
Submitted:
12 July 2024
Posted:
17 July 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Background: Routine childhood vaccination, e.g., for diphtheria-tetanus-pertussis (DTP), might provide an additional protection towards SARS-CoV-2 infection. This concept of heterologous immunity was explored in healthy children receiving both DTP and inactivated SARS-CoV-2 vaccines. Methods: A cross-sectional study was performed on 154 healthy children aged 6–8 years old in Jakarta, Indonesia. Their vaccination status of DTP (including diphtheria-tetanus booster vaccine at 5 years old) and CoronaVac (from 6 years old) were recorded. Peripheral blood samples were collected from all participants, in which anti-diphtheria toxoid IgG, anti-SARS-CoV-2 S-RBD antibodies and T cell-derived IFN-g were measured. Results: Study participants with complete DTP vaccination had significantly higher titer of anti-diphtheria toxoid IgG than the ones without (median=0.9349 versus 0.2113 IU/mL; p<0.0001). Upon stratification based on DTP and CoronaVac vaccination statuses, participants with complete DTP and CoronaVac vaccinations had the highest titer of anti-SARS-CoV-2 S-RBD antibodies (median=1,196 U/mL) and the highest concentration of SARS-CoV-2-specific T cell-derived IFN-g (median=560.9 mIU/mL) among all groups. Conclusion: Healthy children aged 6-8 years old with complete DTP and CoronaVac vaccinations exhibited higher SARS-CoV-2-specific adaptive immune responses, particularly on their T-cell response. This suggests an additional benefit of routine childhood vaccination to generate protection against novel pathogens, presumably via heterologous immunity.
Keywords:
Children
; SARS-CoV-2-specific immune response
; Heterologous immunity
; Diphtheria-tetanus-pertussis vaccine
; Inactivated COVID-19 vaccine
1. Introduction
The coronavirus disease 2019 (COVID-19), caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infection, is now endemic globally. From early 2020 until June 2024, the cumulative COVID-19 cases in the world and Indonesia have risen to 775,583,309 and 6,829,057 confirmed cases, respectively [1]. Furthermore, the COVID-19-associated mortality cases in the world and Indonesia by early June 2024 were already at 7,050,691 and 162,058 deaths, respectively [1], emphasizing the need for effective prevention and treatment methods against COVID-19. With regards to the preventive measurement, vaccination is one of the most important public health tools. Various types of vaccines are available against symptomatic SARS-CoV-2 infection, including, but not limited to, inactivated virus vaccines (Sinovac’s CoronaVac, Sinopharm’s Covilo and Bharat Biotech’s Covaxin), messenger RNA-based vaccines (Moderna’s Spikevax mRNA-1273 and Pfizer–BioNTech’s Comirnaty BNT162b2), adenovirus vector–based vaccines (AstraZeneca’s Vaxzevria and Covishield ChAdOx1 as well as Johnson & Johnson–Janssen’s Ad26.COV2.S) and adjuvanted protein vaccines (Novavax’s Nuvaxovid and Covovax NVX-CoV2373) [2]. The widespread disparity of vaccine distribution, however, impedes the collective effort to mitigate the severity of COVID-19 by administering the vaccines, in which populations from the low- and middle-income countries suffered disproportionately to this inequality [3,4].
One sub-group of those populations is children under 12 years old. This age group is less prioritized in many countries to be vaccinated because of the limited availability of COVID-19 vaccines, the presumed milder symptoms of SARS-CoV-2 infection among children as well as the incomplete knowledge on risks and benefits offered by pediatric COVID-19 vaccines [5,6,7,8,9]. However, this does not exclude a risk that SARS-CoV-2 infection could cause a severe disease or multisystem inflammatory syndrome in children (MIS-C) [10,11,12]. Several low- and middle-income countries, including Indonesia, indeed had reported relatively high incidences of severe COVID-19 among children, particularly prior the circulation of Omicron variant [13,14,15,16,17], stressing an urgent need of COVID-19 vaccination, at least for this age group in those countries. In addition, SARS-CoV-2-infected children, despite their asymptomatic status, could become a potential reservoir for transmission to the vulnerable family members and the evolution of SARS-CoV-2 variants [18]. Taken together, the policy to vaccinate children under 12 years old against SARS-CoV-2 infection is arguably prudent.
The Ministry of Health of Republic of Indonesia has recommended two types of COVID-19 vaccines for children aged 6-11 years old, i.e., CoronaVac and BNT162b2. However, the pediatric COVID-19 vaccination program in Indonesia is facing significant hurdles, including parental hesitancy and shortage of vaccines’ availability [19,20]. As a part of the national COVID-19 vaccination program, CoronaVac vaccine has been administered for Indonesian children aged 6 years old and above since December 2021. Until June 20, 2024, there were approximately 17.6 million children aged 6-11 years old who had received two doses of CoronaVac [21]. However, its availability has become scarce in Indonesia since October 2022 [22,23], prompting the need for improvised methods to keep protecting the pediatric population in Indonesia against SARS-CoV-2 infection.
One of the proposed efforts is by investigating plausible protection against COVID-19 offered by routine vaccines against unrelated pathogens. This concept is known by heterologous immunity, which is an induction of adaptive immune response by a certain pathogen or vaccine against an unrelated pathogen [24]. Three vaccines have been hypothesized to generate heterologous immunity due to their association with reductions in all-cause infant mortality, i.e., bacillus Calmette-Guérin, measles-containing vaccines as well as diphtheria-tetanus-pertussis (DTP) vaccine. DTP vaccination is of interest due to several reasons, i.e., (i) in silico investigation had reported high similarities between antigens of SARS-CoV-2 and DTP vaccine [25]; (ii) diphtheria and tetanus vaccination history had been associated with lower risk of COVID-19 hospitalization among older population in the UK [26]; and (iii) DTP booster vaccination (using diphtheria-tetanus vaccine in Indonesia) is administered at 5 years old [27], which is in a relatively short time-frame with the starting of CoronaVac vaccination at 6 years old. If heterologous immunity does exist, the immunological memory created by DTP vaccination-booster will be strong enough to be recalled by CoronaVac vaccination.
We therefore investigated the potential heterologous immunity offered by DTP (including diphtheria-tetanus booster) and CoronaVac vaccinations in the healthy children aged 6–8 years old living in Jakarta, Indonesia. Peripheral blood samples were collected from all study participants. Anti-diphtheria immunoglobulin G was quantified as an indicator of successful DTP vaccination-booster. SARS-CoV-2-specific adaptive immune responses were assessed as well, i.e., anti-SARS-CoV-2 S-RBD antibodies and T cell-derived interferon gamma to indicate humoral and cellular immunities, respectively.
2. Materials and Methods
2.1. Study Design and Participants
A cross-sectional study on healthy children aged 6–8 years old was conducted between November 2022 until October 2023, preceded by a questionnaire-based data collection on parents of eligible subjects living in the Senen district, Central Jakarta, Indonesia. The questionnaire-based results had been published [20]. Pertaining to the status of DTP vaccination among the study participants, their anti-diphtheria toxoid antibodies had been published as well (manuscript was accepted).
The inclusion criteria were (i) healthy children aged 6–8 years old and (ii) no confirmed history of COVID-19. The exclusion criteria were (i) obese and having poor nutrition status, (ii) receiving primary DTP immunization less than three doses, (iii) receiving COVID-19 vaccination (using CoronaVac vaccine, an inactivated SARS-CoV-2 and aluminum hydroxide-adjuvanted vaccine [Sinovac Life Sciences, Beijing, China]) less than two doses, (iv) receiving other type of immunization less than a month prior the commencement of the study, (v) contracting disease, e.g., primary immunodeficiency disease, autoimmune disease, cancer, chronic or congenital disease, (vi) receiving medication that could alter immune response in the past 4 weeks, e.g., long-term corticosteroid, intravenous immunoglobulin drugs, or blood products. The minimum number of subjects was estimated to be 110 individuals. This study was approved by the Ethics Committee of Faculty of Medicine, Universitas Indonesia and Cipto Mangunkusumo Hospital (KET-1160/UN2.F1/ETIK/PPM.00.02/2022). All parents of recruited study participants had provided their consents to participate.
