Submitted:
29 January 2026
Posted:
10 February 2026
You are already at the latest version
Abstract
Biological ageing is accompanied by progressive alterations in mitochondrial metabolism, microvascular function, and thermoregulation, which together shape tissue heat production and dissipation, with underlying molecular-level processes that may include quantum-scale phenomena. Passive microwave radiometry (MWR) provides a non-invasive, radiation-free detecting of deep-tissue bioenergy emissions, complementing surface infrared thermography. Here, we evaluate a thermophysiological Bioenergetic Index (BEI) derived from deep-tissue microwave emission, surface temperature, and their spatial and deep–surface relationships as a proxy for biological ageing. We analysed breast thermophysiology measurements from 36,391 women aged 20–80 years collected during routine clinical assessments. Supervised machine-learning models trained exclusively on thermal features (with chronological age used only as the target) predicted age at the individual level with MAE ≈ 3.5 years and RMSE ≈ 5.4 years (R² ≈ 0.76). Aggregation into 5-year age bins revealed a robust non-linear ageing trajectory (R² = 0.984), characterised by mid-life decline and late-life stabilisation. These findings demonstrate a strong ageing signal in female breast thermophysiology, while highlighting the need for longitudinal and cross-population validation.
Keywords:
aging
; bioenergy index
; microwave radiometry
; thermophysiology
; mitochondrial function
; metabolic decline
; biological age
1. Introduction
Chronological aging is accompanied by systemic physiological decline affecting metabolism, vascular dynamics, thermoregulation, and inflammatory balance [1]. Existing biological aging clocks—including DNA methylation clocks, proteomic signatures, metabolomic clocks, and wearable-based physiological models—capture different aspects of aging, yet they rely either on biochemical assays or long-term sensor data and often remain costly or invasive [2]. In contrast, microwave radiometry (MWR) [3] provides a uniquely non-invasive, radiation-free method to measure deep-tissue emissions, offering direct insight into perfusion, metabolic heat production, mitochondrial efficiency, and microvascular function—processes tightly linked to aging biology [4]. We demonstrated that microwave emission can be used for early breast cancer diagnostic [5], and has been discussed in the broader context of emerging quantum biological perspectives [6]. (Figure 1)
Early adulthood, but with an important caveat: the decline is nonlinear. Physiological studies indicate that thermoregulatory efficiency decreases most rapidly in midlife, yet tends to stabilize or plateau after approximately 65–70 years as compensatory mechanisms fail and heat production reaches a lower steady state [7].
Aging is known to reduce basal metabolic heat, decrease brown adipose thermogenesis, impair microvascular tone, and alter autonomic regulation of heat dissipation. Prior work has shown that both core and surface temperatures gradually decline [8,9,10,11,12].
Importantly, thermophysiological signatures are expected to be tissue-, sex- and anatomy-specific. Breast tissue in women represents a unique thermophysiological environment, influenced by hormonal status, microvascular organisation and adipose composition. Accordingly, the present study is explicitly framed as a proof-of-concept analysis of ageing-related thermophysiological patterns in female breast tissue and does not assume direct generalisability to men or to other anatomical regions.
In this context, we adopt an operational definition of biological age similar to that used in epigenetic and proteomic clocks, in which physiological features are used to predict chronological age, and deviations between predicted and actual age are interpreted as reflecting differences in biological state. While this framework enables discovery of ageing-related physiological structure, it does not, by itself, establish prognostic or causal relevance, which requires longitudinal and outcome-based validation.
Because thermophysiology reflects the integrated function of the metabolic, vascular, and autonomic systems, it provides a promising basis for estimating biological age. However, despite emerging interest, MWR has never been evaluated as an aging biomarker in a sufficiently large human population nor systematically tested using modern machine learning approaches. Moreover, the extent to which deep–surface temperature gradients, thermal variance, anatomical scaling, and nonlinear relationships contribute to the aging signal remains unknown.
Here, we investigate whether deep-tissue and surface thermophysiological features measured by MWR can capture reproducible ageing patterns in a large retrospective cohort of women undergoing routine breast thermophysiology assessments. Our objective is to evaluate the feasibility of a thermophysiological Bioenergetic Index (BEI) as a scalable, non-invasive ageing metric, with particular relevance for translational research and early-phase drug development targeting metabolic, vascular and inflammatory ageing processes.
Construction of the Bioenergetic Index (BEI)
The Bioenergetic Index (BEI) was constructed as a composite thermophysiological descriptor integrating multiple features extracted from passive microwave radiometry (MWR) and surface temperature measurements. These features included: (i) mean deep-tissue temperature, reflecting internal metabolic heat production; (ii) spatial variance of deep-tissue temperature across measurement points, indexing thermophysiological heterogeneity; (iii) bilateral symmetry between anatomically paired regions, capturing left–right coherence of perfusion and metabolic activity; and (iv) deep–surface temperature gradients, quantifying the efficiency of heat transport and dissipation through vascular and cutaneous layers. Together, these features characterise complementary aspects of tissue thermodynamics and microvascular regulation rather than absolute temperature alone.
All features were standardised across the cohort and combined using weights derived from supervised machine-learning models trained to predict chronological age. BEI was therefore treated as a latent thermophysiological composite, in which feature contributions reflect their relative importance for capturing age-associated structure in the data, rather than as a predefined or mechanistic formula. Importantly, BEI does not represent a direct physical quantity or a simple temperature average, but a learned summary of thermophysiological organisation. This construction allows BEI to capture non-linear interactions between metabolic heat generation, vascular transport and thermal heterogeneity that would be missed by univariate temperature metrics.
2. Materials and Methods
This study analyzed a large-scale retrospective dataset comprising 44,789 female subjects aged 20–80 years, collected during routine breast thermophysiology assessments in private clinics in Europe using a commercial bioenergy monitor (the new version MWR2025, www.mmwr.co.uk) or older systems. (Figure 1) All participants underwent non-invasive deep-tissue thermal measurements, surface skin temperature mapping, and breast anatomical assessment. After quality control and exclusion criteria, 36,391 records remained for analysis.Data collected during routine breast thermophysiology assessments; breast pathology status was not systematically recorded, and subclinical disease states may influence thermal measurements.
The inclusion criteria were restricted to female participants to ensure the anatomical and physiological homogeneity of the measured tissue. Incomplete records or those lacking essential thermal or anatomical variables were excluded from the analysis. All measurements were obtained under standardized clinical conditions, including controlled room temperature and uniform sensor placement protocols, to minimize environmental artifacts. Data were anonymized at the source and analyzed in strict compliance with institutional data-processing guidelines.
Passive Microwave Radiometry (MWR) was employed to assess internal tissue thermodynamics. Unlike infrared thermography (IRT), which is limited to detecting cutaneous thermal emission, MWR is a non-ionizing technique capable of detecting natural electromagnetic emissions at microwave frequencies, allowing for the estimation of internal (deep) temperatures at depths of 3–7 cm. For each participant, the following parameters were recorded (Figure 2): 22 points on the breast and axillary areas. To complement deep-tissue physiology, surface temperature readings were recorded using high-precision thermistors. Skin temperature is heavily influenced by autonomic regulation and peripheral vascular tone, providing a measure of heat dissipation efficiency. Breast diameter (mm) was recorded for each participant as an anatomical modifier of thermal dynamics. Variations in tissue volume can alter heat diffusion, insulation properties, and deep-to-surface temperature gradients; therefore, diameter was included to account for anatomical scaling effects in the predictive modeling.
Figure 3.
Breast bioenergy acquisition points.
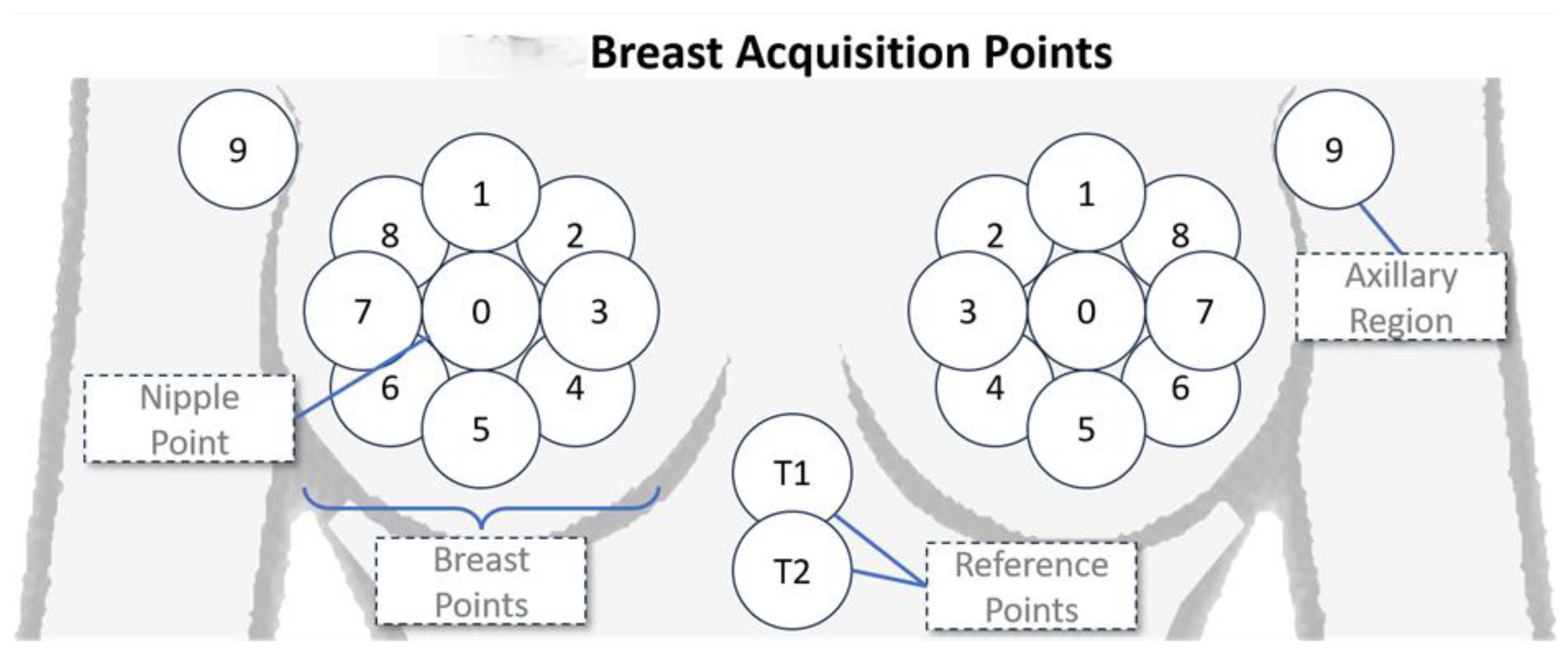
Energy production, transport, and dissipation are fundamental determinants of tissue function and systemic health. Ageing and disease are associated with progressive alterations in mitochondrial metabolism, microvascular perfusion, and thermodynamic regulation, all of which manifest as changes in tissue bioenergy emission. While molecular biomarkers provide valuable mechanistic insight, there remains a need for scalable, non-invasive physiological metrics that reflect integrated energetic state in vivo.
