
Submitted:
07 February 2026
Posted:
09 February 2026
You are already at the latest version
Abstract
Kidney disease is an alarming universal health concern and a leading cause of morbidity and mortality. About 861 million individuals around the world suffer from kidney complications. However, current treatment alternatives are limited. These limi-tations underscore the impending need for new therapeutic approaches. Autophagy is a dynamic and cellular housekeeping mechanism. The use of conditional autopha-gy-related gene knockouts in kidney cells has led to a better understanding of au-tophagy's significance. Basal autophagy in the kidney serves as a quality control mechanism, vital for cellular metabolism and organelle homeostasis. Under stressful conditions, kidney cells adapt their autophagic activity. This process is intricately con-trolled by signalling pathways that control autophagic flux, with sirtuins, AMP-activated protein kinase (AMPK), and mammalian target of rapamycin (mTOR) acting as key regulators. Additionally, autophagy plays a role in the natural aging process of renal tissue. Small-molecule natural products have demonstrated efficacy in regulating au-tophagy and mitigating kidney damage in several experimental studies. However, specific mechanisms by which small molecules regulate autophagy across different renal disorders have yet to be fully understood. This study reviews that the recent advance-ments in using small molecules in autophagy research have reignited interest in the related signalling pathways and their role in the pathophysiology of renal diseases. Further research into autophagy and its regulatory signalling networks could provide new therapeutic targets for small-molecule intervention in renal disorders.
Keywords:
Autophagy
; Autophagy dysfunction
; Herbal medicine
; Kidney diseases
; Small-molecule natural products.
1. Introduction
According to projections, 700 million individuals worldwide currently have chronic kidney disease (CKD) [1]. The prevalence of kidney disease increases to around 850 million people worldwide when acute kidney injury (AKI) and kidney failure, including dialysis patients and kidney transplant recipients, are considered [2,3]. Current treatment alternatives, including dialysis and kidney transplantation, are inadequate, inaccessible, and often accompanied by major complications like cardiovascular disease, stroke, and immunosuppression. Thus, it is essential to investigate the effective therapeutic approaches to cure and avert the progression of kidney diseases.
Autophagy is a dynamic, strictly controlled self-degradative process that breaks down cytosolic components through lysosomal enzymes. It is a pro-survival mechanism that has been conserved through evolution and is found in all eukaryotic organisms, such as yeast, plants, and mammals. It serves constitutively as a cellular housekeeping mechanism during relaxation, facilitating the removal of long-lived macromolecules and damaged organelles to restore homeostasis and cellular integrity [4]. Dr. Yoshinori Ohsumi was granted the 2016 Nobel Prize in Physiology or Medicine for his pioneering research on the regulation and functional importance of autophagy, which opened the door for an extensive amount of study on the role of autophagy in human disorders [5,6]. Research conducted on animal models has demonstrated a strong link between autophagy and the underlying cause of numerous kinds of diseases, emphasizing its function as a crucial mediator in human disorders [7,8,9,10]. Cellular stress induces changes in autophagy within kidney cells, representing an adaptive mechanism that is controlled by key signaling networks, particularly the mTOR, AMPK, and sirtuin pathways [11,12].
Plants are considered to be a source of medicines for various kidney diseases [13]. Several phytochemicals, such as Ferulic acid, dioscin, resveratol, celastrol, hispidulin, catalpol, curcumin, berberine, Asiatic acid, astilbin, wogonin, and trigonelline, etc. showed promising activities by autophagic function modulation [14] and thus controlling diabetic kidney disease (DKD) in different in vivo and in vitro models [15,16,17,18]. The recent reviews show the beneficial effects of small-molecule natural products in the protection against chronic kidney disease (CKD) [19,20]. These findings demonstrate that autophagy is a promising target for therapeutic intervention in kidney disease, while highlighting the potential of traditionally used phytoconstituents. In this review, we explore the mechanisms of action of small-molecule natural products in the context of kidney disease pathogenesis and treatment, highlighting their prospective roles in maintaining autophagic balance.
2. Materials and Methods
A comprehensive literature search was conducted to identify original research articles published in English that focus on therapeutic compounds for kidney diseases and their underlying mechanisms of action. Google Scholar, Web of Science, Scopus, and PubMed are the four main scientific databases that were explored. Natural compounds, kidney disease, phytochemicals, drug delivery approaches, targeted signalling pathways, renal diseases, autophagy, apoptosis, and therapeutic possibilities in kidney treatment were among the relevant keywords and search terms. The retrieval of appropriate information has been improved by using binary operators and combinations of these phrases. All of the figures in this manuscript were drawn using Adobe Illustrator (Adobe Inc., San Jose, CA, United States) for clarity and visual consistency.
3. Autophagy Dysfunction on Pathophysiology in Kidney Diseases
3.1. Autophagy and Oxidative Stress
Oxidative stress mediates kidney injury by promoting inflammation and cellular damage through excessive reactive oxygen species (ROS) production [21]. It activates signaling pathways that stimulate autophagy, creating a feedback loop that can either protect or harm renal cells, depending on the context [21].
Autophagy is typically activated in response to cellular stressors such as glucose or amino acid scarcity, oxidative stress, hypoxic conditions, and xenobiotic compounds [22]. It has gained recognition as a key regulator of pathological processes, closely linked to ROS, which influence both cellular signaling pathways and oxidative damage [23]. Autophagy serves as a cellular recycling mechanism that mitigates oxidative damage by degrading dysfunctional organelles and proteins, while oxidative stress arises from an imbalance in ROS and antioxidant defenses. This interplay is essential for maintaining renal homeostasis and offers potential therapeutic targets for kidney disorders.
Autophagy and oxidative stress are critical factors in the pathogenesis of kidney diseases, influencing both acute and chronic conditions. In AKI, activated autophagy protects tubular cells from apoptosis, promoting recovery [24]. CKD is associated with impaired autophagy, which exacerbates renal damage and dysfunction. Targeting the balance between autophagy and oxidative stress presents novel therapeutic strategies for kidney diseases, including the modulation of autophagy to enhance renal recovery [4]. Research indicates that interventions aimed at reducing oxidative stress and enhancing autophagy could improve outcomes in patients with kidney diseases [24,25].
3.2. Autophagy and Inflammation
As inflammation is widely acknowledged as the core of most kidney-related diseases, the inflammation pathway can be a breathing room for treatment [26]. Inflammation is a major component in kidney dysfunction, distinguishable in AKI [27]. Another condition, CKD, arises due to low-grade systemic inflammation eventually triggers the terminal illness. A few physical states, for instance, metabolic syndrome, diabetes, and heart disease, correlated with low-grade inflammation [28]. In CKD, the elongated inflammation process occurs when an injury leads to the production of proinflammatory cytokines such as tumor necrosis factor-alpha (TNF-α), interleukin-1 (IL-1) later enhances blood circulation, fluctuations in chemical mediators, and apparent infiltration of leukocytes [29].
The kidney requires an impeccable function of autophagy to preserve an integral endogenous atmosphere; however, dysregulations in autophagy can hinder the process. Disruptive autophagy can be a consequence of inflammation, along with ischemia and virulent injury, which can lead to renal cell cancer [30]. There are some crucial observations suggesting the combined role of inflammasome NLRP3 and NLRs, which eventually triggers caspase-1 to express IL-1β and IL-18. This phenomenon is instigated by the inflammasome NLRP3, ambiguously related to autophagy dysregulations, leading to several diseases accompanied by kidney diseases [31].
In vitro model, which consists of transwell co-culture endothelial cells along with kidney organoids and an Atg-7-lacking mouse model, precise to endothelial cells in the role of in vivo representation, demonstrated that autophagy disruption in endothelial cells leads to impaired endothelial barrier, indicating enhanced NLRP3 inflammasome function, resulting in kidney degradation [32].
A clinical study conducted on 60 DKD patients showed a disproportionate rise of inflammatory factors such as AIF-1, miR-34a, NLRP3, IL-1β, and IL-18 in DKD patients’ blood and urine samples. Conversely, the presence of autophagy-related gene ATG4B declined in the sample, hence establishing the impact of extreme inflammation on autophagy dysregulation in kidney disease [33].
3.3. Autophagy and Fibrosis
The characteristic symptoms of fibrosis include the loss of capillary networks and the buildup of inflammatory cells, fibrillary collagens, and activated myofibroblasts [34,35]. This fibrogenic response, which is closely linked to inflammation and occurs after various forms of tissue injury, is the initial attempt at tissue repair in the dynamic wound healing process. Fibroblasts and myofibroblasts deposit extracellular matrix (ECM) components, such as fibrillar collagen, elastin, and fibronectin [36]. Renal fibrosis occurs due to injury to the kidney that triggers local pericytes and fibroblasts to proliferate and transform into myofibroblasts, which exhibit greater motility and ECM deposition [37,38]. After severe or repeated AKI, incomplete or maladaptive tubular repair results in persistent inflammation and renal interstitial fibrosis, which links AKI to the advancement of CKD [39,40,41,42,43]. Progressive CKD often results in tubulointerstitial fibrosis. The latest study demonstrates that renal fibrosis is accelerated by persistent autophagy dysfunction or deficiency [44].
After three days of UUO, autophagy and interstitial fibrosis peaked in the rat kidneys that had been obstructed. The 3-methyladenine (MA)-induced autophagy suppression consequently decreased Akt/mTOR signaling, exacerbated tubular cell death, and interstitial fibrosis [45]. Another study exhibited that Autophagy has a protective function in diabetic nephropathy, as demonstrated by diabetic mice with impaired autophagy and exacerbated renal hypertrophy, tubular damage, inflammation, fibrosis, and albuminuria after Atg7 was ablated from the proximal tubules [46]. Again, pharmacologically inhibiting autophagy in UUO mice significantly decreased the accumulation of lipids in kidney tubules, which resulted in a decrease in tubular cell degeneration and apoptosis, as well as a suppression of kidney interstitial fibrosis [47].
In an in vitro experiment, autophagy inhibitors inhibited the accumulation of fibrotic proteins and the secretion of pro-fibrotic growth factors in renal proximal tubular cells, which were activated by repeated low-dose cisplatin. After cisplatin nephrotoxicity, results demonstrate the crucial role autophagy plays in maladaptive kidney repair and interstitial fibrosis, possibly via improving the generation and release of pro-fibrotic cytokines [48]. Another study showed, TGFβ1 caused primary proximal tubular cells to undergo autophagy, death, and FN1 accumulation in vitro. While autophagy promotion boosted TGFβ1-induced cell death, autophagy suppression decreased both FN1 accumulation and apoptosis. According to these results, renal interstitial fibrosis during unilateral ureteral obstruction (UUO) may be facilitated by persistent autophagy activation in kidney proximal tubules, regulating tubular cell death, interstitial inflammation, and the synthesis of profibrotic factors [49].
A clinical study explains that in chronic renal graft dysfunction, decreased levels of ATG16L expression hinder autophagy, which elevates inflammation (IL-1β, IL-6, and TNF-α) and induces EndMT and renal interstitial fibrosis [50].
3.4. Autophagy and ER Stress
The endoplasmic reticulum (ER) plays a numerous function, which includes the transportation of proteins to the cell surface or various organelles, as well as being associated with the processing, synthesis, and folding of proteins [51]. When excess misfolded proteins accumulate in the ER, ER stress occurs, and the unfolded protein response (UPR) is activated to restore homeostasis [52]. However, if ER stress becomes severe or ongoing, the adaptive UPR pathway may be interfered with, and UPR-related apoptosis may be triggered [53].
Recent studies have shown that ER stress affects kidney function and that increased markers of ER stress in renal biopsies of AKI patients correlate with the severity of AKI [20,54]. Urinary angiogenin was produced by renal tubular cells as a result of activation of the transcription factor XBP1 during ER stress, indicating the incidence of ER stress in this clinical situation [55]. Cysteine-rich with EGF-like domains 2 (CRELD2) is a newly identified urinary biomarker that sensitively detects ER stress and is significantly increased in uromodulin (UMOD)-associated kidney diseases, such as ischemic AKI [56]. Autophagy is the degradation of misfolded proteins and damaged organelles by lysosomes, resulting in the release of essential components for cellular metabolism [57]. The ATG genes, AMP-activated protein kinase (AMPK), and mechanistic target of rapamycin (mTOR) all control it. The autophagy process can be triggered by AMPK activation or mTOR inhibition [12,58]. Upon stimulation, Serine/Threonine Kinase Unc-51-like Kinase-1 (ULK1) activates PI3K-Beclin1 to initiate autophagosome formation from the ER. The phagophore material is encapsulated by Atg12 and LC3-PE to form a mature autophagosome. Microtubules deliver it to the lysosome, where metabolism occurs; LC3-II is the autophagy signaling protein [59]. This correlation is especially important in AKI, where autophagy is activated as a protective mechanism [60,61].