2.2. Humoral Immune Response Assays
Upon blood collection from all participants, sera anti-diphtheria immunoglobulin G (IgG) and SARS-CoV-2 S-RBD antibodies were assessed. Titers of anti-diphtheria IgG were assessed using a commercial enzyme-linked immunosorbent assay (ELISA) of anti-diphtheria toxoid ELISA IgG (EUROIMMUN, Lübeck, Germany). This method utilized inactivated diphtheria toxin as a tracer antigen for quantitative assessment of anti-diphtheria toxoid IgG in serum [28]. The SARS-CoV-2 spike protein receptor-binding domain (S-RBD) total antibodies (including IgG) were assessed using the electrochemiluminescence assay of Roche Elecsys® anti-SARS-CoV-2 with the Cobas e 411 analyzer (Roche Diagnostic, Rotkreuz, Switzerland). According to the manufacture, the measuring range was between 0.4–250 U/mL in undiluted samples, with a value below 0.8 U/mL was considered as negative and a value equal to or above 0.8 U/mL was considered as positive [29]. If an undiluted sample exhibited a value above 250 U/mL, it would be diluted 1:10 until it reached 2,500 U/mL. If its value was still above 2,500 U/mL, a subsequent dilution was carried out at 1:100 (i.e., reaching 25,000 U/mL).
2.3. Cellular Immune Response Assay
The specific cellular immunity was assessed by measuring SARS-CoV-2-specific T cell-derived interferon gamma (IFN-g) using the EUROIMMUN SARS-CoV-2 Quan-T-Cell interferon gamma-release assay (IGRA) (EUROIMMUN, Lübeck, Germany). The assay was used to assess specific production of IFN-g by CD4+ and CD8+ T lymphocytes upon stimulation with SARS-CoV-2 spike protein-derived peptides [30]. Briefly, 500 μL of whole blood in a lithium heparin tube was transferred within an hour into three specific tubes: (i) a SARS-CoV-2 IGRA BLANK with no T-cell activating component and reflecting background T-cell activity, (ii) a SARS-CoV-2 IGRA TUBE coated with a pool of SARS-CoV-2 spike protein-derived peptides able to stimulate specific CD4+ and CD8+ T lymphocytes, and (iii) a SARS-CoV-2 IGRA STIM coated with a mitogen for non-specific T-cell stimulation and used as a control for the viability and stimulation ability of T cells. After six inversions, these tubes were incubated at 37 °C for 20–24 hours and subsequently centrifuged at room temperature for 10 minutes at 12,000 ×g. The supernatant was stored at -20°C until measurement. Pertaining to IFN-g measurement with ELISA (EUROIMMUN, Lübeck, Germany), the supernatant was diluted 1:5 with a dilution buffer. Six calibrators and two controls were used in each run. Specific IFN-g concentrations were obtained after subtracting the BLANK value from the TUBE/STIM value, in which the concentrations were expressed in mIU/mL. Results <100 and >200 mIU/mL were considered negative and positive, respectively. The upper limit of detection was 2,500 mIU/mL.
2.4. Statistical Analysis
The statistical analysis was performed using the IBM SPSS Statistics for Windows version 26.0 (IBM Corp., Armonk, USA). Descriptive data were presented as median with minimum and maximum values for continuous variables, while categorical variables were presented as frequency and percentage. Statistical difference of anti-diphtheria toxoid IgG or anti-SARS-CoV-2 S-RBD antibody titers between study participants who had received DT booster and the ones who had not received was analyzed using Mann-Whitney test. Statistical differences of anti-SARS-CoV-2 S-RBD titers or SARS-CoV-2-specific T cell-derived IFN-g concentrations among groups was assessed using Kruskal-Wallis test. If it was significant (p-value less than 0.05), Dunn’s multiple comparisons test was performed to analyze statistical differences between two groups. Data were visualized using the GraphPad Prism version 10.2.3 (GraphPad Software, Boston, USA).
3. Results
3.1. Characteristic of Study Participants
One hundred and fifty four children were recruited into this study (Table 1), with median age of 92 months and BMI of 14.8 kg/m2, which was considered as underweight [31]. The proportion of boys and girls were 39% and 61%, respectively. Most study participants came from lower-income families. Although a prior diagnosis of COVID-19 excluded eligible subjects from this study, most study participants reported a history of unknown acute respiratory infection in the past 6 months, of whom 31.2% suffered the respiratory infection more than 3 times. Many of them also had family members who had been diagnosed with COVID-19.
The study participants were subsequently classified based on their COVID-19 and DTP vaccination statuses. While study participants were categorized as “yes” for COVID-19 vaccination if they had received two doses of CoronaVac vaccine, the participants were categorized as “yes” for DTP vaccination if they had received three doses of DTP vaccine and one dose of DT booster vaccine at 5 years old, irrespective whether they had received additional dose of DTP vaccine before 2 years old. The study participants were subsequently stratified among four groups, i.e., 39 children in group A (“COVID-19 yes / DTP yes”), 38 children in group B (“COVID-19 yes / DTP no”), 38 children in group C (“COVID-19 no / DTP yes”) and 39 children in group D (“COVID-19 no / DTP no”). Frequency of acute respiratory infection as well as incidence of COVID-19 among other family members were evenly distributed among these 4 groups as well (Supplementary Table).

Table 1.
Characteristics of study participants (n=154).
| Variable | Value |
|---|---|
| Age, months [median (minimum–maximum)] | 92 (81–103) |
| BMI, kg/m2 [median (minimum–maximum)] | 14.8 (12.3–19.7) |
| Sex [n (%)] Male Female |
60 (39) 94 (61) |
| Parental occupation [n (%)] Working Not working |
71 (46.1) 83 (53.9) |
| Parental income [n (%)] Equal or above the minimum wage Below the minimum wage |
24 (15.6) 130 (84.4) |
| History of acute respiratory infection in last 6 months [n (%)] < 3 times ≥ 3 times |
106 (68.8) 48 (31.2) |
| History of COVID-19 disease in other family members [n (%)] No Yes |
73 (47.4) 81 (52.6) |
| Classification based on both vaccination statuses [n (%)] Group A (COVID-19 yes / DTP yes) Group B (COVID-19 yes / DTP no) Group C (COVID-19 no / DTP yes) Group D (COVID-19 no / DTP no) |
39 (25.3) 38 (24.7) 38 (24.7) 39 (25.3) |
The minimum wage in Jakarta for 2024 was IDR 5,067,381 (approximately US$ 325). COVID-19 vaccination status was deemed as yes if a subject received 2 doses of CoronaVac. DTP vaccination status was deemed as yes if a subject received 3 doses of DTP vaccine and a booster DT vaccine at five years old (with or without receiving an additional DTP vaccine before two years old). BMI, body mass index; COVID-19, coronavirus disease 2019; DTP, diphtheria-tetanus-pertussis; DT, diphtheria-tetanus.
3.2. Titers of Anti-Diphtheria Toxoid IgG
DTP primary vaccination (using whole-cell pertussis) along with DT booster vaccination are included in the mandatory national vaccination program in Indonesia. In order to assess whether DTP vaccination could enhance the impact of CoronaVac vaccination to generate COVID-19-specific immune responses, study participants were classified into two groups, i.e., “DTP yes” and “DTP no” (i.e., had received only 3 doses of DTP vaccine). There were 77 children in each group. As a surrogate marker of DTP vaccination-booster status, titer of anti-diphtheria toxoid IgG was measured in all study participants (Figure 1). As expected, study participants in the group “DTP yes” had a significantly higher titer (median=0.9349 IU/mL) as compared to the ones in the group “DTP no” (median=0.2113 IU/mL). Of note, a further sub-stratification of group “DTP yes” based on receiving an additional dose of DTP vaccine before 2 years old (“Full”; n=63; median=0.9349 IU/mL) or not receiving (“Partial”; n=14; median=0.6290 IU/mL) did not show any significant difference (p=0.2630; Supplementary Figure). The group “DTP yes” in subsequent analyses would therefore consist of all study participants receiving DT booster vaccine, irrespective whether they had received an additional dose of DTP vaccine before two years old.
3.3. Titers of Anti-SARS-CoV-2 S-RBD Antibodies
The study participants were subsequently assessed based on their COVID-19 vaccination status, in which they were classified into four groups based on their DTP and COVID-19 vaccination statuses (Table 2). Of note, the administration of CoronaVac was well tolerated by the study participants (data not shown). Overall finding suggested that there was no statistical difference in titers of anti-SARS-CoV-2 S-RBD antibodies among four groups (p=0.089). Nonetheless, as expected, group A (“COVID-19 yes / DTP yes”) had the highest median titer of anti-S-RBD antibodies (median=1,196 U/mL), while group D (“COVID-19 no / DTP no”) had the lowest median titer of anti-S-RBD antibodies (median=527.9 U/mL). Group C (“COVID-19 no / DTP yes”) surprisingly had the second highest median titer of anti-S-RBD antibodies (median=1,163 U/mL), despite the participants did not receive 2 doses of CoronaVac vaccine. Interestingly, group A had a higher median titer than group B (“COVID-19 yes / DTP no”; median=771.2 U/mL), suggesting an enhancing effect of DTP vaccination on CoronaVac vaccine to generate COVID-19-specific humoral immunity. A follow-up analysis was performed by stratifying titers of anti-SARS-CoV-2 S-RBD antibodies based on their DTP vaccination status. Table 3 indeed shows that study participants from the group of “DTP yes” had a higher median titer of anti-SARS-CoV-2 S-RBD antibodies than the ones from the group of “DTP no”.
