Submitted:
07 February 2026
Posted:
09 February 2026
You are already at the latest version
Abstract
Acute and chronic kidney diseases remain significant challenges in regenerative medi-cine, with few therapies capable of reversing tissue injury or preventing progression. Bone marrow mesenchymal stem cell–derived exosomes (BM-MSC-Exos), nanosized vesicles measuring 30–150 nm, have emerged as multifunctional nanotheranostic platforms that integrate targeted therapeutic activity with imaging-enabled monitor-ing. In renal pathophysiology, BM-MSC-Exos exert anti-inflammatory, antifibrotic, angiogenic, and pro-regenerative effects mediated by microRNAs, messenger RNAs, mitochondrial regulators, and bioactive proteins that modulate epithelial repair and immune responses. Advances in bioengineering now allow precise modification of BM-MSC-Exos, including enrichment with synthetic RNAs or gene-editing compo-nents and membrane functionalization to enhance kidney-specific tropism. Parallel developments in fluorescent, bioluminescent, and nanoparticle-based imaging enable real-time visualization of biodistribution and tubular uptake, providing essential in-sights into therapeutic engagement. This review synthesizes current knowledge on BM-MSC-Exos in renal repair, describes contemporary strategies for cargo and surface engineering, examines imaging methodologies for in vivo tracking, and discusses how administration route influences renal targeting. An updated overview of clinical trials applying exosomes as therapeutic agents or biomarkers in nephrology is also present-ed. Collectively, engineered BM-MSC-Exos represent a promising and increasingly so-phisticated nanotheranostic platform for precision-guided treatment of kidney disease, supported by imaging tools that facilitate preclinical evaluation of biodistribution and efficacy.
Keywords:
bone marrow mesenchymal stem cell–derived exosomes
; engineered exosomes
; nanotheranostics
1. Introduction
Extracellular vesicles (EVs) are nanoscale, membrane-bound structures released by virtually all cell types and responsible for transferring bioactive molecules that shape intercellular communication in health and disease [1]. Historically, EVs were categorized into apoptotic bodies, microvesicles, and exosomes based on size and presumed biogenesis; however, the MISEV2023 guidelines emphasize that these categories rarely correspond to experimentally verifiable entities. Instead, they should be interpreted as operational descriptors anchored in measurable physical and molecular properties [2]. Density ranges traditionally associated with EVs subtypes (1.10–1.30 g/cm³) also represent enrichment rather than discrete biological identities. To consolidate experimental requirements introduced by MISEV2023, Table 1 summarizes recommended isolation strategies, characterization markers, and purity metrics.
Beyond size or density, EVs exhibit heterogeneous molecular compositions. Distinct RNA signatures, including miRNAs, long non-coding RNAs (lncRNAs), circular RNAs (circRNAs), and double-stranded DNA fragments, vary across vesicle populations and hold potential for disease stratification. Exosomes commonly express tetraspanins (CD9, CD63, CD81), heat shock proteins, and components of the endosomal sorting complex required for transport (ESCRT), whereas larger EVs may carry cytoskeletal and mitochondrial proteins. Importantly, no single marker is exclusive to a specific EVs subtype, reinforcing the need for multiparametric approaches to EVs identification [1,2,3].
MISEV2023 underscores the need for orthogonal isolation and characterization strategies to overcome longstanding challenges in purity, specificity, and reproducibility [2]. The guidelines recommend combining density-, size-, and affinity-based approaches and mandate transparent reporting of instrument calibration, negative controls, and inter-batch reproducibility. These recommendations seek to harmonize methodological rigor across studies and facilitate accurate interpretation of EVs-mediated biological effects [2].
Exosomes are commonly described as small EVs originating from the endosomal pathway, formed by inward budding of late endosomal membranes to generate intraluminal vesicles (ILVs) within multivesicular bodies (MVBs), which are released upon fusion with the plasma membrane [4,5]. Cargo selection involves ESCRT-dependent and ESCRT-independent mechanisms mediated by ceramides, sphingolipids, and tetraspanins [4,5]. After secretion, small EVs can interact with target cells through clathrin-mediated endocytosis, macropinocytosis, phagocytosis, or direct membrane fusion; however, mechanisms governing cytosolic cargo release remain incompletely defined [4,5]. EV molecular content varies according to parental cell type and physiological state, a concept summarized in Figure 1, which depicts EV composition and endosomal biogenesis while highlighting representative proteins, lipids, and RNA species [1,2,3].
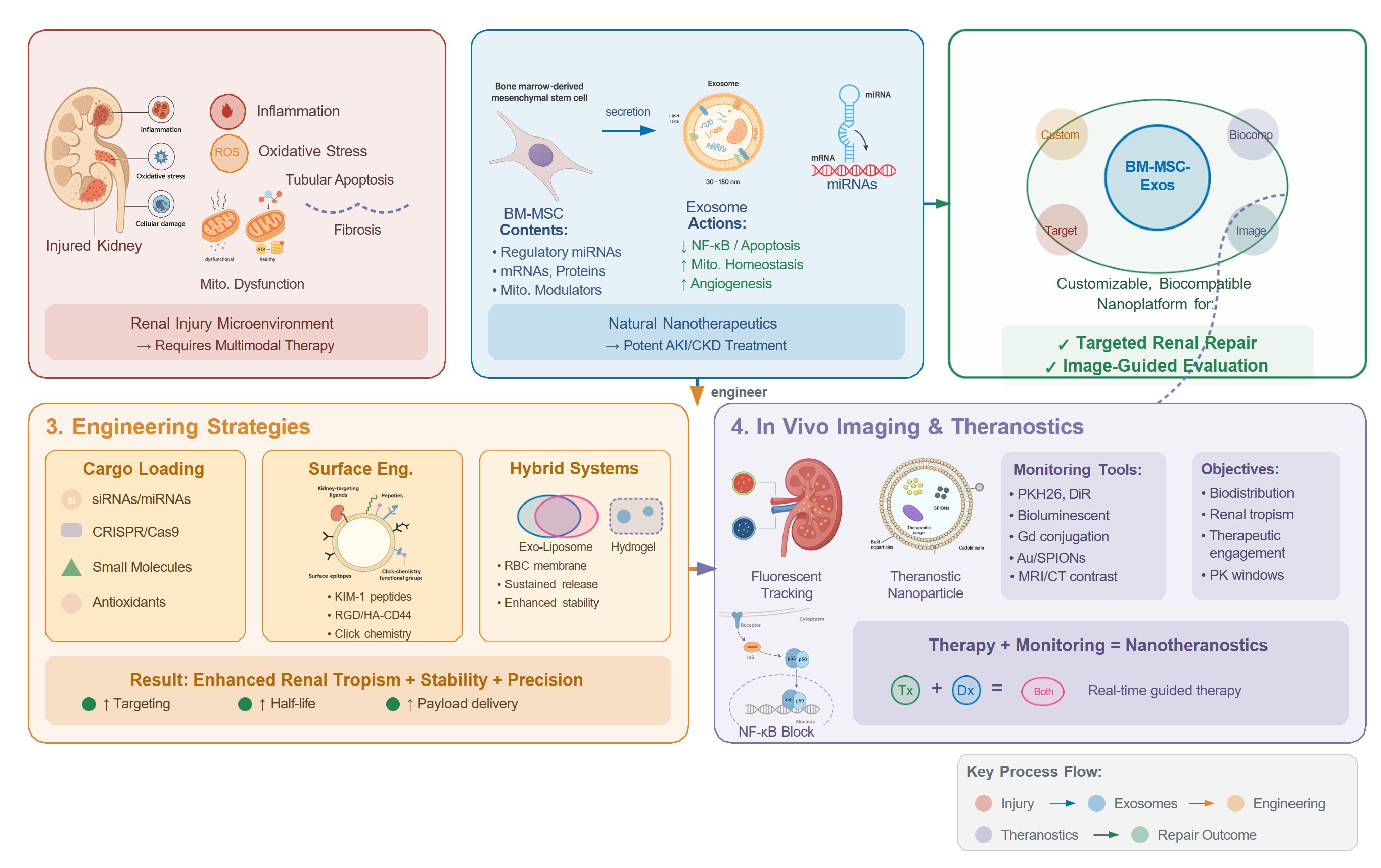
In regenerative medicine, exosomes have gained prominence as principal mediators of the paracrine effects of mesenchymal stem cells (MSCs). They regulate angiogenesis, inflammation, fibrosis, and oxidative stress, providing a cell-free therapeutic alternative with improved safety, stability, and regulatory feasibility compared with direct MSC transplantation [6,7]. These advantages have stimulated interest in bone marrow mesenchymal stem cell–derived exosomes (BM-MSC-Exos) as modular biological agents capable of orchestrating tissue repair across multiple organ systems [8]. Because exosomal cargo reflects the physiological and transcriptional phenotype of donor cells, engineering of BM-MSC-Exos enables enrichment or loading of therapeutic molecules, including miRNAs, small interfering RNAs (siRNAs), messenger RNAs (mRNAs), recombinant proteins, mitochondrial regulators, and epigenetic modifiers [9]. This engineerability supports next-generation nanotheranostic concepts integrating targeted delivery with molecular imaging and biomarker-based monitoring of therapeutic response.
These advances are especially relevant to nephrology, where kidney diseases affect more than 844 million individuals worldwide and represent major drivers of morbidity and premature mortality [10]. Exosomes have demonstrated promise in modulating pathways central to renal injury, including inflammation, fibrosis, apoptosis, mitochondrial dysfunction, and microvascular damage. Urinary and circulating exosomes also represent emerging sources of diagnostic biomarkers for early disease detection [11]. BM-MSC-Exos have shown nephroprotective potential in both acute and chronic kidney injury models and provide a platform for engineering therapeutically enhanced vesicles with improved targeting and functional potency [12].
While recent reviews have discussed MSC-derived extracellular vesicles in renal repair or have separately surveyed exosome engineering technologies, this review advances an integrated renal nanotheranostic engineering-by-design framework for bone marrow MSC-derived exosomes. We connect kidney injury biology and injury-associated targetable cues with the rational selection of engineering strategies (cargo programming, membrane functionalization, and hybrid/biomimetic systems) to improve renal tropism and target engagement. Importantly, we link these design choices to imaging-enabled tracking and route-of-administration considerations to support in vivo verification of biodistribution and therapeutic engagement.
2. Materials and Methods
This manuscript follows a narrative review format. A literature search was conducted to identify peer-reviewed studies related to bone marrow mesenchymal stem cell–derived exosomes/extracellular vesicles (BM-MSC-Exos/EVs), renal injury and regeneration, exosome engineering, and exosome-based nanotheranostics. Searches were performed in PubMed/MEDLINE, Scopus, Web of Science, Google Scholar, and the Directory of Open Access Journals (DOAJ) from database inception to 10 November 2025 (last searched on 10 November 2025). Search terms combined exosome-, MSC-, kidney-, and engineering-related keywords, including but not limited to: “bone marrow MSC exosomes,” “MSC-derived EVs,” “renal exosomes,” “exosome engineering,” “endogenous cargo loading,” “exogenous loading,” “hybrid exosomes,” “EV mimetics,” “exosome MRI,” “gadolinium exosomes,” “gold nanoparticle exosomes,” “radioisotope-labeled EVs,” “kidney regeneration,” “AKI exosomes,” “CKD exosomes,” “exosome theranostics,” “renal nanomedicine,” and “personalized nanotherapy.”
Eligibility criteria were defined a priori. Only peer-reviewed publications written in English were included; editorials, preprints, and conference abstracts were excluded. Records retrieved from all sources were consolidated and deduplicated using DOI/title/author matching, followed by manual verification. Google Scholar and DOAJ were used as supplementary discovery sources; items retrieved through these platforms were included only after confirmation of peer-review status and credible journal provenance. Study relevance was prioritized based on kidney-disease applicability, BM-MSC-Exos/MSC-EVs specificity, mechanistic depth, and translational/engineering significance; items that did not meet these criteria were excluded.
In selected mechanistic subsections addressing rapidly evolving topics, we prioritized post-2024 mechanistic studies to reflect recent advances while retaining seminal earlier work when required for conceptual continuity. Because this is a narrative review, no PRISMA workflow or quantitative pooling of outcomes (meta-analysis) was performed.
A structured query of ClinicalTrials.gov (“exosome OR exosomes”) was performed and last accessed on 10 November 2025. Only studies with recruitment status listed as completed or recruiting were considered for summary, recognizing that individual protocols may span more than one category (e.g., diagnostics and biomarkers).
3. Results and Discussion
3.1. Bone Marrow Mesenchymal Stem Cell–Derived Exosomes (BM-MSC-Exos)
BM-MSC-Exos are enriched in regulatory RNAs, proteins, and lipids that collectively modulate inflammation, fibrosis, mitochondrial homeostasis, and epithelial repair—processes central to acute and chronic kidney injury [12]. In nephrology, they have gained relevance as therapeutic and diagnostic platforms and as engineerable vesicles whose cargo and surface can be tuned to improve renal target engagement [10,11,12,13]. The following sections summarize key properties, mechanisms, and preclinical evidence, and then focus on engineering strategies and translational considerations for kidney disease.
3.1.1. General Properties and Advantages
BM-MSC-Exos have emerged as an alternative to classical MSC therapy by recapitulating key paracrine activities of their parental cells while avoiding several risks and logistical constraints associated with administering live cells [1,15]. As acellular products lacking a replicative nucleus, they cannot proliferate, differentiate, or undergo malignant transformation, and they may reduce infusion-related complications such as pulmonary microvascular entrapment [14]. These features generally support a favorable safety and handling profile and can simplify aspects of manufacturing and oversight compared with cell-based approaches [1,15].
BM-MSC-Exos carry a complex and functionally relevant cargo of RNAs, proteins, and lipids. Proteomic analyses have reported >857 proteins in MSC-Exos, and microarray studies have detected >2,000 miRNAs, with extensive overlap with parental MSC miRNA repertoires (reported at ~98%), supporting biological continuity between donor cells and their vesicular products [16]. This cargo exhibits immunomodulatory, cytoprotective, pro-angiogenic, and antifibrotic activities and may contribute to the regulation of mitochondrial homeostasis, redox balance, and extracellular matrix remodeling—pathways directly implicated in renal injury responses. The nanoscale vesicular structure also helps protect labile molecules (e.g., miRNAs) from extracellular degradation, supporting sustained bioactivity in inflamed or hypoxic microenvironments.
Importantly, the functional profile of MSC-Exos varies depending on the source tissue. Bone marrow–derived mesenchymal stromal cells (BM-MSC) are among the most widely used MSC sources and have been used in more clinical trials than MSC derived from other tissues (clinicaltrials.gov), providing a long-standing benchmark platform with extensive characterization. However, BM-MSC procurement requires an invasive bone marrow aspiration procedure, which has motivated the search for alternative sources that are more easily obtained in larger quantities, including adipose tissue and umbilical cord [1,13,14,15,16]. In line with this source dependence, BM-MSCs are well characterized and strongly immunomodulatory, whereas exosomes from adipose-derived MSCs (AD-MSC-Exos) and umbilical cord–derived MSCs (UC-MSC-Exos) may exhibit distinct angiogenic, anti-inflammatory, or proliferative signatures [13,14,15,16]. These differences arise from intrinsic transcriptional programs, metabolic states, and extracellular cues present in each tissue niche. Furthermore, donor age, sex, and health status influence vesicular content, miRNA abundance, and therapeutic potency, affecting outcomes in renal injury models and underscoring the need for standardized sourcing protocols and potency assays [1,13,14,15,16].
Comparative evidence indicates that adipose-derived MSC are extensively investigated due to ease of isolation and high tissue availability, while umbilical cord–derived MSC are conveniently harvested and efficiently isolated; moreover, UC-MSC show highly overlapping gene-expression profiles compared with BM-MSCs [1,13,14,15,16]. Despite the translational attractiveness of these sources, BM-MSC-Exos were selected as the focus of this review because they represent the most frequently studied reference platform in preclinical nephrology, with a robust mechanistic literature that anchors engineering-by-design and translational discussions. Nonetheless, harmonized head-to-head comparisons across MSC-Exo sources using standardized isolation, dosing metrics, and renal-relevant potency assays remain essential to determine whether source selection yields disease-tailored advantages or broadly interchangeable efficacy in kidney disease.
From a translational perspective, exosomes offer logistical and manufacturing advantages, including long-term storage at −80 °C and potential lyophilization. Compared with BM-MSCs, BM-MSC-Exos are easier to standardize and handle, as summarized in Table 2. Supported by the International Society for Cell and Gene Therapy (ISCT), BM-MSC-Exos are increasingly recognized as distinct biological medicinal products, provided they meet rigorous characterization and comply with Good Manufacturing Practice (GMP) standards [17].
In vivo, BM-MSC-Exos exhibit favorable biodistribution, reaching dense and relatively poorly vascularized tissues such as the kidney, liver, and central nervous system [18,19]. They avoid pulmonary trapping and preferentially accumulate in injured renal tissue, with comparatively reduced hepatic and splenic uptake [20]. This renal tropism is partly mediated by exosomal surface proteins, including integrins (α4β1, α6β4) and tetraspanins (CD9, CD63, CD81), which support vascular recognition and homing to sites of injury [19].
At the membrane level, BM-MSC-Exos are enriched in ceramides, cholesterol, phosphatidylserine, and sphingomyelin, which contribute to structural rigidity, maintain vesicular curvature, and increase resistance to lysosomal degradation [21,22]. This lipid architecture also helps preserve internal cargo (miRNAs, mRNAs, and proteins) from degradation by circulating RNases and proteases, sustaining bioactivity in vivo [21,22].