Abnormal regulation of autophagy associated with pathogenic ER stress causes podocyte injury and tubular cell apoptosis, which accelerates CKD progression [53]. Similarly, in podocyte models, Inositol-requiring enzyme 1 deletion causes microvilli alterations and foot process shrinkage, which increases albuminuria due to reduced autophagy [62]. ER stress does not always promote autophagy. In certain pathological conditions, such as neurodegenerative diseases, ER stress can disrupt the autophagy process [63]. Studies have shown that deletion of the Atg5 gene in podocyte cells of aged mice (20–24 months) results in glomerulosclerosis and podocyte degeneration, ER stress, decreased proteasome activity, accumulation of ubiquitinated proteins, and albuminuria [64]. In contrast, Young mice showed no significant problems and only low levels of albuminuria; multiple unspecified mechanisms may be at work behind protein accumulation when autophagy is disrupted [65].
3.5. Molecular Mechanisms Involved in Autophagy Dysfunction in Kidney Disease
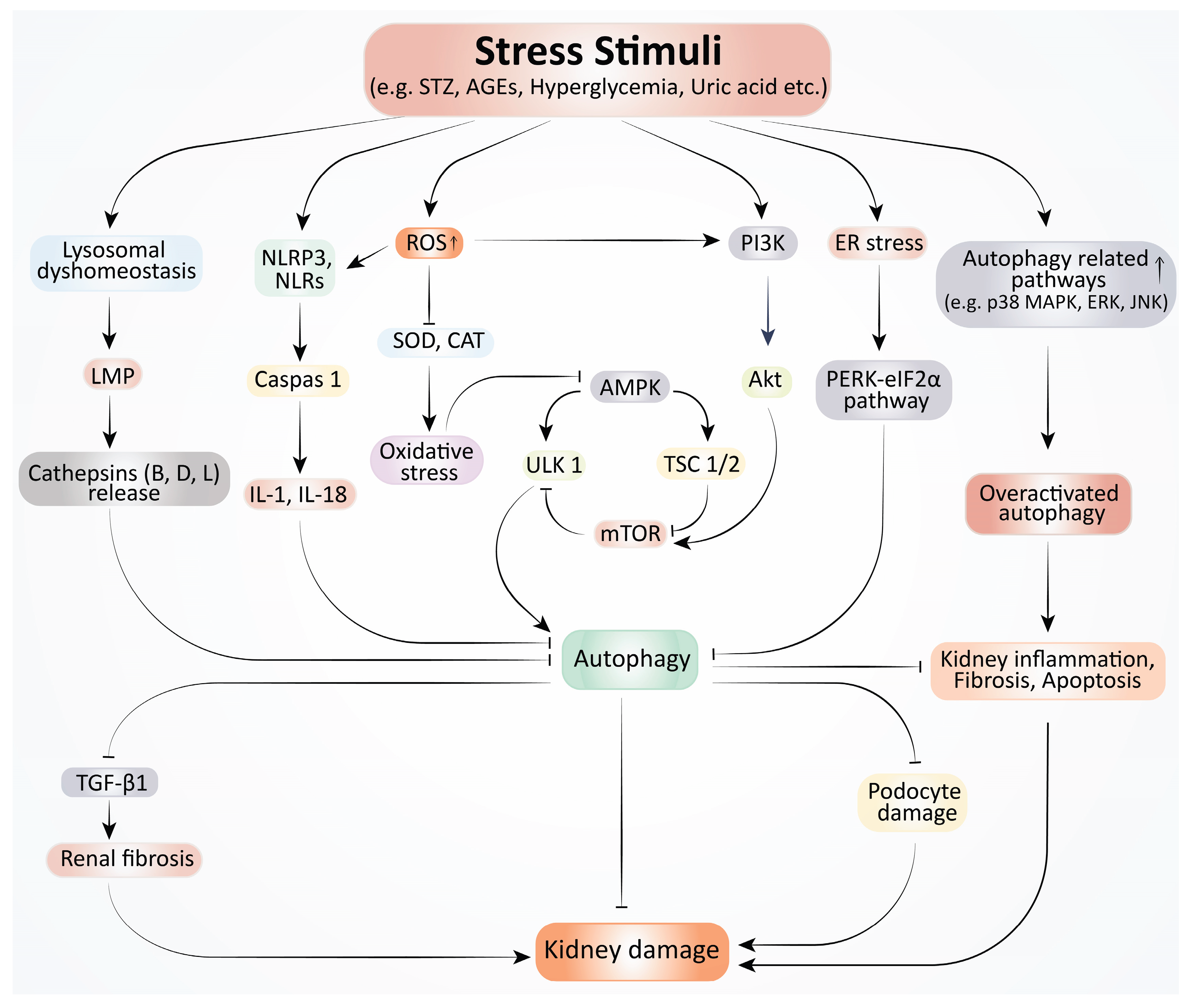
Autophagy, a crucial cellular housekeeping mechanism (Error! Reference source not found.), plays an essential role in maintaining kidney cell health, particularly under stress [11]. It operates through well-characterized regulators, such as mTOR, AMPK, and sirtuins, and is particularly vital in renal cell types, including podocytes, tubular epithelial cells, and immune cells [103]. In healthy kidneys, autophagy removes damaged proteins and organelles, sustaining cellular function and homeostasis. However, impaired or dysregulated autophagy is increasingly recognized as a central contributor to various kidney diseases, including DKD, CKD, and uric acid nephropathy [66,67,68]. In DKD, which is a major cause of end-stage renal failure, high glucose environments disrupt autophagic processes, exacerbating cellular damage [67]. Podocytes and proximal tubular cells are especially vulnerable, with impaired autophagy accelerating their dysfunction [66]. Similarly, lysosomal dyshomeostasis—critical for autophagic flux—has emerged as another layer of dysfunction in DKD, suggesting that strategies to enhance lysosomal function may hold therapeutic promise [69]. Hyperuricemia-induced kidney injury also involves autophagy dysregulation. In urate oxidase knockout rat models, elevated serum urate levels triggered kidney inflammation and fibrosis through autophagy-related pathways, including AMPK, p38 MAPK, ERK, and JNK [68]. Notably, inhibiting autophagy via the PI3K pathway mitigated these effects, underscoring the complex, sometimes detrimental role of autophagy in kidney pathology [68]. In CKD, the scenario is equally nuanced. Impaired autophagy contributes to progressive damage and renal fibrosis, often influenced by cytokines like TGF-β1, which has dual roles—promoting both fibrosis and autophagy depending on the disease context. Protective autophagy activation in proximal tubular cells has been shown to counteract stressors such as proteinuria and ischemia [66]. However, these effects are tightly modulated by multiple intersecting signaling pathways [70,71].
Among these, the PI3K/Akt/mTOR pathway stands out as a central regulator. Frequently overactivated in kidney diseases, it suppresses autophagy and promotes inflammation and fibrosis [71,72]. Conversely, AMPK/ULK1 signaling promotes autophagy under energy stress and shows renoprotective effects, particularly in diabetic and drug-induced nephropathies [73]. Sirtuin 1 (SIRT1)/LC3 interactions further support autophagy by enhancing the clearance of damaged components and reducing oxidative stress [74,75]. Additionally, the ERK, STING1, and PKC pathways demonstrate context-dependent roles in either promoting or inhibiting autophagy, influencing outcomes in inflammation, fibrosis, and toxic injuries [76,77,78]. Autophagy also intersects with regulated cell death pathways, including apoptosis, necroptosis, and ferroptosis, adding further complexity to kidney disease mechanisms [79]. These processes contribute to inflammation and fibrosis, highlighting autophagy as a potential modulator of broader cell fate decisions in the kidney. Despite significant advances, the dual nature of autophagy—as both protective and potentially harmful—underscores the need for precise, context-specific modulation [67,69].
Figure 1.
Autophagy dysfunction in kidney disease. Stress stimuli STZ, AGEs, hyperglycemia, and uric acid, raise ROS production, which lowers antioxidant enzyme (SOD, CAT) activity and causes oxidative stress. As a result, oxidative stress disrupts autophagy by inhibiting the AMPK pathway. Elevated ROS also suppresses autophagy through the activation of the NLRP3 and PI3K pathway. NLRP3 inflammasome activation results in caspase-1 activation and the release of pro-inflammatory cytokines (IL-1, IL-18) that block autophagy. The hyperglycemia condition triggers the PI3K pathway, which in turn triggers the mTOR pathway through the activation of the Akt pathway, finally resulting in a reduction in autophagy. The lysosomal dyshomeostasis causes LMP, which releases cathepsins and suppresses autophagy. ER stress also disrupts autophagy through the activation of the PERK-eIF2α pathway. Both Autophagy disruption and overactivation by several pathways (p38 MAPK, ERK, JNK) cause inflammation, fibrosis, apoptosis, and injury in podocytes. Collectively, these pathological mechanisms lead to kidney damage.
Figure 1.
Autophagy dysfunction in kidney disease. Stress stimuli STZ, AGEs, hyperglycemia, and uric acid, raise ROS production, which lowers antioxidant enzyme (SOD, CAT) activity and causes oxidative stress. As a result, oxidative stress disrupts autophagy by inhibiting the AMPK pathway. Elevated ROS also suppresses autophagy through the activation of the NLRP3 and PI3K pathway. NLRP3 inflammasome activation results in caspase-1 activation and the release of pro-inflammatory cytokines (IL-1, IL-18) that block autophagy. The hyperglycemia condition triggers the PI3K pathway, which in turn triggers the mTOR pathway through the activation of the Akt pathway, finally resulting in a reduction in autophagy. The lysosomal dyshomeostasis causes LMP, which releases cathepsins and suppresses autophagy. ER stress also disrupts autophagy through the activation of the PERK-eIF2α pathway. Both Autophagy disruption and overactivation by several pathways (p38 MAPK, ERK, JNK) cause inflammation, fibrosis, apoptosis, and injury in podocytes. Collectively, these pathological mechanisms lead to kidney damage.
4. Therapeutic Effects of Phytochemicals against Autophagy Dysfunction in Kidney Diseases
Table 1 shows the pharmacological potentials of various phytochemicals against several pathophysiological mechanisms (i.e., oxidative stress, inflammation, fibrosis, and other pathologies) in autophagy dysfunction in kidney diseases.
4.1. Acute kidney Injury
AKI is a condition where kidney function abruptly diminishes to maintain its basic function, which may further lead to CKD with a significant fatality rate. AKI is identified through the increased serum creatinine level and low volume of urine production [90].
Autophagy initiation in proximal tubules illustrated its nephrotoxic utility in AKI; however, dysregulated autophagy eventually leads to decreased cell [91]. A study demonstrated promising impacts of fibroblast growth factor 10 to alleviate IRI-AKI by hindering disproportionate autophagy, thus perceptible as a significant therapeutic objective for AKI [92]. Several studies demonstrated the crucial impact of omitted autophagy protein-related consequences, which ultimately had a detrimental effect on renal function, as well as rising P62 levels and oxidative stress. Moreover, autophagy dysregulation is associated with impaired tubules, indicating dysregulated autophagy pathways hold potential therapeutic sites [93].
Rapamycin has been regarded as an autophagy inducer in AKI therapeutic strategies, along with some autophagy inhibitors, for instance, 3-MA and Chloroquine [94]. Nevertheless, these interventions are futile to ensure a reduced mortality rate in AKI-related cases. At present, despite some interventional therapeutic strategies, renal replacement therapy remains the only effective approach in the minimization of AKI [95]. Since AKI has been regarded as a global health concern, an approach to apply traditional Chinese medicine (TCM), which is enriched in phytochemicals, has validated some splendid clinical outcomes [96]. Phytochemicals present in TCM demonstrated their efficacy by approaching different pathways of AKI, such as oxidative stress reduction, modifying autophagy, and lessening impairment in mitochondria [97]. Berberine (BBR), an active organic compound originating from rhizomacoptidis, exhibits widespread pharmacological impact precisely in mitophagy. By inhibiting NLRP3 inflammasome instigation, BBR initiates mitophagy in contrast-induced AKI (CI-AKI) mice [98]. Another bioactive compound, Tetramethylpyrazine (TMP), exhibited its potential in modulating mitochondrial autophagy in renal tubular cells by overwhelming the CCL2/CCR2 pathway [99]. A recent study revealed the role of Paeoniflorin (PF), derived from Paeonia lactiflora Pal in attenuating the damage of autophagy flux due to intestinal I/R through LKB1/AMPK trail initiation [100]. Furthermore, Neferine (Nef), a bisbenzylisoquinoline alkaloid, which is a bioactive compound, possesses diverse pharmacological potentials, including autophagy stimulation in CIS-Induced AKI [101]. An erstwhile study of Quercetin's impact on prompted initiation of SIRT1, eventually led to p53 deacetylation. These phenomena endorsed autophagy in renal TECs, ultimately decreasing sepsis-induced AKI [75]. In both in vivo and in vitro studies, an investigation concentrated on proteins and genes concomitant with kidney injury, apoptosis, and autophagy, BBR evidently displayed a substantial defensive approach against CI-AKI. BBR initiated autophagy, as per the deviations that occurred in autophagy-related proteins and autophagic flux [102].
4.2. Chronic Kidney Disease
CKD involves progressive loss of kidney function due to structural and functional changes [103]. It is characterized by nephron loss, inflammation, myofibroblast activation, and extracellular matrix deposition. Lipotoxicity, oxidative stress, inflammation, and fibrosis lead to the exacerbation of CKD, which is a global health concern affecting 10% of the population [70,104].
