Submitted:
03 February 2026
Posted:
05 February 2026
You are already at the latest version
Abstract
The integration of 3D bioprinting technology and CRISPR-Cas9 genome editing has become a game-changing method for creating complex organotypic cancer models. This integrated platform overcomes the drawbacks of traditional 2D culture systems by enabling precise genetic modifications within physiologically relevant, biomimetic tumor microenvironments. Researchers can more precisely recreate tumor progression, oncogenic mutations, cellular heterogeneity, and drug resistance mechanisms by utilizing the structural complexity provided by 3D bioprinting and the specificity of CRISPR-Cas9-mediated gene editing. CRISPR-Cas9 enables specific gene modifications, including oncogene knockout (e.g., MYC, KRAS) or immune checkpoint genes (e.g., PD-1, PD-L1), in 3D-bioprinted structures made from tumorigenic or patient-specific cell populations. It has been demonstrated that these modified models maintain important histopathological and molecular characteristics of original tumors, allowing for accurate high-throughput screening of immunotherapeutics and anticancer drugs. Significantly speeding up the modeling of tumorigenesis, studies using prostate cancer organoids showed gene correction efficiencies ranging from 50 to 90 %. Additionally, in 3D cultures, combinatorial CRISPR-Cas9 editing has demonstrated synergistic drug responses in models of lung and breast cancer, underscoring the platform's potential for discovering new therapeutic targets. Biomaterial-based vectors, like hydrogels and nanocarriers, are being improved to reduce off-target effects and increase intracellular uptake to increase the accuracy and safety of CRISPR delivery. However, issues with scalability, reproducibility, and standardization still exist, requiring ongoing interdisciplinary cooperation to improve downstream validation procedures, gene-editing tactics, and bioink formulations. The potential of CRISPR-Cas9-integrated 3D bioprinting as a state-of-the-art technique for drug discovery and cancer modeling is highlighted in this review. It emphasizes how the platform can speed up translational research in oncology, lessen dependency on animal models, and customize treatment plans. The goal of this review is to present a thorough summary of current developments, technical difficulties, and potential paths forward in this quickly developing field.
Keywords:
CRISPR-Cas9
; 3D bioprinting
; cancer modeling
; tumor microenvironment
; gene editing
; personalized medicine
1. Introduction
The current state of cancer modeling is still plagued by serious issues that prevent it from accurately simulating human disease, which impedes the development of new treatments. The basic genomic and physiological differences between species make it difficult for traditional animal models, especially rodent xenografts and genetically modified mice, to forecast the results of clinical trials [1,2]. For example, when implanted in immunocompromised mice, patient-derived xenografts (PDXs) do not accurately replicate human immune responses or stromal interactions, which reduces their translational value. Similar to advanced human tumors, conventional in vitro systems, which range from simple spheroids to two-dimensional (2D) monolayers, lack the microenvironmental complexity and hierarchical cellular heterogeneity. By removing the "holistic" biology of tumors, highly reductionist methods contribute to Eroom's Law, which states that R&D productivity declines. These restrictions are made worse by technical obstacles in primary human tissue models, such as difficulties maintaining clinical metadata, guaranteeing reproducibility, and simulating metastatic variants like MDA-MB-231-LM2-4 [3,4]. The urgent need for next-generation models that more closely reflect the biology of human tumors is highlighted by these gaps taken together [5,6,7].
A more systemic paradigm has replaced the purely genetic one as the key factors influencing cancer progression, metastasis, and treatment resistance. These factors include intratumoral heterogeneity and the tumor microenvironment (TME). A dynamic milieu of stromal cells, such as cancer-associated fibroblasts (CAFs), immune cells (e.g. 3. networks of endothelial and pericyte cells—encased in a dynamic extracellular matrix (ECM), TAMs, and T-cells. The chemical gradients caused by this complex ecosystem—hypoxia, acidity, and nutrient depletion—as well as biophysical stressors like interstitial pressure, all have an impact on cellular phenotypes and treatment outcomes [8,9,10,11]. For instance, CAFs promote angiogenesis and resist chemotherapeutics by co-ordinating ECM remodeling and secreting immunosuppressive cytokines like TGF-β. TAMs and other immune cells further strengthen resistance by forming immunosuppressive niches. However, intratumoral heterogeneity—which includes temporal and spatial variations in genetic, epigenetic, phenotypic, and metabolic profiles—causes treatment failure by producing resistant subclones that avoid therapy. These intricate ecosystems are being broken down by recent developments in single-cell and spatial genomics, which have shown that the TME and tumor-intrinsic characteristics are crucial to disease outcome and heterogeneity [12,13,14]. It is now necessary—not optional—to use models that incorporate cellular diversity and microenvironmental complexity when creating effective treatments.
3D bioprinting and CRISPR-Cas9 are two emerging technologies that have the potential to bridge the gap between the complex reality of human tumors and simplistic models. Modeling aging-related mutations, engineering advanced CAR-T cell therapies, and precisely manipulating cancer-relevant genes are all made possible by CRISPR-Cas9. These genome-editing tools are being used to create mouse models and organoids that include clinically relevant mutations and enable in situ gene function interrogation, improving preclinical accuracy and mechanistic insight [15,16,17]. On the other hand, 3D bioprinting, which involves layer-by-layer deposition of cells and bioinks (supporting cells, hydrogels, and extracellular matrix components), makes it possible to create intricate, repeatable, and physiologically significant structures that more closely resemble in vivo TME characteristics than conventional spheroids. 3D bioprinted constructs can replicate metabolic gradients, metastatic niches, cell-matrix and cell-cell interactions, and reproducible heterogeneity, according to recent reviews [18,19,20,21].
By integrating CRISPR precision and bioprinting realism, researchers can generate multi-cellular constructs that maintain spatial heterogeneity and microenvironmental context, as compared with traditional models. CRISPR allows cancer-specific mutations to be inserted into organoids or iPSCs, and bioprinting puts these genetically defined cells together with immune cells, fibroblasts, and endothelial cells in spatially ordered configurations. These structures allow for the dynamic examination of CAF-mediated drug resistance, ECM remodeling, hypoxia-induced metabolic changes, and immune evasion in a realistic yet controlled TME [22,23,24]. These systems, when paired with microfluidic "tumor-on-chip" platforms, provide scalable, high-throughput models that record continuous biomarker readouts and physiologic flows. Together with single-cell and spatial multi-omics, they enable high-resolution tracking of tumor evolution and the emergence of therapy resistance.
In conclusion, the integration of emerging new technologies into cancer modeling represents a paradigm change. The field of cancer modeling is changing dramatically, moving away from low-fidelity, reductionist systems and toward more integrated, human-relevant platforms. Precise genetic manipulation is now possible thanks to the development of CRISPR-Cas9 technology, and 3D bioprinting makes it easier to reconstruct the intricate microenvironment and tumor architecture. When combined with sophisticated analytical techniques, these technologies' convergence holds great promise for producing models that faithfully capture tumor evolution, heterogeneity, and therapeutic response. In this review, we discuss the integration of these technologies—CRISPR-Cas9 and 3D bioprinting—and their potential to synergistically enhance the fidelity of cancer models. The integration of these approaches, supported by advanced analytical methodologies, holds considerable promise for generating systems that accurately recapitulate tumor evolution, heterogeneity, and therapeutic response. Despite persistent technical and logistical challenges—including issues of reproducibility, scalability, and cost—these innovations represent critical advancements in preclinical cancer research. They offer the potential to develop models that more faithfully replicate human tumor biology and to streamline translational efforts from laboratory discovery to clinical application. Sustained progress in this domain will require interdisciplinary collaboration, methodological standardization, and rigorous validation across both preclinical and clinical settings.
2. CRISPR-Cas9 Technology in Cancer Research
Cancer research has been greatly impacted by the groundbreaking genome-editing technique known as CRISPR-Cas9 (Clustered Regularly Interspaced Short Palindromic Repeats and CRISPR-associated protein 9). The CRISPR-Cas9 system, which was initially inspired by an adaptive immune mechanism in bacteria, allows for precise, targeted modifications of genomic DNA. The Cas9 endonuclease is guided to a particular DNA sequence by base pairing, where the Cas9 enzyme introduces a double-stranded break (DSB), using a synthetic single-guide RNA (sgRNA) [25,26,27]. Targeted gene disruption or insertion results from the cell's endogenous repair pathways, which mainly involve non-homologous end joining (NHEJ) or homology-directed repair (HDR), repairing this break. A vital tool for understanding gene function and simulating genetic changes pertinent to cancer biology, CRISPR-Cas9 is versatile, easy to use, and highly effective [28,29].
The CRISPR/Cas9 system is the third generation of genome editing tools, providing better efficiency and specificity than earlier technologies like transcription activator-like effector nucleases and zinc finger nucleases. Initially identified as an adaptive immune system of bacteria, CRISPR/Cas9 directs the Cas9 endonuclease to complementary DNA sequences for cleavage by using tiny RNA fragments from prior viral infections. In genome editing applications, Cas9 is guided to a particular genomic locus where it introduces a double-strand break (DSB) using a simplified single guide RNA (sgRNA) that combines tracrRNA and crRNA. Cellular processes like homology-directed repair (HDR) or non-homologous end joining (NHEJ) mend this break, enabling precise insertion or gene disruption (Figure 1A) [30]. Because multiple genetic mutations frequently underlie the progression of disease, this system is especially useful in cancer research due to its ability to multiplex—target multiple loci simultaneously using multiple sgRNAs. Compared to earlier nuclease systems or RNA interference, this technological advancement has created new opportunities for oncogene research, cancer modeling, and the development of more precise gene-targeted therapies.
The creation of tumor-specific delivery and expression systems is crucial to the therapeutic use of CRISPR/Cas9 in cancer in order to prevent off-target effects. To improve cancer-targeting effectiveness, a number of tactics are being investigated (Figure 1B) and Table 1 [31]. These include oncolytic viruses, liposomes guided by antibodies or aptamers, targeted extracellular vesicles, and adeno-associated viruses (AAVs) with tissue-specific tropism. It is also possible to use telomerase-dependent mechanisms, miRNA-responsive switches, or cancer-specific promoters to achieve cancer-selective expression of the CRISPR components. These methods guarantee that only malignant tissues have the system functioning. Gene editing techniques have also been developed to target vulnerabilities specific to tumors, like viral oncogene dependencies or synthetic lethality. It has been determined that non-viral delivery platforms are safer than viral vectors (Figure 1C). The CRISPR-Cas9 components can be delivered specifically into cells using these platforms, which are usually nanoparticles. This improves cellular uptake and lowers immunological responses. Stimuli-responsive nanocarriers (Figure 1D) are noteworthy because they can be designed to release CRISPR cargo in response to environmental cues like temperature, pH, or enzymes. These systems help the editing machinery localize itself in the nucleus and escape from endosomes, which increases the accuracy and effectiveness of gene modification. All of these developments are pushing CRISPR/Cas9 closer to clinical use in oncology and present a viable path toward highly targeted and customized cancer treatments [32].
One significant disadvantage of the CRISPR/Cas9 genome editing system is its propensity to cause off-target editing, or the inadvertent cleavage of DNA at genomic locations that share sequence similarities with the intended target. These off-target occurrences have the potential to disrupt gene function or promote carcinogenesis by causing insertion-deletion (indel) mutations and double-strand breaks (DSBs) within important genomic loci. In silico sgRNA design algorithms have been created to improve target specificity and decrease off-target effects in order to lessen this risk [33]. Furthermore, variations with improved specificity that drastically reduce off-target cleavage have been produced by engineering the Cas9 nuclease itself [34]. Notably, methods that use paired Cas9 "nickases," each of which produces a single-strand break, have also shown improved genomic precision and decreased indel formation [35]. Even with these developments, thorough genome-wide off-target screening is still necessary to guarantee safety and efficacy, especially for CRISPR-based cancer treatments.
