
Submitted:
04 February 2026
Posted:
05 February 2026
You are already at the latest version
Abstract
The method of X-ray Footprinting and Mass Spectrometry (XFMS) using high brightness synchrotron X-ray sources has become an established method in structural biology and is based on the radiolytic production of hydroxyl radicals which oxidatively modify protein sidechains. While other methods of producing hydroxyl radicals are available, one benefit of using high flux density sources is that hydroxyl radical scavenging reactions can be minimized, and exposure times kept short to minimize secondary reactions. Here we present an application of the XFMS method using low dose rate X-rays from a commercial instrument. We demonstrate the feasibility of the approach using short peptides, characterizing the oxidative modifications +14, +16, and +32 Da under both aerated and low-oxygen conditions, and we additionally quantify the hydrogen peroxide production for various doses using the low dose rate source. These results provide fundamental information on the oxidative damage to peptides due to hydroxyl radicals using a low dose rate X-ray source.
Keywords:
hydroxyl radical footprinting
; peptide oxidation
; X-ray methods
1. Introduction
With the structural biology method of X-ray footprinting mass spectrometry (XFMS), hydroxyl radicals (•OH) generated by the radiolysis of water produce oxidative modifications to biomolecules in solution. Peptide modifications are typically quantified using liquid chromatography mass spectrometry (LCMS), while further localization of the modifications to specific residues can be determined with MS2 analysis. The map of modifications yields a solvent accessibility map, which is in turn used to determine structural information, such as interaction regions between proteins and their binding partners [1,2,3], determination or validation of antibody-antigen binding sites [4] structural changes during protein complex formation [5,6], or changes in three-dimensional structure during folding or unfolding events [7,8].
Footprinting has also been explored with ionizing radiation sources other than X-rays, such as high energy electrons [9] or gamma-rays [10]. In addition, non-radiolytic methods of labeling proteins, such as using Fenton chemistry [11], are also common. Another high flux density method is the UV-laser-induced •OH labeling method known as Fast Photochemical Oxidation of Proteins (FPOP) [12], which relies on the dissociation of H2O2 to produce hydroxyl radicals. While there are many methods which rely on solvent accessibility as a means of determining structure, XFMS does not require the addition of extraneous reagents, and the irradiation times can be on the order of microseconds. However, to the best of our knowledge, the XFMS method has not yet been explored using low dose rate X-rays.
Fundamentally, the initial interaction of a photon with a water molecule is independent of dose rate: either an electron is ejected from a water molecule (photoelectric effect) or the water molecule goes into an excited state [13]. For the two national XFMS programs – one at the Advanced Light Source (ALS) at Lawrence Berkeley National Laboratory [14] and the other at the National Synchrotron Light Source II (NSLS-II) at Brookhaven National Laboratory [15] – the X-rays are high enough energy that the photoelectric effect dominates. After this initial event, the flux density and the linear energy transfer characteristics of the beam, as well as the concentration of dissolved O2 in solution, can affect the further evolution of radical species and their distribution in solution [16,17]. Therefore, we expect the subsequent radical-radical recombination to be different for low and high dose rate irradiation rates, and this in turn would affect the concentration of products such as solvated electrons, •OH, superoxide, and H2O2 during irradiation. Because these products react with protein side chains to varying degrees [18,19], we might expect to see differences in the amount and types of modification with low dose rate XFMS as compared to the traditional high dose rate XFMS method.
Several previous studies have tabulated modifications to short peptides or single amino acids using various radiation sources [20,21,22]. Here we report on the first use of low dose rate X-rays to generate oxidative modifications to short peptides under conditions used in the XFMS method, namely micromolar concentrations of peptide in phosphate buffer. We examine modification as a function of oxygen availability, and report on the generation of H2O2 in solution as a function of dose.
2. Materials and Methods
Peptide preparation
Peptides were synthesized and purchased from GenScript in lyophilized form and guaranteed to ≥ 98% purity. They were dissolved in 1X phosphate buffer (VWR Chemicals, USA) to a concentration of 10 μM. Samples described as aerobic were prepared under ambient conditions. Samples described as low-oxygen were prepared in a Coy Vinyl Anaerobic Airlock Chamber (Coy Laboratory Products Inc, USA), where each peptide solution, before aliquoting, was bubbled with N2 gas for three minutes to achieve an oxygen concentration of <0.1%, as measured by a FireSting-GO2 oxygen meter (PyroScience, Germany). All samples were aliquoted into 0.25 mL PCR tubes containing 30 μL of peptide solution in each. Twenty-four samples per peptide were prepared (4 radiation doses in triplicate under aerobic and anaerobic conditions). Before transportation, while samples were still in the anaerobic chamber, samples were double-bagged in plastic bags with zip closures; they were first bagged by dose and then again by peptide, both for organizational purposes and to keep samples under anaerobic conditions for as long as possible. Aerobic samples were not bagged. All samples were transported on ice. A parallel test sample was prepared anaerobically as described above, and then allowed to sit outside the anaerobic chamber for 15 minutes, corresponding to the maximum amount of time samples experienced being outside the plastic bags while being irradiated in the XRAD instrument. The test sample was then brought back into the anaerobic chamber and tested for oxygen, which measured at < 0.5%.
Peptide Irradiation
The samples were irradiated at Lawrence Berkeley National Laboratory (LBNL) using the 320-kVp XRAD320 X-ray machine (Precision X-ray, USA). The machine was operated at 300 kVp and 10 mA, with a 0.5 mm Cu beam hardening filter in place during irradiation. Samples were placed on a rotating platform in a circle to be equidistant from the center and irradiated with a dose rate of 0.020 Gy/s at varying times to achieve doses of 5, 10, and 15 Gy, as previously described [23]. Dosimetry was performed with a RadCal 10X6-0.18 ion chamber (RadCal, USA), where a single point ion chamber measurement was taken in the middle of the platform to verify the doses delivered. Following irradiation, samples were immediately moved to a -80°C freezer to minimize any ongoing modification resulting from secondary reactions and longer-lived radical species.