3.4. Concentrations of SARS-CoV-2-Specific T Cell-Derived IFN-g
Upon the observation that the DTP vaccination could modulate the COVID-19-specific humoral immunity, it was of interest to assess the COVID-19-specific cellular immunity as well. The interferon gamma-release assay following ex vivo whole blood stimulation with a peptide pool of SARS-CoV-2 spike protein was deployed because this approach was relatively simple but reliable and it allowed an unbiased analysis on T-cell response towards SARS-CoV-2-specific antigen by using limited amounts of blood [30,32]. As shown in Figure 2, the stratification of study participants based on their COVID-19 and DTP vaccination statuses provided a clear assessment of their COVID-19-specific cellular immunity. The group A indeed had the highest median concentrations of T cell-derived IFN-g, followed by group B, C and D. Indeed, the IFN-g concentration in group A (median=560.9 mIU/mL) was significantly higher than the one in group C (median=230.8 mIU/mL; p=0.0003) or D (median=187.9 mIU/mL; p=0.0027), suggesting a substantial impact of synergistic DTP and CoronaVac vaccinations to generate COVID-19-specific T-cell immunity among these pediatric subjects. Despite no significant difference in the IFN-g concentration was observed between group A and B (median=318.0 mIU/mL; p=0.6634), it was interesting to note that there was an increasing trend of IFN-g production among the study participants who had received both COVID-19 and DTP vaccinations. Taken together, the complete DTP vaccination might be able to boost SARS-CoV-2-specific cellular immunity generated by CoronaVac vaccination.
4. Discussion
We hereby reported the impact of DTP vaccination-booster on SARS-CoV-2-specific adaptive immune responses among 154 healthy children aged 6-8 years old who had received CoronaVac vaccination in Indonesia. Our findings can be summarized in three points. First, despite DTP vaccination-booster has been included in the national pediatric immunization program in Indonesia since 2017 [33], we noticed that only 50% of the study participants had received 3 doses of DTP primary vaccine with 1 dose of DT booster vaccine at 5 years old, resulting in a substantial difference of anti-diphtheria toxoid IgG between children with complete DTP vaccination status and the ones with the incomplete status. Several factors contributed to the low coverage of DTP vaccination, including inadequate health services across Indonesia, vaccine hesitancy and COVID-19 pandemic [34,35]. This low coverage was alarming because Indonesia had recently encountered several outbreaks of diphtheria and pertussis [34,36,37]. Furthermore, it has been postulated that DTP vaccine can generate trained immunity in innate immune cells as well as activate heterologous adaptive immunity to unrelated pathogens [24]. It is unlikely that the heterologous immunity could be sufficiently created if the coverage of DTP vaccination-booster was below the required level of mass vaccination.
Second, we observed that the administration of 2 doses of CoronaVac vaccine was effective to generate B and T cell-mediated immune responses towards SARS-CoV-2 among the study participants. To date, two pediatric COVID-19 vaccines for 6-year-old children and above are approved in Indonesia, i.e., CoronaVac and BNT162b2 [38,39]. However, as the latter vaccine is not yet included in the national childhood vaccination program in Indonesia thus far, we focused our analysis on healthy children receiving the former vaccine. In Indonesia, the CoronaVac is administered twice (@3 mg per 0.5 mL) with an interval of 28 days for children aged 6-17 years old without any booster. We indeed observed that CoronaVac-vaccinated study participants had noticeable higher levels of anti-SARS-CoV-2 S-RBD antibodies and T cell-derived IFN-g as compared to the ones who did not receive COVID-19 vaccination. These findings were in accordance with previous studies among healthy children and adults, showing that CoronaVac, an inactivated SARS-CoV-2 vaccine, could generate both humoral and cellular immune responses [29,38,40,41,42]. While the induced titers of anti-SARS-CoV-2 S-RBD antibodies were relatively low (presumably due to a long duration between the second dose of CoronaVac administration and the blood testing, because substantial waning of the specific antibodies commonly occurred after 3 months post-vaccination), the T cell-specific response upon stimulation with peptides of SARS-CoV-2 spike protein was well maintained in this study, supporting a notion that inactivated SARS-CoV-2 vaccine was better in generating specific cellular immunity that would help to protect against development of severe COVID-19, rather than inducing specific humoral immunity that would help to prevent a symptomatic SARS-CoV-2 infection [41,43]. Of note, it was recently published that upon administration of 2 doses of BNT162b2, healthy children aged 5-12 years old mounted antibody, B cell- and T cell-responses, in which they had higher antibody and T cell responses than adults 6 months after vaccination. Importantly, that study also suggested that T-cell response was the most important predictor of protection against COVID-19 in children [44].
Third, we suggested that the complete DTP vaccination-booster (i.e., had received 3 doses of DTP primary vaccine and 1 dose of DT booster vaccine at 5 years old, irrespective whether had received an additional dose of DTP vaccine before 2 years old) could enhance SARS-CoV-2-specific humoral and cellular immune responses. Despite the low coverage of DTP vaccination among the study cohort, we observed that children with complete DTP vaccination-booster exhibited significantly higher titers of anti-SARS-CoV-2 S-RBD antibodies than the ones with an incomplete DTP vaccination-booster (p=0.026). Although the subsequent stratification based on the COVID-19 and DTP vaccination statuses did not demonstrate any significant difference of anti-SARS-CoV-2 S-RBD antibody titers among the four groups, we speculate that the insignificant difference was presumably due to the relatively low ability of CoronaVac vaccination to generate higher titers of anti-SARS-CoV-2 S-RBD antibodies as well as the substantial waning of specific antibodies several months post-vaccination, as had been reported by our group and others in healthy children and adults [29,38,45,46,47,48,49]. In contrast, we observed a significant difference of T cell-specific IFN-g concentrations among the four groups, suggesting that a combination of COVID-19 and DTP vaccinations (i.e., group A) could enhance T-cell immunity towards SARS-CoV-2. The potential role of heterologous immunity was supported by our observation that the IFN-g concentrations among group B (“COVID-19 yes / DTP no”), C (“COVID-19 no / DTP yes”) and D (“COVID-19 no / DTP no”) were not significantly different. This idea was aligned with published studies, reporting an association between a recent history of DTP vaccination and protection against COVID-19 [26,50]. However, as the IFN-g concentrations between group A and B in this study did not differ significantly, this also suggests that either the heterologous immune response due to DTP and CoronaVac vaccinations might not be that strong to substantially enhance T cell-immunity against SARS-CoV-2 [24,51,52], the choice of DT vaccine (instead of DTP) as a booster at 5 years old might not be the most optimum choice to generate heterologous immune response towards SARS-CoV-2 [25,50], or the longer and varied duration between DT booster vaccination, CoronaVac vaccination and the immunological assays might decrease the impact of heterologous immunity in this study [50]. Nonetheless, it was obvious that the group A had the highest median of IFN-g concentration among all in this study, suggesting a usefulness of childhood routine vaccination program (i.e., DTP) to induce heterologous immunity against SARS-CoV-2 antigens, which subsequently could enhance the generated adaptive immune responses upon vaccination with an inactivated SARS-CoV-2 vaccine in healthy children (Figure 3).
