Submitted:
01 February 2026
Posted:
05 February 2026
You are already at the latest version
Abstract
Obsessive–compulsive disorder (OCD) is a disabling psychiatric condition traditionally framed in terms of serotonergic dysfunction and aberrant cortico–striato–thalamo–cortical (CSTC) circuitry. While influential, these models do not fully explain key features of OCD, including cognitive rigidity, exaggerated error monitoring, impaired resolution of uncertainty, and the persistence of compulsive behaviours despite preserved insight. Here, we propose that the brain histamine system represents an under-recognised neuromodulatory contributor to OCD, with the capacity to shape circuit dynamics rather than encode obsessional content.Histaminergic neurons arising from the tuberomammillary nucleus project diffusely across cortical and subcortical regions implicated in OCD, positioning histamine as a regulator of neural gain, arousal, and learning flexibility at the systems level. Integrating evidence from neuroanatomy, receptor pharmacology, circuit neuroscience, genetics, and computational psychiatry, we outline a framework in which histamine influences the stability and adaptability of CSTC loops. Receptor-specific actions, particularly via presynaptic H₃ receptors that regulate histamine and other neuromodulators, provide a mechanism by which histaminergic signalling may bias inhibitory control and habitual responding.From a computational perspective, histamine is well suited to modulate the trade-off between stability and flexibility in learning under uncertainty, a process increasingly implicated in OCD. We conclude by outlining testable hypotheses and experimental strategies to evaluate histaminergic modulation of CSTC dynamics and its relevance for circuit-informed investigation in OCD.
Keywords:
obsessive–compulsive disorder
; histaminergic system
; cortico–striato–thalamo–cortical circuits
; H3 receptor
; learning under uncertainty
Introduction
Obsessive–compulsive disorder (OCD) is a chronic and disabling psychiatric condition characterised by intrusive, unwanted thoughts and repetitive behaviours performed in an attempt to reduce distress or prevent feared outcomes [1]. Affecting approximately 1–3% of the population, OCD is associated with substantial functional impairment, reduced quality of life, and elevated comorbidity with anxiety, mood, and neurodevelopmental disorders [1]. Despite decades of research and the availability of evidence-based treatments, a significant proportion of patients show only partial response or remain treatment resistant [1]. This persistent unmet clinical need reflects, at least in part, an incomplete understanding of the neurobiological mechanisms that sustain compulsivity, cognitive rigidity, and maladaptive responses to uncertainty [1]. This motivates the exploration of complementary and alternative neurobiological mechanisms that regulate neural gain, learning flexibility, and responses to uncertainty, and that may interact with—rather than replace—classical circuit-based models.
Contemporary models converge on the notion that OCD arises from dysfunction within cortico–striato–thalamo–cortical (CSTC) circuits linking the orbitofrontal cortex, anterior cingulate cortex, striatum, and thalamus [1,2]. Neuroimaging, lesion, and neuromodulation studies consistently implicate these loops in symptom expression and treatment response [2,3]. However, while circuit-level models have provided a powerful anatomical framework, they do not fully explain why CSTC activity becomes persistently rigid, why error and threat signals are over-amplified, or why learning remains maladaptive even when patients retain insight into the irrationality of their symptoms. These limitations suggest that additional, higher-order mechanisms regulating circuit dynamics may be critical to OCD pathophysiology [1].
Historically, neurochemical models of OCD have been dominated by serotonin and, to a lesser extent, dopamine [1,4,5]. The clinical efficacy of selective serotonin reuptake inhibitors and the augmentation benefits of dopamine antagonists underscore the relevance of these systems. Yet serotonergic models alone struggle to account for core cognitive features such as inflexible belief updating, exaggerated uncertainty sensitivity, and habitual responding. Moreover, serotonin-targeting treatments often require high doses and prolonged administration, and many patients fail to achieve full remission. Dopaminergic accounts, while offering a clearer link to habit formation and striatal dysfunction, also face important limitations [5]. Dopamine-centric models primarily explain compulsivity in terms of aberrant reinforcement learning or over-reliance on habitual control, yet they do not readily account for the persistent generation of error signals, heightened monitoring, and subjective sense of incompleteness that characterise OCD, particularly in patients with preserved insight. Moreover, dopaminergic abnormalities in OCD appear heterogeneous and subtle, and dopamine-targeting interventions are typically effective only as adjuncts rather than primary treatments [5]. More broadly, both serotonin and dopamine models tend to focus on synapse- or circuit-specific mechanisms, offering limited explanatory power for why cortico–striato–thalamo–cortical circuits become globally locked into a high-gain, low-flexibility operating regime. Clinically, serotonin-targeting treatments often require high doses and prolonged administration, and a substantial proportion of patients fail to achieve full remission. Together, these limitations have prompted renewed interest in neuromodulatory systems that regulate brain state, learning, and flexibility at a more global level, rather than encoding specific cognitive or motivational content [1].
Within this context, the brain histamine system represents a compelling but underexplored candidate. Neuronal histamine is synthesised by a small population of neurons in the tuberomammillary nucleus of the posterior hypothalamus, which project diffusely across cortical and subcortical regions, including all major nodes of CSTC circuits [6]. Rather than conveying specific sensory or motivational content, histamine modulates arousal, vigilance, attentional gain, and the readiness of neural circuits to update in response to change [6,7]. These functions map closely onto clinical and cognitive features of OCD, which is marked by heightened alertness, persistent monitoring for errors or threats, and a paradoxical coexistence of hypervigilance and behavioural rigidity [8].
At the synaptic level, histamine acts through multiple receptor subtypes with distinct anatomical distributions and functional effects [8]. H1 and H2 receptors primarily exert postsynaptic actions that influence cortical excitability and plasticity, whereas H3 receptors function as presynaptic auto- and heteroreceptors regulating the release of histamine itself as well as other key neurotransmitters, including dopamine, serotonin, noradrenaline, glutamate, and GABA [9]. This unique pharmacology positions histamine, and H3 receptors in particular, as a meta-modulatory system capable of shaping the balance of excitation, inhibition, and neuromodulatory tone within CSTC loops. Such a role is especially relevant to disorders of compulsivity, where subtle shifts in gain and inhibition can have outsized behavioural consequences.
Recent advances in computational psychiatry further strengthen the case for histamine in OCD. Increasingly, OCD is conceptualised as a disorder of learning under uncertainty, characterised by aberrant estimates of environmental volatility, persistent prediction errors, and a failure to appropriately update internal models and action policies [10]. Histamine is well suited to regulate these processes, as it is posed to dynamically adjust acknowledging brain responsiveness to novelty and change while interacting closely with dopaminergic and noradrenergic systems that encode prediction errors and uncertainty. Dysregulated histaminergic signalling could therefore contribute to a pathological state in which error signals are repeatedly generated but never fully resolved, driving compulsive behaviours as attempts to regain a sense of control.
Despite these converging theoretical considerations, the role of histamine in OCD has received comparatively little systematic attention [11]. Existing evidence is scattered across basic neuroscience, genetics, pharmacology, and clinical observations, and has rarely been integrated into a coherent framework. The aim of this review is to address this gap by synthesising current knowledge on the brain histamine system and evaluating its relevance to OCD from a circuit, computational, and translational perspective. We argue that histamine is not a peripheral or ancillary player, but a candidate regulator, complementary axis of neural gain and flexibility whose dysregulation may help explain key features of OCD that remain insufficiently accounted for by existing models. By integrating molecular, circuit-level, and computational insights, we aim to provide a foundation for future experimental work and to highlight novel therapeutic opportunities, including the targeted modulation of histaminergic signalling in OCD.
