Submitted:
04 February 2026
Posted:
04 February 2026
You are already at the latest version
Abstract
Adult neurogenesis is a highly regulated form of brain plasticity shaped by the interaction between hormonal systems and environmental context. Social experience has emerged as a powerful modulator of neuronal proliferation, differentiation, and survival across the lifespan. This review synthesizes evidence showing how diverse social behaviors—including isolation, social hierarchy, parenting, sexual interaction, social buffering, and social learning—engage neuroendocrine, neurochemical, and stress-related pathways to regulate hippocampal and olfactory neurogenesis. Affiliative and reproductive behaviors generally promote neurogenesis through gonadal hormones, oxytocin, vasopressin, and neurotrophic signaling, whereas chronic isolation or social defeat reliably suppress neurogenic processes, particularly within stress-sensitive regions of the ventral dentate gyrus. Sex differences further shape these effects, reflecting distinct hormonal environments and behavioral strategies. Comparative studies in prairie voles, eusocial mole-rats, nonhuman primates, songbirds, and teleost fish reveal that sociality can either enhance or constrain adult neurogenesis depending on ecological demands and social organization. Together, these findings position adult neurogenesis as a plastic process deeply embedded within the social lives of vertebrates, offering a framework for linking social behavior, brain plasticity, and adaptive function.
Keywords:
adult neurogenesis
; social behavior
; neuroendocrine modulation
1. Introduction
Neurogenesis is a continuous process in the brain that remains highly sensitive to a wide range of internal and external factors, making it critical for mammalian maturation, development, adaptation, and evolution [1,2]. The neurogenic process comprises multiple stages that are modulated by shared neurochemical and hormonal systems [3]. Endogenous factors include hormonal and neurochemical signals that promote the proliferation, growth, survival, and functional integration of new neurons, whereas exogenous factors encompass environmental elements—both stressful and appetitive—that can either trigger or inhibit neurogenic mechanisms [4,5].
A wide body of research has examined key external modulators of neurogenesis. Physical activity, particularly cardiovascular exercise, promotes the generation of new neurons through neurotrophic pathways [6,7,8] and enhances memory and learning functions [9]. Environmental enrichment is likewise associated with increased neuronal density in the dentate gyrus of the hippocampus [4,10]. More recently, social experience has emerged as a potent regulator of neurogenesis, influencing each stage of the process [11,12]. A deeper understanding of these pathways offers opportunities to develop novel stimulation- and treatment-based strategies for multiple human conditions.
Comprehensive frameworks of adult neurogenesis have been previously described [13]. Building on this foundation, the present review focuses specifically on how social interaction shapes neurogenic processes across different stages of life.
2. Socialization
Social interaction is associated with numerous neuroprotective, cognitive, and mood-related benefits, particularly by stimulating and enhancing cognitive functions such as learning and memory [14]. It also plays a key role in reducing the impact of stress and decreasing anxiety- and depression-like behaviors [15]. This review synthesizes evidence on the relationship between social behavior and neurogenesis, identifying different forms of social behavior and their underlying mechanisms—ranging from neurotrophic factors to the hormonal pathways governed by the hypothalamic–pituitary–gonadal and hypothalamic–pituitary–adrenal axes.
Several elements contribute to social behavior, along with their associated mechanisms and manifestations, including social hierarchy, sexual interaction, parenting, and living in groups or in isolation. All of these domains have been examined in relation to the generation, survival, and functional integration of neurons. However, interactions with other variables—such as age, sex, and species—produce heterogeneous findings, underscoring the need for studies that systematically incorporate these moderating factors.
Social behavior is broadly understood to comprise both reinforcing and aversive components, although most behaviors involve a mixture of the two. For instance, receiving aggression exemplifies an aversive social experience, whereas mating typically represents a highly reinforcing one. Social status, in turn, reflects a combination of both motivational components.
3. Isolation
In most wildlife species, living in a social group provides several advantages, including improved access to territory, greater protection, enhanced performance, and more reliable food sources—factors that collectively contribute to well-being and survival [16]. To avoid misinterpretations of the effects of social behavior, it is essential to identify both the social role of individuals and the contextual characteristics of their environment, as isolation can be stressful in one situation yet relatively neutral—or even rewarding—in another [17,18].
Social isolation consistently produces measurable effects on brain function and plasticity, most notably by disrupting synaptic transmission and impairing neurogenesis [19,20,21]. However, these outcomes depend strongly on developmental stage. Because neurogenesis varies across the lifespan—adolescents generate and integrate new neurons at substantially higher rates than adults [22]—isolation during early life versus adulthood leads to different consequences.
Importantly, the neurogenic and behavioral impact of social isolation is strongly modulated by developmental stage and duration of exposure. Adolescence represents a particularly sensitive window, during which chronic isolation induces persistent behavioral alterations and long-lasting vulnerability. In contrast, short-term isolation in adulthood produces relatively limited behavioral effects, whereas prolonged isolation leads to measurable impairments in hippocampal plasticity and cognition. These age-dependent patterns underscore that isolation does not exert uniform effects across the lifespan, but instead interacts with baseline neurogenic capacity and circuit maturation (Figure 1).
Conceptual overview of age- and duration-dependent effects of social isolation on behavior and hippocampal plasticity.
DG, dentate gyrus; CA3, cornu ammonis 3; mPFC, medial prefrontal cortex; BDNF, brain-derived neurotrophic factor.
Ibi et al. [19] conducted an experiment in which 3-week-old male mice were isolated for 4 weeks. When BrdU (bromodeoxyuridine) was administered at the end of the fourth week, no significant differences in cell proliferation were found between isolated and control groups. However, when BrdU was administered at the beginning of the fourth week, isolated mice showed significantly lower concentrations of BrdU-labeled cells in the dentate gyrus, indicating that isolation impaired cell differentiation and survival but not proliferation. Isolation produces different effects depending on natural fluctuations in gonadal hormones, sex, and specific features of the isolation paradigm, including onset and duration [23,24]. McCormick et al. [25] reported decreased cell proliferation and survival in the dentate gyrus during breeding [26] and adolescence [27]. These reductions, however, could be reversed by re-exposure to social housing during these developmental windows, consistent with heightened neuroplasticity during early life. In contrast, male mice isolated after weaning exhibited no changes in dentate gyrus cell proliferation [19], highlighting that the neurogenic impact of early isolation depends on precise developmental timing.
Additional evidence shows that isolation decreases doublecortin expression—a marker of neuronal immaturity and ongoing neurogenesis [28,29]—indicating reduced numbers of newly generated neurons.
In adult female prairie voles, six weeks of isolation similarly reduced cell proliferation and survival; notably, this reduction extended beyond the dentate gyrus to include the amygdala, medial preoptic area, and ventromedial hypothalamus [30].
In contrast, several studies indicate that social or group housing can buffer the impact of chronic stress on hippocampal neurogenesis. Social housing reduces or prevents stress-induced decreases in BrdU-labeled cells in the dentate gyrus [31], and social enrichment following a period of isolation restores hippocampal BDNF and NGF expression as well as neurogenesis in the dentate gyrus [32]. Additionally, chronic social isolation reduces hippocampal BDNF levels relative to pair-housed rats [33], supporting the notion that group living may exert a neuroprotective effect through neurotrophic mechanisms.
Galea et al. [24] exposed male and female adult rats to an electric-shock stress paradigm under two housing conditions: isolated or group-housed. A reduction in BrdU-labeled cells was observed in isolated animals, whereas group-housed rats showed increased cell proliferation; importantly, sex differences emerged, indicating modulation by gonadal hormones. These changes were not evident in the subventricular zone, even though exposure to a social olfactory stimulus can mitigate stress effects in the dentate gyrus [17]. Social interaction also increases the expression of neurotrophic factors such as brain-derived neurotrophic factor (BDNF), its TrkB receptor [34], and nerve growth factor (NGF) [35], all of which play essential roles in neurogenesis.
Importantly, hippocampal responses to isolation are anatomically dissociable. The dorsal dentate gyrus (dDG) is primarily involved in spatial and contextual memory, whereas the ventral dentate gyrus (vDG) regulates stress, social behavior, and emotional processing. Adult neurogenesis in the vDG has been shown to confer resilience to chronic stress by inhibiting stress-responsive granule cells, whereas suppression of neurogenesis in this region increases stress susceptibility [36]. Emerging data further indicate that social isolation produces selective reductions in cell proliferation in the vDG (but not the dDG), at least in female rodents, following adolescent isolation; these changes persist into adulthood and are not reversed by a resocialization period [37]. These findings suggest that many of the behavioral and affective consequences of isolation likely involve vDG-mediated circuits rather than dorsal hippocampal pathways.
These findings indicate that the effects of social isolation on hippocampal neurogenesis are not uniformly distributed along the dentate gyrus. Instead, isolation preferentially targets the ventral dentate gyrus—a subregion critically involved in stress regulation, emotional processing, and social behavior—while sparing the dorsal dentate gyrus, which primarily supports spatial and contextual memory. This functional–anatomical dissociation provides a mechanistic link between reduced neurogenesis and the affective and behavioral consequences of isolation (Figure 2).
Conceptual schematic highlighting differential involvement of dorsal and ventral dentate gyrus in cognitive versus affective responses to social isolation.
dDG, dorsal dentate gyrus; vDG, ventral dentate gyrus.
Moreover, longer or repeated periods of isolation appear more likely to yield persistent structural or neurogenic alterations, consistent with reports showing that even intermediate isolation protocols (e.g., ~14 days) can produce measurable reductions in hippocampal neurogenesis and associated behavioral changes in adult rodents [38].
Table 1 provides an overview of empirical findings describing the behavioral and neurobiological consequences of social isolation across early life, adolescence, and adulthood in both animal models and human populations. Early-life isolation is associated with long-term anxiety-like phenotypes and broad neuroendocrine and neurochemical disruptions [39]. Adolescence represents a particularly sensitive window, with chronic isolation yielding persistent alterations in reward-related behavior, anxiety-like responses, ethanol consumption, and fear extinction [40,41]. In early adulthood, longitudinal analyses indicate that loneliness or social isolation significantly predict depressive episodes and deteriorated mental health [42]. Isolation initiated in adulthood induces limited but measurable behavioral effects alongside transcriptomic changes in monoaminergic pathways [43], whereas long-term isolation leads to neuroendocrine and behavioral alterations in rodent models [44]. Human data from short-term quarantine further support these associations, showing elevated anxiety and depressive symptoms influenced by behavioral and sociodemographic factors [45].
4. Social Hierarchy
It is well documented that social groups exhibit a distribution of roles that emerges from competition for limited resources. Dominance and submission have been studied using several paradigms, including the visible burrow system [46]. Dominance not only confers priority access to food, mates, and territory but also produces measurable effects at the brain level. Kozorovitskiy and Gould [47] reported increased production of new neurons in the dentate gyrus of dominant male rats compared to subordinates; however, there is no evidence of neurogenesis suppression in subordinate individuals [48,49,50]. This suggests that hierarchy-related variation in neurogenesis depends on circulating hormone concentrations. For example, dominant rats exhibit high levels of testosterone and luteinizing hormone [51], whereas subordinate rats show elevated corticosteroid levels [46,51].
Behaviorally, subordinate rats display reduced aggression and decreased locomotor, sexual, and social activity [52], which may limit their engagement in behaviors that themselves promote neurogenesis, such as sexual interaction. Notably, increased adult neurogenesis has also been observed in dominant baboons compared with subordinates, even when both have access to sexual interaction [53], indicating a specific effect of hierarchical status independent of mating opportunities.
Finally, disrupting an established social status produces divergent effects. Subordinate naked mole-rats, when removed from their colony and isolated, show increased proliferation in the ventral dentate gyrus—likely reflecting the release of social and reproductive suppression [54]. In contrast, disruption of dominant status in other social species is associated with stress-related reductions in hippocampal plasticity, including decreased neurogenesis and altered expression of neuromodulatory systems involved in social behavior. Although specific effects vary across taxa, loss of dominance generally engages glucocorticoid-dependent mechanisms that suppress neuronal survival and remodel limbic circuits supporting social and emotional regulation.
Repeated social defeat in rodents produces robust and persistent social avoidance and, in susceptible mice, reduced sucrose preference indicative of anhedonia. Chronic defeat also induces anxiety-like behaviors. Molecular profiling reveals marked transcriptional adaptations in mesolimbic reward regions (NAc, VTA) and the medial prefrontal cortex (mPFC) that sharply differentiate susceptible from resilient animals. These circuit-level and molecular changes have been consistently observed across studies using the standardized social defeat paradigm [55,56]. Pharmacological studies further show that glutamatergic NMDA signaling in the basolateral amygdala is required for both the acquisition and expression of conditioned defeat, whereas GABAA-mediated inactivation of the ventromedial prefrontal cortex abolishes dominance-related resistance to defeat [57,58].
In contrast, adolescent social instability stress—brief daily isolation paired with frequent changes of cage partners—produces more subtle yet enduring alterations in social behavior, accompanied by subregion-specific dendritic and synaptic remodeling within the medial amygdala and lateral septum [59]. This paradigm also induces pronounced sex-dependent changes in the neuroendocrine–immune–gut axis, with females showing the strongest alterations in inflammatory signaling, microbial composition, and stress responsivity [60]. Finally, single-unit recordings indicate that anxiety-related firing patterns in the mPFC are tightly coupled to ventral hippocampal input during exploration of anxiogenic environments, highlighting a broader vHPC–mPFC–amygdala circuit through which both defeat and social instability stress may bias emotional processing and social behavior [61].