For large-scale production, each MSC can release approximately 10⁴–10⁵ vesicles per day, enabling scalable manufacturing using closed-loop bioreactors and three-dimensional culture systems under GMP conditions [17,23,24]. These platforms support controlled purity, sterility, molecular identity, and potency, facilitating clinical translation.
After administration, BM-MSC-Exos are internalized by renal tubular epithelial cells via clathrin-mediated endocytosis, caveolin-dependent uptake, macropinocytosis, phagocytosis, and direct membrane fusion; uptake is enhanced by phosphatidylserine exposure and injury-induced receptor expression in renal tissues [22,23,24,25]. This enables intracellular delivery of regulatory molecules that modulate inflammation, mitochondrial homeostasis, oxidative stress, and apoptotic signaling.
BM-MSC-Exos maintain integrity after repeated freeze–thaw cycles, supporting practical storage and transport workflows. Their naturally encoded cargo includes anti-inflammatory miRNAs (miR-21, miR-146a, miR-223), pro-angiogenic mRNAs, and anti-apoptotic proteins [26,27,28]. Preconditioning strategies can further enhance exosomal bioactivity. Hypoxia, serum deprivation, three-dimensional culture, lipopolysaccharide (LPS) stimulation, and nitric oxide donors alter MSC gene expression and enrich exosomes with survival- and repair-promoting molecules such as HIF-1α and Bcl-2 family proteins [1,29].
Despite these advantages, limitations remain, including non-standardized isolation and quantification methods, variability in yield and purity, and batch-to-batch differences, particularly with donor-derived MSC sources [1,2,17]. Potency assays are still under development, regulatory frameworks continue to evolve, and purification workflows may co-isolate non-vesicular contaminants (e.g., lipoproteins or protein aggregates), affecting reproducibility and safety [2,17]. Nevertheless, available evidence supports BM-MSC-Exos as a clinically promising nanotherapeutic platform for kidney disease, with advantages over cell-based therapy summarized in Table 2. The following sections review roles in AKI and CKD and outline engineering strategies to enhance activity through cargo modulation.
3.1.2. BM-MSC-Exos: Acute Kidney Injury (AKI)
AKI is characterized by a rapid decline in renal function triggered by heterogeneous insults such as ischemia–reperfusion injury (IRI), nephrotoxicity, sepsis, hypoxia, or rhabdomyolysis [30,31]. BM-MSC-Exos are attractive candidates in this setting because their nanoscale size may improve access to injured renal microenvironments where edema, microvascular dysfunction, and impaired perfusion can limit delivery and effectiveness of cellular interventions [1,30,31]. Importantly, their engineerability enables renal-targeting and mechanistic probing in a disease context defined by biological heterogeneity and narrow therapeutic windows [13].
Mechanistically, BM-MSC-Exos convey regulatory miRNAs and proteins to tubular, endothelial, and immune cells, modulating pathways involved in apoptosis, mitochondrial homeostasis, oxidative stress, and inflammation [32,33]. Their lipid-enriched membranes, including ceramides and sphingomyelin, facilitate docking and intracellular trafficking, enabling delivery of bioactive cargo [32,33]. Many mechanistic insights remain associative; therefore, claims are framed according to strength of evidence (causal, supportive, or correlative), with emphasis on variability related to model design and dosing strategies.
To identify representative studies on BM-MSC-Exos in AKI, a targeted PubMed query using terms related to mesenchymal exosomes and acute kidney injury yielded 64 records. After excluding 24 reviews, 40 original research articles were evaluated, prioritizing recent and mechanistically informative studies published from 2024 onward.
Among these, Ji et al. [34] reported protective effects of exosomal miR-127-3p in an IRI model, associated with KIF3B and Gli1 suppression, reduced Hedgehog signaling, and modulation of autophagy-related proteins (ATG5, ATG7). This axis is considered supportive or partially causal (Tier 2) and would benefit from further validation and replication across non-IRI models.
Taghavi et al. [35] developed microneedle patches for perirenal delivery, achieving better functional outcomes (BUN, creatinine) and lower oxidative/apoptotic markers than intravenous administration. Kong et al. [36], in a nephrotoxic setting, demonstrated that exosomal delivery of miR-874-3p mitigated cisplatin-induced injury, implicating activation of Wnt/β-catenin signaling in protecting tubular and vascular compartments. Given the context-dependent nature of this pathway in kidney injury, confirming miRNA dependency and pathway specificity is essential [36].
In inflammatory AKI, Yang et al. [37] showed that curcumin-preconditioned BM-MSC-Exos conferred protection in sepsis-associated injury via FTO-mediated m6A regulation of OXSR1, with concurrent reductions in oxidative damage and inflammatory signaling. At the same time, these findings suggest an epitranscriptomic axis; the proposed causal chain (FTO, m6A, and OXSR1) warrants further support through functional validation. Çam et al. [38], using a human AKI-on-a-chip platform, found that allogeneic BM-MSC-Exos restored tubular barrier integrity and promoted cell proliferation under hypoxic conditions, emphasizing the translational value of human-relevant models and precise dosing kinetics.
Chen and Hou [39] introduced a distinct strategy for rhabdomyolysis-associated AKI, engineering magnetic nanoparticle-loaded BM-MSC-Exos to target upstream muscle injury. Under magnetic guidance, vesicles localized to damaged muscle, reduced myoglobin release, and attenuated secondary renal damage, illustrating the potential of indirect targeting in multi-organ AKI scenarios.
Across preclinical AKI models (IRI, nephrotoxicity, sepsis, and rhabdomyolysis), BM-MSC-Exos consistently reduce tubular injury and improve renal function via convergent mechanisms that involve mitochondrial stability, redox control, apoptosis regulation, and immunomodulation [32,33,34,35,36,37,38,39]. However, cross-study comparability is limited by differences in etiology, timing, delivery route (systemic vs. local), dose metrics (particle number vs. protein mass), and outcome measures. To enhance translational relevance, future studies should report particle quantification methods, dose and purity criteria, administration protocols, and, when possible, biodistribution data. Overall, these findings support BM-MSC-Exos as versatile, cell-free candidates for AKI therapy, while underscoring the need for standardized protocols and robust causal validation as the field progresses toward clinical application [32,33,34,35,36,37,38,39].
3.1.3. BM-MSC-Exos: Chronic Kidney Disease (CKD)
CKD evolves through an interconnected set of pathogenic processes, including persistent low-grade inflammation, redox imbalance, mitochondrial dysfunction, maladaptive cell-death programs, and excessive extracellular matrix deposition. These alterations converge on interstitial fibrosis and glomerulosclerosis, the histopathological hallmarks of progressive nephron loss [40,41]. Because these pathways are nonlinear and mutually reinforcing, interventions capable of modulating both immunometabolic stress and profibrotic signaling are particularly promising, given the limited efficacy of single-node targeting in CKD [40,41].
In this context, BM-MSC-derived exosomes (BM-MSC-Exos) have emerged as nanoscale, cell-free platforms capable of transferring bioactive miRNAs, proteins, and lipids that modulate disease-relevant signaling networks across CKD etiologies [12,42]. However, mechanistic interpretation and translational comparability remain limited by heterogeneity in CKD models (autoimmune, metabolic, infectious), timing of intervention, route of administration, dose normalization (particle number vs. protein mass), and endpoints (albuminuria, fibrosis, inflammation, and functional markers).
To capture current trends, a structured PubMed search was conducted using terms for mesenchymal exosomes and chronic kidney disease, yielding 109 studies. After excluding 32 reviews, 77 original research articles were retained, and the most recent, mechanistically informative publications (2024 onward) were prioritized.
Among these, Tian et al. [43] engineered MSC-Exos loaded with cerium oxide nanoparticles and rapamycin (CEX@Rapa) for the treatment of lupus nephritis. In murine models, CEX@Rapa accumulated in inflamed renal tissue and reduced oxidative stress and cytokine levels, suggesting antioxidant and immunosuppressive synergy. However, claims of renal specificity should be interpreted in light of biodistribution data and potential off-target accumulation inherent to nanoparticle-EV hybrids.
In metabolic CKD, Zhang et al. [44] reported that BM-MSC-Exos attenuated ferroptosis-associated injury in a type 1 diabetes model, marked by increased GPX4, reduced lipid peroxidation, and improved early renal function. These findings suggest ferroptosis involvement, but the mechanism remains correlational (Tier 3) without pathway-specific manipulation, such as ferroptosis inhibition, GPX4 modulation, or rescue experiments [44]. Furthermore, systemic metabolic improvements complicate attribution of renal effects to direct exosome activity versus secondary endocrine influences.
Wang et al. [45] demonstrated that BM-MSC-Exos mitigate diabetic kidney injury by suppressing NOD2-mediated sterile inflammation. In high-glucose-stimulated podocytes, exosomes reduced apoptosis, oxidative stress, and cytokine production, with in vivo evidence of functional and structural improvement. While this NOD2 axis appears mechanistically plausible (Tier 2), it awaits confirmation through loss-of-function or rescue strategies in target cells or whole-animal models [45].
In chronic inflammatory glomerulopathies, Chen et al. [46] found that BM-MSC-Exos enriched with miR-223-3p downregulated HDAC2 and reduced STAT3 phosphorylation in a hepatitis B virus-associated glomerulonephritis model, thereby decreasing lipid peroxidation and iron accumulation and improving podocyte viability. Although this miRNA-pathway interaction is consistent with Tier 2 evidence, further validation via independent replication and explicit miRNA dependency is warranted [46].
Collectively, these studies support the potential of BM-MSC-Exos as modulators of inflammatory stress, redox imbalance, and profibrotic remodeling in CKD through miRNA- and protein-mediated mechanisms [40,41,42,43,44,45,46]. Nonetheless, consistent interpretation across studies requires attention to model timing (early vs. advanced disease), route of administration, exposure kinetics, and standardized outcome measures. Future efforts should prioritize harmonization of dose metrics and purification protocols, and the inclusion of biodistribution and target engagement analyses, alongside rigorous causal validation, to strengthen the translational readiness of BM-MSC-Exos in CKD [12,40,41,42,43,44,45,46].
3.1.4. Exosomes as Biosensors and Biomarkers in Kidney Disease
Although BM-MSC-Exos are not directly applicable for clinical diagnostics due to their localization in the bone marrow niche and the invasiveness of collection, they exhibit remarkable sensitivity to systemic physiological alterations. In response to inflammation, hypoxia, oxidative stress, metabolic imbalance, and aging, BM-MSCs dynamically remodel the RNA, protein, and lipid cargo of their exosomes [1,26,47]. As such, even if clinically inaccessible, BM-MSC-Exos function as systemic biosensors reflecting organism-wide pathophysiological states, which is particularly relevant in CKD.
This biosensory capacity was demonstrated by Di Trapani et al. [48], who showed that MSCs exposed to different immune stimuli secreted exosomes with distinct immunomodulatory profiles. Pro-inflammatory cytokine exposure enhanced the immunosuppressive molecular signature of MSC-Exos, confirming that exosomal cargo mirrors the inflammatory and metabolic environment of the parent cell.
Further supporting this role, Li et al. [49] observed that the pro-osteogenic and pro-angiogenic effects of BM-MSC-Exos were significantly attenuated in steroid-induced osteonecrosis, indicating that disease-related stress can constrain exosomal bioactivity. Similarly, Luiz et al. [26] showed that BM-MSC-Exos from physically trained animals were enriched in miR-296 and other pro-angiogenic molecules, which, in a unilateral ureteral obstruction (UUO) model, reduced inflammation, improved vascular homeostasis, and mitigated hypoxia-induced injury. These findings reinforce the concept that BM-MSC-Exos act as dynamic biosensors of physiological health status.
In contrast, urinary exosomes (uEVs) are readily obtainable and have emerged as robust non-invasive biomarkers. Derived from diverse nephron segments (including podocytes, proximal and distal tubules, loop of Henle, collecting ducts) as well as from the urothelium and prostate, uEVs offer a segment-specific molecular snapshot of renal physiology and pathology [50,51]. Approximately 99.96% of uEV proteins originate from the urinary tract, and these vesicles also carry mitochondrial DNA, mRNAs, miRNAs, long non-coding RNAs, and lipids implicated in inflammation, fibrosis, apoptosis, oxidative stress, and mitochondrial dysfunction [11,52,53].
Segment-specific markers within uEVs include Sodium/Hydrogen Exchanger 3 (NHE3) from the proximal tubule; Aquaporin-1, uromodulin, and Tamm–Horsfall protein from the loop of Henle; and Aquaporin-2 from the collecting duct [51,52,53,54]. miRNAs such as miR-21, miR-29, miR-192, and miR-200 (frequently detected in uEVs) have been consistently associated with renal fibrosis, podocyte injury, diabetic nephropathy, and acute kidney injury, supporting their use in diagnosis, risk stratification, and disease monitoring [50,51,52,53,54].
The clinical viability of exosome-based diagnostics was exemplified by the FDA approval of the ExoDx Prostate (IntelliScore) test, which uses urinary exosomal RNA to detect clinically significant prostate cancer non-invasively. This success underscores the translational feasibility of exosome-based assays and suggests potential applications in nephrology [56].
In summary, uEVs provide a non-invasive platform for segment-specific molecular profiling across the spectrum of kidney diseases. While BM-MSC-Exos function primarily as therapeutic vesicles and systemic biosensors of inflammation, oxidative stress, and metabolic dysfunction, uEVs complement this by enabling accurate diagnosis, prognosis, and longitudinal monitoring. This functional convergence lays the groundwork for future nanotheranostic strategies in nephrology. However, the clinical translation of urinary exosome biomarkers remains limited by challenges in standardizing isolation procedures, ensuring reproducibility, and interpreting molecular signatures across heterogeneous populations.
3.1.5. Exosomes: Clinical Trials
The growing body of preclinical evidence supporting the therapeutic potential of exosomes has spurred their inclusion in numerous clinical trials. Successful translation to clinical settings, however, requires strict adherence to Good Manufacturing Practice (GMP) standards [17,56], which govern donor selection, culture conditions, media formulation, and bioprocessing parameters. Isolated exosomes must undergo multi-step purification and rigorous structural, biochemical, and functional characterization to ensure reproducibility, potency, and safety. Regulatory guidance from agencies such as the European Medicines Agency (EMA), particularly in the context of Advanced Therapy Medicinal Products (ATMPs), is critical to the development of engineered or cargo-enhanced exosome therapeutics [57].
BM-MSC-Exos dosing in clinical studies varies significantly due to inconsistent quantification strategies (e.g., particle count vs. total protein), divergent isolation protocols, and limited standardization of potency assays [1,32,58,59]. As a result, no universally accepted therapeutic dose has been established, and while intravenous administration remains the most common route, alternative delivery strategies such as microneedle patches, localized injections, and targeted formulations are under active investigation [58,59].
A search of the ClinicalTrials.gov database using the query “exosome” OR “exosomes” identified four major clinical categories: diagnostics, therapy, biomarkers, and drug development. Only studies listed as completed or recruiting were included, regardless of disease subtype. The search yielded 42 completed and 60 recruiting diagnostic trials; 45 completed and 72 ongoing therapeutic trials; 21 completed and 24 recruiting biomarker-based trials; and 34 completed and 45 recruiting drug development studies. Because many clinical protocols are multipurpose, a single study may fall into more than one category. Figure 2 summarizes the percentage distribution of ClinicalTrials.gov-registered exosome trials. Among completed studies, therapeutic (32%) and diagnostic (29%) applications predominated, consistent with an initial focus on safety and biomarker discovery. Recruiting studies showed a shift toward therapy (36%) and diagnostics (30%), suggesting growing confidence in exosomes as both therapeutic agents and diagnostic platforms. Drug delivery and biomarker trials are also increasing, underscoring expanding interest in engineered and precision-medicine applications of exosomes
CKD remains a significant global healthcare burden, frequently arising from underlying conditions such as diabetic nephropathy, lupus nephritis, hypertensive nephrosclerosis, and chronic allograft dysfunction. Within ClinicalTrials.gov, 14 exosome-related clinical trials addressing renal disorders were identified. These include studies registered under “kidney diseases” (9 trials), “diabetic nephropathy” (2), “lupus nephritis” (2), and “graft fibrosis” (1). Collectively, they highlight the expanding interest in exosome-based diagnostics and molecular monitoring in nephrology. Examples of exosome-related clinical trials in nephrology include:
- 1. NCT03503461 (France): Urinary exosomal NCC and pendrin evaluated as biomarkers of post-transplant hypertension in calcineurin inhibitor–treated kidney transplant recipients.
- 2. NCT05695573 (Egypt): UMOD gene and uromodulin protein in urinary exosomes assessed as early biomarkers for type 2 diabetes–related nephropathy.
- 3. NCT04142138 (USA): Proteomic profiling of urinary exosomes in individuals with prehypertension following DASH diet intervention.
- 4. NCT04536688 (USA): RGLS4326 therapy in ADPKD patients monitored through urinary exosomal PC- 1 and PC-2 profiles.
- 5. NCT03227055 (Taiwan): Urinary exosomal miRNAs as early cardiovascular–renal risk markers in pediatric CKD (G1–G4).
- 6. NCT03202212 (Italy): Changes in plasma EVs-RNA content and inflammatory markers in dialysis patients undergoing mixed hemodiafiltration.