Neuroanatomy of the Brain Histamine System and Its Relevance to OCD Circuits
Any attempt to understand the role of histamine in obsessive–compulsive disorder must begin with its highly distinctive neuroanatomical organization [6,7] (summary in Figure 1). Unlike most neurotransmitter systems, which comprise multiple nuclei with partially segregated projection fields, the entire central histaminergic system originates almost exclusively from a compact cluster of neurons in the tuberomammillary nucleus of the posterior hypothalamus. From this anatomically circumscribed source, histaminergic axons project diffusely and extensively throughout the brain, innervating virtually all cortical and subcortical structures implicated in OCD, including the orbitofrontal cortex, anterior cingulate cortex, dorsomedial and ventral striatum, globus pallidus, thalamus, hippocampus, and cerebellum [6,7,9].
This pattern of widespread innervation has important functional implications. Histamine is thought to not be organised to transmit specific information between defined nodes, but to impose a global modulatory context within which local computations unfold [12]. In CSTC circuits, where small shifts in excitation–inhibition balance can profoundly alter action selection and behavioural output, such diffuse modulation is particularly powerful [13,14]. It is an empirical question to what extent histamine can simultaneously influence cortical monitoring systems, striatal gating mechanisms, and thalamic relay fidelity, thereby coordinating how these elements interact rather than acting on any single component in isolation.
Within the cortex, histaminergic fibres densely innervate regions central to OCD symptomatology [15]. The orbitofrontal cortex, which encodes value, outcome expectations, and error signals, receives strong histaminergic input that modulates pyramidal neuron excitability and interneuron function [16]. Similarly, the anterior cingulate cortex, a key node in performance monitoring and conflict detection, is richly innervated by histaminergic projections [17]. Through these pathways, histamine is positioned to influence the intensity and persistence of error-related signalling, a hallmark feature of OCD in both behavioural and neuroimaging studies.
In the striatum, histamine interacts with the principal machinery of action selection [18]. Medium spiny neurons of both the direct and indirect pathways express histamine receptors, and histaminergic terminals are closely interwoven with dopaminergic inputs from the substantia nigra and ventral tegmental area [19,20,21]. This anatomical convergence is probably already present during early postdevelopment [22] and highly relevant to compulsivity. CSTC models of OCD emphasise an imbalance between facilitation and suppression of action tendencies, leading to repetitive, habitual behaviours that are difficult to inhibit. By modulating dopaminergic tone and synaptic integration within striatal microcircuits, histamine can bias this balance toward either flexible updating or perseverative repetition, depending on receptor engagement and overall system state [23].
The thalamus represents another critical interface [24,25]. Acting as both a relay and a gatekeeper, the thalamus determines which cortical signals are amplified and which are suppressed. Histaminergic projections to thalamic nuclei modulate thalamocortical excitability and sensory throughput, influencing how strongly internally generated signals, such as intrusive thoughts or error representations, gain access to conscious awareness [24,25]. In OCD, where thalamocortical hyperconnectivity has been repeatedly observed, altered histaminergic modulation may contribute to excessive reverberation within CSTC loops [26].
Importantly, the diffuse nature of histaminergic projections allows for coordinated modulation across these regions [6,7]. This stands in contrast to more spatially constrained neuromodulatory systems and suggests that histamine may synchronise the functional state of CSTC loops as a whole. In OCD, such synchronisation may become maladaptive, locking the system into a high-gain, low-flexibility regime in which intrusive signals dominate and behavioural output becomes stereotyped.
This anatomical framework also helps to explain why histamine is particularly relevant to state-dependent features of OCD. Symptoms often fluctuate with stress, sleep disruption, and arousal—conditions known to strongly influence histaminergic activity [27]. The tuberomammillary nucleus is tightly coupled to circadian and homeostatic mechanisms regulating wakefulness, linking histamine not only to cognition and behaviour, but also to the temporal organisation of brain states [28]. Dysregulation at this level could therefore amplify CSTC dysfunction during periods of heightened arousal or fatigue, contributing to symptom exacerbation.
Taken together, the neuroanatomy of the brain histamine system positions it as a plausible contributor to the regulation of distributed circuits implicated in OCD. Its diffuse projections, convergence with dopaminergic and serotonergic systems, and presence across cortical, striatal, and thalamic nodes indicate that histamine is well suited to modulate the global operating conditions under which CSTC loops function, rather than to encode circuit-specific information or behavioural content. Importantly, diffuse innervation alone does not imply coordinated control across these elements; whether and how histaminergic signalling exerts integrative effects across CSTC circuits remains an open empirical question. Nevertheless, this anatomical organisation provides a necessary—though not sufficient—substrate for exploring how receptor-specific actions may influence circuit dynamics and computational processes relevant to OCD, which we address in the following section.
Histamine Receptor Pharmacology and Microcircuit Mechanisms in OCD
The relevance of histamine to obsessive–compulsive disorder becomes clearer when considering how its receptor subtypes shape neural processing at the microcircuit level (Figure 2). Histamine does not exert a unitary effect on the brain; instead, it operates through multiple receptors with distinct signalling properties, cellular localisations, and functional consequences [9,29]. This receptor heterogeneity allows histamine to fine-tune excitability, plasticity, and neuromodulatory balance across cortico–striato–thalamo–cortical (CSTC) loops, providing a mechanistic substrate for the alterations in gain, flexibility, and habit formation observed in OCD.
H1 receptors are widely expressed in the cerebral cortex, particularly on pyramidal neurons and subsets of interneurons [30]. Activation of these receptors generally increases neuronal excitability through depolarising effects and modulation of intracellular calcium signalling [30]. In regions such as the orbitofrontal cortex and anterior cingulate cortex, this translates into enhanced responsiveness to incoming signals and increased salience of internally generated representations [30]. Within the context of OCD, such effects are likely to amplify error-related and threat-related signals, contributing to the subjective intensity and intrusiveness of obsessive thoughts. Importantly, this amplification does not necessarily improve behavioural accuracy or learning; rather, it may increase the persistence of neural activity patterns associated with doubt and perceived incompleteness, reinforcing the subjective need to act.
H2 receptors, which are also expressed postsynaptically in cortex and hippocampus, exert slower modulatory effects that facilitate synaptic plasticity and long-term potentiation [9,31]. Through these mechanisms, H2 receptor activation can bias learning toward consolidation rather than revision. In CSTC circuits, this property is particularly relevant to the formation and maintenance of habits (though we still lack clear evidence that those effects observed in the hippocampus and in some cortical regions can be readily transferable to other regions). While plasticity is essential for adaptive learning, excessive stabilisation of stimulus–response associations can lead to behavioural inflexibility. In OCD, where compulsions often begin as goal-directed strategies to reduce distress but gradually become habitual and insensitive to outcome, altered H2-mediated plasticity may contribute to the persistence of maladaptive routines [13].