In parallel to behavioral and neurogenic differences, social hierarchy is also shaped by hormonal and molecular mechanisms that modulate plasticity within stress- and reward-related circuits. Dominant males exhibit elevated testosterone and luteinizing hormone, whereas subordinate individuals show sustained increases in corticosterone [46,51]—a profile consistent with divergent engagement of androgen-responsive pathways that promote neuronal survival in the dentate gyrus [62,63] versus glucocorticoid-dependent suppression of plasticity under chronic subordination [64]. At the molecular level, hierarchy-related differences in neurotrophic and transcriptional signaling have been described across social stress paradigms. Chronic social defeat produces robust remodeling of mesolimbic circuits, including differential activation of BDNF- and CREB-related pathways in susceptible versus resilient animals [34,55,56]. These adaptations intersect with glucocorticoid-receptor–dependent mechanisms known to regulate stress responsivity and long-term behavioral outcomes [64]. Together, these endocrine and molecular mechanisms provide a coherent framework linking hierarchical status to stress vulnerability, circuit plasticity, and individual differences in resilience.
5. Social Buffering
Social buffering refers to the reduction of physiological and behavioral stress responses that occurs when an individual encounters a threat in the presence of an affiliative conspecific or caregiver. Across development and species, supportive social partners reliably dampen HPA-axis activity and reduce cortisol/corticosterone output [65,66]. In rodents, the presence of an adult conspecific during threat exposure markedly suppresses freezing and attenuates corticosterone responses, demonstrating that social cues buffer both behavioral and endocrine components of the stress reaction [67,68]. Human studies similarly show that attachment figures reduce cortisol reactivity to social-evaluative stress, indicating an evolutionarily conserved mechanism for regulating HPA-axis responsivity [65].
At the mechanistic level, social buffering operates by dampening stress-hormone signaling and preserving plasticity within stress-sensitive hippocampal circuits. In the absence of affiliative social contact, chronic stress leads to sustained activation of the HPA axis, elevated glucocorticoids, and suppression of cell proliferation and survival in the dentate gyrus. By contrast, the presence of an affiliative conspecific attenuates corticosterone responses, maintains neurotrophic support, and protects hippocampal neurogenesis from stress-induced disruption (Figure 3).
Conceptual overview illustrating how affiliative social interactions attenuate stress-hormone signaling and preserve hippocampal neurogenesis.
DG, dentate gyrus; HPA, hypothalamic–pituitary–adrenal axis.
At the neural and plasticity level, social buffering mitigates the deleterious effects of chronic stress on mesolimbic and hippocampal circuits. Rewarding social interaction reverses stress-induced behavioral and molecular alterations in limbic regions [15], whereas socially impoverished or unstable environments reduce hippocampal neurogenesis and dysregulate stress-hormone profiles—often in sex-specific ways [25,26,31]. These findings converge with evidence that increased neurogenesis in the ventral dentate gyrus promotes resilience by inhibiting stress-responsive granule cells and limiting pathological engagement of stress circuits [36]. Together, these results position social buffering as a multilevel protective mechanism—hormonal, neurocircuit-level, and neurogenic—that stabilizes HPA-axis output, maintains ventral hippocampal plasticity, and reduces vulnerability to stress-related psychopathology.
6. Social Learning & Social Memory
Social learning depends on the ability to acquire information from conspecifics, a process grounded in social recognition memory—the capacity to discriminate familiar from unfamiliar individuals and to encode socially relevant cues. This form of memory is strongly regulated by the neuropeptides oxytocin (OXT) and vasopressin (AVP), which modulate olfactory–limbic circuits supporting identity recognition [69,70]. Extensive reviews show that disrupting OXT/AVP signaling—whether pharmacologically, genetically, or circuit-specifically—reliably impairs social recognition across rodent models, underscoring the central role of these neuropeptide systems in encoding and retrieving social identities [69,70,71].
Within the hippocampal formation, the CA2 subregion and ventral CA1 constitute key nodes of a circuit specialized for social memory. Converging evidence shows that disrupting CA2 function—or its direct input from the lateral entorhinal cortex—selectively impairs the ability of rodents to discriminate familiar from novel conspecifics, while leaving non-social memory domains intact [72,73]. In particular, optogenetic silencing of LEC→CA2 projections abolishes social recognition memory without affecting object recognition, indicating that CA2-dependent computations are specifically tuned to socially relevant cues [73]. As reviewed by Wang & Zhan [72], social memory representations in CA2 and vCA1 interface with broader limbic and motivational circuits—including septal, prefrontal, and nucleus accumbens pathways—that encode social salience, identity, and motivational value, situating CA2 as a central hub within the social information-processing network.
Adult hippocampal neurogenesis provides an additional layer of plasticity for social information processing. Experimental suppression of adult-born granule cells impairs social recognition memory in rodents, indicating that new neurons contribute to the stability, maintenance, and updating of social representations [74]. Conversely, environmental enrichment—which enhances adult neurogenesis—restores social memory deficits induced by previous isolation, and this improvement is abolished when cell proliferation is pharmacologically blocked, supporting a causal role for neurogenesis in social memory persistence [75]. Complementary systematic reviews highlight that social behaviors both influence and depend on neurogenesis within hippocampal and olfactory circuits, integrating adult-born neurons into a broader network supporting social learning [11]. These mechanisms also shape how rodents adapt to socially structured environments, including the formation of stable dominance hierarchies. Dominant males exhibit greater survival of adult-born neurons in the dentate gyrus compared with subordinate males, indicating that hierarchical status modulates hippocampal plasticity. Although this study did not directly examine social memory or identity encoding, adult-born granule cells are broadly implicated in contextual and affective processing, suggesting that neurogenesis may influence how animals navigate complex social situations. However, the specific encoding of individual identity, rank, or social network structure requires evidence from other research domains and is beyond the scope of Kozorovitskiy & Gould [47].
7. Sex Differences and Sexual Dimorphism
Sex differences and sexual dimorphism shape how mammals integrate social experience, stress, and reproductive demands into neural plasticity and behavior. In this context, sex differences refer to quantitative variations in traits shared by both sexes—such as the magnitude of hormonal responses, levels of neurogenesis, or behavioral intensity—whereas sexual dimorphism denotes qualitative distinctions in phenotype, physiology, or behavior that arise from sex-specific developmental pathways, leading males and females to express partially non-overlapping strategies or neural specializations.
Males and females exhibit distinct neuroendocrine and behavioral responses to social stress. Males often show stress-induced reductions in hippocampal neurogenesis and alterations in defensive behavior, whereas females display sex-specific changes in anxiety, social motivation, and stress responsivity [31,49,76,77]. These dimorphisms extend to parental care, mating, and aggression, where the hormonal milieu modulates hippocampal plasticity and social circuits. In males, androgens enhance adult-born neuron survival and influence sexually motivated behavior and aggression [62,78,79]. In females, neurogenesis is dynamically regulated across pregnancy, lactation, and the postpartum period by estrogens, progesterone, adrenal steroids, and prolactin [80,81,82].
Emerging evidence also indicates that males and females rely on partially distinct neural pathways for processing social information, social memory, and social buffering, reflecting dimorphic organization of corticolimbic and hypothalamic systems [12,23,83].
Collectively, these findings support the growing field of sex-informed neuroscience, which conceptualizes sex as a biological variable that systematically modulates neurogenesis, hormone–brain interactions, stress vulnerability, parental and reproductive strategies, and the adaptive use of social environments across the lifespan.
8. Parenting
Like other social behaviors, parenting is strongly modulated by hormonal changes [77], particularly during pregnancy and throughout contact with the infant, including breastfeeding [12]. However, its effects on neurogenesis vary depending on the specific stage of caregiving. During lactation [84] and the postpartum period [85], elevated corticosterone levels have been reported, suppressing the generation of new neurons and suggesting that the stressful component of parenting can negatively impact hippocampal plasticity.
Conversely, other findings point to positive neuroplastic effects associated with maternity. Leuner and Gould [81] reported increased dendritic spine density in the hippocampus and medial prefrontal cortex, and a buffering effect of motherhood on stress-induced learning deficits has also been described [86]. In addition, parenting has been shown to stimulate both the proliferation and survival of new neurons [82].
Taken together, these results suggest that parenting exerts bidirectional effects on neurogenesis: positive, reward-related influences driven by the hedonic and affiliative components of caregiving, and negative influences associated with the physiological stress inherent to parental demands.
Beyond the general effects of parenting on hippocampal plasticity, important distinctions emerge between maternal and paternal care. In mothers, pregnancy, lactation, and infant contact produce marked fluctuations in estrogens, progesterone, oxytocin, and prolactin—a hormone that directly stimulates neural progenitor proliferation in the subventricular zone and facilitates maternal behavioral responsiveness [80,87].
In fathers, parenting induces different forms of neural plasticity: paternal experience suppresses adult hippocampal neurogenesis in Peromyscus californicus, whereas fatherhood increases dendritic spine density in dentate gyrus granule cells and CA1 pyramidal neurons and reduces anxiety-like behavior—changes that reflect circuit-level adaptations supporting paternal care [88,89,90].
Human EEG and neuroimaging studies similarly show that the postpartum period involves experience-dependent changes in prefrontal–parietal synchronization and reward-related activity when mothers process infant cues, revealing cortical plasticity that enhances sensitivity to infant signals [91,92].
Taken together, parenting integrates both a stress component—reflected in corticosterone-associated suppression of neurogenesis—and a rewarding component mediated by prolactin-, oxytocin-, and dopamine-related mechanisms that promote attachment and motivation [12,86,93]. The interplay between these stress-related and hedonic pathways determines the direction and magnitude of plastic changes across hippocampal and prefrontal systems during the parental period.
To synthesize these findings and highlight the temporal dynamics of endocrine and neural plasticity during caregiving, Table 2 organizes key hormonal profiles and associated neurobiological effects across distinct stages of parental behavior. This overview emphasizes how pregnancy, postpartum transitions, lactation, and paternal involvement each engage unique constellations of hormones and plasticity mechanisms—from prolactin-driven proliferation in the SVZ to experience-dependent modulation of prefrontal and hippocampal circuits—while illustrating the shared balance between stress-related and reward-related pathways that shape parental adaptations.
9. Sexual Interaction
Sexual interaction is a potent social reinforcer that engages motivational and reward circuits while producing hormone-dependent effects on adult hippocampal plasticity. In rodents and other mammals, copulation and sexually relevant cues recruit mesolimbic dopamine pathways—particularly within the nucleus accumbens—in ways that parallel other natural rewards. Dopaminergic signaling contributes to the incentive value of sexual stimuli and to the formation of conditioned partner and place preferences, even though dopamine is not required for the motor execution of copulation itself [94,95]. Through these mechanisms, sexual interaction influences not only reproductive outcomes but also the motivational and affective components of social behavior.
In females, estradiol exerts rapid and robust proliferative effects on neural progenitors in the dentate gyrus. Estradiol surges—whether naturally occurring during proestrus or experimentally induced—transiently increase the number of dividing hippocampal cells [96,97]. These effects fluctuate across the estrous cycle, with proestrus/high-estradiol phases enhancing both sexual receptivity and hippocampal cell proliferation. Because estradiol also facilitates appetitive aspects of sexual behavior and modulates dopaminergic responses to sexual stimuli, estrous-cycle variation likely coordinates reward-circuit recruitment with periods of heightened neurogenic plasticity [94,98].
In males, testosterone acts primarily on the survival and maturation of adult-born granule neurons. Experimental studies demonstrate that testosterone and dihydrotestosterone increase the survival of newly generated neurons through androgen receptor–dependent mechanisms [62,79]. Sexual activity elevates testosterone and engages these pathways, potentially linking mating to long-term changes in hippocampal circuit integration [78]. Thus, estradiol predominantly influences proliferative phases in females, whereas testosterone modulates survival and functional incorporation of new neurons in males.
Sexual experience additionally affects stress reactivity and emotional resilience. Although mating can acutely elevate glucocorticoids, repeated sexual interaction increases hippocampal neurogenesis and improves anxiety- and stress-related behaviors [81]. These effects likely arise from interactions among mesolimbic dopamine, gonadal steroids, and hippocampal plasticity mechanisms. Sexual reward recruits neuromodulatory systems that can buffer stress-related suppression of neurogenesis and promote adaptive emotional regulation [12].
Collectively, sexual interaction influences adult neurogenesis through four coordinated mechanisms:
- recruitment of mesolimbic dopamine circuits as a socially rewarding stimulus;
- estradiol-dependent increases in progenitor proliferation across the estrous cycle;
- testosterone-dependent enhancement of new-neuron survival; and
- experience-driven modulation of reward and stress pathways that jointly support adaptive hippocampal plasticity.