Heterologous immunity is a memory T cell-response induced by a particular pathogen, which could be directed against another pathogen as well. This could result in an enhanced immunity against the novel pathogen [53,54]. This hypothesis becomes an attractive concept when there is an outbreak of a novel pathogen and there is a lack of effective vaccine to control the outbreak, probably due to ongoing research and development or due to distribution inequality of vaccines globally [3,4]. As routine vaccination (particularly the childhood vaccination) has become an integral part of the national public health program across the globe, including Indonesia, capabilities of routine vaccines (e.g., DTP, BCG or MMR) to generate heterologous immunity against COVID-19 had been investigated. Several in silico findings had reported that certain pathogenic bacteria, including DTP, could generate cross-reactive immunity to SARS-CoV-2, because DTP and SARS-CoV-2 shared multiple epitopes, including numerous CD8+ and CD4+ T-cell epitopes with broad protection coverage [25,55]. This finding was substantiated by other studies, reporting that elderlies with registered diphtheria or tetanus vaccinations were less likely to develop severe COVID-19 in the UK and USA [26,50]. The in vitro data also had suggested a high correlation between T-cell response towards Tdap and SARS-CoV-2 antigens [50]. However, another study had reported a conflicting result, showing that DTP vaccination was not associated with a lower incidence of SARS-CoV-2 infection across various age groups in the USA [51]. This difference could be attributed to (i) T-cell immunity, as the primary result of heterologous immune response, is more involved in the protection against development of severe COVID-19, instead of the prevention of SARS-CoV-2 infection [41,43], (ii) the Tdap vaccine is commonly used as a booster every 10 years in developed countries, despite the recent finding that acellular pertussis antigen (in contrast to whole-cell pertussis antigen) only shared a few epitopes with SARS-CoV-2 antigen [25], (iii) the time interval between routine vaccination, COVID-19 vaccination (or even SARS-CoV-2 infection) and immunological testing, as well as (iv) the selected age groups (e.g., older individuals might have lower immunity than younger individuals).
Our study therefore had an advantage by focusing on DTP and CoronaVac vaccinations among healthy children aged 6-8 years old. As DTP vaccination and DT booster at 5 years old are parts of the national childhood vaccination program, we were able to narrow the time interval between DT booster and CoronaVac vaccination (administered as early as 6 years old) to increase the probability of inducing heterologous immunity. Our study demonstrated additional benefits of the routine childhood vaccination program, i.e., vaccination against certain bacteria might be able to boost immune responses against a novel virus. This emphasizes the rationale as well as the importance to continue vaccinating populations against common pathogens, despite the disruption of routine childhood vaccination program due to the pandemic [56]. Furthermore, our study compared the production of specific antibodies and IFN-g against SARS-CoV-2 to discern any enhancing impact of DTP vaccination on CoronaVac vaccination to generate SARS-CoV-2-specific adaptive immunity. This is important because many published studies had only measured humoral immunity (e.g., titers of specific antibodies) against COVID-19, resulting in an incomplete analysis and understanding of adaptive immune responses towards SARS-CoV-2.
Our study had several limitations. The nature of cross-sectional study did not allow us to determine any causality between DTP and CoronaVac vaccinations. We also did not measure clinical outcome of the study participants pertaining to SARS-CoV-2 infection and/or COVID-19 severity. Nevertheless, two published studies in Chile had reported the effectiveness of CoronaVac vaccination among children aged 3-16 years old to protect against severe COVID-19, either due to Delta or Omicron variant [57,58], suggesting that CoronaVac vaccination among healthy children in Indonesia might result in a similar efficacy. In addition, the relatively long duration between the second dose of COVID-19 vaccination and the immunological testing in this study might obscure the distinction of COVID-19 vaccine-related effect among tested groups, particularly on titers of anti-SARS-CoV-2 S-RBD antibodies. Although we only recruited healthy children aged 6-8 years old without any confirmed history of COVID-19, we could not exclude a possibility that the study participants might already contract COVID-19 without exhibiting any serious symptoms or without being diagnosed. This was possible because SARS-CoV-2 infection among pediatric patients were usually milder than the ones among adults [5,59]. Next, CoronaVac as an inactivated SARS-CoV-2 vaccine would generate spike-, membrane-, as well as nucleoprotein-specific T cells (particularly CD4+ T cells) [41,42]. Thus, by only using peptide pools from the spike protein, we would not be able to assess the overall T-cell immune response against all SARS-CoV-2 antigens. Lastly, our study only measured secreted IFN-g as a marker of T-cell response among vaccinated children. This mono-functionality would overlook the complete capability of virus-specific T cells because SARS-CoV-2-specific T cells appeared to be poly-functional by able in secreting various cytokines (e.g., IL-2 or TNF-a in addition to IFN-g) [32,60]. The limited blood volume from the children also did not allow us to investigate antigen-specific T cells by using flow cytometry. Nonetheless, a recent study on healthy children aged 5-12 years old vaccinated with BNT162b2 vaccine had reported that T-cell response as indicated by IFN-g release was the most important predictor for protection against symptomatic SARS-CoV-2 infection [44].
5. Conclusions
In this study, we analyzed the potential of complete DTP vaccination-booster to generate heterologous immunity that could enhance SARS-CoV-2-specific adaptive immunity induced by an administration of inactivated SARS-CoV-2 vaccine among healthy children aged 6-8 years old. Upon stimulation with peptides of SARS-CoV-2 spike protein, we particularly observed higher concentrations of IFN-g secreted by T cells in study participants with complete DTP and CoronaVac vaccination statuses, which could provide protection against COVID-19 in pediatric population. Our findings support the usefulness of pediatric COVID-19 vaccination as it generated SARS-CoV-2-specific adaptive immune responses, hence could serve as an impetus for policymakers in Indonesia to continuously provide COVID-19 vaccines for children. We also suggest an additional benefit of routine childhood vaccination, e.g., DTP, to generate protection against novel pathogens, further emphasizing the importance for policymakers to maintain routine childhood immunization despite of new epidemic. Further studies will be required to elucidate exact mechanism of heterologous adaptive immunity and to enhance it as a priming to protect against novel pathogens.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Supplementary Table S1. Characteristics of study participants as stratified in four groups; Supplementary Figure S1. Titers of anti-diphtheria toxoid immunoglobulin G of study participants. Study participants who had received 3 doses of DTP vaccine and 1 dose of DT booster (n=77) were classified into two groups, i.e., Full (had also received 1 additional dose of DTP vaccine before 2 years old; n=63) and Partial (had not received 1 additional dose of DTP vaccine before 2 years old; n=14). Solid horizontal line within each box-and-whiskers refers to the median value. The whiskers refer to respective minimum and maximum values. Mann-Whitney test was performed with p<0.05 was considered as statistically significant.
Author Contributions
Conceptualization, T.S., J.J., Z.M., Y.V. and B.H.; methodology, T.S., J.J., A.R.H., R.A.W., Z.M. and B.H.; software, T.S and J.J..; validation, T.S., J.J., A.R.H. and B.H.; formal analysis, T.S., J.J., A.R.H., R.A.W., S.R.S.H., A.P., Z.M. and B.H.; investigation, T.S., J.J., A.R.H., Z.M. and B.H.; resources, T.S., A.R.H. and B.H.; data curation, T.S., J.J., A.R.H., Z.M. and B.H.; writing—original draft preparation, T.S. and J.J.; writing—review and editing, T.S., J.J., A.R.H., R.A.W., S.R.S.H., I.N.S, A.P., Z.M., Y.V. and B.H.; visualization, T.S., J.J. and A.R.H.; supervision, A.P., Z.M. and B.H.; project administration, T.S. and B.H.; funding acquisition, T.S. and B.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Directorate of Research and Development, Universitas Indonesia under Hibah PUTI 2023 (Grant No. NKB-361/UN2.RST/HKP.05.00/2023).
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Ethics Committee of Universitas Indonesia, No. KET-1160/UN2.F1/ETIK/PPM.00.02/2022.
Informed Consent Statement
Informed consent was obtained from all parents of the study participants involved in the study. Written informed consent has been obtained from parents of the study participants to publish this paper.
Data Availability Statement
Data are available upon request to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- World Health Organization WHO COVID-19 Dashboard Available online: https://data.who.int/dashboards/covid19/cases (accessed on 22 June 2024).