Ultimately, the primary cause of CKD progression to end-stage renal disease (ESRD) is renal fibrosis. Autophagy can be activated as an endogenous defense mechanism in podocytes and renal tubular epithelial cells, and CKD progresses when the autophagic process is compromised [105]. In kidney cells, oxidative stress, inflammation, and mitochondrial dysfunction alter autophagy activation and inhibition, which results in dysfunctional cellular recycling [106]. Since autophagy may either protect or damage cells depending on the experimental setup, targeting autophagy malfunction in CKD patients with phytochemicals is a possible therapeutic approach [107].
A recent computational study reported that potential plant flavonoids may have potential against kidney fibrosis targeting TGFβR-1 [19]. Curcumin could potentially treat CKD by improving renal autophagy in rats with experimental membrane nephropathy via controlling the Nrf2/HO-1 and PI3K/AKT/mTOR signaling pathways [108]. Similarly, in renal tubular epithelial cells, ginsenoside Rb1 (G-Rb1) improves autophagy through the AMPK/mTOR pathway both in vitro and in vivo [109]. In addition, Astragaloside IV boosted autophagy activation and markedly suppressed PI3K/AKT/AS160 pathway activity in both in vitro and in vivo conditions [110]. Likewise, Isovitexin (IV) alleviated renal injury and inflammation by promoting protective autophagy primarily through its anti-ROS production, anti-inflammation, and anti-pyroptosis in mice [111]. Notably, Phytosterols (PS), especially stigmasterol and β-sitosterol, alleviate CKD-related renal damage by stimulating PINK1/Parkin-mediated mitophagy and reducing inflammation, highlighting the role of PS and autophagy in CKD therapy [112]. Furthermore, Danggui Shaoyao San (DSS) reduces TGF-β1-induced apoptosis and fibrosis in NRK-52E renal cells by promoting autophagy, highlighting the potential of phytochemicals targeting autophagy to slow CKD progression [113]. Moreover, Pterostilbene (PT) induces autophagy, which may help prevent renal fibrosis by attenuating NLRP3 inflammasome activation and epithelial-mesenchymal transition, indicating its potential clinical application for better CKD management [114]. Meanwhile, Sulforaphane (SFN) reduces renal fibrosis in UUO mice by dual-regulating mTOR-mediated autophagy, increasing autophagy in renal fibroblasts and decreasing it in tubular epithelial cells, emphasizing SFN’s potential in CKD treatment through autophagy modulation [115].
By contrast, Rhein suppresses autophagy in rat renal tubular cells by modulating AMPK/mTOR, p38/Erk MAPK, and Akt-independent pathways, suggesting its potential therapeutic role in treating CKD patients in the clinic [116]. Finally, Rhubarb-Astragalus may improve CKD, possibly by inhibiting autophagy via the p38-MAPK/TGF-β1 and p38-MAPK/smad2/3 pathways [117].
4.3. Obesity-Related Nephropathy
Obesity-related nephropathy (ORN) is a growing health concern due to its rising incidence and progressive impact on kidney function. Obesity acts as a potent risk factor for CKD, primarily through the interplay of lipotoxicity, insulin resistance, systemic inflammation, and adipocyte dysfunction that ultimately impair renal structure and function [118]. Histopathological features such as obesity-related glomerulopathy (ORG) emerge as specific manifestations, characterized by glomerular hypertrophy, focal segmental glomerulosclerosis, and altered renal hemodynamics [119]. Additionally, the adipose–renal axis—involving pro-inflammatory adipokines and metabolic disturbances—contributes significantly to ORN progression [120].
Despite current treatment strategies—including renin-angiotensin-aldosterone system (RAAS) inhibitors, sodium-glucose co-transporter 2 (SGLT2) inhibitors, glucagon-like peptide-1 (GLP-1) receptor agonists, and bariatric surgery—many patients continue to experience progressive renal decline, as these therapies do not specifically target the underlying mechanisms of obesity-induced kidney injury [120,121]. Given this limitation, phytochemicals have attracted considerable attention for their potential to modulate oxidative stress, inflammation, and autophagy dysfunction—key processes in obesity-related kidney damage. This therapeutic gap has spurred interest in phytochemicals for their ability to influence key pathological pathways such as oxidative stress, inflammation, and autophagy dysfunction, all of which contribute to obesity-related nephropathy. Among these, Wedelolactone (WDL)—a bioactive compound isolated from Eclipta prostrata—has demonstrated anti-obesity effects in preclinical models, primarily through the activation of the SIRT1, AMPK, and peroxisome proliferator-activated receptor alpha (PPARα) signaling axis. This activation not only improves energy metabolism and glucose tolerance but may also support kidney health indirectly by alleviating obesity-induced metabolic strain [122]. Similarly, sulforaphane, found in broccoli sprouts, enhances nuclear factor erythroid 2 (NRF2) activity, thereby reducing hepatic gluconeogenesis and systemic oxidative stress. In diabetic and obese models, sulforaphane supplementation lowered fasting glucose and improved HbA1c, pointing to its relevance in managing obesity-related metabolic and renal dysfunctions [123]. NRF2 is a key transcription factor involved in regulating the cellular antioxidant response and protecting renal tissues from oxidative stress and inflammation. Natural bioactive compounds such as resveratrol, curcumin, and quercetin have been found to stimulate NRF2 activation and promote its translocation to the nucleus. This activation leads to the upregulation of detoxifying and antioxidant enzymes, including heme oxygenase-1 (HO-1) and glutathione S-transferase (GST). Additionally, these phytochemicals can suppress the expression of pro-inflammatory cytokines like IL-6 and TNF-α. Through these mechanisms, NRF2 activation may play a crucial role in mitigating autophagy dysfunction associated with obesity-related kidney damage [124,125]. Furthermore, bilberries and blueberries, rich in anthocyanins and polyphenols, demonstrate the ability to reduce inflammation, improve cytokine profiles, and ameliorate glucose and lipid metabolism in obese models. In high-fat diet-fed mice, blueberry polyphenols embedded in defatted soybean flour (BB-DSF) reduced weight gain, improved glucose tolerance, and lowered fasting glucose and serum cholesterol levels. In vitro studies further revealed their ability to suppress hepatic glucose production—highlighting the potential of polyphenol-enriched foods in mitigating metabolic dysfunctions underlying ORN [126,127]. Betaine, a naturally derived osmoprotective compound present in dietary sources such as beets, spinach, wheat, and certain seafood, supports cellular homeostasis and overall metabolic function. Endogenously, it is produced via the oxidation of choline. Studies using high-fat diet-induced models have shown that betaine supplementation can enhance metabolic performance and reduce insulin resistance [128]. Moreover, individuals with CKD often exhibit reduced circulating betaine levels, which are associated with metabolic syndrome and unfavorable clinical outcomes, indicating its relevance as a potential therapeutic or diagnostic marker in CKD management [129]. Dietary patterns rich in phytochemicals—fruits, vegetables, legumes, and whole grains—have been inversely associated with the risk of diabetic nephropathy, a condition that shares overlapping mechanisms with obesity-related kidney disease. The protective effects are attributed to improved glucose tolerance, reduced inflammation, and blood pressure control [130,131]. Cinnamon, widely used in traditional medicine, contains active compounds that reduce oxidative stress, modulate gut microbiota, and improve metabolic profiles—traits that make it a potential complementary therapy for obesity-related CKD [132].
Despite their promise, the therapeutic efficacy of phytochemicals is influenced by their metabolism, bioavailability, and interaction with gut microbiota. Processes like glucuronidation, methylation, and hepatic transformation determine their systemic availability and renal protective effects. Understanding this pharmacokinetics is essential for optimizing dosing and delivery systems, potentially paving the way for personalized phytochemical-based therapies in metabolic and renal disorders [133]. These findings suggest that phytochemicals, through modulation of autophagy, metabolic signaling pathways, and antioxidant defense, offer a promising adjunct or preventive approach against obesity-related nephropathy. However, further clinical validation and mechanistic studies are warranted to clarify their bioavailability, dosage, and long-term effects in human subjects.
4.4. Diabetic Nephropathy
Diabetic nephropathy (DN) is a long-term complication of diabetes, and its clinical and pathological characteristics take many years to develop in the human body [134]. DN occurs in patients with both type I and type II diabetes, and its main clinical features are proteinuria and progressive deterioration of glomerular filtration rate [135]. Mortality in patients with DN is about 30 times higher than in patients with diabetes but without DN [136]. In primary DN, there is glomerular enlargement, mesangial expansion, and basement membrane thickening. Advanced DN exhibits nodular glomerulosclerosis, mesangiolysis, and tubulointerstitial fibrosis [137]. Nutrient-sensing signaling pathways regulate Autophagy activity and are disrupted by metabolic stress in diabetes [138,139]. Reduced autophagy activity contributes to DN pathogenesis, and an auspicious therapeutic target for DN may be the restoration of autophagy activity [140,141]. Recently, phytochemicals have been increasingly measured against autophagy dysfunction in kidney diseases.
BBR, derived from Rhizoma Coptidis and Phellodendron amurense Rupr., can stimulate autophagy by activating the AMPK signaling pathway, and can reduce podocyte cell death (apoptosis) caused by a high glucose environment [142]. Similarly, Curcumin reduces cell death (apoptosis) and increases protective autophagy by phosphorylating the PI3K/Akt signaling pathway in tubular epithelial cells of AGE-induced rats [143]. Also, the phenolic-rich fraction of Golden berry (P. peruviana L.) may help prevent diabetic nephropathy by increasing autophagy and reducing cell death through the AMPK/mTOR signaling pathway [144]. In vivo studies have found that Keluoxin Capsule regulates autophagy in podocytes and that it works by increasing the expression of LC3-II and p62 proteins [145]. Furthermore, the Dendrobium mixture is effective in protecting the kidney in the streptozotocin (STZ)-induced rat model, which suppresses PI3K/Akt/mTOR signaling pathway and increases autophagy in the kidney [146]. Besides, Yishen Capsule increases the expression of Beclin-1, SIRT1, and LC3-II proteins and simultaneously reduces the expression of NF-κB p65 [147]. Researchers have shown that flavonoids are effective in preventing the development of STZ-induced experimental diabetic nephropathy, inhibiting disease progression by reducing the expression of AGEs, collagen IV, laminin, as well as TGF-β1, p-Smad 2/3, and CTGF proteins in mice [148]. In vitro experiments have shown consistent results in podocytes in a high-glucose environment [149], Celastrol has been reported to protect podocytes from inflammatory responses and restore podocyte viability by activating heme oxygenase-1-mediated autophagy [150]. Astragaloside IV has been reported to play a critical role in reducing the overproduction of ECM proteins and stimulating autophagy through the regulation of Sirt1/NF-κB signaling in KK-ay mice [151]. As well as Resveratrol also reduces kidney cell death by stimulating autophagy in podocytes of db/db mice in vitro, and reduces damage in DKD by suppressing miR-383-5p and increasing miR-18a-5p [152,153]. Leaf extract from Cassia auriculata ameliorates kidney damage by reducing autophagy-related necroptosis through RIP-1/RIP-3-p-p38MAPK signaling in glomerular endothelial cells of STZ-induced rats and rats induced by high glucose [154]. A recent study showed that Radix astragali delayed the development of DKD by increasing podocyte autophagy and possibly suppressing the PI3K/Akt/mTOR pathway [155]. In vivo experiments showed that Tongluo Digui decoction protected podocytes and reduced proteinuria by inhibiting mTOR phosphorylation and increasing autophagy in STZ-induced mice [156].
4.5. Hypertensive Nephropathy
Hypertensive nephropathy (HN) is a main cause of CKD and ESRD, which has a vital role in morbidity, mortality, and rising healthcare costs. Long-term high blood pressure can be responsible for HN, which can damage the kidneys primarily. It can lead to glomerulosclerosis, glomerular hypertrophy, tubular atrophy, interstitial fibrosis, and inflammation [157]. The NLRP3 inflammasome, which is a predominant source of activated IL-1β, has been found as responsible for the development of hypertensive kidney injury [158]. Essential hypertension can damage renal structure and function, which leads to renal inflammation [159]. On this stage, the renin-angiotensin-aldosterone system (RAAS) becomes activated, which upregulates the expression of angiotensin II (AngII). It promotes dysfunction of renal endothelial cells that release different inflammatory factors and promote renal inflammation. AngII is an important mediator of HN [160]. Different studies have shown that hydrogen sulfide donors can relieve renal damage by regulating autophagy, oxidative stress through different signaling pathways [161]. On the other hand, many studies show that hydrogen sulfide also regulates miRNAs to improve cardiac and renal dysfunction [162,163]. Studies are being conducted to find out which miRNAs and mRNAs control autophagy in hypertensive kidney disease, and the effect of hydrogen sulfide donors to elucidate molecular-level treatment [164].
A phytochemical named YGYSG is an herbal medicine that works on kidney damage caused by hypertension on a specific cellular process in the PI3K/AKT/mTOR signaling pathway. Network pharmacology identified autophagy targets and ten core components in the YGYSG treatment of Hypertensive Renal Disease. Here, YGYSG is involved with the PI3K/AKT/mTOR signaling pathway, also with the autophagy process. The treatment was introduced to kidney cells with AngII, where it effectively reverses the AngII function and protects the cells. As it normalizes autophagy in a cell line, it may also work in vivo [165].