A significant percentage of genetic mutations in cancer are single-nucleotide alterations in genes such as KRAS [41], EGFR [42], and NOTCH [43,44]. Base editors (BEs), which are CRISPR-related RNA-programmable deaminases, may be able to target these point mutations, as shown in Table 1 and Table 2. BEs use a deaminase to enzymatically change the base after unwinding a brief section of DNA known as the "editing window" at the precise location [45]. One major benefit of base editing is that, in contrast to traditional CRISPR/Cas9, it avoids the introduction of DSBs, which stops the NHEJ pathway from being activated and indel mutations from occurring. Additionally, because BEs don't depend on HDR repair, higher editing efficiencies can be sought and proven, and mRNA-based delivery systems are made simpler because no DNA donor template is needed [46]. Currently, there are two primary categories of BEs: Cystosine base editors (CBEs), which produce the opposite change of C-G base pairs to T-A base pairs, and Adenine base editors (ABEs), which convert A-T base pairs to G-C base pairs.
The effective and accurate delivery of CRISPR/Cas9 into target cells is also necessary for its clinical utility. Navigating both the nuclear and cellular membranes is necessary for successful genome editing, which poses significant difficulties. Because of their ease of use and effectiveness, non-specific methods like electroporation or microinjection of plasmids encoding Cas9 and sgRNA have been widely used in vitro [49]. Although hydrodynamic tail vein injection in murine models and other in vivo plasmid-based delivery methods have been investigated, their effectiveness is limited, and they cannot be applied to humans [47,50,51]. The CRISPR/Cas9 system must be encapsulated in an appropriate carrier for therapeutic delivery, such as viral vectors (e.g., G. AAV or lentivirus) or non-viral systems like extracellular vesicles or lipid nanoparticles. The safety and effectiveness of the CRISPR/Cas9 system in treating cancer have been evaluated in about 25 clinical trials to date, thanks to continuous technological advancements targeted at resolving significant obstacles in the clinical translation of CRISPR-based therapies, as shown in Table 4. Currently in Phase I or II, these studies mainly investigate methods to prevent cancer cells from acting malignantly. CRISPR/Cas9-edited immune cells, which are genetically altered to precisely target and impede tumor progression in patients with advanced-stage cancers, are frequently transplanted autologously.
CRISPR/Cas9 can be transported by these carriers in several different forms, including DNA plasmids that need intracellular transcription and translation, preassembled Cas9-sgRNA ribonucleoprotein (RNP) complexes, or mRNA encoding Cas9 and sgRNA[52,53] [42,43]. As Table 3 illustrates, each format has unique benefits and drawbacks with regard to effectiveness, stability, and immunogenicity. But improving targeted delivery methods is a top priority in the creation of CRISPR-based cancer treatments. By reducing off-target effects in healthy tissues, selective targeting seeks to maximize uptake by cancerous cells. Current research is looking into ligand- or antibody-mediated targeting, delivery vehicles that respond to the tumor microenvironment, and biomaterial-based systems that can take advantage of physiological conditions or surface markers unique to cancer. To fully realize the therapeutic potential of CRISPR/Cas9 in oncology, these new methods highlight how crucial it is to optimize delivery specificity.
The effective and accurate delivery of CRISPR/Cas9 into target cells is also necessary for its clinical utility. Navigating both the nuclear and cellular membranes is necessary for successful genome editing, which poses significant difficulties. Because of their ease of use and effectiveness, non-specific methods like electroporation or microinjection of plasmids encoding Cas9 and sgRNA have been widely used in vitro [49]. Although hydrodynamic tail vein injection in murine models and other in vivo plasmid-based delivery methods have been investigated, their effectiveness is limited, and they cannot be applied to humans [47,50,51]. The CRISPR/Cas9 system must be encapsulated in an appropriate carrier for therapeutic delivery, such as viral vectors (e.g., G. AAV or lentivirus) or non-viral systems like extracellular vesicles or lipid nanoparticles. The safety and effectiveness of the CRISPR/Cas9 system in treating cancer have been evaluated in about 25 clinical trials to date, thanks to continuous technological advancements targeted at resolving significant obstacles in the clinical translation of CRISPR-based therapies, as shown in Table 4. Currently in Phase I or II, these studies mainly investigate methods to prevent cancer cells from acting malignantly. CRISPR/Cas9-edited immune cells, which are genetically altered to precisely target and impede tumor progression in patients with advanced-stage cancers, are frequently transplanted autologously.
CRISPR/Cas9 can be transported by these carriers in several different forms, including DNA plasmids that need intracellular transcription and translation, preassembled Cas9-sgRNA ribonucleoprotein (RNP) complexes, or mRNA encoding Cas9 and sgRNA[52,53] [42,43]. As Table 3 illustrates, each format has unique benefits and drawbacks with regard to effectiveness, stability, and immunogenicity. But improving targeted delivery methods is a top priority in the creation of CRISPR-based cancer treatments. By reducing off-target effects in healthy tissues, selective targeting seeks to maximize uptake by cancerous cells. Current research is looking into ligand- or antibody-mediated targeting, delivery vehicles that respond to the tumor microenvironment, and biomaterial-based systems that can take advantage of physiological conditions or surface markers unique to cancer. To fully realize the therapeutic potential of CRISPR/Cas9 in oncology, these new methods highlight how crucial it is to optimize delivery specificity.

Table 4.
Overview of ongoing and completed CRISPR-based clinical trials in cancer therapy.
| Pathological Condition | Phase | Status | CRISPR-Engineered Therapeutic Agent | Other Therapies | Identifier |
|---|---|---|---|---|---|
| B-cell acute lymphoblastic leukemia | Phase 1 | Recruiting | Allogenic transplantation of engineered T cells—PBLTT52CAR19 | – | NCT04557436 |
| CD19+ leukemia and lymphoma | Phase 1 | Withdrawn | Allogenic transplantation of engineered T cells—PACE CART19 | – | NCT05037669 |
| Gastrointestinal cancers | Ph½ | Recruiting | Autologous transplantation of CISH-CRISPR TILs | Cyclophosphamide, fludarabine, aldesleukin | NCT04426669 |
| HIV + hematologic malignancies | N/A | Unknown | CRISPR/Cas9-modified CD34+ hematopoietic stem/progenitor cells (CCR5 knockout) | – | NCT03164135 |
| HPV-related malignant neoplasm | Phase 1 | Unknown | Local application of HPV16/18 E6/E7 CRISPR | – | NCT03057912 |
| EBV-associated malignancies | Ph½ | Recruiting | Autologous transplantation of PD-1 knockout EBV-CTL cells | Cyclophosphamide, fludarabine, interleukin-2 | NCT03044743 |
| Non-small cell lung cancer | Phase 1 | Completed | Autologous PD-1 knockout T cells | Cyclophosphamide | NCT02793856 |
| Renal cell carcinoma | Phase 1 | Withdrawn (no funding) | Autologous PD-1 knockout T cells | Cyclophosphamide, interleukin-2 | NCT02867332 |
| Prostate cancer | Phase 1 | Withdrawn (no funding) | Autologous PD-1 knockout T cells | Cyclophosphamide, interleukin-2 | NCT02867345 |
| Bladder cancer | Phase 1 | Withdrawn (no funding) | Autologous PD-1 knockout T cells | Cyclophosphamide, interleukin-2 | NCT02863913 |
| Hepatocellular carcinoma | Phase 1 | Recruiting | Autologous PD-1 knockout T cells | TACE | NCT04417764 |
| Esophageal cancer | Phase 1 | Completed | Autologous PD-1 knockout T cells | – | NCT03081715 |
In the basic and translational domains of cancer research, CRISPR-Cas9 has enabled impressive advancements. With the use of high-throughput loss-of-function or gain-of-function screens, the system enables the methodical screening of tumor suppressor genes and oncogenes. Finding new cancer drivers, artificially created lethal interactions, and possible treatment targets has been made possible thanks in large part to these functional genomic studies. Additionally, by making it possible to engineer particular mutations or chromosomal rearrangements that mimic the complexity of human tumors, CRISPR has sped up the development of complex cancer models, such as in vitro organoids and in vivo patient-derived xenografts (PDX). CRISPR is being investigated therapeutically for ex vivo editing of immune cells, including T cells that have been modified to express chimeric antigen receptors (CAR-T cells), which will improve anti-tumor immunity. Additionally, preclinical research has shown that targeting oncogenic mutations with CRISPR in vivo is feasible, providing a way forward for precision cancer treatment [54,55].
The clinical translation of CRISPR-Cas9 in oncology is hampered by a number of issues, despite its revolutionary potential. Off-target effects are a serious worry since they can result in genotoxicity and carcinogenesis by editing unintended genomic loci. Developments in high-fidelity Cas9 variants and sgRNA design algorithms (e.g., A. eSpCas9 and HiFi Cas9), as well as base-editing systems, have reduced off-target activity and increased specificity. Another major obstacle, particularly for in vivo applications, is getting CRISPR components into target cells or tissues. Although viral vectors, like adeno-associated viruses (AAV), provide effective delivery, their immunogenicity and cargo size are constrained. Alternative non-viral delivery methods like extracellular vesicles and lipid nanoparticles are being researched. Furthermore, the effectiveness of CRISPR-based therapies may be restricted by the complexity of tumor heterogeneity and the tumor microenvironment, requiring combinatorial methods and context-specific validation [56,57,58]. Innovations in CRISPR technology are rapidly overcoming these limitations and expanding its utility in cancer research. To reduce genomic instability, prime editing and CRISPR-associated transposases provide more accurate and adaptable genome engineering capabilities without causing DSBs. By enabling transcriptional regulation of endogenous genes without changing DNA sequences, CRISPR interference (CRISPRi) and CRISPR activation (CRISPRa) offer a reversible and adjustable method for functional research. Moreover, researchers can examine epigenetic dysregulation in carcinogenesis thanks to CRISPR-based epigenome editing tools, which modify DNA methylation and histone marks. Combinatorial gene editing, which is crucial for simulating intricate genetic interactions and resistance mechanisms in cancer, is made easier by multiplexed CRISPR systems and barcoding techniques. All of these developments are working together to make CRISPR a more reliable platform for target validation, cancer modeling, and treatment development [59,60,61].
In conclusion, the CRISPR-Cas9 system has transformed cancer research by making it possible to create more complex cancer models, to evaluate a functional gene, and to make precise genomic modifications. Even though issues with specificity, delivery, and tumor complexity still exist, new developments in technology are removing these obstacles and increasing CRISPR's usefulness in oncology. The development of new cancer treatments and individualized treatment plans is expected to be accelerated by the ongoing integration of CRISPR with cutting-edge disciplines like single-cell genomics, immuno-oncology, and artificial intelligence.
3.3D Bioprinting for Cancer Modeling
In cancer research, 3D bioprinting has become one of the game-changing tools because it provides previously unheard-of capabilities for creating physiologically accurate tumor models. The intricate cellular interactions, spatial heterogeneity, and tumor microenvironment (TME) observed in human cancers are frequently not replicated by conventional two-dimensional (2D) cell cultures and animal models. By contrast, 3D bioprinting makes it possible to create biomimetic tumor tissues through the precise spatial deposition of cells, biomaterials, and signaling factors. By facilitating the integration of stromal components, vasculature, and patient-derived cells, this technique offers a more precise platform for researching drug responses, metastasis, and cancer biology. Advanced preclinical platforms for therapeutic testing and personalized medicine are provided by 3D bioprinted models, which replicate tumor architecture and microenvironmental gradients [62,63,64].
The fundamentals of 3D bioprinting include the layer-by-layer deposition of bioinks—which are made of supportive biomaterials and living cells—using methods like extrusion, inkjet, and laser-assisted bioprinting. Droplets of cell-laden bioinks are used in inkjet bioprinting, which is appropriate for high-throughput applications but may be constrained by viscosity issues. Extrusion-based bioprinting, the most popular technique, allows for the deposition of dense cellular structures and works with a wide range of viscosities. Laser-assisted bioprinting is perfect for creating complex tumor microarchitectures because it provides high resolution and cell viability. These methods, which are frequently combined with computer-aided design (CAD) programs, enable the creation of tumor models that more accurately depict the behavior of tumors in vivo by customizing their geometry, cell density, and matrix composition.
A new therapeutic agent must normally be developed over a period of fifteen years and cost between $2 and $6 billion, making the traditional drug development pipeline both time-consuming and prohibitively expensive [65]. Traditionally, animal models and two-dimensional (2D) cell culture systems are used for preclinical evaluation of drug safety and efficacy. Nevertheless, these platforms frequently fall short in accurately simulating physiological reactions in humans. The unnatural growth conditions in 2D cultures cause cells to change in shape and behavior, and the translational relevance of animal models is limited by interspecies variations in anatomy and physiology. To increase the predictive accuracy of preclinical testing, more human-relevant in vitro platforms are desperately needed. Organoids made from bioengineered human cells offer a promising substitute that could increase clinical trial success rates by producing data unique to each individual [66,67,68].