- Barouch, D.H. Covid-19 Vaccines — Immunity, Variants, Boosters. N. Engl. J. Med. 2022, 387, 1011–1020. [CrossRef]
- Tatar, M.; Shoorekchali, J.M.; Faraji, M.R.; Seyyedkolaee, M.A.; Pagán, J.A.; Wilson, F.A. COVID-19 Vaccine Inequality: A Global Perspective. J. Glob. Health 2022, 12, 10–13. [CrossRef]
- Gozzi, N.; Chinazzi, M.; Dean, N.E.; Longini, I.M.; Halloran, M.E.; Perra, N.; Vespignani, A. Estimating the Impact of COVID-19 Vaccine Inequities: A Modeling Study. Nat. Commun. 2023, 14, 1–10. [CrossRef]
- Steinman, J.B.; Lum, F.M.; Ho, P.P.K.; Kaminski, N.; Steinman, L. Reduced Development of COVID-19 in Children Reveals Molecular Checkpoints Gating Pathogenesis Illuminating Potential Therapeutics. Proc. Natl. Acad. Sci. U. S. A. 2020, 117, 24620–24626. [CrossRef]
- Zimmermann, P.; Curtis, N. Why Is COVID-19 Less Severe in Children? A Review of the Proposed Mechanisms Underlying the Age-Related Difference in Severity of SARS-CoV-2 Infections. Arch. Dis. Child. 2021, 106, 429–439. [CrossRef]
- Axfors, C.; Pezzullo, A.M.; Contopoulos-Ioannidis, D.G.; Apostolatos, A.; Ioannidis, J.P.A. Differential COVID-19 Infection Rates in Children, Adults, and Elderly: Systematic Review and Meta-Analysis of 38 Pre-Vaccination National Seroprevalence Studies. J. Glob. Health 2023, 13. [CrossRef]
- Powell, A.A.; Dowell, A.C.; Moss, P.; Ladhani, S.N. Current State of COVID-19 in Children: 4 Years On. J. Infect. 2024, 88. [CrossRef]
- Munro, A.P.S.; Jones, C.E.; Faust, S.N. Vaccination against COVID-19 — Risks and Benefits in Children. Eur. J. Pediatr. 2024, 183, 1107–1112. [CrossRef]
- Castagnoli, R.; Votto, M.; Licari, A.; Brambilla, I.; Bruno, R.; Perlini, S.; Rovida, F.; Baldanti, F.; Marseglia, G.L. Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Infection in Children and Adolescents: A Systematic Review. JAMA Pediatr. 2020, 174, 882–889. [CrossRef]
- Feldstein, L.R.; Rose, E.B.; Horwitz, S.M.; Collins, J.P.; Newhams, M.M.; Son, M.B.F.; Newburger, J.W.; Kleinman, L.C.; Heidemann, S.M.; Martin, A.A.; et al. Multisystem Inflammatory Syndrome in U.S. Children and Adolescents. N. Engl. J. Med. 2020, 383, 334–346. [CrossRef]
- Hoste, L.; Paemel, R. Van; Haerynck, F. Multisystem Inflammatory Syndrome in Children Related to COVID-19: A Systematic Review.Eur J Pediatr.2021, Jul;180(7):2019-2034. [CrossRef]
- Pudjiadi, A.H.; Putri, N.D.; Sjakti, H.A.; Yanuarso, P.B.; Gunardi, H.; Roeslani, R.D.; Pasaribu, A.D.; Nurmalia, L.D.; Sambo, C.M.; Ugrasena, I.D.G.; et al. Pediatric COVID-19: Report From Indonesian Pediatric Society Data Registry. Front. Pediatr. 2021, 9, 1–7. [CrossRef]
- Nachega, J.B.; Sam-Agudu, N.A.; MacHekano, R.N.; Rabie, H.; Van Der Zalm, M.M.; Redfern, A.; Dramowski, A.; O’Connell, N.; Pipo, M.T.; Tshilanda, M.B.; et al. Assessment of Clinical Outcomes among Children and Adolescents Hospitalized with COVID-19 in 6 Sub-Saharan African Countries. JAMA Pediatr. 2022, 176, E216436. [CrossRef]
- Ortiz-Prado, E.; Izquierdo-Condoy, J.S.; Fernandez-Naranjo, R.; Vasconez, J.; Dávila Rosero, M.G.; Revelo-Bastidas, D.; Herrería-Quiñonez, D.; Rubio-Neira, M. The Deadly Impact of COVID-19 among Children from Latin America: The Case of Ecuador. Front. Pediatr. 2023, 11, 1–12. [CrossRef]
- Martínez-Valdez, L.; Richardson, V.L.; Bautista-Márquez, A.; Camacho Franco, M.A.; Cruz Cruz, V.; Hernández Ávila, M. Three Years of COVID-19 in Children That Attend the Mexican Social Security Institute’s 1,350 Child Day-Care Centers, 2020–2023. Front. Pediatr. 2023, 11, 1–12. [CrossRef]
- Kulkarni, D.; Ismail, N.F.; Zhu, F.; Wang, X.; del Carmen Morales, G.; Srivastava, A.; Allen, K.E.; Spinardi, J.; Rahman, A.E.; Kyaw, M.H.; et al. Epidemiology and Clinical Features of SARS-CoV-2 Infection in Children and Adolescents in the Pre-Omicron Era: A Global Systematic Review and Meta-Analysis. J. Glob. Health 2024, 14, 1–23. [CrossRef]
- Yonker, L.M.; Boucau, J.; Regan, J.; Choudhary, M.C.; Burns, M.D.; Young, N.; Farkas, E.J.; Davis, J.P.; Moschovis, P.P.; Bernard Kinane, T.; et al. Virologic Features of Severe Acute Respiratory Syndrome Coronavirus 2 Infection in Children. J. Infect. Dis. 2021, 224, 1821–1829. [CrossRef]
- Hartantri, Y.; Debora, J.; Widyatmoko, L.; Giwangkancana, G.; Suryadinata, H.; Susandi, E.; Hutajulu, E.; Hakiman, A.P.A.; Pusparini, Y.; Alisjahbana, B. Clinical and Treatment Factors Associated with the Mortality of COVID-19 Patients Admitted to a Referral Hospital in Indonesia. Lancet Reg. Heal. - Southeast Asia 2023, 11, 100167. [CrossRef]
- Santi, T.; Hegar, B.; Munasir, Z.; Prayitno, A.; Werdhani, R.A.; Bandar, I.N.S.; Jo, J.; Uswa, R.; Widia, R.; Vandenplas, Y. Factors Associated with Parental Intention to Vaccinate Their Preschool Children against COVID-19: A Cross-Sectional Survey in Urban Area of Jakarta, Indonesia. Clin. Exp. Vaccine Res. 2023, 12, 240–248. [CrossRef]
- Ministry of Health Republic of Indonesia Vaksinasi COVID-19 Nasional.
- Widianto, S. Indonesia to Start Vaccinating Children Aged 6-11 against COVID-19 Available online: https://www.reuters.com/business/healthcare-pharmaceuticals/indonesia-start-vaccinating-children-aged-6-11-against-covid-19-2021-12-13/ (accessed on 22 June 2024).
- Suhenda, D. COVID-19 Vaccine Shortages Reported across Country Available online: https://www.thejakartapost.com/indonesia/2022/10/21/covid-19-vaccine-shortages-reported-across-country.html (accessed on 22 June 2024).
- Messina, N.L.; Zimmermann, P.; Curtis, N. The Impact of Vaccines on Heterologous Adaptive Immunity. Clin. Microbiol. Infect. 2019, 25, 1484–1493. [CrossRef]
- Reche, P.A. Potential Cross-Reactive Immunity to SARS-CoV-2 From Common Human Pathogens and Vaccines. Front. Immunol. 2020, 11, 1–13. [CrossRef]
- Monereo-Sánchez, J.; Luykx, J.J.; Pinzón-Espinosa, J.; Richard, G.; Motazedi, E.; Westlye, L.T.; Andreassen, O.A.; van der Meer, D. Diphtheria And Tetanus Vaccination History Is Associated With Lower Odds of COVID-19 Hospitalization. Front. Immunol. 2021, 12, 1–7. [CrossRef]
- Nicholson, L.; Adkins, E.; Karyanti, M.R.; Ong-Lim, A.; Shenoy, B.; Huoi, C.; Vargas-Zambrano, J.C. What Is the True Burden of Diphtheria, Tetanus, Pertussis and Poliovirus in Children Aged 3–18 Years in Asia? A Systematic Literature Review. Int. J. Infect. Dis. 2022, 117, 116–129. [CrossRef]
- Zasada, A.A.; Rastawicki, W.; Śmietańska, K.; Rokosz, N.; Jagielski, M. Comparison of Seven Commercial Enzyme-Linked Immunosorbent Assays for the Detection of Anti-Diphtheria Toxin Antibodies. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 891–897. [CrossRef]
- Jo, J.; Sanjaya, A.; Pinontoan, R.; Aruan, M.; Wahyuni, R.M.; Viktaria, V. Assessment on Anti-SARS-CoV-2 Receptor-Binding Domain Antibodies among CoronaVac-Vaccinated Indonesian Adults. Clin. Exp. Vaccine Res. 2022, 11, 116. [CrossRef]
- Saad Albichr, I.; Mzougui, S.; Devresse, A.; Georgery, H.; Goffin, E.; Kanaan, N.; Yombi, J.C.; Belkhir, L.; De Greef, J.; Scohy, A.; et al. Evaluation of a Commercial Interferon-γ Release Assay for the Detection of SARS-CoV-2 T-Cell Response after Vaccination. Heliyon 2023, 9. [CrossRef]
- Rappaport, E.B. Identifying and Evaluating the Metabolic Syndrome in Children and Adolescents. Ethn. Dis. 2007, 17.