- 7. NCT03034265 (Switzerland): Urinary sodium, channel exosomal profiles, and plasma angiotensin peptides assessed in resistant hypertension.
- 8. NCT02226055 (Sri Lanka): Urinary exosomes, proteomics, genotoxic markers, and vascular stiffness evaluated in CKDu pathogenesis.
- 9. NCT02822131 (Switzerland): Urinary exosomal transporter proteins (NCC, NaPi, IIa) assessed after controlled phosphate intake.
- 10. NCT06123871 (China): Urinary exosomal miR-136-5p investigated as an early biomarker of diabetic nephropathy.
-
11. NCT06097351 (China): Diagnostic potential of urinary exosomal miR-142-3p in diabetic kidney disease.12. NCT04894695 (China): Identification of urinary exosomal biomarkers in lupus nephritis.
- 13. NCT04534647 (Latin America): Validation of exosomal signatures for lupus nephritis activity and progression.
- 14. NCT03487861 (South Korea): Exosomal transglutaminase 2 (TG2) evaluated as a biomarker of chronic allograft fibrosis after kidney transplantation.
Collectively, these studies reflect progress toward integrating exosome-based biomarkers into precision nephrology. Nonetheless, translation is limited by heterogeneity in EV isolation, lack of standardized quantification and dosing metrics, and absence of validated diagnostic thresholds [1,32,58,59]. Harmonized protocols, renal-relevant potency assays, and clinically meaningful endpoints are essential for moving exosome-based tools into routine practice.
Despite encouraging early clinical advances, manufacturing remains a significant barrier for BM-MSC-Exos in renal therapeutics. Upstream challenges include donor and passage variability, inconsistent yield, and dependence on culture conditions such as serum-free or EV-depleted media. These limitations have led to the development of closed, scalable expansion platforms, such as multilayer flasks and bioreactors. Downstream, ultracentrifugation remains difficult to scale, while filtration and chromatography-based purification must be optimized to preserve vesicle integrity and remove contaminants. GMP-grade production requires stringent release criteria, including assessments of identity, purity, sterility, endotoxin levels, residual DNA/protein content, stability, and renal-targeted potency assays [17,56].
Reported dosing of BM-MSC-Exos varies widely across studies, including particle number and protein mass, which complicates cross-study interpretation and translation [1,32,58,59]. To improve comparability, we recommend a harmonized reporting framework aligned with MISEV2023 principles [2]. Particle dose and protein mass should be reported concurrently, including methodological details for particle quantification and protein measurement [2]. The particle-to-protein ratio should be provided as a practical indicator of preparation quality [2]. In vivo dosing should be normalized to recipient body weight and accompanied by explicit reporting of administration route, infusion volume, dosing schedule, and formulation [2]. For engineered exosomes, cargo dose should be reported when feasible, such as drug mass or nucleic-acid content per administered dose. Physical dose metrics should be complemented by a fit-for-purpose potency assay and, when possible, a functional dose expressed in potency units relevant to renal repair mechanisms to enable batch comparability and clinical-grade lot release [2].
3.2. Engineering BM-MSC and Their Exosomes for Kidney Diseases
Native BM-MSC-derived exosomes (MSC-Exos) naturally carry miRNAs, proteins, and lipids with anti-inflammatory, antifibrotic, and pro-regenerative activities. Their therapeutic profile can be further enhanced through engineering strategies that modify their cargo or membrane to improve kidney-specific delivery, bioactivity, and diagnostic potential. These approaches transform MSC-Exos into versatile nanotheranostic platforms with increasing relevance in nephrology [13].
Exosome engineering strategies are broadly classified into biological, chemical, and physical approaches. Biological engineering involves genetic modification of donor BM-MSCs to enrich exosomal cargo with specific therapeutic agents, such as overexpressed miRNAs, regulatory proteins, or targeting peptides. Chemical methods include surface conjugation of ligands, imaging probes, or synthetic linkers to the exosomal membrane, enhancing targeting specificity, biodistribution, and potential for multimodal diagnostics [13].
Native exosomes contain lipid rafts, adhesion molecules, MHC proteins, tetraspanins, heat shock proteins, and nucleic acids, mediating intercellular communication through membrane fusion, receptor engagement, and endocytic uptake. Engineered exosomes may be functionalized with covalent ligands, targeting peptides, linker chemistries, nanotheranostic probes, growth factors, or therapeutic nucleic acids (miRNA/siRNA), enabling enhanced drug delivery and disease-specific targeting.
Physical engineering techniques such as sonication, electroporation, extrusion, freeze–thaw cycles, membrane permeabilization, and hypotonic dialysis are widely used to facilitate efficient loading of therapeutic cargo. These approaches allow the incorporation of nucleic acids, proteins, and small molecules with high efficiency, broadening the functional scope of engineered exosomes and enabling synergistic use of biological, chemical, and physical engineering methods [13] (Figure 3).
The kidney is an especially favorable target for engineered exosomes, as specific receptors are upregulated during injury. These include KIM-1, CD44, and various integrins, along with increased tubular permeability observed in both AKI and CKD [62,63]. However, each engineering approach carries limitations. Physical methods may compromise vesicle integrity, biological strategies introduce variability and regulatory complexity, and chemical modifications may leave behind cytotoxic reagents [13,64]. These considerations must be addressed to ensure safety, reproducibility, and clinical applicability.
Together, these foundational strategies support the development of next-generation exosomal systems for kidney disease. The following sections will explore key engineering modalities in greater depth, including surface functionalization, cargo manipulation, and the integration of hybrid or biomimetic designs aimed at optimizing targeting specificity, therapeutic potency, and in vivo traceability in nephrology applications.
3.2.1. Surface Engineering: Membrane Modification
Surface engineering is a key strategy to enhance the biodistribution, stability, and cellular specificity of exosomes in vivo [65]. The exosomal membrane, enriched in lipids, proteins, and glycoconjugates essential for intercellular communication, provides an ideal platform for functionalization. This can be achieved via non-covalent, covalent, or membrane-protein–based approaches, enabling attachment of targeting peptides, ligands, nucleic acids, or imaging probes. Such modifications increase renal tropism, improve circulation time, and reduce off-target effects, thereby advancing precision therapies for kidney disease [65].
Non-Covalent Engineering
Non-covalent engineering exploits hydrophobic, electrostatic, and ligand–receptor interactions to functionalize exosomes while preserving their native membrane architecture. Hydrophobic anchoring is widely applied using cholesterol, 1,2-distearoyl-sn-glycero-3-phosphoethanolamine (DSPE), polyethylene glycol (PEG) conjugates, and 1,2-dioleoyl-sn-glycero-3-phosphoethanolamine (DOPE)-PEG conjugates, which spontaneously insert into the lipid bilayer. This allows stable incorporation of oligonucleotides, aptamers, antibodies, or imaging agents. In nephrology, BM-MSC-Exos (bone marrow–derived mesenchymal stem cell–derived exosomes) enriched with cholesterol-modified miR-let-7c significantly attenuated renal fibrosis in the unilateral ureteral obstruction (UUO) model by reducing TGF-β1, collagen type I, and fibronectin expression [65,66].
Electrostatic approaches leverage the negatively charged exosomal surface, which is enriched in phosphatidylserine and phosphate groups. Cationic polymers such as polyethylenimine (PEI), poly(amidoamine) dendrimers (PAMAM), and chitosan bind reversibly to vesicles, improving colloidal stability and increasing RNA loading efficiency. Tang et al. [67] developed positively charged KIM-1-targeted EVs that deliver siRNAs targeting P65 and Snail1, achieving high proximal tubular uptake and antifibrotic activity in AKI models. This RNAi-based approach provides Tier 1 (causal) evidence, as the therapeutic phenotype is linked to delivery of defined siRNA cargo targeting RelA/p65 and Snai1 in a pathway-specific manner.
Amphipathic and membrane-penetrating peptides such as the trans-activator of transcription peptide (TAT), penetratin, rabies virus glycoprotein peptide (RVG), and other cell-penetrating peptides (CPPs) adsorb spontaneously onto exosomal membranes, enhancing internalization and tissue tropism [68]. Kidney-specific targeting has been achieved using ligand-targeting peptides such as LTH, which bind to KIM-1, and hyaluronic acid, which promotes CD44-mediated uptake during inflammation. RGD peptides have also been used to target integrins upregulated in fibrotic kidney tissue [69].
Aptamers represent another class of highly specific ligands that bind exosomes via hydrophobic and electrostatic interactions. Luo et al. [70] demonstrated aptamer-functionalized BM-MSC-Exos for targeted bone regeneration, a strategy that holds feasibility for renal targeting. Anti-LRP2 (megalin) aptamers are particularly promising due to the high expression and active endocytosis of LRP2 in proximal tubular cells [71]
Phosphatidylserine (PS), exposed on the exosomal surface, enables non-covalent anchoring of lactadherin C1C2 domains, annexins, and anti-PS nanobodies [65]. PS also plays a central role in renal biodistribution. KIM-1, acting as a PS receptor, confers to injured tubular epithelial cells, and this mechanism, first described by Ichimura et al. [62], helps to explain the natural renal tropism of EVs in AKI.
Modified lipids, such as DSPE-PEG or DOPE-PEG, conjugated to peptides, aptamers, or fluorophores, can spontaneously insert into the exosomal membrane. This enhances vesicle stability, prolongs circulation time, and supports targeted delivery. PEGylation also reduces opsonization and early clearance, improving pharmacokinetics, an effect widely documented in both extracellular vesicle and nanoparticle engineering [72].
However, extensive PEGylation or heavy surface decoration may partially mask native exosomal ligands, including tetraspanins, integrins, and phosphatidylserine, potentially attenuating intrinsic renal tropism and modifying biodistribution profiles [73]. This phenomenon has been reported across multiple exosomes platforms and must be considered when designing kidney-targeted formulations [73].
Safety considerations are critical for all non-covalent strategies. Cationic polymers may induce dose-dependent cytotoxicity, and exogenous peptides or ligands may elicit immunogenic responses [74]. Therefore, rigorous optimization, purification, and toxicological validation are essential prerequisites for successful clinical translation.
Covalent Surface Engineering: Click Chemistry
Covalent surface engineering via click chemistry has emerged as one of the most precise and bioorthogonal strategies for modifying BM-MSC-Exos, enabling functionalization under mild, selective conditions that preserve membrane integrity [75].
Click reactions offer high specificity, rapid kinetics, and minimal toxic byproducts, allowing stable conjugation of targeting ligands onto exosomal surfaces for applications in delivery, imaging, and theranostics. Widely used platforms include copper-catalyzed azide–alkyne cycloaddition (CuAAC) and copper-free chemistries such as dibenzocyclooctyne (DBCO)–azide and tetrazine–trans-cyclooctene (TCO) reactions, which provide superior compatibility with physiological environments [75]. These systems enable covalent attachment of peptides, antibodies, fluorophores, chemokines, and diagnostic probes, expanding the functionality of BM-MSC-Exos for kidney-targeted applications.
The renal relevance of click-based engineering has been demonstrated by Li et al. [76], who functionalized exosomes with the LTH peptide, a high-affinity ligand for KIM-1. Using DBCO–azide chemistry, LTH-modified vesicles accumulated selectively within damaged renal epithelia and attenuated inflammation and fibrosis in models of tubular injury. Because BM-MSC-Exos already possess intrinsic renal tropism driven by phosphatidylserine–KIM-1 interactions and anti-inflammatory cargo, click chemistry acts synergistically by enhancing targeting precision.
Beyond enhancing renal homing, covalent modification can also be exploited to redirect biodistribution. exosomes, peptides, or chemokine nanocarriers engineered via click reactions have demonstrated preferential accumulation in ischemic neural tissue while reducing renal uptake, illustrating the bidirectional utility of click chemistry to either promote or limit kidney tropism depending on the therapeutic objective [77]. This adaptability is particularly useful when renal retention is beneficial, as in AKI and CKD, or undesirable during systemic therapies.
Click chemistry has also been integrated into hybrid platforms that anchor BM-MSC-Exos to biomaterials, such as hydrogels, collagen matrices, or implantable scaffolds, via CuAAC or thiol–ene reactions [78,79]. These systems enhance local retention, prolong vesicle availability within injured tissue, and allow sustained release of therapeutic RNAs, proteins, or small molecules. Such strategies are especially promising in chronic kidney injury, renal regeneration, and transplant immunomodulation, where spatially confined delivery is critical.
Additionally, click-based fluorophore conjugation, such as with Alexa Fluor or Cy5.5, enables high-resolution in vivo imaging, facilitating real-time monitoring of renal accumulation, biodistribution, pharmacokinetics, and therapeutic windows [80]. These tools are essential for optimizing clinical dosing and evaluating the effectiveness of engineered targeting strategies.
Despite their versatility, covalent approaches carry notable limitations. CuAAC requires careful purification to remove residual copper ions, which can induce oxidative damage or compromise membrane integrity [74,81,82]. Excessive surface decoration with ligands may also obscure native exosomal membrane proteins, including tetraspanins, integrins, and phosphatidylserine, potentially impairing intrinsic renal tropism or modifying uptake mechanisms [81,82]. Moreover, peptide- or chemokine-functionalized vesicles may pose immunogenicity risks, and scaling click-modified formulations under GMP standards remains technically and financially challenging [81,82].
In summary, click chemistry provides a powerful, modular platform for covalent engineering of BM-MSC-Exos. It enables enhanced renal targeting, tunable biodistribution, improved therapeutic efficacy, advanced imaging applications, and integration with biomaterials. Click-engineered BM-MSC-Exos thus represent a foundational tool for next-generation precision nanotherapies and theranostic strategies in renal medicine.
Protein-Based Membrane Engineering
Protein-based membrane engineering of BM-MSC-Exos leverages intrinsic transmembrane proteins such as CD63, CD81, CD9, and especially lysosome-associated membrane glycoprotein 2b (Lamp2b) as structural scaffolds for displaying targeting peptides, single-chain antibodies (scFvs), therapeutic proteins, or imaging reporters [13,84,85]. Because these proteins are naturally enriched on exosomal membranes and present outward-facing domains, they enable selective redirection of BM-MSC-Exos toward specific renal cell populations, improved tissue retention, and enhanced modulation of pathophysiological pathways relevant to kidney injury [13,84,85].
Lamp2b-based gene fusion strategies have advanced rapidly, as Lamp2b offers high exosomal abundance and favorable extracellular topology. Fusion constructs such as Lamp2b–internalizing Arginine-Glycine-Aspartic acid peptide (iRGD), designed ankyrin repeat protein (DARPin), and scFvs have improved targeting to ischemic and inflamed tissues in multiple disease models [85]. Translational extensions to nephrology include Lamp2b fused to albumin-binding peptides (ABPs), which prolong systemic circulation and enhance renal accumulation of therapeutic vesicles carrying curcumin or hepatocyte growth factor (HGF) [86].
Protein engineering also enables targeted delivery of active therapeutic payloads. A notable example is the exosomal delivery of the super-repressor of IκB alpha (srIκBα), engineered by Kim et al. [87], which attenuated NF-κB–driven inflammation, tubular apoptosis, and renal dysfunction following IRI injury. Although this strategy involves cargo engineering, its success depends on membrane protein design for vesicle stabilization and immune cell targeting, demonstrating the interdependence between membrane and cargo engineering.
In addition, membrane proteins serve as anchors for aptamer-based engineering. Han and colleagues [88] created the Exosomal Anchor Aptamer (EAA), which shows wide-ranging binding to transmembrane proteins and enables robust loading of nucleic acid therapeutics, including PMOs and chemically stabilized oligonucleotides, onto EVs surfaces. Although tested outside nephrology, this approach is directly applicable to BM-MSC-Exos for targeted RNA delivery to injured renal epithelia.
Membrane-associated domains that recognize phosphatidylserine (PS), such as lactadherin C1C2 or anti-PS nanobodies, have also been used to functionalize EVs surfaces [89]. PS exposure is markedly increased on injured tubular epithelial cells, and KIM-1 is a PS-binding phagocytic receptor [62,90]. Thus, PS-interacting engineered exosomes can exploit this injury-specific uptake route.
Exosomal tetraspanins, particularly CD63 and CD81, also constitute robust scaffold proteins for luminescent or fluorescent reporters, as demonstrated by Verweij et al. [91] and Kowal et al. [92]. These tools allow high-resolution tracking of BM-MSC-Exos in vivo, enabling quantitative analysis of renal biodistribution, tubular uptake, and immune cell interaction, parameters essential for optimizing therapeutic design [93].
Despite their promise, protein-based engineering strategies face notable challenges. The use of viral or plasmid vectors introduces manufacturing complexity and potential immunogenicity; engineered fusion proteins may undergo lysosomal degradation after endocytosis; and inefficient endosomal escape remains a significant bottleneck for cytosolic delivery of therapeutic cargo [94]. Nevertheless, protein-based membrane engineering remains one of the most potent methods for conferring precise renal tropism and enhanced therapeutic potency to BM-MSC-Exos.
3.2.2. Cargo EngineeringEndogenous Engineering (Genetic /Cellular Reprogramming)
Endogenous cargo engineering modifies BM-MSC-Exos, either genetically or epigenetically, so that they secrete exosomes enriched with defined therapeutic RNAs, proteins, or epigenetic regulators. Because BM-MSC naturally respond to inflammatory and metabolic cues and load regulatory molecules into their vesicles, this strategy leverages intrinsic sorting pathways to generate exosomes with predictable biologically coherent cargo profiles. Foundational work by Bruno et al. [20] demonstrated that MSC-derived vesicles naturally contain mRNAs that promote tubular regeneration, epithelial repair, and immunomodulation, enabling renal functional recovery in murine AKI models.