Furthermore, by facilitating patient-specific therapeutic screening, these organoid models have great potential for personalized medicine, especially when it comes to complex diseases like cancer. The inability to produce large quantities of morphologically uniform organoids compatible with standard assay formats, such as 96- and 384-well plates, has limited the integration of organoids into high-throughput drug screening workflows, despite their benefits. To address this limitation, we present an immersion bioprinting strategy capable of generating consistent tissue organoids within 96-well plates, thereby enhancing the throughput of 3D drug screening platforms. In this approach, a composite hydrogel bioink, comprising hyaluronic acid and collagen, is deposited into a viscous gelatin support bath [69,70,71,72]. This support medium prevents interaction between the bioink and the well surfaces and maintains the spherical architecture of the printed constructs. The method was initially validated using established cancer cell lines and subsequently applied to patient-derived glioblastoma (GBM) and sarcoma biospecimens for therapeutic screening, as illustrated in Figure 2.
The formulation and selection of bioinks is a crucial step in 3D bioprinting for cancer modeling. Similar to the extracellular matrix (ECM) of native tissues, bioinks must promote cell viability, proliferation, and differentiation. Natural hydrogels with bioactivity and biocompatibility, like collagen, gelatin, hyaluronic acid, and decellularized extracellular matrix (dECM), are frequently utilized. Although Pluronic F127 and polyethylene glycol (PEG), two synthetic polymers, have mechanical properties that can be adjusted, they frequently need to be chemically modified to improve cell interactions. To balance biofunctionality and printability, composite bioinks—which combine natural and synthetic components—are commonly used for tumor modeling. Furthermore, the incorporation of immune cells, endothelial cells, and cancer-associated fibroblasts (CAFs) into bioinks facilitates the development of more representative organotypic tumor models, thereby bolstering research on angiogenesis and tumor-stroma interactions [73,74,75,76].
The physiological relevance of in vitro cancer models has been greatly increased by recent developments in organotypic tumor constructs. In order to replicate the complexity of tumors, bioprinted constructs now incorporate vascular networks, hypoxic gradients, and multicellular architectures. For instance, patient-derived tumor cells embedded in brain-mimicking dECM have been used to create glioblastoma models, which allow researchers to examine drug resistance mechanisms and invasive behavior. Similar to this, stromal and immune components have been added to breast and pancreatic tumor models, offering insights into immunological evasion and treatment response. The application of these constructs for assessing drug perfusion and metastatic potential is further improved by the creation of vascularized tumor models using endothelialized microchannels. Furthermore, dynamic monitoring of tumor responses to chemotherapeutics under controlled physiological conditions has been made easier by the integration of 3D bioprinting with microfluidic organ-on-a-chip platforms [77,78,79].
In conclusion, with its ability to produce high-fidelity, customizable tumor constructs that outperform conventional models, 3D bioprinting is a paradigm shift in cancer modeling. The spatial, cellular, and molecular complexities of human tumors can now be replicated by researchers using organotypic tissue engineering, optimized bioinks, and sophisticated bioprinting techniques. These advancements have the potential to advance personalized therapies, screen for anticancer agents, and reveal new mechanisms of cancer progression. To improve bioprinted tumor models and turn them into clinically useful applications, biomaterials scientists, cancer biologists, and biomedical engineers must continue their interdisciplinary cooperation.
4. Integration of CRISPR-Cas9 with 3D Bioprinted Cancer Models
In oncology research, the integration of CRISPR-Cas9 gene editing technology and 3D bioprinted cancer models is revolutionary because it allows for previously unheard-of precision in modifying genetic and microenvironmental parameters within physiologically relevant constructs. Researchers can introduce, remove, or alter particular genes implicated in tumorigenesis, drug resistance, and immune evasion thanks to CRISPR-Cas9's high efficiency and specificity in targeted genome editing. This approach makes it possible to create in vitro tumor models that more closely mimic the heterogeneity and spatial complexity of native tumors when paired with 3D bioprinting, which makes it possible to create intricate tissue-like structures using cancer cells, stromal components, and extracellular matrices. These integrated platforms provide strong tools for screening tailored treatment plans, validating oncogenic pathways, and clarifying gene function in situ [80,81,82].
There are a number of methods for successfully combining 3D bioprinting technology with CRISPR-Cas9. Before adding them to bioinks for 3D bioprinting, one method entails pre-editing patient-derived cells or cancer cell lines using CRISPR-Cas9. Using this technique guarantees that stable genetic alterations are maintained throughout the bioprinting process and permits the accurate spatial organization of genetically altered cell populations inside the construct. An alternative is to directly edit genes within the bioprinted structure after post-printing. This would enable real-time gene expression modification in response to therapeutic interventions or microenvironmental cues. Delivery methods for this strategy include viral vectors, nanoparticles, or electroporation, as well as inducible CRISPR systems. These techniques provide spatiotemporal control, which is essential for simulating dynamic aspects of tumor progression like angiogenesis, metastatic potential, and the epithelial-to-mesenchymal transition (EMT) [83,84,85]. Utilizing inducible systems, like CRISPR activation (CRISPRa) or interference (CRISPRi), improves the capacity to reversibly alter gene expression in the three-dimensional tumor model. The schematic workflow of an integrated CRISPR-Cas9 and 3D bioprinting approach for genome-wide screening in breast cancer organoids is described in Figure 3.
Using CRISPR-Cas9 integrated bioprinted models presents a special chance to create tumor microenvironments (TMEs) with architectural and genetic accuracy. TMEs that closely resemble in vivo tumor conditions can be recreated by researchers by modifying genes involved in extracellular matrix remodeling, immune modulation, or angiogenic signaling. For example, hypoxia-induced vascular remodeling can be mimicked by overexpressing stromal cell-derived factors like VEGF or knocking out matrix metalloproteinases (MMPs) in the printed construct's stromal or endothelial compartments. Additionally, the co-printing of immune cells that have had their genes altered, like T cells that have immune checkpoint genes knocked out (e.g., A. The study of immune-tumor interactions and immunotherapy responses is made possible by the interaction of PD-1 or CTLA-4 with tumor cells. To comprehend resistance mechanisms and assess novel treatment approaches, a thorough recapitulation of tumor-stroma-immune crosstalk is necessary. These models also offer a platform for examining gene-environment interactions in solid tumor-specific nutrient gradients, hypoxia, and mechanical stress scenarios [86,87,88].
The importance of integrating 3D bioprinting and CRISPR-Cas9 in improving cancer biology has been highlighted by recent studies. For instance, using CRISPR-edited glioma stem cells devoid of PTEN or TP53, scientists have created 3D glioblastoma models, allowing for the tissue-mimetic study of tumorigenic mechanisms and drug responses. In a different study, CRISPR-modified cells were used to bioprint breast cancer organoids in order to evaluate the impact of BRCA1 mutations on tumor progression and treatment sensitivity. These platforms have also been used to model uncommon cancers and their metastases, like pancreatic ductal adenocarcinoma and uveal melanoma, which frequently show unique mutational signatures and interactions with the microenvironment. Furthermore, these genetically customized models now have more cellular heterogeneity and spatial resolution thanks to developments in bioprinting methods like multi-material extrusion and microfluidic-assisted droplet bioprinting. These developments are speeding up biomarker-driven drug development and the identification of actionable mutations [89,90,91].
In conclusion, an integrating platform to study tumor biology with high genetic and structural fidelity is being made possible by the combination of 3D bioprinting and CRISPR-Cas9 gene editing, which is transforming the creation of next-generation cancer models. By manipulating particular genetic components within physiologically relevant architectures, these engineered constructs enable mechanistic insights into immunological evasion, drug resistance, and oncogenesis. Additionally, the creation of high-throughput drug screening platforms and customized treatment plans is facilitated by this integrative approach. Future advancements in gene editing effectiveness, bioprinting resolution, and multicellular modeling should increase the systems' applicability in translational oncology as well as fundamental research.
5. Modeling Tumor Progression and Metastasis by Integration of CRISPR-Cas9 with 3D Bioprinted Cancer Models
The integration of CRISPR-Cas9 gene-editing technology with three-dimensional (3D) bioprinted cancer models offers a powerful platform to dissect the complex genetic and microenvironmental factors underlying tumor progression and metastasis. Fundamentally, genetic changes that build up over time and produce diverse cancer cell populations with unique phenotypic characteristics are what propel tumor evolution. Researchers can functionally interrogate genetic drivers that promote oncogenesis, therapeutic resistance, and metastatic potential by using CRISPR-Cas9 to precisely manipulate particular genes within 3D bioprinted tumor constructs. In contrast to conventional two-dimensional (2D) cultures, 3D bioprinted models provide a more physiologically relevant context for studying tumor biology by more accurately simulating the spatial organization, cell-cell, and cell-matrix interactions of in vivo tumors. This combination of biomimetic modeling and precise genome editing has great potential to improve our comprehension of tumor evolution and find new therapeutic targets [92,93,94].
The dynamic interaction between cancer cells and the surrounding stromal components, such as fibroblasts, immune cells, endothelial cells, and the extracellular matrix (ECM), is essential to the progression of tumors. Tumor growth, angiogenesis, immune evasion, and metastatic dissemination are all dependent on these tumor–stroma interactions. Multiple cell types can be co-cultured in predetermined architectures thanks to 3D bioprinting's ability to reconstruct these intricate cellular microenvironments with high spatial fidelity. Certain signaling pathways in stromal and tumor cells can be altered via CRISPR-Cas9-mediated gene editing to examine their functional contributions to tumor progression and reciprocal crosstalk. For instance, altering genes that control immune checkpoint expression or ECM remodeling can clarify how the stroma controls tumor invasiveness and immune suppression. Thus, this integrated approach bridges the gap between in vitro research and in vivo tumor behavior by allowing the controlled dissection of heterogeneous tumor ecosystems [95,96,97].
In cancer, metastasis—the movement of cancer cells from the original tumor to other organs—continues to be the main cause of death. To comprehend the progression of metastatic disease and resistance to treatment, it is crucial to model metastatic niches, which are the specialized microenvironments in secondary organs that facilitate the colonization and proliferation of transplanted tumor cells. By creating vascular networks, stromal populations, and tissue-specific extracellular matrix compositions that resemble organ microenvironments like the liver, lung, or bone marrow, 3D bioprinting provides an unprecedented ability to engineer metastatic niches. When used in conjunction with CRISPR-Cas9 technology, tumor cells can be genetically altered to evaluate the roles of genes that promote metastasis in colonization and dormancy. This platform makes it possible to track tumor cell invasion, extravasation, and niche adaptation in real time in response to specific mechanical and biochemical cues. Such biomimetic metastatic models serve as sophisticated drug screening platforms to assess anti-metastatic treatments individually for each patient, in addition to facilitating mechanistic research [98,99,100].
In conclusion, a revolutionary development in cancer research is the integration of CRISPR-Cas9 gene editing and 3D bioprinted cancer models. This integrated approach offers unparalleled insight into the intricate biology of metastatic niches, the crucial tumor–stroma interactions, and the genetic drivers of tumor evolution by enabling precise genetic manipulation within physiologically relevant tumor microenvironments. For mechanistic research, biomarker identification, and therapeutic development, this platform provides a reliable experimental system that overcomes the drawbacks of traditional models. These models' translational relevance will be further increased by future initiatives aimed at advancing genome editing specificity, enhancing bioprinting technologies, and integrating patient-derived cells, which will hasten the transition to personalized cancer treatment.