The H3 receptor occupies a uniquely strategic position in this system. Predominantly presynaptic, H3 receptors act as autoreceptors on histaminergic terminals and as heteroreceptors on axon terminals releasing dopamine, serotonin, noradrenaline, glutamate, and GABA [9]. Through these actions, H3 receptors regulate not only histamine release itself but also the broader neuromodulatory environment within which CSTC circuits operate [9]. This property is particularly relevant to OCD, which is characterised by complex interactions between serotonergic control, dopaminergic habit learning, and noradrenergic arousal [32].
In the striatum, H3 receptors modulate dopamine release and influence the integration of cortical inputs by medium spiny neurons [23]. Subtle shifts in this balance can alter the relative dominance of direct and indirect pathways, biasing action selection toward repetition or suppression [18]. Dysregulated H3 signalling may therefore promote a state in which habitual responses are overly reinforced while inhibitory control is weakened, a core feature of compulsive behaviour. At the same time, cortical H3 receptors influence prefrontal dopamine and noradrenaline tone, affecting cognitive flexibility, working memory, and the ability to disengage from perseverative thought patterns [18].
Crucially, these receptor-specific effects do not operate in isolation. The net impact of histamine on CSTC microcircuits likely depends on the balance between receptor subtypes, regional expression patterns, and the current brain state [9]. Under conditions of heightened arousal or stress, histaminergic tone increases, potentially shifting circuits into a high-gain mode characterised by enhanced signal amplification and reduced tolerance for uncertainty [28]. If this shift is not accompanied by effective updating of internal models, the result may be a pathological amplification of error signals without resolution, driving repetitive checking or ritualised behaviour.
We also argue that this receptor-level framework might help reconcile seemingly contradictory aspects of OCD, such as the coexistence of hyperarousal and rigidity [33]. Through H1-mediated excitation, histamine can increase vigilance and attentional focus, while through H2- and H3-mediated mechanisms it can stabilise existing patterns of neural activity and neuromodulatory balance. The result is a brain state that is alert but inflexible, sensitive to perceived errors yet unable to disengage from maladaptive responses. Such a configuration aligns closely with phenomenological descriptions of OCD, in which patients report feeling constantly “on edge” while simultaneously trapped in repetitive mental and behavioural loops. This is nevertheless a speculative model deserving further exploration in experimental studies.
Understanding histamine receptor pharmacology at the microcircuit level thus provides a mechanistic bridge between diffuse neuromodulatory influences and the specific circuit dysfunctions observed in OCD. It suggests that histamine contributes not by encoding obsessional content, but by shaping the operating regime of CSTC circuits—determining how strongly signals are weighted, how easily habits are reinforced, and how resistant these patterns are to change. This sets the stage for integrating histamine into computational models of OCD, where altered learning rates, volatility estimation, and prediction error processing offer a unifying account of compulsivity and cognitive rigidity, which we address in the next section.
Histamine, Uncertainty, and Learning: A Computational Framework for OCD
The contribution of histamine to OCD becomes particularly salient when framed within contemporary computational models of learning and decision-making [34,35,36]. Increasingly, OCD is understood not simply as a disorder of abnormal beliefs or habits, but as a disorder of how the brain learns under uncertainty. Patients with OCD tend to behave as if the environment is persistently unreliable, generating repeated checks and corrective actions even in situations that are objectively stable [35]. At the same time, they show a marked resistance to updating beliefs and action policies, resulting in rigid, repetitive behaviours that persist despite negative feedback or insight [34]. This apparent contradiction—heightened sensitivity to uncertainty coexisting with inflexible learning—has proven difficult to reconcile within classical neurochemical frameworks, but aligns naturally with the known computational roles of histaminergic modulation.
In computational terms, adaptive behaviour requires balancing stability and flexibility [37,38,39]. Learning systems must estimate not only the value of actions or beliefs, but also the volatility of the environment—that is, how likely it is that the underlying rules have changed. When volatility is high, learning rates should increase, allowing rapid updating; when volatility is low, learning rates should decrease, protecting stable representations from noise [37]. In OCD, converging behavioural and modelling studies suggest a breakdown in this calibration. Patients often show exaggerated prediction error signals and inflated estimates of uncertainty, yet fail to translate these signals into effective belief updating. As a result, prediction errors are repeatedly generated but never fully resolved, sustaining a chronic sense of doubt and incompleteness [39].
Histamine is well suited to influence precisely this level of computation [7]. As a neuromodulator tightly linked to arousal, vigilance, and novelty detection, histamine dynamically adjusts the gain of neural responses to incoming information [9]. Increased histaminergic tone amplifies the impact of sensory and internal signals, effectively increasing the weight of prediction errors. Under normal conditions, this facilitates exploration and rapid adaptation to change. However, if histaminergic signalling is dysregulated—either excessive, poorly timed, or uncoupled from downstream plasticity mechanisms—the system may enter a state in which errors are amplified without being integrated into updated models of the world [8].
Within cortico–striato–thalamo–cortical loops, this imbalance can have profound consequences [6]. Cortical regions involved in monitoring performance and outcomes, such as the orbitofrontal and anterior cingulate cortices, generate strong error and conflict signals. Histamine-driven gain modulation may exaggerate these signals, increasing their salience and persistence [40]. At the same time, striatal learning mechanisms responsible for updating action values and suppressing maladaptive habits may fail to adjust appropriately, particularly if dopaminergic and serotonergic modulation is constrained by altered presynaptic control [18]. The result is a system that repeatedly signals that something is wrong, yet cannot converge on a stable corrective policy.
From this perspective, compulsions can be understood as behavioural attempts to resolve unresolved prediction errors [39,41]. Repetitive checking, washing, or ordering behaviours temporarily reduce uncertainty or error signals, providing short-lived relief. However, because the underlying learning parameters remain unchanged, the relief does not consolidate into a stable belief that the environment is safe or the action is complete. Instead, the cycle restarts, reinforcing habitual responses without resolving the computational mismatch [39]. Histamine, by sustaining a high-gain state without effective updating, may therefore contribute to both the urgency of compulsions and their resistance to extinction.
This framework also offers insight into state-dependent fluctuations in OCD symptoms. Histaminergic activity varies with sleep–wake state, circadian rhythm, stress, and environmental novelty [7,9]. Periods of sleep deprivation, heightened stress, or sensory overload—conditions known to increase histaminergic tone—are frequently reported to exacerbate OCD symptoms [42]. Sleep disturbances are common in people with OCD [43]. Computationally, these states may push the system further toward excessive gain and volatility estimation, increasing the frequency and intensity of intrusive thoughts and compulsive urges. Conversely, interventions that stabilise brain state and reduce global gain may alleviate symptoms by dampening error amplification and allowing learning mechanisms to regain balance.
Importantly, this computational view reframes histamine not as a primary driver of obsessional content, but as a regulator of how strongly and how persistently the brain treats its own errors and uncertainties. It suggests that the core dysfunction in OCD lies in the misalignment between prediction error signalling and belief updating, a process that histamine is uniquely positioned to influence. This perspective also generates clear, testable predictions: histaminergic modulation should alter learning rates, volatility estimates, and the coupling between prediction errors and behavioural updating, particularly in tasks that probe uncertainty and cognitive flexibility.