Building on this foundation, endogenous reprogramming has been used to enhance mitochondrial resilience, a central determinant of susceptibility to AKI and CKD progression. Zhao et al. [95] engineered MSC to overexpress mitochondrial transcription factor A (TFAM), resulting in exosomes enriched with intact mtDNA fragments. These vesicles attenuated mitochondrial injury, preserved tubular bioenergetics, and reduced systemic inflammation under oxidative stress, illustrating coordinated co-delivery of mRNA and mtDNA for mitochondrial network repair. TFAM-driven, mtDNA-enriched exosomes provide Tier 2 evidence for a causal mitochondrial-protective effect, although the relative contribution of mtDNA versus co-loaded RNAs/proteins warrants further mechanistic dissection.
Endogenous manipulation of miRNAs is another powerful strategy for reshaping the epigenetic landscape of renal disease. Grange et al. [96] enriched BM-MSC with antifibrotic miRNAs, producing exosomes capable of halting and reversing fibrosis in diabetic nephropathy by restoring glomerular and tubular structure.
Endogenous engineering also supports the controlled expression of siRNAs for pathway-specific silencing. The seminal discovery by Valadi et al. [97] that exosomes mediate the horizontal transfer of mRNAs and miRNAs provides the biological rationale for this approach.
Cargo loading is further refined by endogenous sorting machinery [98,99,100]. RNA incorporation into exosomes is actively regulated by proteins such as hnRNPA2B1 (recognizing the EXOmotif “GGAG”), YBX1 (critical for packaging miR-223 and other regulatory miRNAs), and ALIX (involved in both miRNA sorting and ILV biogenesis). Modulating these molecules enhances the selectivity and reproducibility of vesicle cargo composition.
Within this mechanistic framework, Tang et al. [67] reprogrammed MSC to express siRNAs targeting RelA/p65 and Snai1, producing vesicles that naturally packaged these RNAs and, when directed to KIM-1–positive tubules, delivered robust anti-inflammatory and antifibrotic effects in AKI.
CRISPR/Cas9 technology has further expanded endogenous engineering. CRISPR can be loaded into BM-MSC-Exos as a low-immunogenic gene-editing modality for renal cells or used directly to edit the parental MSC, enabling stable overexpression of protective genes, repression of pathogenic pathways, or generation of modified regulatory RNAs incorporated into vesicles. Dashti et al. [101] highlighted that CRISPR-edited MSC serve as renewable and standardized sources of therapeutic exosomes with reduced batch variability.
CRISPR also enables the creation of immortalized BM-MSC lines for large-scale and GMP-compatible vesicle production. Hu et al. [102] inserted SV40 large T antigen into the Rosa26 locus via HDR, generating reversibly immortalized MSC that maintain multipotency and secrete stable EVs populations.
Finally, CRISPR-edited kidney organoids have emerged as physiologically relevant platforms for evaluating the uptake, molecular actions, and functional rescue mediated by engineered BM-MSC-Exos [103]. Organoid models replicating polycystic kidney disease (PKD1/PKD2), congenital nephrotic syndrome (NPHS1), and podocyte structural disorders (PODXL) accelerate mechanistic discovery and translational optimization of miRNA-enriched or gene-edited exosomes [103,104,105,106].
Overall, endogenous cargo engineering constitutes a biologically coherent and programmable strategy to enhance the therapeutic profile of BM-MSC-Exos. By reprogramming producer cells to enrich defined RNAs and regulatory proteins, and by leveraging endogenous RNA-sorting machinery, this approach can increase cargo selectivity and improve the consistency of exosome-mediated bioactivity across renal contexts [95,96,97,98,99,100]. When integrated with human-relevant validation systems, including CRISPR-edited kidney organoids, endogenous engineering supports more mechanistically grounded evaluation of uptake, pathway engagement, and functional rescue for kidney disease applications [103,104,105,106].
Although BM-MSC-Exos are acellular, the use of CRISPR-edited or immortalized parental MSCs introduces additional regulatory and biosafety requirements because the manufacturing platform relies on a genetically modified cell source [101,102]. Practical barriers include demonstrating identity, genomic integrity, and stability of the producer cell bank across passages, including edit specificity, off-target assessment, and karyotypic stability, as well as controlling vector- and transgene-associated risks and excluding process-related impurities in the final EVs preparation [101]. From a product-release perspective, programs using genetically modified producer cells generally require fit-for-purpose controls to support sterility, mycoplasma and endotoxin safety, residual host-cell DNA and transgene-fragment limits, and batch comparability aligned with the proposed mechanism of action and clinical context [101]. These considerations are particularly salient for immortalization strategies that use oncogenic elements such as SV40 large T, which can trigger heightened safety scrutiny even when the final therapeutic modality is cell-free [102].
Exogenous Cargo Loading
Exogenous cargo loading enhances the therapeutic versatility of exosomes by enabling the post-secretory incorporation of defined bioactive molecules. In contrast to endogenous loading, which is constrained by cellular machinery and yields variable cargo content, exogenous methods offer precise control over payload composition and dosing. This is a critical advantage for regenerative and anti-inflammatory applications, including renal injury models in which modulation of mitochondrial, inflammatory, or fibrotic pathways has demonstrated therapeutic benefit [84].
These techniques fall broadly into two categories (Figure 4): passive loading, driven by spontaneous physicochemical interactions with the exosomal membrane, and active loading, which employs transient membrane permeabilization to improve cargo entry [84].
Although numerous exogenous loading strategies have been applied across the extracellular vesicle field, only a subset has been experimentally validated in MSC-Exos, particularly in renal injury. Among the most compelling examples, miR-30-loaded MSC-Exos, introduced via physical permeabilization, have demonstrated potent nephroprotective effects. In ischemia-reperfusion injury (IRI), these engineered vesicles inhibited the dynamin-related protein 1 (Drp1)–mitochondrial fission one protein (Fis1) axis, thereby preserving mitochondrial homeostasis, reducing tubular apoptosis and inflammation, and ultimately improving renal function [107].
Although direct examples of exogenous loading in BM-MSC-Exos for renal injury remain limited, the preferential renal accumulation of MSC-Exos as a broader class provides strong biological justification for their use as therapeutic carriers. Following tubular injury, epithelial cells externalize phosphatidylserine and upregulate receptors such as KIM-1/TIM-1, ICAM-1, and multiple integrins, all of which naturally recognize phospholipid domains and tetraspanin-enriched surfaces characteristic of exosomes derived from diverse MSC-sources [20,62,86,96]. These conserved features shared across BM-MSC-, UC-MSC-, and WJ-MSC-derived exosomes facilitate efficient retention and internalization within damaged tubules, conferring an inherent targeting advantage over synthetic nanocarriers such as liposomes, dendrimers, and chitosan-based systems [20,62,86,96,107].
Furthermore, as comprehensively discussed by Huang et al. [108], MSC-Exos surpass synthetic platforms in biocompatibility, immune evasion, microenvironmental stability, and preferential accumulation in injured renal tissue. Thus, even in the absence of extensive renal studies specifically employing exogenously loaded BM-MSC-Exos, the well-established mechanistic basis of MSC-Exo renal tropism strongly supports their application as optimized delivery vehicles for kidney regeneration.
Nonetheless, several translational barriers persist. Physical loading can destabilize the exosomal membrane, alter the zeta potential, or diminish the surface expression of tetraspanins and integrins, which are essential for renal targeting, as highlighted by Batrakova and Kim [109]. Electroporation-based approaches, while widely used, frequently induce RNA aggregation and generate size-dependent loading artifacts unless rigorously optimized, a limitation clearly demonstrated by Lamichhane et al. [110]. Chemical permeabilization methods introduce additional challenges, including the need to remove residual reagents to stringent GMP standards.
Sonication, extrusion, and related mechanical strategies increase loading efficiency but often yield heterogeneous vesicle populations or disrupt membrane-associated proteins, as observed by Haney et al. [111]. Hybrid engineering strategies, such as liposome–exosome fusion, partially address these issues but may alter vesicle identity, as shown by Sato et al. [112]. Together, these limitations underscore the need for next-generation hybrid and biomimetic systems that integrate BM-MSC-Exos with synthetic nanomaterials to enhance stability, target fidelity, and scalability for renal regenerative applications.
Hybrid Systems, Biomimetic and Artificial EVs Systems (EVs-Mimetics)
Hybrid systems represent an emerging strategy to overcome the intrinsic limitations of natural exosomes, including limited loading capacity, limited structural stability, and batch heterogeneity, while preserving their biological advantages, such as low immunogenicity, compatibility with inflammatory microenvironments, and the well-characterized renal tropism of BM-MSC-Exos [13].
These platforms combine exosomal membranes with liposomes, erythrocyte membranes, or biocompatible polymers through controlled membrane fusion or surface assembly, generating modular nanosystems capable of transporting therapeutic RNAs, proteins, small molecules, or CRISPR-associated components with improved stability and pharmacokinetics [13].
Among these approaches, exosome-liposome hybrids (Exo-Liposome systems) are the most extensively studied. Fusion achieved through incubation, freeze-thaw cycles, sonication, or extrusion produces vesicles with enlarged internal volume and enhanced encapsulation of hydrophilic agents, large nucleic acids, or genome-editing tools. In foundational work, Sato et al. [112] demonstrated efficient membrane fusion, highlighting the importance of lipid composition for vesicle reproducibility and structural stability.
Natural phospholipids offer biomimetic properties but introduce variability, whereas synthetic glycerophosphocholine-based lipids exhibit superior consistency and storage stability, as described by van Hoogevest and Wendel [113]. Building on this, Lin et al. [114] successfully delivered Cas9 and sgRNA into MSC using Exo-Liposome hybrids, achieving high editing efficiency with lower immunogenicity than viral vectors, an approach with direct translational implications for renal gene correction strategies targeting monogenic kidney disorders or profibrotic signaling pathways.
A second modality involves erythrocyte membrane-integrated systems (Exo-RBC hybrids). These constructs exploit the natural stealth properties of red blood cell membranes, thereby extending circulation time and reducing clearance by the mononuclear phagocyte system.
Critically, Tang et al. [67] demonstrated that RBC-mimetic vesicles functionalized with the KIM-1-binding peptide LTH selectively targeted injured proximal tubules in ischemic and obstructive AKI. These hybrids efficiently delivered siRNAs targeting RelA/p65 and Snail-1, resulting in significant reductions in inflammation, apoptosis, and fibrosis. Although the system employed EVs and RBC membranes rather than BM-MSC-Exos specifically, the study provides compelling proof of concept for kidney-targeted hybrid nanocarriers.
A third category includes polymer-integrated hybrid systems (Exo-Polymer platforms), in which BM-MSC-Exos or MSC-Exos are coated or embedded within PEG, PLGA, chitosan, PAMAM dendrimers, or hydrogel matrices. These materials improve vesicle mechanical stability, prolong renal retention, and enable ligand functionalization with RGD peptides or hyaluronic acid, which bind to receptors upregulated during tubular injury.
As summarized by Li et al. [115], hydrogels incorporating exosomes enhance the delivery of therapeutic miRNAs, such as let-7a-5p, thereby supporting autophagy regulation and reducing tubular apoptosis after IRI. Although most studies have not specifically employed BM-MSC-Exos in polymer-integrated systems for kidney disease, the mechanistic rationale and preclinical success in other tissues strongly support their future application to renal regenerative therapeutics.
Despite the current absence of studies directly integrating BM-MSC-Exos into hybrid or mimetic platforms for renal applications, the intrinsic renal tropism, cargo complexity, and immunomodulatory potential of these vesicles suggest a compelling opportunity for future translational research. The synergy between biological fidelity and engineering precision could yield next generation nanotherapeutics for AKI and CKD.
While hybrid systems retain biologically secreted exosomal components, biomimetic and artificial extracellular vesicle systems (collectively termed EV mimetics) are engineered de novo through physical-mechanical or synthetic approaches. These systems are designed to mimic the structural features, surface markers, and functional behavior of natural exosomes, without relying on cellular secretion pathways [13]. EVs-mimetics offer key advantages for scalable nanotherapeutic development, including uniform vesicle size, tunable cargo composition, and enhanced batch-to-batch reproducibility [13].
They are commonly fabricated via extrusion, sonication, membrane fragmentation, or chemical assembly using lipid bilayers, cell-derived membranes (e.g., RBC, MSC), or polymeric scaffolds such as PLGA or PEG. Due to their customizable architecture and surface chemistry, EVs-mimetics are increasingly explored for kidney-targeted therapies, particularly where precise dosing, stability, and regulatory standardization are critical [13].
As noted by Li et al. [115], the choice of membrane source, such as erythrocytes, platelets, leukocytes, or MSC, strongly influences biodistribution, immune compatibility, and interactions with inflamed renal tissue. MSC- or epithelial-derived membranes are particularly relevant for kidney disease because they present integrins and CD44, which bind to receptors upregulated during tubular injury [119].
Beyond cell-derived mimetics, polymer-based artificial vesicles, such as PLGA, PEG–PLGA, chitosan, or low-generation PAMAM dendrimers, can be coated with lipid bilayers to emulate exosomal curvature and surface charge [113,115]. These artificial vesicles exhibit higher loading efficiency for hydrophilic drugs, nucleic acids, and chemically fragile therapeutics, while synthetic lipid components eliminate co-isolated contaminants and avoid the immunological variability associated with biologically sourced materials.
Despite the advantages of these platforms, as emphasized by Lener et al. [116] and Reiner et al. [18], regulatory classification becomes more complex when biological membranes are combined with synthetic materials, complicating GMP standardization. Recent analyses, such as those by Zhu et al. [117], further underscore the need for careful evaluation of biomimetic carriers to ensure long-term safety and functional fidelity.
3.3. Safety and Regulatory Considerations
Collectively, the engineering strategies discussed above reinforce the central role of safety and regulatory risk assessment in the clinical translation of extracellular vesicle-based therapeutics, including BM-MSC-Exos [94,116]. Surface functionalization approaches, including peptides, antibodies, polymers, PEGylation, and covalent chemistries, can alter immune recognition and immunogenicity risk, and can also shift biodistribution, potentially increasing off-target accumulation, particularly in organs involved in particle clearance [65,74,94]. Because biodistribution and delivery efficiency are formulation- and route-dependent, fit-for-purpose in vivo biodistribution and safety assessments remain important, ideally supported by robust tracking strategies and model-matched dosing schedules [73,91,93,94].
Covalent conjugation workflows, including click-chemistry-based strategies, introduce additional process-related risks that require explicit control, as residual catalysts and unreacted reagents can compromise vesicle integrity and oxidative stability if not adequately removed [75,80]. Scale-up of modified EVs formulations under GMP, therefore, requires validated purification and in-process controls, together with standardized analytical characterization and release specifications aligned with community guidance [2,17,18,56,116]. In parallel, long-term clearance and retention profiles remain incompletely defined for many engineered EVs formats, particularly when modifications are intended to prolong circulation time or increase tissue retention, reinforcing the need for stability testing and batch comparability criteria suitable for the intended clinical use [18,94].
Hybrid nanoparticle-exosome systems introduce further toxicology variables that depend on material composition, residual reagents, and potential membrane perturbation during fabrication, and these constructions may require additional attention to material-linked toxicity and the reproducibility of hybrid assembly [24,72,112]. Accordingly, release testing should address identity, purity, and potency, together with sterility, mycoplasma, and endotoxin controls, as well as residual process-related impurities, under GMP-aligned workflows [2,17,56].
3.4. Tracking and Imaging of Engineered Exosomes
BM-MSC-Exos possess intrinsic therapeutic potential but can be transformed into fully theranostic nanosystems when engineered with imaging agents such as fluorophores, gadolinium chelates, gold nanoparticles, luciferase reporters, or radioisotopes. These modifications enable non-invasive, real-time tracking of biodistribution, renal targeting, and therapeutic engagement, which are critical for validating treatment mechanisms in both AKI and CKD. It is essential, however, that labeling strategies preserve vesicle integrity, the natural tropism toward injured proximal tubules, and biological activity [13,67].
Physical, chemical, and genetic labeling methods enable multimodal visualization via fluorescence, bioluminescence, magnetic resonance imaging (MRI), computed tomography (CT), and nuclear imaging (SPECT/PET). In models of kidney injury, such tracking techniques are essential to confirm selective accumulation of BM-MSC-Exos within the proximal tubular microenvironment, a key correlate of therapeutic efficacy [13].
Gold nanoparticles enable CT imaging with excellent contrast. For example, Betzer et al. [118] demonstrated that labeled exosomes retained selective tropism and supported in vivo deep-tissue visualization. Lipophilic tracers such as PKH26, DiI, and DiR remain popular tools, mainly owing to their straightforward application and high signal intensity; however, dye aggregation and residual carry-over can produce false-positive signals, requiring careful experimental controls [118]. Despite this, DiR-labeled MSC-EVs have demonstrated preferential accumulation in injured renal tubules in AKI models, confirming their natural renal targeting ability [93].
Bioluminescent reporters offer high sensitivity with minimal background noise. Lai et al. [119] showed that EVs tagged with Gaussia luciferase enabled precise kinetic tracking of vesicle trafficking and clearance. Radioisotope labeling with agents such as ⁹⁹ᵐTc, ¹¹¹In, ¹²⁴I, or ⁶⁴Cu provides absolute quantification and is among the most clinically translatable imaging modalities.