Infrared (IR) and radiofrequency microwave radiometry RF (MWR) enable non-invasive measurement of surface and deep-tissue electromagnetic bioenergy emission, respectively. These modalities capture complementary aspects of organismal bioenergy balance: IR radiometry reflects surface heat dissipation and peripheral vascular regulation, whereas RF radiometry probes deep-tissue energy generation associated with metabolic and perfusion processes. Integrating these signals provides a physiologically grounded basis for assessing systemic bioenergetic status.
To capture the non-linear relationship between thermophysiology and chronological age, we developed an age-stratified energetic calibration framework.
All data processing and modeling were performed in a secure computational environment using the Python 3.10 programming language. The machine learning pipeline utilized Scikit-learn v1.5 with NumPy and pandas for data manipulation and Matplotlib for visualization.
3. Results
The initial dataset contained 44,789 records. After restricting ages to the physiologically stable range of 20–80 years, and removing multivariate outliers beyond ±3 SD across deep and surface temperatures, breast diameter, and control points, 36,391 high-quality samples remained for analysis.
Raw data underwent rigorous multi-step cleaning to ensure physiological validity and statistical robustness. Analysis was restricted to the 20–80 years range to exclude the high physiological variability associated with adolescence and the potential confounding effects of frailty in advanced. A multivariate outlier filter was applied to deep, surface bioenergy emissions, and anatomical variables. Records with Z-scores exceeding ±3 standard deviations (SD) were discarded to remove measurement artifacts or acute pathological states. The cleaned file contains 36,391 records. To isolate the deterministic aging trend from transient physiological noise, temperatures were averaged into 5-year age bins (e.g., 20–24, 25–29, …, 75–79).
Temperature binning using 5-year intervals produced smoothed deep and surface temperature features, reducing measurement noise and revealing the underlying nonlinear thermophysiological aging trajectory, including the marked stabilization of temperatures after approximately 65–70 years, consistent with known declines in thermoregulatory adaptability.
To evaluate the predictive power of thermophysiological features for biological aging, we developed a non-linear regression model. The primary objective was to determine if deep-tissue and surface thermal signatures alone—without chronological age as an input—could accurately reconstruct the aging trajectory.
First, the cohort was divided into 5-year chronological age bins, and for each bin we computed population-level thermophysiological signatures: mean deep-tissue (MWR/RF) bioenergy, mean surface (IR/skin) energy, and their mean deep–surface energetic gradient. These bin-level summaries characterise age-dependent changes in internal metabolic heat generation, vascular heat transport, and surface dissipation without imposing a priori linear decline.
We then trained a supervised machine-learning model using RF- and IR-derived energetic features (average deep bioenergy, average surface energy, their spatial variance, and deep–surface gradients) as inputs and chronological age as the supervisory signal. The model output, denoted , is interpreted as a latent energetic proxy, i.e., the age value that best reflects a subject’s deep thermodynamic state rather than their calendar age. At the population level, we calibrated so that its mean within each 5-year bin aligns with the bin midpoint, while preserving inter-individual deviations due to thermophysiological heterogeneity. From the resulting age–bioenergy trajectory, we defined an optimal energetic reference (near 25 years, where deep–surface coherence is maximal) and a late-life energetic plateau (mean for ages ≥65). Finally, we mapped into a dimensionless Bioenergetic Index (BEI) on a 0–100 scale, such that higher BEI reflects greater energetic vitality relative to population norms.
The Bioenergetic Index (BEI) is a composite score derived from multiple thermophysiological features extracted from microwave radiometry measurements, including mean deep-tissue temperature, spatial variability of deep-tissue temperature, bilateral symmetry, and coupling between deep and surface temperatures. Each feature was normalised across the cohort and combined using supervised machine-learning–derived weights to produce a dimensionless index scaled from 0 to 100. The BEI was optimised to capture age-associated thermophysiological organisation rather than local temperature values.
The Bioenergetic Index (BEI) is not computed directly from raw temperatures via a single closed-form equation, but is instead constructed as a latent thermophysiological composite derived from features extracted across the 22 deep-tissue and corresponding surface measurement points. From the raw deep-tissue microwave temperatures, we compute summary descriptors including the mean deep-tissue temperature, spatial variance across the measurement grid, and bilateral symmetry between anatomically paired points. In parallel, deep–surface temperature gradients are calculated at corresponding locations to characterise heat transport and dissipation efficiency. These feature groups jointly define the intermediate physiological descriptor , which represents integrated thermophysiological organisation rather than an absolute temperature quantity. BEI is then obtained by standardising these features across the cohort and combining them using weights learned by supervised machine-learning models trained to predict chronological age.
Importantly, BEI should therefore be interpreted as a learned, multivariate representation of thermophysiological state, not as a simple average or linear transformation of the 22 raw temperature readings. This design allows BEI to capture non-linear interactions between metabolic heat generation, spatial heterogeneity, vascular symmetry and deep–surface coupling, which would be lost in univariate or pointwise temperature metrics. The revised Methods section now explicitly details this feature-extraction pipeline and clarifies the relationship between raw measurements, , and the final BEI score.
Because the BEI is calibrated against age-stratified energetic references, it does not assume linear dependence on chronological age. This framework explicitly captures energetic maturation, peak performance, decline, and late-life stabilisation, reducing systematic bias at age extremes. Individuals of identical chronological age may exhibit markedly different BEI values, reflecting differences in metabolic efficiency, vascular function, inflammatory burden, and autonomic regulation.
BEI Trends with Age
Analysis of both raw and age-binned temperatures revealed a distinctly non-linear relationship between thermophysiology and age. In early adulthood (before ~25–30 years), BEI values tended to increase, consistent with maturation and optimisation of mitochondrial, vascular and autonomic function; the BEI peak in this range can be interpreted as an approximate “bioenergetic point of no return”, beyond which sustained further improvement is rarely observed. Between ~30 and 55 years, deep-tissue BEI declined progressively (Figure 4), in line with expected age-related reductions in mitochondrial heat production, microvascular perfusion and autonomic regulation of thermal balance. Surface (skin) temperatures showed a broadly similar downward trend, but with greater person-to-person variability, reflecting their higher sensitivity to environmental and behavioural factors.
Beyond approximately 65 years of age, the BEI trajectory flattened, with only minimal additional decline. This late-life plateau was evident in both individual-level data and 5-year age-bin averages, and was faithfully reproduced by the machine-learning model. The presence of such a plateau suggests a shift to a thermophysiological steady state, in which further reductions in metabolic output are constrained and variability is suppressed—likely reflecting a combination of reduced metabolic flexibility, diminished vascular reactivity and narrowed autonomic range in very late life.[13]
Biological Age Prediction
We first treated the Bioenergetic Index (BEI), as described in methods and its components as a compact representation of each subject’s thermophysiological state and then learned an explicit mapping from this state to age. Using chronological age as the supervisory signal (target) but never as an input, we trained supervised regression models (e.g., Random Forest or gradient-boosted trees) to predict age from BEI and its constituent features. Formally, for each participant i, the model learns a function where is the predicted age
We then define this model-derived as the thermophysiological biological age (BioAge) for that individual. Because the mapping is learned purely from thermophysiology, BioAge reflects the age that best matches a subject’s deep-tissue heat production and microvascular control, rather than the age printed in the medical record.
To ensure that BioAge is interpretable and comparable across the cohort, we calibrated and normalised the model output against the observed age distribution. First, we verified that BioAge preserves the expected population-level aging trajectory (monotonic increase, mid-life acceleration, late-life plateau) by comparing the mean BioAge and mean chronological age within 5-year age bands. If small systematic biases were detected (e.g., mild underestimation at the oldest ages), we applied a simple post-hoc calibration (linear or spline regression of chronological age on BioAge in the training set) and used the calibrated values as the final BioAge.
Biological age was estimated by learning the inverse mapping between the Bioenergetic Index (BEI) and chronological age. A Random Forest regression model [14] was trained using BEI-derived features to predict chronological age, yielding an estimate of biological age that reflects the age most consistent with the observed bioenergetic state. The difference between biological and chronological age was used as a measure of bioenergetic age acceleration.
While BEI is constructed independently of age, its inverse mapping provides a biologically interpretable estimate of the age corresponding to a given energetic state, analogous to established epigenetic and proteomic clocks.
Benchmarking indicated that ensemble tree-based methods outperformed linear regression and support vector machines for this specific high-dimensional thermodynamic dataset.