4.6. Obstructed Nephropathy
Obstructed nephropathy is a common disease in all ages, from children to older people. It is a renal disease mainly caused by the impaired flow of urine or tubular fluid [166]. Many studies have shown that AngII has a vital role in the initiation and development of obstructed nephropathy, which directly or indirectly promotes the production of molecules that can contribute to renal injury [167]. Inhibition of angiotensin production can cause renal injury, which is responsible for the obstructed nephropathy [168]. The pathophysiology of obstructed nephropathy is multifactorial and has multiple interactions with hemodynamic, inflammatory, and fibrotic pathways. Primary response to acute, full ureteral obstruction is a temporary rise in renal blood flow, followed by profound vasoconstriction [169]. This is mediated by several vasoactive compounds, such as AngII, thromboxane A2, and antidiuretic hormone [166].
Most of the studies in the case of autophagy in renal interstitials have been run using the UUO model [170]. For renal interstitial fibrosis, UUO is a perfect model of renal interstitial fibrosis, which can respond to inflammation, ECM deposition, and tubular atrophy [45,171]. Some studies have shown that autophagy has antifibrotic effects in UUO-associated renal fibrosis. It can reduce the extent of renal fibrosis by decreasing ECM deposition and renal tubular atrophy [45]. Autophagy also has an effect on renal fibrosis by regulating the TGF-β1 and NLRP3 inflammatory vesicle signaling pathways. Autophagy can slow down renal interstitial fibrosis through negative regulation of TGF-β1 by promoting mature TGF-β1 degradation in the UUO kidney [140]. A study found that treatment with rapamycin, a known drug to increase autophagy, successfully increased autophagy in the kidney of a rat model with obstructed kidney disease, and this led to a significant reduction of kidney fibrosis and also decreased disease progression [172].
Researchers found that curcumin can reduce interstitial fibrosis effectively in mice with obstructive nephropathy. They found two mechanisms, one is by suppressing the inflammatory response that remarkably reduces key inflammatory proteins (IL-6, IL-1β, and TNF-α). Another one is by inhibiting the epithelial-mesenchymal transition that drives fibrosis by reducing vimentin marker expression. Here, curcumin mainly blocked the LR4/NF-κβ and PI3K/AKT signaling pathways, which are the main drivers for inflammation and fibrosis [173].
5. Recent Updates on Bioactive Compounds for Autophagy in Kidney Diseases
Several phytochemicals, including alkaloids, flavonoids, polyphenols, phytosterols, etc., ameliorate autophagy dysfunction in kidney disease. In a mouse model of AKI, BBR, an alkaloid, had therapeutic potential. Nevertheless, BBR has extremely limited solubility and bioavailability. Because of this, substantial doses of BBR are needed to achieve a therapeutic impact, which may have negative effects on the digestive system [174,175]. An advanced drug delivery system would be a viable option to address this problem. A study showed that the BBR carried by Janus nanoparticles consisting of Fe3O4 head was effective in treating hepatocellular carcinoma [176]. Another study reported that Linaxiang microemulsion of BBR chloride increased the bioavailability [177]. Another phytochemical, TMP, with lower bioavailability, permeability, and solubility, has been shown to be very effective in treating AKI [178,179]. According to a study, the encapsulation rate of TMP delivered by liposome carriers was higher both in vitro and ex vivo [180]. Another investigation on rats with spinal cord injuries revealed that TMP nanoparticles containing HIV-1 transcription factor increased the drug's half-life and, as a result, its prolonged activity [181]. The bioactivity of Nef, an alkaloid, is limited because of its low bioavailability. A study on NaNO2-induced anoxia in mice showed that Nef nanoliposomes were effective in protecting against cerebral embolism [182]. Similarly, due to quercetin's poorer solubility and bioavailability, a study recommended delivering it via nanoparticles to treat kidney disease [183].
6. Prospects and Limitations
In renal pathophysiology, autophagy plays an important role in treating kidney disease. Numerous phytochemicals such as BBR, TMP, PF, Nef, Quercetin, Curcumin, G-Rb1, Astragaloside IV, Isovitexin, stigmasterol and β-sitosterol, PT, and SFN improve autophagic pathways in AKI, CKD, and DN. Despite their promising therapeutic potentials, some limitations that should be addressed. First, some phytochemicals are associated with low toxicity and adverse effects. PF showed low toxicity [184]. The LD50 with rats was 14.55 g/kg, suggesting a safety margin [84]. BBR can interact with other drugs and increase the nephrotoxicity. For instance, in combinatorial therapy with vancomycin, the immune system and cause nephrotoxicity [185]. Second, the human body metabolizes SFN very quickly. In order to get a therapeutic effect, a dose determination study is necessary [186]. Third, despite numerous studies on animal models, clinical research is very limited. Clinical trials are needed for evaluating safety, optimal dosing, and pharmacokinetics. By resolving these issues, it will become possible to harness the autophagy-modulating potential of these bioactive phytochemicals to treat kidney diseases.
7. Conclusions
Autophagy function must be maintained in order to improve a number of illnesses, including kidney disease. This review highlighted the pathophysiology of autophagy dysregulation in kidney disease and explored the potential of small bioactive natural molecules to maintain the autophagic process. Collectively, this study suggests that small-molecule natural products would be a viable option for therapeutic intervention for renal disease. The autophagy-targeted treatment is highly recommended using the advanced drug delivery technique, such as a nano-guided system. Further studies on the mode of action and therapeutic effects of phytochemicals are required to establish them as an alternative therapeutic agent for kidney disease.
Author Contributions
MJU and AM conceptualized the manuscript. NA, MF, MTAR, MS, AIH, and MSA collected the data and wrote the preliminary draft of the manuscript. TZS and MMI corrected and improved the draft of the manuscript. MJU and AM reviewed the scientific contents. All the authors read and approved the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
No specific funding was received for this work from any organization.
Data Availability Statement
Not applicable.
Acknowledgments
None.
Conflicts of Interest
There is no conflict of interest among the authors.
References
- Global, regional, and national burden of chronic kidney disease, 1990-2017: a systematic analysis for the Global Burden of Disease Study 2017. Lancet 2020, 395, 709–733. [CrossRef] [PubMed]
- Jager, K.J.; Kovesdy, C.; Langham, R.; Rosenberg, M.; Jha, V.; Zoccali, C. A single number for advocacy and communication-worldwide more than 850 million individuals have kidney diseases. Nephrol Dial Transplant 2019, 34, 1803–1805. [Google Scholar] [CrossRef]
- Francis, A.; Harhay, M.N.; Ong, A.C.M.; Tummalapalli, S.L.; Ortiz, A.; Fogo, A.B.; Fliser, D.; Roy-Chaudhury, P.; Fontana, M.; Nangaku, M.; et al. Chronic kidney disease and the global public health agenda: an international consensus. Nat Rev Nephrol 2024, 20, 473–485. [Google Scholar] [CrossRef]
- Sureshbabu, A.; Ryter, S.W.; Choi, M.E. Oxidative stress and autophagy: crucial modulators of kidney injury. Redox Biol 2015, 4, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, N. A brief history of autophagy from cell biology to physiology and disease. Nat Cell Biol 2018, 20, 521–527. [Google Scholar] [CrossRef]
- Levine, B.; Klionsky, D.J. Autophagy wins the 2016 Nobel Prize in Physiology or Medicine: Breakthroughs in baker's yeast fuel advances in biomedical research. Proc Natl Acad Sci U S A 2017, 114, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Levine, B.; Kroemer, G. Biological Functions of Autophagy Genes: A Disease Perspective. Cell 2019, 176, 11–42. [Google Scholar] [CrossRef] [PubMed]
- Leidal, A.M.; Levine, B.; Debnath, J. Autophagy and the cell biology of age-related disease. Nat Cell Biol 2018, 20, 1338–1348. [Google Scholar] [CrossRef] [PubMed]
- Ravikumar, B.; Sarkar, S.; Davies, J.E.; Futter, M.; Garcia-Arencibia, M.; Green-Thompson, Z.W.; Jimenez-Sanchez, M.; Korolchuk, V.I.; Lichtenberg, M.; Luo, S.; et al. Regulation of mammalian autophagy in physiology and pathophysiology. Physiol Rev 2010, 90, 1383–1435. [Google Scholar] [CrossRef] [PubMed]
- Levine, B.; Kroemer, G. Autophagy in the pathogenesis of disease. Cell 2008, 132, 27–42. [Google Scholar] [CrossRef]
- Tang, C.; Livingston, M.J.; Liu, Z.; Dong, Z. Autophagy in kidney homeostasis and disease. Nat Rev Nephrol 2020, 16, 489–508. [Google Scholar] [CrossRef]
- Sohn, M.; Kim, K.; Uddin, M.J.; Lee, G.; Hwang, I.; Kang, H.; Kim, H.; Lee, J.H.; Ha, H. Delayed treatment with fenofibrate protects against high-fat diet-induced kidney injury in mice: the possible role of AMPK autophagy. American journal of physiology. Renal physiology 2017, 312, F323–f334. [Google Scholar] [CrossRef]
- Pandiri, I.; Biotechnology, A.M.J.J.o.A.; Therapeutics, E. Ocimum herb species: a potential treatment strategy for diabetic kidney disease. 2018. [Google Scholar] [CrossRef]
- Rahman, M.A.; Hannan, M.A.; Dash, R.; Rahman, M.H.; Islam, R.; Uddin, M.J.; Sohag, A.A.M.; Rahman, M.H.; Rhim, H. Phytochemicals as a Complement to Cancer Chemotherapy: Pharmacological Modulation of the Autophagy-Apoptosis Pathway. 2021, 12–2021. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.; Ghosh, S.; Das, A.K.; Sil, P.C. Ferulic Acid Protects Hyperglycemia-Induced Kidney Damage by Regulating Oxidative Insult, Inflammation and Autophagy. Front Pharmacol 2019, 10, 27. [Google Scholar] [CrossRef]
- Liu, P.; Zhu, W.; Wang, Y.; Ma, G.; Zhao, H.; Li, P. Chinese herbal medicine and its active compounds in attenuating renal injury via regulating autophagy in diabetic kidney disease. Front Endocrinol (Lausanne) 2023, 14, 1142805. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, Q.; Wang, C.; Lou, Z.; Li, Q. Trigonelline reduced diabetic nephropathy and insulin resistance in type 2 diabetic rats through peroxisome proliferator-activated receptor-gamma. Exp Ther Med 2019, 18, 1331–1337. [Google Scholar] [CrossRef]
- Putra, I.; Fakhrudin, N.; Nurrochmad, A.; Wahyuono, S. A Review of Medicinal Plants with Renoprotective Activity in Diabetic Nephropathy Animal Models. Life (Basel) 2023, 13. [Google Scholar] [CrossRef]
- Rahman, M.A.; Akter, S.; Dorotea, D.; Mazumder, A.; Uddin, M.N.; Hannan, M.A.; Hossen, M.J.; Ahmed, M.S.; Kim, W.; Kim, B.; et al. Renoprotective potentials of small molecule natural products targeting mitochondrial dysfunction. 2022, 13–2022. [Google Scholar] [CrossRef]
- Uddin, M.J.; Kim, E.H.; Hannan, M.A.; Ha, H. Pharmacotherapy against Oxidative Stress in Chronic Kidney Disease: Promising Small Molecule Natural Products Targeting Nrf2-HO-1 Signaling. Antioxidants (Basel) 2021, 10. [Google Scholar] [CrossRef]
- Kaushal, G.P.; Chandrashekar, K.; Juncos, L.A. Molecular Interactions Between Reactive Oxygen Species and Autophagy in Kidney Disease. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef]
- Sridhar, S.; Botbol, Y.; Macian, F.; Cuervo, A.M. Autophagy and disease: always two sides to a problem. J Pathol 2012, 226, 255–273. [Google Scholar] [CrossRef]
- Wible, D.J.; Bratton, S.B. Reciprocity in ROS and autophagic signaling. Curr Opin Toxicol 2018, 7, 28–36. [Google Scholar] [CrossRef]
- Lin, T.A.; Wu, V.C.; Wang, C.Y. Autophagy in Chronic Kidney Diseases. Cells 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Ai, S.; Li, Y.; Zheng, H.; Wang, Z.; Liu, W.; Tao, J.; Li, Y.; Wang, Y. Global research trends and hot spots on autophagy and kidney diseases: a bibliometric analysis from 2000 to 2022. Front Pharmacol 2023, 14, 1275792. [Google Scholar] [CrossRef]
- Anders, H.J.; Schaefer, L. Beyond tissue injury-damage-associated molecular patterns, toll-like receptors, and inflammasomes also drive regeneration and fibrosis. J Am Soc Nephrol 2014, 25, 1387–1400. [Google Scholar] [CrossRef]
- Kinsey, G.R.; Li, L.; Okusa, M.D. Inflammation in acute kidney injury. Nephron Exp Nephrol 2008, 109, e102-107. [Google Scholar] [CrossRef] [PubMed]
- Akchurin, O.M.; Kaskel, F. Update on inflammation in chronic kidney disease. Blood Purif 2015, 39, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Pahwa, R.; Goyal, A.; Jialal, I. Chronic Inflammation. In StatPearls; StatPearls Publishing.