10. Neurochemical Pathways of Socialization
As noted above, diverse forms of social behavior influence adult neurogenesis through multiple physiological, hormonal, and neurochemical pathways. These effects are particularly evident in the two principal neurogenic niches of the adult brain—the subventricular zone (SVZ), which supplies newborn neurons to the olfactory bulb, and the dentate gyrus of the hippocampus [99,100]. Socially induced changes in neurogenesis are broadly regulated by neuroendocrine systems and by the synthesis, release, and receptor-mediated actions of several neuromodulators and trophic factors.
Social interaction can be either reinforcing or aversive, and correspondingly may promote or suppress neurogenesis. These bidirectional effects are largely orchestrated by hormones of the hypothalamic–pituitary–adrenal (HPA) and hypothalamic–pituitary–gonadal (HPG) axes [24]. Reinforcing social experiences—such as mating, bonding, or parental contact—tend to engage gonadal-steroid and neuropeptide systems that facilitate neurogenic processes, whereas aversive experiences, including social defeat or chronic isolation, activate glucocorticoid pathways that inhibit neurogenesis.
Gonadal hormones exert particularly strong and dynamic influences on neural progenitor proliferation and survival. Estradiol is closely associated with increased production of new neurons [12]. Acute estradiol administration stimulates hippocampal cell proliferation within 2–4 hours, although this effect disappears by 48 hours [101]. However, high-dose estradiol [96,102] or chronic administration at any dose does not enhance proliferation [98]. These suppressive outcomes observed at elevated estradiol levels appear to be mediated by glucocorticoids [103]. Moreover, estradiol’s effects on neurogenesis and dendritic spine density vary as a function of sex [104], age, and reproductive status [105]. For instance, females in proestrus display higher rates of hippocampal cell proliferation than males [97], although this pattern is not preserved across all species. The hippocampus contains abundant receptors for gonadal steroids, oxytocin, luteinizing hormone (LH), and prolactin—hormones strongly implicated in sexual behavior and parental care [24]. Prolactin receptors are present in both the SVZ and the hippocampus [106]. Exposure to pheromones from a dominant male increases cell proliferation in the olfactory bulb and hippocampus of female mice [87], while pregnancy induces pronounced proliferation in the SVZ [80]. These effects appear to be mediated by prolactin in the olfactory bulb and by LH in the hippocampus [107]. Additionally, prolactin may confer neuroprotective effects during chronic stress. Notably, however, early postnatal administration of prolactin (postnatal day 14) reduces neurogenesis, indicating that the hormone’s effects are strongly age-dependent.
Adult neurogenesis is tightly regulated by endocrine signals that differentially influence distinct stages of the neurogenic process. Gonadal hormones, prolactin, and glucocorticoids exert stage-specific and context-dependent effects on progenitor proliferation and neuronal survival, with outcomes shaped by dose, duration of exposure, developmental stage, and behavioral state. A comparative overview of these hormone-specific actions is provided in Figure 4.
Schematic overview of hormone-specific effects on neurogenic stages under different physiological contexts.
Beyond hormonal and classical neurochemical pathways, adverse social experiences also recruit immune–glial mechanisms that modulate adult neurogenesis. Long-term social isolation in middle-aged mice decreases ΔFosB expression in the dentate gyrus and reduces BDNF levels in CA3, without increasing hippocampal oxidative damage. These findings indicate a state of reduced neuronal activation and diminished trophic support that likely contributes to impaired plasticity [44]. Shorter isolation periods in adult rats induce transcriptional reorganization of monoaminergic and peptidergic signaling in the medial prefrontal cortex—downregulating RGS9, HTR2C, Pdyn, among others—alongside measurable alterations in social behavior, demonstrating that isolation stress rapidly reshapes prefrontal neuromodulatory dynamics [43]. Complementary evidence from chronic social defeat shows that repeated social stress suppresses cell proliferation in the dentate gyrus and elicits robust microglial activation in hippocampal and prefrontal regions, including hypertrophic, cytokine-expressing phenotypes [108]. Taken together, these findings support the conclusion that microglial and inflammatory processes—acting in concert with monoaminergic and neurotrophic changes—form a critical interface through which adverse social environments remodel synaptic function and constrain adult neurogenic plasticity.
Estrogens influence every stage of neurogenesis—including proliferation, differentiation, migration, and maturation. Administration of selective agonists for the alpha (ERα) and beta (ERβ) estrogen receptors regulates cell proliferation in adult female rats [109]. The survival of newly generated neurons depends on the type of estrogen administered, the timing of administration, and the behavioral context in which it occurs. For instance, administration of 17β-estradiol (the most potent endogenous estrogen) increases the survival of newborn neurons in rats performing a maze task, whereas administration of estrone decreases neuronal survival [110].
Androgens constitute another major class of gonadal hormones influencing adult neurogenesis, and their levels increase following sexual interaction [78]. The dentate gyrus as well as the CA1 and CA3 subfields of the hippocampus contain androgen receptors [62], which mediate their specific neurogenic effects. Testosterone and its metabolite dihydrotestosterone enhance the survival of newly generated neurons but do not increase proliferation [12]. Conversely, castrated rats exhibit reduced neuronal survival [79], while adolescent macaques that underwent gonadectomy show increased hippocampal neuronal survival [111]. These findings indicate that, similar to estrogens, androgen effects vary across species, age, sex, and treatment duration. For example, testosterone treatment lasting fewer than 30 days has been associated with an inhibitory effect on neuronal survival [112].
Hormones of the hypothalamic–pituitary–adrenal (HPA) axis also regulate stress effects on neurogenesis and interact bidirectionally with hormones of the hypothalamic–pituitary–gonadal (HPG) axis. Spritzer et al. [63] reported that castrated adult male rats show fewer dentate gyrus cells after one month of isolation compared with intact males. Increased glucocorticoid levels are typically associated with reduced cell proliferation and heightened anxiety [64,113], as well as elevated corticosterone and estradiol levels during aging [103]. However, elevated glucocorticoids do not invariably produce negative effects on neurogenesis; for example, the interaction between sexual activity and stress can promote neurogenesis despite increased glucocorticoid release—a phenomenon likely driven by the hedonic component of sexual interaction [12]. As with other modulators of neurogenesis, these effects are not unidirectional: outcomes depend on complex interactions among hormonal, environmental, and individual variables.
Together, these findings indicate that social experience does not act on adult neurogenesis through a single pathway, but rather through coordinated endocrine, neurochemical, and environmental mechanisms that converge on distinct stages of the neurogenic process. Socially reinforcing behaviors, such as mating, affiliation, and parental interaction, preferentially engage gonadal hormones and neurotrophic signaling to promote neuronal survival and integration, whereas aversive conditions—including chronic isolation and social stress—activate glucocorticoid-dependent pathways that suppress proliferation and impair maturation. Importantly, these effects are stage-specific and context-dependent, highlighting adult neurogenesis as a dynamic interface between social experience and brain plasticity (Figure 5).
Adult neurogenesis proceeds through sequential stages of proliferation, differentiation, survival, and functional integration, which are dynamically regulated by internal endocrine systems and external environmental factors. Social experiences can either promote or suppress neurogenic plasticity by engaging gonadal hormones, neurotrophic signaling, or stress-related glucocorticoid pathways, thereby shaping hippocampal and subventricular neurogenesis in a context-dependent manner.
DG, dentate gyrus; SVZ, subventricular zone; BDNF, brain-derived neurotrophic factor; NGF, nerve growth factor.
To integrate the diverse findings reviewed, it is useful to summarize how distinct hormonal, neurochemical, and stress-related systems jointly regulate adult neurogenesis in social contexts. Social behaviors—from mating and parenting to affiliation, competition, and isolation—are embedded within coordinated endocrine and neuromodulatory responses that target the dentate gyrus and the subventricular zone. These systems do not operate in isolation; rather, they converge on shared cellular processes such as progenitor proliferation, neuronal survival, dendritic remodeling, and synaptic plasticity, while simultaneously shaping social motivation, recognition, and stress responsivity. Table 3 synthesizes the principal hormonal and neurochemical pathways implicated in these interactions, highlighting their specific effects on DG/SVZ plasticity as well as their functional relevance to social behavior.
11. Comparative Models
Comparative models across mammals, birds, and other vertebrates provide crucial insights into how social structure, mating systems, and communication demands shape adult neurogenesis. Species such as prairie voles, naked mole-rats, nonhuman primates, and songbirds illustrate the diversity of neural adaptations supporting monogamy, eusociality, hierarchical organization, and vocal-based social learning. These models reveal that sociality can either amplify or suppress neurogenic activity depending on ecological pressures and behavioral specialization, offering an evolutionary lens through which to interpret rodent findings.
Comparative species models—from socially monogamous prairie voles and eusocial mole-rats to primates, songbirds, and teleost fish—demonstrate that social systems and communication demands profoundly shape adult neurogenesis. Across these taxa, social bonding, hierarchical complexity, and vocal or shoaling behaviors are associated with either up- or down-regulation of neurogenic niches, as summarized in Table 4.
12. Conclusions and Future Directions
Across the evidence reviewed, social experience emerges as a central regulator of adult neurogenesis, influencing every stage of the neurogenic trajectory—from progenitor proliferation to neuronal survival and functional integration. Social interaction, parenting, sexual behavior, hierarchy formation, and group living consistently modulate neurogenesis through coordinated endocrine and neurochemical pathways involving gonadal and adrenal hormones, oxytocin- and vasopressin-dependent systems, neurotrophic signaling, and monoaminergic modulation. Conversely, adverse conditions such as social isolation or chronic social stress reliably disrupt hippocampal plasticity, particularly within ventral dentate gyrus circuits that govern emotional regulation and stress responsivity.
A core conclusion is that the neurogenic impact of social behavior is highly context-dependent, shaped by developmental stage, sex, hormonal milieu, species-specific ecology, and the duration and valence (reinforcing vs. aversive) of the social experience. Parenting illustrates this duality: prolactin-, oxytocin-, and reward-related mechanisms can enhance progenitor activity, whereas corticosterone elevations during stressful caregiving demands suppress hippocampal neurogenesis. Similarly, hierarchy formation and social status modulate neurogenesis through contrasting hormonal profiles, with dominant individuals often showing enhanced neuronal survival.
Comparative models reinforce this diversity. Socially monogamous voles, eusocial mole-rats, primates with complex hierarchies, songbirds with vocal learning systems, and teleost fish with high regenerative capacity demonstrate that sociality can either amplify or constrain neurogenesis depending on evolutionary pressures and behavioral specialization. These species illustrate that adult neurogenesis is not a monolithic phenomenon, but a plastic process embedded within ecological and social adaptations.
Future work should refine how specific social behaviors and hormonal contexts generate distinct neurogenic outcomes across development, sex, and species. Integrating cellular, endocrine, and circuit-level perspectives will be essential for clarifying how socially regulated neurogenesis contributes to social memory, stress resilience, and behavioral adaptation. As shown throughout this review, understanding the mechanisms by which social experience engages or suppresses adult neurogenesis provides a powerful framework for interpreting both the diversity of social behaviors and the adaptive capacity of the adult brain.
Author Contributions
A.T.-J. conceived the initial idea and preliminary structure; J.M.-L. expanded and refined this framework, adding thematic sections and reorganizing the conceptual flow. Both authors conducted literature analysis and co-wrote all manuscript sections. J.M.-L. led the editorial revision and narrative integration. M.H.B.-J. created all figures, contributed technical and theoretical guidance, and critically reviewed the manuscript. All authors approved the final version.