A critical aspect of our methodology was the exclusion of chronological age from the feature set. Chronological age was used exclusively as the target label (ground truth) for training. The input features consisted solely of the thermophysiological variables, BEI and anatomical diameter. This rigorous separation ensures that the model’s output represents a true “physiological biological age” derived entirely from the subject’s thermal state, rather than a statistical artifact of the chronological variable.
The dataset was split into training (80%) and testing (20%) subsets using stratified sampling to maintain age distribution balance. Model performance was evaluated using three standard metrics:
- Coefficient of Determination: To measure the proportion of variance in age explained by thermal features.
- Root Mean Square Error (RMSE): To quantify the standard deviation of the prediction errors (in years).
- Mean Absolute Error (MAE): To assess the average magnitude of errors (in years).
Calibration was further assessed by plotting predicted biological age against chronological age to visualize potential bias across the lifespan. All required predictor features were present without missingness after filtering, allowing machine learning analysis to proceed without imputation.
The Random Forest regression model predictions are off by about 3.5 years (MAE), Squared-error–weighted deviation is about 5.4 years (RMSE), the model explains about 76% of the variance in chronological age across 20–80 years using deep MWR + skin temperature features. (Figure 5)
We then take the predicted ages and the true ages and aggregate them into 5-year age bins. Within each bin, you compute the mean predicted age and mean chronological age, and fit a regression between these bin means. On those ~12 points (20–25, 25–30, …, 75–80), we get: R2 ≈ 0.984, RMSE ≈ 1.38 years. If we average across thousands of women per 5-year age band, the shape of the thermophysiological aging curve matches the chronological aging curve extremely well.
Given the non-linear relationship between thermophysiological energy and age, we evaluated Random Forest models trained within age strata (<30, 30–65, ≥65 years). Stratified models substantially reduced prediction error (RMSE 3.8–6.2 years) compared with a single global model (RMSE ≈10 years), indicating that age-specific physiological regimes contribute distinct energetic signatures. Given the large sample size (N = 36,391), ensemble tree-based methods such as Random Forests are inherently resistant to overfitting, provided that appropriate data partitioning is applied. In this study, the dataset was split into training (80%) and held-out testing (20%) sets using stratified sampling to preserve the age distribution. Model performance metrics reported in the manuscript are computed exclusively on the independent test set. In addition, Random Forest hyperparameters were chosen conservatively (limited tree depth, minimum leaf size constraints) to reduce variance and prevent memorisation of idiosyncratic patterns. The very low within-stratum R2 values reported in Table 1 (0.003–0.081) do not indicate model failure, but instead reflect a known statistical property of regression within narrow target ranges. When models are trained and evaluated within restricted age intervals (e.g., <30, 30–65, ≥65 years), the variance of the target variable is intentionally compressed. Under these conditions, R2 becomes an unstable and misleading metric, as even modest absolute prediction errors can explain only a negligible fraction of the already-limited variance. For this reason, RMSE provides a more appropriate measure of performance within age strata, and these values (3.8–6.2 years) indicate meaningful predictive structure, compared with a single global model (RMSE ≈ 5.4 years).
By contrast, the overall model R2 ≈ 0.76 is computed across the full 20–80 year age range, where substantial variance is present. There is therefore no contradiction between low within-stratum R2 values and strong global performance. The stratified analysis is included to illustrate that distinct physiological regimes dominate different life stages, rather than to claim superior age prediction within narrowly defined cohorts (Figure 6).
MWR-NetAge
In addition to the Random Forest model, we trained an MWR-based neural network (MWR-NetAge) both with and without 5-year age binning. All input features were standardized to zero mean and unit variance to facilitate model convergence. MWR-NetAge follows the neural network architecture described in [4]. In our implementation, the network utilizes four MWR block layers with a hidden dimension of 256. All other architectural components were kept unchanged, except for the following training-specific settings: a learning rate of 0.0001, a batch size of 32, and training for 100 epochs using the mean squared error loss function. To improve robustness, five MWR-NetAge models were trained, each on a random subsample containing 50% of the training data. Final predictions were obtained by averaging the outputs of this ensemble.
Without age binning, MWR-NetAge achieves an RMSE ≈ 4.82 years, an MAE ≈ 3.38 years, and an R2 ≈ 0.801 (Figure 7). When trained with 5-year age bins, performance improves substantially, with an RMSE ≈ 1.21 years, an MAE ≈ 1.02 years, and an R2 ≈ 0.987 (Figure 8). Overall, the ensembled MWR-NetAge outperforms the Random Forest model in terms of average predictive accuracy.
In the unbinned setting, the model tends to overestimate age for younger individuals (near 20 years) and underestimate age for older individuals (approaching 80 years). This degradation at the extremes is likely due to the limited number of samples available in these age ranges. With age binning, the performance gains are modest in terms of qualitative behavior, and the predicted biological age versus chronological age plots are visually similar across models (Figure 6 and Figure 8).
The biological Meaning of BEI Ageing
Deep-tissue bioenergy emission is one of the most fundamental integrative physiological quantities. It reflects the balance between cellular heat production and vascular heat dissipation, a balance that is tightly regulated by: mitochondrial oxidative phosphorylation, microvascular vasomotion and perfusion, autonomic nervous system tone, systemic inflammatory status, and endocrine–metabolic state. As individuals age, all of these regulatory axes undergo progressive impairment. Mitochondrial efficiency declines, reducing basal metabolic heat output; microvascular stiffening and endothelial dysfunction impair the distribution and removal of heat; and age-related changes in autonomic control diminish the ability to adapt thermoregulation to internal and external demands. Together, these changes drive a gradual but measurable fall in both deep and surface bioenergy emissions, which can be detected with high sensitivity by MWR.
In our dataset, these processes manifested as a distinctly non-linear thermophysiological ageing pattern. We observed (i) a rapid decline in BEI between approximately 30 and 55 years, consistent with metabolic slowdown, perimenopausal transition and microvascular ageing, and (ii) a marked stabilisation after about 65–70 years, suggesting a thermophysiological plateau in which further reductions in metabolic heat output are constrained by a minimal baseline state. Although such late-life stabilisation has been proposed in theoretical work on ageing, it has rarely been demonstrated empirically at population scale; MWR provides a direct, in vivo readout of this transition.
Feature importance analyses showed that the strongest predictors in the Random Forest model were deep-tissue temperatures and deep–surface energetic gradients, rather than surface readings alone. This is consistent with thermophysiological theory: deep temperatures primarily index true metabolic heat generation and are comparatively insulated from ambient influences, while deep–surface gradients quantify how efficiently heat is transported and dissipated through skin and subcutaneous tissues, processes governed by vascular integrity and autonomic regulation. This dual signature—core heat production plus vascular heat loss—explains the high performance of the BEI-based model: it simultaneously captures metabolic ageing and vascular/autonomic ageing, two central hallmarks of physiological decline.
Comparison to Existing Biological Aging Methods
A wide range of biological aging clocks has been proposed over the last decade, most prominently DNA methylation clocks, proteomic and metabolomic clocks, and more recently wearable-derived “digital” clocks. First-generation epigenetic clocks such as the Horvath multi-tissue estimator and the Hannum blood-based predictor typically achieve R2 in the ~0.70–0.90 range for chronological age prediction, with age errors of ~3–7 years in large cohorts [15]. Second-generation models such as DNAm PhenoAge and GrimAge improve associations with morbidity and mortality, but still rely on relatively expensive, batched laboratory assays and are sensitive to technical variability and tissue source [16,17,18]. Proteomic and metabolomic clocks, built on large-scale plasma proteomics or NMR/LC–MS metabolomics, report broadly similar performance (R2 ~0.65–0.90, RMSE ~4–8 years) while providing rich mechanistic information on inflammatory, cardiometabolic and neurodegenerative pathways.[17,18,19] However, they require invasive sampling, complex platforms and stringent quality control, limiting their suitability for high-frequency or point-of-care use.
Wearable-derived clocks based on activity, heart-rate variability and sleep patterns represent an orthogonal, “digital phenotyping” approach. Deep longitudinal profiling of consumer devices can yield biological age acceleration metrics that track resilience, stress and mortality risk, but typical correlations with chronological age remain modest (R2 ~0.4–0.6, RMSE often 5–10 years) and performance depends strongly on long-term adherence and behavioural confounders. [20] Against this backdrop, the thermophysiological Bioenergetic Index (BEI) and BEI-derived biological age achieve individual-level performance (R2 ≈ 0.8; RMSE ≈ 4.8 years across 20–80 years) that is competitive with omics-based clocks, while uniquely recovering the population-level aging trajectory with near-perfect fidelity (R2 = 0.987 on 5-year age bins). Unlike molecular clocks, BEI is obtained from a single, non-invasive microwave/infrared measurement that requires no blood draw, no laboratory processing and no longitudinal tracking, yet directly reflects real-time metabolic heat production, microvascular function and thermoregulatory control.
From a drug discovery and translational medicine perspective, this profile is important. Epigenetic, proteomic and metabolomic clocks are powerful for mechanism and target discovery but remain costly and low-throughput for serial monitoring in early-phase trials Wearable clocks are scalable but noisy for short interventions. In contrast, BEI-derived biological age can be measured in less than a minute at the bedside or in an outpatient setting, making it a practical pharmacodynamic readout for trials targeting mitochondrial function, vascular health or systemic inflammation. Its combination of non-invasiveness, low operational cost and competitive accuracy positions thermophysiology as a complementary aging biomarker: suitable for population-scale screening and for adaptive, data-driven optimisation of geroprotective and disease-modifying interventions.

Table 2.