- Copyright © 2025. StatPearls Publishing LLC.: Treasure Island (FL), 2025.
- Banerjee, P.; Basu, A.; Wegiel, B.; Otterbein, L.E.; Mizumura, K.; Gasser, M.; Waaga-Gasser, A.M.; Choi, A.M.; Pal, S. Heme oxygenase-1 promotes survival of renal cancer cells through modulation of apoptosis- and autophagy-regulating molecules. J Biol Chem 2012, 287, 32113–32123. [Google Scholar] [CrossRef]
- Ding, Y.; Fu, X.; Wang, Q.; Liu, H.; Wang, H.; Wu, D. The Complex Interplay between Autophagy and NLRP3 Inflammasome in Renal Diseases. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Kim, J.W.; Nam, S.A.; Koh, E.S.; Kim, H.W.; Kim, S.; Woo, J.J.; Kim, Y.K. The Impairment of Endothelial Autophagy Accelerates Renal Senescence by Ferroptosis and NLRP3 Inflammasome Signaling Pathways with the Disruption of Endothelial Barrier. Antioxidants (Basel) 2024, 13. [Google Scholar] [CrossRef]
- Jianbing, H.; Xiaotian, L.; Jie, T.; Xueying, C.; Honge, J.; Bo, Z.; Lirong, H.; Lei, Z. The Effect of Allograft Inflammatory Factor-1 on Inflammation, Oxidative Stress, and Autophagy via miR-34a/ATG4B Pathway in Diabetic Kidney Disease. Oxid Med Cell Longev 2022, 2022, 1668000. [Google Scholar] [CrossRef]
- Kato, H.; Gruenwald, A.; Suh, J.H.; Miner, J.H.; Barisoni-Thomas, L.; Taketo, M.M.; Faul, C.; Millar, S.E.; Holzman, L.B.; Susztak, K. Wnt/β-catenin pathway in podocytes integrates cell adhesion, differentiation, and survival. Journal of Biological Chemistry 2011, 286, 26003–26015. [Google Scholar] [CrossRef]
- Fabian, S.L.; Penchev, R.R.; St-Jacques, B.; Rao, A.N.; Sipilä, P.; West, K.A.; McMahon, A.P.; Humphreys, B.D. Hedgehog-Gli pathway activation during kidney fibrosis. The American journal of pathology 2012, 180, 1441–1453. [Google Scholar] [CrossRef] [PubMed]
- White, E.S.; Mantovani, A.R. Inflammation, wound repair, and fibrosis: reassessing the spectrum of tissue injury and resolution. J Pathol 2013, 229, 141–144. [Google Scholar] [CrossRef]
- Kuppe, C.; Ibrahim, M.M.; Kranz, J.; Zhang, X.; Ziegler, S.; Perales-Patón, J.; Jansen, J.; Reimer, K.C.; Smith, J.R.; Dobie, R. Decoding myofibroblast origins in human kidney fibrosis. Nature 2021, 589, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Yanagita, M. Resident fibroblasts in the kidney: a major driver of fibrosis and inflammation. Inflammation and regeneration 2017, 37, 1–7. [Google Scholar] [CrossRef]
- Ferenbach, D.A.; Bonventre, J.V. Mechanisms of maladaptive repair after AKI leading to accelerated kidney ageing and CKD. Nature Reviews Nephrology 2015, 11, 264–276. [Google Scholar] [CrossRef] [PubMed]
- Venkatachalam, M.A.; Weinberg, J.M.; Kriz, W.; Bidani, A.K. Failed tubule recovery, AKI-CKD transition, and kidney disease progression. Journal of the American Society of Nephrology 2015, 26, 1765–1776. [Google Scholar] [CrossRef]
- Basile, D.P.; Bonventre, J.V.; Mehta, R.; Nangaku, M.; Unwin, R.; Rosner, M.H.; Kellum, J.A.; Ronco, C. Progression after AKI: understanding maladaptive repair processes to predict and identify therapeutic treatments. Journal of the American Society of Nephrology 2016, 27, 687–697. [Google Scholar] [CrossRef]
- Kumar, S. Cellular and molecular pathways of renal repair after acute kidney injury. Kidney international 2018, 93, 27–40. [Google Scholar] [CrossRef]
- Humphreys, B.D. Mechanisms of renal fibrosis. Annual review of physiology 2018, 80, 309–326. [Google Scholar] [CrossRef]
- Podestà, M.A.; Faravelli, I.; Ponticelli, C. Autophagy in lupus nephritis: a delicate balance between regulation and disease. Autoimmunity Reviews 2022, 21, 103132. [Google Scholar] [CrossRef]
- KIM, W.Y.; Nam, S.A.; Song, H.C.; Ko, J.S.; Park, S.H.; Kim, H.L.; Choi, E.J.; KIM, Y.S.; Kim, J.; Kim, Y.K. The role of autophagy in unilateral ureteral obstruction rat model. Nephrology 2012, 17, 148–159. [Google Scholar] [CrossRef]
- Ma, Z.; Li, L.; Livingston, M.J.; Zhang, D.; Mi, Q.; Zhang, M.; Ding, H.-F.; Huo, Y.; Mei, C.; Dong, Z. p53/microRNA-214/ULK1 axis impairs renal tubular autophagy in diabetic kidney disease. The Journal of clinical investigation 2020, 130, 5011–5026. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Song, Y.; Zhang, L.; Chen, Z.; Yang, C.; Liu, S.; Yuan, X.; Gao, H.; Ding, G.; Wang, H. Autophagy activation contributes to lipid accumulation in tubular epithelial cells during kidney fibrosis. Cell death discovery 2018, 4, 39. [Google Scholar] [CrossRef]
- Fu, Y.; Xiang, Y.; Wu, W.; Cai, J.; Tang, C.; Dong, Z. Persistent activation of autophagy after cisplatin nephrotoxicity promotes renal fibrosis and chronic kidney disease. Frontiers in Pharmacology 2022, 13, 918732. [Google Scholar] [CrossRef]
- Livingston, M.J.; Ding, H.-F.; Huang, S.; Hill, J.A.; Yin, X.-M.; Dong, Z. Persistent activation of autophagy in kidney tubular cells promotes renal interstitial fibrosis during unilateral ureteral obstruction. Autophagy 2016, 12, 976–998. [Google Scholar] [CrossRef] [PubMed]
- Gui, Z.; Suo, C.; Wang, Z.; Zheng, M.; Fei, S.; Chen, H.; Sun, L.; Han, Z.; Tao, J.; Ju, X. Impaired ATG16L-dependent autophagy promotes renal interstitial fibrosis in chronic renal graft dysfunction through inducing EndMT by NF-κB signal pathway. Frontiers in Immunology 2021, 12, 650424. [Google Scholar] [CrossRef] [PubMed]
- Hetz, C.; Zhang, K.; Kaufman, R.J. Mechanisms, regulation and functions of the unfolded protein response. Nature reviews Molecular cell biology 2020, 21, 421–438. [Google Scholar] [CrossRef]
- Wang, M.; Chen, Z.; Tang, Z.; Tang, S. Natural products derived from traditional Chinese medicines targeting ER stress for the treatment of kidney diseases. Renal Failure 2024, 46, 2396446. [Google Scholar] [CrossRef]
- Hasegawa, S.; Inagi, R. Organelle stress and crosstalk in kidney disease. Kidney360 2020, 1, 1157–1164. [Google Scholar] [CrossRef]
- Fan, Y.; Xiao, W.; Lee, K.; Salem, F.; Wen, J.; He, L.; Zhang, J.; Fei, Y.; Cheng, D.; Bao, H. Inhibition of reticulon-1a–mediated endoplasmic reticulum stress in early AKI attenuates renal fibrosis development. Journal of the American Society of Nephrology 2017, 28, 2007–2021. [Google Scholar] [CrossRef] [PubMed]
- Tavernier, Q.; Mami, I.; Rabant, M.; Karras, A.; Laurent-Puig, P.; Chevet, E.; Thervet, E.; Anglicheau, D.; Pallet, N. Urinary angiogenin reflects the magnitude of kidney injury at the infrahistologic level. Journal of the American Society of Nephrology 2017, 28, 678–690. [Google Scholar] [CrossRef]
- Yan, M.; Shu, S.; Guo, C.; Tang, C.; Dong, Z. Endoplasmic reticulum stress in ischemic and nephrotoxic acute kidney injury. Annals of medicine 2018, 50, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Matsui, T. Molecular mechanisms of macroautophagy, microautophagy, and chaperone-mediated autophagy. Journal of Nippon Medical School 2024, 91, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Aman, Y.; Schmauck-Medina, T.; Hansen, M.; Morimoto, R.I.; Simon, A.K.; Bjedov, I.; Palikaras, K.; Simonsen, A.; Johansen, T.; Tavernarakis, N. Autophagy in healthy aging and disease. Nature aging 2021, 1, 634–650. [Google Scholar] [CrossRef]
- Liu, F.; Zhao, L.; Wu, T.; Yu, W.; Li, J.; Wang, W.; Huang, C.; Diao, Z.; Xu, Y. Targeting autophagy with natural products as a potential therapeutic approach for diabetic microangiopathy. Frontiers in Pharmacology 2024, 15, 1364616. [Google Scholar] [CrossRef]
- Liu, J.; Livingston, M.J.; Dong, G.; Tang, C.; Su, Y.; Wu, G.; Yin, X.-M.; Dong, Z. Histone deacetylase inhibitors protect against cisplatin-induced acute kidney injury by activating autophagy in proximal tubular cells. Cell death & disease 2018, 9, 322. [Google Scholar]
- Tang, C.; Han, H.; Yan, M.; Zhu, S.; Liu, J.; Liu, Z.; He, L.; Tan, J.; Liu, Y.; Liu, H. PINK1-PRKN/PARK2 pathway of mitophagy is activated to protect against renal ischemia-reperfusion injury. Autophagy 2018, 14, 880–897. [Google Scholar] [CrossRef]
- Kaufman, D.R.; Papillon, J.; Larose, L.; Iwawaki, T.; Cybulsky, A.V. Deletion of inositol-requiring enzyme-1α in podocytes disrupts glomerular capillary integrity and autophagy. Molecular biology of the cell 2017, 28, 1636–1651. [Google Scholar] [CrossRef]
- Rashid, H.-O.; Yadav, R.K.; Kim, H.-R.; Chae, H.-J. ER stress: Autophagy induction, inhibition and selection. Autophagy 2015, 11, 1956–1977. [Google Scholar] [CrossRef] [PubMed]
- Hartleben, B.; Gödel, M.; Meyer-Schwesinger, C.; Liu, S.; Ulrich, T.; Köbler, S.; Wiech, T.; Grahammer, F.; Arnold, S.J.; Lindenmeyer, M.T. Autophagy influences glomerular disease susceptibility and maintains podocyte homeostasis in aging mice. The Journal of clinical investigation 2010, 120, 1084–1096. [Google Scholar] [CrossRef] [PubMed]
- Cybulsky, A.V. The intersecting roles of endoplasmic reticulum stress, ubiquitin–proteasome system, and autophagy in the pathogenesis of proteinuric kidney disease. Kidney international 2013, 84, 25–33. [Google Scholar] [CrossRef]
- Ruby, M.; Gifford, C.C.; Pandey, R.; Raj, V.S.; Sabbisetti, V.S.; Ajay, A.K. Autophagy as a Therapeutic Target for Chronic Kidney Disease and the Roles of TGF-β1 in Autophagy and Kidney Fibrosis. Cells 2023, 12. [Google Scholar] [CrossRef]
- Zhang, Z.; Sun, Y.; Xue, J.; Jin, D.; Li, X.; Zhao, D.; Lian, F.; Qi, W.; Tong, X. The critical role of dysregulated autophagy in the progression of diabetic kidney disease. Front Pharmacol 2022, 13, 977410. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Ma, Y.; Chen, X.; Liang, N.; Qu, S.; Chen, H. Hyperuricemia causes kidney damage by promoting autophagy and NLRP3-mediated inflammation in rats with urate oxidase deficiency. Dis Model Mech 2021, 14. [Google Scholar] [CrossRef]
- Wu, M.; Zhang, M.; Zhang, Y.; Li, Z.; Li, X.; Liu, Z.; Liu, H.; Li, X. Relationship between lysosomal dyshomeostasis and progression of diabetic kidney disease. Cell Death Dis 2021, 12, 958. [Google Scholar] [CrossRef]
- Yuan, Q.; Tang, B.; Zhang, C. Signaling pathways of chronic kidney diseases, implications for therapeutics. Signal Transduct Target Ther 2022, 7, 182. [Google Scholar] [CrossRef]
- Wang, H.; Gao, L.; Zhao, C.; Fang, F.; Liu, J.; Wang, Z.; Zhong, Y.; Wang, X. The role of PI3K/Akt signaling pathway in chronic kidney disease. Int Urol Nephrol 2024, 56, 2623–2633. [Google Scholar] [CrossRef]
- Miricescu, D.; Balan, D.G.; Tulin, A.; Stiru, O.; Vacaroiu, I.A.; Mihai, D.A.; Popa, C.C.; Papacocea, R.I.; Enyedi, M.; Sorin, N.A.; et al. PI3K/AKT/mTOR signalling pathway involvement in renal cell carcinoma pathogenesis (Review). Exp Ther Med 2021, 21, 540. [Google Scholar] [CrossRef]