LC-MS/MS and data analysis
Peptides were analyzed using a Vanquish Flex UHPLC system coupled to an Orbitrap Exploris 480 mass spectrometer (ThermoFisher Scientific Inc, USA). Separation was performed on a Thermo Scientific Hypersil GOLD Peptide C18 column (2.1 x 100 mm, 1.9 µm particle size, maintained at 55°C) at a flow rate of 0.400 mL/min with a mobile phase consisting of solvent A (0.1% formic acid in water) and solvent B (99.9% acetonitrile, 0.1% formic acid in water). The injection volume was 10 µL and peptides were eluted with the following gradient: initial condition was 1% solvent B for a 3 min wash step with flow diverted to waste, after which solvent B was increased to 10% over 1.5 min, then increased to 35% over 10 min followed by a ramp-up to 80% over 0.5 min, and held at 80% solvent B for 1.5 min at a flow rate of 0.600 mL/min. The flow rate was reduced to 0.400 mL/min at 2% solvent B for column equilibration. The mass spectrometer was operated in positive mode with full scan parameters set to an Orbitrap resolution of 60,000; mass range 300-1200 m/z; RF lens at 50%; AGC target at 3.0e6; maximum injection time 60 ms. The top 10 most intense ions per MS scan were selected for HCD fragmentation with an intensity threshold of 5.0e3 and dynamic exclusion set at 2 s. Data-dependent MS2 scan parameters included an isolation window of 2 m/z; 30% normalized collision energy; Orbitrap resolution set to 15,000; AGC target at 1.0e5; maximum injection time 50 ms.
Peptide analysis was performed using the Byos® software platform from Protein Metrics Inc., which combines the Byonic™ MS/MS search engine and the Byologic® peptide analysis software. The Oxidative Footprinting workflow has been customized to search for, identify, and quantify peptide modifications. Peptide modifications are reported as a percentage of modified peptide relative to unmodified (or native) peptide based on their respective extracted ion chromatogram (XIC) peak areas, and is calculated by the following formula:
Hydrogen peroxide assay
AmplexRed (ThermoFisher Scientific Inc, USA) was used according to manufacturer protocol for quantification of H2O2 in solution. Milli-Q water samples were prepared in triplicate. For low-oxygen conditions, the samples were brought into a Coy Vinyl Anaerobic Airlock Chamber (Coy Laboratory Products Inc, USA) and N2 gas was bubbled through samples for 5 minutes, after which O2 levels were measured using a FireSting-GO2 oxygen meter (PyroScience, Germany). All samples measured less than 1% O2 when bagged for transport to the XRAD instrument, as previously described [24]. After irradiation, 50 µl of each sample was pipetted into a 96-well plate, and 50 µl of the working buffer (AmplexRed solution) was added to each well. A BioTek Synergy H1 plate reader (Agilent, USA) was used to measure fluorescence, with excitation at 530 nm and emission measured at 590 nm. Hydrogen peroxide calibrants were prepared fresh for each experiment and used to generate a standard curve.
H2O2 was also used in concentrations ranging from 1 µM to 10 mM and incubated directly with the peptide for the amount of time corresponding to the maximum dose used in the peptide irradiation experiments. LCMS/MS was then conducted as described above.
3. Results
3.1. Measurement of Hydrogen Peroxide Due to Radiolysis
Radiolysis of aqueous solutions results in a complex ROS environment composed of many species, including electrons, hydroxyl radicals, super oxide, hydrogen ions, hydroperoxyl radicals, and hydrogen peroxide [25,26] (Table S1). Simulations have been used to predict the evolution and lifetime of various species under different dose rates and radiation types, and experimental methods to quantify yields of specific species have included fluorescence readouts, EPR spin trapping, and scavenging reactions [16,27,28]. Hydrogen peroxide, although one of the least reactive species to proteins, is stable and long-lasting, can be measured after irradiation using the standard fluorescence assay AmplexRed (ThermoFisher Scientific Inc, USA). Using this assay, previous studies using electron, carbon ion, and proton sources have shown that under irradiation, samples low in dissolved oxygen will produce less H2O2 than fully aerated samples [16,29,30,31,32]. However, results are complicated by the fact that the dose rate and LET value of the radiation will affect the production and evolution of radiolytic species. The presence of radical scavengers and dissolved gases such as CO2 will also affect the final yield of H2O225, and simulations have produced conflicting results[33,34,35,36].
Because of the long exposure times necessary when using a low dose rate source, and the corresponding potential buildup of H2O2, we characterized the H2O2 yield for the exposures used in this study under both aerobic and low-oxygen conditions. For a given dose, aerated milli-Q produced higher yields of H2O2 than low-oxygen samples under X-ray irradiation, with higher doses leading to a larger difference in final yield (Figure 1). We also compared milli-Q water with 10 mM Tris buffer and 10 mM Phosphate buffer and observed no differences in H2O2 yield (Table S2). To further test the effect of the ROS environment on protein side chains, we then designed short peptides and characterized their oxidation under X-ray irradiation with different doses and two different dissolved oxygen conditions as described below.
3.2. Peptide Design
We designed short peptides for this study in order to minimize secondary structure. Single amino acids have varying reactivities to hydroxyl radicals, with the highest reactivity observed for the sulfur-containing residues and the aromatic residues, and the lowest observed for Gly. We therefore kept the sequence of the peptides identical except for one residue, which varied an order of magnitude in reactivity. We also included specific residues to maintain solubility of the peptide, and hydrophobic residues to ensure good LC coverage. The specific sequence was EDLAXLK, in which X represents residues F, G, H, I, M, P, R, or Y.