Funding
The División de Investigación y Posgrado of the Universidad Iberoamericana Ciudad de México funded this research. J.M.-L, A.T.-J. and M.H.B.-J. were supported by Sistema Nacional de Investigadoras e Investigadores grants from the Secretaría de Ciencia, Humanidades, Tecnología e Innovación of México.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| AHN | Adult Hippocampal Neurogenesis |
| AVP | Arginine Vasopressin |
| BDNF | Brain-Derived Neurotrophic Factor |
| BrdU | Bromodeoxyuridine |
| CA1 | Cornu Ammonis 1 |
| CA2 | Cornu Ammonis 2 |
| CA3 | Cornu Ammonis 3 |
| DCX | Doublecortin |
| DG | Dentate Gyrus |
| dDG | Dorsal Dentate Gyrus |
| DHT | Dihydrotestosterone |
| EEG | Electroencephalography |
| ERα | Estrogen Receptor Alpha |
| ERβ | Estrogen Receptor Beta |
| HPA axis | Hypothalamic–Pituitary–Adrenal Axis |
| HPG axis | Hypothalamic–Pituitary–Gonadal Axis |
| HVC | High Vocal Center |
| LH | Luteinizing Hormone |
| mPFC | Medial Prefrontal Cortex |
| NAc | Nucleus Accumbens |
| NGF | Nerve Growth Factor |
| OXT | Oxytocin |
| PFC | Prefrontal Cortex |
| RA | Robust Nucleus of the Arcopallium |
| RNA-seq | RNA Sequencing |
| SVZ | Subventricular Zone |
| TrkB | Tropomyosin Receptor Kinase B |
| vDG | Ventral Dentate Gyrus |
| VTA | Ventral Tegmental Area |
| ΔFosB | Truncated FosB Transcription Factor |
References
- Taverna, E.; Götz, M.; Huttner, W.B. The Cell Biology of Neurogenesis: Toward an Understanding of the Development and Evolution of the Neocortex. Annu. Rev. Cell Dev. Biol. 2014, 30, 465–502. [Google Scholar] [CrossRef] [PubMed]
- Gross, C.G. Neurogenesis in the Adult Brain: Death of a Dogma. Nat Rev Neurosci 2000, 1, 67–73. [Google Scholar] [CrossRef]
- Jorgensen, C.; Wang, Z. Hormonal Regulation of Mammalian Adult Neurogenesis: A Multifaceted Mechanism. Biomolecules 2020, 10, 1151. [Google Scholar] [CrossRef] [PubMed]
- Kempermann, G. Environmental Enrichment, New Neurons and the Neurobiology of Individuality. Nat Rev Neurosci 2019, 20, 235–245. [Google Scholar] [CrossRef]
- Jorgensen, C. Adult Mammalian Neurogenesis and Motivated Behaviors. Integrative Zoology 2018, 13, 655–672. [Google Scholar] [CrossRef]
- Liu, P.Z.; Nusslock, R. Exercise-Mediated Neurogenesis in the Hippocampus via BDNF. Front. Neurosci. 2018, 12, 52. [Google Scholar] [CrossRef]
- Choi, D.-H.; Lee, K.-H.; Lee, J. Effect of Exercise-Induced Neurogenesis on Cognitive Function Deficit in a Rat Model of Vascular Dementia. Molecular Medicine Reports 2016, 13, 2981–2990. [Google Scholar] [CrossRef]
- Ma, C.-L.; Ma, X.-T.; Wang, J.-J.; Liu, H.; Chen, Y.-F.; Yang, Y. Physical Exercise Induces Hippocampal Neurogenesis and Prevents Cognitive Decline. Behavioural Brain Research 2017, 317, 332–339. [Google Scholar] [CrossRef]
- Duzel, E.; Van Praag, H.; Sendtner, M. Can Physical Exercise in Old Age Improve Memory and Hippocampal Function? Brain 2016, 139, 662–673. [Google Scholar] [CrossRef]
- Bekinschtein, P.; Oomen, C.A.; Saksida, L.M.; Bussey, T.J. Effects of Environmental Enrichment and Voluntary Exercise on Neurogenesis, Learning and Memory, and Pattern Separation: BDNF as a Critical Variable? Seminars in Cell & Developmental Biology 2011, 22, 536–542. [Google Scholar] [CrossRef] [PubMed]
- García-Gómez, L.; Castillo-Fernández, I.; Perez-Villalba, A. In the Pursuit of New Social Neurons. Neurogenesis and Social Behavior in Mice: A Systematic Review. Front. Cell Dev. Biol. 2022, 10, 1011657. [Google Scholar] [CrossRef]
- Opendak, M.; Briones, B.A.; Gould, E. Social Behavior, Hormones and Adult Neurogenesis. Frontiers in Neuroendocrinology 2016, 41, 71–86. [Google Scholar] [CrossRef]
- Leal-Galicia, P.; Chávez-Hernández, M.E.; Mata, F.; Mata-Luévanos, J.; Rodríguez-Serrano, L.M.; Tapia-de-Jesús, A.; Buenrostro-Jáuregui, M.H. Adult Neurogenesis: A Story Ranging from Controversial New Neurogenic Areas and Human Adult Neurogenesis to Molecular Regulation. IJMS 2021, 22, 11489. [Google Scholar] [CrossRef]
- Leblanc, H.; Ramirez, S. Linking Social Cognition to Learning and Memory. J. Neurosci. 2020, 40, 8782–8798. [Google Scholar] [CrossRef]
- Lemos, C.; Salti, A.; Amaral, I.M.; Fontebasso, V.; Singewald, N.; Dechant, G.; Hofer, A.; El Rawas, R. Social Interaction Reward in Rats Has Anti-stress Effects. Addiction Biology 2021, 26, e12878. [Google Scholar] [CrossRef] [PubMed]
- Seebacher, F.; Krause, J. Physiological Mechanisms Underlying Animal Social Behaviour. Phil. Trans. R. Soc. B 2017, 372, 20160231. [Google Scholar] [CrossRef] [PubMed]
- Holmes, M.M. Social Regulation of Adult Neurogenesis: A Comparative Approach. Frontiers in Neuroendocrinology 2016, 41, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Bell, M.R.; De Lorme, K.C.; Figueira, R.J.; Kashy, D.A.; Sisk, C.L. Adolescent Gain in Positive Valence of a Socially Relevant Stimulus: Engagement of the Mesocorticolimbic Reward Circuitry. Eur J of Neuroscience 2013, 37, 457–468. [Google Scholar] [CrossRef]
- Ibi, D.; Takuma, K.; Koike, H.; Mizoguchi, H.; Tsuritani, K.; Kuwahara, Y.; Kamei, H.; Nagai, T.; Yoneda, Y.; Nabeshima, T.; et al. Social Isolation Rearing-induced Impairment of the Hippocampal Neurogenesis Is Associated with Deficits in Spatial Memory and Emotion-related Behaviors in Juvenile Mice. Journal of Neurochemistry 2008, 105, 921–932. [Google Scholar] [CrossRef]
- Rivera, D.S.; Lindsay, C.B.; Oliva, C.A.; Codocedo, J.F.; Bozinovic, F.; Inestrosa, N.C. Effects of Long-Lasting Social Isolation and Re-Socialization on Cognitive Performance and Brain Activity: A Longitudinal Study in Octodon Degus. Sci Rep 2020, 10, 18315. [Google Scholar] [CrossRef]
- Arzate-Mejía, R.G.; Lottenbach, Z.; Schindler, V.; Jawaid, A.; Mansuy, I.M. Long-Term Impact of Social Isolation and Molecular Underpinnings. Front. Genet. 2020, 11, 589621. [Google Scholar] [CrossRef]
- He, J.; Crews, F.T. Neurogenesis Decreases during Brain Maturation from Adolescence to Adulthood. Pharmacology Biochemistry and Behavior 2007, 86, 327–333. [Google Scholar] [CrossRef]
- Duarte-Guterman, P.; Yagi, S.; Chow, C.; Galea, L.A.M. Hippocampal Learning, Memory, and Neurogenesis: Effects of Sex and Estrogens across the Lifespan in Adults. Hormones and Behavior 2015, 74, 37–52. [Google Scholar] [CrossRef]
- Galea, L.A.M.; Wainwright, S.R.; Roes, M.M.; Duarte-Guterman, P.; Chow, C.; Hamson, D.K. Sex, Hormones and Neurogenesis in the Hippocampus: Hormonal Modulation of Neurogenesis and Potential Functional Implications. J Neuroendocrinology 2013, 25, 1039–1061. [Google Scholar] [CrossRef]
- McCormick, C.M.; Nixon, F.; Thomas, C.; Lowie, B.; Dyck, J. Hippocampal Cell Proliferation and Spatial Memory Performance after Social Instability Stress in Adolescence in Female Rats. Behavioural Brain Research 2010, 208, 23–29. [Google Scholar] [CrossRef]
- Lu, L.; Bao, G.; Chen, H.; Xia, P.; Fan, X.; Zhang, J.; Pei, G.; Ma, L. Modification of Hippocampal Neurogenesis and Neuroplasticity by Social Environments. Experimental Neurology 2003, 183, 600–609. [Google Scholar] [CrossRef]
- Rizzi, S.; Bianchi, P.; Guidi, S.; Ciani, E.; Bartesaghi, R. Neonatal Isolation Impairs Neurogenesis in Thedentate Gyrus of the Guinea Pig. Hippocampus 2007, 17, 78–91. [Google Scholar] [CrossRef] [PubMed]
- Bloch, J.; Kaeser, M.; Sadeghi, Y.; Rouiller, E.M.; Redmond, D.E.; Brunet, J. Doublecortin-positive Cells in the Adult Primate Cerebral Cortex and Possible Role in Brain Plasticity and Development. J of Comparative Neurology 2011, 519, 775–789. [Google Scholar] [CrossRef] [PubMed]
- Dranovsky, A.; Picchini, A.M.; Moadel, T.; Sisti, A.C.; Yamada, A.; Kimura, S.; Leonardo, E.D.; Hen, R. Experience Dictates Stem Cell Fate in the Adult Hippocampus. Neuron 2011, 70, 908–923. [Google Scholar] [CrossRef] [PubMed]
- Lieberwirth, C.; Liu, Y.; Jia, X.; Wang, Z. Social Isolation Impairs Adult Neurogenesis in the Limbic System and Alters Behaviors in Female Prairie Voles. Hormones and Behavior 2012, 62, 357–366. [Google Scholar] [CrossRef]
- Westenbroek, C.; Den Boer, J.A.; Veenhuis, M.; Ter Horst, G.J. Chronic Stress and Social Housing Differentially Affect Neurogenesis in Male and Female Rats. Brain Research Bulletin 2004, 64, 303–308. [Google Scholar] [CrossRef]
- Biggio, F.; Mostallino, M.C.; Talani, G.; Locci, V.; Mostallino, R.; Calandra, G.; Sanna, E.; Biggio, G. Social Enrichment Reverses the Isolation-Induced Deficits of Neuronal Plasticity in the Hippocampus of Male Rats. Neuropharmacology 2019, 151, 45–54. [Google Scholar] [CrossRef]
- Scaccianoce, S.; Del Bianco, P.; Paolone, G.; Caprioli, D.; Modafferi, A.M.E.; Nencini, P.; Badiani, A. Social Isolation Selectively Reduces Hippocampal Brain-Derived Neurotrophic Factor without Altering Plasma Corticosterone. Behavioural Brain Research 2006, 168, 323–325. [Google Scholar] [CrossRef]
- Kowiański, P.; Lietzau, G.; Czuba, E.; Waśkow, M.; Steliga, A.; Moryś, J. BDNF: A Key Factor with Multipotent Impact on Brain Signaling and Synaptic Plasticity. Cell Mol Neurobiol 2018, 38, 579–593. [Google Scholar] [CrossRef]
- Castrén, E.; Antila, H. Neuronal Plasticity and Neurotrophic Factors in Drug Responses. Mol Psychiatry 2017, 22, 1085–1095. [Google Scholar] [CrossRef]
- Anacker, C.; Luna, V.M.; Stevens, G.S.; Millette, A.; Shores, R.; Jimenez, J.C.; Chen, B.; Hen, R. Hippocampal Neurogenesis Confers Stress Resilience by Inhibiting the Ventral Dentate Gyrus. Nature 2018, 559, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Hanif, S.; Sclar, M.; Lee, J.; Nichols, C.; Likhtik, E.; Burghardt, N.S. Social Isolation during Adolescence Differentially Affects Spatial Learning in Adult Male and Female Mice. Learn. Mem. 2025, 32, a054059. [Google Scholar] [CrossRef]
- Karimian, M.; Famitafreshi, H.; Fanaei, H.; Attari, F.; Fatima, S. Social Isolation Is Associated with Reduced Neurogenesis, Impaired Spatial Working Memory Performance, and Altered Anxiety Levels in Male Rats. OAAP 2015, 87. [Google Scholar] [CrossRef]
- Dimonte, S.; Sikora, V.; Bove, M.; Morgese, M.G.; Tucci, P.; Schiavone, S.; Trabace, L. Social Isolation from Early Life Induces Anxiety-like Behaviors in Adult Rats: Relation to Neuroendocrine and Neurochemical Dysfunctions. Biomedicine & Pharmacotherapy 2023, 158, 114181. [Google Scholar] [CrossRef]