- Muratsubaki, S.; Kuno, A.; Tanno, M.; Miki, T.; Yano, T.; Sugawara, H.; Shibata, S.; Abe, K.; Ishikawa, S.; Ohno, K.; et al. Suppressed autophagic response underlies augmentation of renal ischemia/reperfusion injury by type 2 diabetes. Scientific Reports 2017, 7, 5311. [Google Scholar] [CrossRef]
- R, H.; Y, X.; W, W.; X, S.; J, Q.; Z, Y.; B, L.; C, C.; T, Z.; J, L.-S.; et al. Deacetylation of nuclear LC3 drives autophagy initiation under starvation - PubMed. Molecular cell 02/05/2015, 57. [Google Scholar] [CrossRef]
- Sun, M.; Li, J.; Mao, L.; Wu, J.; Deng, Z.; He, M.; An, S.; Zeng, Z.; Huang, Q.; Chen, Z. p53 Deacetylation Alleviates Sepsis-Induced Acute Kidney Injury by Promoting Autophagy. Front Immunol 2021, 12, 685523. [Google Scholar] [CrossRef]
- Deng, J.K.; Zhang, X.; Wu, H.L.; Gan, Y.; Ye, L.; Zheng, H.; Zhu, Z.; Liu, W.J.; Liu, H.F. ROS-ERK Pathway as Dual Mediators of Cellular Injury and Autophagy-Associated Adaptive Response in Urinary Protein-Irritated Renal Tubular Epithelial Cells. J Diabetes Res 2021, 2021, 6614848. [Google Scholar] [CrossRef]
- Yang, J.; Yuan, L.; Liu, F.; Li, L.; Liu, J.; Chen, Y.; Lu, Y.; Yuan, Y. Molecular mechanisms and physiological functions of autophagy in kidney diseases. Front Pharmacol 2022, 13, 974829. [Google Scholar] [CrossRef]
- Wang, T.; Liu, C.; Jia, L. The roles of PKCs in regulating autophagy. J Cancer Res Clin Oncol 2018, 144, 2303–2311. [Google Scholar] [CrossRef]
- Sanz, A.B.; Sanchez-Niño, M.D.; Ramos, A.M.; Ortiz, A. Regulated cell death pathways in kidney disease. Nat Rev Nephrol 2023, 19, 281–299. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Mou, L.; Yi, Z.; Lin, Q.; Banu, K.; Wei, C.; Yu, X. Integrative informatics analysis identifies that ginsenoside Re improves renal fibrosis through regulation of autophagy. J Nat Med 2024, 78, 722–731. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Jin, Q.; Yang, L.; Mao, H.; Ma, F.; Wang, Y.; Li, P.; Zhan, Y. Regulation of autophagy by natural polyphenols in the treatment of diabetic kidney disease: therapeutic potential and mechanism. Front Endocrinol (Lausanne) 2023, 14, 1142276. [Google Scholar] [CrossRef]
- Jia, Z.; Wang, K.; Zhang, Y.; Duan, Y.; Xiao, K.; Liu, S.; Ding, X. Icariin Ameliorates Diabetic Renal Tubulointerstitial Fibrosis by Restoring Autophagy via Regulation of the miR-192-5p/GLP-1R Pathway. Front Pharmacol 2021, 12, 720387. [Google Scholar] [CrossRef]
- Matboli, M.; Ibrahim, D.; Hasanin, A.H.; Hassan, M.K.; Habib, E.K.; Bekhet, M.M.; Afifi, A.M.; Eissa, S. Epigenetic modulation of autophagy genes linked to diabetic nephropathy by administration of isorhamnetin in Type 2 diabetes mellitus rats. Epigenomics 2021, 13, 187–202. [Google Scholar] [CrossRef]
- Ou, X.; Yu, Z.; Pan, C.; Zheng, X.; Li, D.; Qiao, Z.; Zheng, X. Paeoniflorin: a review of its pharmacology, pharmacokinetics and toxicity in diabetes. Frontiers in Pharmacology 2025, 16–2025. [Google Scholar] [CrossRef]
- Zhang, X.X.; Jiang, C.H.; Liu, Y.; Lou, D.X.; Huang, Y.P.; Gao, M.; Zhang, J.; Yin, Z.Q.; Pan, K. Cyclocarya paliurus triterpenic acids fraction attenuates kidney injury via AMPK-mTOR-regulated autophagy pathway in diabetic rats. Phytomedicine 2019, 64, 153060. [Google Scholar] [CrossRef]
- Teh, Y.M.; Mualif, S.A.; Mohd Noh, N.I.; Lim, S.K. The Potential of Naturally Derived Compounds for Treating Chronic Kidney Disease: A Review of Autophagy and Cellular Senescence. Int J Mol Sci 2024, 26. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Zhang, H.; Lv, S.; Zhu, C.; Gong, S.; Yu, X.; Wang, Y.; Huang, X.; Yuan, S.; Ding, X.; et al. Sulforaphane alleviates renal fibrosis through dual regulation on mTOR-mediated autophagy pathway. Int Urol Nephrol 2025, 57, 1277–1287. [Google Scholar] [CrossRef] [PubMed]
- Akaras, N.; Gur, C.; Kucukler, S.; Kandemir, F.M. Zingerone reduces sodium arsenite-induced nephrotoxicity by regulating oxidative stress, inflammation, apoptosis and histopathological changes. Chem Biol Interact 2023, 374, 110410. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Fan, Q.; Wang, X.; Li, L.; Lu, X.; Yue, Y.; Cao, X.; Liu, J.; Zhao, X.; Wang, L. Ursolic acid improves podocyte injury caused by high glucose. Nephrol Dial Transplant 2017, 32, 1285–1293. [Google Scholar] [CrossRef]
- Turgut, F.; Awad, A.S.; Abdel-Rahman, E.M. Acute Kidney Injury: Medical Causes and Pathogenesis. J Clin Med 2023, 12. [Google Scholar] [CrossRef]
- Zeng, Y.; Li, S.; Wu, J.; Chen, W.; Sun, H.; Peng, W.; Yu, X.; Yang, X. Autophagy inhibitors promoted aristolochic acid I induced renal tubular epithelial cell apoptosis via mitochondrial pathway but alleviated nonapoptotic cell death in mouse acute aritolochic acid nephropathy model. Apoptosis 2014, 19, 1215–1224. [Google Scholar] [CrossRef]
- Tan, X.; Zhu, H.; Tao, Q.; Guo, L.; Jiang, T.; Xu, L.; Yang, R.; Wei, X.; Wu, J.; Li, X.; et al. FGF10 Protects Against Renal Ischemia/Reperfusion Injury by Regulating Autophagy and Inflammatory Signaling. Front Genet 2018, 9, 556. [Google Scholar] [CrossRef]
- Kaushal, G.P.; Shah, S.V. Autophagy in acute kidney injury. Kidney international 2016, 89, 779–791. [Google Scholar] [CrossRef]
- Djavaheri-Mergny, M.; Amelotti, M.; Mathieu, J.; Besançon, F.; Bauvy, C.; Codogno, P. Regulation of autophagy by NFkappaB transcription factor and reactives oxygen species. Autophagy 2007, 3, 390–392. [Google Scholar] [CrossRef] [PubMed]
- Vijayan, A. Tackling AKI: prevention, timing of dialysis and follow-up. Nature Reviews Nephrology 2020, 17. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wang, Q.; Li, T.; Li, L.; Wang, C.; Xu, B.; Gong, X. Exploring therapeutic mechanisms of Chuan Huang Fang-II in the treatment of acute kidney injury on chronic kidney disease patients from the perspective of lipidomics. Ren Fail 2024, 46, 2356021. [Google Scholar] [CrossRef]
- Shi, Y.; Shi, X.; Zhao, M.; Chang, M.; Ma, S.; Zhang, Y. Ferroptosis: A new mechanism of traditional Chinese medicine compounds for treating acute kidney injury. Biomed Pharmacother 2023, 163, 114849. [Google Scholar] [CrossRef]
- Qi, J.; Xue, Q.; Kuang, L.; Xie, L.; Luo, R.; Nie, X. Berberine alleviates cisplatin-induced acute kidney injury by regulating mitophagy via PINK 1/Parkin pathway. Transl Androl Urol 2020, 9, 1712–1724. [Google Scholar] [CrossRef]
- Gong, X.; Duan, Y.; Zheng, J.; Ye, Z.; Hei, T.K. Tetramethylpyrazine Prevents Contrast-Induced Nephropathy via Modulating Tubular Cell Mitophagy and Suppressing Mitochondrial Fragmentation, CCL2/CCR2-Mediated Inflammation, and Intestinal Injury. Oxid Med Cell Longev 2019, 2019, 7096912. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Xu, B.; Sun, Y.; Lian, M.; Li, Y.; Lin, Y.; Chen, D.; Diao, Y.; Almoiliqy, M.; Wang, L. Paeoniflorin protects against intestinal ischemia/reperfusion by activating LKB1/AMPK and promoting autophagy. Pharmacol Res 2019, 146, 104308. [Google Scholar] [CrossRef]
- Li, H.; Tang, Y.; Wen, L.; Kong, X.; Chen, X.; Liu, P.; Zhou, Z.; Chen, W.; Xiao, C.; Xiao, P.; et al. Neferine reduces cisplatin-induced nephrotoxicity by enhancing autophagy via the AMPK/mTOR signaling pathway. Biochem Biophys Res Commun 2017, 484, 694–701. [Google Scholar] [CrossRef]
- Zuo, Z.; Li, Q.; Zhou, S.; Yu, R.; Wu, C.; Chen, J.; Xiao, Y.; Chen, H.; Song, J.; Pan, Y.; et al. Berberine ameliorates contrast-induced acute kidney injury by regulating HDAC4-FoxO3a axis-induced autophagy: In vivo and in vitro. Phytother Res 2024, 38, 1761–1780. [Google Scholar] [CrossRef]
- Kalantar-Zadeh, K.; Jafar, T.H.; Nitsch, D.; Neuen, B.L.; Perkovic, V. Chronic kidney disease. The lancet 2021, 398, 786–802. [Google Scholar] [CrossRef] [PubMed]
- Levin, A.; Okpechi, I.G.; Caskey, F.J.; Yang, C.-W.; Tonelli, M.; Jha, V. Perspectives on early detection of chronic kidney disease: the facts, the questions, and a proposed framework for 2023 and beyond. Kidney International 2023, 103, 1004–1008. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Choi, M.E. Autophagy in diabetic nephropathy. The Journal of endocrinology 2014, 224, R15. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.-A.; Wu, V.C.-C.; Wang, C.-Y. Autophagy in chronic kidney diseases. Cells 2019, 8, 61. [Google Scholar] [CrossRef]
- Periyasamy-Thandavan, S.; Jiang, M.; Schoenlein, P.; Dong, Z. Autophagy: molecular machinery, regulation, and implications for renal pathophysiology. American Journal of Physiology-Renal Physiology 2009, 297, F244–F256. [Google Scholar] [CrossRef]
- Di Tu, Q.; Jin, J.; Hu, X.; Ren, Y.; Zhao, L.; He, Q. Curcumin improves the renal autophagy in rat experimental membranous nephropathy via regulating the PI3K/AKT/mTOR and Nrf2/HO-1 signaling pathways. BioMed research international 2020, 2020, 7069052. [Google Scholar] [CrossRef]
- Liu, X.; Chen, J.; Sun, N.; Li, N.; Zhang, Z.; Zheng, T.; Li, Z. Ginsenoside Rb1 ameliorates autophagy via the AMPK/mTOR pathway in renal tubular epithelial cells in vitro and in vivo. International Journal of Biological Macromolecules 2020, 163, 996–1009. [Google Scholar] [CrossRef]
- Lu, R.; Chen, J.; Liu, B.; Lin, H.; Bai, L.; Zhang, P.; Chen, D.; Li, H.; Li, J.; Pang, Y. Protective role of Astragaloside IV in chronic glomerulonephritis by activating autophagy through PI3K/AKT/AS160 pathway. Phytotherapy Research 2020, 34, 3236–3248. [Google Scholar] [CrossRef]
- Tseng, C.-Y.; Yu, P.-R.; Hsu, C.-C.; Lin, H.-H.; Chen, J.-H. The effect of isovitexin on lipopolysaccharide-induced renal injury and inflammation by induction of protective autophagy. Food and Chemical Toxicology 2023, 172, 113581. [Google Scholar] [CrossRef]
- Yang, F.; Gao, Y.; Xie, S.; Yang, W.; Wang, Q.; Ye, W.; Sun, L.; Zhou, J.; Feng, X. Dietary phytosterol supplementation mitigates renal fibrosis via activating mitophagy and modulating the gut microbiota. Food & Function 2025, 16, 2316–2334. [Google Scholar] [CrossRef]
- Fan, L.; Chen, H.-h.; Liu, H.-j.; Chen, H.-j.; Zhu, L.-l.; Zhang, T. Experimental study on danggui shaoyao san improving renal fibrosis by promoting autophagy. Evidence-Based Complementary and Alternative Medicine 2022, 2022, 6761453. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.-J.; Lee, Y.-H.; Yeh, Y.-L.; Wu, W.-S.; Ho, C.-T.; Li, C.-Y.; Wang, B.-J.; Wang, Y.-J. Autophagy-inducing effect of pterostilbene: A prospective therapeutic/preventive option for skin diseases. journal of food and drug analysis 2017, 25, 125–133. [Google Scholar] [CrossRef]