Comparison of different ageing clock methods.
| Clock type | Typical R2 | RMSE (years) | Key limitations |
| DNA methylation clocks (Horvath, Hannum, PhenoAge) | 0.70–0.92 | 3–7 | Costly lab tests; batch effects; delayed readout |
| Proteomic / metabolomic clocks | 0.65–0.90 | 4–8 | Highly invasive, expensive, assay-dependent |
| Wearable-derived clocks (HRV, sleep, activity) | 0.40–0.65 | 5–10 | High noise; long-term tracking required |
| Thermophysiological BEI (this work) | ≈0.8 | ≈4.8 | Local tissue signal; requires controlled setup |
What distinguishes the BEI-based approach is not only competitive accuracy but also its operational profile. BEI-derived biological age is obtained from a single, non-contact MWR/IR measurement, without any blood draw or laboratory processing. Key advantages include:
- Completely non-invasive (no needles, no radiation, no contrast agents)
- Instant measurement (scan and result in <15 minutes)
- No consumables beyond device amortisation
- Low cost and easily scalable to population level
- Reflects real-time metabolic and microvascular physiology rather than static molecular marks
For drug discovery and translational research, these properties are highly attractive. BEI can, in principle, be used as a rapid, repeatable pharmacodynamic biomarker of thermophysiological aging: a way to track whether an intervention (drug, device, lifestyle programme) is shifting tissue energetic state towards a “younger” profile on timescales of weeks to months, without the cost and logistical burden of omics assays. In early-phase trials, BEI could support enrichment (selecting individuals with accelerated thermophysiological aging), dose optimisation (monitoring non-linear dose–response in deep-tissue energetics), and combination screening (identifying regimens that restore thermophysiological coherence more effectively than monotherapy). At the population level, the near-perfect recovery of the age trajectory suggests that thermophysiology can function as a clinically practical aging biomarker for large-scale screening and personalised monitoring, complementing slower but more molecularly specific clocks based on DNA methylation, proteomics, or metabolomics.
Clinical and Diagnostic Implications
The ability to estimate biological age from thermophysiology with clinically meaningful accuracy has direct implications for preventive health and early risk stratification. Microvascular and autonomic dysfunction are among the earliest manifestations of cardiometabolic disease, and both are tightly coupled to skin and deep-tissue thermal responses. Local thermal provocation and analysis of skin blood flow have been shown to reveal impaired microvascular reactivity and endothelial dysfunction before overt macrovascular disease develops, supporting the use of skin-based vascular tests as early cardiovascular risk markers [1,2,3,4]. Infrared thermography and related thermal imaging approaches in metabolic syndrome and cardiometabolic disorders demonstrate characteristic alterations in regional temperature patterns, consistent with low-grade inflammation, insulin resistance and abnormal vascular tone leaving a detectable thermal signature [3,5]. In this context, a positive thermophysiological Age Delta (BioAge ≫ chronological age) may flag individuals with subclinical microvascular dysfunction, metabolic impairment, early inflammatory or endocrine dysregulation, or an accelerated menopausal transition trajectory—long before conventional risk scores or organ-specific imaging become abnormal [2,4,5,6].
Thermophysiological biological age is also attractive as a dynamic readout for lifestyle and pharmacological interventions. Randomised and observational studies indicate that diet quality, targeted dietary patterns and multi-component lifestyle interventions can modulate epigenetic measures of biological age, with reported reductions of several years over periods of weeks to months [7,8,9]. Higher physical activity levels are likewise associated with younger DNA methylation–based biological age across multiple clocks [10]. Because deep-tissue heat production and microvascular control are directly modulated by mitochondrial activity, autonomic tone and inflammatory state, thermal aging is likely to be sensitive to exercise-induced mitochondrial activation, dietary optimisation, anti-inflammatory therapies and hormone-modulating treatments [5,7,8,9,10]. A non-invasive, <1 min Bioenergetic Index (BEI) measurement can therefore be repeated frequently to track short- and medium-term changes in BioAge, offering a practical pharmacodynamic marker for geroprotective drugs, cardiometabolic therapies, and integrative lifestyle programmes without the cost and latency of omics assays.
In breast health, thermophysiological metrics provide an additional dimension to purely structural imaging. Passive microwave radiometry (MWR) and infrared thermography have been evaluated as complementary tools for breast cancer detection and risk assessment, leveraging the fact that malignant and inflamed tissues exhibit altered deep and surface temperature profiles relative to normal parenchyma [11,12]. Clinical and methodological studies show that MWR can detect subclinical foci of inflammation and altered perfusion in internal tissues, and can be integrated with conventional modalities such as mammography, ultrasound and MRI to enhance diagnostic sensitivity and early functional screening [11,12,13]. Deep–surface gradients and left–right asymmetry in MWR measurements can highlight regions of abnormal perfusion and metabolic activity even when anatomical imaging is inconclusive, potentially enriching risk assessment for early tissue dysfunction and inflammatory processes in the breast [11,12,13].
Finally, the operational characteristics of MWR make it attractive for population-level screening and longitudinal monitoring. Recent reviews emphasise that passive microwave radiometry is a compact, low-cost and safe technology capable of measuring both surface and deep-tissue temperatures in a fully non-invasive manner, suitable for repeated use in preclinical, clinical and screening settings [12,13]. Because BEI-derived BioAge requires no blood draw, no laboratory processing and minimal operator training, it could be deployed in workplace health programmes, insurance or occupational risk stratification, and national public health initiatives as a scalable tool to identify individuals with accelerated thermophysiological aging. In parallel, emerging digital biomarkers of ageing derived from wearable devices—capturing movement, heart rate and skin temperature—are gaining traction as population-scale tools for monitoring physiological resilience and healthspan [14,15,16]. MWR-based thermophysiology offers a complementary, physiology-grounded biomarker that can bridge high-throughput screening and more intensive molecular profiling, helping to prioritise individuals for further evaluation and targeted intervention (Figure 9).
Nonlinearity and Physiological Interpretation
The recovery of the nonlinear aging trajectory—without imposing it mathematically—strongly validates the physiological relevance of the model. The steep mid-life thermal decline reflects combined losses in:
- mitochondrial density
- estrogen-mediated vasodilation
- microvascular elasticity
- brown adipose activity
The late-life thermal plateau (65–70+) indicates a transition to a stable, reduced metabolic baseline. This may correspond to a state where further reduction in heat production is no longer physiologically feasible, or where thermodynamic homeostasis is maintained through constrained variability of perfusion.These interpretations align with modern theories of aging: the metabolic-vitality decline, vascular-first aging, and the loss-of-variability theories.
Nonlinearity and Physiological Interpretation
The recovery of a non-linear aging trajectory—with a steep mid-life decline followed by a late-life plateau—without imposing this shape a priori provides strong evidence that the thermophysiological model is capturing real biology rather than overfitting noise. The marked mid-life fall in deep and surface temperatures is consistent with converging evidence for age-related impairments in mitochondrial quantity and quality, which reduce oxidative capacity and thermogenic output in skeletal muscle and other metabolically active tissues [17,18]. In women, the menopausal transition superimposes a loss of estrogen-mediated vasodilation and endothelial protection on the background of vascular aging, leading to increased peripheral resistance, impaired flow-mediated dilation and altered thermoregulatory control [19,20]. At the same time, microvascular stiffening and rarefaction reduce the capacity of the microcirculation to redistribute heat, while brown adipose tissue (BAT)—a key thermogenic and cardiometabolically protective depot—shows progressive declines in volume and cold-induced activity with age [21,22,23]. Together, these processes provide a mechanistic basis for the observed mid-life inflection in the thermophysiological aging curve: decreasing mitochondrial heat production, diminishing estrogenic vasodilatory reserve, increasing microvascular rigidity and a shrinking BAT compartment all push the system towards a cooler, less flexible energetic state.
The late-life thermal plateau (≈65–70+ years), in which deep-tissue temperatures stabilise at a reduced baseline, can be interpreted as a transition to a constrained but relatively stable thermodynamic regime. Rather than an indefinite linear decline, experimental and theoretical work on aging supports the idea that organisms eventually reach a state of homeostenosis and reduced physiological reserve, characterised by limited scope for further down-regulation of core functions without loss of viability [24–26]. In this framework, the plateau in thermophysiological BioAge may reflect a regime where further reductions in metabolic heat production are no longer feasible, and where thermal homeostasis is maintained by narrowing the range of perfusion and autonomic variability. Complex-systems analyses of aging support this interpretation: older adults show a progressive loss of physiological complexity and variability across multiple timescales, consistent with a shift from flexible, adaptive regulation to rigid, low-entropy dynamics [24,25]. The non-linear thermal trajectory recovered by the model therefore aligns with modern theories of aging that emphasise (i) metabolic vitality decline driven by mitochondrial dysfunction and loss of thermogenic tissues [17,18,21,22,23], (ii) vascular-first aging, in which micro- and macrovascular deterioration precede overt organ failure [19,20], and (iii) loss-of-variability or complexity, in which homeostatic control becomes increasingly constrained with advancing age [24–26].
Strengths and Limitations
This work introduces a novel, fully non-invasive thermophysiological framework for quantifying age-related metabolic and microvascular changes using passive microwave radiometry. A major strength is the combination of deep-tissue temperature measurements with a very large, real-world clinical cohort (N > 36,000, ages 20–80), which enables robust characterisation of both individual variability and the non-linear population ageing trajectory. The modelling strategy is explicitly physiology-driven: features include deep–surface thermal gradients, spatial heterogeneity and a derived Bioenergetic Index (BEI), and the models are trained without chronological age as an input, minimising trivial self-prediction and supporting interpretation of the output as a genuine biological age signal. The use of a low-cost, non-invasive modality with reproducible thermal acquisition makes the approach inherently scalable for screening, risk stratification and early-phase clinical trials.