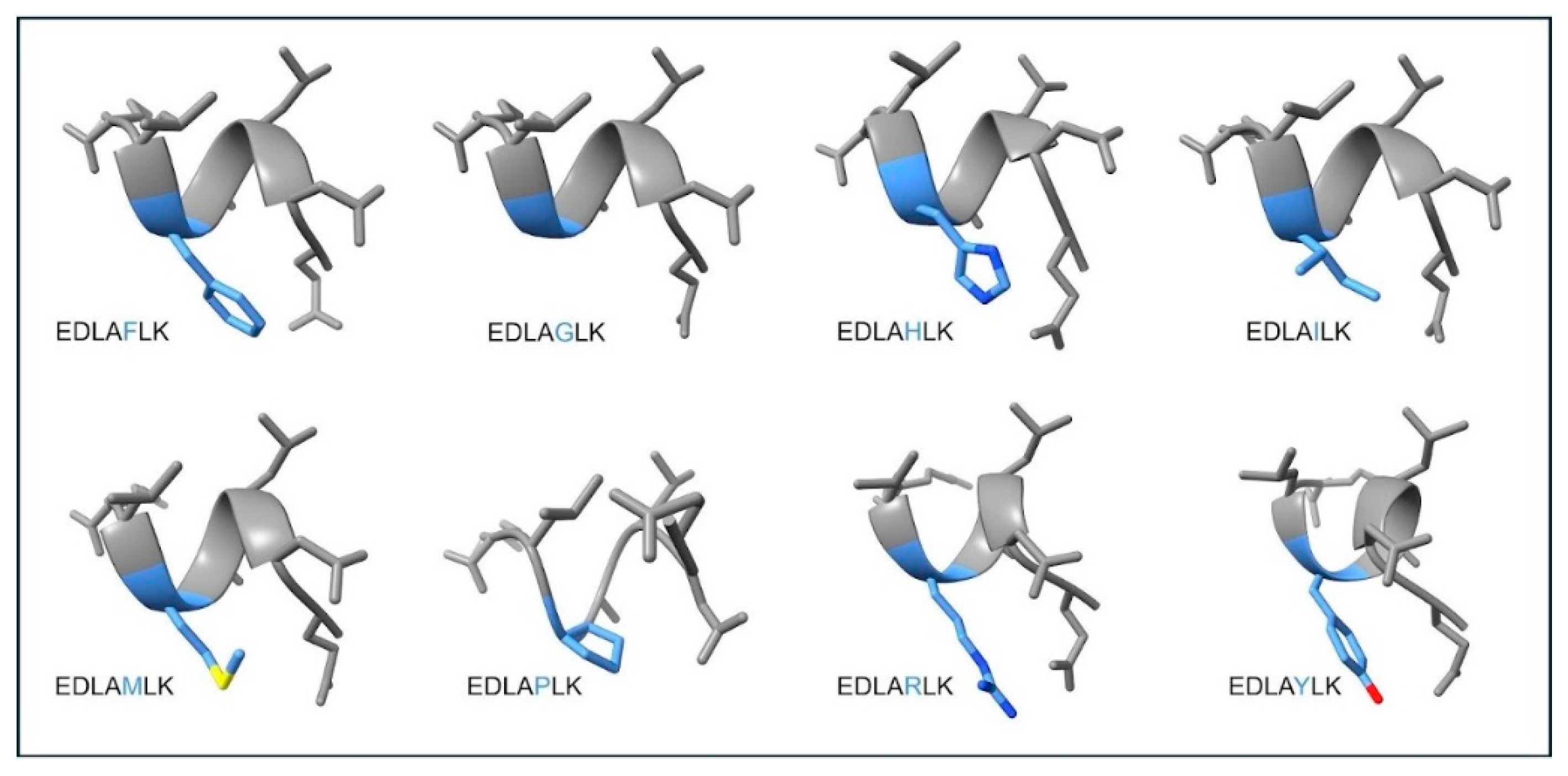
We ran these eight peptides through the PEP-FOLD3 server [37] to predict their three-dimensional structures (Figure 2). The top five models in each case had nearly identical sOPEP scores, which ranged by peptide from -4.59 to -9.17.
To test whether H2O2 contributed to oxidative modification, we subjected the most reactive peptide (EDLAMLK, or peptM) to H2O2 in a concentration similar to that determined using the AmplexRed assay as well as higher concentrations of H2O2. At 1 µM concentration, we observed no increase in oxidation on peptM over background oxidation, and only at concentrations of 1 mM and higher did we observe any significant oxidation over background (Table S3). While Met can be oxidized directly by H2O2, the reaction rate is ~10−2 M−1s−1 [19] which is 7-8 orders of magnitude lower than the reaction rate with hydroxyl radical11.
Another mechanism by which oxidation of peptides and proteins in solution might occur is through hydroxyl radicals generated by reaction of H2O2 with trace metals in solution [38]. However, because we did not observe oxidation of peptM in solution containing H2O2 at the concentrations generated during X-ray irradiation, we conclude that oxidation of the peptides in this study was primarily due to attack by hydroxyl radicals. This is also the assumption in typical XFMS experiments using high dose rate X-rays [14].
3.3. Peptide Oxidation Under Aerated Conditions
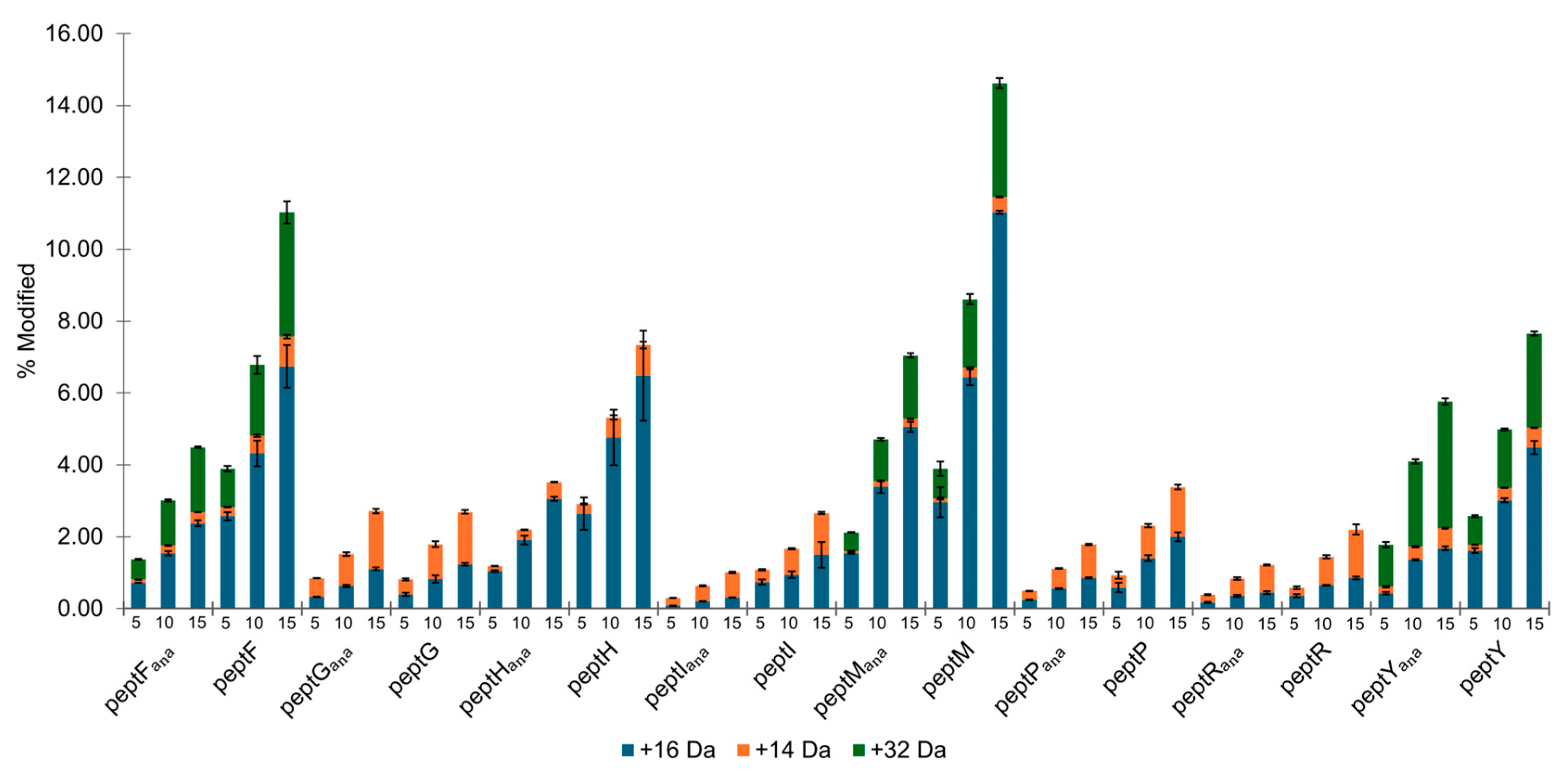
Figure 3 and Table S4 show percent modification for each peptide as a function of dose (5, 10 and 15 Gy), oxygen availability, and type of oxidative modification (+14, +16, and +32 Da). The +16 Da modification is a hydroxylation and is the most common modification reported in XFMS and FPOP studies[39]. However, reaction mechanisms for the various types of modifications are distinct for each amino acid and can include the +14, +32 modifications in addition to higher mass additions and some subtractions. The +14 Da oxidation, referred to as a carbonyl modification, results from formation of a C=O bond after hydroxyl radical abstraction of an H from a saturated carbon, and subsequent attack by O2. The +32 Da modification in this study refers to a double hydroxyl addition at two different carbons within a single residue, or in the case of Met, to a double oxidation of the sulfur resulting in a sulfone.