- Zhang, D.; Zhang, H.; Lv, S.; Zhu, C.; Gong, S.; Yu, X.; Wang, Y.; Huang, X.; Yuan, S.; Ding, X. Sulforaphane alleviates renal fibrosis through dual regulation on mTOR-mediated autophagy pathway. International Urology and Nephrology 2025, 57, 1277–1287. [Google Scholar] [CrossRef]
- Tu, Y.; Gu, L.; Chen, D.; Wu, W.; Liu, H.; Hu, H.; Wan, Y.; Sun, W. Rhein inhibits autophagy in rat renal tubular cells by regulation of AMPK/mTOR signaling. Scientific reports 2017, 7, 43790. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Qin, X.; Xu, W.; Zhang, H.; Huang, S.; Yang, Y.; Qin, M.; Mi, Z.; Zhong, X. Herb pair of Rhubarb-Astragalus mitigates renal interstitial fibrosis through downregulation of autophagy via p38-MAPK/TGF-β1 and p38-MAPK/smad2/3 pathways. The International Journal of Biochemistry & Cell Biology 2024, 169, 106549. [Google Scholar]
- Kounatidis, D.; Vallianou, N.G.; Stratigou, T.; Voukali, M.; Karampela, I.; Dalamaga, M. The Kidney in Obesity: Current Evidence, Perspectives and Controversies. Curr Obes Rep 2024, 13, 680–702. [Google Scholar] [CrossRef]
- Martínez-Montoro, J.I.; Morales, E.; Cornejo-Pareja, I.; Tinahones, F.J.; Fernández-García, J.C. Obesity-related glomerulopathy: Current approaches and future perspectives. Obes Rev 2022, 23, e13450. [Google Scholar] [CrossRef]
- Hao, M.; Lv, Y.; Liu, S.; Guo, W. The New Challenge of Obesity - Obesity-Associated Nephropathy. Diabetes Metab Syndr Obes 2024, 17, 1957–1971. [Google Scholar] [CrossRef] [PubMed]
- Yau, K.; Kuah, R.; Cherney, D.Z.I.; Lam, T.K.T. Obesity and the kidney: mechanistic links and therapeutic advances. Nat Rev Endocrinol 2024, 20, 321–335. [Google Scholar] [CrossRef]
- Yao, E.; Yang, X.; Huang, X.; Mi, Y.; Wu, X.; Fang, M.; Huang, J.; Qiu, Y.; Hong, X.; Peng, L.; et al. Phytochemical wedelolactone reverses obesity by prompting adipose browning through SIRT1/AMPK/ PPARα pathway via targeting nicotinamide N-methyltransferase. Phytomedicine 2022, 94, 153843. [Google Scholar] [CrossRef]
- Axelsson, A.S.; Tubbs, E.; Mecham, B.; Chacko, S.; Nenonen, H.A.; Tang, Y.; Fahey, J.W.; Derry, J.M.J.; Wollheim, C.B.; Wierup, N.; et al. Sulforaphane reduces hepatic glucose production and improves glucose control in patients with type 2 diabetes. Sci Transl Med 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Bian, Z.; Zhang, M. Targeting the Nrf2 signaling pathway using phytochemical ingredients: A novel therapeutic road map to combat neurodegenerative diseases. Phytomedicine 2023, 109, 154582. [Google Scholar] [CrossRef]
- Mafra, D.; Borges, N.A.; Lindholm, B.; Shiels, P.G.; Evenepoel, P.; Stenvinkel, P. Food as medicine: targeting the uraemic phenotype in chronic kidney disease. Nat Rev Nephrol 2021, 17, 153–171. [Google Scholar] [CrossRef] [PubMed]
- Mykkänen, O.T.; Huotari, A.; Herzig, K.H.; Dunlop, T.W.; Mykkänen, H.; Kirjavainen, P.V. Wild blueberries (Vaccinium myrtillus) alleviate inflammation and hypertension associated with developing obesity in mice fed with a high-fat diet. PLoS One 2014, 9, e114790. [Google Scholar] [CrossRef] [PubMed]
- Roopchand, D.E.; Kuhn, P.; Rojo, L.E.; Lila, M.A.; Raskin, I. Blueberry polyphenol-enriched soybean flour reduces hyperglycemia, body weight gain and serum cholesterol in mice. Pharmacol Res 2013, 68, 59–67. [Google Scholar] [CrossRef]
- Du, J.; Shen, L.; Tan, Z.; Zhang, P.; Zhao, X.; Xu, Y.; Gan, M.; Yang, Q.; Ma, J.; Jiang, A.; et al. Betaine Supplementation Enhances Lipid Metabolism and Improves Insulin Resistance in Mice Fed a High-Fat Diet. Nutrients 2018, 10. [Google Scholar] [CrossRef]
- Mafra, D.; Esgalhado, M.; Borges, N.A.; Cardozo, L.; Stockler-Pinto, M.B.; Craven, H.; Buchanan, S.J.; Lindholm, B.; Stenvinkel, P.; Shiels, P.G. Methyl Donor Nutrients in Chronic Kidney Disease: Impact on the Epigenetic Landscape. J Nutr 2019, 149, 372–380. [Google Scholar] [CrossRef]
- Bahrampour, N.; Mirzababaei, A.; Hosseininasab, D.; Abaj, F.; Clark, C.C.T.; Mirzaei, K. High intake of dietary phytochemical index may be related to reducing risk of diabetic nephropathy: a case-control study. BMC Nutr 2023, 9, 14. [Google Scholar] [CrossRef]
- Urmila, A.; Armash, M.; Pavan, S.; Rachana, N.; Patil, R. Anti-Inflammatory phytochemicals for the treatment of diabetic nephropathy. Current Functional Foods 2023. [Google Scholar]
- Moreira, L.S.G.; Brum, I.; de Vargas Reis, D.C.M.; Trugilho, L.; Chermut, T.R.; Esgalhado, M.; Cardozo, L.; Stenvinkel, P.; Shiels, P.G.; Mafra, D. Cinnamon: an aromatic condiment applicable to chronic kidney disease. Kidney Res Clin Pract 2023, 42, 4–26. [Google Scholar] [CrossRef]
- Dinda, B.; Chakraborty, A. Pharmacokinetics and Metabolism of Phytochemicals Having Anti-obesity and Antidiabetic Activity; 2022; pp. 469–505. [Google Scholar]
- Wang, W.; Sun, W.; Cheng, Y.; Xu, Z.; Cai, L. Role of sirtuin-1 in diabetic nephropathy. J Mol Med (Berl) 2019, 97, 291–309. [Google Scholar] [CrossRef]
- Kim, H.; Dusabimana, T.; Kim, S.R.; Je, J.; Jeong, K.; Kang, M.C.; Cho, K.M.; Kim, H.J.; Park, S.W. Supplementation of Abelmoschus manihot Ameliorates Diabetic Nephropathy and Hepatic Steatosis by Activating Autophagy in Mice. Nutrients 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Sagoo, M.K.; Gnudi, L. Diabetic Nephropathy: An Overview. Methods Mol Biol 2020, 2067, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, J.A.; Shankland, S.J.; Pichler, R.H. Proteinuria in diabetic kidney disease: a mechanistic viewpoint. Kidney Int 2008, 74, 22–36. [Google Scholar] [CrossRef]
- Kume, S.; Koya, D. Autophagy: A Novel Therapeutic Target for Diabetic Nephropathy. Diabetes Metab J 2015, 39, 451–460. [Google Scholar] [CrossRef]
- Kume, S.; Koya, D.; Uzu, T.; Maegawa, H. Role of nutrient-sensing signals in the pathogenesis of diabetic nephropathy. Biomed Res Int 2014, 2014, 315494. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Kim, S.; Lee, S.Y.; Koo, J.K.; Wang, Z.; Choi, M.E. Autophagy regulates TGF-β expression and suppresses kidney fibrosis induced by unilateral ureteral obstruction. J Am Soc Nephrol 2014, 25, 2835–2846. [Google Scholar] [CrossRef]
- Kume, S.; Yamahara, K.; Yasuda, M.; Maegawa, H.; Koya, D. Autophagy: emerging therapeutic target for diabetic nephropathy. Semin Nephrol 2014, 34, 9–16. [Google Scholar] [CrossRef]
- Jin, Y.; Liu, S.; Ma, Q.; Xiao, D.; Chen, L. Berberine enhances the AMPK activation and autophagy and mitigates high glucose-induced apoptosis of mouse podocytes. Eur J Pharmacol 2017, 794, 106–114. [Google Scholar] [CrossRef]
- Wei, Y.; Gao, J.; Qin, L.; Xu, Y.; Shi, H.; Qu, L.; Liu, Y.; Xu, T.; Liu, T. Curcumin suppresses AGEs induced apoptosis in tubular epithelial cells via protective autophagy. Exp Ther Med 2017, 14, 6052–6058. [Google Scholar] [CrossRef]
- Ezzat, S.M.; Abdallah, H.M.I.; Yassen, N.N.; Radwan, R.A.; Mostafa, E.S.; Salama, M.M.; Salem, M.A. Phenolics from Physalis peruviana fruits ameliorate streptozotocin-induced diabetes and diabetic nephropathy in rats via induction of autophagy and apoptosis regression. Biomed Pharmacother 2021, 142, 111948. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Han, X.; Wen, Q.; Qiu, X.; Deng, H.; Chen, Q. Protective Effect of Keluoxin against Diabetic Nephropathy in Type 2 Diabetic Mellitus Models. Evid Based Complement Alternat Med 2021, 2021, 8455709. [Google Scholar] [CrossRef]
- Chen, Y.; Zheng, Y.F.; Lin, X.H.; Zhang, J.P.; Lin, F.; Shi, H. Dendrobium mixture attenuates renal damage in rats with diabetic nephropathy by inhibiting the PI3K/Akt/mTOR pathway. Mol Med Rep 2021, 24. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, W.; Zhang, Z.; Hu, Y.; Zhang, X.; Sun, Y.; Lei, Q.; Sun, D.; Liu, T.; Fan, Y.; et al. Yishen capsule promotes podocyte autophagy through regulating SIRT1/NF-κB signaling pathway to improve diabetic nephropathy. Ren Fail 2021, 43, 128–140. [Google Scholar] [CrossRef]
- Amanat, M.; Lal, K.; Singh, T.G.; Singh, R. Molecular insights of diabetic nephropathy and chemical constituents-based treatment approach. In Phytochemistry Reviews; 2025. [Google Scholar] [CrossRef]
- Zhan, X.; Yan, C.; Chen, Y.; Wei, X.; Xiao, J.; Deng, L.; Yang, Y.; Qiu, P.; Chen, Q. Celastrol antagonizes high glucose-evoked podocyte injury, inflammation and insulin resistance by restoring the HO-1-mediated autophagy pathway. Mol Immunol 2018, 104, 61–68. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, H.; Wang, Q.; Zhou, X.; Lu, X.; Liu, T.; Zhan, Y.; Li, P. Chinese Herbal Medicine in Ameliorating Diabetic Kidney Disease via Activating Autophagy. J Diabetes Res 2019, 2019, 9030893. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Gao, Y.; Tian, N.; Zhu, Z.; Wang, T.; Xu, J.; Wu, B.; Zhang, N. Astragaloside IV represses high glucose-induced mesangial cells activation by enhancing autophagy via SIRT1 deacetylation of NF-κB p65 subunit. Drug Des Devel Ther 2018, 12, 2971–2980. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.H.; Ding, D.F.; Yong, H.J.; Dong, C.L.; You, N.; Ye, X.L.; Pan, M.L.; Ma, J.H.; You, Q.; Lu, Y.B. Resveratrol transcriptionally regulates miRNA-18a-5p expression ameliorating diabetic nephropathy via increasing autophagy. Eur Rev Med Pharmacol Sci 2017, 21, 4952–4965. [Google Scholar]
- Huang, S.S.; Ding, D.F.; Chen, S.; Dong, C.L.; Ye, X.L.; Yuan, Y.G.; Feng, Y.M.; You, N.; Xu, J.R.; Miao, H.; et al. Resveratrol protects podocytes against apoptosis via stimulation of autophagy in a mouse model of diabetic nephropathy. Sci Rep 2017, 7, 45692. [Google Scholar] [CrossRef]
- Al Shahrani, M.; Chandramoorthy, H.C.; Alshahrani, M.; Abohassan, M.; Eid, R.A.; Ravichandran, K.; Rajagopalan, P. Cassia auriculata leaf extract ameliorates diabetic nephropathy by attenuating autophagic necroptosis via RIP-1/RIP-3-p-p38MAPK signaling. J Food Biochem 2021, 45, e13810. [Google Scholar] [CrossRef]
- Yang, F.; Qu, Q.; Zhao, C.; Liu, X.; Yang, P.; Li, Z.; Han, L.; Shi, X. Paecilomyces cicadae-fermented Radix astragali activates podocyte autophagy by attenuating PI3K/AKT/mTOR pathways to protect against diabetic nephropathy in mice. Biomed Pharmacother 2020, 129, 110479. [Google Scholar] [CrossRef]
- Han, J.; Zhang, Y.; Shi, X.; Peng, Z.; Xing, Y.; Pang, X. Tongluo Digui decoction treats renal injury in diabetic rats by promoting autophagy of podocytes. J Tradit Chin Med 2021, 41, 125–132. [Google Scholar] [CrossRef]