Integrating biodistribution readouts with functional outcomes supports the rationale for imaging-enabled BM-MSC-Exos as nanotheranostic platforms in kidney disease. Nonetheless, clinical adoption of exosome imaging remains limited, and additional validation in large-animal models and early-phase human studies is needed. Labeling protocols also require careful optimization to preserve vesicle integrity and bioactivity and to minimize artifacts arising from free label or label transfer. Hybrid and biomimetic vesicle designs may improve compatibility with specific imaging agents while maintaining renal tropism and therapeutic performance. At present, imaging should be framed primarily as an enabling layer for mechanistic and pharmacology validation, confirming renal uptake, biodistribution, and target engagement, rather than as a mature standalone clinical diagnostic modality.
4. Future Directions
Engineered BM-MSC-Exos have rapidly progressed from experimental biologics to promising therapeutic and nanotheranostic platforms for kidney diseases. Their natural biocompatibility, low immunogenicity, and inherent tropism toward injured proximal tubules form a unique biological foundation that can be enhanced through membrane modification, cargo engineering, and hybrid or biomimetic designs.
These strategies enable targeted silencing of inflammatory and profibrotic pathways, preservation of mitochondrial function, modulation of epithelial repair, and long-term functional recovery in models of AKI and CKD. Parallel advances in imaging have transformed BM-MSC-Exos into traceable nanosystems capable of coupling therapeutic delivery with real-time biodistribution analysis, which is essential for mechanistic validation, dose refinement, and precision nephrotherapy.
Although significant strides have been made, substantial barriers remain that currently limit movement toward clinical implementation. Isolation and engineering workflows require strict standardization to ensure reproducibility, potency, and batch consistency across manufacturing platforms. Comprehensive safety evaluations must address immunogenicity, off-target accumulation, membrane alterations induced by engineering methods, and interactions with inflamed renal environments. The long-term fate, biodegradation, and metabolic clearance of engineered exosomes also remain poorly understood. Additionally, regulatory frameworks must be adapted to accommodate increasingly complex biohybrid products that combine natural vesicles with synthetic materials, genome-editing components, or targeting ligands.
Looking ahead, several emerging directions are expected to shape the future of exosome-based renal therapeutics, as shown in Figure 5.
Together, these advances position engineered BM-MSC-Exos as next-generation precision-guided biologics with transformative potential for treating acute and chronic kidney diseases. As bioengineering technologies, computational modeling, organoid platforms, and regulatory science continue to evolve, the clinical implementation of personalized, imaging-capable exosome therapeutics becomes increasingly achievable and promising.
5. Conclusions
This review integrates recent evidence on bone marrow mesenchymal stromal/stem cell–derived exosomes (BM-MSC-Exos) in acute and chronic kidney injury and highlights how engineering choices shape bioactivity, delivery, and translational feasibility. Across representative AKI and CKD models, BM-MSC-Exos are most consistently associated with improved renal outcomes through convergent effects on mitochondrial homeostasis, redox balance, apoptosis, and immune-inflammatory signaling. However, mechanistic depth varies substantially, and many proposed pathways remain supported primarily by associative readouts rather than definitive causal validation.
Accordingly, engineering should be framed as a strategy to improve specificity, reproducibility, and interpretability, rather than solely to amplify therapeutic effects. Producer-cell programming and cargo modulation can generate vesicles with more defined functional profiles and can be interrogated in human-relevant systems (e.g., organoids) to strengthen mechanistic inference. Delivery-oriented modifications and hybrid theranostic designs may enhance renal exposure and enable target-engagement readouts. Still, they also raise requirements for purification, stability, and safety, particularly regarding immunogenicity, off-target accumulation, and long-term retention. In parallel, route and exposure kinetics emerge as key determinants of efficacy and should be optimized alongside cargo and surface design.
Major translational bottlenecks include scalable GMP-compatible manufacturing, batch consistency, and harmonized dosing and potency frameworks aligned with MISEV2023. Clinically, EV biomarker development, especially in urine, appears to be progressing more rapidly than interventional renal EV therapeutics and imaging-enabled applications. In this setting, tracking and imaging are best positioned as enabling layers that strengthen translational rigor by confirming biodistribution and target engagement and by informing dose and schedule refinement, rather than as established diagnostic modalities.
Author Contributions
The following statements should be used “Conceptualization, M.B.C. and F.T.B.; methodology, M.B.C. and F.T.B.; software, M.B.C.; validation, M.B.C. and F.T.B.; formal analysis, M.B.C. and F.T.B.; investigation, M.B.C. and F.T.B.; resources, M.B.C. and F.T.B.; data curation, M.B.C. and F.T.B.; writing—original draft preparation, M.B.C.; writing—review and editing, F.T.B.; visualization, M.B.C. and F.T.B.; supervision, F.T.B.; project administration, F.T.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
During the preparation of this manuscript, the authors used Grammarly and Ginger exclusively for grammar checking and language refinement. All tool-assisted outputs were reviewed and edited by the authors, who take full responsibility for the content of this publication.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Convento, M.B.; Borges, F.T. Challenges and Opportunities in the Study of Bone Marrow-Derived Mesenchymal Stromal Cells: From Where We Stand to Where We Go. In Handbook of Regenerative Medicine: Stem Cell-based Approach; Haider, K.H., Ed.; CRC Press: Boca Raton, FL, USA, 2025; pp 181–198. [CrossRef]
- Welsh, J.A.; Goberdhan, D.C.I.; O’Driscoll, L.; Buzas, E.I.; Blenkiron, C.; Bussolati, B.; Cai, H.; Di Vizio, D.; Driedonks, T.A.P.; Erdbrügger, U.; et al. Minimal Information for Studies of Extracellular Vesicles (MISEV2023): From Basic to Advanced Approaches. J. Extracell. Vesicles 2024, 13, e12404. [CrossRef]
- Maguire, G. Stem Cell Therapy without the Cells. Commun. Integr. Biol. 2013, 6, e26631. [CrossRef]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding Light on the Cell Biology of Extracellular Vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [CrossRef]
- Kowal, J.; Tkach, M.; Théry, C. Biogenesis and Secretion of Exosomes. Curr. Opin. Cell Biol. 2014, 29, 116–125. [CrossRef]
- Bianco, P.; Cao, X.; Frenette, P.S.; Mao, J.J.; Robey, P.G.; Simmons, P.J.; Wang, C.Y. The Meaning, the Sense and the Significance: Translating the Science of Mesenchymal Stem Cells into Medicine. Nat. Med. 2013, 19, 35–42. [CrossRef]
- Li, D.; Li, D.; Wang, Z.; Li, J.; Shahzad, K.A.; Wang, Y.; Tan, F. Signaling Pathways Activated and Regulated by Stem Cell-Derived Exosome Therapy. Cell Biosci. 2024, 14, 105. [CrossRef]
- Lotfy, A.; AboQuella, N.M.; Wang, H. Mesenchymal Stromal/Stem Cell (MSC)-Derived Exosomes in Clinical Trials. Stem Cell Res. Ther. 2023, 14, 66. [CrossRef]
- Butkiene, G.; Daugelaite, A.M.; Poderys, V.; Marin, R.; Steponkiene, S.; Kazlauske, E.; Uzieliene, I.; Daunoravicius, D.; Jaque, D.; Rotomskis, R.; et al. Synergistic Enhancement of Photodynamic Cancer Therapy with Mesenchymal Stem Cells and Theranostic Nanoparticles. ACS Appl. Mater. Interfaces 2024, 16, 49092–49103. [CrossRef]
- Pesce, F.; Vadalà, M.; Almeida, E.; Fernandez, B.; Fouque, D.; Malyszko, J.; Schmidt-Ott, K.; Stenvinkel, P.; Wheeler, D.C.; Seidu, S.; et al. International Nephrology Masterclass in Chronic Kidney Disease: Rationale, Summary, and Future Perspectives. Life 2024, 14, 1668. [CrossRef]
- Delrue, C.; De Bruyne, S.; Speeckaert, R.; Speeckaert, M.M. Urinary Extracellular Vesicles in Chronic Kidney Disease: From Bench to Bedside? Diagnostics 2023, 13, 443. [CrossRef]
- Himanshu; Gunjan; Pandey, R.P.; Mukherjee, R.; Chang, C.M. Meta-Analysis Study of the Therapeutic Impact of Mesenchymal Stem Cell–Derived Exosomes for Chronic Kidney Diseases. Biochem. Biophys. Rep. 2025, 43, 102072. [CrossRef]
- Bahadorani, M.; Nasiri, M.; Dellinger, K.; Aravamudhan, S.; Zadegan, R. Engineering Exosomes for Therapeutic Applications: Decoding Biogenesis, Content Modification, and Cargo Loading Strategies. Int. J. Nanomed. 2024, 19, 7137–7164. [CrossRef]
- Maldonado, V.V.; Patel, N.H.; Smith, E.E.; Barnes, C.L.; Gustafson, M.P.; Rao, R.R.; Samsonraj, R.M. Clinical Utility of Mesenchymal Stem/Stromal Cells in Regenerative Medicine and Cellular Therapy. J. Biol. Eng. 2023, 17, 44. [CrossRef]
- Ma, Z.J.; Yang, J.J.; Lu, Y.B.; Liu, Z.Y.; Wang, X.X. Mesenchymal Stem Cell-Derived Exosomes: Toward Cell-Free Therapeutic Strategies in Regenerative Medicine. World J. Stem Cells 2020, 12, 814–840. [CrossRef]
- Zhang, B.; Tian, X.; Hao, J.; Xu, G.; Zhang, W. Mesenchymal Stem Cell-Derived Extracellular Vesicles in Tissue Regeneration. Cell Transplant. 2020, 29, 963689720908500. [CrossRef]
- Lötvall, J.; Hill, A.F.; Hochberg, F.; Buzás, E.I.; Di Vizio, D.; Gardiner, C.; Gho, Y.S.; Kurochkin, I.V.; Mathivanan, S.; Quesenberry, P.; et al. Minimal Experimental Requirements for Definition of Extracellular Vesicles and Their Functions: A Position Statement from the International Society for Extracellular Vesicles. J. Extracell. Vesicles 2014, 3, 26913. [CrossRef]
- Reiner, A.T.; Witwer, K.W.; van Balkom, B.W.M.; de Beer, J.; Brodie, C.; Corteling, R.L.; Gabrielsson, S.; Gimona, M.; Ibrahim, A.G.; de Kleijn, D.; et al. Concise Review: Developing Best-Practice Models for the Therapeutic Use of Extracellular Vesicles. Stem Cells Transl. Med. 2017, 6, 1730–1773. [CrossRef]
- Barile, L.; Vassalli, G. Exosomes: Therapy Delivery Tools and Biomarkers of Diseases. Pharmacol. Ther. 2017, 174, 63–78. [CrossRef]
- Bruno, S.; Grange, C.; Deregibus, M.C.; Calogero, R.A.; Saviozzi, S.; Collino, F.; Morando, L.; Busca, A.; Falda, M.; Bussolati, B.; et al. Mesenchymal Stem Cell-Derived Microvesicles Protect against Acute Tubular Injury. J. Am. Soc. Nephrol. 2009, 20, 1053–1067. [CrossRef]
- Skotland, T.; Sandvig, K.; Llorente, A. Lipids in Exosomes: Current Knowledge and the Way Forward. Prog. Lipid Res. 2017, 66, 30–41. [CrossRef]
- Record, M.; Silvente-Poirot, S.; Poirot, M.; Wakelam, M.J.O. Extracellular Vesicles: Lipids as Key Components of Their Biogenesis and Functions. J. Lipid Res. 2018, 59, 1316–1324. [CrossRef]
- Dalal, V.; Lata, H.; Kharkwal, G.; Jotwani, G. Regulation of Clinical Research for Cellular and Gene Therapy Products in India. Adv. Exp. Med. Biol. 2023, 1430, 135–154. [CrossRef]
- Al-Jipouri, A.; Almurisi, S.H.; Al-Japairai, K.; Bakar, L.M.; Doolaanea, A.A. Liposomes or Extracellular Vesicles: A Comprehensive Comparison of Both Lipid Bilayer Vesicles for Pulmonary Drug Delivery. Polymers (Basel) 2023, 15, 318. [CrossRef]
- Pan, L.; Garcia, S.G.; Font-Morón, M.; Sanroque-Muñoz, M.; Clos-Sansalvador, M.; Garcia, G.M.; Borràs, F.E.; Franquesa, M. Beyond Preclinical Promise: Can Mesenchymal Stromal Cell-Derived Extracellular Vesicles Reliably Target Tubular Epithelial Cells? Extracell. Vesicles Circ. Nucleic Acids 2025, 6, 580–593. [CrossRef]
- Luiz, R.D.S.; Rampaso, R.R.; Dos Santos, A.A.C.; Convento, M.B.; Barbosa, D.A.; da Fonseca, C.D.; de Oliveira, A.S.; Caires, A.; Furlan, A.; Schor, N.; et al. BM-MSC–Derived Small Extracellular Vesicles (sEV) from Trained Animals Present Nephroprotective Potential in a Unilateral Ureteral Obstruction Model. J. Venom Anim. Toxins Incl. Trop. Dis. 2021, 27, e20200187. [CrossRef]