- Tan, A.T.; Lim, J.M.E.; Le Bert, N.; Kunasegaran, K.; Chia, A.; Qui, M.D.C.; Tan, N.; Chia, W.N.; de Alwis, R.; Ying, D.; et al. Rapid Measurement of SARS-CoV-2 Spike T Cells in Whole Blood from Vaccinated and Naturally Infected Individuals. J. Clin. Invest. 2021, 131. [CrossRef]
- Indonesian Pediatric Society Proceeding Book Childhood Immunization Update 2023; 2023;
- Harapan, H.; Anwar, S.; Dimiati, H.; Hayati, Z.; Mudatsir, M. Diphtheria Outbreak in Indonesia, 2017: An Outbreak of an Ancient and Vaccine-Preventable Disease in the Third Millennium. Clin. Epidemiol. Glob. Heal. 2019, 7, 261–262. [CrossRef]
- Itiakorit, H.; Sathyamoorthi, A.; O’Brien, B.E.; Nguyen, D. COVID-19 Impact on Disparity in Childhood Immunization in Low- and Middle-Income Countries Through the Lens of Historical Pandemics. Curr. Trop. Med. Reports 2022, 9, 225–233. [CrossRef]
- Arguni, E.; Karyanti, M.R.; Satari, H.I.; Hadinegoro, S.R. Diphtheria Outbreak in Jakarta and Tangerang, Indonesia: Epidemiological and Clinical Predictor Factors for Death. PLoS One 2021, 16, 1–11. [CrossRef]
- Sunarno, S.; Sofiah, S.N.; Amalia, N.; Hartoyo, Y.; Rizki, A.; Puspandari, N.; Saraswati, R.D.; Febriyana, D.; Febrianti, T.; Susanti, I.; et al. Laboratory and Epidemiology Data of Pertussis Cases and Close Contacts: A 5-Year Case-Based Surveillance of Pertussis in Indonesia, 2016–2020. PLoS One 2022, 17, 1–9. [CrossRef]
- Han, B.; Song, Y.; Li, C.; Yang, W.; Ma, Q.; Jiang, Z.; Li, M.; Lian, X.; Jiao, W.; Wang, L.; et al. Safety, Tolerability, and Immunogenicity of an Inactivated SARS-CoV-2 Vaccine (CoronaVac) in Healthy Children and Adolescents: A Double-Blind, Randomised, Controlled, Phase 1/2 Clinical Trial. Lancet Infect. Dis. 2021, 21, 1645–1653. [CrossRef]
- Walter, E.B.; Talaat, K.R.; Sabharwal, C.; Gurtman, A.; Lockhart, S.; Paulsen, G.C.; Barnett, E.D.; Muñoz, F.M.; Maldonado, Y.; Pahud, B.A.; et al. Evaluation of the BNT162b2 Covid-19 Vaccine in Children 5 to 11 Years of Age. N. Engl. J. Med. 2022, 386, 35–46. [CrossRef]
- Zeng, G.; Wu, Q.; Pan, H.; Li, M.; Yang, J.; Wang, L.; Wu, Z.; Jiang, D.; Deng, X.; Chu, K.; et al. Immunogenicity and Safety of a Third Dose of CoronaVac, and Immune Persistence of a Two-Dose Schedule, in Healthy Adults: Interim Results from Two Single-Centre, Double-Blind, Randomised, Placebo-Controlled Phase 2 Clinical Trials. Lancet Infect. Dis. 2022, 22, 483–495. [CrossRef]
- Lim, J.M.E.; Hang, S.K.; Hariharaputran, S.; Chia, A.; Tan, N.; Lee, E.S.; Chng, E.; Lim, P.L.; Young, B.E.; Lye, D.C.; et al. A Comparative Characterization of SARS-CoV-2-Specific T Cells Induced by MRNA or Inactive Virus COVID-19 Vaccines. Cell Reports Med. 2022, 3, 100793. [CrossRef]
- Huang, R.; Ying, L.; Wang, J.; Xia, J.; Zhang, Y.; Mao, H.; Zhang, R.; Zang, R.; Le, Z.; Shu, Q.; et al. Non-Spike and Spike-Specific Memory T Cell Responses after the Third Dose of Inactivated COVID-19 Vaccine. Front. Immunol. 2023, 14, 1–10. [CrossRef]
- Bertoletti, A.; Le Bert, N.; Tan, A.T. SARS-CoV-2-Specific T Cells in the Changing Landscape of the COVID-19 Pandemic. Immunity 2022, 55, 1764–1778. [CrossRef]
- Zhong, Y.; Kang, A.Y.H.; Tay, C.J.X.; Li, H.E.; Elyana, N.; Tan, C.W.; Yap, W.C.; Lim, J.M.E.; Le Bert, N.; Chan, K.R.; et al. Correlates of Protection against Symptomatic SARS-CoV-2 in Vaccinated Children. Nat. Med. 2024, 30. [CrossRef]
- Santi, T.; Sungono, V.; Kamarga, L.; Samakto, B. De; Hidayat, F.; Hidayat, F.K.; Satolom, M.; Permana, A.; Yusuf, I.; Suriapranata, I.M.; et al. Heterologous Prime-Boost with the MRNA-1273 Vaccine among CoronaVac-Vaccinated Healthcare Workers in Indonesia. Clin. Exp. Vaccine Res. 2022, 11, 209–216.
- Barin, B.; Kasap, U.; Selçuk, F.; Volkan, E.; Uluçkan, Ö. Comparison of SARS-CoV-2 Anti-Spike Receptor Binding Domain IgG Antibody Responses after CoronaVac, BNT162b2, ChAdOx1 COVID-19 Vaccines, and a Single Booster Dose: A Prospective, Longitudinal Population-Based Study. The Lancet Microbe 2022, 3, e274–e283. [CrossRef]
- Clemens, S.A.C.; Weckx, L.; Clemens, R.; Mendes, A.V.A.; Souza, A.R.; Silveira, M.B. V.; da Guarda, S.N.F.; de Nobrega, M.M.; Pinto, M.I. de M.; Gonzalez, I.G.S.; et al. Heterologous versus Homologous COVID-19 Booster Vaccination in Previous Recipients of Two Doses of CoronaVac COVID-19 Vaccine in Brazil (RHH-001): A Phase 4, Non-Inferiority, Single Blind, Randomised Study. Lancet 2022, 399, 521–529. [CrossRef]
- Fadlyana, E.; Setiabudi, D.; Kartasasmita, C.B.; Putri, N.D.; Rezeki Hadinegoro, S.; Mulholland, K.; Sofiatin, Y.; Suryadinata, H.; Hartantri, Y.; Sukandar, H.; et al. Immunogenicity and Safety in Healthy Adults of Full Dose versus Half Doses of COVID-19 Vaccine (ChAdOx1-S or BNT162b2) or Full-Dose CoronaVac Administered as a Booster Dose after Priming with CoronaVac: A Randomised, Observer-Masked, Controlled Trial in I. Lancet Infect. Dis. 2023, 23, 545–555. [CrossRef]
- Sauré, D.; O’Ryan, M.; Torres, J.P.; Zuñiga, M.; Soto-Rifo, R.; Valiente-Echeverría, F.; Gaete-Argel, A.; Neira, I.; Saavedra, V.; Acevedo, M.L.; et al. COVID-19 Lateral Flow IgG Seropositivity and Serum Neutralising Antibody Responses after Primary and Booster Vaccinations in Chile: A Cross-Sectional Study. The Lancet Microbe 2023, 4, e149–e158. [CrossRef]
- Mysore, V.; Cullere, X.; Settles, M.L.; Ji, X.; Kattan, M.W.; Desjardins, M.; Durbin-Johnson, B.; Gilboa, T.; Baden, L.R.; Walt, D.R.; et al. Protective Heterologous T Cell Immunity in COVID-19 Induced by the Trivalent MMR and Tdap Vaccine Antigens. Med 2021, 2, 1050-1071.e7. [CrossRef]
