, Liu X, Liu Y, Yu T, Chen Z, Gao Y, Li H, He Q. Appearance of macrolide-resistant B. pertussis strains in China. Antimicrob Agents Chemother. 2013t;57(10):5193-5194. [CrossRef]
- Fry NK, Duncan J, Vaghji L, George RC, Harrison TG. Antimicrobial susceptibility testing of historical and recent clinical isolates of B. pertussis in the United Kingdom using the Etest method. Eur J Clin Microbiol Infect Dis. 2010;29(9):1183-1185. [CrossRef]
- Mai Q, Wen J, Luo Y, Guo J, Qin Y, Lai W, Deng W, Ji C, Mai R, Zheng M, Chen Z, Chen Y, Gu C, Guo L, Li H, Tang Y, Huang D, Luo M. Molecular epidemiology and increasing macrolide resistance of B. pertussisisolates in Guangzhou, China. BMC Infect Dis. 2025;25(1):1152. [CrossRef]
- Iwasaki T, Koide K, Kido T, Nakagawa S, Goto M, Kenri T, Suzuki H, Otsuka N, Takada H. Fatal case of macrolide-resistant B. pertussis infection, Japan, 2024. J Infect Chemother. 2025 Jul;31(7):102727. [CrossRef] [PubMed]
- Mooi FR. B. pertussis and vaccination: the persistence of a genetically monomorphic pathogen. Infect Genet Evol. 2010;10(1):36-49. [CrossRef]
- Wardlaw AC, Parton R. B. pertussistoxins. Pharmacol Ther. 1982;19(1):1-53. [CrossRef]
- Tuomanen E, Weiss A. Filamentous hemagglutinin and pertussis toxin promote adherence of B. pertussis to cilia. Infect Immun. 1985;50(3):575-579.
- Centers for Disease Control and Prevention (CDC). Treatment of Pertussis (Whooping Cough): Clinical Care. Updated Dec 2, 2025. Available from: https://www.cdc.gov/pertussis/hcp/clinical-care/index.html.
- Tiwari T, Murphy TV, Moran J; National Immunization Program, CDC. Recommended antimicrobial agents for the treatment and postexposure prophylaxis of pertussis: 2005 CDC Guidelines. MMWR Recomm Rep. 2005;54(RR-14):1-16.
- Eberly MD, Eide MB, Thompson JL, Nylund CM. Azithromycin in early infancy and pyloric stenosis. Pediatrics. 2015 Mar;135(3):483-8. [CrossRef] [PubMed] [PubMed Central]
- Bartkus JM, Juni BA, Ehresmann K, Miller CA, Sanden GN, Cassiday PK, Saubolle M, Lee B, Long J, Harrison AR Jr, Besser JM. Identification of a mutation associated with erythromycin resistance in Bordetella pertussis: implications for surveillance of antimicrobial resistance. J Clin Microbiol. 2003 Mar;41(3):1167-72. [CrossRef]
- Ivaska L, Barkoff AM, Mertsola J, He Q. Macrolide Resistance in Bordetella pertussis: Current Situation and Future Challenges. Antibiotics (Basel). 2022;11(11):1570. [CrossRef]
- European Centre for Disease Prevention and Control (ECDC). Antimicrobial susceptibility testing of B. pertussis– Technical report. ECDC; 2022. https://www.ecdc.europa.eu/sites/default/files/documents/bordetella-pertussis-antimicrobial-susceptibility-testing-2022_0.pdf.
- Morse SI, Morse JH. Isolation and properties of the leukocytosis- and lymphocytosis-promoting factor of Bordetella pertussis. J Exp Med. 1976;143(6):1483-1502. [CrossRef]
- Weiss AA, Hewlett EL. Virulence factors of Bordetella pertussis. Annu Rev Microbiol. 1986;40:661-686. [CrossRef]
- Redhead K, Watkins J, Barnard A, Mills KH. Effective immunization against B. pertussis respiratory infection in mice is dependent on induction of cell-mediated immunity. Infect Immun. 1993;61(8):3190-3198. [CrossRef]
- Mills KH, Barnard A, Watkins J, Redhead K. Cell-mediated immunity to Bordetella pertussis: role of Th1 cells in bacterial clearance in a murine respiratory infection model. Infect Immun. 1993;61(2):399-410. [CrossRef]
- Hellwig SM, van Spriel AB, Schellekens JF, Mooi FR, van de Winkel JG. Immunoglobulin A-mediated protection against B. pertussis infection. Infect Immun. 2001;69(8):4846-4850. [CrossRef]
- Flak TA, Goldman WE. Autotoxicity of nitric oxide in airway disease. Am J Respir Crit Care Med. 1996;154(4 Pt 1):S202-S206.
Figure 1.
Pathogenesis and host immune modulation during B. pertussis infection.