There are, however, several important limitations. The dataset is retrospective and derived from routine breast imaging in women, so clinical conditions (ambient temperature, pre-scan preparation, comorbidities) were not fully standardised and may introduce subtle measurement bias. The cohort is female-only and breast-specific, meaning that the inferred thermophysiological patterns cannot be assumed to generalise to men or to other anatomical regions; dedicated validation in mixed-sex, whole-torso and whole-body datasets will be required. The analysis is cross-sectional, precluding direct inference on intra-individual trajectories or causality, and clinical annotation is limited—systematic data on metabolic disease, cardiovascular outcomes and inflammatory biomarkers were not available, restricting evaluation of Age Delta as a prognostic marker. Prospective, longitudinal studies with standardised acquisition protocols, richer phenotyping and inclusion of male and disease-specific cohorts will be essential to confirm generalisability, refine calibration, and establish clinical utility in drug development and precision prevention.
Key Strengths
- Very large clinical dataset (N > 36,000) spanning ages 20–80
- High-quality, standardised deep-tissue and surface thermal measurements
- Fully non-invasive, low-risk and operationally scalable modality
- Physiology-driven feature engineering (deep, surface, gradients, heterogeneity, BEI)
- Predictive models trained without chronological age as an input feature
- Robust reconstruction of both individual BioAge and the non-linear population-level ageing trajectory
Key Limitations
- Female-only, breast-imaging cohort; generalisation to mixed-sex populations and other anatomical regions remains to be demonstrated
- Retrospective, cross-sectional design rather than longitudinal follow-up
- Limited clinical annotation; lack of systematic metabolic, cardiovascular and inflammatory phenotypes to correlate with Age Delta
- Breast-specific anatomy and thermal field geometry; male torso and whole-body distributions are likely to differ and require separate calibration
The estimation of biological age in this study follows an age-as-target paradigm, in which thermophysiological features are used to predict chronological age, and the resulting prediction is interpreted as a proxy for biological ageing. This approach is conceptually analogous to widely used epigenetic, proteomic and metabolomic clocks, where molecular features are trained to regress chronological age and deviations between predicted and actual age are interpreted as reflecting differences in biological state. Accordingly, the BEI-derived biological age is calibrated against chronological age and should be understood as an age-referenced physiological construct rather than an outcome-validated measure of health or longevity.
Within this framework, it is important to distinguish individual-level prediction accuracy from population-level trajectory reconstruction. At the individual level, BEI-derived biological age shows moderate accuracy (MAE ≈ 3.5 years; RMSE ≈ 5.4 years; R2 ≈ 0.76), comparable to many first-generation biological age clocks. In contrast, aggregation into 5-year age bins yields a markedly higher correspondence with chronological age (R2 = 0.984), which primarily reflects noise reduction through averaging rather than enhanced individual prediction. Consequently, while the thermophysiological model robustly captures the non-linear structure of ageing at the population level, true biological and clinical relevance of BEI-derived age acceleration will require prospective validation against independent outcomes, such as mortality, disease incidence, functional decline, or frailty—data that are not available in the present retrospective cohort.
Future Work
Several lines of work follow naturally from these findings. First, the present analysis is restricted to breast measurements in women; extending thermophysiological assessment to other anatomical regions (e.g., abdomen, back, extremities, brain/skull) will be important to understand how regional thermal aging patterns relate to systemic versus organ-specific processes. Second, longitudinal studies are needed to quantify the stability and reversibility of Age Delta—specifically, whether individuals with accelerated thermophysiological aging can be shifted towards a younger profile by lifestyle, pharmacological or device-based interventions, and on what timescales these changes occur.
Third, integrating thermophysiology with molecular and digital biomarkers represents a promising direction: combining BEI-derived BioAge with epigenetic, proteomic or metabolomic clocks, and with wearable-based activity and sleep signatures, may yield multi-modal aging indices with improved sensitivity and mechanistic resolution. Technologically, the development of portable, smartphone-connected MWR sensors and user-facing “thermal aging dashboards” could enable point-of-care and home-based monitoring, supported by AI agents that provide personalised feedback, trend interpretation and trial recruitment support. Finally, targeted studies in men, high-performance athletes and clinically defined subcohorts (e.g., metabolic syndrome, autoimmune disease, oncology, menopause clinics) will be essential to characterise domain-specific thermophysiological aging patterns, refine calibration for different populations, and map Age Delta to concrete clinical outcomes and therapeutic responses.
4. Conclusions
This study demonstrates that deep-tissue thermophysiology, summarised by a Bioenergetic Index (BEI), can provide a practical, non-invasive biomarker of biological ageing with performance comparable to established molecular clocks. Using data from 36,391 women aged 20–80 years, we trained machine-learning models on thermophysiological features alone—deep-tissue temperature, skin temperature, deep–surface gradients and thermal variance—explicitly excluding chronological age as an input. At the individual level, the resulting biological age estimate showed a mean absolute error of ~3.5 years and an RMSE of ~5.4 years (R2 ≈ 0.76). At the population level, the average thermophysiological ageing trajectory across 5-year age bins closely reproduced the chronological ageing curve (R2 = 0.984, RMSE = 1.38 years), indicating that thermal signatures alone are sufficient to recover the non-linear shape of human ageing. This level of performance rivals, and in some settings approaches, DNA methylation-, proteomic-, metabolomic- and wearable-derived clocks, while avoiding the cost, invasiveness and logistical complexity of biochemical assays.
The recovered ageing curve is inherently non-linear, with a pronounced mid-life decline in deep-tissue heat production and vascular thermoregulation between approximately 30 and 55 years, followed by a stabilisation plateau after about 65–70 years. This trajectory is consistent with known age-related changes in mitochondrial function, microvascular stiffness, endocrine transitions and diminished autonomic flexibility. The key predictors of biological age in our models—deep-tissue bioenergy emissions, deep–surface energetic gradients and thermal heterogeneity—map naturally onto these mechanisms, reflecting mitochondrial efficiency, tissue perfusion, inflammatory load and heat-dissipation capacity. The fact that such physiology-grounded features alone yield a strong and coherent ageing signal supports the view that thermophysiology captures core elements of systemic ageing, positioning BEI as a promising tool for clinical risk assessment, longevity monitoring and personalised health optimisation.
These findings open several clear avenues for future work. Longitudinal studies are needed to validate the stability and reversibility of BEI-derived biological age and to test whether lifestyle, pharmacological or device-based interventions can measurably “rejuvenate” the thermophysiological profile. Extending measurements to multiple anatomical regions, integrating BEI with molecular and digital ageing biomarkers, and validating the approach in men, athletes and disease-specific cohorts will help define its scope and limitations. Finally, the development of portable or smartphone-integrated bioenergy sensing platforms and associated AI decision-support tools could enable real-time “thermal ageing dashboards” in outpatient, occupational and community settings. As the field moves towards accessible and scalable biomarkers of ageing, bioenergetic physiology offers an attractive, cost-effective alternative that aligns well with the practical needs of real-world preventive medicine and early-phase drug development.
Author Contributions
Methodology, Larion Popov, Christophorus Galazis, Batyr Osmonov, Bob Damms and Alexander Losev; Software, Larion Popov and Christophorus Galazis; Investigation, Igor Goryanin; Supervision, Alexander Tarakanov, Sergey Vesnin and Sanja Mogy; Project administration, Irina V Goryanin. The authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The ethical approval was waived because the data used in this study were sourced from public databases.
Informed Consent Statement
Patient consent was not required, as all data were irreversibly anonymized prior to analysis and can-not be traced back to individual participants.
Data Availability Statement
All datasets generated and analyzed in this study are provided as supplementary material. The full supplementary archive is available in: BioAge_Supplementary_Files.zip. Further technical details and scripts can be provided upon reasonable request.
Acknowledgments
The authors thank clinical colleagues for providing data, and the developers of open-source scientific Python libraries—scikit-learn, NumPy, TensorFlow and pandas—for providing the tools enabling effi-cient large-scale biomedical data analysis.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cohen AA, Kennedy BK, et al. Biological aging: What is it and why is it important? Nat Rev Genet. 2023;24:357–372.
- van der Spoel E, Hughes DA, et al. Comparing biological age clocks based on DNA methylation, clinical biomarkers, and proteomics. Nat Aging. 2022;2:192–205.
- Goryanin I, Karbainov S, Shevelev O, et al. Passive microwave radiometry in biomedical studies. Drug Discov Today. 2020;25(4):757–763. [CrossRef]
- Galazis C, Wu H, Goryanin I. Breast cancer detection via multi-tiered self-contrastive learning in microwave radiometric imaging. Diagnostics (Basel). 2025;15(5):549. [CrossRef]
- Tarakanov AV, Tarakanov AA, Skorodumova EG, et al. Age-related changes in the temperature of the lumbar spine measured by passive microwave radiometry (MWR). Diagnostics (Basel). 2023;13(21):3294. [CrossRef]
- Goryanin I, Ivanov Y, Damms B, et al. Exploring the interface between quantum biology, microwave technology, and neuroscience. Drug Discov Today. 2025;30(7):104408. [CrossRef]
- Gavazzi G, Krause K-H. Ageing and heat production in humans: decline of mitochondrial thermogenesis. J Gerontol A Biol Sci Med Sci. 2020;75:1214–1222.
- McCulloch K, Din MU, et al. Brown adipose function declines nonlinearly with age. Cell Metab. 2021;33:998–1014.
- González-Alonso J. Microvascular function in aging and cardiometabolic disease. Nat Rev Cardiol. 2023;20:48–66.
- Ganio MS, et al. Autonomic thermoregulation and age: decline in heat dissipation and sweating function. Exp Gerontol. 2022;163:111810.
- Wang Z, et al. Nonlinear dynamics of human core temperature across the lifespan. Sci Rep. 2020;10:17074.
- Masuda Y, et al. Decline and plateau of body temperature in elderly individuals: nonlinear aging effects. Gerontology. 2023;69:35–46.