Developmental and Genetic Considerations: Histamine as a Vulnerability Factor for Compulsivity
Although much of the evidence linking histamine to obsessive–compulsive disorder (OCD) comes from adult neurobiology and computational models, developmental and genetic findings provide an important complementary perspective. OCD often emerges in childhood or adolescence, shows a fluctuating course, and overlaps substantially with tic disorders and other neurodevelopmental conditions characterised by compulsivity and repetitive behaviours [1]. This pattern suggests that vulnerability may arise from early alterations in the maturation of cortico–striato–thalamo–cortical (CSTC) circuits, processes in which neuromodulatory systems play a critical role [1].
Histamine is increasingly recognised as an important regulator of brain development [44,45]. During prenatal and early postnatal periods, histaminergic signalling influences neuronal differentiation, migration, synaptogenesis, and the maturation of excitatory–inhibitory balance [45]. Histamine receptors are expressed early across cortical and subcortical regions, including the striatum and thalamus, and modulate synaptic refinement during sensitive developmental windows. Disruption of histaminergic tone during these periods could therefore bias CSTC circuits toward reduced flexibility and increased rigidity, conferring long-term vulnerability to compulsive behavioural patterns [22,45].
Genetic evidence, while still limited, is consistent with this view. Rare loss-of-function mutations in histidine decarboxylase (HDC)—the enzyme responsible for histamine synthesis—have been robustly associated with Tourette syndrome, a condition that frequently co-occurs with OCD and shares overlapping CSTC circuitry [46,47,48]. Rather than pointing to histamine as a disorder-specific mechanism, these findings suggest that altered histaminergic signalling confers a broader liability to compulsivity-related phenotypes, with clinical expression shaped by developmental timing and interactions with other neuromodulatory systems.
Beyond rare variants, common genetic variation affecting histamine receptors, transport, and metabolism may contribute to individual differences in cognitive flexibility, habit learning, and sensitivity to uncertainty [33]. Such variation is unlikely to cause OCD in isolation, but may act as a risk modifier, biasing CSTC circuits toward maladaptive operating regimes when combined with serotonergic, dopaminergic, or stress-related vulnerabilities.
A developmental framework also helps explain heterogeneity in OCD. Early-onset forms are more often associated with tics and neurodevelopmental traits, whereas later-onset OCD shows stronger links to stress and affective symptoms [33]. Histaminergic dysregulation may contribute to both, but through different mechanisms: altered signalling during development may bias circuit maturation toward rigidity, whereas state-dependent perturbations of histamine in adulthood—related to stress, sleep disruption, or circadian dysregulation—may destabilise already vulnerable circuits [22].
Together, developmental and genetic evidence supports the view that histamine is not merely a downstream modulator of OCD symptoms, but part of the vulnerability architecture shaping how CSTC circuits develop and respond to challenge. By influencing synaptic maturation, neuromodulatory balance, and learning under uncertainty, histaminergic signalling may predispose individuals to enduring patterns of compulsivity that later manifest as OCD.
Alternative Neuromodulatory Accounts and Why Histamine Might Have Been Overlooked
Although the present review argues for a central role of histaminergic modulation in shaping cortico–striato–thalamo–cortical (CSTC) dynamics in obsessive–compulsive disorder (OCD), histamine has remained comparatively peripheral in dominant neurobiological models of the disorder. Several factors likely account for this relative neglect. First, histamine lacks a clear clinical foothold in OCD treatment. In contrast to serotonin and dopamine—whose relevance has been reinforced by the efficacy of selective serotonin reuptake inhibitors and antipsychotic augmentation—there are no established OCD treatments that directly target histamine receptors. This absence of a translational feedback loop has likely limited sustained experimental attention to histaminergic mechanisms in OCD.
Second, histamine primarily regulates global brain state variables such as arousal, vigilance, and neural gain, rather than encoding specific cognitive or affective content [9]. Historically, OCD research has focused on neurotransmitter systems that could be more directly linked to obsessions, anxiety, or compulsive actions themselves [33]. As a result, neuromodulatory systems whose effects are diffuse, context-dependent, and difficult to capture using standard task-based paradigms may have been underappreciated, despite their potential importance for shaping circuit operating regimes [12].
Third, methodological limitations have constrained the in vivo study of histaminergic signalling in humans. Compared with dopaminergic or serotonergic systems, there are relatively few positron emission tomography ligands available for histamine receptors, and large-scale neuroimaging studies of OCD have rarely incorporated direct measures of histaminergic function. Consequently, histamine has often been absent from data-driven circuit models, reinforcing its marginal position in the literature [49].
It is also important to recognise that histamine is not the only neuromodulatory system capable of influencing neural gain, uncertainty processing, and learning flexibility. The noradrenergic system, originating in the locus coeruleus, plays a well-established role in arousal and adaptive gain modulation, and heightened noradrenergic tone has been linked to vigilance and salience processing [50]. From this perspective, noradrenergic dysregulation could plausibly contribute to exaggerated error or threat signals in OCD. However, noradrenaline is most strongly associated with phasic responses to salient or unexpected external events, whereas OCD is characterised by persistent, internally generated signals of doubt and incompleteness [51]. This distinction suggests that noradrenergic mechanisms alone may not fully account for the chronicity and rigidity of OCD symptoms.
Similarly, the cholinergic system has been implicated in attentional control, learning, and habit formation, particularly within cortico-striatal circuits [52,53]. Cholinergic modulation can influence the balance between goal-directed and—habitual responding and may therefore contribute to compulsive behaviour. Nevertheless, cholinergic effects tend to be more spatially and functionally specific, shaping processing within defined circuits rather than imposing a global brain state.
Within this broader neuromodulatory landscape, histamine may occupy a complementary role. Rather than acting as a primary driver of specific signals, histaminergic modulation may set the tonic background conditions—such as arousal level, gain, and responsiveness—under which other neuromodulatory systems operate [54]. From this perspective, gain dysregulation in OCD is unlikely to reflect dysfunction in a single neuromodulatory system. Instead, it may arise from maladaptive interactions among multiple systems, with histamine contributing to the overall operating regime that determines how strongly errors are amplified, how persistent they become, and how readily learning and behavioural updating can occur.
Translational and Clinical Implications: Histamine as a Candidate Target for Circuit-Informed Investigation
Framing histamine as a potential regulator of neural gain and learning flexibility in obsessive–compulsive disorder (OCD) generates testable hypotheses rather than an established treatment strategy. If histaminergic signalling contributes to the operating conditions of cortico–striato–thalamo–cortical (CSTC) circuits, experimental modulation of this system may help probe how changes in neuromodulatory tone influence learning, flexibility, and error processing in OCD.