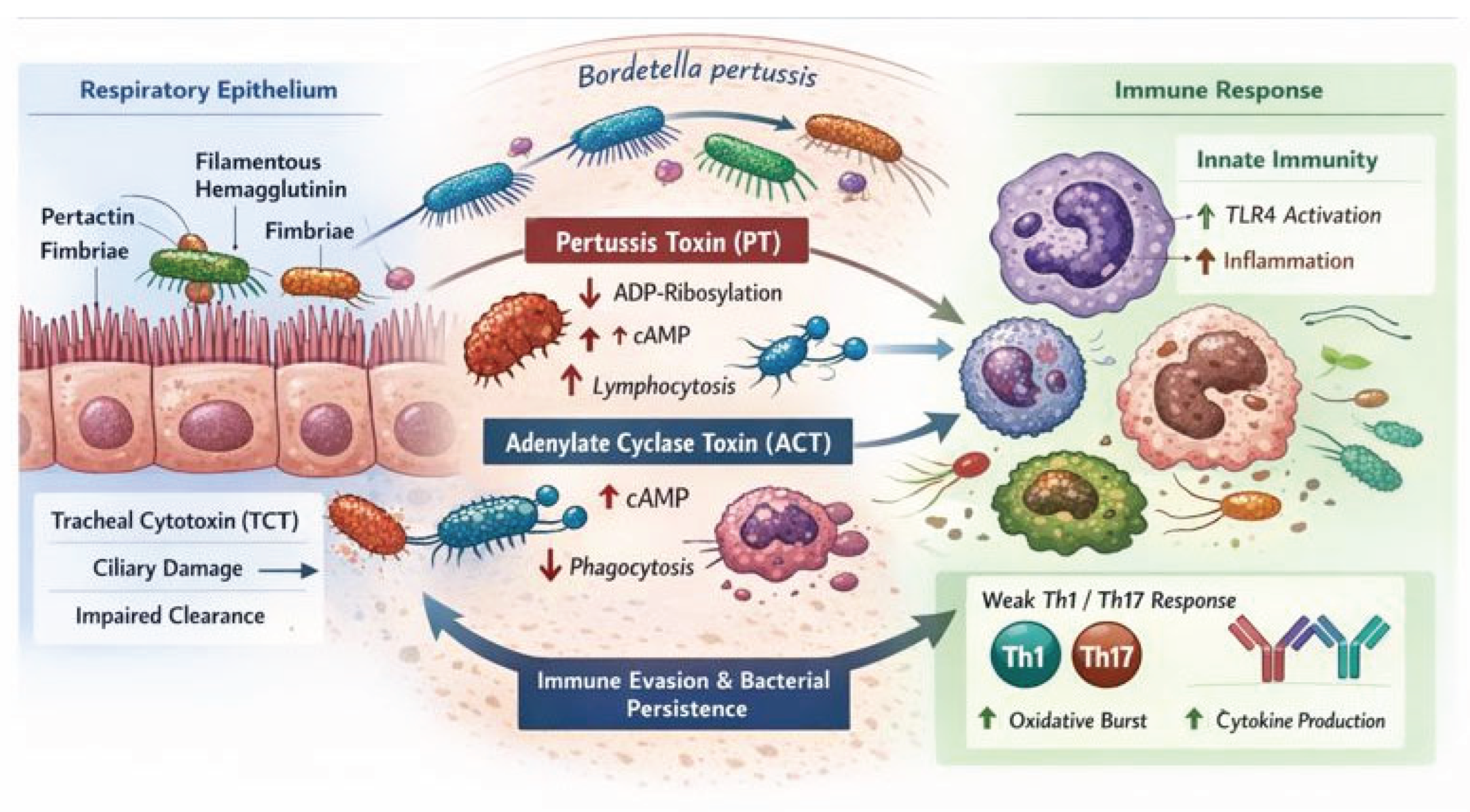

Table 1.
Comparison of licensed and next-generation pertussis vaccine platforms with respect to immune correlates and prevention of colonization and transmission.
Table 1.
Comparison of licensed and next-generation pertussis vaccine platforms with respect to immune correlates and prevention of colonization and transmission.