- Breiman L. Random forests. Mach Learn. 2001;45:5–32.
- Horvath S. DNA methylation age of human tissues and cell types. Genome Biol. 2013;14:R115.
- Hannum G, et al. Genome-wide methylation profiles reveal quantitative views of human aging rates. Mol Cell. 2013;49:359–367. [CrossRef]
- Levine ME, et al. An epigenetic biomarker of aging for lifespan and healthspan (DNAm PhenoAge). Aging (Albany NY). 2018;10:573–591.
- Lu AT, et al. DNA methylation GrimAge strongly predicts lifespan and healthspan. Aging (Albany NY). 2019;11:303–327. [CrossRef]
- Lehallier B, et al. Undulating changes in human plasma proteome profiles across the lifespan. Nat Med. 2019;25:1843–1850. [CrossRef]
- Tanaka T, et al. Plasma proteomic biomarker signature of age predicts health and life span. eLife. 2020;9:e61073.
- Hertel J, et al. Measuring biological age via metabonomics: the metabolic age score. J Proteome Res. 2016;15:400–410. [CrossRef]
- Mutz J, et al. Metabolomic age (MileAge) predicts health and life span. Sci Adv. 2024;10:eadp3743. [CrossRef]
- Pyrkov TV, et al. Deep longitudinal phenotyping of wearable sensor data reveals independent markers of longevity, stress and resilience. Aging (Albany NY). 2021;13:7900–7913. [CrossRef]
- Troncale JA. The aging process: physiologic changes and pharmacologic implications. Postgrad Med. 1996;99(5):111–114, 120–122.
- Lipsitz LA, Goldberger AL. Loss of “complexity” and aging: potential applications of fractals and chaos theory to senescence. JAMA. 1992;267(13):1806–1809.
- Goldberger AL, Peng C-K, Lipsitz LA. What is physiologic complexity and how does it change with aging and disease? Neurobiol Aging. 2002;23(1):23–26. [CrossRef]
Figure 1.
Thermophysiological framework for bioenergetic aging. Deep-tissue radiofrequency (RF) radiometry captures bioenergy emission associated with metabolic activity and vascular heat transport, whereas infrared (IR) radiometry reflects surface energy dissipation. Aging is associated with a progressive reduction in deep-tissue bioenergy emission and altered deep–surface energetic gradients, providing a physiologically interpretable basis for biological age estimation.
Figure 1.
Thermophysiological framework for bioenergetic aging. Deep-tissue radiofrequency (RF) radiometry captures bioenergy emission associated with metabolic activity and vascular heat transport, whereas infrared (IR) radiometry reflects surface energy dissipation. Aging is associated with a progressive reduction in deep-tissue bioenergy emission and altered deep–surface energetic gradients, providing a physiologically interpretable basis for biological age estimation.
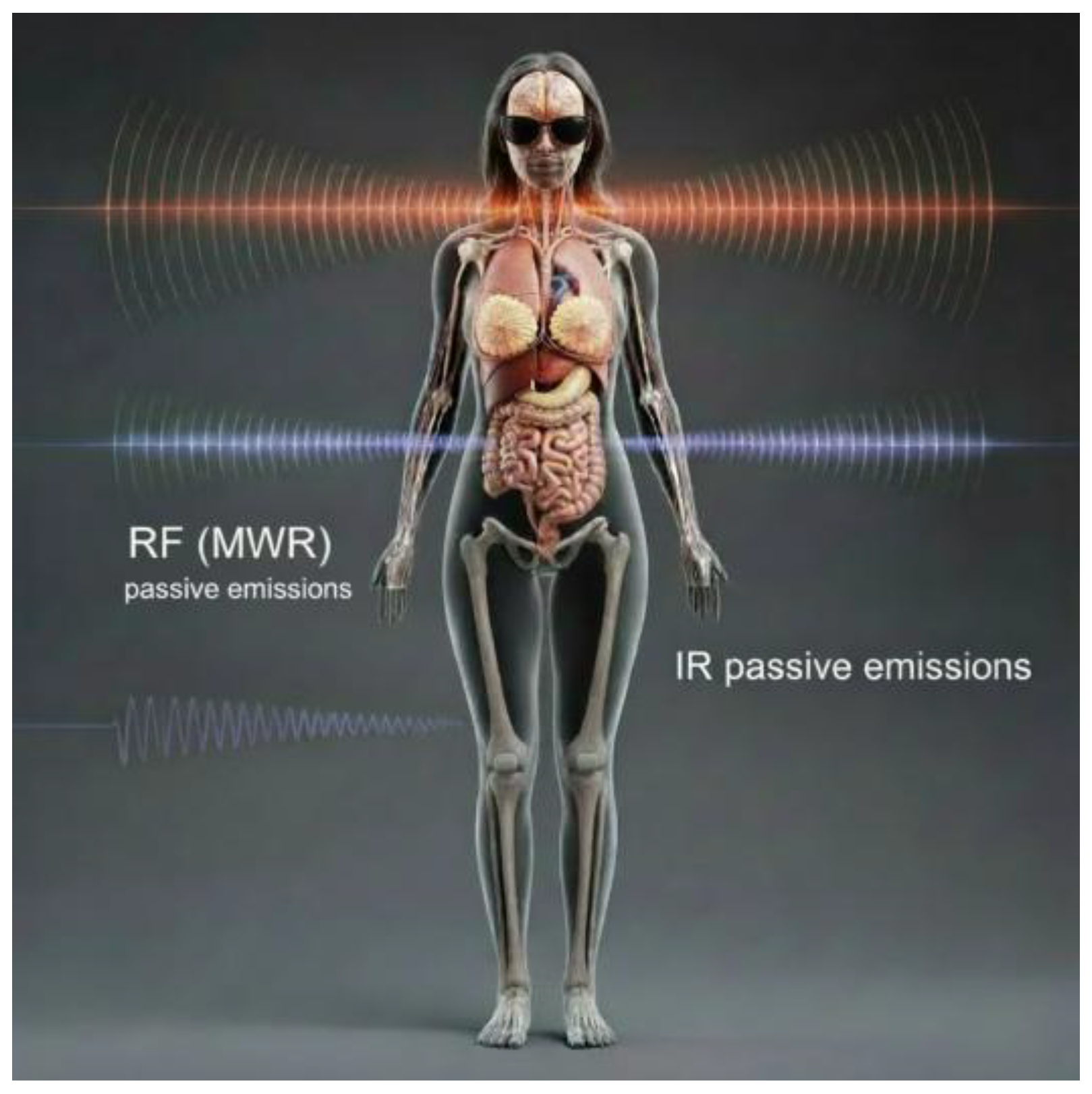
Figure 2.
MWR2025. Bioenergy monitor.

Figure 4.
Bioenergetic Index (BEI) across age. Mean ± SEM BEI values by age bin confirm normalisation behaviour and post-65 stabilisation.
Figure 4.
Bioenergetic Index (BEI) across age. Mean ± SEM BEI values by age bin confirm normalisation behaviour and post-65 stabilisation.
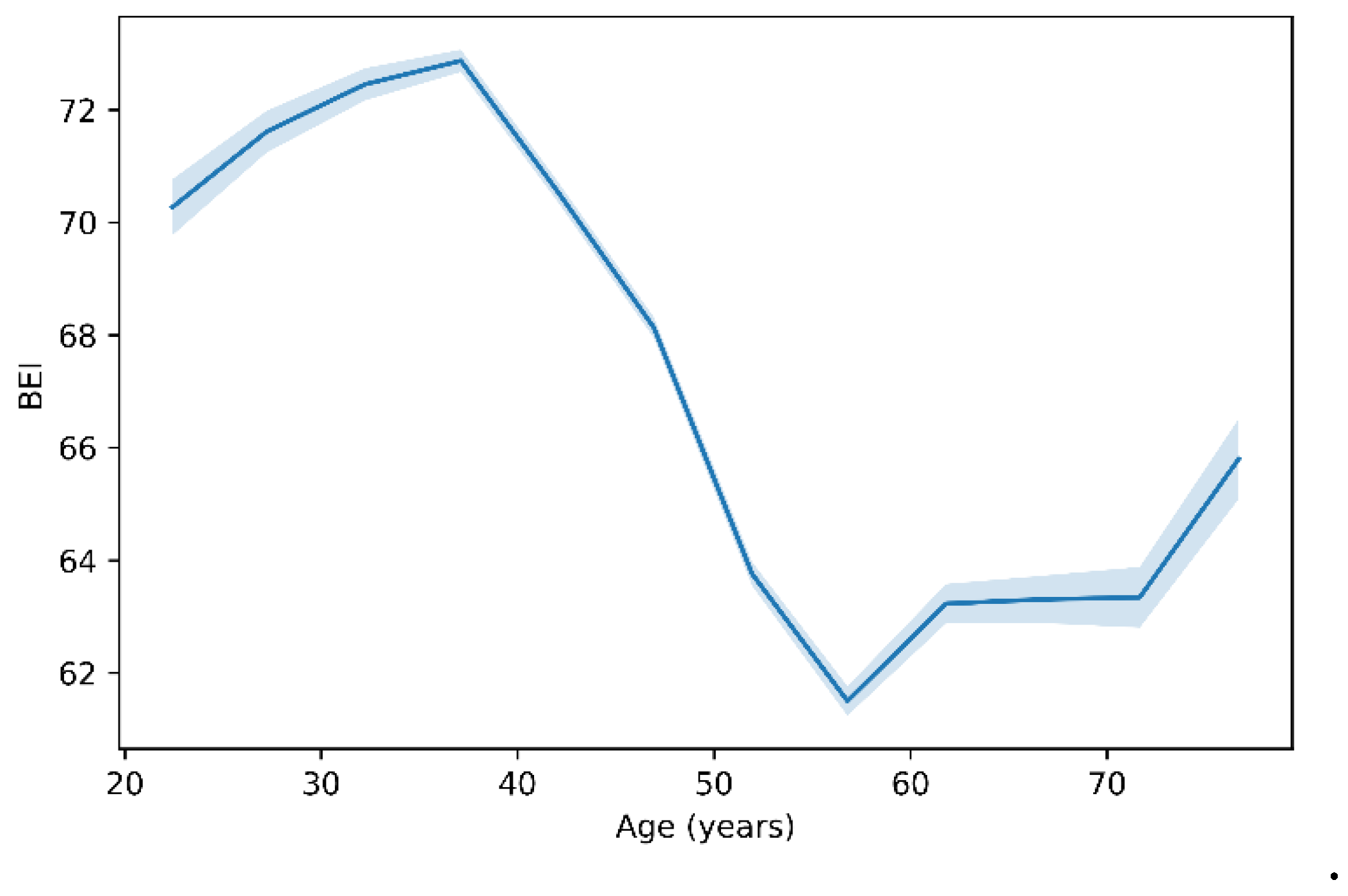
Figure 5.