- Ahmadian, S.; Jafari, N.; Tamadon, A.; Ghaffarzadeh, A.; Rahbarghazi, R.; Mahdipour, M. Different Storage and Freezing Protocols for Extracellular Vesicles: A Systematic Review. Stem Cell Res. Ther. 2024, 15, 453. [CrossRef]
- Li, R.; Tao, H.; Pan, K.; Li, R.; Guo, Z.; Chen, X.; Li, Z. Extracellular Vesicles Derived from Mesenchymal Stem Cells Alleviate Renal Fibrosis via the miR-99b-5p/mTOR/Autophagy Axis in Diabetic Kidney Disease. Stem Cell Res. Ther. 2025, 16, 142. [CrossRef]
- Yoo, D.; Jung, S.Y.; Go, D.; Park, J.Y.; You, D.G.; Jung, W.K.; Li, Y.; Ding, J.; Park, J.H.; Um, W. Functionalized Extracellular Vesicles of Mesenchymal Stem Cells for Regenerative Medicine. J. Nanobiotechnol. 2025, 23, 219. [CrossRef]
- Zuk, A.; Bonventre, J.V. Acute Kidney Injury. Annu. Rev. Med. 2016, 67, 293–307. [CrossRef]
- Malek, M.; Nematbakhsh, M. Renal Ischemia/Reperfusion Injury: From Pathophysiology to Treatment. J. Renal Inj. Prev. 2015, 4, 20–27. [CrossRef]
- Mulcahy, L.A.; Pink, R.C.; Carter, D.R. Routes and Mechanisms of Extracellular Vesicle Uptake. J. Extracell. Vesicles 2014, 3, 24641. [CrossRef]
- Costa Verdera, H.; Gitz-Francois, J.J.; Schiffelers, R.M.; Vader, P. Cellular Uptake of Extracellular Vesicles Is Mediated by Clathrin-Independent Endocytosis and Macropinocytosis. J. Control. Release 2017, 266, 100–108. [CrossRef]
- Ji, X.; Nie, S.; Li, X.; Liu,H.; Du, X.; Fan, L. MSC-Derived Exosomal miR-127-3p Alleviates Acute Kidney Ischemia–Reperfusion Injury via Suppressing ATG5/ATG7-Mediated Autophagy. Nephrology 2025, 30, e70054. [CrossRef]
- Taghavi, S.; Keshtkar, S.; Abedanzadeh, M.; Hashemi, M.; Heidari, R.; Abolmaali, S.S.; Dara, M.; Aghdaei, M.H.; Sabegh, A.; Azarpira, N. Exosome Loaded in Microneedle Patch Ameliorates Renal Ischemia–Reperfusion Injury in a Mouse Model. Stem Cells Int. 2025, 2025, 3106634. [CrossRef]
- Kong, D.; Yang, Z.; Zhang, X.; Song, Y.; Anayyat, U.; Li, Y.; Liu, H.; Ou, Z.; Pang, S.; Wang, X. Cisplatin-Induced Acute Kidney Injury Is Alleviated by BMSC-Derived Exosomes via mmu-miR-874-3p–Mediated Activation of the Wnt/β-Catenin Signaling Pathway. Regen. Ther. 2025, 30, 719–729. [CrossRef]
- Yang, T.; Yu, H.; Xie, Z. Curcumin-Induced Exosomal FTO from Bone Marrow Stem Cells Alleviates Sepsis-Associated Acute Kidney Injury by Modulating the m6A Methylation of OXSR1. Kaohsiung J. Med. Sci. 2025, 41, e12923. [CrossRef]
- Çam, S.B.; Çiftci, E.; Gürbüz, N.; Altun, B.; Korkusuz, P. Allogeneic Bone Marrow Mesenchymal Stem Cell-Derived Exosomes Alleviate Human Hypoxic AKI-on-a-Chip within a Tight Treatment Window. Stem Cell Res. Ther. 2024, 15, 105. [CrossRef]
- Chen, Y.; Hou, S. Targeted Treatment of Rat AKI Induced by Rhabdomyolysis Using BMSC-Derived Magnetic Exosomes and Its Mechanism. Nanoscale Adv. 2024, 6, 4180–4195. [CrossRef]
- Dong, Y.; Tong, Y. The Pathogenesis of Chronic Kidney Disease (CKD) and the Preventive and Therapeutic Effects of Natural Products. Curr. Issues Mol. Biol. 2025, 47, 853. [CrossRef]
- Kandel, R.; Roy, P.; Singh, K.P. Molecular Basis of Oxidative Stress-Induced Acute Kidney Injury, Kidney Fibrosis, Chronic Kidney Disease, and Clinical Significance of Targeting Reactive Oxygen Species-Regulated Pathways to Treat Kidney Disease. Front. Biosci. (Schol. Ed.) 2025, 17 (3), 38963. [CrossRef]
- Lv, Y.; Hua, Z.; Lu, X. Protective Effects and Possible Mechanisms of Mesenchymal Stem Cells and Mesenchymal Stem Cell-Derived Extracellular Vesicles against Kidney Fibrosis in Animal Models: A Systematic Review and Meta-Analysis. Front. Pharmacol. 2025, 15, 1511525. [CrossRef]
- Tian, J.; Wang, Z.; Tang, Z.; Zhu, H.; Sun, L. Engineered Exosomes Conferred with ROS Regulation and Immunosuppression for Ameliorating Lupus Nephritis. J. Nanobiotechnol. 2025, 23, 639. [CrossRef]
- Zhang, J.; Wang, L.; Liu, D.; Liu, Z.; Liu, Z.; Zhou, S. Bone Marrow Mesenchymal Stem Cell-Derived Exosomes Protect against β-Cell Destruction Models and Kidney Injury by Suppressing Ferroptosis. Sci. Rep. 2025, 15, 40644. [CrossRef]
- Wang, Y.; Lu, D.; Lv, S.; Liu, X.; Liu, G. Mesenchymal Stem Cell-Derived Exosomes Ameliorate Diabetic Kidney Disease through the NOD2 Signaling Pathway. Ren. Fail. 2024, 46, 2381597. [CrossRef]
- Chen, Y.; Yang, X.; Feng, M.; Yu, Y.; Hu, Y.; Jiang, W. Exosomal miR-223-3p from Bone Marrow Mesenchymal Stem Cells Targets HDAC2 to Downregulate STAT3 Phosphorylation and Alleviate HBx-Induced Ferroptosis in Podocytes. Front. Pharmacol. 2024, 15, 1327149. [CrossRef]
- Convento, M.B.; de Oliveira, A.S.; Boim, M.A.; Borges, F.T. Mesenchymal Stromal Cell Nanovesicles Carry microRNA with Nephroprotective Properties Regardless of Aging. Curr. Aging Sci. 2024, 17, 118–126. [CrossRef]
- Di Trapani, M.; Bassi, G.; Midolo, M.; Gatti, A.; Kamga, P.T.; Cassaro, A.; Carusone, R.; Adamo, A.; Krampera, M. Differential and Transferable Modulatory Effects of Mesenchymal Stromal Cell-Derived Extracellular Vesicles on T, B and NK Cell Functions. Sci. Rep. 2016, 6, 24120. [CrossRef]
- Li, J.; Ge, Z.; Ji, W.; Yuan, N.; Wang, K. The Proosteogenic and Proangiogenic Effects of Small Extracellular Vesicles Derived from Bone Marrow Mesenchymal Stem Cells Are Attenuated in Steroid-Induced Osteonecrosis of the Femoral Head. Biomed. Res. Int. 2020, 2020, 4176926. [CrossRef]
- Pisitkun, T.; Shen, R.F.; Knepper, M.A. Identification and Proteomic Profiling of Exosomes in Human Urine. Proc. Natl. Acad. Sci. U.S.A. 2004, 101, 13368–13373. [CrossRef]
- Sun, I.O.; Lerman, L.O. Urinary Extracellular Vesicles as Biomarkers of Kidney Disease: From Diagnostics to Therapeutics. Diagnostics 2020, 10, 311. [CrossRef]
- Grange, C.; Dalmasso, A.; Cortez, J.J.; Spokeviciute, B.; Bussolati, B. Exploring the Role of Urinary Extracellular Vesicles in Kidney Physiology, Aging, and Disease Progression. Am. J. Physiol. Cell Physiol. 2023, 325, C1439–C1450. [CrossRef]
- Takizawa, K.; Ueda, K.; Sekiguchi, M.; Nakano, E.; Nishimura, T.; Kajiho, Y.; Kanda, S.; Miura, K.; Hattori, M.; Hashimoto, J.; et al. Urinary Extracellular Vesicles Signature for Diagnosis of Kidney Disease. iScience 2022, 25, 105416. [CrossRef]
- Barreiro, K.; Dwivedi, O.P.; Rannikko, A.; Holthöfer, H.; Tuomi, T.; Groop, P.H.; Puhka, M. Capturing the Kidney Transcriptome by Urinary Extracellular Vesicles—From Pre-Analytical Obstacles to Biomarker Research. Genes 2023, 14, 1415. [CrossRef]
- Tutrone, R.; Donovan, M.J.; Torkler, P.; Tadigotla, V.; McLain, T.; Noerholm, M.; Skog, J.; McKiernan, J. Clinical Utility of the Exosome-Based ExoDx Prostate (IntelliScore) EPI Test in Men Presenting for Initial Biopsy with a PSA of 2–10 ng/mL. Prostate Cancer Prostatic Dis. 2020, 23, 607–614. [CrossRef]
- Chen, Y.S.; Lin, E.Y.; Chiou, T.W.; Harn, H.J. Exosomes in Clinical Trial and Their Production in Compliance with Good Manufacturing Practice. Tzu Chi Med. J. 2020, 32, 113–120. [CrossRef]
- Rezaie, J.; Feghhi, M.; Etemadi, T. A Review on Exosomes Application in Clinical Trials: Perspective, Questions, and Challenges. Cell Commun. Signal. 2022, 20, 145. [CrossRef]
- Yadav, K.; Sahu, K.K.; Sucheta; Minz, S.; Pradhan, M. Unlocking Exosome Therapeutics: The Critical Role of Pharmacokinetics in Clinical Applications. Tissue Cell 2025, 93, 102749. [CrossRef]
- Lenzini, S. Establishing a Working Range for Effective MSC–EV Dose. RoosterBio, 2021. (accessed Nov 10, 2025).
- ClinicalTrials.gov. Search Results for “Exosome OR Exosomes”. U.S. National Library of Medicine. (accessed Nov 10, 2025).
- Tan, F.; Li, X.; Wang, Z.; Li, J.; Shahzad, K.; Zheng, J. Clinical Applications of Stem Cell-Derived Exosomes. Signal Transduct. Target. Ther. 2024, 9, 17. [CrossRef]
- Ichimura, T.; Asseldonk, E.J.P.; Humphreys, B.D.; Gunaratnam, L.; Duffield, J.S.; Bonventre, J.V. Kidney Injury Molecule-1 Is a Phosphatidylserine Receptor That Confers a Phagocytic Phenotype on Epithelial Cells. J. Clin. Invest. 2008, 118, 1657–1668. [CrossRef]
- Matsushita, K.; Toyoda, T.; Akane, H.; Morikawa, T.; Ogawa, K. CD44 Expression in Renal Tubular Epithelial Cells in the Kidneys of Rats with Cyclosporine-Induced Chronic Kidney Disease. J. Toxicol. Pathol. 2024, 37, 55–67. [CrossRef]
- Ferenbach, D.A.; Bonventre, J.V. Mechanisms of Maladaptive Repair after AKI Leading to Accelerated Kidney Ageing and CKD. Nat. Rev. Nephrol. 2015, 11, 264–276. [CrossRef]
- Chao, T.; Zhao, J.; Gao, R.; Wang, H.; Guo, J.; Gao, Z.; Wang, Y. Exosome Surface Modification and Functionalization: A Narrative Review of Emerging Technologies and Their Application Potential in Precision Medicine. Adv. Technol. Neurosci. 2025, 2, 27–33. [CrossRef]
- Wang, B.; Yao, K.; Huuskes, B.M.; Shen, H.H.; Zhuang, J.; Godson, C.; Brennan, E.P.; Wilkinson-Berka, J.L.; Wise, A.F.; Ricardo, S.D. Mesenchymal Stem Cells Deliver Exogenous microRNA-let7c via Exosomes to Attenuate Renal Fibrosis. Mol. Ther. 2016, 24, 1290–1301. [CrossRef]
- Tang, T.T.; Wang, B.; Li, Z.L.; Wen, Y.; Feng, S.T.; Wu, M.; Liu, D.; Cao, J.Y.; Yin, Q.; Yin, D.; et al. Kim-1 Targeted Extracellular Vesicles: A New Therapeutic Platform for RNAi to Treat AKI. J. Am. Soc. Nephrol. 2021, 32 (10), 2467–2483. [CrossRef]
- Birtwistle, L.; Chen, X.M.; Pollock, C. Mesenchymal Stem Cell-Derived Extracellular Vesicles to the Rescue of Renal Injury. Int. J. Mol. Sci. 2021, 22 (12), 6596. [CrossRef]
- Zhou, X.; Chen, Q.; Guo, C.; Su, Y.; Guo, H.; Cao, M.; Liu, Z.; Zhang, D.; Diao, N.; Fan, H.; et al. CD44 Receptor-Targeted and Reactive Oxygen Species-Responsive H2S Donor Micelles Based on Hyaluronic Acid for the Therapy of Renal Ischemia/Reperfusion Injury. ACS Omega 2022, 7, 42339–42346. [CrossRef]
- Luo, Z.W.; Li, F.X.; Liu, Y.W.; Rao, S.S.; Yin, H.; Huang, J.; Chen, C.Y.; Hu, Y.; Zhang, Y.; Tan, Y.J.; et al. Aptamer-Functionalized Exosomes from Bone Marrow Stromal Cells Target Bone to Promote Bone Regeneration. Nanoscale 2019, 11, 20884–20892. [CrossRef]
- Gao, S.; Hein, S.; Dagnæs-Hansen, F.; Weyer, K.; Yang, C.; Nielsen, R.; Christensen, E.I.; Fenton, R.A.; Kjems, J. Megalin-Mediated Specific Uptake of Chitosan/siRNA Nanoparticles in Mouse Kidney Proximal Tubule Epithelial Cells Enables AQP1 Gene Silencing. Theranostics 2014, 4, 1039–1051. [CrossRef]
- Knudsen, K.B.; Northeved, H.; Kumar, P.E.; Permin, A.; Gjetting, T.; Andresen, T.L.; Larsen, S.; Wegener, K.M.; Lykkesfeldt, J.; Jantzen, K.; et al. In Vivo Toxicity of Cationic Micelles and Liposomes. Nanomedicine 2015, 11, 467–477. [CrossRef]
- Smyth, T.; Kullberg, M.; Malik, N.; Smith-Jones, P.; Graner, M.W.; Anchordoquy, T.J. Biodistribution and Delivery Efficiency of Unmodified Tumor-Derived Exosomes. J. Control. Release 2015, 199, 145–155. [CrossRef]
- Vader, P.; Mol, E.A.; Pasterkamp, G.; Schiffelers, R.M. Extracellular Vesicles for Drug Delivery. Adv. Drug Deliv. Rev. 2016, 106, 148–156. [CrossRef]
- Li, X.; Xiong, Y. Application of “Click” Chemistry in Biomedical Hydrogels. ACS Omega 2022, 7, 36918–36928. [CrossRef]
- Li, B.; Qi, C.; Zhang, Y.; Shi, L.; Zhang, J.; Qian, H.; Ji, C. Frontier Role of Extracellular Vesicles in Kidney Disease. J. Nanobiotechnol. 2024, 22, 583. [CrossRef]
- Ruan, H.; Li, Y.; Wang, C.; Jiang, Y.; Han, Y.; Li, Y.; Zheng, D.; Ye, J.; Chen, G.; Yang, G.Y.; et al. Click Chemistry Extracellular Vesicle/Peptide/Chemokine Nanocarriers for Treating Central Nervous System Injuries. Acta Pharm. Sin. B 2023, 13, 2202–2218. [CrossRef]
- Kim, J.; Lee, N.; Hyeon, T. Recent Development of Nanoparticles for Molecular Imaging. Philos. Trans. A Math. Phys. Eng. Sci. 2017, 375, 20170022. [CrossRef]
- Stichler, S.; Jungst, T.; Schamel, M.; Zilkowski, I.; Kuhlmann, M.; Böck, T.; Blunk, T.; Teßmar, J.; Groll, J. Thiol–Ene Clickable Poly(glycidol) Hydrogels for Biofabrication. Ann. Biomed. Eng. 2017, 45, 273–285. [CrossRef]
- Smyth, T.; Petrova, K.; Payton, N.M.; Persaud, I.; Redzic, J.S.; Graner, M.W.; Smith-Jones, P.; Anchordoquy, T.J. Surface Functionalization of Exosomes Using Click Chemistry. Bioconjugate Chem. 2014, 25, 1777–1784. [CrossRef]