| Platform (typical route) | Mucosal IgA | Th1/Th17 | Airway T_RM | Prevents colonization / transmission | Key evidence (selected) |
|---|---|---|---|---|---|
| Licensed acellular pertussis (aP; IM) |
Low | Weak (Th2-leaning with alum) | Limited | Poor (protection mainly against disease, not carriage) | Strong serum anti-toxin IgG and reduced severe disease but limited mucosal immunity and weak Th1/Th17; does not reliably block carriage/transmission [56,72,73]. |
| OMV-based pertussis vaccines (IN or IM; often IN in next-gen designs) | High with intranasal delivery | Robust Th17 (and Th1) in preclinical models | Airway-local T-cell memory (preclinical) | Improved; intranasal OMV prevented nasal and lung colonization in mice | Intranasal OMV vaccination induced mucosal IgA and Th17 responses and prevented colonization in both nasal cavity and lungs in mice [78]. |
| BPZE1 live-attenuated nasal vaccine (IN) | High (mucosal + systemic responses) | Th1/Th17-skewed, infection-like programming | Expected strong airway T_RM (supported in models) | Yes/strong signal; reduces or prevents colonization in controlled settings | Human trials show acceptable safety and immunogenicity [79]; a controlled human infection model phase 2b study showed that a single intranasal dose could prevent or substantially reduce colonization by virulent B. pertussis and markedly lower bacterial burden versus placebo [80]. |
| Intranasal subunit strategies (e.g., aP antigens reformulated for IN delivery ± PRR agonists) | Moderate-High (route-dependent) | Improved when paired with innate-programming adjuvants (e.g., TLR agonists) | Potential to seed airway memory (data emerging) | Partial; reduction in upper-airway burden in animals; clinical evidence limited | Intranasal delivery of acellular antigens can elicit mucosal IgA and IL-17A and reduce bacterial burden in upper and lower airways in mice, highlighting the importance of route and adjuvanting to restore mucosal correlates [74,75]. |
Overall, platforms that deliver antigens to the nasal mucosa and provide infection-like innate cues (OMV formulations and live-attenuated BPZE1) most consistently induce the combination of sIgA, Th17-associated mucosal immunity, and airway-local memory associated with control of colonization, whereas standard IM aP vaccination predominantly generates systemic antibodies that mitigate disease severity but leave residual carriage and transmission.
Table 2.
Risk stratification and therapeutic options in high-MRBP settings.
| Population / scenario | Preferred agent(s) when MRBP suspected/confirmed | Key safety constraints | Role of susceptibility testing | Notes in high-MRBP regions |
|---|---|---|---|---|
| Neonate (<1 month) | Azithromycin remains the only guideline-endorsed macrolide option; if MRBP confirmed, individualized management with specialist input | TMP–SMX contraindicated <2 months; erythromycin linked to infantile hypertrophic pyloric stenosis; limited safety/efficacy data for alternatives | Prioritize rapid genotyping (A2047G) + culture/qPCR where feasible | If MRBP is confirmed, there is no well-validated oral alternative; focus on supportive care, infection control, and expert consultation; consider investigational/locally recommended parenteral options only within protocolized care [68,91,92,93] |
| Infant (1–<2 months) | Azithromycin (standard of care); if MRBP confirmed, specialist-led individualized approach | TMP–SMX not recommended <2 months; tetracyclines/fluoroquinolones generally avoided | Genotype/culture to confirm MRBP whenever possible | Highest-risk group for severe disease; hospital monitoring often warranted; prioritize eradication of household source cases [68,91,92,93] |
| ≥2 months (children/adolescents) | TMP–SMX (alternative agent when MRBP suspected/confirmed) | Avoid in sulfonamide allergy; monitor for adverse effects; avoid in late pregnancy if adolescent is pregnant | Genotyping useful to avoid ineffective macrolides; phenotypic MIC testing may support local surveillance | If TMP–SMX cannot be used, evidence for alternatives is limited; some reports suggest in vitro/in vivo activity of certain β-lactam/β-lactamase inhibitor combinations, but these are not standard guideline therapies [54,92,94,95] |
| Adults (non-pregnant) | TMP–SMX when MRBP suspected/confirmed; macrolides only if susceptibility likely/confirmed | Contraindications as above; consider drug interactions | Genotyping/culture where available to guide therapy and prophylaxis | For non-severe disease, the main antibiotic goal is to reduce transmission; if MRBP is circulating, public-health measures and targeted prophylaxis strategies become more important [54,92,94,95] |
| Pregnancy / immediate postpartum | Macrolide (azithromycin) is typically preferred when susceptible; in high-MRBP settings, prioritize susceptibility testing and individualized risk–benefit decisions | TMP–SMX generally avoided (especially 1st trimester and near term); tetracyclines/fluoroquinolones generally avoided in pregnancy | Rapid genotyping is particularly valuable to avoid prolonged ineffective therapy | Because robust MRBP-specific pregnancy-safe alternatives are lacking, prevention (maternal vaccination) + early case identification and infection control are critical; consult obstetrics/ID for any non-standard regimens [91,92,93] |
| Immunocompromised hosts | Treat early; TMP–SMX may be used when MRBP suspected/confirmed if not contraindicated | Higher risk of complications; drug–drug interactions common; monitor closely | Lower threshold for microbiologic confirmation and follow-up sampling | Consider longer observation and follow-up to ensure clearance; integrate with infection control to reduce nosocomial transmission [92,94,95,96] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