Among histamine receptors, the H3 receptor represents a mechanistically tractable experimental target. As a presynaptic auto- and heteroreceptor, H3 regulates histamine release as well as dopaminergic, serotonergic, and noradrenergic transmission [23,32,55]. Pharmacological manipulation of H3 receptors therefore offers a controlled means of perturbing neuromodulatory balance, although whether this meaningfully alters CSTC dynamics or behavioural flexibility in OCD remains an open empirical question. In preclinical models, selective activation of presynaptic H3 heteroreceptors on glutamatergic afferents to the nucleus accumbens core reduced glutamatergic transmission and ameliorated both anxiety- and obsessive-compulsive-like behaviours induced by stress, demonstrating that H3 receptor signalling can influence compulsive phenotypes via modulation of specific glutamatergic circuits [11]. However, the study is based on stress-induced behavioural phenotypes in rodents, which capture aspects of compulsive-like behaviour but only partially model obsessive–compulsive disorder, particularly with respect to insight, cognitive rigidity, and uncertainty processing. In addition, the manipulation targets a specific glutamatergic circuit within the nucleus accumbens rather than distributed cortico–striato–thalamo–cortical loops, and therefore does not provide evidence for global circuit-level modulation. Finally, the reported effects were obtained through activation of H3 receptors, whereas much of the proposed translational interest in OCD centres on H3 antagonism or inverse agonism, highlighting a mismatch in pharmacological directionality that complicates direct extrapolation.
Pitolisant, an H3 inverse agonist approved for narcolepsy, should be viewed as a tool for experimental medicine studies, not as a candidate treatment for OCD [56,57]. Its potential relevance lies in enabling mechanistic tests of how transient changes in arousal and neuromodulatory tone affect cognitive flexibility, perseveration, and learning under uncertainty.
More broadly, histaminergic modulation is unlikely to act as a stand-alone intervention. If relevant, it may be most informative when used to interrogate or facilitate learning processes engaged by psychological therapies. Neuroimaging and computational approaches, particularly pharmacological challenge paradigms combined with tasks probing learning and uncertainty, will be essential to determine whether histamine plays a meaningful role in OCD-relevant circuit dynamics.
Outstanding Questions for Future Research
Despite growing theoretical and translational interest, the role of the brain histamine system in obsessive–compulsive disorder remains incompletely characterised. One of the most fundamental open questions concerns the direction and nature of histaminergic dysregulation in OCD. It is still unclear whether OCD is associated with chronically elevated histaminergic tone, deficient signalling, or—more plausibly—a state-dependent dysregulation in which histamine release is exaggerated in some contexts (e.g., stress, uncertainty, sleep deprivation) but insufficiently coupled to downstream plasticity and learning mechanisms. Resolving this issue will require multimodal approaches integrating postmortem molecular data, in vivo neuroimaging, and physiological markers of arousal and brain state.
A second unresolved question relates to regional and receptor-specific effects. Histamine receptors are heterogeneously distributed across cortex, striatum, and thalamus, and their functional roles differ markedly across microcircuits. It remains unknown whether OCD is characterised by a global shift in histaminergic modulation or by regionally specific alterations, for example preferential effects in orbitofrontal–striatal loops versus motor or limbic circuits. Disentangling H1-, H2-, and H3-mediated effects in humans, particularly within CSTC networks, represents a major challenge but is essential for refining mechanistic models and therapeutic targets.
From a computational perspective, an important open question is how histamine shapes learning parameters in OCD. While theoretical accounts suggest that histamine modulates neural gain, learning rates, and volatility estimation, direct empirical evidence in patients is sparse. It remains to be established whether histaminergic modulation selectively alters the coupling between prediction errors and belief updating, whether it affects habit formation versus goal-directed control, or whether its primary role lies in regulating the persistence and salience of error signals. Carefully designed behavioural tasks combined with computational modelling and pharmacological manipulation will be critical to address these issues.
Developmental timing represents another key gap in knowledge. Although genetic and neurodevelopmental data suggest that histamine may act as a vulnerability factor for compulsivity, it is not yet clear when in development histaminergic alterations exert their greatest impact. Determining whether histamine-related effects primarily influence early circuit maturation, adolescent refinement of CSTC loops, or later stress sensitivity has important implications for prevention and early intervention. Longitudinal studies spanning childhood to adulthood, ideally integrating sleep, circadian, and neuromodulatory measures, are needed to clarify these trajectories.
At the clinical level, a major unanswered question concerns patient stratification. OCD is a heterogeneous disorder, and it is unlikely that histaminergic dysfunction contributes equally across all symptom dimensions and clinical subtypes. Whether histamine-related mechanisms are more relevant to checking and doubt, symmetry and ordering, or tic-related compulsivity remains unknown. Identifying cognitive, behavioural, or neurobiological markers that index histaminergic involvement could enable more targeted application of histamine-based interventions and avoid non-specific treatment approaches.
Finally, there are important translational questions regarding therapeutic implementation and integration. It remains to be determined whether histaminergic agents are best deployed as monotherapies, adjuncts to serotonergic treatment, or facilitators of psychological interventions such as exposure and response prevention. Equally important is understanding the temporal dynamics of their effects: whether benefits arise acutely through state modulation or require longer-term changes in learning and circuit plasticity. Addressing these questions will require experimental medicine studies that move beyond symptom reduction to focus on mechanism-specific endpoints.
Taken together, these outstanding questions highlight both the promise and the current limitations of a histaminergic framework for OCD. Addressing them will require close integration of molecular neuroscience, systems and computational approaches, and carefully designed clinical studies. Such efforts have the potential not only to clarify the role of histamine in OCD, but also to advance a broader understanding of how neuromodulatory systems regulate flexibility, uncertainty, and compulsivity in the human brain.
Conclusions
Obsessive–compulsive disorder has classically been conceptualised in terms of cortico–striato–thalamo–cortical dysfunction and serotonergic imbalance. While influential, these models leave key features insufficiently explained, including the coexistence of heightened vigilance with cognitive rigidity, persistent error signals despite insight, and resistance to behavioural updating. In this review, we argue that the brain histamine system provides a unifying and underexplored framework for these phenomena.
Through its diffuse projections, receptor heterogeneity, and presynaptic regulation of multiple neuromodulatory systems, histamine acts as a meta-modulator of neural gain, arousal, and learning flexibility. Histaminergic dysregulation may therefore bias CSTC circuits toward a high-gain, low-flexibility regime, amplifying uncertainty and entrenching habitual responding without specifying the content of obsessions or compulsions. However, this remains as a computational hypothesis.
This perspective reframes compulsivity as a maladaptive response to unresolved uncertainty within inflexible circuits, with important translational implications. Targeting histaminergic signalling—particularly via H3 receptors—offers a putative mechanistically grounded avenue to address cognitive rigidity and perseveration, dimensions often poorly targeted by existing treatments. More broadly, the histamine framework underscores the importance of neuromodulatory control of brain state in psychiatric disorders, aligning OCD with emerging systems-level and computational models of psychopathology.