- Skelly, M.J.; Chappell, A.E.; Carter, E.; Weiner, J.L. Adolescent Social Isolation Increases Anxiety-like Behavior and Ethanol Intake and Impairs Fear Extinction in Adulthood: Possible Role of Disrupted Noradrenergic Signaling. Neuropharmacology 2015, 97, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Flashner, B.; Chiu, M.; Ver Hoeve, E.; Luz, S.; Bhatnagar, S. Social Isolation in Adolescence Alters Behaviors in the Forced Swim and Sucrose Preference Tests in Female but Not in Male Rats. Physiology & Behavior 2012, 105, 269–275. [Google Scholar] [CrossRef]
- Hysing, M.; Petrie, K.J.; Harvey, A.G.; Sivertsen, B. From Loneliness to Depression: A Longitudinal Diagnostic Study among Norwegian University Students. Soc Psychiatry Psychiatr Epidemiol 2025. [Google Scholar] [CrossRef]
- Csikós, V.; Dóra, F.; Láng, T.; Darai, L.; Szendi, V.; Tóth, A.; Cservenák, M.; Dobolyi, A. Social Isolation Induces Changes in the Monoaminergic Signalling in the Rat Medial Prefrontal Cortex. Cells 2024, 13, 1043. [Google Scholar] [CrossRef]
- Benfato, I.D.; Quintanilha, A.C.S.; Henrique, J.S.; Souza, M.A.; Rosário, B.D.A.; Beserra Filho, J.I.A.; Santos, R.L.O.; Ribeiro, A.M.; Le Sueur Maluf, L.; De Oliveira, C.A.M. Effects of Long-Term Social Isolation on Central, Behavioural and Metabolic Parameters in Middle-Aged Mice. Behavioural Brain Research 2022, 417, 113630. [Google Scholar] [CrossRef]
- Paz, C.; Mascialino, G.; Adana-Díaz, L.; Rodríguez-Lorenzana, A.; Simbaña-Rivera, K.; Gómez-Barreno, L.; Troya, M.; Paez, M.I.; Cárdenas, J.; Gerstner, R.M.; et al. Behavioral and Sociodemographic Predictors of Anxiety and Depression in Patients under Epidemiological Surveillance for COVID-19 in Ecuador. PLoS ONE 2020, 15, e0240008. [Google Scholar] [CrossRef]
- Blanchard, D.C.; Spencer, R.L.; Weiss, S.M.; Blanchard, R.J.; McEwen, B.; Sakai, R.R. Visible Burrow System as a Model of Chronic Social Stress: Behavioral and Neuroendocrine Correlates. Psychoneuroendocrinology 1995, 20, 117–134. [Google Scholar] [CrossRef] [PubMed]
- Kozorovitskiy, Y.; Gould, E. Dominance Hierarchy Influences Adult Neurogenesis in the Dentate Gyrus: Figure 1. J. Neurosci. 2004, 24, 6755–6759. [Google Scholar] [CrossRef] [PubMed]
- Pallé, A.; Zorzo, C.; Luskey, V.E.; Mcgreevy, K.R.; Fernández, S.; Trejo, J.L. Social Dominance Differentially Alters Gene Expression in the Medial Prefrontal Cortex without Affecting Adult Hippocampal Neurogenesis or Stress and Anxiety-like Behavior. The FASEB Journal 2019, 33, 6995–7008. [Google Scholar] [CrossRef] [PubMed]
- Falconer, E.M.; Galea, L.A.M. Sex Differences in Cell Proliferation, Cell Death and Defensive Behavior Following Acute Predator Odor Stress in Adult Rats. Brain Research 2003, 975, 22–36. [Google Scholar] [CrossRef]
- Holmes, M.M.; Galea, L.A.M. Defensive Behavior and Hippocampal Cell Proliferation: Differential Modulation by Naltrexone during Stress. Behavioral Neuroscience 2002, 116, 160–168. [Google Scholar] [CrossRef]
- Hardy, M.P.; Sottas, C.M.; Ge, R.; McKittrick, C.R.; Tamashiro, K.L.; McEwen, B.S.; Haider, S.G.; Markham, C.M.; Blanchard, R.J.; Blanchard, D.C.; et al. Trends of Reproductive Hormones in Male Rats During Psychosocial Stress: Role of Glucocorticoid Metabolism in Behavioral Dominance1. Biology of Reproduction 2002, 67, 1750–1755. [Google Scholar] [CrossRef]
- Blanchard, R.J.; Yudko, E.; Dulloog, L.; Blanchard, D.C. Defense Changes in Stress Nonresponsive Subordinate Males in a Visible Burrow System. Physiology & Behavior 2001, 72, 635–642. [Google Scholar] [CrossRef]
- Wu, M.V.; Shamy, J.L.; Bedi, G.; Choi, C.-W.J.; Wall, M.M.; Arango, V.; Boldrini, M.; Foltin, R.W.; Hen, R. Impact of Social Status and Antidepressant Treatment on Neurogenesis in the Baboon Hippocampus. Neuropsychopharmacol 2014, 39, 1861–1871. [Google Scholar] [CrossRef]
- Peragine, D.E.; Simpson, J.A.; Mooney, S.J.; Lovern, M.B.; Holmes, M.M. Social Regulation of Adult Neurogenesis in a Eusocial Mammal. Neuroscience 2014, 268, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Golden, S.A.; Covington, H.E.; Berton, O.; Russo, S.J. A Standardized Protocol for Repeated Social Defeat Stress in Mice. Nat Protoc 2011, 6, 1183–1191. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, V.; Han, M.-H.; Graham, D.L.; Berton, O.; Renthal, W.; Russo, S.J.; LaPlant, Q.; Graham, A.; Lutter, M.; Lagace, D.C.; et al. Molecular Adaptations Underlying Susceptibility and Resistance to Social Defeat in Brain Reward Regions. Cell 2007, 131, 391–404. [Google Scholar] [CrossRef]
- Morrison, K.E.; Bader, L.R.; McLaughlin, C.N.; Cooper, M.A. Defeat-Induced Activation of the Ventral Medial Prefrontal Cortex Is Necessary for Resistance to Conditioned Defeat. Behavioural Brain Research 2013, 243, 158–164. [Google Scholar] [CrossRef]
- Jasnow, A.M.; Cooper, M.A.; Huhman, K.L. N-Methyl-d-Aspartate Receptors in the Amygdala Are Necessary for the Acquisition and Expression of Conditioned Defeat. Neuroscience 2004, 123, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Hodges, T.E.; Louth, E.L.; Bailey, C.D.C.; McCormick, C.M. Adolescent Social Instability Stress Alters Markers of Synaptic Plasticity and Dendritic Structure in the Medial Amygdala and Lateral Septum in Male Rats. Brain Struct Funct 2019, 224, 643–659. [Google Scholar] [CrossRef]
- McCormick, C.M.; Smith, K.; Baumbach, J.L.; De Lima, A.P.N.; Shaver, M.; Hodges, T.E.; Marcolin, M.L.; Ismail, N. Adolescent Social Instability Stress Leads to Immediate and Lasting Sex-Specific Changes in the Neuroendocrine-Immune-Gut Axis in Rats. Hormones and Behavior 2020, 126, 104845. [Google Scholar] [CrossRef]
- Adhikari, A.; Topiwala, M.A.; Gordon, J.A. Single Units in the Medial Prefrontal Cortex with Anxiety-Related Firing Patterns Are Preferentially Influenced by Ventral Hippocampal Activity. Neuron 2011, 71, 898–910. [Google Scholar] [CrossRef]
- Hamson, D.K.; Wainwright, S.R.; Taylor, J.R.; Jones, B.A.; Watson, N.V.; Galea, L.A.M. Androgens Increase Survival of Adult-Born Neurons in the Dentate Gyrus by an Androgen Receptor-Dependent Mechanism in Male Rats. Endocrinology 2013, 154, 3294–3304. [Google Scholar] [CrossRef] [PubMed]
- Spritzer, M.D.; Ibler, E.; Inglis, W.; Curtis, M.G. Testosterone and Social Isolation Influence Adult Neurogenesis in the Dentate Gyrus of Male Rats. Neuroscience 2011, 195, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Schoenfeld, T.J.; Gould, E. Stress, Stress Hormones, and Adult Neurogenesis. Experimental Neurology 2012, 233, 12–21. [Google Scholar] [CrossRef]
- Hostinar, C.E.; Sullivan, R.M.; Gunnar, M.R. Psychobiological Mechanisms Underlying the Social Buffering of the Hypothalamic–Pituitary–Adrenocortical Axis: A Review of Animal Models and Human Studies across Development. Psychological Bulletin 2014, 140, 256–282. [Google Scholar] [CrossRef]
- Gunnar, M.R.; Donzella, B. Social Regulation of the Cortisol Levels in Early Human Development. Psychoneuroendocrinology 2002, 27, 199–220. [Google Scholar] [CrossRef]
- Kiyokawa, Y.; Takeuchi, Y.; Mori, Y. Two Types of Social Buffering Differentially Mitigate Conditioned Fear Responses. Eur J of Neuroscience 2007, 26, 3606–3613. [Google Scholar] [CrossRef]
- Kiyokawa, Y.; Hiroshima, S.; Takeuchi, Y.; Mori, Y. Social Buffering Reduces Male Rats’ Behavioral and Corticosterone Responses to a Conditioned Stimulus. Hormones and Behavior 2014, 65, 114–118. [Google Scholar] [CrossRef]
- Caldwell, H.K. Oxytocin and Vasopressin: Powerful Regulators of Social Behavior. Neuroscientist 2017, 23, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Bielsky, I.F.; Young, L.J. Oxytocin, Vasopressin, and Social Recognition in Mammals. Peptides 2004, 25, 1565–1574. [Google Scholar] [CrossRef]
- Cilz, N.I.; Cymerblit-Sabba, A.; Young, W.S. Oxytocin and Vasopressin in the Rodent Hippocampus. Genes Brain and Behavior 2019, 18, e12535. [Google Scholar] [CrossRef]
- Wang, X.; Zhan, Y. Regulation of Social Recognition Memory in the Hippocampal Circuits. Front. Neural Circuits 2022, 16, 839931. [Google Scholar] [CrossRef]
- Lopez-Rojas, J.; De Solis, C.A.; Leroy, F.; Kandel, E.R.; Siegelbaum, S.A. A Direct Lateral Entorhinal Cortex to Hippocampal CA2 Circuit Conveys Social Information Required for Social Memory. Neuron 2022, 110, 1559–1572.e4. [Google Scholar] [CrossRef]
- Cope, E.C.; Waters, R.C.; Diethorn, E.J.; Pagliai, K.A.; Dias, C.G.; Tsuda, M.; Cameron, H.A.; Gould, E. Adult-Born Neurons in the Hippocampus Are Essential for Social Memory Maintenance. eNeuro 2020, 7, ENEURO.0182–20. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, B.M.M.; Moreira, F.A.; Massensini, A.R.; Moraes, M.F.D.; Pereira, G.S. Enriched Environment Increases Neurogenesis and Improves Social Memory Persistence in Socially Isolated Adult Mice: Neurogenesis Supports Social Memory Persistence. Hippocampus 2014, 24, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Bangasser, D.A.; Wicks, B. Sex-specific Mechanisms for Responding to Stress. J of Neuroscience Research 2017, 95, 75–82. [Google Scholar] [CrossRef]
- Gobinath, A.R.; Mahmoud, R.; Galea, L.A.M. Influence of Sex and Stress Exposure across the Lifespan on Endophenotypes of Depression: Focus on Behavior, Glucocorticoids, and Hippocampus. Front. Neurosci. 2015, 8. [Google Scholar] [CrossRef] [PubMed]
- Bonilla-Jaime, H.; Vázquez-Palacios, G.; Arteaga-Silva, M.; Retana-Márquez, S. Hormonal Responses to Different Sexually Related Conditions in Male Rats. Hormones and Behavior 2006, 49, 376–382. [Google Scholar] [CrossRef]
- Spritzer, M.D.; Galea, L.A.M. Testosterone and Dihydrotestosterone, but Not Estradiol, Enhance Survival of New Hippocampal Neurons in Adult Male Rats. Developmental Neurobiology 2007, 67, 1321–1333. [Google Scholar] [CrossRef]
- Shingo, T.; Gregg, C.; Enwere, E.; Fujikawa, H.; Hassam, R.; Geary, C.; Cross, J.C.; Weiss, S. Pregnancy-Stimulated Neurogenesis in the Adult Female Forebrain Mediated by Prolactin. Science 2003, 299, 117–120. [Google Scholar] [CrossRef]