- Nakase, I.; Akita, H.; Kogure, K.; Gräslund, A.; Langel, U.; Harashima, H.; Futaki, S. Efficient Intracellular Delivery of Nucleic Acid Pharmaceuticals Using Cell-Penetrating Peptides. Acc. Chem. Res. 2012, 45, 1132–1139. [CrossRef]
- Meng, W.; He, C.; Hao, Y.; Wang, L.; Li, L.; Zhu, G. Prospects and Challenges of Extracellular Vesicle-Based Drug Delivery System: Considering Cell Source. Drug Deliv. 2020, 27 (1), 585–598. [CrossRef]
- Shao, H.; Chung, J.; Balaj, L.; Charest, A.; Bigner, D.D.; Carter, B.S.; Hochberg, F.H.; Breakefield, X.O.; Weissleder, R.; Lee, H. Protein Typing of Circulating Microvesicles Allows Real-Time Monitoring of Glioblastoma Therapy. Nat. Med. 2012, 18, 1835–1840. [CrossRef]
- Mediratta, K.; Diab, M.D.; Han, P.; Hu, H.; Wang, L. Emerging Strategies for Cargo Loading and Engineering of Extracellular Vesicles for Breast Cancer Treatment. Nanomaterials 2025, 15, 1418. [CrossRef]
- Qiao, L.; Hu, J.; Qiu, X.; Wang, C.; Peng, J.; Zhang, C.; Zhang, M.; Lu, H.; Chen, W. LAMP2A, LAMP2B and LAMP2C: Similar Structures, Divergent Roles. Autophagy 2023, 19, 2837–2852. [CrossRef]
- Liu, S.; Lv, K.; Wang, Y.; Lou, P.; Zhou, P.; Wang, C.; Li, L.; Liao, G.; Zhang, Y.; Chen, Y.; et al. Improving the Circulation Time and Renal Therapeutic Potency of Extracellular Vesicles Using an Endogenous Ligand Binding Strategy. J. Control. Release 2022, 352, 1009–1023. [CrossRef]
- Kim, S.; Lee, S.A.; Yoon, H.; Kim, M.Y.; Yoo, J.K.; Ahn, S.H.; Park, C.H.; Park, J.; Nam, B.Y.; Park, J.T.; et al. Exosome-Based Delivery of Super-Repressor IκBα Ameliorates Kidney Ischemia–Reperfusion Injury. Kidney Int. 2021, 100, 570–584. [CrossRef]
- Han, G.; Zhang, Y.; Zhong, L.; Wang, B.; Qiu, S.; Song, J.; Lin, C.; Zou, F.; Wu, J.; Yu, H.; et al. Generalizable Anchor Aptamer Strategy for Loading Nucleic Acid Therapeutics on Exosomes. EMBO Mol. Med. 2024, 16, 1027–1045. [CrossRef]
- Kooijmans, S.A.A.; Gitz-François, J.J.J.M.; Schiffelers, R.M.; Vader, P. Recombinant Phosphatidylserine-Binding Nanobodies for Targeting of Extracellular Vesicles to Tumor Cells: A Plug-and-Play Approach. Nanoscale 2018, 10, 2413–2426. [CrossRef]
- Humphreys, B.D.; Xu, F.; Sabbisetti, V.; Grgic, I.; Movahedi Naini, S.; Wang, N.; Chen, G.; Xiao, S.; Patel, D.; Henderson, J.M.; et al. Chronic Epithelial Kidney Injury Molecule-1 Expression Causes Murine Kidney Fibrosis. J. Clin. Invest. 2013, 123 (9), 4023–4035. [CrossRef]
- Verweij, F.J.; Revenu, C.; Arras, G.; Dingli, F.; Loew, D.; Pegtel, D.M.; Follain, G.; Allio, G.; Goetz, J.G.; Zimmermann, P.; et al. Live Tracking of Inter-Organ Communication by Endogenous Exosomes In Vivo. Dev. Cell 2019, 48, 573–589.e4. [CrossRef]
- Kowal, J.; Arras, G.; Colombo, M.; Jouve, M.; Morath, J.P.; Primdal-Bengtson, B.; Dingli, F.; Loew, D.; Tkach, M.; Théry, C. Proteomic Comparison Defines Novel Markers to Characterize Heterogeneous Populations of Extracellular Vesicle Sub-Types. Proc. Natl. Acad. Sci. U.S.A. 2016, 113, E968–E977. [CrossRef]
- Grange, C.; Tapparo, M.; Bruno, S.; Chatterjee, D.; Quesenberry, P.J.; Tetta, C.; Camussi, G. Biodistribution of Mesenchymal Stem Cell-Derived Extracellular Vesicles in a Model of Acute Kidney Injury Monitored by Optical Imaging. Int. J. Mol. Med. 2014, 33, 1055–1063. [CrossRef]
- György, B.; Hung, M.E.; Breakefield, X.O.; Leonard, J.N. Therapeutic Applications of Extracellular Vesicles: Clinical Promise and Open Questions. Annu. Rev. Pharmacol. Toxicol. 2015, 55, 439–464. [CrossRef]
- M.; Liu, S.; Wang, C.; Wang, Y.; Wan, M.; Liu, F.; Gong, M.; Yuan, Y.; Chen, Y.; Cheng, J.; et al. Mesenchymal Stem Cell-Derived Extracellular Vesicles Attenuate Mitochondrial Damage and Inflammation by Stabilizing Mitochondrial DNA. ACS Nano 2021, 15, 1519–1538. [CrossRef]
- Grange, C.; Tritta, S.; Tapparo, M.; Cedrino, M.; Tetta, C.; Camussi, G.; Brizzi, M.F. Stem Cell-Derived Extracellular Vesicles Inhibit and Revert Fibrosis Progression in a Mouse Model of Diabetic Nephropathy. Sci. Rep. 2019, 9, 4468. [CrossRef]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-Mediated Transfer of mRNAs and microRNAs Is a Novel Mechanism of Genetic Exchange Between Cells. Nat. Cell Biol. 2007, 9, 654–659. [CrossRef]
- Villarroya-Beltri, C.; Gutiérrez-Vázquez, C.; Sánchez-Cabo, F.; Pérez-Hernández, D.; Vázquez, J.; Martin-Cofreces, N.; Martinez-Herrera, D.J.; Pascual-Montano, A.; Mittelbrunn, M.; Sánchez-Madrid, F. Sumoylated hnRNPA2B1 Controls the Sorting of miRNAs into Exosomes through Binding to Specific Motifs. Nat. Commun. 2013, 4, 2980. [CrossRef]
- Shurtleff, M.J.; Temoche-Diaz, M.M.; Karfilis, K.V.; Ri, S.; Schekman, R. Y-Box Protein 1 Is Required to Sort microRNAs into Exosomes in Cells and in a Cell-Free Reaction. eLife 2016, 5, e19276. [CrossRef]
- Géminard, C.; De Gassart, A.; Blanc, L.; Vidal, M. Degradation of AP2 during Reticulocyte Maturation Enhances Binding of hsc70 and Alix to a Common Site on TFR for Sorting into Exosomes. Traffic 2004, 5, 181–193. [CrossRef]
- Dashti, M.; Mohammaddust Sarab, M.; Shad, F.; Dehnavi, S. CRISPR-Mediated Engineering of Mesenchymal Stromal/Stem Cells: A Summary of Recent Progress in Immunological Applications for Regenerative Medicine and Cancer Therapy. Stem Cell Res. Ther. 2025, 16, 592. [CrossRef]
- Hu, X.; Li, L.; Yu, X.; Zhang, R.; Yan, S.; Zeng, Z.; Shu, Y.; Zhao, C.; Wu, X.; Lei, J.; et al. CRISPR/Cas9-Mediated Reversibly Immortalized Mouse Bone Marrow Stromal Stem Cells (BMSCs) Retain Multipotent Features of Mesenchymal Stem Cells (MSCs). Oncotarget 2017, 8, 111847–111865. [CrossRef]
- D’Ambrosio, V.; Huimei, C.; Vo, N.; Siew, K.; Evans, R.D.R.; Freedman, B.; Pesce, F. CRISPR and Gene Editing for Kidney Diseases: Where Are We? Clin. Kidney J. 2025, 18, sfaf246. [CrossRef]
- Freedman, B.S.; Brooks, C.R.; Lam, A.Q.; Fu, H.; Morizane, R.; Agrawal, V.; Saad, A.F.; Li, M.K.; Hughes, M.R.; van Werff, R.; et al. Modelling Kidney Disease with CRISPR-Mutant Kidney Organoids Derived from Human Pluripotent Epiblast Spheroids. Nat. Commun. 2015, 6, 8715. [CrossRef]
- Vishy, C.E.; Thomas, C.; Vincent, T.; Crawford, D.K.; Goddeeris, M.M.; Freedman, B.S. Genetics of Cystogenesis in Base-Edited Human Organoids Reveal Therapeutic Strategies for Polycystic Kidney Disease. Cell Stem Cell 2024, 31, 537–553.e5. [CrossRef]
- Kim, Y.K.; Refaeli, I.; Brooks, C.R.; Jing, P.; Gulieva, R.E.; Hughes, M.R.; Cruz, N.M.; Liu, Y.; Churchill, A.J.; Wang, Y.; et al. Gene-Edited Human Kidney Organoids Reveal Mechanisms of Disease in Podocyte Development. Stem Cells 2017, 35, 2366–2378. [CrossRef]
- Gu, D.; Zou, X.; Ju, G.; Zhang, G.; Bao, E.; Zhu, Y. Mesenchymal Stromal Cells–Derived Extracellular Vesicles Ameliorate Acute Renal IRI by Inhibition of Mitochondrial Fission through miR-30. Stem Cells Int. 2016, 2016, 2093940. [CrossRef]
- Huang, L.F.; Ye, Q.R.; Chen, X.C.; Huang, X.R.; Zhang, Q.F.; Wu, C.Y.; Liu, H.F.; Yang, C. Research Progress of Drug Delivery Systems Targeting the Kidneys. Pharmaceuticals 2024, 17, 625. [CrossRef]
- Batrakova, E.V.; Kim, M.S. Using Exosomes, Naturally Equipped Nanocarriers, for Drug Delivery. J. Control. Release 2015, 219, 396–405. [CrossRef]
- Lamichhane, T.N.; Raiker, R.S.; Jay, S.M. Exogenous DNA Loading into Extracellular Vesicles via Electroporation Is Size-Dependent and Enables Limited Gene Delivery. Mol. Pharm. 2015, 12, 3650–3657. [CrossRef]
- Haney, M.J.; Klyachko, N.L.; Zhao, Y.; Gupta, R.; Plotnikova, E.G.; He, Z.; Patel, T.; Piroyan, A.; Sokolsky, M.; Kabanov, A.V.; et al. Exosomes as Drug Delivery Vehicles for Parkinson’s Disease Therapy. J. Control. Release 2015, 207, 18–30. [CrossRef]
- Sato, Y.; Umezaki, K.; Sawada, S.; Sasaki, Y.; Harada, N.; Shiku, H.; Akiyoshi, K. Engineering Hybrid Exosomes by Membrane Fusion with Liposomes. Sci. Rep. 2016, 6, 21933. [CrossRef]
- van Hoogevest, P.; Wendel, A. The Use of Natural and Synthetic Phospholipids as Pharmaceutical Excipients. Eur. J. Lipid Sci. Technol. 2014, 116, 1088–1107. [CrossRef]
- Lin, Y.; Wu, J.; Gu, W.; Huang, Y.; Tong, Z.; Huang, L.; Tan, J. Exosome–Liposome Hybrid Nanoparticles Deliver CRISPR/Cas9 System in MSCs. Adv. Sci. 2018, 5, 1700611. [CrossRef]
- Li, Y.J.; Wu, J.Y.; Liu, J.; Xu, W.; Qiu, X.; Huang, S.; Hu, X.B.; Xiang, D.X. Artificial Exosomes for Translational Nanomedicine. J. Nanobiotechnol. 2021, 19, 242. [CrossRef]
- Lener, T.; Gimona, M.; Aigner, L.; Börger, V.; Buzas, E.; Camussi, G.; Chaput, N.; Chatterjee, D.; Court, F.A.; Del Portillo, H.A.; et al. Applying Extracellular Vesicles Based Therapeutics in Clinical Trials—An ISEV Position Paper. J. Extracell. Vesicles 2015, 4, 30087. [CrossRef]
- Zhu, K.; Huang, Y.; Li, K.; Zhang, K. Biomimetic Erythrocyte-Based Drug Delivery Systems for Organ-Targeted Therapy. Front. Bioeng. Biotechnol. 2025, 13, 1663092. [CrossRef]
- Betzer, O.; Perets, N.; Angel, A.; Motiei, M.; Sadan, T.; Yadid, G.; Offen, D.; Popovtzer, R. In Vivo Neuroimaging of Exosomes Using Gold Nanoparticles. ACS Nano 2017, 11 (11), 10883–10893. [CrossRef]
- Lai, C.P.; Kim, E.Y.; Badr, C.E.; Weissleder, R.; Mempel, T.R.; Tannous, B.A.; Breakefield, X.O. Visualization and Tracking of Tumour Extracellular Vesicle Delivery and RNA Translation Using Multiplexed Reporters. Nat. Commun. 2015, 6, 7029. [CrossRef]
Figure 1.
Overview of exosome composition and endosomal biogenesis. Exosomes contain membrane proteins (tetraspanins, adhesion molecules, signaling mediators), cytosolic components (HSPs, ESCRT proteins, cytoskeletal proteins, Rab-GTPases, annexins), and nucleic acids (miRNAs, lncRNAs, circRNAs, DNA). The figure also illustrates their formation within multivesicular bodies and provides examples of parental cell-specific exosomal cargo. Abbreviations: Ras-associated binding GTPase (Rab-GTPase); Heat Shock Protein 60 (HSP60); Transforming Growth Factor beta (TGF-β); Hypoxia-Inducible Factor 1 (HIF-1); Cancer Stem Cells (CSCs); Mesenchymal Stem Cell (MSC).
Figure 1.
Overview of exosome composition and endosomal biogenesis. Exosomes contain membrane proteins (tetraspanins, adhesion molecules, signaling mediators), cytosolic components (HSPs, ESCRT proteins, cytoskeletal proteins, Rab-GTPases, annexins), and nucleic acids (miRNAs, lncRNAs, circRNAs, DNA). The figure also illustrates their formation within multivesicular bodies and provides examples of parental cell-specific exosomal cargo. Abbreviations: Ras-associated binding GTPase (Rab-GTPase); Heat Shock Protein 60 (HSP60); Transforming Growth Factor beta (TGF-β); Hypoxia-Inducible Factor 1 (HIF-1); Cancer Stem Cells (CSCs); Mesenchymal Stem Cell (MSC).
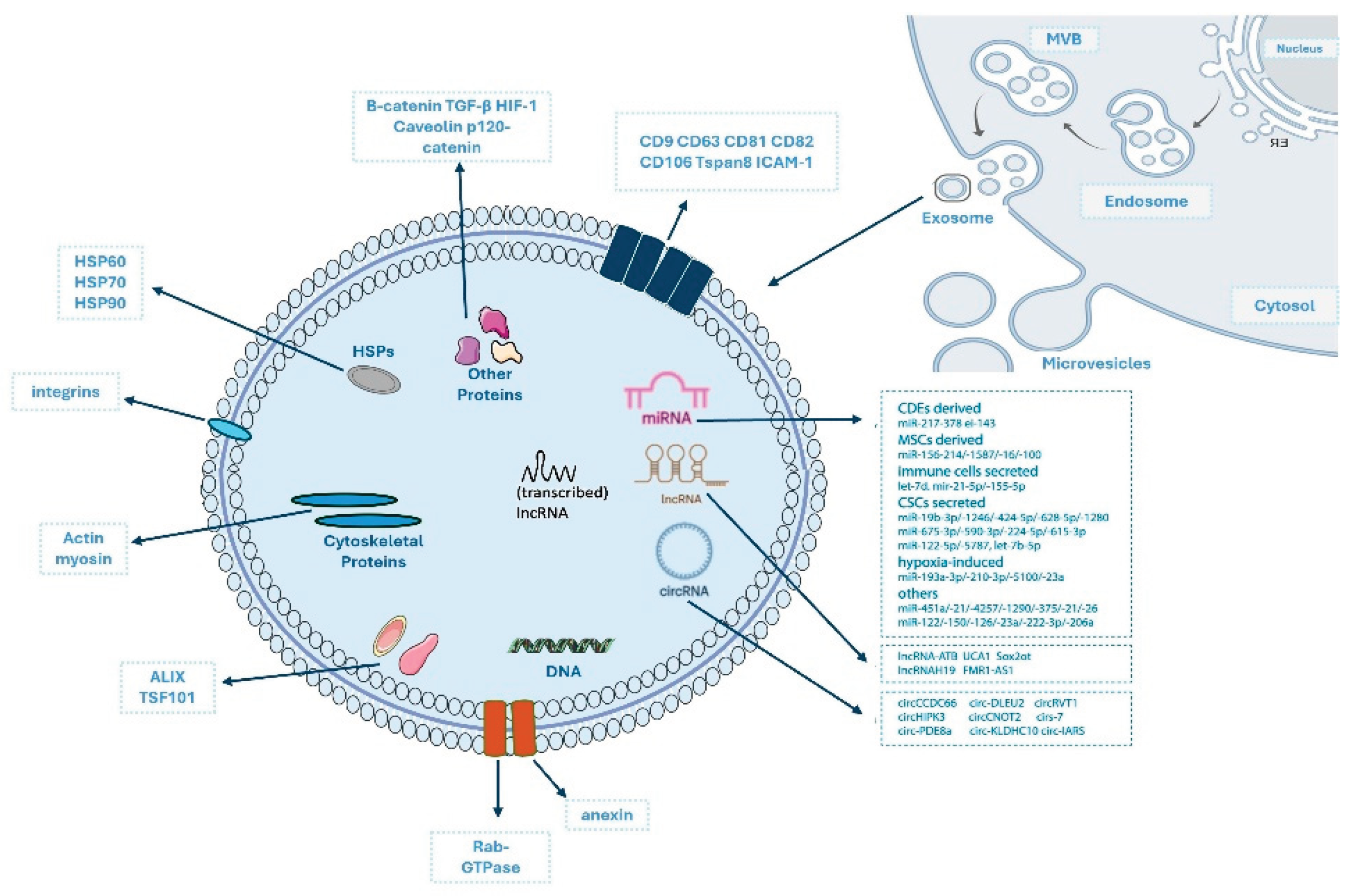
Figure 2.
Percentage distribution of clinical trials involving exosomes registered on ClinicalTrials.gov, displayed as completed (left) and recruiting (right) across four categories: diagnostics, therapy, biomarkers, and drug delivery.
Figure 2.
Percentage distribution of clinical trials involving exosomes registered on ClinicalTrials.gov, displayed as completed (left) and recruiting (right) across four categories: diagnostics, therapy, biomarkers, and drug delivery.
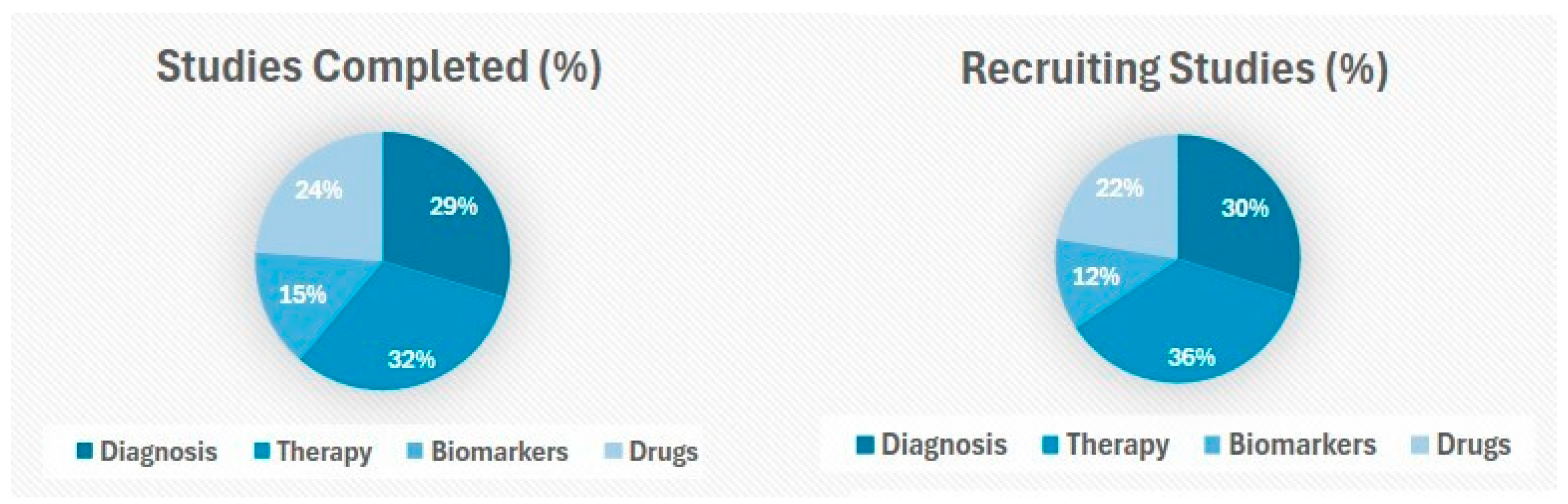
Figure 3.
Overview of exosome composition, modification strategies, and isolation methods. Schematic representation illustrating major components of native exosomes, common engineering approaches used to generate modified vesicles, available purification techniques, and principal cellular interaction pathways. Abbreviation: immunoaffinity capture (IAC). Figure adapted from Tan et al. [61].
Figure 3.
Overview of exosome composition, modification strategies, and isolation methods. Schematic representation illustrating major components of native exosomes, common engineering approaches used to generate modified vesicles, available purification techniques, and principal cellular interaction pathways. Abbreviation: immunoaffinity capture (IAC). Figure adapted from Tan et al. [61].