In this study, modification of a given type (+14, +16, or +32 Da) ranged from 0.05-11%, with modification increasing linearly with dose. For high dose rate XFMS, exposures are also chosen to produce a linear dose response, and are kept deliberately low to reduce unfolding of the protein due to oxidation[14]. Here, the highest amount of oxidation (5-11%) was observed for the +16 Da oxidation on the peptides containing Met or an aromatic ring (the peptF, peptH, and peptY peptides). For these peptides, the +16 Da modification was on the order of 10-fold higher than the +14 Da modification and approximately double that of the +32 Da modification. For all the peptides characterized in this study, the +14 Da modifications ranged from 0.5 to 1.5% for the highest X-ray dose. Of the peptides that did not contain S or an aromatic ring, the hydroxylation modification was significantly lower (0.8-1.5% at the highest X-ray dose), and in the case of the peptG and peptR peptides, on the order of or even lower than the carbonyl modification. Previous studies have shown that depending on sequence, the oxidation of short peptides can result in higher carbonyl products than hydroxyl products, and that even a Gly residue can exhibit carbonyl modification (technically a backbone modification) in substantial amounts[20]. We also found that the amount of hydroxylation of peptF is higher than hydroxylation of peptY (6.7 vs 4.5 %). While Phe has been reported to have a lower reactivity to hydroxyl radicals than Tyr when in the free amino acid form[13], other studies have shown that the yield of hydroxylated Phe due to hydroxyl radical attack can increase when in a short peptide20.
Figure 3.
Peptide-level percent modified by modification type and radiation dose. Organized by peptide, where peptX (X representing the fifth position residue which distinguishes each peptide) refers to aerobically prepared samples, and peptXana refers to anaerobically prepared samples. For each peptide, from left to right, percent modified is shown for each dose: 5, 10, and 15 Gy. As this data has been averaged and normalized to 0% modification at 0 Gy, that dose is not shown on this plot. Error bars represent the standard deviation between triplicate data points.
Figure 3.
Peptide-level percent modified by modification type and radiation dose. Organized by peptide, where peptX (X representing the fifth position residue which distinguishes each peptide) refers to aerobically prepared samples, and peptXana refers to anaerobically prepared samples. For each peptide, from left to right, percent modified is shown for each dose: 5, 10, and 15 Gy. As this data has been averaged and normalized to 0% modification at 0 Gy, that dose is not shown on this plot. Error bars represent the standard deviation between triplicate data points.
3.4. Peptide Oxidation Under Low Oxygen Conditions
When prepared and irradiated under low oxygen, the peptides generally exhibited lower oxidation at each dose relative to the aerobically prepared peptides, as has been previously observed in high dose rate XFMS [4]. The +16 Da oxidation ranged from 0.3 to 5%, with the highest yields observed on peptM and the peptides containing an aromatic ring (Table S4), similar to the trend for the aerobic peptides.
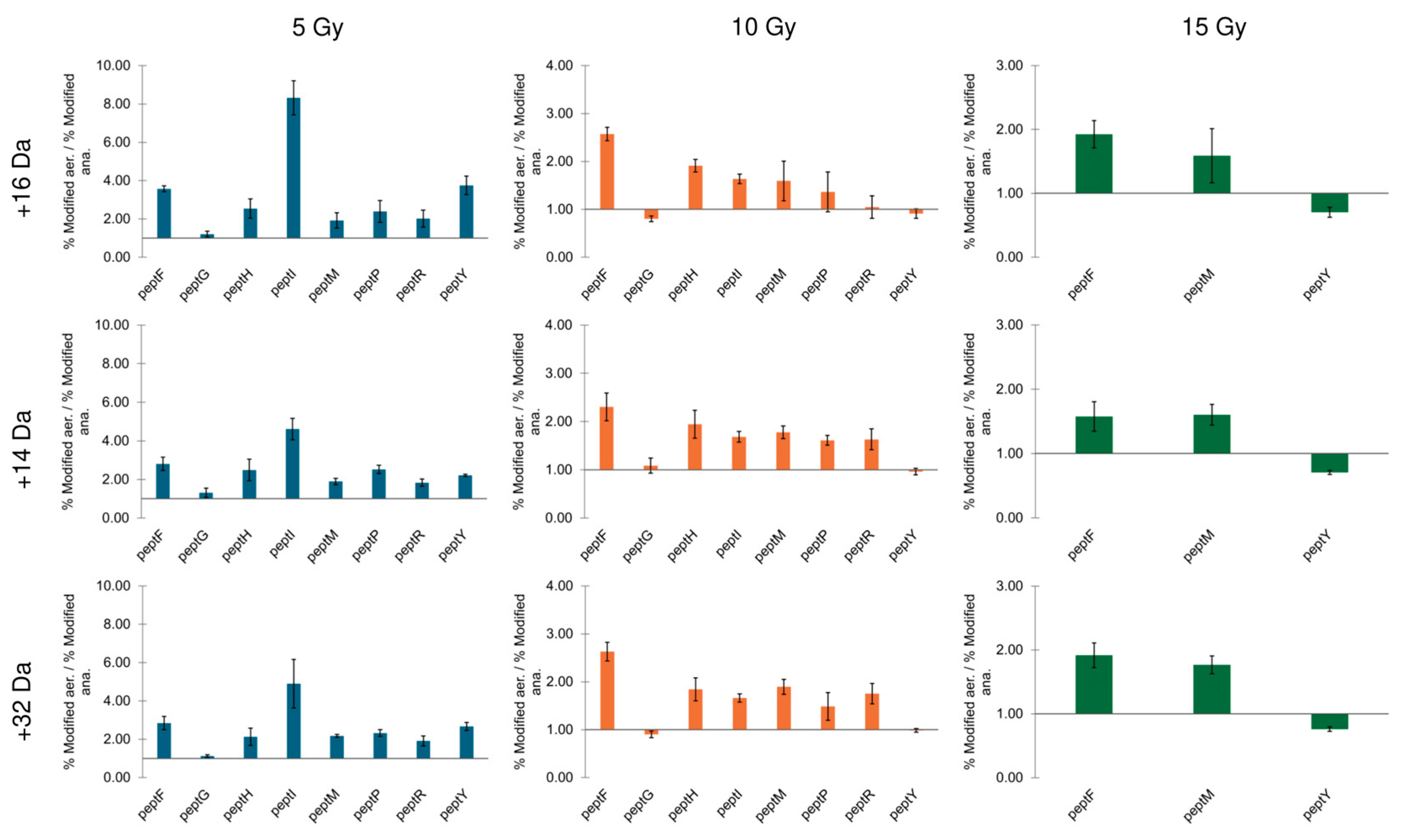
The calculated ratio of percent oxidation for aerobic versus anaerobic preparation for each peptide in this study are shown in Figure 4, separated by modification type and dose. The general pattern of oxidation does not change with the dose (5, 10, or 15 Gy). For most of the peptides and modification type, ratios were above one, indicating more oxidation under aerobic conditions, with some exceptions. One notable exception is peptY, which showed more +32 Da oxidation under low oxygen conditions. The yield of the first hydroxylation of peptY is comparable to the double hydroxylation of peptF: For peptY, 4.5% (aerated) to 1.7% (low-oxygen), and for peptF, 3.8% (aerated) to 1.8% (low oxygen). However, the second hydroxylation of peptY is 2.7% (aerated) to 3.5% (low oxygen). The reaction scheme for hydroxylation of an aromatic ring has been delineated in several sources [13] and begins with a direct attack by a hydroxyl radical on the aromatic ring. The resulting carbon-centered radical is in turn susceptible to attack by O2, which is followed by hydroperoxide elimination, leaving the hydroxylated form of the amino acid. Previous studies on free amino acids or three-residue peptides have found that in the absence of O2, the final hydroxylated product is still formed, but in lower yield, presumably because the hydroxylated radical intermediate is not stable [13]. A previous study using O18 labelling showed that oxidation of Phe and Tyr could occur through multiple mechanisms, not all involving oxygen [40]. We have seen in a previous study [24] that some Phe residues in a protein show increased hydroxylation under low-oxygen conditions, while others do not, indicating that the local environment plays a significant role in hydroxylation of an aromatic ring. It is possible that for the peptides used in this study, the neighboring residues play a role in stabilizing the initial OH addition to the ring, mediating the dependency on O2.