- Leuner, B.; Glasper, E.R.; Gould, E. Parenting and Plasticity. Trends in Neurosciences 2010, 33, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Pawluski, J.L.; Galea, L.A.M. Reproductive Experience Alters Hippocampal Neurogenesis during the Postpartum Period in the Dam. Neuroscience 2007, 149, 53–67. [Google Scholar] [CrossRef]
- Bale, T.L.; Epperson, C.N. Sex Differences and Stress across the Lifespan. Nat Neurosci 2015, 18, 1413–1420. [Google Scholar] [CrossRef] [PubMed]
- Leuner, B.; Mirescu, C.; Noiman, L.; Gould, E. Maternal Experience Inhibits the Production of Immature Neurons in the Hippocampus during the Postpartum Period through Elevations in Adrenal Steroids. Hippocampus 2007, 17, 434–442. [Google Scholar] [CrossRef]
- Hillerer, K.M.; Neumann, I.D.; Couillard-Despres, S.; Aigner, L.; Slattery, D.A. Lactation-induced Reduction in Hippocampal Neurogenesis Is Reversed by Repeated Stress Exposure. Hippocampus 2014, 24, 673–683. [Google Scholar] [CrossRef] [PubMed]
- Maeng, L.Y.; Shors, T.J. Once a Mother, Always a Mother: Maternal Experience Protects Females from the Negative Effects of Stress on Learning. Behavioral Neuroscience 2012, 126, 137–141. [Google Scholar] [CrossRef]
- Torner, L. Actions of Prolactin in the Brain: From Physiological Adaptations to Stress and Neurogenesis to Psychopathology. Front. Endocrinol. 2016, 7. [Google Scholar] [CrossRef]
- Saltzman, W.; Ziegler, T.E. Functional Significance of Hormonal Changes in Mammalian Fathers. J Neuroendocrinology 2014, 26, 685–696. [Google Scholar] [CrossRef]
- Glasper, E.R.; Kozorovitskiy, Y.; Pavlic, A.; Gould, E. Paternal Experience Suppresses Adult Neurogenesis without Altering Hippocampal Function in Peromyscus Californicus. J of Comparative Neurology 2011, 519, 2271–2281. [Google Scholar] [CrossRef]
- Glasper, E.R.; Hyer, M.M.; Katakam, J.; Harper, R.; Ameri, C.; Wolz, T. Fatherhood Contributes to Increased Hippocampal Spine Density and Anxiety Regulation in California Mice. Brain and Behavior 2016, 6, e00416. [Google Scholar] [CrossRef]
- Cárdenas, E.F.; Kujawa, A.; Humphreys, K.L. Neurobiological Changes during the Peripartum Period: Implications for Health and Behavior. Social Cognitive and Affective Neuroscience 2020, 15, 1097–1110. [Google Scholar] [CrossRef]
- Aguirre, R.M.H.; González, M.H.; Hernández, M.P.; Gutiérrez, C.D.C.A.; Guevara, M.Á. Observing Baby or Sexual Videos Changes the Functional Synchronization between the Prefrontal and Parietal Cortices in Mothers in Different Postpartum Periods. Social Neuroscience 2020, 15, 489–504. [Google Scholar] [CrossRef]
- Kim, P.; Leckman, J.F.; Mayes, L.C.; Feldman, R.; Wang, X.; Swain, J.E. The Plasticity of Human Maternal Brain: Longitudinal Changes in Brain Anatomy during the Early Postpartum Period. Behavioral Neuroscience 2010, 124, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Pfaus, J.G. Pathways of Sexual Desire. The Journal of Sexual Medicine 2009, 6, 1506–1533. [Google Scholar] [CrossRef]
- Paredes, R.G.; Ågmo, A. Has Dopamine a Physiological Role in the Control of Sexual Behavior? Progress in Neurobiology 2004, 73, 179–225. [Google Scholar] [CrossRef]
- Barha, C.K.; Lieblich, S.E.; Galea, L.A.M. Different Forms of Oestrogen Rapidly Upregulate Cell Proliferation in the Dentate Gyrus of Adult Female Rats. J Neuroendocrinology 2009, 21, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Tanapat, P.; Hastings, N.B.; Reeves, A.J.; Gould, E. Estrogen Stimulates a Transient Increase in the Number of New Neurons in the Dentate Gyrus of the Adult Female Rat. J. Neurosci. 1999, 19, 5792–5801. [Google Scholar] [CrossRef]
- Pawluski, J.L.; Brummelte, S.; Barha, C.K.; Crozier, T.M.; Galea, L.A.M. Effects of Steroid Hormones on Neurogenesis in the Hippocampus of the Adult Female Rodent during the Estrous Cycle, Pregnancy, Lactation and Aging. Frontiers in Neuroendocrinology 2009, 30, 343–357. [Google Scholar] [CrossRef]
- Fares, J.; Bou Diab, Z.; Nabha, S.; Fares, Y. Neurogenesis in the Adult Hippocampus: History, Regulation, and Prospective Roles. International Journal of Neuroscience 2019, 129, 598–611. [Google Scholar] [CrossRef]
- Nicolis Di Robilant, V.; Scardigli, R.; Strimpakos, G.; Tirone, F.; Middei, S.; Scopa, C.; De Bardi, M.; Battistini, L.; Saraulli, D.; Farioli Vecchioli, S. Running-Activated Neural Stem Cells Enhance Subventricular Neurogenesis and Improve Olfactory Behavior in P21 Knockout Mice. Mol Neurobiol 2019, 56, 7534–7556. [Google Scholar] [CrossRef] [PubMed]
- Overall, R.W.; Walker, T.L.; Leiter, O.; Lenke, S.; Ruhwald, S.; Kempermann, G. Delayed and Transient Increase of Adult Hippocampal Neurogenesis by Physical Exercise in DBA/2 Mice. PLoS ONE 2013, 8, e83797. [Google Scholar] [CrossRef]
- Barha, C.K.; Ishrat, T.; Epp, J.R.; Galea, L.A.M.; Stein, D.G. Progesterone Treatment Normalizes the Levels of Cell Proliferation and Cell Death in the Dentate Gyrus of the Hippocampus after Traumatic Brain Injury. Experimental Neurology 2011, 231, 72–81. [Google Scholar] [CrossRef]
- Ormerod, B.K.; Lee, T.T.; Galea, L.A.M. Estradiol Initially Enhances but Subsequently Suppresses (via Adrenal Steroids) Granule Cell Proliferation in the Dentate Gyrus of Adult Female Rats. J. Neurobiol. 2003, 55, 247–260. [Google Scholar] [CrossRef]
- Mahmoud, R.; Wainwright, S.R.; Galea, L.A.M. Sex Hormones and Adult Hippocampal Neurogenesis: Regulation, Implications, and Potential Mechanisms. Frontiers in Neuroendocrinology 2016, 41, 129–152. [Google Scholar] [CrossRef]
- Nugent, B.; Tobet, S.; Lara, H.; Lucion, A.; Wilson, M.; Recabarren, S.; Paredes, A. Hormonal Programming Across the Lifespan. Horm Metab Res 2012, 44, 577–586. [Google Scholar] [CrossRef]
- Nogami, H.; Hoshino, R.; Ogasawara, K.; Miyamoto, S.; Hisano, S. Region-Specific Expression and Hormonal Regulation of the First Exon Variants of Rat Prolactin Receptor mRNA in Rat Brain and Anterior Pituitary Gland. J Neuroendocrinology 2007, 19, 583–593. [Google Scholar] [CrossRef]
- Mak, G.K.; Enwere, E.K.; Gregg, C.; Pakarainen, T.; Poutanen, M.; Huhtaniemi, I.; Weiss, S. Male Pheromone–Stimulated Neurogenesis in the Adult Female Brain: Possible Role in Mating Behavior. Nat Neurosci 2007, 10, 1003–1011. [Google Scholar] [CrossRef] [PubMed]
- Czéh, B.; Müller-Keuker, J.I.H.; Rygula, R.; Abumaria, N.; Hiemke, C.; Domenici, E.; Fuchs, E. Chronic Social Stress Inhibits Cell Proliferation in the Adult Medial Prefrontal Cortex: Hemispheric Asymmetry and Reversal by Fluoxetine Treatment. Neuropsychopharmacol 2007, 32, 1490–1503. [Google Scholar] [CrossRef] [PubMed]
- Mazzucco, C.A.; Lieblich, S.E.; Bingham, B.I.; Williamson, M.A.; Viau, V.; Galea, L.A.M. Both Estrogen Receptor α and Estrogen Receptor β Agonists Enhance Cell Proliferation in the Dentate Gyrus of Adult Female Rats. Neuroscience 2006, 141, 1793–1800. [Google Scholar] [CrossRef] [PubMed]
- McClure, R.E.S.; Barha, C.K.; Galea, L.A.M. 17β-Estradiol, but Not Estrone, Increases the Survival and Activation of New Neurons in the Hippocampus in Response to Spatial Memory in Adult Female Rats. Hormones and Behavior 2013, 63, 144–157. [Google Scholar] [CrossRef]
- Allen, K.M.; Fung, S.J.; Rothmond, D.A.; Noble, P.L.; Shannon Weickert, C. Gonadectomy Increases Neurogenesis in the Male Adolescent Rhesus Macaque Hippocampus: Gonadectomy And Adolescent Neurogenesis. Hippocampus 2014, 24, 225–238. [Google Scholar] [CrossRef]
- Carrier, N.; Kabbaj, M. Extracellular Signal-Regulated Kinase 2 Signaling in the Hippocampal Dentate Gyrus Mediates the Antidepressant Effects of Testosterone. Biological Psychiatry 2012, 71, 642–651. [Google Scholar] [CrossRef]
- Schoenfeld, T.J.; Rada, P.; Pieruzzini, P.R.; Hsueh, B.; Gould, E. Physical Exercise Prevents Stress-Induced Activation of Granule Neurons and Enhances Local Inhibitory Mechanisms in the Dentate Gyrus. J. Neurosci. 2013, 33, 7770–7777. [Google Scholar] [CrossRef] [PubMed]
- Potretzke, S.; Ryabinin, A.E. The Prairie Vole Model of Pair-Bonding and Its Sensitivity to Addictive Substances. Front. Psychol. 2019, 10, 2477. [Google Scholar] [CrossRef] [PubMed]
- Castro, A.E.; Domínguez-Ordoñez, R.; Young, L.J.; Camacho, F.J.; Ávila-González, D.; Paredes, R.G.; Díaz, N.F.; Portillo, W. Pair-Bonding and Social Experience Modulate New Neurons Survival in Adult Male and Female Prairie Voles (Microtus Ochrogaster). Front. Neuroanat. 2022, 16, 987229. [Google Scholar] [CrossRef]
- Amrein, I.; Becker, A.S.; Engler, S.; Huang, S.; Müller, J.; Slomianka, L.; Oosthuizen, M.K. Adult Neurogenesis and Its Anatomical Context in the Hippocampus of Three Mole-Rat Species. Front. Neuroanat. 2014, 8. [Google Scholar] [CrossRef]
- Oosthuizen, M.K.; Amrein, I. Trading New Neurons for Status: Adult Hippocampal Neurogenesis in Eusocial Damaraland Mole-Rats. Neuroscience 2016, 324, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xu, N.-N.; Hao, Z.-Z.; Liu, S. Zoological Research 2023, 44, 315–322. [CrossRef]
- Kornack, D.R.; Rakic, P. Continuation of Neurogenesis in the Hippocampus of the Adult Macaque Monkey. Proc. Natl. Acad. Sci. U.S.A. 1999, 96, 5768–5773. [Google Scholar] [CrossRef]
- Larson, T.A.; Wang, T.-W.; Gale, S.D.; Miller, K.E.; Thatra, N.M.; Caras, M.L.; Perkel, D.J.; Brenowitz, E.A. Postsynaptic Neural Activity Regulates Neuronal Addition in the Adult Avian Song Control System. Proc. Natl. Acad. Sci. U.S.A. 2013, 110, 16640–16644. [Google Scholar] [CrossRef]
- Brenowitz, E.A.; Larson, T.A. Neurogenesis in the Adult Avian Song-Control System. Cold Spring Harb Perspect Biol 2015, 7, a019000. [Google Scholar] [CrossRef] [PubMed]
- Ganz, J.; Brand, M. Adult Neurogenesis in Fish. Cold Spring Harb Perspect Biol 2016, 8, a019018. [Google Scholar] [CrossRef] [PubMed]
- Teles, M.C.; Faustino, F.; Chanfana, C.; Cunha, A.; Esteves, M.; Oliveira, R.F. Social Enhancement of Adult Neurogenesis in Zebrafish Is Not Regulated by Cortisol. Neuroscience 2023, 509, 51–62. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Age-dependent effects of social isolation on adult neurogenesis and behavior.
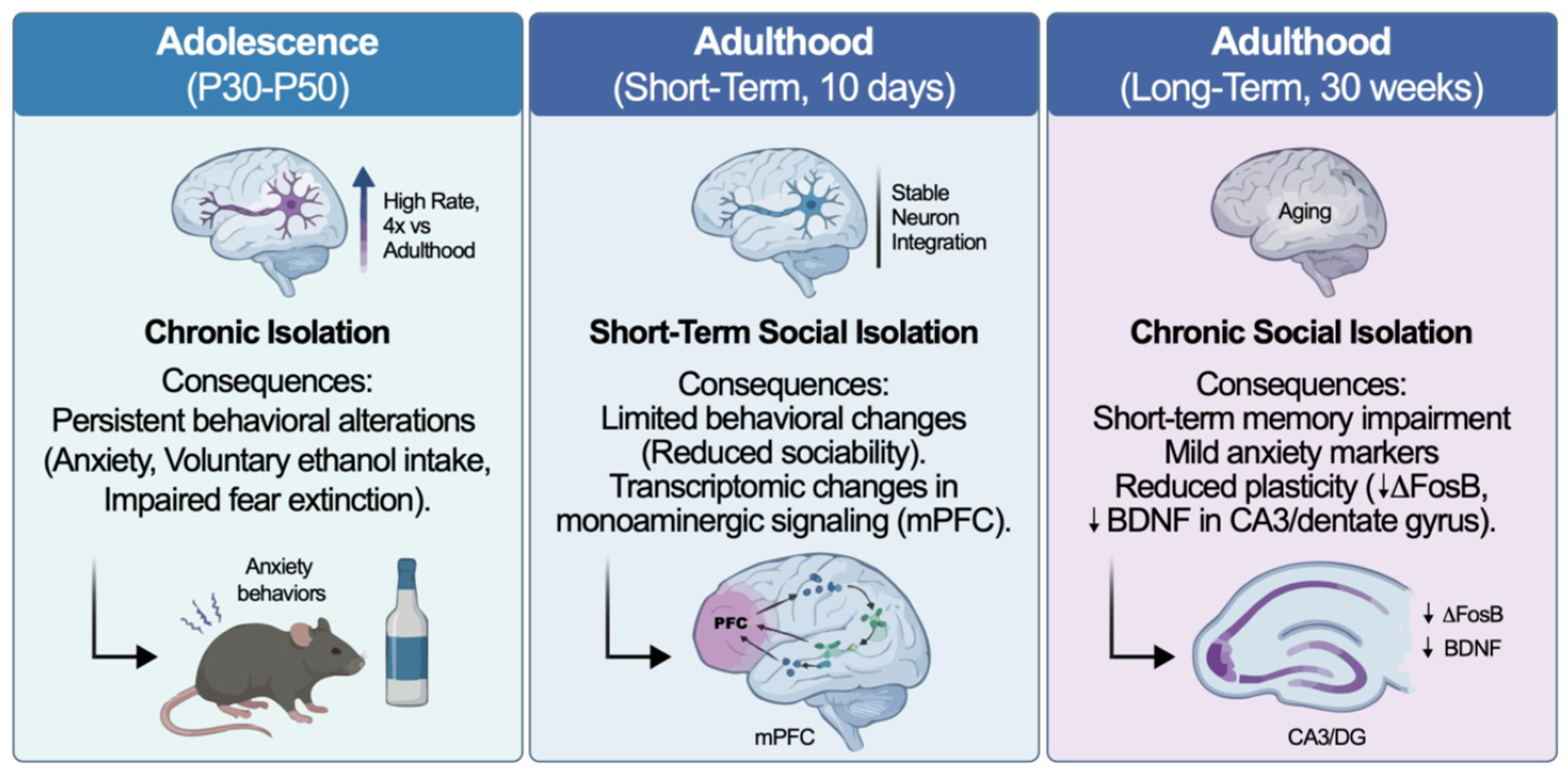
Figure 2.