The predicted individual biological age against chronological age. Model performance metrics RMSE ≈ 5.37 y, MAE ≈ 3.53 y, R2 ≈ 0.757.
Figure 5.
The predicted individual biological age against chronological age. Model performance metrics RMSE ≈ 5.37 y, MAE ≈ 3.53 y, R2 ≈ 0.757.
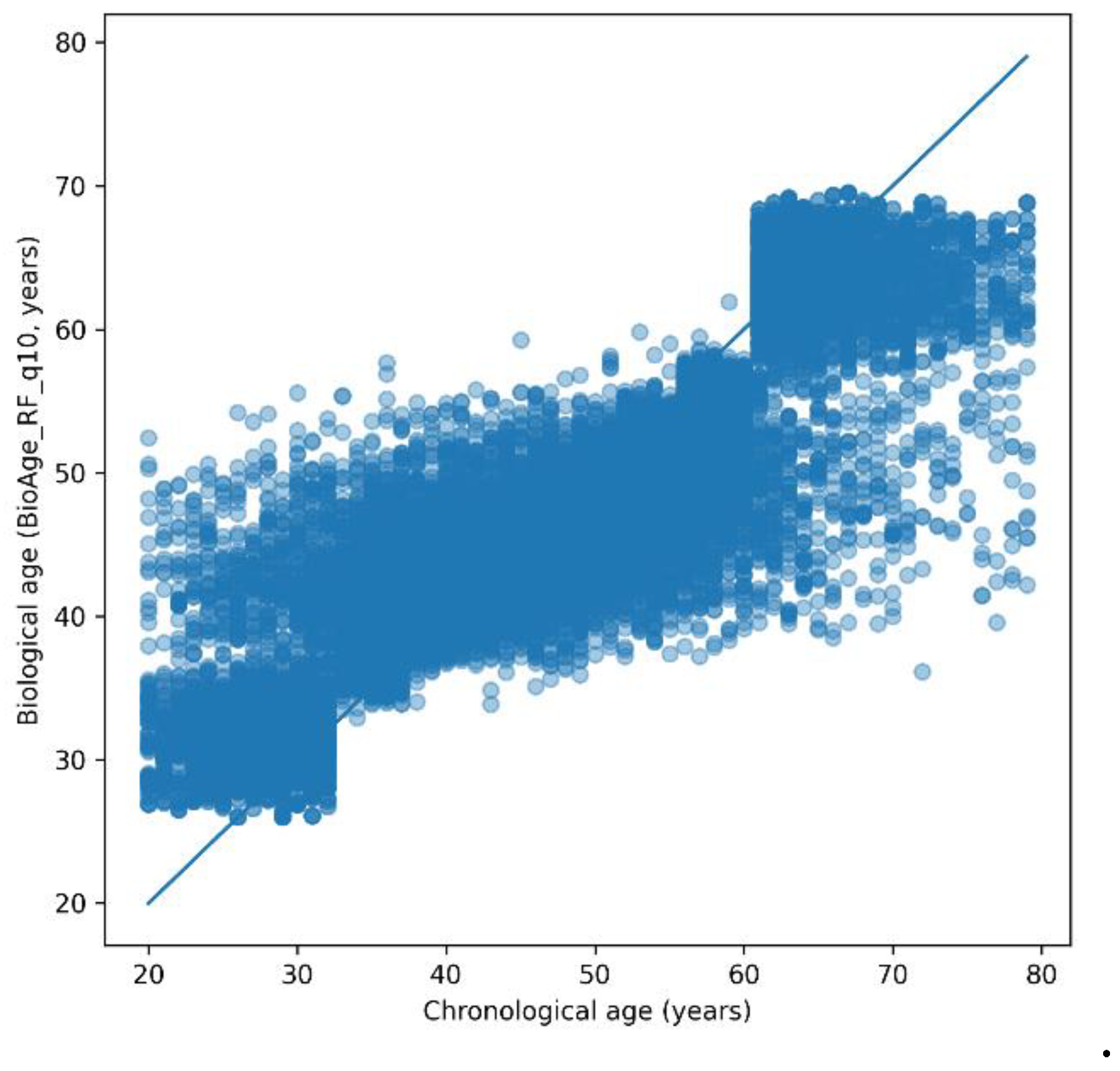
Figure 6.
The predicted ages and the true ages and aggregated to 5-year age bins.
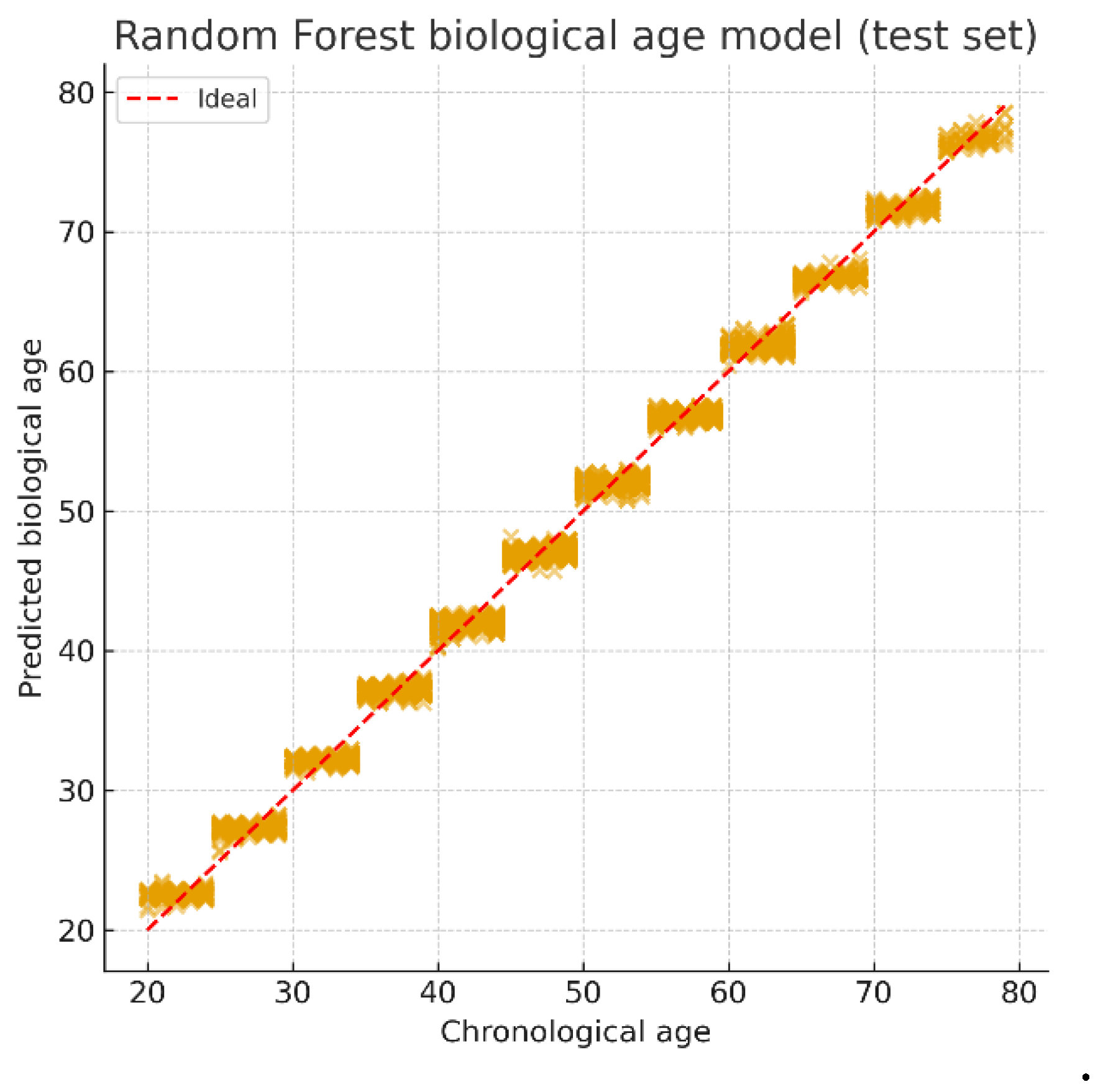
Figure 7.
The predicted individual biological age against chronological age using ensembled MWR-NetAge. Model performance metrics: RMSE ≈ 4.82 y, MAE ≈ 3.38 y, R2 ≈ 0.801.
Figure 7.
The predicted individual biological age against chronological age using ensembled MWR-NetAge. Model performance metrics: RMSE ≈ 4.82 y, MAE ≈ 3.38 y, R2 ≈ 0.801.