Figure 4.
Ratios of peptide-level percent of modification under aerobic to low-oxygen conditions, separated by type of modification and dose. The blue bars (top row) show only +16 Da, the orange bars (middle row) show only +14 Da, and the green bars (bottom row) show only the +32 Da modification. From left to right, columns are 5, 10, and 15 Gy exposures. Error bars represent the standard deviation between triplicate data points.
Figure 4.
Ratios of peptide-level percent of modification under aerobic to low-oxygen conditions, separated by type of modification and dose. The blue bars (top row) show only +16 Da, the orange bars (middle row) show only +14 Da, and the green bars (bottom row) show only the +32 Da modification. From left to right, columns are 5, 10, and 15 Gy exposures. Error bars represent the standard deviation between triplicate data points.
4. Discussion
Free amino acids have been demonstrated to react at different rates to different hydroxyl radicals, H2O2 and electrons with the reactivities of the amino acids specifically to hydroxyl radicals ranked as follows: Tyr>Met>Phe>His>Arg>Ile>Leu>Pro>Lys>Glu>Ala>Asp>Gly [13]. Since the peptides used in this study differed only in their fifth residue, we assume that the change in peptide-level modification between the eight peptide sequences is due to the amino acid occupying the fifth position on the peptide. If we rank the peptides based on total peptide-level percent modified observed, then we list the peptides as follows: peptM>peptF>peptY>peptH>peptP>peptG>peptI>peptR. That this does not completely align with the ranking of reported reactivities of individual amino acids to hydroxyl radicals is perhaps not surprising, given that the local sequence of a residue can alter peptide reactivity/modification, as has been noted in some previous studies[20,21].
Here we have shown that peptide modification under low dose rate X-ray irradiation using a commercial instrument (0.02 Gy/s) results in modification types of +14, +16, +32 Da, and amounts of up to 11% at a given dose. These are sufficient for XFMS analysis; however, it should be noted that the low dose rate source required much longer irradiation times than at high dose rate beamlines (minutes versus milliseconds) to achieve the same dose. For proteins under long irradiation times, we might expect to see secondary oxidation reactions accumulating, which would limit the structural information obtainable using a low dose rate source. We also measured the concentration of hydrogen peroxide resulting from low dose rate exposures in order to rule out the possibility that prolonged exposure to hydrogen peroxide formed during exposure might contribute to oxidation products. We found that for the highest dose (15 Gy), the H2O2 produced was 1.0-1.3 µM, depending on oxygen availability. Exposing the most reactive peptide, peptM, to up to 100 µM concentration of H2O2 for the longest exposure time (15 minutes) did not result in oxidation over background. In addition, we found that the production of H2O2 was similar in milli-Q water, Phosphate buffer, and Tris buffer. One of the mechanisms for production of H2O2 is the combination of two hydroxyl radicals, so it is noteworthy that the H2O2 yield in Tris, a known hydroxyl radical scavenger, was comparable to the yield in pure water or Phosphate buffer. This points to other H2O2-producing mechanisms possibly playing a greater role, such as the combination of hydroperoxyl radicals or ions with protons or hydrogen ions, all of which are produced under radiolysis of aqueous solutions (Table S1). However, the production of hydroperoxyls depends to some extent on the presence of O2, which may explain the lower H2O2 yield in solutions with low dissolved O2. Molecular oxygen also serves as an electron scavenger, and since electrons react with hydroxyl radicals to produce hydroxyl ions, reducing the available electrons in solution can increase the amount of hydroxyl radicals, possibly increasing in turn the yield of H2O2. These considerations are relevant for structural biology studies using the method of hydroxyl radical footprinting, since this method relies on hydroxyl radical induced modifications of protein sidechains, and is based on the assumption that protein modifications are not affected by H2O2 generated during radiolysis. The measurement of H2O2 yield under different oxygen levels is also relevant for radiation oncology, since the tumor environment is generally more hypoxic than healthy tissue, and H2O2 is one of the oxidatively damaging species produced during radiation. Future studies on the comparison of low dose rate irradiation versus high dose rate irradiation on peptides and proteins will be of interest to the radiation oncology field, in which the dose rate effect on damage to biomolecules is currently of great interest.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Radiolysis reactions and rate constants in aqueous buffer; Table S2: Production of hydrogen peroxide in water, Phosphate, and Tris buffers; Table S3: Percentage oxidation of EDLAMLK peptide under exposure to hydrogen peroxide; Table S4: Percent of modified peptide by modification type, radiation dose and oxygen availability.