References
- Stein, D.J. Obsessive-compulsive disorder. Nat Rev Dis Primers 2019, 5(1), p. 52. [Google Scholar] [CrossRef]
- Calza, J. Altered Cortico-Striatal Functional Connectivity During Resting State in Obsessive-Compulsive Disorder. Front Psychiatry 2019, 10, 319. [Google Scholar] [CrossRef]
- Cotovio, G. Obsessive-compulsive disorder secondary to focal brain lesions: from lesions to networks. medRxiv 2025. [Google Scholar] [CrossRef]
- Baumgarten, H.G.; Grozdanovic, Z. Role of serotonin in obsessive-compulsive disorder. Br J Psychiatry Suppl 1998, 35, 13–20. [Google Scholar] [CrossRef]
- Mota, P.; Pico-Perez, M.; Morgado, P. Mapping dopamine’s role in obsessive-compulsive disorder: A scoping review of neural circuits, brain regions, and behavioral implications. J Psychiatr Res 2025, 194, 11–20. [Google Scholar] [CrossRef]
- Haas, H.; Panula, P. The role of histamine and the tuberomamillary nucleus in the nervous system. Nat Rev Neurosci 2003, 4(2), 121–30. [Google Scholar] [CrossRef]
- Green, M. Histamine in the central nervous system. Proc West Pharmacol Soc 1978, 21, 337–9. [Google Scholar] [PubMed]
- Baronio, D. Histaminergic system in brain disorders: lessons from the translational approach and future perspectives. Ann Gen Psychiatry 2014, 13(1), p. 34. [Google Scholar] [CrossRef]
- Brown, R.E.; Stevens, D.R.; Haas, H.L. The physiology of brain histamine. Prog Neurobiol 2001, 63(6), 637–72. [Google Scholar] [CrossRef] [PubMed]
- Bruckner, R.; Heekeren, H.R.; Nassar, M.R. Understanding learning through uncertainty and bias. Commun Psychol 2025, 3(1), p. 24. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y. Targeting presynaptic H3 heteroreceptor in nucleus accumbens to improve anxiety and obsessive-compulsive-like behaviors. Proc Natl Acad Sci U S A 2020, 117(50), 32155–32164. [Google Scholar] [CrossRef]
- Panula, P.; Nuutinen, S. The histaminergic network in the brain: basic organization and role in disease. Nat Rev Neurosci 2013, 14(7), 472–87. [Google Scholar] [CrossRef]
- Yin, H.H.; Knowlton, B.J. The role of the basal ganglia in habit formation. Nat Rev Neurosci 2006, 7(6), 464–76. [Google Scholar] [CrossRef]
- Gittis, A.H.; Kreitzer, A.C. Striatal microcircuitry and movement disorders. Trends Neurosci 2012, 35(9), 557–64. [Google Scholar] [CrossRef]
- Airaksinen, M.S.; Panula, P. The histaminergic system in the guinea pig central nervous system: an immunocytochemical mapping study using an antiserum against histamine. J Comp Neurol 1988, 273(2), 163–86. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M. The representation of decision variables in orbitofrontal cortex is longitudinally stable. Cell Rep 2024, 43(10), p. 114772. [Google Scholar] [CrossRef]
- Carter, C.S. Anterior cingulate cortex, error detection, and the online monitoring of performance. Science 1998, 280(5364), 747–9. [Google Scholar] [CrossRef] [PubMed]
- Bolam, J.P.; Ellender, T.J. Histamine and the striatum. Neuropharmacology 2016, 106, 74–84. [Google Scholar] [CrossRef]
- Wall, N.R. Differential innervation of direct- and indirect-pathway striatal projection neurons. Neuron 2013, 79(2), 347–60. [Google Scholar] [CrossRef]
- Peng, J.Y. Receptor and Ionic Mechanism of Histamine on Mouse Dorsolateral Striatal Neurons. Mol Neurobiol 2023, 60(1), 183–202. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.X. Histamine Modulation of the Basal Ganglia Circuitry in the Motor Symptoms of Parkinson’s Disease. CNS Neurosci Ther 2025, 31(2), p. e70308. [Google Scholar] [CrossRef]
- Han, S. Histaminergic Control of Corticostriatal Synaptic Plasticity during Early Postnatal Development. J Neurosci 2020, 40(34), 6557–6571. [Google Scholar] [CrossRef] [PubMed]
- Ellenbroek, B.A. Histamine H(3) receptors, the complex interaction with dopamine and its implications for addiction. Br J Pharmacol 2013, 170(1), 46–57. [Google Scholar] [CrossRef]
- Lee, K.H. Histamine modulates thalamocortical activity by activating a chloride conductance in ferret perigeniculate neurons. Proc Natl Acad Sci U S A 2004, 101(17), 6716–21. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.Y.; Kalimo, H.; Panula, P. The histaminergic system in human thalamus: correlation of innervation to receptor expression. Eur J Neurosci 2002, 15(7), 1125–38. [Google Scholar] [CrossRef]
- Weeland, C.J. The thalamus and its subnuclei-a gateway to obsessive-compulsive disorder. Transl Psychiatry 2022, 12(1), p. 70. [Google Scholar] [CrossRef] [PubMed]