Functional and anatomical dissociation of dorsal and ventral dentate gyrus in social isolation.
Figure 2.
Functional and anatomical dissociation of dorsal and ventral dentate gyrus in social isolation.
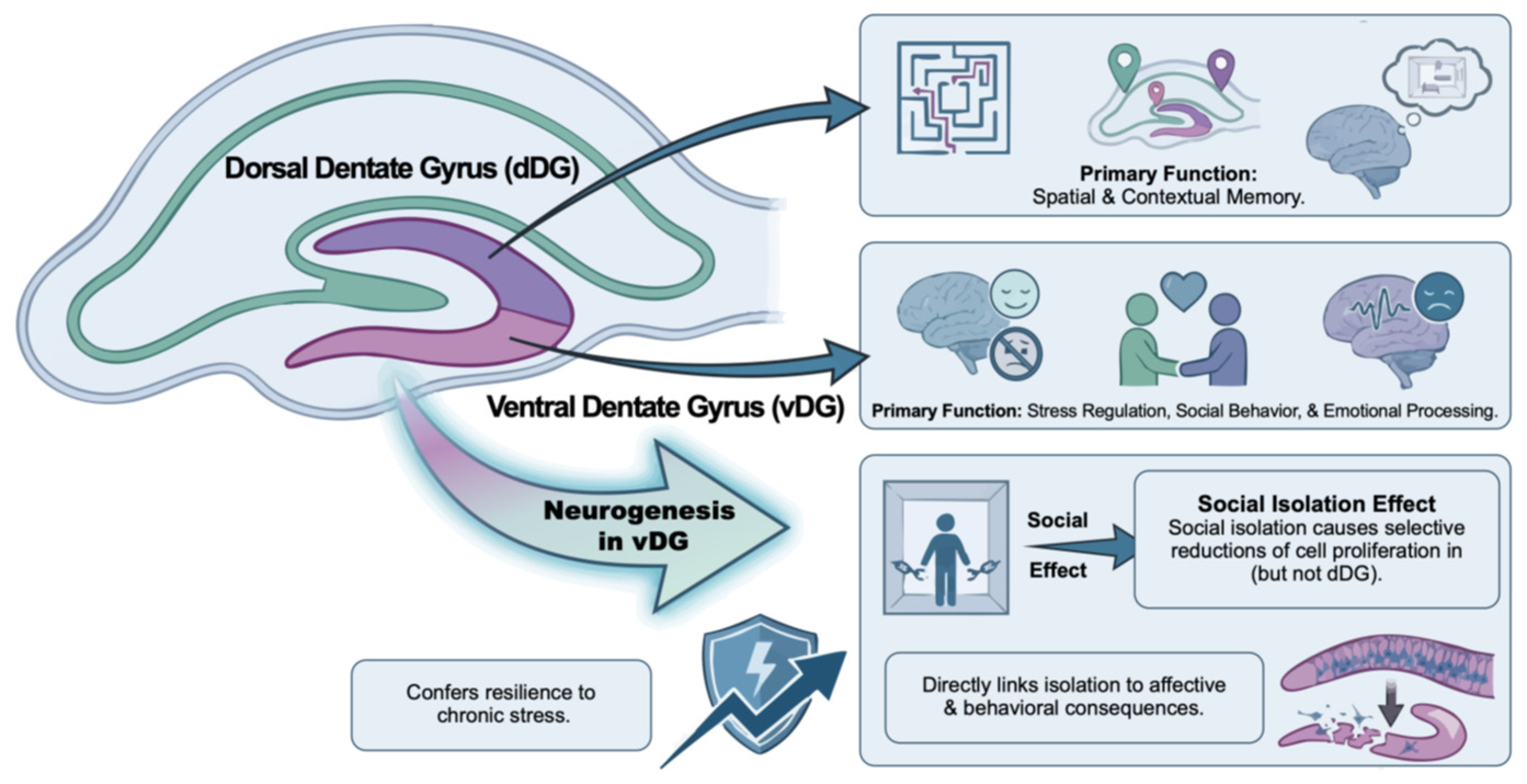
Figure 3.
Social buffering as a neuroprotective mechanism against stress-induced suppression of neurogenesis.
Figure 3.
Social buffering as a neuroprotective mechanism against stress-induced suppression of neurogenesis.
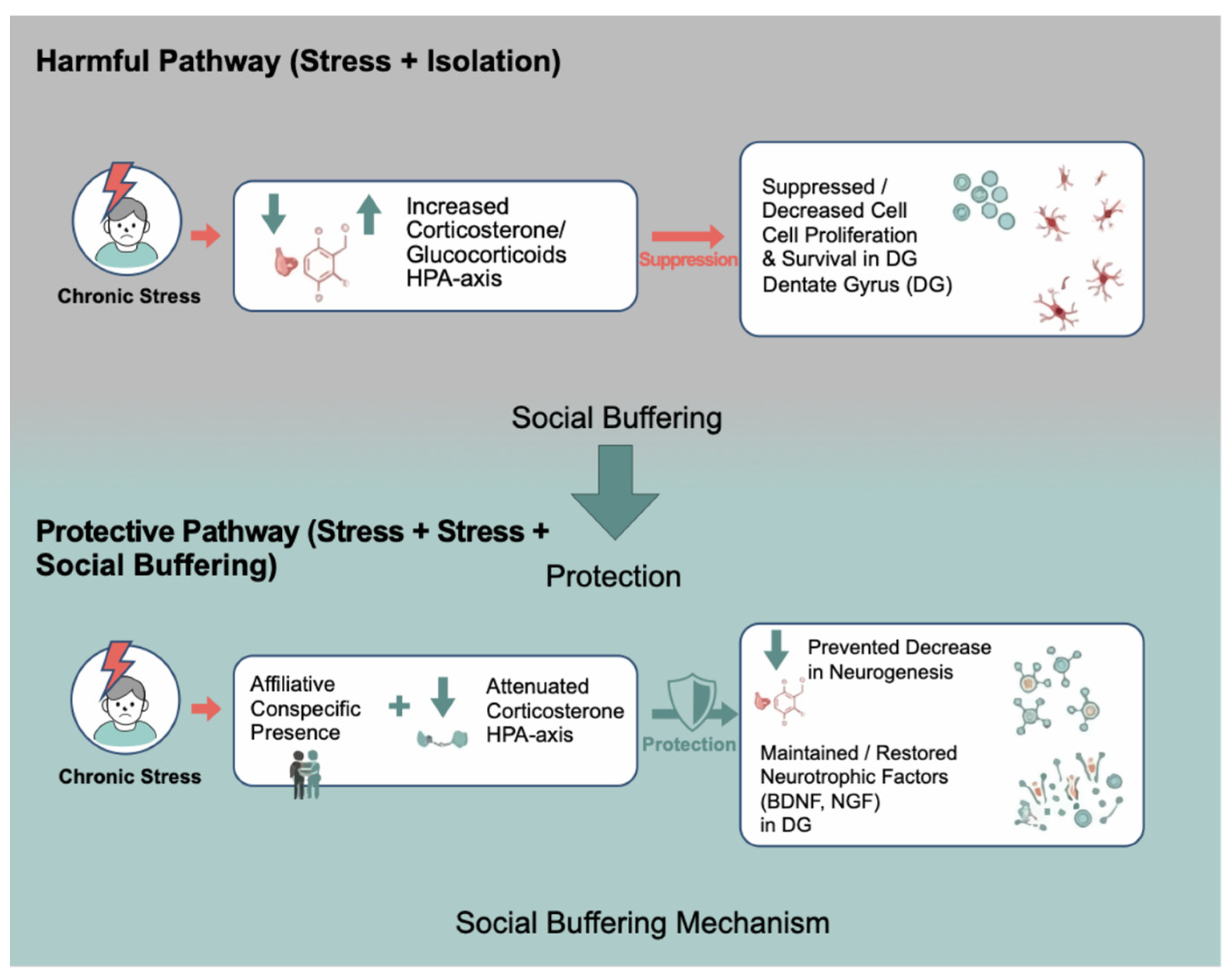
Figure 4.
Hormonal modulation of adult neurogenesis.
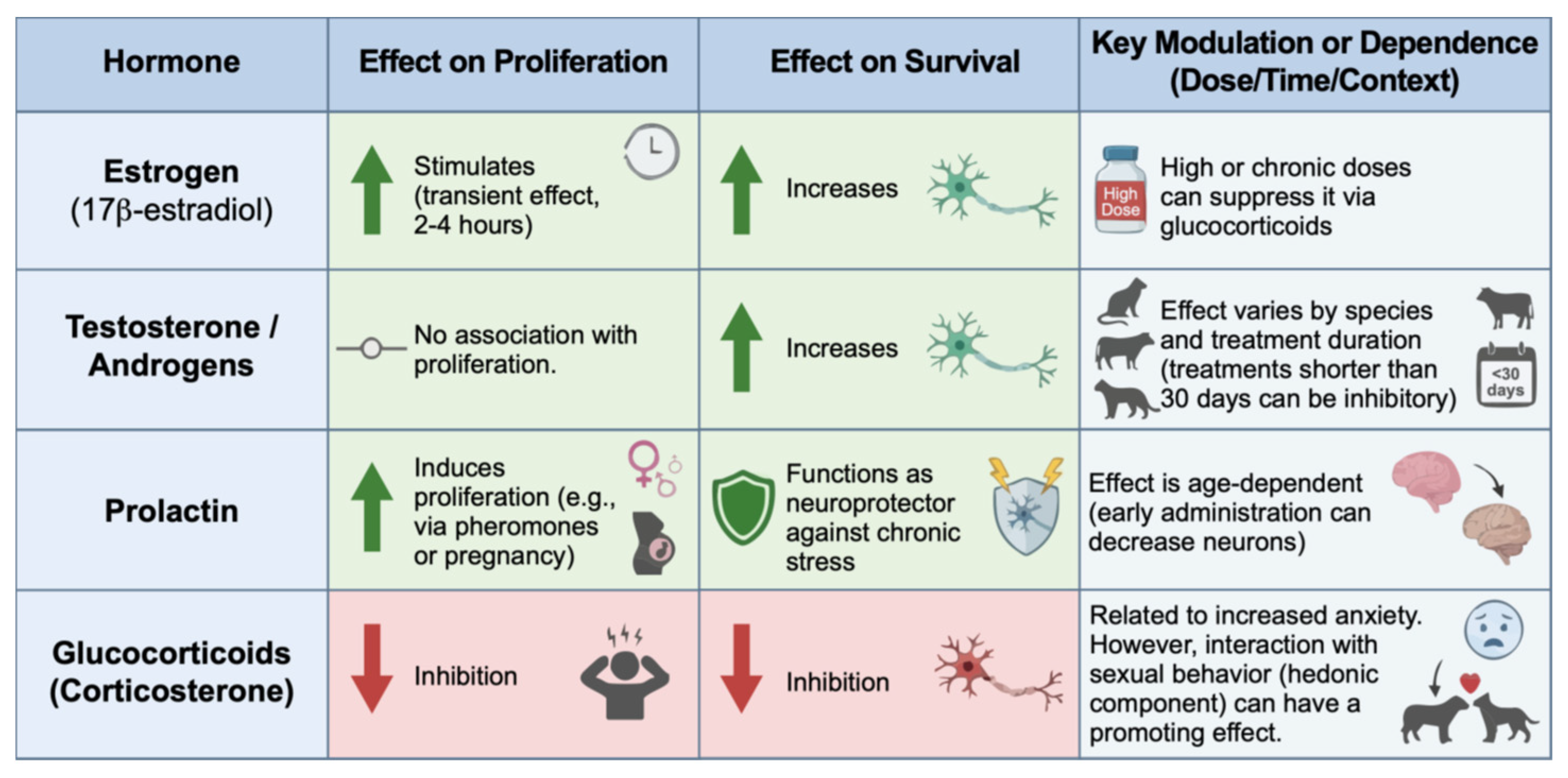
Figure 5.
Social and endocrine modulation of adult neurogenesis across neurogenic stages.