Author Contributions
Conceptualization, S.K., S.S., S.G. and C.Y.R.; methodology, J.L.I, L.O.H. S.G., L.G.K., S.S., T.M. and C.Y.R.; validation, S.K. and L.G.K.; formal analysis, S.K., T.M. and L.G.K.; writing—original draft preparation, S.K.; writing—review and editing, S.K., S.G. and C.Y.R.; supervision, S.G. and C.Y.R.; project administration, C.Y.R.; funding acquisition, C.Y.R, J.L.I. and L.O.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported in part by National Institute of Health grants P30 GM124169 and 1R01GM126218 (C.Y.R.), and Laboratory Directed Research and Development funding (C.Y.R., J.L.I. and L.O.H.). Work conducted at the Advanced Light Source and the Molecular Foundry (Office of Basic Energy Sciences) was supported by the Office of Science of the U.S. DOE under contract DE-AC02-05CH1123. T.M. was supported by the Community College Internship program.
Data Availability Statement
All data are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| LCMS | Liquid Chromatography Mass Spectrometry |
| XFMS | X-ray Footprinting Mass Spectrometry |
References
- He, L.; McAndrew, R.; Barbu, R.; Gifford, G.; Halacoglu, C.; Drouin-Allaire, C.; Weber, L.; Kristensen, L. G.; Gupta, S.; Chen, Y.; et al. Structure and Interactions of HIV-1 gp41 CHR-NHR Reverse Hairpin Constructs Reveal Molecular Determinants of Antiviral Activity. J Mol Biol 2024, 436(16), 168650. [Google Scholar] [CrossRef]
- Dominguez-Martin, M. A.; Hammel, M.; Gupta, S.; Lechno-Yossef, S.; Sutter, M.; Rosenberg, D. J.; Chen, Y.; Petzold, C. J.; Ralston, C. Y.; Polivka, T.; Kerfeld, C. A. Structural analysis of a new carotenoid-binding protein: the C-terminal domain homolog of the OCP. Sci Rep 2020, 10(1), 15564. [Google Scholar] [CrossRef]
- Wheatley, P.; Gupta, S.; Pandini, A.; Chen, Y.; Petzold, C. J.; Ralston, C. Y.; Blair, D. F.; Khan, S. Allosteric Priming of E. coli CheY by the Flagellar Motor Protein FliM. Biophys J 2020, 119(6), 1108–1122. [Google Scholar] [CrossRef]
- Kristensen, L. G.; Gupta, S.; Chen, Y.; Petzold, C. J.; Ralston, C. Y. Residue-Specific Epitope Mapping of the PD-1/Nivolumab Interaction Using X-ray Footprinting Mass Spectrometry. Antibodies 2024, 13(3). [Google Scholar] [CrossRef]
- Sommer, M.; Sutter, M.; Gupta, S.; Kirst, H.; Turmo, A.; Lechno-Yossef, S.; Burton, R. L.; Saechao, C.; Sloan, N. B.; Cheng, X.; et al. Heterohexamers Formed by CcmK3 and CcmK4 Increase the Complexity of Beta Carboxysome Shells. Plant Physiol 2019, 179(1), 156–167. [Google Scholar] [CrossRef]
- Sutter, M.; Utschig, L. M.; Niklas, J.; Paul, S.; Kahan, D. N.; Gupta, S.; Poluektov, O. G.; Ferlez, B. H.; Tefft, N. M.; TerAvest, M. A.; et al. Electrochemical cofactor recycling of bacterial microcompartments. Proc Natl Acad Sci U S A 2024, 121(49), e2414220121. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Bavro, V. N.; D’Mello, R.; Tucker, S. J.; Venien-Bryan, C.; Chance, M. R. Conformational changes during the gating of a potassium channel revealed by structural mass spectrometry. Structure 2010, 18(7), 839–846. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S. Using X-ray Footprinting and Mass Spectrometry to Study the Structure and Function of Membrane Proteins. Protein Pept Lett 2019, 26(1), 44–54. [Google Scholar] [CrossRef] [PubMed]
- Watson, C.; Janik, I.; Zhuang, T.; Charvatova, O.; Woods, R. J.; Sharp, J. S. Pulsed electron beam water radiolysis for submicrosecond hydroxyl radical protein footprinting. Anal Chem 2009, 81(7), 2496–2505. [Google Scholar] [CrossRef]
- Hayes, J. J.; Kam, L.; Tullius, T. D. Footprinting protein-DNA complexes with gamma-rays. Methods Enzymol 1990, 186, 545–549. [Google Scholar] [CrossRef]
- Shcherbakova, I.; Mitra, S.; Beer, R. H.; Brenowitz, M. Fast Fenton footprinting: a laboratory-based method for the time-resolved analysis of DNA, RNA and proteins. Nucleic Acids Res 2006, 34(6), 48–49. [Google Scholar] [CrossRef] [PubMed]