6. Integration of CRISPR-Cas9 with 3D Bioprinted Models for Evaluation of Treatment Response
A major development in personalized oncology is the integration of CRISPR-Cas9 gene-editing technology with 3D bioprinted tumor models, which provides high-resolution control over genetic manipulation in physiologically relevant microenvironments. Complex tissue-like structures that mimic the extracellular matrix, architecture, and heterocellular makeup of natural tumors can now be created thanks to 3D bioprinting. These models can be genetically modified to carry oncogenic drivers or patient-specific mutations when paired with CRISPR-Cas9, allowing for a more thorough investigation of tumorigenic pathways and therapeutic vulnerabilities. In addition to making it possible to model tumor behavior more realistically, this convergence makes it easier to create predictive platforms for drug screening and mechanistic resistance research. Drug screening with CRISPR-enhanced 3D bioprinted models is a practical and trustworthy way to assess treatment efficacy and investigate resistance mechanisms [101,102,103]. Using CRISPR-Cas9, scientists can introduce particular gene knockouts or knock-ins in a spatially ordered bioprinted tumor model to mimic various oncogenic scenarios and assess drug responses under predefined genetic conditions. It is possible to introduce loss-of-function mutations in TP53 or gain-of-function mutations in KRAS to examine their impact on drug sensitivity in colorectal or pancreatic cancer models. Additionally, the 3D microenvironment adds yet another level of complexity by simulating hypoxic gradients, drug penetration barriers, and stromal interactions—all of which are commonly absent in traditional 2D systems. It is feasible to perform high-throughput pharmacological profiling and identify gene-drug interactions that result in intrinsic or acquired resistance thanks to this complex integration [104].
CRISPR-bioprinted tumor platforms provide a potent method for assessing immunotherapy responses in a regulated, genetically defined environment, in addition to chemotherapeutic and targeted agents. The expression of immune checkpoints can be altered in genetically modified tumor cells (e.g., G. T cell infiltration, activation, and cytotoxicity in co-culture systems with immune cells can be evaluated thanks to PD-L1, CTLA-4, or neoantigens. These models are especially useful for analyzing the function of immune evasion tactics used by tumors, such as MHC downregulation and immunosuppressive cytokine secretion. Researchers can systematically test the effects of immune checkpoint inhibitors, adoptive T cell therapies, and bispecific antibodies by creating isogenic tumor variants that differ only in immune-modulatory gene expression. Furthermore, the physiological relevance of immune-oncology research can be improved by designing bioprinted models to include elements of the tumor microenvironment, such as fibroblasts, myeloid-derived suppressor cells (MDSCs), and regulatory T cells (Tregs) [16,105,106,107].
The predictive value of CRISPR-enhanced 3D bioprinted tumor models lies in their capacity to recapitulate the genetic and phenotypic diversity observed in patient tumors while enabling iterative functional validation. These platforms can be used to stratify treatments, find biomarkers, and track the evolution of resistance in real time. CRISPR screens, for example, can find new druggable targets, artificially created lethal interactions, or compensatory pathways that are triggered by therapy in bioprinted constructs. Additionally, clonal evolution can be modeled by bioprinting and editing patient-derived cells, which captures intra-tumoral heterogeneity and its effect on therapeutic response. Transcriptomic profiling, proteomic analysis, and real-time imaging can all be integrated into these systems to create dynamic tools for clinical outcome prediction and precision treatment strategy customization [108,109,110,111].
In conclusion, a revolutionary platform for cancer research and treatment development is provided by the synergistic integration of CRISPR-Cas9 gene editing and 3D bioprinted tumor models. Through precise genetic manipulation in physiologically relevant microenvironments, this method enables nuanced assessment of immunotherapeutic efficacy, resistance mechanisms, and drug response. CRISPR-enhanced bioprinted models have enormous potential to speed up translational research and increase the predictive capacity of preclinical studies in oncology by bridging the gap between genetic engineering and tissue engineering.
7. Integration of CRISPR-Cas9 with 3D Bioprinting: Challenges and Limitations
The integration of CRISPR-Cas9 gene-editing technology with 3D bioprinting represents a groundbreaking frontier in biomedical research, with potential applications in personalized medicine, regenerative therapies, and disease modeling. Nevertheless, there are substantial biological and technical obstacles to this convergence. A significant obstacle is the stability and delivery of CRISPR-Cas9 components inside 3D bioprinted structures. For gene editing to be effective, the Cas9 nuclease and guide RNA must be delivered to target cells precisely and effectively without endangering the structural integrity or viability of the cells. This delivery gets more complicated in 3D bioprinted tissues, which frequently have an extracellular matrix that is dense and has diverse cell populations. Even though they are effective, viral vectors can cause immunogenicity and insertional mutagenesis, whereas non-viral vectors, like nanoparticles, can only spread temporarily and have limited penetration. Furthermore, preserving gene-editing effectiveness and cell functionality after bioprinting is still a major challenge, especially when taking functional integration and long-term tissue maturation into account[112,113,114].
Other challenges in this integrative approach include clinical translation, reproducibility, and scalability. Standardized bioink formulations, consistent bioprinting parameters, and uniform gene delivery are critical to the reproducibility of CRISPR-Cas9 editing in 3D printed tissues. The creation of trustworthy tissue models for translational research can be hampered by variations in scaffold architecture, cell distribution, and local microenvironments, which can result in inconsistent editing outcomes. Furthermore, the difficulty of creating vascularized, multicellular constructs that maintain stable gene edits over long culture times continues to restrict the scalability of this integration. This raises questions about batch-to-batch variability and the bioprinting process's resilience under Good Manufacturing Practice (GMP) guidelines for clinical applications. Furthermore, the regulatory pathways for bioprinted products and CRISPR-edited tissues are still developing, which makes their path to therapeutic application more difficult [115,116,117].
The integration of CRISPR-Cas9 with 3D bioprinting presents a particularly complex ethical and regulatory environment. Even though each technology presents important ethical issues on its own, when combined, they create previously unheard-of problems. The application of CRISPR in human-derived bioprinted tissues, for example, raises concerns about the limits of genetic modification that are acceptable, particularly when germline cells or heritable edits are involved [118,119,120]. These debates are heightened by worries about producing human-animal chimeras, improving human characteristics, or building organs that resemble humans and have neural functions. Regulatory bodies must assess the effectiveness and safety of gene-edited constructs while also considering the wider societal ramifications. Standardized international guidelines are needed, particularly since scientific discoveries are outpacing the creation of policies. The main moral and legal issues in this area are compiled in Table 5.
In conclusion, A number of technological, biological, ethical, and legal restrictions prevent the revolutionary potential of integrating CRISPR-Cas9 with 3D bioprinting. Collaboration between bioengineers, geneticists, ethicists, and policymakers across disciplinary boundaries will be necessary to address these issues. Developments in tissue engineering standards, bioink optimization, and delivery technologies may aid in removing present obstacles. At the same time, a strong regulatory framework that strikes a balance between creativity and moral obligation is necessary for the safe and just application of this potent biotechnological fusion. As the field develops, transferring these technologies from the lab to clinical and societal use will require responsible innovation.
8. Integration of CRISPR-Cas9 with 3D Bioprinted Cancer Models: Future Perspectives
A revolutionary approach to oncology is presented by the integration of 3D bioprinted cancer models and CRISPR-Cas9 genome editing technology, which provides previously unheard-of precision in comprehending tumor biology and promoting personalized medicine. While CRISPR-Cas9 allows targeted gene modification to analyze gene function, model disease mutations, and evaluate therapeutic vulnerabilities, three-dimensional bioprinting allows the creation of intricate, multicellular constructs that closely mimic the tumor microenvironment. When combined, these technologies offer a synergistic platform that speeds up the creation of specialized treatment plans and improves the physiological relevance of in vitro cancer models [16,57,121].
A significant development in this field is the use of patient-derived cells to customize cancer models. Individualized tumor architectures that preserve the genetic and phenotypic diversity of native tumors can be replicated by researchers using primary tumor cells or patient-derived organoids in bioprinted constructs. Oncogenic pathways can be studied in a patient-contextual framework by using CRISPR-Cas9 to introduce or correct mutations unique to a patient. Functional precision oncology may benefit from the use of such customized models in determining drug responses and resistance mechanisms unique to individual patients. The results of genome editing can also be evaluated in real time in a physiologically relevant microenvironment thanks to these systems[122,123,124].
The effectiveness of CRISPR-edited 3D bioprinted models is further increased by integrating them with multi-omics and artificial intelligence (AI) technologies. AI systems are capable of processing high-dimensional datasets from proteomic, metabolomic, transcriptomic, and genomic analyses in order to find important molecular signatures and forecast the results of treatment. The use of AI in conjunction with CRISPR screening in 3D models can direct the discovery of new therapeutic targets and synthetic lethal interactions. High-throughput omics profiling of modified models also makes it possible to map cellular responses in great detail, which enhances our understanding of how genes function in the complex temporal and spatial dynamics of tumors. The combination of experimental and computational platforms has great potential to reveal the complex nature of cancer [125,126,127].
The use of CRISPR-Cas9 in bioprinted cancer models is a critical step toward clinical translation and the realization of precision oncology. In a controlled but clinically relevant environment, bioprinted models provide scalable, repeatable platforms for drug screening and gene-editing strategy validation. Crucially, in these models, CRISPR-based functional assays are able to forecast both therapeutic efficacy and resistance, allowing for patient stratification for focused interventions. Ethics and regulations are still important, particularly when applying results from engineered models to clinical settings. Current translational barriers, however, might be overcome by improving CRISPR delivery systems and creating bioprinting protocols that adhere to good manufacturing practice (GMP) [70,128,129,130].
In conclusion, a significant advancement in cancer research and treatment is the combination of 3D bioprinted cancer models and CRISPR-Cas9 genome editing. Strong platforms for functional genomics and drug discovery are provided by personalized models made with patient-derived cells that are improved by AI and omics integration. The combined potential of these technologies to advance precision oncology and enhance clinical results is becoming more and more apparent as they develop. Transforming this promising paradigm into standard clinical practice will require ongoing interdisciplinary cooperation and technological advancement.
9. Conclusion
The integration of 3D bioprinted cancer models and CRISPR-Cas9 genome editing represents a paradigm shift in cancer research and treatment advancement. An original, multifaceted method for deconstructing intricate tumor biology in physiologically significant microenvironments is provided by this convergence. Researchers can now more accurately model oncogenic mutations, tumor heterogeneity, and drug responses than they could with conventional 2D cultures or animal models thanks to the ability to precisely modify genes in spatially organized, patient-specific constructs. This development is especially significant in the age of precision oncology, where it is critical to comprehend interpatient variability and forecast each patient's unique therapeutic response. Furthermore, the creation of customized cancer models that accurately represent the genetic and phenotypic complexity of clinical tumors is made possible by the integration of patient-derived cells into bioprinted constructs in conjunction with CRISPR-based gene editing.
These platforms allow for high-throughput screening of gene functions and therapeutic vulnerabilities and offer thorough insights into molecular changes when further combined with artificial intelligence and omics technologies. In addition to improving translational research, these systems aid in the discovery of new biomarkers and the creation of customized treatment plans. Even though there are still issues with scalability, regulatory compliance, and the effectiveness of CRISPR delivery, these issues are being addressed by continuous technological advancements. This integrated platform is well-positioned to close the gap between lab research and patient-specific therapy as bioprinting protocols advance toward clinical-grade manufacturing standards and CRISPR applications become more accurate and secure. In the end, the combination of CRISPR-Cas9 and 3D bioprinted cancer models provides a potent toolkit for developing the upcoming generation of tailored, focused cancer therapies and transforming clinical oncology.