- Pawlowski, C.; Puranik, A.; Bandi, H.; Venkatakrishnan, A.J.; Agarwal, V.; Kennedy, R.; O’Horo, J.C.; Gores, G.J.; Williams, A.W.; Halamka, J.; et al. Exploratory Analysis of Immunization Records Highlights Decreased SARS-CoV-2 Rates in Individuals with Recent Non-COVID-19 Vaccinations. Sci. Rep. 2021, 11, 1–20. [CrossRef]
- Haddad-Boubaker, S.; Othman, H.; Touati, R.; Ayouni, K.; Lakhal, M.; Ben Mustapha, I.; Ghedira, K.; Kharrat, M.; Triki, H. In Silico Comparative Study of SARS-CoV-2 Proteins and Antigenic Proteins in BCG, OPV, MMR and Other Vaccines: Evidence of a Possible Putative Protective Effect. BMC Bioinformatics 2021, 22, 1–14. [CrossRef]
- Balz, K.; Trassl, L.; Härtel, V.; Nelson, P.P.; Skevaki, C. Virus-Induced T Cell-Mediated Heterologous Immunity and Vaccine Development. Front. Immunol. 2020, 11, 1–8. [CrossRef]
- Gil, A.; Kenney, L.L.; Mishra, R.; Watkin, L.B.; Aslan, N.; Selin, L.K. Vaccination and Heterologous Immunity: Educating the Immune System. Trans. R. Soc. Trop. Med. Hyg. 2014, 109, 62–69. [CrossRef]
- Eggenhuizen, P.J.; Ng, B.H.; Chang, J.; Cheong, R.M.Y.; Yellapragada, A.; Wong, W.Y.; Ting, Y.T.; Monk, J.A.; Gan, P.Y.; Holdsworth, S.R.; et al. Heterologous Immunity Between SARS-CoV-2 and Pathogenic Bacteria. Front. Immunol. 2022, 13, 1–18. [CrossRef]
- Basu, S.; Ashok, G.; Debroy, R.; Ramaiah, S.; Livingstone, P.; Anbarasu, A. Impact of the COVID-19 Pandemic on Routine Vaccine Landscape: A Global Perspective. Hum. Vaccines Immunother. 2023, 19, 1–8. [CrossRef]
- Jara, A.; Undurraga, E.A.; Zubizarreta, J.R.; González, C.; Acevedo, J.; Pizarro, A.; Vergara, V.; Soto-Marchant, M.; Gilabert, R.; Flores, J.C.; et al. Effectiveness of CoronaVac in Children 3–5 Years of Age during the SARS-CoV-2 Omicron Outbreak in Chile. Nat. Med. 2022, 1–4. [CrossRef]
- Jara, A.; Undurraga, E.A.; Flores, J.C.; Zubizarreta, J.R.; González, C.; Pizarro, A.; Ortuño-Borroto, D.; Acevedo, J.; Leo, K.; Paredes, F.; et al. Effectiveness of an Inactivated SARS-CoV-2 Vaccine in Children and Adolescents: A Large-Scale Observational Study. Lancet Reg. Heal. - Am. 2023, 21, 100487. [CrossRef]
- Ludvigsson, J.F. Systematic Review of COVID-19 in Children Shows Milder Cases and a Better Prognosis than Adults. Acta Paediatr. Int. J. Paediatr. 2020, 109, 1088–1095. [CrossRef]
- Le Bert, N.; Clapham, H.E.; Tan, A.T.; Chia, W.N.; Tham, C.Y.L.; Lim, J.M.; Kunasegaran, K.; Tan, L.W.L.; Dutertre, C.A.; Shankar, N.; et al. Highly Functional Virus-Specific Cellular Immune Response in Asymptomatic SARS-CoV-2 Infection. J. Exp. Med. 2021, 218. [CrossRef]
Figure 1.
Titers of anti-diphtheria toxoid immunoglobulin G of study participants. Study participants were classified into two groups based on their DTP vaccination status, i.e., Yes (had received 3 doses of DTP primary vaccine with 1 dose of DT booster vaccine at 5 years old, irrespective whether receiving an additional dose of DTP vaccine before 2 years old) and No (had only received 3 doses of DTP primary vaccine). Solid horizontal line within each violin plot refers to the median value. Mann-Whitney test was performed to determine any statistical difference between two groups, in which **** marked p<0.0001. .
Figure 1.
Titers of anti-diphtheria toxoid immunoglobulin G of study participants. Study participants were classified into two groups based on their DTP vaccination status, i.e., Yes (had received 3 doses of DTP primary vaccine with 1 dose of DT booster vaccine at 5 years old, irrespective whether receiving an additional dose of DTP vaccine before 2 years old) and No (had only received 3 doses of DTP primary vaccine). Solid horizontal line within each violin plot refers to the median value. Mann-Whitney test was performed to determine any statistical difference between two groups, in which **** marked p<0.0001. .

Figure 2.
Concentration of SARS-CoV-2-specific T cell-derived interferon gamma of study participants. Study participants were classified into four groups based on their COVID-19 and DTP vaccination statuses, i.e., A (“COVID-19 yes / DTP yes”), B (“COVID-19 yes / DTP no”), C (“COVID-19 no / DTP yes”) and D (“COVID-19 no / DTP no”). Solid horizontal line within each box-and-whiskers refers to the median value. The whiskers refer to respective the 10 and 90 percentile values. Kruskal-Wallis test was performed to determine any statistical difference among four groups. If it was significant (p<0.05), Dunn’s multiple comparisons test was subsequently performed. The *** and ** marked p<0.001 and p<0.01, respectively. .
Figure 2.
Concentration of SARS-CoV-2-specific T cell-derived interferon gamma of study participants. Study participants were classified into four groups based on their COVID-19 and DTP vaccination statuses, i.e., A (“COVID-19 yes / DTP yes”), B (“COVID-19 yes / DTP no”), C (“COVID-19 no / DTP yes”) and D (“COVID-19 no / DTP no”). Solid horizontal line within each box-and-whiskers refers to the median value. The whiskers refer to respective the 10 and 90 percentile values. Kruskal-Wallis test was performed to determine any statistical difference among four groups. If it was significant (p<0.05), Dunn’s multiple comparisons test was subsequently performed. The *** and ** marked p<0.001 and p<0.01, respectively. .
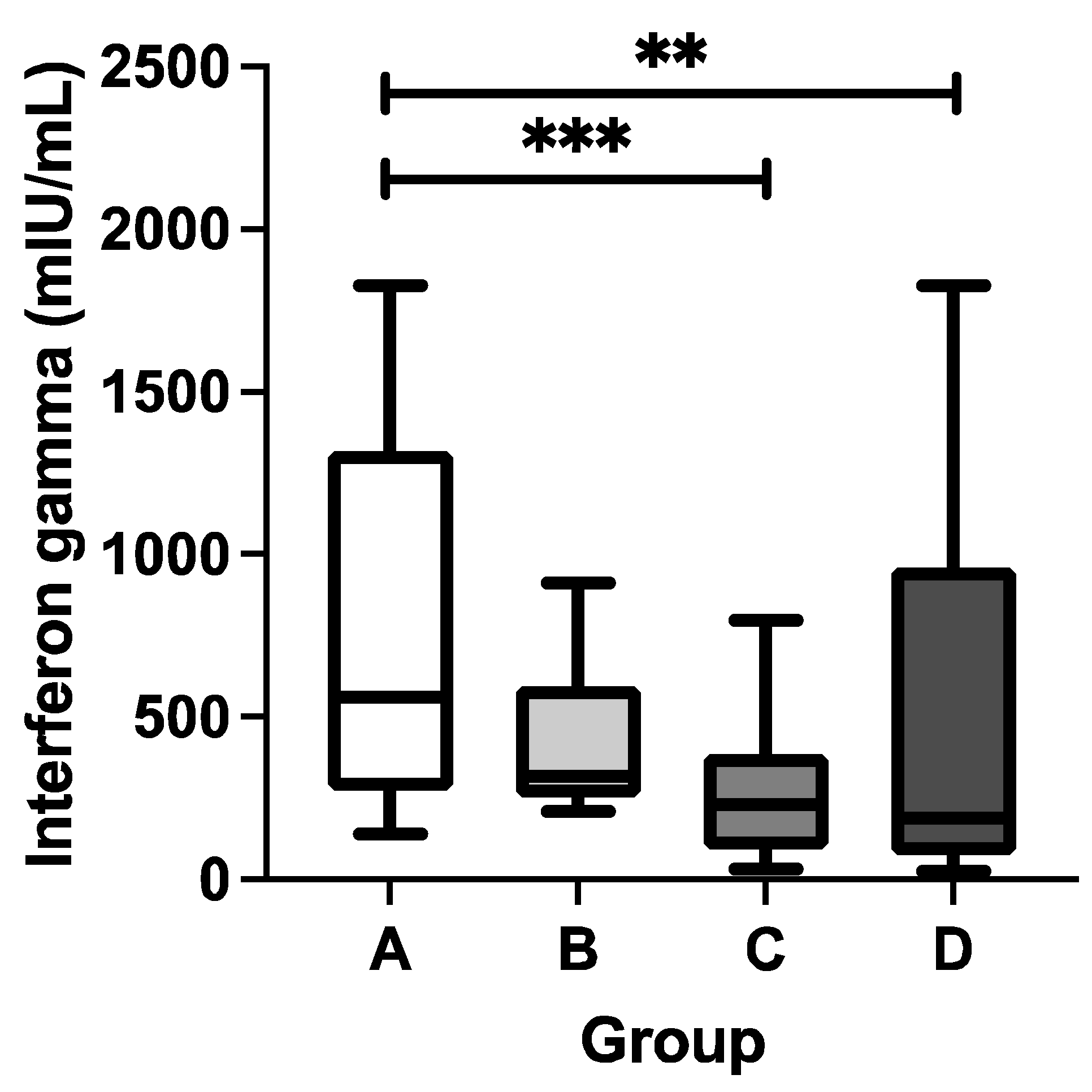
Figure 3.