- Sharp, J. S.; Becker, J. M.; Hettich, R. L. Analysis of protein solvent accessible surfaces by photochemical oxidation and mass spectrometry. Anal Chem 2004, 76(3), 672–683. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Chance, M. R. Hydroxyl radical-mediated modification of proteins as probes for structural proteomics. Chem Rev 2007, 107(8), 3514–3543. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Celestre, R.; Petzold, C. J.; Chance, M. R.; Ralston, C. Development of a microsecond X-ray protein footprinting facility at the Advanced Light Source. J Synchrotron Radiat 2014, 21 Pt 4, 690–699. [Google Scholar] [CrossRef]
- Asuru, A.; Farquhar, E. R.; Sullivan, M.; Abel, D.; Toomey, J.; Chance, M. R.; Bohon, J. The XFP (17-BM) beamline for X-ray footprinting at NSLS-II. J Synchrotron Radiat 2019, 26 Pt 4, 1388–1399. [Google Scholar] [CrossRef]
- Kusumoto, T.; Danvin, A.; Mamiya, T.; Arnone, A.; Chefson, S.; Galindo, C.; Peaupardin, P.; Raffy, Q.; Kamiguchi, N.; Amano, D.; et al. Dose Rate Effects on Hydrated Electrons, Hydrogen Peroxide, and a OH Radical Molecular Probe Under Clinical Energy Protons. Radiat Res 2024, 201(4), 287–293. [Google Scholar] [CrossRef]
- Sorensen, B. S.; Overgaard, J.; Bassler, N. In vitro RBE-LET dependence for multiple particle types. Acta Oncol 2011, 50(6), 757–762. [Google Scholar] [CrossRef]
- Dean, R. T.; Fu, S.; Stocker, R.; Davies, M. J. Biochemistry and pathology of radical-mediated protein oxidation. Biochem J 1997, 324, 1–18. [Google Scholar] [CrossRef]
- Davies, M. J. Protein oxidation and peroxidation. Biochem J 2016, 473(7), 805–825. [Google Scholar] [CrossRef]
- Morgan, P. E.; Pattison, D. I.; Davies, M. J. Quantification of hydroxyl radical-derived oxidation products in peptides containing glycine, alanine, valine, and proline. Free Radic Biol Med 2012, 52(2), 328–339. [Google Scholar] [CrossRef]
- Sharp, J. S.; Tomer, K. B. Effects of anion proximity in peptide primary sequence on the rate and mechanism of leucine oxidation. Anal Chem 2006, 78(14), 4885–4893. [Google Scholar] [CrossRef]
- Galano, A.; Cruz-Torres, A. OH radical reactions with phenylalanine in free and peptide forms. Org Biomol Chem 2008, 6(4), 732–738. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Inman, J. L.; Chant, J.; Obst-Huebl, L.; Nakamura, K.; Costello, S. M.; Marqusee, S.; Mao, J. H.; Kunz, L.; Paisley, R.; et al. A Novel Platform for Evaluating Dose Rate Effects on Oxidative Damage to Peptides: Toward a High-Throughput Method to Characterize the Mechanisms Underlying the FLASH Effect. Radiat Res 2023, 200(6), 523–530. [Google Scholar] [CrossRef]
- Kristensen, L. G.; Holton, J. M.; Rad, B.; Chen, Y.; Petzold, C. J.; Gupta, S.; Ralston, C. Y. Hydroxyl radical mediated damage of proteins in low oxygen solution investigated using X-ray footprinting mass spectrometry. J Synchrotron Radiat 2021, 28, 1333–1342. [Google Scholar] [CrossRef] [PubMed]
- Boyd, A.; Carver, M.; Dixon, R. Computed and Experimental Product Concentrations in the Radiolysis of Water. Radiat. Phys. Chem. 1980, 15, 8. [Google Scholar] [CrossRef]
- Garrison, W. M. Reaction Mechanisms in the Radiolysis of Peptides, Polypeptides, and Proteins. Chem Rev 1986, 87, 18. [Google Scholar]
- Megroureche, J.; Bekerat, H.; Bian, J.; Bui, A.; Sankey, J.; Childress, L.; Enger, S. A. Development of a hydrated electron dosimeter for radiotherapy applications: A proof of concept. Med Phys 2023, 50(11), 7245–7251. [Google Scholar] [CrossRef]
- Murphy, M. P.; Bayir, H.; Belousov, V.; Chang, C. J.; Davies, K. J. A.; Davies, M. J.; Dick, T. P.; Finkel, T.; Forman, H. J.; Janssen-Heininger, Y.; et al. Guidelines for measuring reactive oxygen species and oxidative damage in cells and in vivo. Nat Metab 2022, 4(6), 651–662. [Google Scholar] [CrossRef]