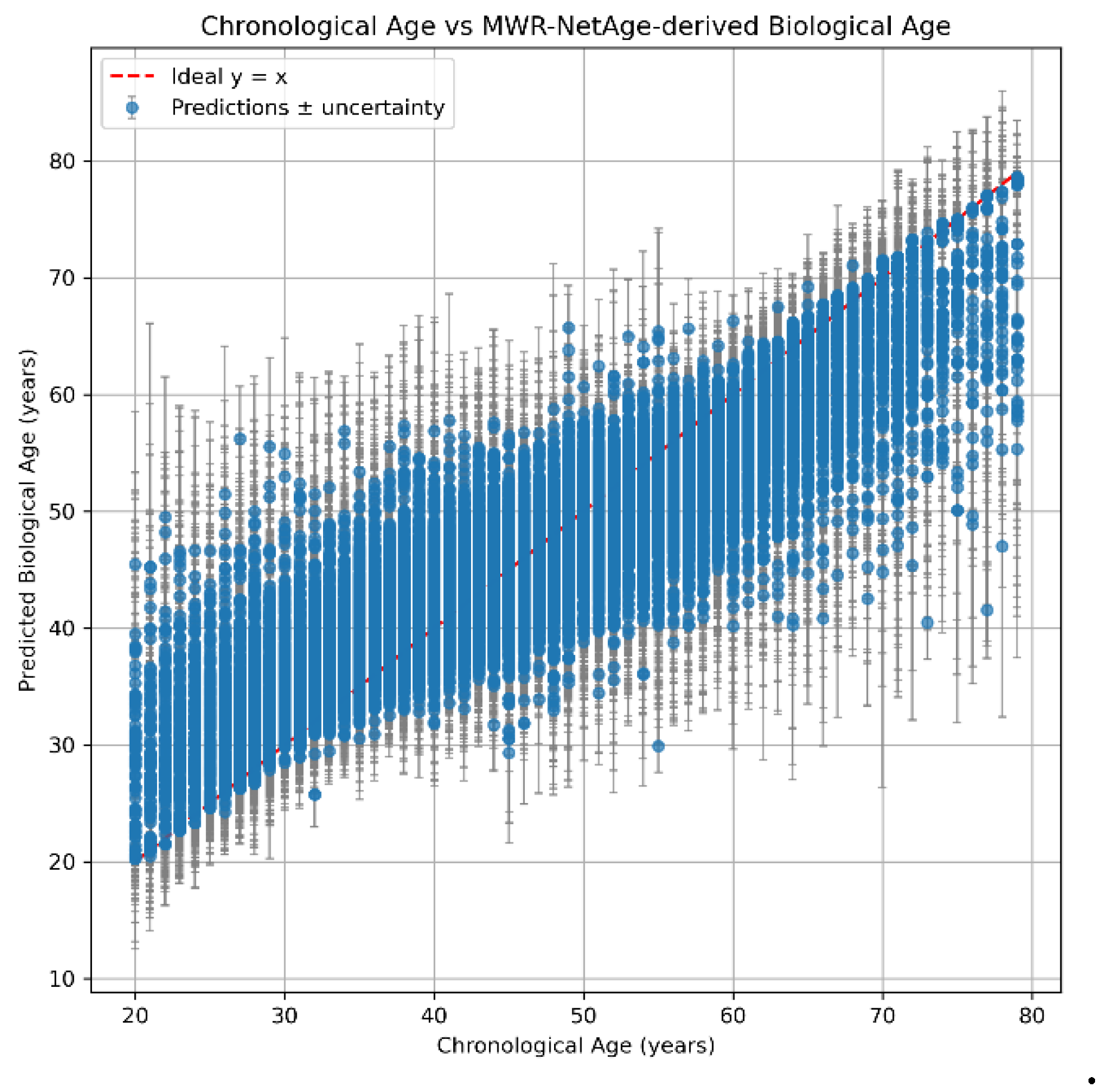
Figure 8.
The predicted 5-year age bins biological age against chronological age using ensembled MWR-NetAge. Model performance metrics: RMSE ≈ 1.21 y, MAE ≈ 1.02 y, R2 ≈ 0.987.
Figure 8.
The predicted 5-year age bins biological age against chronological age using ensembled MWR-NetAge. Model performance metrics: RMSE ≈ 1.21 y, MAE ≈ 1.02 y, R2 ≈ 0.987.
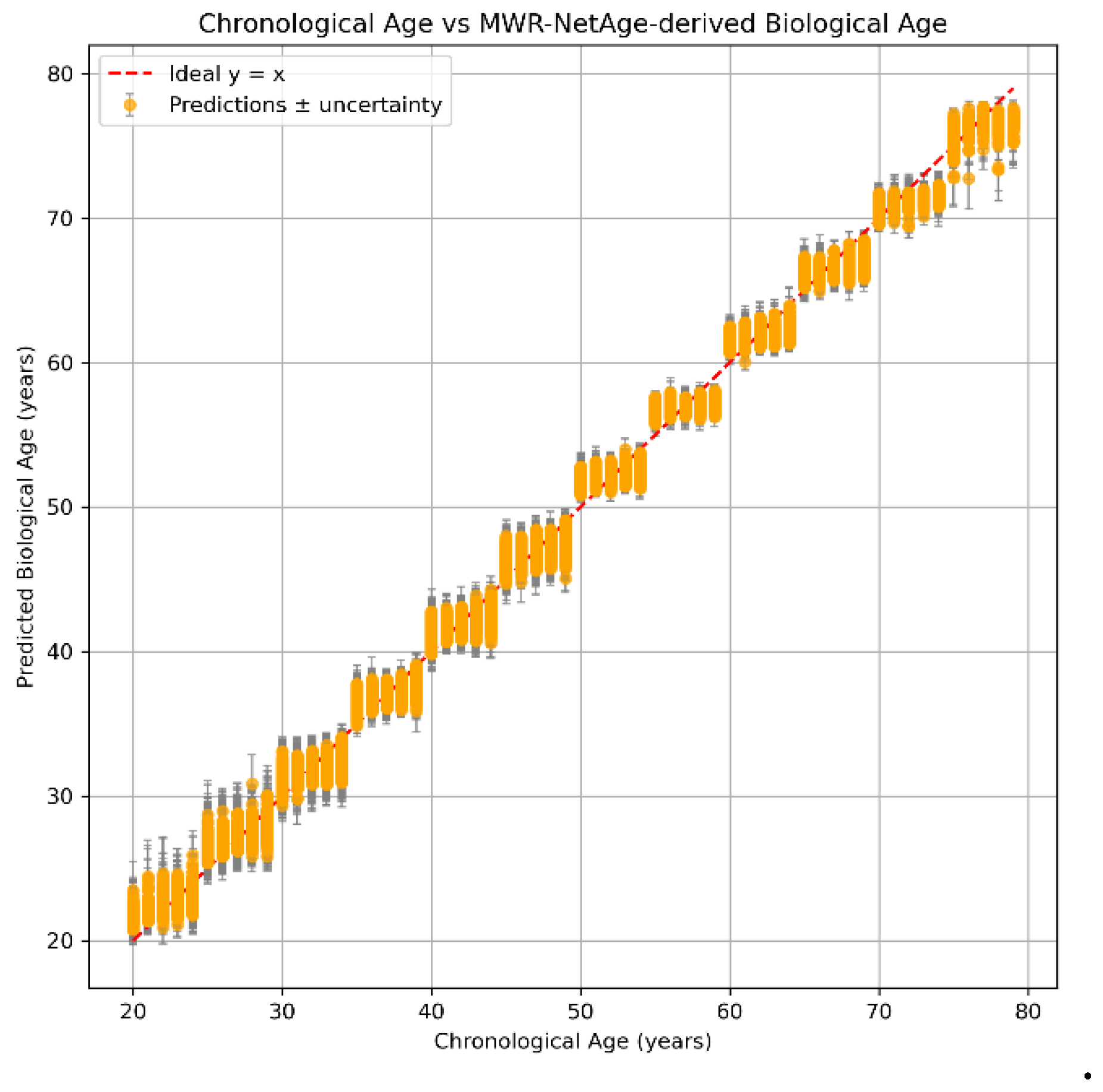
Figure 9.
Translational applications of thermophysiological biological age. Non-invasive estimation of biological age from energetic signatures enables patient stratification, longitudinal monitoring, and integration into AI-driven drug discovery and translational research.
Figure 9.
Translational applications of thermophysiological biological age. Non-invasive estimation of biological age from energetic signatures enables patient stratification, longitudinal monitoring, and integration into AI-driven drug discovery and translational research.
Table 1.
Performance of Random Forest models trained and evaluated within age strata (<30, 30–65, ≥65 years). Stratified models highlight distinct thermophysiological regimes across the lifespan but are not intended to optimise age prediction within narrow age ranges. Because chronological age variance is intentionally restricted within each stratum, the coefficient of determination (R2) becomes unstable and is not an appropriate measure of predictive quality in this context; correspondingly low R2 values are expected. Root mean square error (RMSE) provides a more informative metric within strata and indicates meaningful age-associated structure despite compressed target variance.
Table 1.
Performance of Random Forest models trained and evaluated within age strata (<30, 30–65, ≥65 years). Stratified models highlight distinct thermophysiological regimes across the lifespan but are not intended to optimise age prediction within narrow age ranges. Because chronological age variance is intentionally restricted within each stratum, the coefficient of determination (R2) becomes unstable and is not an appropriate measure of predictive quality in this context; correspondingly low R2 values are expected. Root mean square error (RMSE) provides a more informative metric within strata and indicates meaningful age-associated structure despite compressed target variance.
| Age group. | N (subjects) | RMSE (years) | R2 |
|---|---|---|---|
| < 30 years | 9,845 | 4.92 | 0.003 |
| 30–65 years | 24,464 | 6.18 | 0.081 |
| ≥ 65 years | 2,081 | 3.81 | 0.009 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