- Zhang, Y.; Arzaghi, H.; Ma, Z.; Roye, Y.; Musah, S. Epigenetics of Hypertensive Nephropathy. Biomedicines 2024, 12. [Google Scholar] [CrossRef] [PubMed]
- Pitzer, A.; Elijovich, F.; Laffer, C.L.; Ertuglu, L.A.; Sahinoz, M.; Saleem, M.; Krishnan, J.; Dola, T.; Aden, L.A.; Sheng, Q.; et al. DC ENaC-Dependent Inflammasome Activation Contributes to Salt-Sensitive Hypertension. Circ Res 2022, 131, 328–344. [Google Scholar] [CrossRef]
- Bidani, A.K.; Griffin, K.A. Pathophysiology of hypertensive renal damage: implications for therapy. Hypertension 2004, 44, 595–601. [Google Scholar] [CrossRef]
- Laffer, C.L.; Elijovich, F.; Sahinoz, M.; Pitzer, A.; Kirabo, A. New Insights Into the Renin-Angiotensin System in Chronic Kidney Disease. Circ Res 2020, 127, 607–609. [Google Scholar] [CrossRef]
- Ngowi, E.E.; Sarfraz, M.; Afzal, A.; Khan, N.H.; Khattak, S.; Zhang, X.; Li, T.; Duan, S.F.; Ji, X.Y.; Wu, D.D. Roles of Hydrogen Sulfide Donors in Common Kidney Diseases. Front Pharmacol 2020, 11, 564281. [Google Scholar] [CrossRef]
- Hackfort, B.T.; Mishra, P.K. Emerging role of hydrogen sulfide-microRNA crosstalk in cardiovascular diseases. Am J Physiol Heart Circ Physiol 2016, 310, H802–812. [Google Scholar] [CrossRef] [PubMed]
- Weber, G.J.; Pushpakumar, S.B.; Sen, U. Hydrogen sulfide alleviates hypertensive kidney dysfunction through an epigenetic mechanism. Am J Physiol Heart Circ Physiol 2017, 312, H874–h885. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Ren, M.; Fan, D.; Mao, Y.; Zhu, Y.Z. Identification and Validation of the miR/RAS/RUNX2 Autophagy Regulatory Network in AngII-Induced Hypertensive Nephropathy in MPC5 Cells Treated with Hydrogen Sulfide Donors. Antioxidants (Basel) 2024, 13. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Hu, T.; Gao, B.; Xia, R.; Dai, X.; Wang, J. Network pharmacology and in vitro experiments reveal the autophagy mechanism of Yanggan-Yishui granules in improving hypertensive renal injury. Archives of Medical Science 2024. [Google Scholar] [CrossRef]
- Klahr, S. Obstructive nephropathy. Intern Med 2000, 39, 355–361. [Google Scholar] [CrossRef]
- Chevalier, R.L.; Cachat, F. Role of Angiotensin II in Chronic Ureteral Obstruction. Contributions to nephrology 2001, 135, 250–260. [Google Scholar] [CrossRef]
- Ichikawa, I.; Kuwayama, F.; Pope, J.C.t.; Stephens, F.D.; Miyazaki, Y. Paradigm shift from classic anatomic theories to contemporary cell biological views of CAKUT. Kidney Int 2002, 61, 889–898. [Google Scholar] [CrossRef]
- Bander, S.J.; Buerkert, J.E.; Martin, D.; Klahr, S. Long-term effects of 24-hr unilateral ureteral obstruction on renal function in the rat. Kidney Int 1985, 28, 614–620. [Google Scholar] [CrossRef]
- Chevalier, R.L.; Forbes, M.S.; Thornhill, B.A. Ureteral obstruction as a model of renal interstitial fibrosis and obstructive nephropathy. Kidney Int 2009, 75, 1145–1152. [Google Scholar] [CrossRef]
- Li, L.; Zepeda-Orozco, D.; Black, R.; Lin, F. Autophagy is a component of epithelial cell fate in obstructive uropathy. Am J Pathol 2010, 176, 1767–1778. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Ru, F.; Chen, X.; Chen, Z. Autophagy attenuates renal fibrosis in obstructive nephropathy through inhibiting epithelial-to-mesenchymal transition. Zhong Nan Da Xue Xue Bao Yi Xue Ban 2021, 46, 601–608. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, Z.; Li, B.; Zhang, B.; Du, Y.; Liu, Y.; He, Y.; Chen, X. Curcumin attenuates renal interstitial fibrosis of obstructive nephropathy by suppressing epithelial-mesenchymal transition through inhibition of the TLR4/NF-кB and PI3K/AKT signalling pathways. Pharm Biol 2020, 58, 828–837. [Google Scholar] [CrossRef]
- Liu, Y.-F.; Wang, H.-H.; Geng, Y.-H.; Han, L.; Tu, S.-H.; Wang, H. Advances of berberine against metabolic syndrome-associated kidney disease: Regarding effect and mechanism. Frontiers in Pharmacology 2023, 14–2023. [Google Scholar] [CrossRef]
- Malhotra, B.; Kulkarni, G.T.; Dhiman, N.; Joshi, D.D.; Chander, S.; Kharkwal, A.; Sharma, A.K.; Kharkwal, H. Recent advances on Berberis aristata emphasizing berberine alkaloid including phytochemistry, pharmacology and drug delivery system. Journal of Herbal Medicine 2021, 27, 100433. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Y.-s.; Chang, Z.-m.; Li, L.; Zhang, Y.; Lu, M.-m.; Zheng, X.; Li, M.; Shao, D.; Li, J.; et al. Berberine-loaded Janus nanocarriers for magnetic field-enhanced therapy against hepatocellular carcinoma. Chem Biol Drug Des 2017, 89, 464–469. [Google Scholar] [CrossRef]
- Lu, W.-B.; Chen, W.-W.; Li, R.-L. Determination of berberine chloride in mouse plasma after intragastric administration of Lianxiang microemulsion. Zhong yao cai = Zhongyaocai = Journal of Chinese medicinal materials 2010, 33, 957–960. [Google Scholar]
- Mei, D.; Mao, S.; Sun, W.; Wang, Y.; Kissel, T. Effect of chitosan structure properties and molecular weight on the intranasal absorption of tetramethylpyrazine phosphate in rats. European Journal of Pharmaceutics and Biopharmaceutics 2008, 70, 874–881. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, G.; Cui, W.; Zhang, Y.; Liang, X. Regulatory mechanisms of tetramethylpyrazine on central nervous system diseases: A review. Frontiers in Pharmacology 2022, 13–2022. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; Cheng, Z.; Cheng, Y.; Xu, Y. Investigating the passage of tetramethylpyrazine-loaded liposomes across blood-brain barrier models in vitro and ex vivo. Materials Science and Engineering: C 2016, 69, 1010–1017. [Google Scholar] [CrossRef]
- Li, J.; Wei, J.; Wan, Y.; Du, X.; Bai, X.; Li, C.; Lin, Y.; Liu, Z.; Zhou, M.; Zhong, Z. TAT-modified tetramethylpyrazine-loaded nanoparticles for targeted treatment of spinal cord injury. Journal of Controlled Release 2021, 335, 103–116. [Google Scholar] [CrossRef] [PubMed]
- TAN Zuo, H.W. Effects of neferine nanoliposomes on cerebral anoxia and cerebral embolism in mice. Journal of Guangdong College of Pharmacy 2010, 377–379. [Google Scholar]
- Chen, Y.-Q.; Chen, H.-Y.; Tang, Q.-Q.; Li, Y.-F.; Liu, X.-S.; Lu, F.-H.; Gu, Y.-Y. Protective effect of quercetin on kidney diseases: From chemistry to herbal medicines. Frontiers in Pharmacology 2022, 13–2022. [Google Scholar] [CrossRef]
- Cai, T.; Wang, X.; Li, B.; Xiong, F.; Wu, H.; Yang, X. Deciphering the synergistic network regulation of active components from SiNiSan against irritable bowel syndrome via a comprehensive strategy: Combined effects of synephrine, paeoniflorin and naringin. Phytomedicine 2021, 86, 153527. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Fang, C.; Jiang, M.J. The therapeutic effect of Berberine on rodent models of kidney injury: a systematic review and meta - analysis. BMC Nephrology 2025, 26, 315. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Yan, Y. Therapeutic potential of sulforaphane in liver diseases: a review. Front. Pharmacol. 2023, 14–2023. [Google Scholar] [CrossRef] [PubMed]

Table 1.
Effects of phytochemicals on autophagy dysfunction in kidney diseases.
| Animal Models with Disease | Phytochemicals | Doses and times | Alterations in Autophagy and renal outcome | Alterations in mechanism/pathway involved | References |
|---|---|---|---|---|---|
| Sepsis-induced AKI in rats | Resveratrol | 30 mg/kg for 5 days | Autophagy activation reduced renal tubular damage | Induction of p53 deacetylation, activation of deacetylase Sirt1 | [75] |
| Renal fibrosis, AKI models | Ginsenoside Re | - | Improves renal function, reduces fibrosis, and reduces autophagy | [80] | |
| DKD in experimental rats | Ferulic acid |
50 mg/kg, orally, daily for 8 weeks |
Ameliorates kidney injury, reduces apoptosis, inflammation, and defective autophagy | Modulates AGEs, MAPKs, NF-κB, induces autophagy | [15,81] |
| DKD | Polyphenols (general) |
NA |
Improve autophagy and lysosomal function, reduce fibrosis, apoptosis, and inflammation | SIRT1, mTOR, AMPK, TFEB nuclear transfer | [81] |
| STZ-induced rats | Icariin | 20, 40, and 80 mg/kg/d, orally for 12 weeks | Restore autophagy, inhibit tubulointerstitial fibrosis | GLP-1R activate, inhibit mTOR phosphorylation. | [82] |
| STZ/HFD-induced T2DM rats | Isorhamnetin | 50 mg/kg/d) for 4 or 8 weeks | Enhances renal function, improves glucose/lipid metabolism and autophagy | ↑ FYCO1, ULK-1, TECPR1, WIPI | [83] |
| SD rats + UNx + STZ | Emodin | 20–40 mg/kg, 12 weeks | Reduces apoptosis, podocyte injury, and fibrosis | Activates autophagy via AMPK/mTOR | [84] |
| Male Sprague Dawley rats with STZ | Cyclocarya paliurus triterpenic acids | 40–160 mg/kg (CPTL) and 160 mg/kg (CPTH) for 10 weeks | ↑ Autophagy; ↓ kidney injury and apoptosis | ↑ p-AMPK; ↓ p-mTOR | [85] |
| DKD mice serum | Tripterygium glycoside | In vitro: 1.25 μg/mL, 72 h | ↑ Autophagy; ↓ EMT and podocyte apoptosis | mTOR/Twist1 pathway | [84] |
| STZ-induced diabetic mice | Isorhapontigenin (polyphenol) |
- |
Activates autophagy, reduces oxidative stress, improves podocyte/endothelial cell damage | ↑AMPK/Nrf2 pathway, ↑Beclin-1, ↑Atg5, ↓P62 | [81] |
| DKD, nephrotic syndrome | Plantago asiatica/major (Hispidulin) |
- |
↑Autophagy; Prevents podocyte apoptosis via autophagy, improves proteinuria and kidney function | MAPK pathway (animal models) | [86] |
| Renal fibrosis (CKD model) | Sulforaphane |
- |
Alleviates renal fibrosis via dual regulation of autophagy in fibroblasts and tubular epithelial cells | mTOR-mediated autophagy pathway | [87] |
| Post-renal transplant, nephrotoxicity | Schisandra sphenanthera (deoxyschizandrin) | - | Enhances tacrolimus effect, nephroprotection via autophagy (limited human data) | Nrf2 pathway | [86] |
| Arsenic-induced nephrotoxicity in rats | Zingerone | 25 and 50 mg/kg | Decreases oxidative stress, inflammation, apoptosis, and kidney damage; Reduces excessive autophagy | Modulate AKT2 and FOXO1, Reduce NF-κB and IL-1β, TNF, IL-6, iNOS, COX-2, MAPK14, MAPK15, JNK | [88] |
| High glucose-induced podocyte injury | Ursolic acid |
In vitro: 5 μmol/L for 24 h (in vitro) |
Increases autophagy, improves podocyte injury | ↓ miR-21 → ↑ PTEN → ↓ PI3K/Akt/mTOR pathway | [89] |
DKD: Diabetic Kidney Disease; CKD: Chronic Kidney Disease; AKI: Acute Kidney Injury; MAPKs: Mitogen-Activated Protein Kinases; STZ: Streptozotocin; HFD: High-fat diet; T2DM: Type 2 diabetes mellitus; SD: Sprague Dawley; and UNx: Uninephrectomy.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