- Zhang, T.; Stengl, C.; Derksen, L.; Palskis, K.; Koritsidis, K.; Zink, K.; Adeberg, S.; Major, G.; Weishaar, D.; Theiss, U.; et al. Analysis of hydrogen peroxide production in pure water: Ultrahigh versus conventional dose-rate irradiation and mechanistic insights. Med Phys 2024, 51(10), 7439–7452. [Google Scholar] [CrossRef]
- Thomas, W.; Sunnerberg, J.; Reed, M.; Gladstone, D. J.; Zhang, R.; Harms, J.; Swartz, H. M.; Pogue, B. W. Proton and Electron Ultrahigh-Dose-Rate Isodose Irradiations Produce Differences in Reactive Oxygen Species Yields. Int J Radiat Oncol Biol Phys 2024, 118(1), 262–267. [Google Scholar] [CrossRef]
- Blain, G.; Vandenborre, J.; Villoing, D.; Fiegel, V.; Fois, G. R.; Haddad, F.; Koumeir, C.; Maigne, L.; Metivier, V.; Poirier, F.; et al. Proton Irradiations at Ultra-High Dose Rate vs. Conventional Dose Rate: Strong Impact on Hydrogen Peroxide Yield. Radiat Res 2022, 198(3), 318–324. [Google Scholar] [CrossRef]
- Kacem, H.; Psoroulas, S.; Boivin, G.; Folkerts, M.; Grilj, V.; Lomax, T.; Martinotti, A.; Meer, D.; Ollivier, J.; Petit, B.; et al. Comparing radiolytic production of H(2)O(2) and development of Zebrafish embryos after ultra high dose rate exposure with electron and transmission proton beams. Radiother Oncol 2022, 175, 197–202. [Google Scholar] [CrossRef]
- D-Kondo, J. N.; Garcia-Garcia, O. R.; LaVerne, J. A.; Faddegon, B.; Schuemann, J.; Shin, W. G.; Ramos-Mendez, J. An integrated Monte Carlo track-structure simulation framework for modeling inter and intra-track effects on homogenous chemistry. Phys Med Biol 2023, 68(12). [Google Scholar] [CrossRef] [PubMed]
- Derksen, L.; Flatten, V.; Engenhart-Cabillic, R.; Zink, K.; Baumann, K. S. A method to implement inter-track interactions in Monte Carlo simulations with TOPAS-nBio and their influence on simulated radical yields following water radiolysis. Phys Med Biol 2023, 68(13). [Google Scholar] [CrossRef]
- Abolfath, R.; Grosshans, D.; Mohan, R. Oxygen depletion in FLASH ultra-high-dose-rate radiotherapy: A molecular dynamics simulation. Med Phys 2020, 47(12), 6551–6561. [Google Scholar] [CrossRef]
- Lai, Y.; Jia, X.; Chi, Y. Modeling the effect of oxygen on the chemical stage of water radiolysis using GPU-based microscopic Monte Carlo simulations, with an application in FLASH radiotherapy. Phys Med Biol 2021, 66(2), 025004. [Google Scholar] [CrossRef]
- Lamiable, A.; Thevenet, P.; Rey, J.; Vavrusa, M.; Derreumaux, P.; Tuffery, P. PEP-FOLD3: faster de novo structure prediction for linear peptides in solution and in complex. Nucleic Acids Res 2016, 44(W1), W449–454. [Google Scholar] [CrossRef] [PubMed]
- Vogt, W. Oxidation of methionyl residues in proteins: tools, targets, and reversal. Free Radic Biol Med 1995, 18(1), 93–105. [Google Scholar] [CrossRef] [PubMed]
- Wecksler, A. T.; Wang, L.; Bernstein, L. J.; Huang, R. Y. C.; Gupta, S.; Kristensen, L. G.; Ralston, C. Y.; Sobott, F.; Gross, M. L.; Jones, L. M.; et al. Recommendations and Considerations for Hydroxyl Radical Protein Footprinting - Mass Spectrometry. Nature Methods 2026, in press. [Google Scholar]
- Liu, X. R.; Zhang, M. M.; Zhang, B.; Rempel, D. L.; Gross, M. L. Hydroxyl-Radical Reaction Pathways for the Fast Photochemical Oxidation of Proteins Platform As Revealed by (18)O Isotopic Labeling. Anal Chem 2019, 91(14), 9238–9245. [Google Scholar] [CrossRef]
Figure 1.
Hydrogen peroxide production in aerated (blue) or low-oxygen (orange) Milli-Q water as a function of dose using 0.02 Gy/s dose rate X-rays. Error bars represent the standard deviation between triplicate data points.
Figure 1.
Hydrogen peroxide production in aerated (blue) or low-oxygen (orange) Milli-Q water as a function of dose using 0.02 Gy/s dose rate X-rays. Error bars represent the standard deviation between triplicate data points.
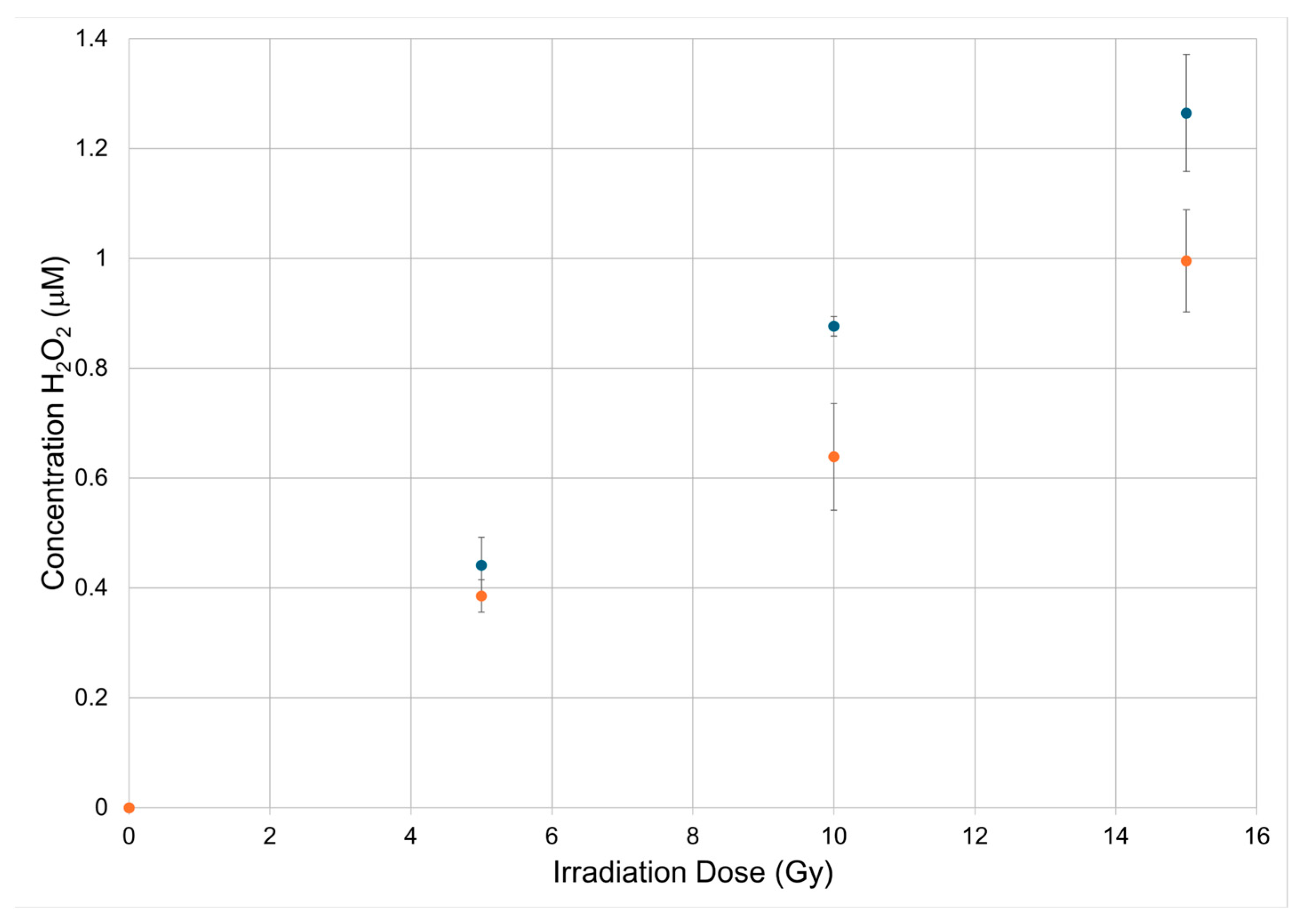
Figure 2.
Structure prediction of each peptide used in the study, as predicted by PEPFOLD3.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