- Tasaka, K. Excitatory effect of histamine on the arousal system and its inhibition by H1 blockers. Brain Res Bull 1989, 22(2), 271–5. [Google Scholar] [CrossRef]
- Thakkar, M.M. Histamine in the regulation of wakefulness. Sleep Med Rev 2011, 15(1), 65–74. [Google Scholar] [CrossRef]
- Thangam, E.B. The Role of Histamine and Histamine Receptors in Mast Cell-Mediated Allergy and Inflammation: The Hunt for New Therapeutic Targets. Front Immunol 2018, 9, 1873. [Google Scholar] [CrossRef]
- Tashiro, M. Roles of histamine in regulation of arousal and cognition: functional neuroimaging of histamine H1 receptors in human brain. Life Sci 2002, 72(4-5), 409–14. [Google Scholar] [CrossRef]
- Luo, T.; Leung, L.S. Endogenous histamine facilitates long-term potentiation in the hippocampus during walking. J Neurosci 2010, 30(23), 7845–52. [Google Scholar] [CrossRef]
- Schlicker, E. Modulation of neurotransmitter release via histamine H3 heteroreceptors. Fundam Clin Pharmacol 1994, 8(2), 128–37. [Google Scholar] [CrossRef] [PubMed]
- Pauls, D.L. Obsessive-compulsive disorder: an integrative genetic and neurobiological perspective. Nat Rev Neurosci 2014, 15(6), 410–24. [Google Scholar] [CrossRef] [PubMed]
- Gillan, C.M. Functional neuroimaging of avoidance habits in obsessive-compulsive disorder. Am J Psychiatry 2015, 172(3), 284–93. [Google Scholar] [CrossRef]
- Voon, V. Disorders of compulsivity: a common bias towards learning habits. Mol Psychiatry 2015, 20(3), 345–52. [Google Scholar] [CrossRef]
- Vaghi, M.M. Action-Outcome Knowledge Dissociates From Behavior in Obsessive-Compulsive Disorder Following Contingency Degradation. Biol Psychiatry Cogn Neurosci Neuroimaging 2019, 4(2), 200–209. [Google Scholar] [CrossRef] [PubMed]
- Behrens, T.E. Learning the value of information in an uncertain world. Nat Neurosci 2007, 10(9), 1214–21. [Google Scholar] [CrossRef]
- Mathys, C.D. Uncertainty in perception and the Hierarchical Gaussian Filter. Front Hum Neurosci 2014, 8, 825. [Google Scholar] [CrossRef]
- Fradkin, I. Doubting what you already know: Uncertainty regarding state transitions is associated with obsessive compulsive symptoms. PLoS Comput Biol 2020, 16(2), e1007634. [Google Scholar] [CrossRef]
- Scammell, T.E. Histamine: neural circuits and new medications. Sleep 2019, 42(1). [Google Scholar] [CrossRef]
- Szechtman, H.; Woody, E. Obsessive-compulsive disorder as a disturbance of security motivation. Psychol Rev 2004, 111(1), 111–27. [Google Scholar] [CrossRef]
- Porkka-Heiskanen, T. The effect of REM sleep deprivation on histamine concentrations in different brain areas. Life Sci 1994, 54(22), 1719–26. [Google Scholar] [CrossRef]
- Segalas, C. Sleep disturbances in obsessive-compulsive disorder: influence of depression symptoms and trait anxiety. BMC Psychiatry 2021, 21(1), 42. [Google Scholar] [CrossRef] [PubMed]
- Panula, P.; Sundvik, M.; Karlstedt, K. Developmental roles of brain histamine. Trends Neurosci 2014, 37(3), 159–68. [Google Scholar] [CrossRef] [PubMed]
- Carthy, E.; Ellender, T. Histamine, Neuroinflammation and Neurodevelopment: A Review. Front Neurosci 2021, 15, 680214. [Google Scholar] [CrossRef]
- Ercan-Sencicek, A.G. L-histidine decarboxylase and Tourette’s syndrome. N Engl J Med 2010, 362(20), 1901–8. [Google Scholar] [CrossRef]
- Baldan, L.C. Histidine decarboxylase deficiency causes tourette syndrome: parallel findings in humans and mice. Neuron 2014, 81(1), 77–90. [Google Scholar] [CrossRef]
- Jindachomthong, K. White matter abnormalities in the Hdc knockout mouse, a model of tic and OCD pathophysiology. Front Mol Neurosci 2022, 15, 1037481. [Google Scholar] [CrossRef]
- Yanai, K. Histamine H1 receptors in human brain visualized in vivo by [11C]doxepin and positron emission tomography. Neurosci Lett 1992, 137(2), 145–8. [Google Scholar] [CrossRef]
- Sara, S.J.; Bouret, S. Orienting and reorienting: the locus coeruleus mediates cognition through arousal. Neuron 2012, 76(1), 130–41. [Google Scholar] [CrossRef]
- Howells, F.M.; Stein, D.J.; Russell, V.A. Synergistic tonic and phasic activity of the locus coeruleus norepinephrine (LC-NE) arousal system is required for optimal attentional performance. Metab Brain Dis 2012, 27(3), 267–74. [Google Scholar] [CrossRef]
- Sarter, M. Unraveling the attentional functions of cortical cholinergic inputs: interactions between signal-driven and cognitive modulation of signal detection. Brain Res Brain Res Rev 2005, 48(1), 98–111. [Google Scholar] [CrossRef]
- Huang, Z. Dynamic responses of striatal cholinergic interneurons control behavioral flexibility. Sci Adv 2024, 10(51), p. eadn2446. [Google Scholar] [CrossRef]