Author Contributions
Conceptualization, AKS, SDD, SS and RKM.; methodology, SS, AMU, AKS, SDD, and RKM.; writing—original draft preparation, A.K.S.; writing—review and editing, SS, AMU, AKS, SDD, and RKM.; visualization, SS, AMU, AKS, SDD, and RKM.; management, SS, AKS, and RKM. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Walrath, J.C.; Hawes, J.J.; Van Dyke, T.; Reilly, K.M. Genetically engineered mouse models in cancer research. Adv Cancer Res. 2010, 106, 113–64. [Google Scholar]
- Xiao, W.; Jiang, W.; Chen, Z.; Huang, Y.; Mao, J.; Zheng, W.; et al. Advance in peptide-based drug development: delivery platforms, therapeutics and vaccines. Signal Transduction and Targeted Therapy 2025, 10, 74. [Google Scholar] [CrossRef]
- Selicharova, I.; Sanda, M.; Mladkova, J.; Ohri, S.S.; Vashishta, A.; Fusek, M.; et al. 2-DE analysis of breast cancer cell lines 1833 and 4175 with distinct metastatic organ-specific potentials: comparison with parental cell line MDA-MB-231. Oncol Rep. 2008, 19, 1237–44. [Google Scholar] [CrossRef]
- Ran, R.; Harrison, H.; Syamimi Ariffin, N.; Ayub, R.; Pegg, H.J.; Deng, W.; et al. A role for CBFβ in maintaining the metastatic phenotype of breast cancer cells. Oncogene 2020, 39, 2624–37. [Google Scholar] [CrossRef]
- Sajjad, H.; Imtiaz, S.; Noor, T.; Siddiqui, Y.H.; Sajjad, A.; Zia, M. Cancer models in preclinical research: A chronicle review of advancement in effective cancer research. Animal Model Exp Med. 2021, 4, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.Y.C.; Lin, D.; Gout, P.W.; Collins, C.C.; Xu, Y.; Wang, Y. Lessons from patient-derived xenografts for better in vitro modeling of human cancer. Advanced Drug Delivery Reviews 2014, 79-80, 222–37. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Koo, B.-K.; Knoblich, J.A. Human organoids: model systems for human biology and medicine. Nature Reviews Molecular Cell Biology 2020, 21, 571–84. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.C.; Liang, S.B.; Luo, M.; Wang, X.P. Intratumoral heterogeneity and drug resistance in cancer. Cancer Cell Int. 2025, 25, 103. [Google Scholar] [CrossRef]
- Yang, D.; Liu, J.; Qian, H.; Zhuang, Q. Cancer-associated fibroblasts: from basic science to anticancer therapy. Experimental & Molecular Medicine 2023, 55, 1322–32. [Google Scholar] [CrossRef]
- de Visser, K.E.; Joyce, J.A. The evolving tumor microenvironment: From cancer initiation to metastatic outgrowth. Cancer Cell. 2023, 41, 374–403. [Google Scholar] [CrossRef]
- Quail, D.F.; Joyce, J.A. Microenvironmental regulation of tumor progression and metastasis. Nat Med. 2013, 19, 1423–37. [Google Scholar] [CrossRef]
- Glabman, R.A.; Choyke, P.L.; Sato, N. Cancer-Associated Fibroblasts: Tumorigenicity and Targeting for Cancer Therapy. Cancers (Basel) 2022, 14(16). [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Ma, Y.; Li, D.; Wei, J.; Chen, K.; Zhang, E.; et al. Cancer associated fibroblasts and metabolic reprogramming: unraveling the intricate crosstalk in tumor evolution. J Hematol Oncol. 2024, 17, 80. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhou, Y.; Wang, Y.; Yang, J.; Li, Z.; Liu, F.; et al. Overcoming cancer treatment resistance: Unraveling the role of cancer-associated fibroblasts. Journal of the National Cancer Center 2025, 5, 237–51. [Google Scholar] [CrossRef]
- Makesh, K.Y.; Navaneethan, A.; Ajay, M.; Munuswamy-Ramanujam, G.; Chinnasamy, A.; Gnanasampanthapandian, D.; et al. A Concise Review of Organoid Tissue Engineering: Regenerative Applications and Precision Medicine. Organoids 2025, 4, 16. [Google Scholar] [CrossRef]
- Feng, X.; Li, Z.; Liu, Y.; Chen, D.; Zhou, Z. CRISPR/Cas9 technology for advancements in cancer immunotherapy: from uncovering regulatory mechanisms to therapeutic applications. Exp Hematol Oncol. 2024, 13, 102. [Google Scholar] [CrossRef]
- Pasupuleti, V.; Vora, L.; Prasad, R.; Nandakumar, D.N.; Khatri, D.K. Glioblastoma preclinical models: Strengths and weaknesses. Biochimica et Biophysica Acta (BBA) - Reviews on Cancer 2024, 1879, 189059. [Google Scholar] [CrossRef]
- Dell, A.C.; Wagner, G.; Own, J.; Geibel, J.P. 3D Bioprinting Using Hydrogels: Cell Inks and Tissue Engineering Applications. Pharmaceutics 2022, 14(12). [Google Scholar] [CrossRef]
- Gogoi, D.; Kumar, M.; Singh, J. A comprehensive review on hydrogel-based bio-ink development for tissue engineering scaffolds using 3D printing. Annals of 3D Printed Medicine 2024, 15, 100159. [Google Scholar] [CrossRef]
- Tripathi, S.; Mandal, S.S.; Bauri, S.; Maiti, P. 3D bioprinting and its innovative approach for biomedical applications. MedComm (2020) 2023, 4, e194. [Google Scholar] [CrossRef]
- Fatimi, A.; Okoro, O.V.; Podstawczyk, D.; Siminska-Stanny, J.; Shavandi, A. Natural Hydrogel-Based Bio-Inks for 3D Bioprinting in Tissue Engineering: A Review. Gels 2022, 8, 179. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Zhu, T.; Cui, H.; Cui, H. Integrating 3D Bioprinting and Organoids to Better Recapitulate the Complexity of Cellular Microenvironments for Tissue Engineering. Adv Healthc Mater. 2025, 14, e2403762. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.H.; Liu, N.; Zhang, H.; Yin, Z.S.; Zha, Z.G. From cells to organs: progress and potential in cartilaginous organoids research. J Transl Med. 2023, 21, 926. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Guo, F.; Jin, Y.; Ma, Y. Applications of human organoids in the personalized treatment for digestive diseases. Signal Transduction and Targeted Therapy 2022, 7, 336. [Google Scholar] [CrossRef]
- Aljabali, A.A.A.; El-Tanani, M.; Tambuwala, M.M. Principles of CRISPR-Cas9 technology: Advancements in genome editing and emerging trends in drug delivery. Journal of Drug Delivery Science and Technology 2024, 92, 105338. [Google Scholar] [CrossRef]
- Balasubramanian, A.; Veluswami, K.; Rao, S.; Aggarwal, S.; Mani, S. Exploring Clustered Regularly Interspaced Short Palindromic Repeats-CRISPR-Associated Protein 9 (CRISPR-Cas9) as a Therapeutic Modality for Cancer: A Scoping Review. Cureus 2024, 16, e64324. [Google Scholar] [CrossRef]
- Shamjetsabam, N.D.; Rana, R.; Malik, P.; Ganguly, N.K. CRISPR/Cas9, an overview of recent developments and applications in cancer research. Int J Surg. 2024, 110, 6198–213. [Google Scholar] [CrossRef]
- Li, H.; Yang, Y.; Hong, W.; Huang, M.; Wu, M.; Zhao, X. Applications of genome editing technology in the targeted therapy of human diseases: mechanisms, advances and prospects. Signal Transduct Target Ther. 2020, 5, 1. [Google Scholar] [CrossRef]
- Schiermeyer, A.; Schneider, K.; Kirchhoff, J.; Schmelter, T.; Koch, N.; Jiang, K.; et al. Targeted insertion of large DNA sequences by homology-directed repair or non-homologous end joining in engineered tobacco BY-2 cells using designed zinc finger nucleases. Plant Direct 2019, 3, e00153. [Google Scholar] [CrossRef]
- Nie, D.; Guo, T.; Yue, M.; Li, W.; Zong, X.; Zhu, Y.; et al. Research Progress on Nanoparticles-Based CRISPR/Cas9 System for Targeted Therapy of Tumors. Biomolecules 2022, 12, 1239. [Google Scholar] [CrossRef]
- Balon, K.; Sheriff, A.; Jacków, J.; Łaczmański, Ł. Targeting Cancer with CRISPR/Cas9-Based Therapy. International Journal of Molecular Sciences 2022, 23, 573. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Rho, W.-Y.; Kim, Y.-H.; Chang, H.; Jun, B.-H. CRISPR-Cas9 Gene Therapy: Non-Viral Delivery and Stimuli-Responsive Nanoformulations. Molecules 2025, 30, 542. [Google Scholar] [CrossRef] [PubMed]
- Ran, F.A.; Hsu, P.D.; Lin, C.-Y.; Gootenberg, J.S.; Konermann, S.; Trevino, A.E.; et al. Double nicking by RNA-guided CRISPR Cas9 for enhanced genome editing specificity. Cell. 2013, 154, 1380–9. [Google Scholar] [CrossRef] [PubMed]
- Jacków, J.; Guo, Z.; Hansen, C.; Abaci, H.E.; Doucet, Y.S.; Shin, J.U.; et al. CRISPR/Cas9-based targeted genome editing for correction of recessive dystrophic epidermolysis bullosa using iPS cells. Proceedings of the National Academy of Sciences 2019, 116, 26846–52. [Google Scholar] [CrossRef]
- Kocher, T.; Wagner, R.N.; Klausegger, A.; Guttmann-Gruber, C.; Hainzl, S.; Bauer, J.W.; et al. Improved double-nicking strategies for COL7A1-editing by homologous recombination. Molecular Therapy Nucleic Acids 2019, 18, 496–507. [Google Scholar] [CrossRef]
- Zhang, M.; Eshraghian, E.A.; Al Jammal, O.; Zhang, Z.; Zhu, X. CRISPR technology: The engine that drives cancer therapy. Biomedicine & pharmacotherapy 2021, 133, 111007. [Google Scholar]
- Chen, Z.-H.; Yu, Y.P.; Zuo, Z.-H.; Nelson, J.B.; Michalopoulos, G.K.; Monga, S.; et al. Targeting genomic rearrangements in tumor cells through Cas9-mediated insertion of a suicide gene. Nature biotechnology 2017, 35, 543–50. [Google Scholar] [CrossRef]
- Kwon, D.Y.; Zhao, Y.-T.; Lamonica, J.M.; Zhou, Z. Locus-specific histone deacetylation using a synthetic CRISPR-Cas9-based HDAC. Nature communications 2017, 8, 15315. [Google Scholar] [CrossRef]
- Wang, J.; Balan, V.; Marincola, F. CRISPR technology for immuno-oncology applications. Methods in Enzymology 2020, 635, 251–66. [Google Scholar]
- Azangou-Khyavy, M.; Ghasemi, M.; Khanali, J.; Boroomand-Saboor, M.; Jamalkhah, M.; Soleimani, M.; et al. CRISPR/Cas: from tumor gene editing to T cell-based immunotherapy of cancer. Frontiers in Immunology 2020, 11, 2062. [Google Scholar] [CrossRef]
- Perincheri, S.; Hui, P. KRAS mutation testing in clinical practice. Expert review of molecular diagnostics 2015, 15, 375–84. [Google Scholar] [CrossRef] [PubMed]
- Cheung, A.H.-K.; Chow, C.; Zhang, J.; Zhou, Y.; Huang, T.; Ng, K.C.-K.; et al. Specific targeting of point mutations in EGFR L858R-positive lung cancer by CRISPR/Cas9. Laboratory Investigation 2018, 98, 968–76. [Google Scholar] [CrossRef] [PubMed]
- Weng, A.P.; Ferrando, A.A.; Lee, W.; Morris, I.V.J.P.; Silverman, L.B.; Sanchez-Irizarry, C.; et al. Activating mutations of NOTCH1 in human T cell acute lymphoblastic leukemia. Science 2004, 306, 269–71. [Google Scholar] [CrossRef] [PubMed]
- Misiorek, J.O.; Przybyszewska-Podstawka, A.; Kałafut, J.; Paziewska, B.; Rolle, K.; Rivero-Müller, A.; et al. Context matters: NOTCH signatures and pathway in cancer progression and metastasis. Cells 2021, 10, 94. [Google Scholar] [CrossRef]
- Komor, A.C.; Kim, Y.B.; Packer, M.S.; Zuris, J.A.; Liu, D.R. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature 2016, 533, 420–4. [Google Scholar] [CrossRef]