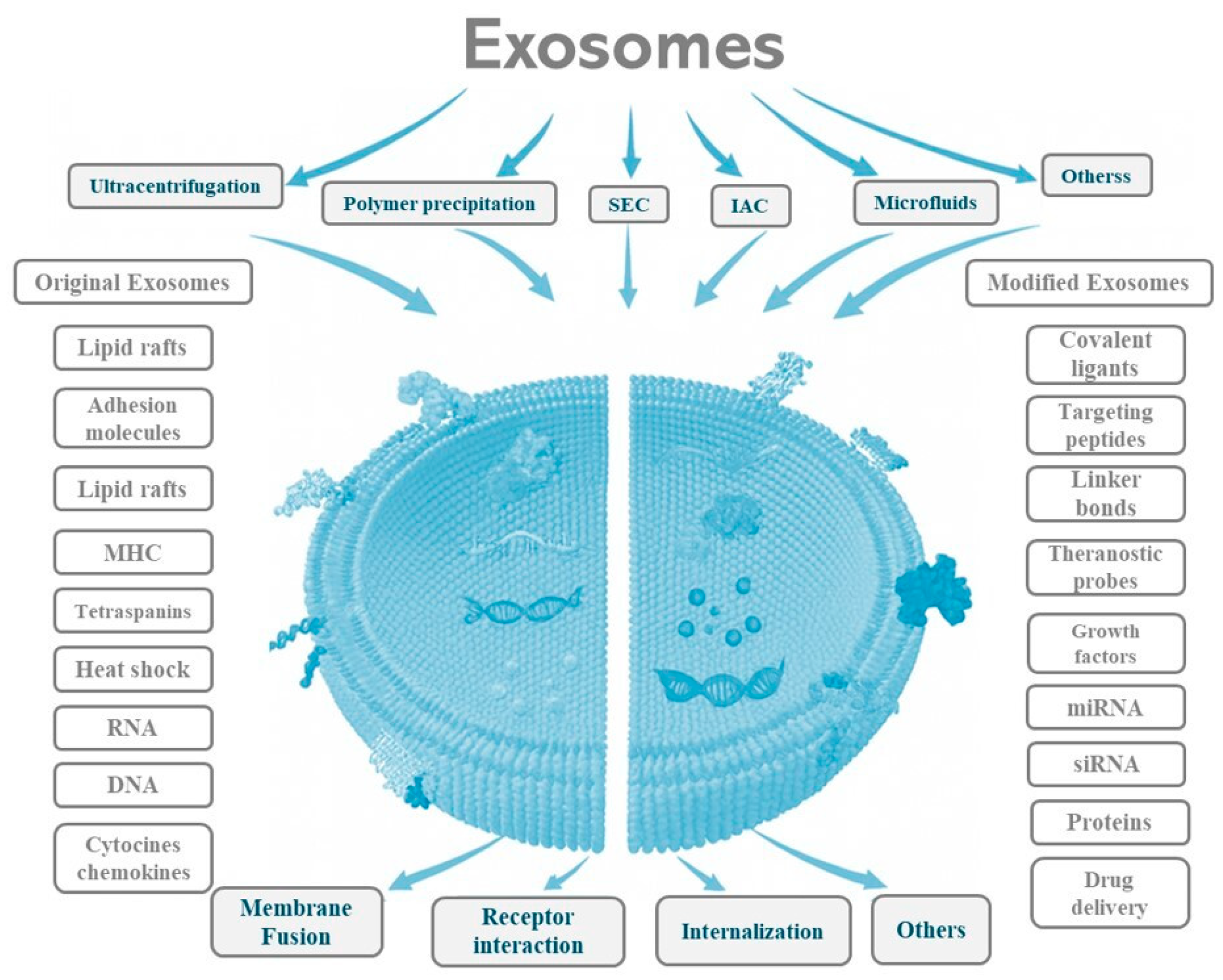
Figure 4.
Overview of exogenous cargo loading techniques for exosomes. Methods are grouped into passive loading (green background), which preserves membrane integrity but yields low loading efficiency; active loading (red background), which relies on membrane-disruptive processes and provides higher efficiency; and specialized techniques, which do not fall strictly into passive or active categories.
Figure 4.
Overview of exogenous cargo loading techniques for exosomes. Methods are grouped into passive loading (green background), which preserves membrane integrity but yields low loading efficiency; active loading (red background), which relies on membrane-disruptive processes and provides higher efficiency; and specialized techniques, which do not fall strictly into passive or active categories.
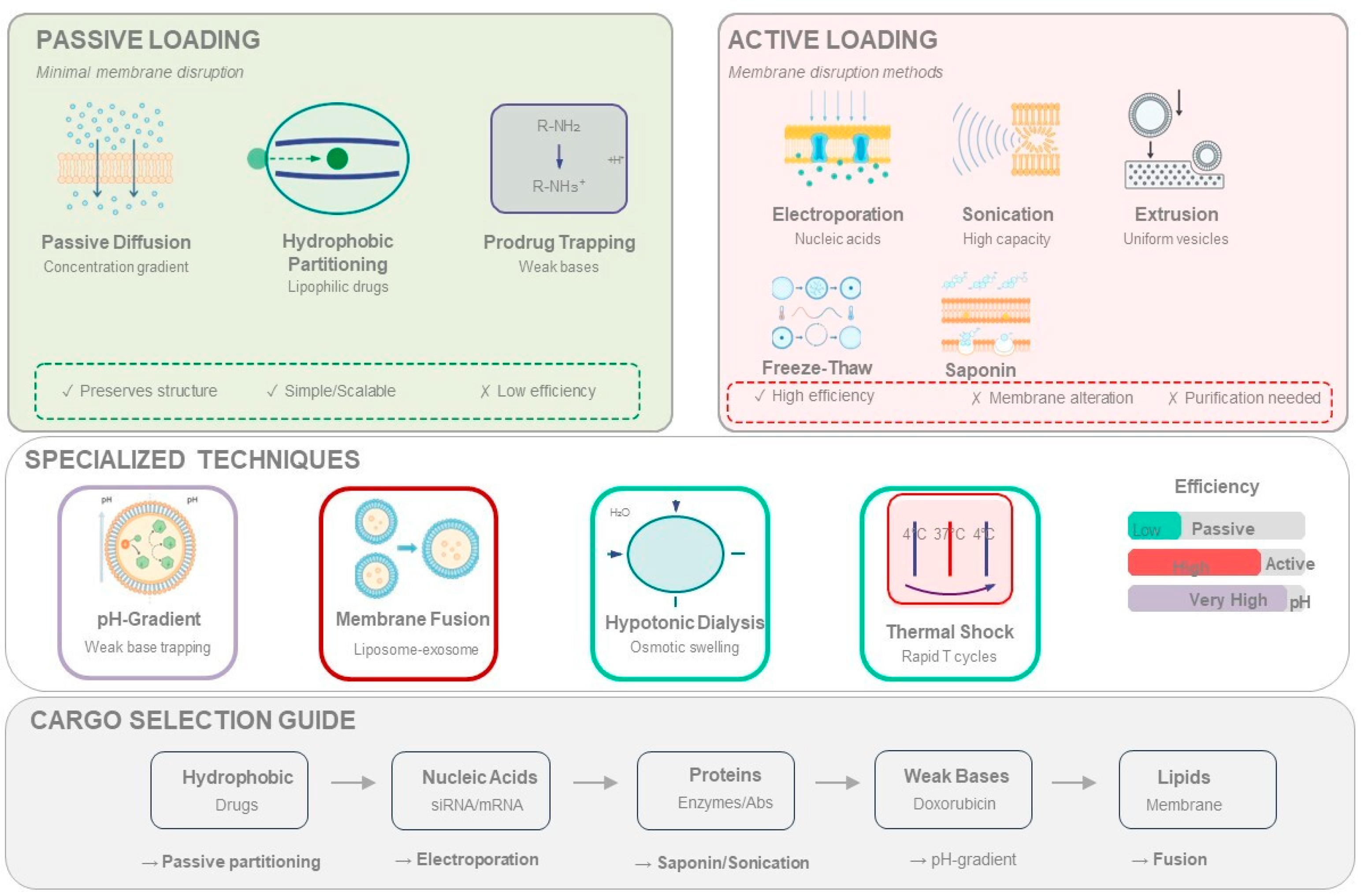
Figure 5.
Future directions for exosome-based renal therapeutics. Schematic overview of emerging research and translational pathways expected to shape the development of exosome-based interventions in nephrology. Near-term priorities include advances in targeting and delivery strategies, gene-editing applications, and diagnostic innovations. Mid-term progress is anticipated through validation in human-relevant preclinical models, AI-driven optimization, and establishment of manufacturing quality standards. These parallel developments converge toward long-term goals of clinical translation, including early-phase clinical trials focused on pharmacokinetics, pharmacodynamics, dosing, and safety.
Figure 5.
Future directions for exosome-based renal therapeutics. Schematic overview of emerging research and translational pathways expected to shape the development of exosome-based interventions in nephrology. Near-term priorities include advances in targeting and delivery strategies, gene-editing applications, and diagnostic innovations. Mid-term progress is anticipated through validation in human-relevant preclinical models, AI-driven optimization, and establishment of manufacturing quality standards. These parallel developments converge toward long-term goals of clinical translation, including early-phase clinical trials focused on pharmacokinetics, pharmacodynamics, dosing, and safety.
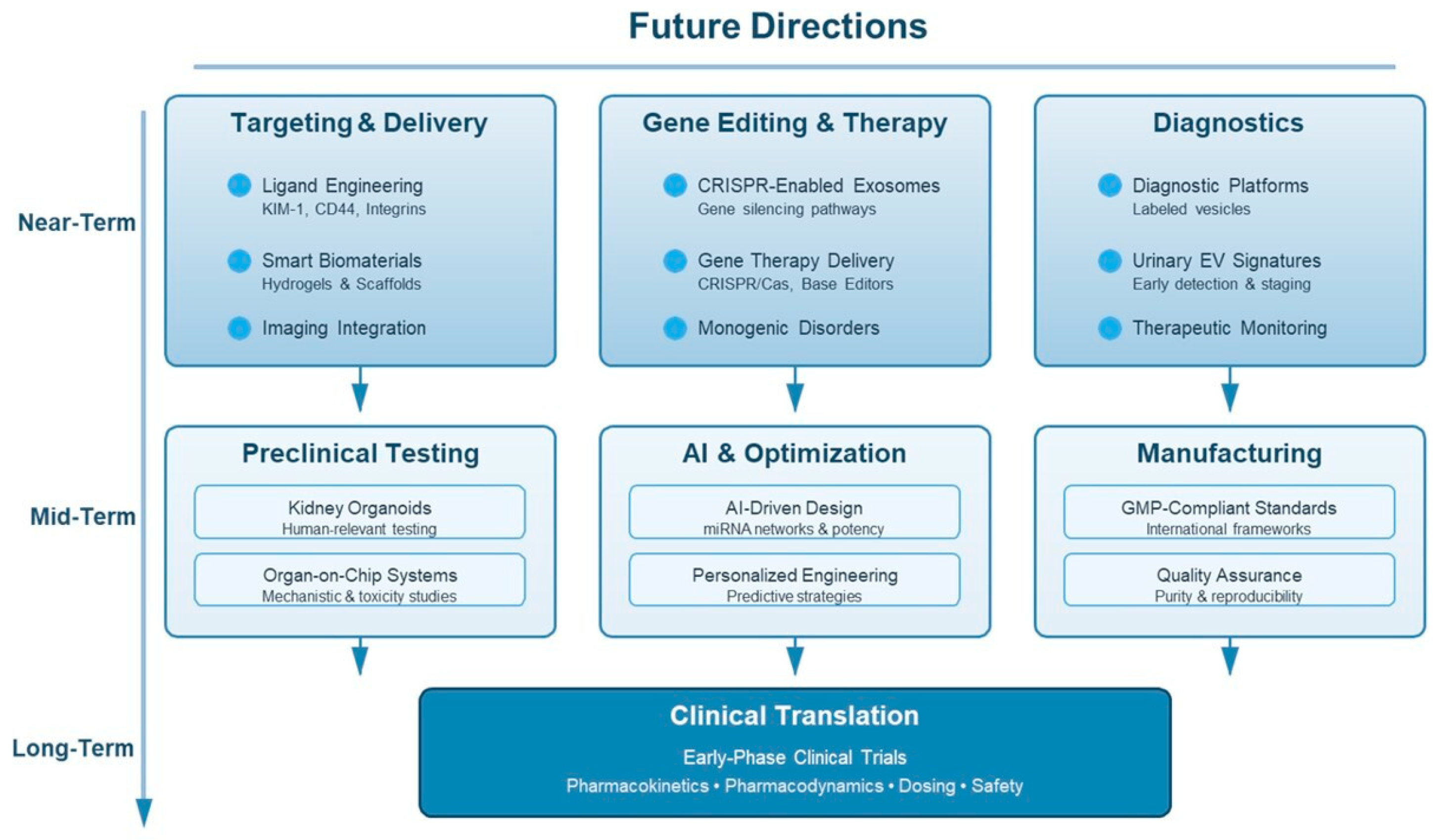

Table 1.
Summary of MISEV2023 recommendations for extracellular vesicle (EVs) isolation, characterization, and analysis [2]. Abbreviations: cluster of differentiation (CD); tumor susceptibility gene 101 (TSG101); ALG-2-interacting protein X (ALIX); heat shock protein (HSP); apolipoprotein (Apo); Golgi matrix protein 130 (GM130); ultracentrifugation (UC); polyethylene glycol (PEG); size exclusion chromatography (SEC); nanoparticle tracking analysis (NTA); asymmetric flow field-flow fractionation (AF4); tunable resistive pulse sensing (TRPS); dynamic light scattering (DLS); tangential flow filtration (TFF); bicinchoninic acid assay (BCA); transmission electron microscopy (TEM); scanning electron microscopy (SEM); cryogenic electron microscopy (Cryo-EM); atomic force microscopy (AFM); Super-resolution microscopy (SRM); flow cytometry (FCM).
Table 1.
Summary of MISEV2023 recommendations for extracellular vesicle (EVs) isolation, characterization, and analysis [2]. Abbreviations: cluster of differentiation (CD); tumor susceptibility gene 101 (TSG101); ALG-2-interacting protein X (ALIX); heat shock protein (HSP); apolipoprotein (Apo); Golgi matrix protein 130 (GM130); ultracentrifugation (UC); polyethylene glycol (PEG); size exclusion chromatography (SEC); nanoparticle tracking analysis (NTA); asymmetric flow field-flow fractionation (AF4); tunable resistive pulse sensing (TRPS); dynamic light scattering (DLS); tangential flow filtration (TFF); bicinchoninic acid assay (BCA); transmission electron microscopy (TEM); scanning electron microscopy (SEM); cryogenic electron microscopy (Cryo-EM); atomic force microscopy (AFM); Super-resolution microscopy (SRM); flow cytometry (FCM).
| Category | MISEV2023 Recommendations | Purpose |
|---|---|---|
|
Positive markers (Membrane-associated) |
CD63, CD9, CD81 | Confirm EVs membrane origin |
|
Positive markers (Cytosolic) |
TSG101, ALIX, HSP70, HSP90 | Validate endosome-derived or EVs-enriched cytosolic cargo |
| Negative markers | ApoA1/ApoB, Albumin, Calnexin, GM130 | Exclude non-EVs contaminants |
| Isolation approaches | Differential UC, Cushion, UC, Density gradients, SEC, TFF, Ultrafiltration, AF4, Immunocapture | Achieve selective enrichment using orthogonal strategies |
| Precipitation methods | PEG, Protamine, Ammonium sulfate, Organic solvents |
High yield but low purity, requiring validation with negative markers |
| Particle quantification | NTA, DLS, TRPS | EVs concentration and size distribution; report the particle-to-protein ratio |
| Protein quantification | BCA, Bradford, Qubit | Estimate total protein levels |
| Morphological validation | TEM, SEM, Cryo-EM, AFM, SRM | EVs structure and membrane integrity |
| Advanced phenotyping | High-sensitivity FCM, Nano-flow FCM | Assess EVs markers and heterogeneity |
| Quality controls | Negative controls, Batch reproducibility, Calibration reporting | Ensure methodological rigor and comparability |
| Category | BM-MSC-Exos | BM-MSC |
|---|---|---|
| Composition | Acellular, containing bioactive molecules | Living cells are capable of self-renewal and differentiation |
| Mechanism of Action | Paracrine communication through the delivery of RNAs, proteins, and lipids | Cell differentiation and tissue integration, in addition to paracrine effects |
| Safety | Low immunogenic risk and no tumorigenic potential | Potential for tumorigenicity, immune rejection, and embolism |
| Therapeutic Effectiveness | More predictable paracrine effects; dependent on vesicle yield and purity | Highly potent but variable due to donor heterogeneity and constraints on cell survival. |
| Production & Scalability | Less complex process, though challenged by purification and yield | Requires cell culture, expansion, differentiation, and cryopreservation |
| Stability & Transport | Storage possible at −80 °C; amenable to lyophilization | Requires cryopreservation and transport in liquid nitrogen |
| Quality Control | High potential for standardization, though batch variability exists | High donor variability; more complex manipulation |
| Regulation | Regulatory frameworks under development; lack of standardized clinical protocols | More established regulations and clinical guidelines |
| Ethical Aspects | Ethically low risk; cell-free products from adult donors | Ethically acceptable but requires invasive bone marrow collection and stringent donor oversight |
| Clinical Applications | Used as personalized therapeutic carriers with wide applicability | Broad applications, primarily in regenerative medicine |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