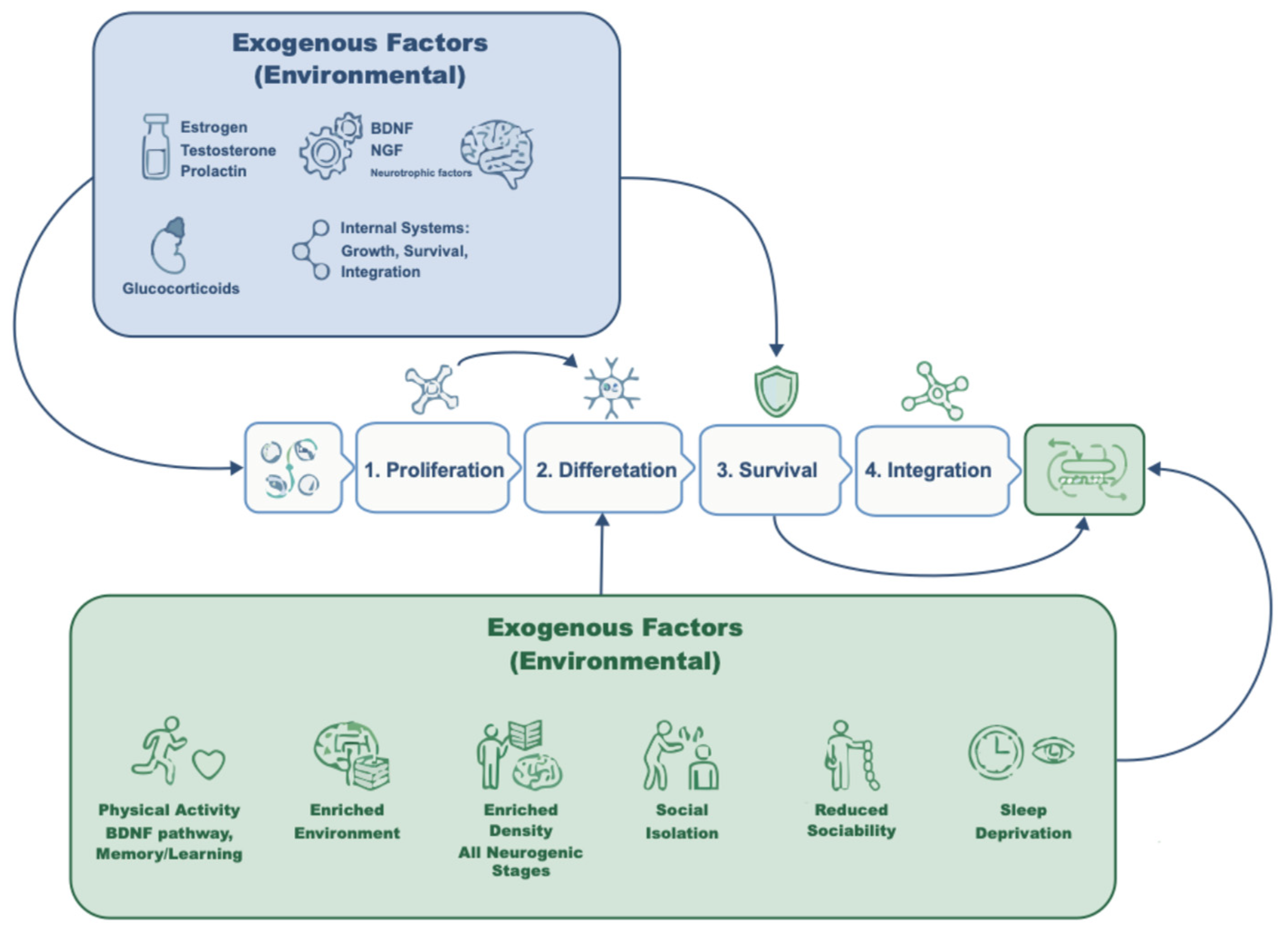

Table 1.
Behavioral and Neurobiological Effects of Social Isolation Across Developmental Stages.
| Age / Stage | Species / Model | Duration / Timing of Isolation | Observed Effects | Reference |
| Early life | Rats | From early life (postnatal) into development; long-term. | Early-life social isolation leads to anxiety-like behavior in adulthood, plus marked neuroendocrine and neurochemical alterations (e.g., oxytocin, ghrelin, melatonin, monoamines). | [39] |
| Adolescence | Rats | Chronic isolation P30–P50 (adolescence). | Adolescent isolation produces stable changes in adult behavior, with altered forced swim and sucrose preference (active coping and reward-related behavior), particularly in females. | [41] |
| Rats | Chronic isolation during adolescence. | Adolescent social isolation increases anxiety-like behavior, voluntary ethanol intake, and difficults conditioned fear extinction on adulthood. | [40] | |
| Early Adulthood | Humans (young adults / students) | Longitudinal exposure to high loneliness / isolation. | Loneliness and social isolation predict major depressive episodes and poorer mental health over time, even after controlling for baseline mental health. | [42] |
| Adulthood | Rats | Short-term isolated housing in adulthood | Short-term social isolation (10 days) produced limited behavioral changes (mostly reduced sociability), while RNA-seq of mPFC revealed significant differential expression of genes related to dopaminergic and monoaminergic signalling. | [43] |
| Mice | Long term (30 weeks). | Long-term adult social isolation impaired short-term memory and produced mild anxiety-related markers, together with reduced ΔFosB activity in the dentate gyrus and lower BDNF expression in CA3, indicating diminished hippocampal plasticity. | [44] | |
| Humans | Short-term mandatory at-home quarantine. | Elevated anxiety and depressive symptoms; behavioral (physical inactivity, poor sleep, increased media use) and sociodemographic factors (female sex, lower education) significantly predicted symptom severity. | [45] |
Abbreviations: mPFC, medial prefrontal cortex; RNA-seq, RNA sequencing; ΔFosB, truncated FosB transcription factor; BDNF, brain-derived neurotrophic factor; CA3, cornu ammonis 3 (hippocampal subfield).
Table 2.
Endocrine and neurobiological changes across stages of parental behavior.
| Phase | Dominant Hormones | Neural / Behavioral effects |
| Pregnancy | Rise in estrogens, progesterone, cortisol, and a strong prolactin surge. | Prolactin-driven SVZ proliferation and increased olfactory bulb neurogenesis prepare maternal circuits. |
| Early Postpartum (0–3 months) | Sharp drop in estrogens/progesterone; prolactin and oxytocin remain elevated; cortisol peaks around birth then declines. | Enhanced PFC–parietal and reward-circuit activity to infant cues; heightened maternal sensitivity. |
| Lactation | Sustained prolactin and oxytocin; altered HPA activity with elevated or modulated glucocorticoids. | Reduced hippocampal neurogenesis under baseline conditions; stress exposure can normalize neurogenesis and glucocorticoid levels. |
| Later Postpartum (≥4 months) | Gradual normalization of prolactin, oxytocin, and cortisol; increasing hormonal variability across women. | Refinement of prefrontal–parietal synchronization and continued tuning of maternal cortical networks. |
| Paternal Care | Increased prolactin and oxytocin; reduced testosterone; species-specific glucocorticoid/vasopressin changes. | Paternal experience suppresses hippocampal neurogenesis; fatherhood increases dendritic spine density and reduces anxiety-like behavior. |
Table 3.
Hormonal and Neurochemical Systems Influencing Adult DG/SVZ Plasticity and Social Behavior.
Table 3.
Hormonal and Neurochemical Systems Influencing Adult DG/SVZ Plasticity and Social Behavior.
| Hormone / system | Effect on DG/SVZ neurogenesis / plasticity | Relation to social behavior | References |
| Estradiol (ovarian estrogens) | Rapid but transient increase in progenitor proliferation in the dentate gyrus after acute administration; effect peaks within hours and disappears by 24–48 h. | Estradiol levels fluctuate across the estrous cycle and reproductive states, coordinating hippocampal plasticity with female reproductive physiology and sexual receptivity. | [96] |
| Androgens (testosterone, DHT) | Do not enhance proliferation, but increase survival of adult-born granule cells in the DG through androgen receptor–dependent mechanisms. | Androgen-sensitive pathways link hippocampal plasticity to male reproductive status and social/sexual contexts, where testosterone fluctuates with competition and mating. | [79] |
| Prolactin | Pregnancy-stimulated proliferation in the SVZ and increased neurogenesis in the olfactory bulb; effects are blocked by prolactin antagonism and mimicked by prolactin infusion. | These changes are interpreted as preparatory adaptations for maternal behavior, enhancing olfactory processing of offspring-related cues. | [80] |
| Oxytocin / vasopressin | Modulate hippocampal synaptic transmission and plasticity (e.g., CA2, DG) rather than directly altering proliferation; influence how existing circuits encode socially relevant information. | Oxytocin and vasopressin signaling in the hippocampus and related regions is critical for social recognition and social memory, supporting affiliative and pair-bonding behaviors. | [71] |
| BDNF and neurotrophins | Support survival, dendritic growth and functional integration of adult-born neurons in DG, and mediate activity-dependent reorganization of hippocampal circuits. | BDNF levels are strongly modulated by experience (exercise, enrichment, stress) and are tightly linked to learning, memory, and affective regulation in social contexts. | [34] |
| Monoaminergic and stress/inflammatory pathways (dopamine, serotonin, glucocorticoids, cytokines) | Chronic social isolation and social defeat reduce DG cell proliferation and neuronal activity and decrease BDNF expression in hippocampal subfields (DG ΔFosB↓, CA3 BDNF↓), while acute pharmacological modulation of monoamines can reverse or buffer some of these effects. | Social reward and stress are transduced by dopaminergic and monoaminergic circuits: dopamine in VTA–NAc supports sexual and social reward; long-term isolation or defeat alters monoaminergic gene expression in mPFC and produces anxiety/depression-like behavior, with associated reductions in hippocampal plasticity. | [43,44,95,108] |
Abbreviations: DG, dentate gyrus; SVZ, subventricular zone; CA2/CA3, cornu ammonis subfields (hippocampal subfields); ΔFosB, truncated FosB transcription factor; VTA, ventral tegmental area; NAc, nucleus accumbens; mPFC, medial prefrontal cortex; DHT, dihydrotestosterone.
Table 4.
Comparative species models linking social organization to adult neurogenesis.
| Model / species group | Type of sociality | Adult neurogenesis – key findings | Evolutionary / functional particularities | References |
| Prairie vole (Microtus ochrogaster) | Social monogamy; stable pair bonds; biparental care | Adult-born cells in the dentate gyrus and olfactory bulb are modulated by cohabitation with mating and opposite-sex social exposure. These social conditions enhance survival and differentiation of new neurons, with sex-dependent effects, aligning with mechanisms underlying pair-bond formation. | Specialized oxytocin and vasopressin systems in mesolimbic–social circuits support selective partner preference and long-term attachments, making prairie voles a canonical model of mammalian social bonding. | [114,115] |
| Naked mole-rat and eusocial mole-rats | Eusociality: reproductive castes; cooperative colonies | Eusocial mole-rats show markedly reduced adult hippocampal neurogenesis relative to surface-dwelling rodents, despite preserved progenitor markers. In naked mole-rats, new cells exhibit slow maturation, and neurogenesis is low but sustained across the lifespan, consistent with exceptional longevity. | Subterranean, hypoxic ecology and extreme lifespan correlate with a small, developmentally protracted hippocampus and atypical expression of calcium-binding proteins. These features are often interpreted as indicators of neoteny-like brain organization in mole-rats. | [116,117] |
| Nonhuman primates (macaques and other monkeys) | Complex dominance hierarchies; alliances; long-term social relationships | Adult hippocampal neurogenesis is clearly present but occurs at ~10-fold lower rates than in rodents. New granule cells mature over many months, and AHN declines with age, reaching very low levels in older primates. | Large brains, extended developmental trajectories, and sophisticated social cognition rely more on synaptic and structural plasticity than on high neuronal turnover, positioning primates as a key translational bridge for understanding human AHN. | [118,119] |
| Songbirds (oscine songbirds: canaries, zebra finches, etc.) | Learned vocal communication; seasonal territoriality; mate attraction via song | Song-control nuclei (HVC, RA, Area X) exhibit robust adult neurogenesis. Recruitment and survival of new neurons are regulated by photoperiod, gonadal hormones, singing activity, and social context, tightly linking neurogenesis to song learning and seasonal song plasticity. | Songbirds are the classic demonstration that adult-born projection neurons directly support a socially relevant learned behavior. Strong seasonal dynamics make them an ideal model to study endocrine, environmental, and social regulation of neurogenesis at a circuit level. | [120,121] |
| Teleost fish (e.g., zebrafish, trout) | Schooling / shoaling; group living; rich social signaling | Teleost fish display extensive adult neurogenesis across multiple proliferative zones along the rostro–caudal brain. Although the cited work on social plasticity does not measure neurogenesis directly, it demonstrates that social context modulates neural activity and gene expression in circuits involved in social behavior, highlighting the high adaptability of teleost neural systems. | Their lifelong brain growth and exceptional regenerative capacity make teleosts a valuable comparative model for studying neural plasticity, offering a clear contrast with the limited neurogenic potential observed in adult mammals. | [122,123] |
Abbreviations: AHN, adult hippocampal neurogenesis; HVC, high vocal center; RA, robust nucleus of the arcopallium; Area X, basal-ganglia–related song nucleus.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