- Anzalone, A.V.; Koblan, L.W.; Liu, D.R. Genome editing with CRISPR–Cas nucleases, base editors, transposases and prime editors. Nature biotechnology 2020, 38, 824–44. [Google Scholar] [CrossRef]
- Liu, C.; Zhang, L.; Liu, H.; Cheng, K. Delivery strategies of the CRISPR-Cas9 gene-editing system for therapeutic applications. Journal of Controlled Release 2017, 266, 17–26. [Google Scholar] [CrossRef]
- Yip, B.H. Recent advances in CRISPR/Cas9 delivery strategies. Biomolecules 2020, 10, 839. [Google Scholar] [CrossRef]
- Wang, L.; Zheng, W.; Liu, S.; Li, B.; Jiang, X. Delivery of CRISPR/Cas9 by novel strategies for gene therapy. ChemBioChem 2019, 20, 634–43. [Google Scholar] [CrossRef]
- Liu, B.; Saber, A.; Haisma, H.J. CRISPR/Cas9, a powerful tool for identification of new targets for cancer treatment. Drug Discovery Today 2019, 24, 955–70. [Google Scholar] [CrossRef]
- Yao, S.; He, Z.; Chen, C. CRISPR/Cas9-Mediated Genome Editing of Epigenetic Factors for Cancer Therapy. Human Gene Therapy 2015, 26, 463–71. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Alphonse, M.; Liu, Q. Strategies for nonviral nanoparticle-based delivery of CRISPR/Cas9 therapeutics. WIREs Nanomedicine and Nanobiotechnology 2020, 12, e1609. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Wan, T.; Xin, H.; Li, D.; Pan, H.; Wu, J.; et al. Delivery of CRISPR/Cas9 for therapeutic genome editing. The Journal of Gene Medicine 2019, 21, e3107. [Google Scholar] [CrossRef]
- Samareh Salavatipour, M.; Poursalehi, Z.; Hosseini Rouzbahani, N.; Mohammadyar, S.; Vasei, M. CRISPR-Cas9 in basic and translational aspects of cancer therapy. Bioimpacts 2024, 14, 30087. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Li, Z.; Liu, Y.; Chen, D.; Zhou, Z. CRISPR/Cas9 technology for advancements in cancer immunotherapy: from uncovering regulatory mechanisms to therapeutic applications. Experimental Hematology & Oncology 2024, 13, 102. [Google Scholar] [CrossRef]
- Allemailem, K.S.; Alsahli, M.A.; Almatroudi, A.; Alrumaihi, F.; Alkhaleefah, F.K.; Rahmani, A.H.; et al. Current updates of CRISPR/Cas9-mediated genome editing and targeting within tumor cells: an innovative strategy of cancer management. Cancer Commun (Lond) 2022, 42, 1257–87. [Google Scholar] [CrossRef]
- Li, T.; Li, S.; Kang, Y.; Zhou, J.; Yi, M. Harnessing the evolving CRISPR/Cas9 for precision oncology. J Transl Med. 2024, 22, 749. [Google Scholar] [CrossRef]
- Seijas, A.; Cora, D.; Novo, M.; Al-Soufi, W.; Sánchez, L.; Arana, Á.J. CRISPR/Cas9 Delivery Systems to Enhance Gene Editing Efficiency. International Journal of Molecular Sciences 2025, 26, 4420. [Google Scholar] [CrossRef]
- Laurent, M.; Geoffroy, M.; Pavani, G.; Guiraud, S. CRISPR-Based Gene Therapies: From Preclinical to Clinical Treatments. Cells 2024, 13(10). [Google Scholar] [CrossRef]
- Xu, Y.; Le, H.; Wu, Q.; Wang, N.; Gong, C. Advancements in CRISPR/Cas systems for disease treatment. Acta Pharmaceutica Sinica B 2025, 15, 2818–44. [Google Scholar] [CrossRef]
- Lei, T.; Wang, Y.; Zhang, Y.; Yang, Y.; Cao, J.; Huang, J.; et al. Leveraging CRISPR gene editing technology to optimize the efficacy, safety and accessibility of CAR T-cell therapy. Leukemia 2024, 38, 2517–43. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.-X.; Wu, Z.; Hou, Y.-Y.; Fang, Z.-X.; Deng, Y.; Wu, H.-T.; et al. Application of three-dimensional (3D) bioprinting in anti-cancer therapy. Heliyon 2023, 9, e20475. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Jiao, J.; Yang, L.; Wang, Y.; Jiang, W.; Yu, T.; et al. Advanced tumor organoid bioprinting strategy for oncology research. Mater Today Bio 2024, 28, 101198. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, J.; Xu, S.; Wang, J. 3D Bioprinting: An Important Tool for Tumor Microenvironment Research. Int J Nanomedicine 2023, 18, 8039–57. [Google Scholar] [CrossRef]
- Sun, D.; Gao, W.; Hu, H.; Zhou, S. Why 90% of clinical drug development fails and how to improve it? Acta Pharm Sin B 2022, 12, 3049–62. [Google Scholar] [CrossRef]
- Wang, H.; Brown, P.C.; Chow, E.C.Y.; Ewart, L.; Ferguson, S.S.; Fitzpatrick, S.; et al. 3D cell culture models: Drug pharmacokinetics, safety assessment, and regulatory consideration. Clin Transl Sci. 2021, 14, 1659–80. [Google Scholar] [CrossRef]
- Milner, E.; Ainsworth, M.; McDonough, M.; Stevens, B.; Buehrer, J.; Delzell, R.; et al. Emerging Three-Dimensional Hepatic Models in Relation to Traditional Two-Dimensional In Vitro Assays for Evaluating Drug Metabolism and Hepatoxicity. Medicine in Drug Discovery 2020, 8, 100060. [Google Scholar] [CrossRef]
- Kapałczyńska, M.; Kolenda, T.; Przybyła, W.; Zajączkowska, M.; Teresiak, A.; Filas, V.; et al. 2D and 3D cell cultures - a comparison of different types of cancer cell cultures. Arch Med Sci. 2018, 14, 910–9. [Google Scholar] [CrossRef]
- Close, D.A.; Johnston, P.A. Miniaturization and characterization of patient derived hepatocellular carcinoma tumor organoid cultures for cancer drug discovery applications. SLAS Discovery 2025, 30, 100201. [Google Scholar] [CrossRef]
- Zhao, Z.; Chen, X.; Dowbaj, A.M.; Sljukic, A.; Bratlie, K.; Lin, L.; et al. Organoids. Nature Reviews Methods Primers 2022, 2, 94. [Google Scholar] [CrossRef]
- Yang, H.; Sun, L.; Liu, M.; Mao, Y. Patient-derived organoids: a promising model for personalized cancer treatment. Gastroenterol Rep (Oxf) 2018, 6, 243–5. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Yuan, F.; Zuo, X.; Li, M. Breakthroughs and challenges of organoid models for assessing cancer immunotherapy: a cutting-edge tool for advancing personalised treatments. Cell Death Discov. 2025, 11, 222. [Google Scholar] [CrossRef] [PubMed]
- Gungor-Ozkerim, P.S.; Inci, I.; Zhang, Y.S.; Khademhosseini, A.; Dokmeci, M.R. Bioinks for 3D bioprinting: an overview. Biomater Sci. 2018, 6, 915–46. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.B.; Fazel Anvari-Yazdi, A.; Duan, X.; Zimmerling, A.; Gharraei, R.; Sharma, N.K.; et al. Biomaterials / bioinks and extrusion bioprinting. Bioactive Materials 2023, 28, 511–36. [Google Scholar] [CrossRef]
- Wang, H.; Yu, H.; Zhou, X.; Zhang, J.; Zhou, H.; Hao, H.; et al. An Overview of Extracellular Matrix-Based Bioinks for 3D Bioprinting. Front Bioeng Biotechnol. 2022, 10, 905438. [Google Scholar] [CrossRef]
- Johari, N.; Adabavazeh, Z.; Baino, F. PVA-based bioinks for 3D bioprinting: A comprehensive review of their applications in tissue engineering. Bioprinting 2025, 49, e00419. [Google Scholar] [CrossRef]
- Fang, L.; Liu, Y.; Qiu, J.; Wan, W. Bioprinting and its Use in Tumor-On-A-Chip Technology for Cancer Drug Screening: A Review. Int J Bioprint 2022, 8, 603. [Google Scholar] [CrossRef]
- Monteiro, M.V.; Zhang, Y.S.; Gaspar, V.M.; Mano, J.F. 3D-bioprinted cancer-on-a-chip: level-up organotypic in vitro models. Trends Biotechnol. 2022, 40, 432–47. [Google Scholar] [CrossRef]
- Shukla, A.K.; Yoon, S.; Oh, S.-O.; Lee, D.; Ahn, M.; Kim, B.S. Advancement in Cancer Vasculogenesis Modeling through 3D Bioprinting Technology. Biomimetics 2024, 9, 306. [Google Scholar] [CrossRef]
- Anari, E.; Anari, R.; Pazoki-Toroudi, H. A decade of scientific advancements and collaborations on CRISPR-Cas9 application in cancer research: a bibliometric review. Discov Oncol. 2025, 16, 1232. [Google Scholar] [CrossRef]
- Li, T.; Yang, Y.; Qi, H.; Cui, W.; Zhang, L.; Fu, X.; et al. CRISPR/Cas9 therapeutics: progress and prospects. Signal Transduction and Targeted Therapy 2023, 8, 36. [Google Scholar] [CrossRef] [PubMed]
- Chehelgerdi, M.; Chehelgerdi, M.; Khorramian-Ghahfarokhi, M.; Shafieizadeh, M.; Mahmoudi, E.; Eskandari, F.; et al. Comprehensive review of CRISPR-based gene editing: mechanisms, challenges, and applications in cancer therapy. Molecular Cancer 2024, 23, 9. [Google Scholar] [CrossRef] [PubMed]
- Shukla, A.K.; Gao, G.; Kim, B.S. Applications of 3D Bioprinting Technology in Induced Pluripotent Stem Cells-Based Tissue Engineering. Micromachines (Basel) 2022, 13(2). [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhou, G.; Jiang, Q.; Li, L. In situ 3D bioprinting: The future of regenerative medicine. In Fundamental Research; 2025. [Google Scholar]
- Mallya, D.; Gadre, M.A.; Varadharajan, S.; Vasanthan, K.S. 3D bioprinting for the construction of drug testing models-development strategies and regulatory concerns. Front Bioeng Biotechnol. 2025, 13, 1457872. [Google Scholar] [CrossRef]
- Cordeiro, S.; Oliveira, B.B.; Valente, R.; Ferreira, D.; Luz, A.; Baptista, P.V.; et al. Breaking the mold: 3D cell cultures reshaping the future of cancer research. Front Cell Dev Biol. 2024, 12, 1507388. [Google Scholar] [CrossRef]
- Ahmed, T. Biomaterial-based in vitro 3D modeling of glioblastoma multiforme. Cancer Pathogenesis and Therapy 2023, 1, 177–94. [Google Scholar] [CrossRef]
- Yamanaka, Y. CRISPR/Cas9 Genome Editing as a Strategy to Study the Tumor Microenvironment in Transgenic Mice. Methods Mol Biol. 2016, 1458, 261–71. [Google Scholar]
- Begagić, E.; Bečulić, H.; Đuzić, N.; Džidić-Krivić, A.; Pugonja, R.; Muharemović, A.; et al. CRISPR/Cas9-Mediated Gene Therapy for Glioblastoma: A Scoping Review. Biomedicines 2024, 12(1). [Google Scholar] [CrossRef]
- Slika, H.; Karimov, Z.; Alimonti, P.; Abou-Mrad, T.; De Fazio, E.; Alomari, S.; et al. Preclinical Models and Technologies in Glioblastoma Research: Evolution, Current State, and Future Avenues. International Journal of Molecular Sciences 2023, 24, 16316. [Google Scholar] [CrossRef]
- Lu, P.; Ruan, D.; Huang, M.; Tian, M.; Zhu, K.; Gan, Z.; et al. Harnessing the potential of hydrogels for advanced therapeutic applications: current achievements and future directions. Signal Transduction and Targeted Therapy 2024, 9, 166. [Google Scholar] [CrossRef]
- Porter, R.J.; Murray, G.I.; McLean, M.H. Current concepts in tumour-derived organoids. British Journal of Cancer 2020, 123, 1209–18. [Google Scholar] [CrossRef] [PubMed]
- Dave, R.; Pandey, K.; Patel, R.; Gour, N.; Bhatia, D. Leveraging 3D cell culture and AI technologies for next-generation drug discovery. Cell Biomaterials 2025, 1(3). [Google Scholar] [CrossRef]
- Stefanoudakis, D. Integrating CRISPR Technology with Key Genetic Markers in Pancreatic Cancer: A New Frontier in Targeted Therapies. SynBio 2025, 3, 1. [Google Scholar] [CrossRef]