- Marder, E. Neuromodulation of neuronal circuits: back to the future. Neuron 2012, 76(1), 1–11. [Google Scholar] [CrossRef]
- Esbenshade, T.A. The histamine H3 receptor: an attractive target for the treatment of cognitive disorders. Br J Pharmacol 2008, 154(6), 1166–81. [Google Scholar] [CrossRef]
- Zou, Y. Pitolisant alleviates brain network dysfunction and cognitive deficits in a mouse model of Alzheimer’s disease. Transl Psychiatry 2025, 15(1), p. 126. [Google Scholar] [CrossRef]
- Li, S.; Yang, J. Pitolisant for treating patients with narcolepsy. Expert Rev Clin Pharmacol 2020, 13(2), 79–84. [Google Scholar] [CrossRef]
Figure 1.
Diffuse histaminergic modulation of cortico–striato–thalamo–cortical (CSTC) circuits. Left: Schematic of the brain histamine system, illustrating widespread projections arising from the tuberomammillary nucleus of the posterior hypothalamus and innervating cortex, basal ganglia, thalamus, brainstem, cerebellum, and spinal cord. This architecture highlights histamine’s role as a global neuromodulator rather than a point-to-point transmitter. Right: Core CSTC loops implicated in obsessive–compulsive disorder (OCD), including orbitofrontal, prefrontal, and cingulate cortices, striatum, thalamus, and midbrain. Through its diffuse innervation and receptor-specific actions across these nodes, histamine is positioned to coordinate cortical monitoring, striatal gating, and thalamic relay fidelity, biasing the overall operating regime of CSTC circuits and influencing action selection and behavioural flexibility.
Figure 1.
Diffuse histaminergic modulation of cortico–striato–thalamo–cortical (CSTC) circuits. Left: Schematic of the brain histamine system, illustrating widespread projections arising from the tuberomammillary nucleus of the posterior hypothalamus and innervating cortex, basal ganglia, thalamus, brainstem, cerebellum, and spinal cord. This architecture highlights histamine’s role as a global neuromodulator rather than a point-to-point transmitter. Right: Core CSTC loops implicated in obsessive–compulsive disorder (OCD), including orbitofrontal, prefrontal, and cingulate cortices, striatum, thalamus, and midbrain. Through its diffuse innervation and receptor-specific actions across these nodes, histamine is positioned to coordinate cortical monitoring, striatal gating, and thalamic relay fidelity, biasing the overall operating regime of CSTC circuits and influencing action selection and behavioural flexibility.
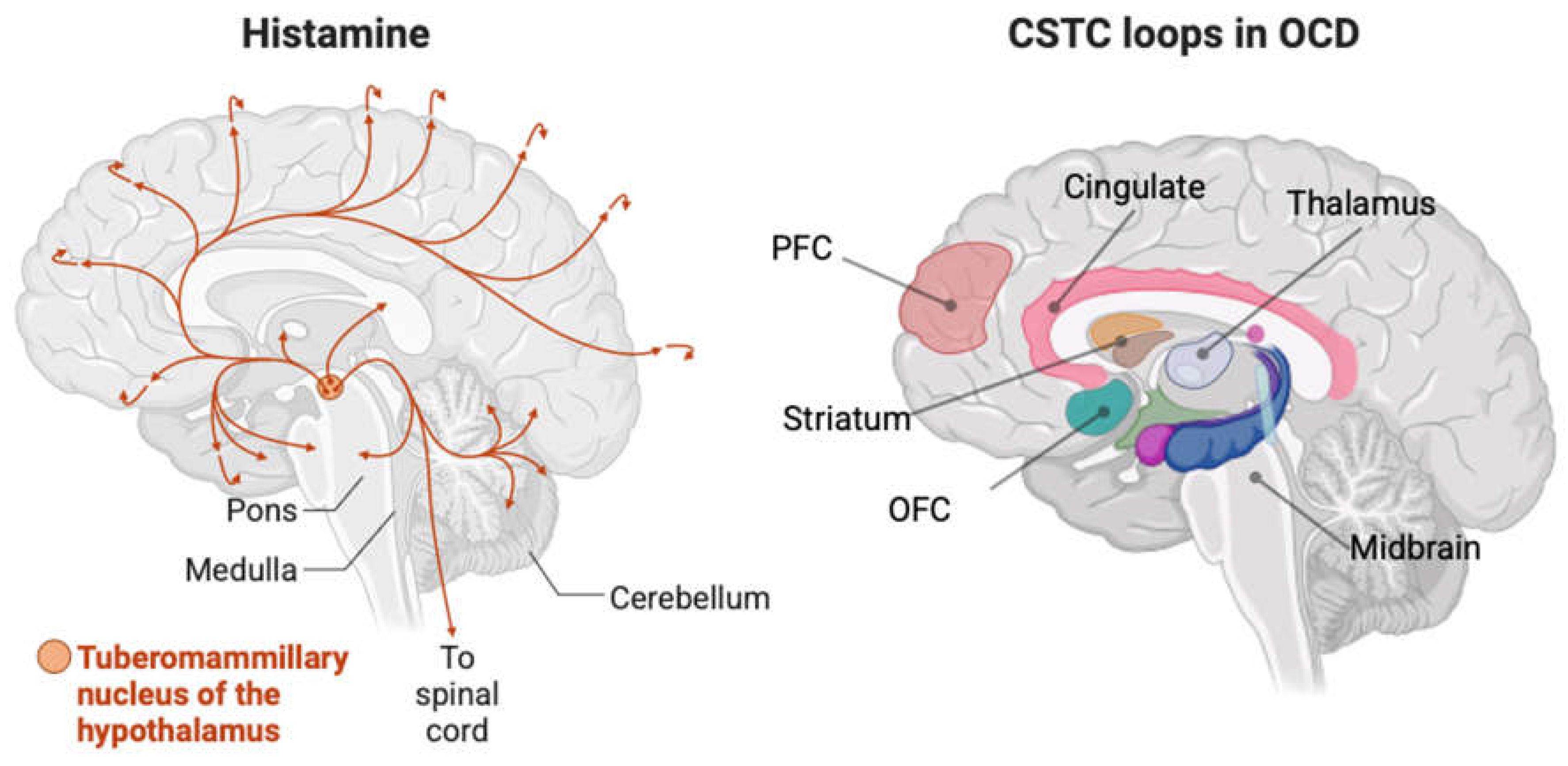
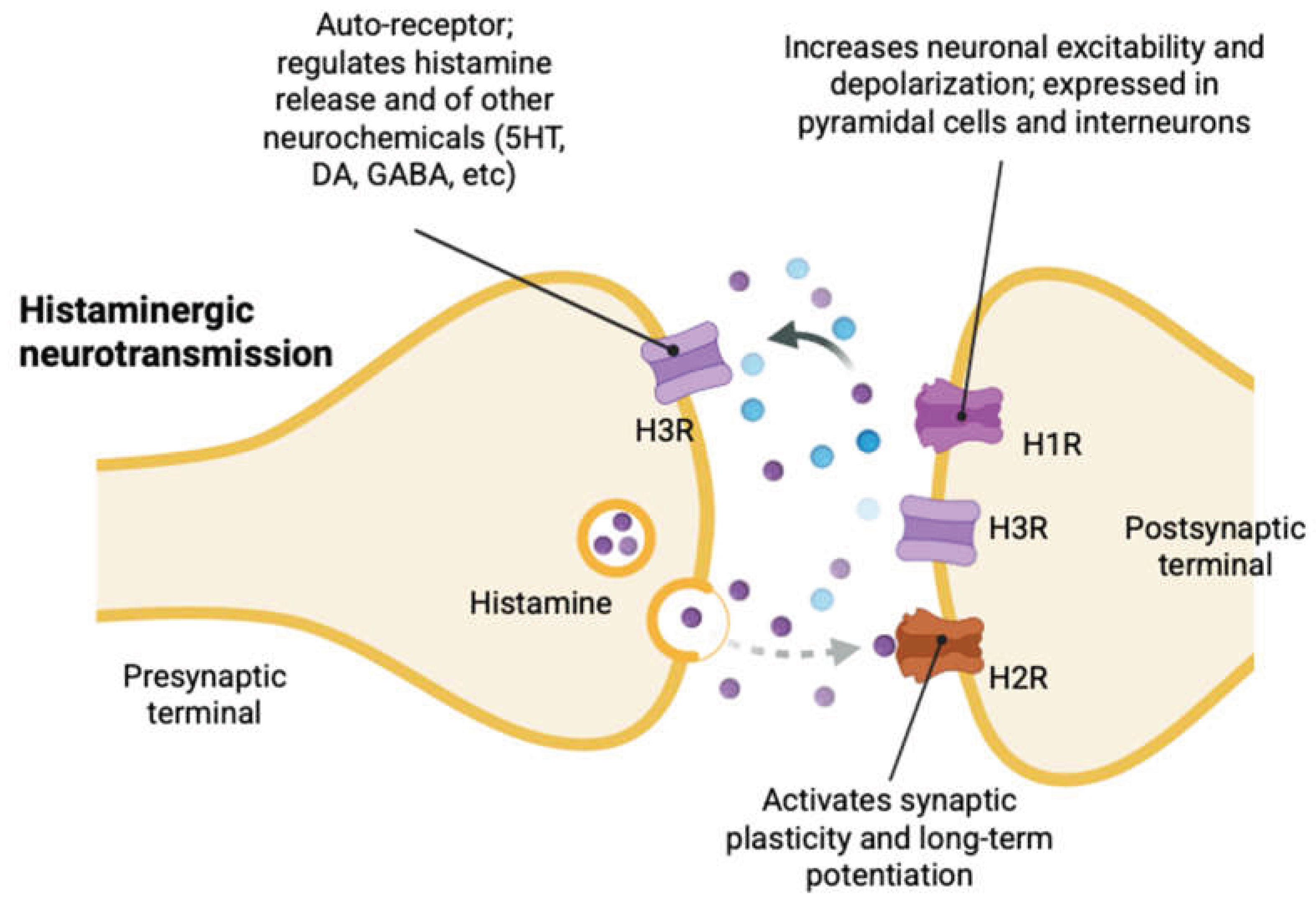
Figure 2.
Receptor-specific mechanisms of histaminergic neurotransmission. Schematic representation of histamine signalling at the synapse, highlighting distinct pre- and postsynaptic actions mediated by histamine receptor subtypes. Presynaptic H3 receptors act as autoreceptors that constrain histamine synthesis and release and heteroreceptors that regulate the release of other neuromodulators, including serotonin, dopamine, GABA, and acetylcholine. Postsynaptically, H1 receptors increase neuronal excitability and depolarisation in both pyramidal neurons and interneurons, while H2 receptors facilitate synaptic plasticity and long-term potentiation through second-messenger signalling in the hippocampus and in some cortical contexts. Together, these receptor-specific actions allow histamine to modulate local excitation–inhibition balance and synaptic gain, linking global arousal signals to circuit-level computation and behavioural flexibility.
Figure 2.
Receptor-specific mechanisms of histaminergic neurotransmission. Schematic representation of histamine signalling at the synapse, highlighting distinct pre- and postsynaptic actions mediated by histamine receptor subtypes. Presynaptic H3 receptors act as autoreceptors that constrain histamine synthesis and release and heteroreceptors that regulate the release of other neuromodulators, including serotonin, dopamine, GABA, and acetylcholine. Postsynaptically, H1 receptors increase neuronal excitability and depolarisation in both pyramidal neurons and interneurons, while H2 receptors facilitate synaptic plasticity and long-term potentiation through second-messenger signalling in the hippocampus and in some cortical contexts. Together, these receptor-specific actions allow histamine to modulate local excitation–inhibition balance and synaptic gain, linking global arousal signals to circuit-level computation and behavioural flexibility.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