Proposed mechanism of heterologous immunity by diphtheria-tetanus-pertussis vaccination in enhancing immune responses generated by inactivated SARS-CoV-2 vaccine in healthy children. (A) There are similar epitopes shared by diphtheria-tetanus-pertussis and inactivated SARS-CoV-2 vaccines, particularly with SARS-CoV-2 spike protein. (B) Upon administration of diphtheria-tetanus-pertussis vaccine or diphtheria-tetanus booster, certain peptide antigens will form the peptide-MHC complex to generate diphtheria-/tetanus-/pertussis-specific T cells. (C) Subsequently, upon administration of inactivated SARS-CoV-2 vaccine, certain diphtheria-/tetanus-/pertussis-specific T cells would respond to certain antigens of SARS-CoV-2 spike protein. This will generate a secondary immune response, resulting in heterologous immunity. (D) The heterologous immunity would also stimulate, at least, SARS-CoV-2 spike protein-specific T cells to produce various cytokines, including interferon gamma, at higher concentrations. (E) The activated CD4+ T cells would stimulate specific B cells to mature and release immunoglobulins. This figure was created with BioRender.com.
Figure 3.
Proposed mechanism of heterologous immunity by diphtheria-tetanus-pertussis vaccination in enhancing immune responses generated by inactivated SARS-CoV-2 vaccine in healthy children. (A) There are similar epitopes shared by diphtheria-tetanus-pertussis and inactivated SARS-CoV-2 vaccines, particularly with SARS-CoV-2 spike protein. (B) Upon administration of diphtheria-tetanus-pertussis vaccine or diphtheria-tetanus booster, certain peptide antigens will form the peptide-MHC complex to generate diphtheria-/tetanus-/pertussis-specific T cells. (C) Subsequently, upon administration of inactivated SARS-CoV-2 vaccine, certain diphtheria-/tetanus-/pertussis-specific T cells would respond to certain antigens of SARS-CoV-2 spike protein. This will generate a secondary immune response, resulting in heterologous immunity. (D) The heterologous immunity would also stimulate, at least, SARS-CoV-2 spike protein-specific T cells to produce various cytokines, including interferon gamma, at higher concentrations. (E) The activated CD4+ T cells would stimulate specific B cells to mature and release immunoglobulins. This figure was created with BioRender.com.
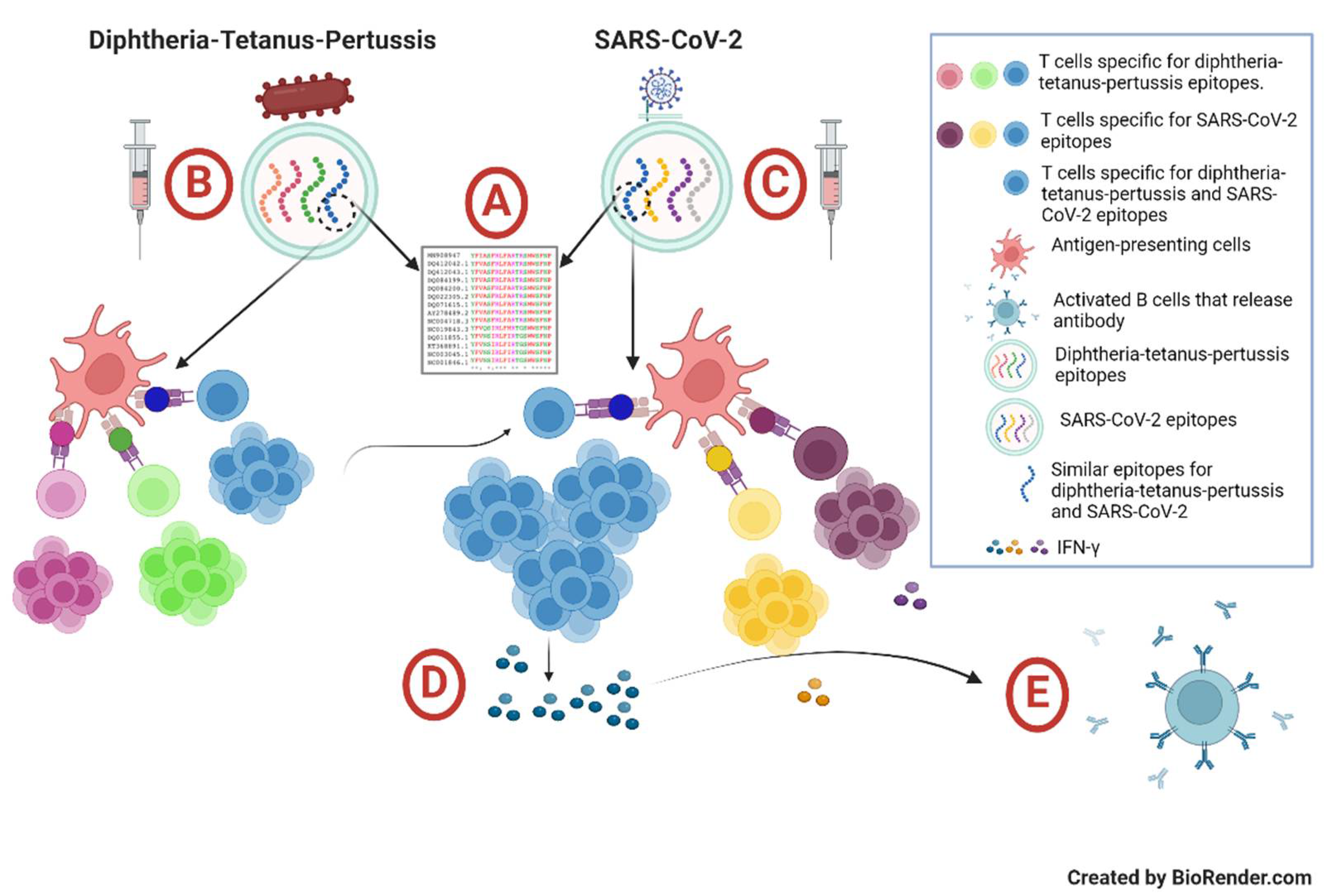
Table 2.
Titers of anti-SARS-CoV-2 S-RBD antibodies among the study participants.
| Group | n | Anti-SARS-CoV-2 S-RBD (U/mL) median (min–max) |
p-value |
|---|---|---|---|
| A [COVID-19 yes / DTP yes] | 39 | 1,196 (16–15,561) | 0.089 |
| B [COVID-19 yes / DTP no] | 38 | 771.2 (0.3–14,589) | |
| C [COVID-19 no / DTP yes] | 38 | 1,162.5 (0.3–22,269) | |
| D [COVID-19 no / DTP no] | 39 | 527.9 (0.3–9,060) |
Kruskal-Wallis test was performed with p-value <0.05 was considered as statistically significant.
Table 3.
Titers of anti-SARS-CoV-2 S-RBD antibodies based on DTP vaccination status.
| DTP Vaccination Status | n | Anti-SARS-CoV-2 S-RBD (U/mL) median (min–max) |
p-value |
|---|---|---|---|
| Yes | 77 | 1,182 (0.3–22,269) | 0.026 |
| No | 77 | 612.5 (0.3–14,589) |
Mann-Whitney test was performed with p-value <0.05 was considered as statistically significant.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