- Seager, R.J.; Hajal, C.; Spill, F.; Kamm, R.D.; Zaman, M.H. Dynamic interplay between tumour, stroma and immune system can drive or prevent tumour progression. Converg Sci Phys Oncol. 2017, 3. [Google Scholar] [CrossRef]
- Kozlova, N.; Grossman, J.E.; Iwanicki, M.P.; Muranen, T. The Interplay of the Extracellular Matrix and Stromal Cells as a Drug Target in Stroma-Rich Cancers. Trends Pharmacol Sci. 2020, 41, 183–98. [Google Scholar] [CrossRef]
- El-Tanani, M.; Rabbani, S.A.; Babiker, R.; Rangraze, I.; Kapre, S.; Palakurthi, S.S.; et al. Unraveling the tumor microenvironment: Insights into cancer metastasis and therapeutic strategies. Cancer Letters 2024, 591, 216894. [Google Scholar] [CrossRef]
- Castaneda, M.; den Hollander, P.; Kuburich, N.A.; Rosen, J.M.; Mani, S.A. Mechanisms of cancer metastasis. Seminars in Cancer Biology 2022, 87, 17–31. [Google Scholar] [CrossRef]
- Liu, Q.; Zhang, H.; Jiang, X.; Qian, C.; Liu, Z.; Luo, D. Factors involved in cancer metastasis: a better understanding to "seed and soil" hypothesis. Mol Cancer 2017, 16, 176. [Google Scholar] [CrossRef]
- Shi, X.; Wang, X.; Yao, W.; Shi, D.; Shao, X.; Lu, Z.; et al. Mechanism insights and therapeutic intervention of tumor metastasis: latest developments and perspectives. Signal Transduction and Targeted Therapy 2024, 9, 192. [Google Scholar] [CrossRef]
- Khawar, M.B.; Wang, Y.; Majeed, A.; Afzal, A.; Haneef, K.; Sun, H. Mini-organs with big impact: Organoids in liver cancer studies. Oncol Res. 2023, 31, 677–88. [Google Scholar] [CrossRef]
- Hu, Y.; Peng, Z.; Qiu, M.; Xue, L.; Ren, H.; Wu, X.; et al. Developing biotechnologies in organoids for liver cancer. Biomedical Technology 2025, 9, 100067. [Google Scholar] [CrossRef]
- Peng, Z.; Lv, X.; Sun, H.; Zhao, L.; Huang, S. 3D tumor cultures for drug resistance and screening development in clinical applications. Mol Cancer 2025, 24, 93. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, A.; Saga, K.; Hisatomi, Y.; Li, Y.; Kaneda, Y.; Nimura, K. A simple method using CRISPR-Cas9 to knock-out genes in murine cancerous cell lines. Sci Rep. 2020, 10, 22345. [Google Scholar] [CrossRef] [PubMed]
- Imtiaz, S.; Ferdous, U.T.; Nizela, A.; Hasan, A.; Shakoor, A.; Zia, A.W.; et al. Mechanistic study of cancer drug delivery: Current techniques, limitations, and future prospects. European Journal of Medicinal Chemistry 2025, 290, 117535. [Google Scholar] [CrossRef]
- Wang, X.-H.; Wang, W.-Y.; Sun, Z.-J. Immune organoid for cancer immunotherapy. Acta Pharmaceutica Sinica B 2025, 15, 3419–35. [Google Scholar] [CrossRef]
- Alves, E.; Taifour, S.; Dolcetti, R.; Chee, J.; Nowak, A.K.; Gaudieri, S.; et al. Reprogramming the anti-tumor immune response via CRISPR genetic and epigenetic editing. Mol Ther Methods Clin Dev. 2021, 21, 592–606. [Google Scholar] [CrossRef]
- Al-Kabani, A.; Huda, B.; Haddad, J.; Yousuf, M.; Bhurka, F.; Ajaz, F.; et al. Exploring Experimental Models of Colorectal Cancer: A Critical Appraisal from 2D Cell Systems to Organoids, Humanized Mouse Avatars, Organ-on-Chip, CRISPR Engineering, and AI-Driven Platforms—Challenges and Opportunities for Translational Precision Oncology. Cancers 2025, 17, 2163. [Google Scholar]
- Gnatowski, P.; Piłat, E.; Kucińska-Lipka, J.; Saeb, M.R.; Hamblin, M.R.; Mozafari, M. Recent advances in 3D bioprinted tumor models for personalized medicine. Transl Oncol. 2023, 37, 101750. [Google Scholar] [CrossRef]
- Ireson, C.R.; Alavijeh, M.S.; Palmer, A.M.; Fowler, E.R.; Jones, H.J. The role of mouse tumour models in the discovery and development of anticancer drugs. British Journal of Cancer 2019, 121, 101–8. [Google Scholar] [CrossRef]
- Zhang, W.; Ding, Y.; He, H.; Chen, K.; Zeng, Q.; Cao, X.; et al. Prospects and challenges of ovarian cancer organoids in chemotherapy research (Review). Oncol Lett. 2025, 29, 198. [Google Scholar] [CrossRef]
- Marei, H.E. Stem cell therapy: a revolutionary cure or a pandora's box. Stem Cell Res Ther. 2025, 16, 255. [Google Scholar] [CrossRef] [PubMed]
- Sadiq, I.Z.; Abubakar, F.S.; Katsayal, B.S.; Ibrahim, B.; Adamu, A.; Usman, M.A.; et al. Stem cells in regenerative medicine: Unlocking therapeutic potential through stem cell therapy, 3D bioprinting, gene editing, and drug discovery. Biomedical Engineering Advances 2025, 9, 100172. [Google Scholar] [CrossRef]
- Chehelgerdi, M.; Behdarvand Dehkordi, F.; Chehelgerdi, M.; Kabiri, H.; Salehian-Dehkordi, H.; Abdolvand, M.; et al. Exploring the promising potential of induced pluripotent stem cells in cancer research and therapy. Mol Cancer 2023, 22, 189. [Google Scholar] [CrossRef] [PubMed]
- Eskandar, K. 3D bioprinting for facial reconstruction: Advances and challenges. Regenesis Repair Rehabilitation 2025, 1, 16–23. [Google Scholar] [CrossRef]
- Gopalakrishnan, M.; Kannan, D.; Elumalai, K.; Karunakar, K.; Jayaraj, S.; Devaraji, M.; et al. Advanced 3D biomaterials and bioprinting strategies for in vitro modeling of neurodegenerative diseases. Biomedical Technology 2025, 11, 100089. [Google Scholar] [CrossRef]
- Hoang, V.T.; Nguyen, Q.T.; Phan, T.T.K.; Pham, T.H.; Dinh, N.T.H.; Anh, L.P.H.; et al. Tissue Engineering and Regenerative Medicine: Perspectives and Challenges. MedComm (2020) 2025, 6, e70192. [Google Scholar] [CrossRef]
- Ayanoğlu, F.B.; Elçin, A.E.; Elçin, Y.M. Bioethical issues in genome editing by CRISPR-Cas9 technology. Turk J Biol. 2020, 44, 110–20. [Google Scholar] [CrossRef]
- Farag, V.E.; Devey, E.A.; Leong, K.W. The Interface of Gene Editing with Regenerative Medicine. Engineering 2025, 46, 73–100. [Google Scholar] [CrossRef]
- Brokowski, C.; Adli, M. CRISPR Ethics: Moral Considerations for Applications of a Powerful Tool. J Mol Biol. 2019, 431, 88–101. [Google Scholar] [CrossRef]
- Jose, A.; Kulkarni, P.; Thilakan, J.; Munisamy, M.; Malhotra, A.G.; Singh, J.; et al. Integration of pan-omics technologies and three-dimensional in vitro tumor models: an approach toward drug discovery and precision medicine. Molecular Cancer 2024, 23, 50. [Google Scholar] [CrossRef]
- Qu, S.; Xu, R.; Yi, G.; Li, Z.; Zhang, H.; Qi, S.; et al. Patient-derived organoids in human cancer: a platform for fundamental research and precision medicine. Mol Biomed. 2024, 5, 6. [Google Scholar] [CrossRef]
- Lv, J.; Du, X.; Wang, M.; Su, J.; Wei, Y.; Xu, C. Construction of tumor organoids and their application to cancer research and therapy. Theranostics 2024, 14, 1101–25. [Google Scholar] [CrossRef] [PubMed]
- Khorsandi, D.; Yang, J.-W.; Foster, S.; Khosravi, S.; Hosseinzadeh Kouchehbaghi, N.; Zarei, F.; et al. Patient-Derived Organoids as Therapy Screening Platforms in Cancer Patients. Advanced Healthcare Materials 2024, 13, 2302331. [Google Scholar] [CrossRef] [PubMed]
- Parvin, N.; Joo, S.W.; Jung, J.H.; Mandal, T.K. Multimodal AI in Biomedicine: Pioneering the Future of Biomaterials, Diagnostics, and Personalized Healthcare. Nanomaterials 2025, 15, 895. [Google Scholar] [CrossRef] [PubMed]
- Imani, S.; Li, X.; Chen, K.; Maghsoudloo, M.; Jabbarzadeh Kaboli, P.; Hashemi, M.; et al. Computational biology and artificial intelligence in mRNA vaccine design for cancer immunotherapy. Front Cell Infect Microbiol. 2024, 14, 1501010. [Google Scholar] [CrossRef]
- Wang, H.; Li, X.; You, X.; Zhao, G. Harnessing the power of artificial intelligence for human living organoid research. Bioactive Materials 2024, 42, 140–64. [Google Scholar] [CrossRef]
- Ravichandran, M.; Maddalo, D. Applications of CRISPR-Cas9 for advancing precision medicine in oncology: from target discovery to disease modeling. Front Genet. 2023, 14, 1273994. [Google Scholar] [CrossRef]
- Li, T.; Li, S.; Kang, Y.; Zhou, J.; Yi, M. Harnessing the evolving CRISPR/Cas9 for precision oncology. Journal of Translational Medicine 2024, 22, 749. [Google Scholar] [CrossRef]
- Cao, R.; Fomina, A.; McGuigan, A.P. Tissue-engineered Cancer Models in Drug Screening. In Biomaterial Based Approaches to Study the Tumour; Winter, J.O., Rao, S., Eds.; The Royal Society of Chemistry: Microenvironment, 2022; p. 0. [Google Scholar]
Figure 1.
(A) Schematic representation of the CRISPR/Cas9 system and its mechanism of action. The CRISPR/Cas9 complex consists of a Cas9 endonuclease guided by a single-guide RNA (sgRNA), which directs the complex to a complementary DNA sequence adjacent to a 3′ protospacer adjacent motif (PAM). Upon recognition and binding, Cas9 induces a double-stranded DNA break (DSB), which is subsequently repaired through either the error-prone non-homologous end joining (NHEJ) pathway or the more precise homology-directed repair (HDR) mechanism [30]. (B) Emerging strategies for cancer-specific gene editing using CRISPR/Cas9. Targeted delivery of CRISPR/Cas9 to cancer cells has been achieved using a variety of vectors, including adeno-associated viruses (AAVs) with tissue-specific tropism, antibody- or aptamer-conjugated liposomes, tumor-selective oncolytic viruses, and extracellular vesicles engineered with surface ligands derived from donor cells. Cancer-specific expression of CRISPR/Cas9 components can be regulated via: (i) miRNA switches responsive to tumor-specific miRNAs that control Cas9 mRNA stability; (ii) logic gate systems using dual promoters specific to both cancer type and tissue-of-origin to independently regulate components of the system; and (iii) telomerase-dependent promoters enabling Cas9 expression predominantly in highly proliferative, telomerase-active tumor tissues. Therapeutic gene targets include those involved in synthetic lethality—wherein disruption of gene pairs selectively induces cancer cell death—and oncogenes derived from tumor-promoting viruses [31]. (C) Overview of non-viral delivery platforms for CRISPR/Cas9-mediated gene editing. This panel summarizes various non-viral carriers designed for loading and intracellular transport of CRISPR/Cas9 components, highlighting their potential to overcome limitations associated with viral vectors [32]. (D) Mechanistic insights into stimuli-responsive nanoformulations for CRISPR/Cas9 delivery. This panel illustrates how responsive nanocarriers enable controlled release and activation of CRISPR/Cas9 machinery in response to specific stimuli (e.g., pH, redox, and enzymatic), thereby enhancing gene editing specificity and efficiency within target cells [32]. This section is adapted from under the Creative Commons Attribution 4.0 (CC BY 4.0) license.
Figure 1.
(A) Schematic representation of the CRISPR/Cas9 system and its mechanism of action. The CRISPR/Cas9 complex consists of a Cas9 endonuclease guided by a single-guide RNA (sgRNA), which directs the complex to a complementary DNA sequence adjacent to a 3′ protospacer adjacent motif (PAM). Upon recognition and binding, Cas9 induces a double-stranded DNA break (DSB), which is subsequently repaired through either the error-prone non-homologous end joining (NHEJ) pathway or the more precise homology-directed repair (HDR) mechanism [30]. (B) Emerging strategies for cancer-specific gene editing using CRISPR/Cas9. Targeted delivery of CRISPR/Cas9 to cancer cells has been achieved using a variety of vectors, including adeno-associated viruses (AAVs) with tissue-specific tropism, antibody- or aptamer-conjugated liposomes, tumor-selective oncolytic viruses, and extracellular vesicles engineered with surface ligands derived from donor cells. Cancer-specific expression of CRISPR/Cas9 components can be regulated via: (i) miRNA switches responsive to tumor-specific miRNAs that control Cas9 mRNA stability; (ii) logic gate systems using dual promoters specific to both cancer type and tissue-of-origin to independently regulate components of the system; and (iii) telomerase-dependent promoters enabling Cas9 expression predominantly in highly proliferative, telomerase-active tumor tissues. Therapeutic gene targets include those involved in synthetic lethality—wherein disruption of gene pairs selectively induces cancer cell death—and oncogenes derived from tumor-promoting viruses [31]. (C) Overview of non-viral delivery platforms for CRISPR/Cas9-mediated gene editing. This panel summarizes various non-viral carriers designed for loading and intracellular transport of CRISPR/Cas9 components, highlighting their potential to overcome limitations associated with viral vectors [32]. (D) Mechanistic insights into stimuli-responsive nanoformulations for CRISPR/Cas9 delivery. This panel illustrates how responsive nanocarriers enable controlled release and activation of CRISPR/Cas9 machinery in response to specific stimuli (e.g., pH, redox, and enzymatic), thereby enhancing gene editing specificity and efficiency within target cells [32]. This section is adapted from under the Creative Commons Attribution 4.0 (CC BY 4.0) license.
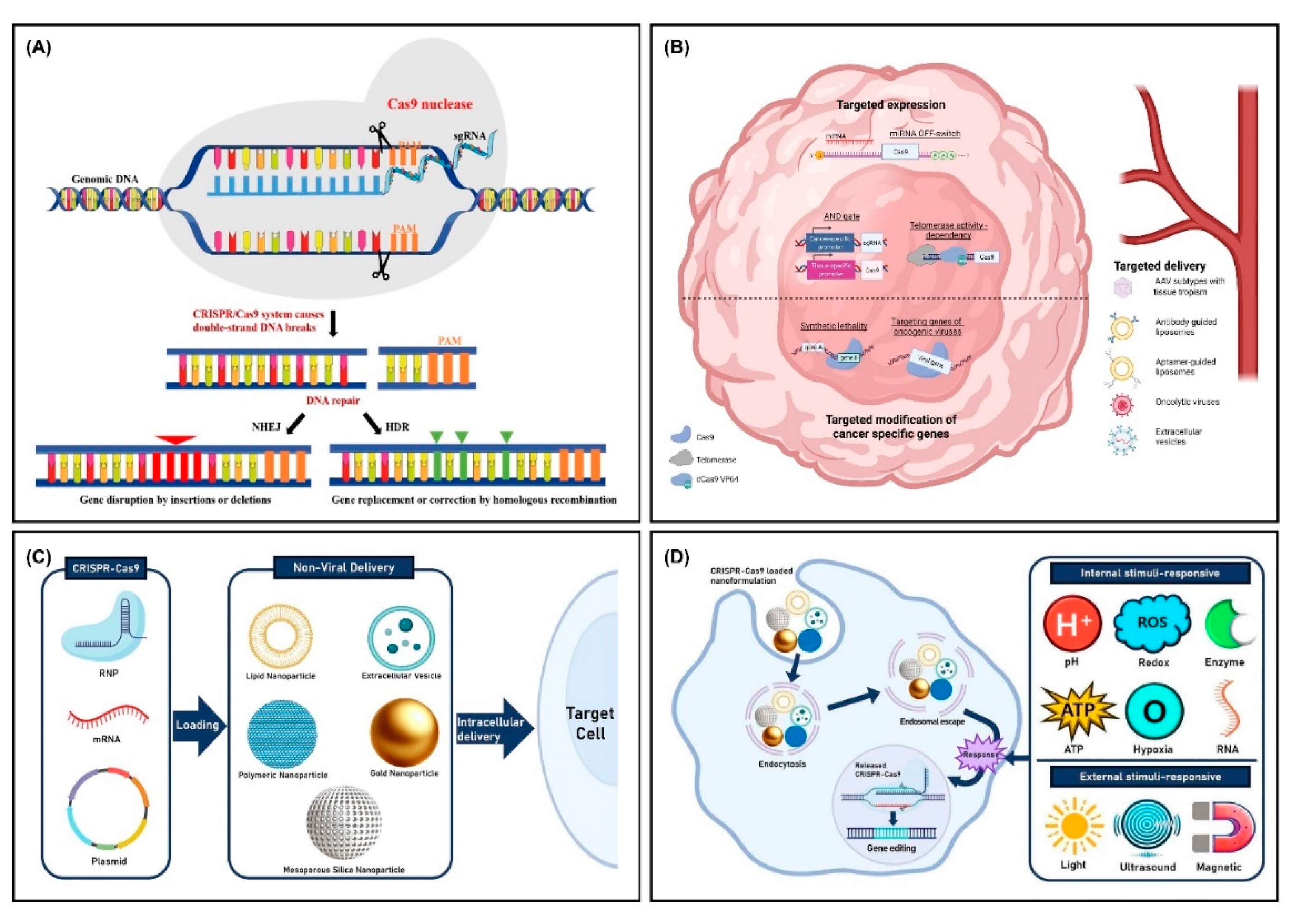
Figure 2.
Immersion 3D bioprinting of cancer organoids for high-throughput drug screening using patient-derived cancer cells.
Figure 2.
Immersion 3D bioprinting of cancer organoids for high-throughput drug screening using patient-derived cancer cells.
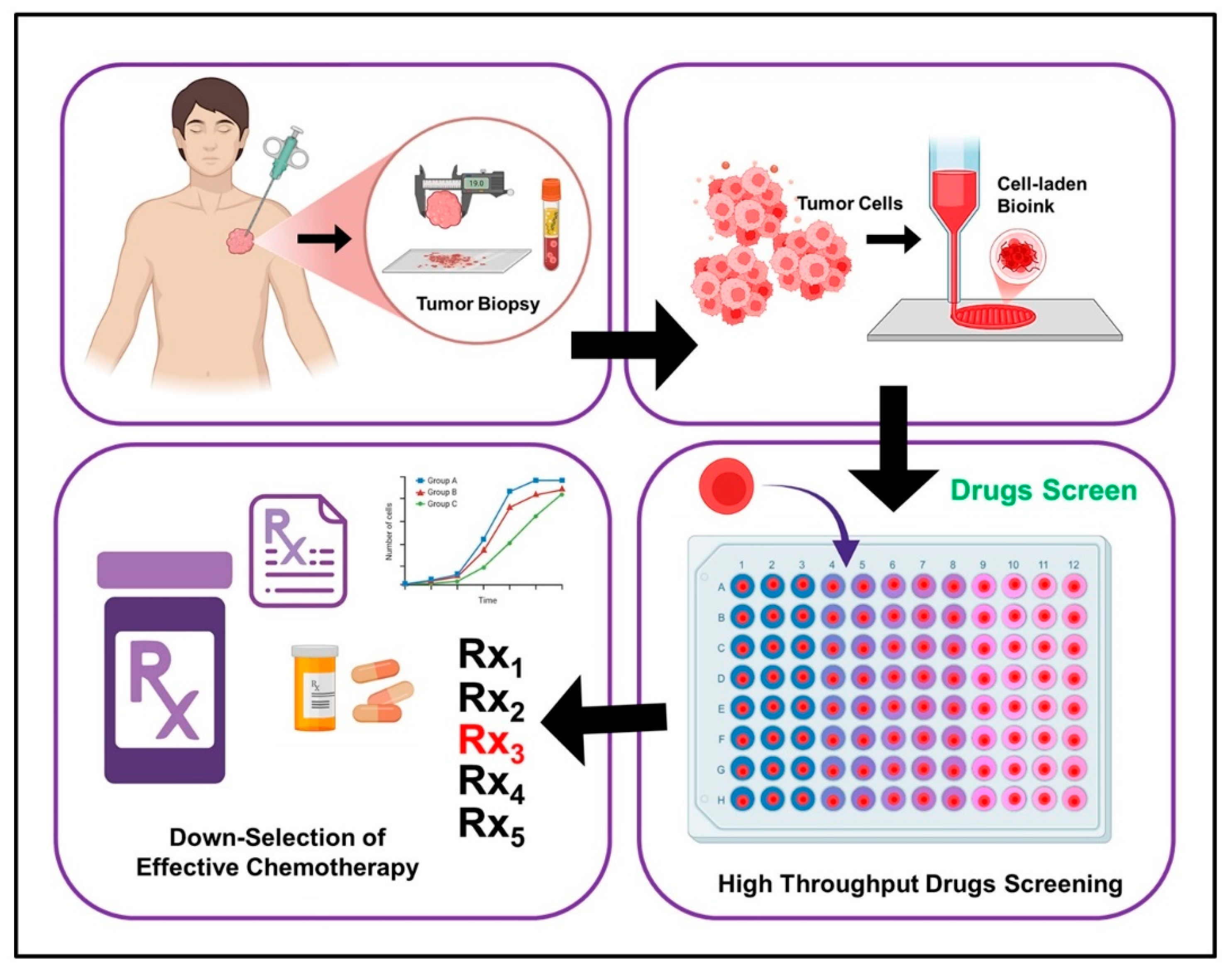
Figure 3.
Schematic workflow of an integrated CRISPR-Cas9 and 3D bioprinting approach for genome-wide screening in breast cancer organoids. A tumor biopsy obtained from a breast cancer patient is utilized to derive cancer cells for downstream applications. A human genome-wide CRISPR/Cas9 knock-out library containing single-guide RNAs (sgRNAs) is packaged into lentiviral vectors and transduced into Cas9-overexpressing breast cancer cells to generate a pool of mutant cells. These genetically modified cells are subjected to selection, followed by treatment with chemotherapeutic agents or DMSO as a control. Using a 3D bioprinting platform, the mutant breast cancer cells are biofabricated into organoid structures to better recapitulate the tumor microenvironment for drug screening. Post-treatment, genomic DNA is extracted from the organoids, and the integrated sgRNA sequences are amplified via PCR. High-throughput next-generation sequencing (NGS) is performed to identify sgRNA enrichment or depletion, enabling genome-wide screening. Bioinformatic analysis is subsequently conducted to identify mutations, including those in BRCA1/2, to inform personalized therapeutic strategies for breast cancer patients.
Figure 3.
Schematic workflow of an integrated CRISPR-Cas9 and 3D bioprinting approach for genome-wide screening in breast cancer organoids. A tumor biopsy obtained from a breast cancer patient is utilized to derive cancer cells for downstream applications. A human genome-wide CRISPR/Cas9 knock-out library containing single-guide RNAs (sgRNAs) is packaged into lentiviral vectors and transduced into Cas9-overexpressing breast cancer cells to generate a pool of mutant cells. These genetically modified cells are subjected to selection, followed by treatment with chemotherapeutic agents or DMSO as a control. Using a 3D bioprinting platform, the mutant breast cancer cells are biofabricated into organoid structures to better recapitulate the tumor microenvironment for drug screening. Post-treatment, genomic DNA is extracted from the organoids, and the integrated sgRNA sequences are amplified via PCR. High-throughput next-generation sequencing (NGS) is performed to identify sgRNA enrichment or depletion, enabling genome-wide screening. Bioinformatic analysis is subsequently conducted to identify mutations, including those in BRCA1/2, to inform personalized therapeutic strategies for breast cancer patients.
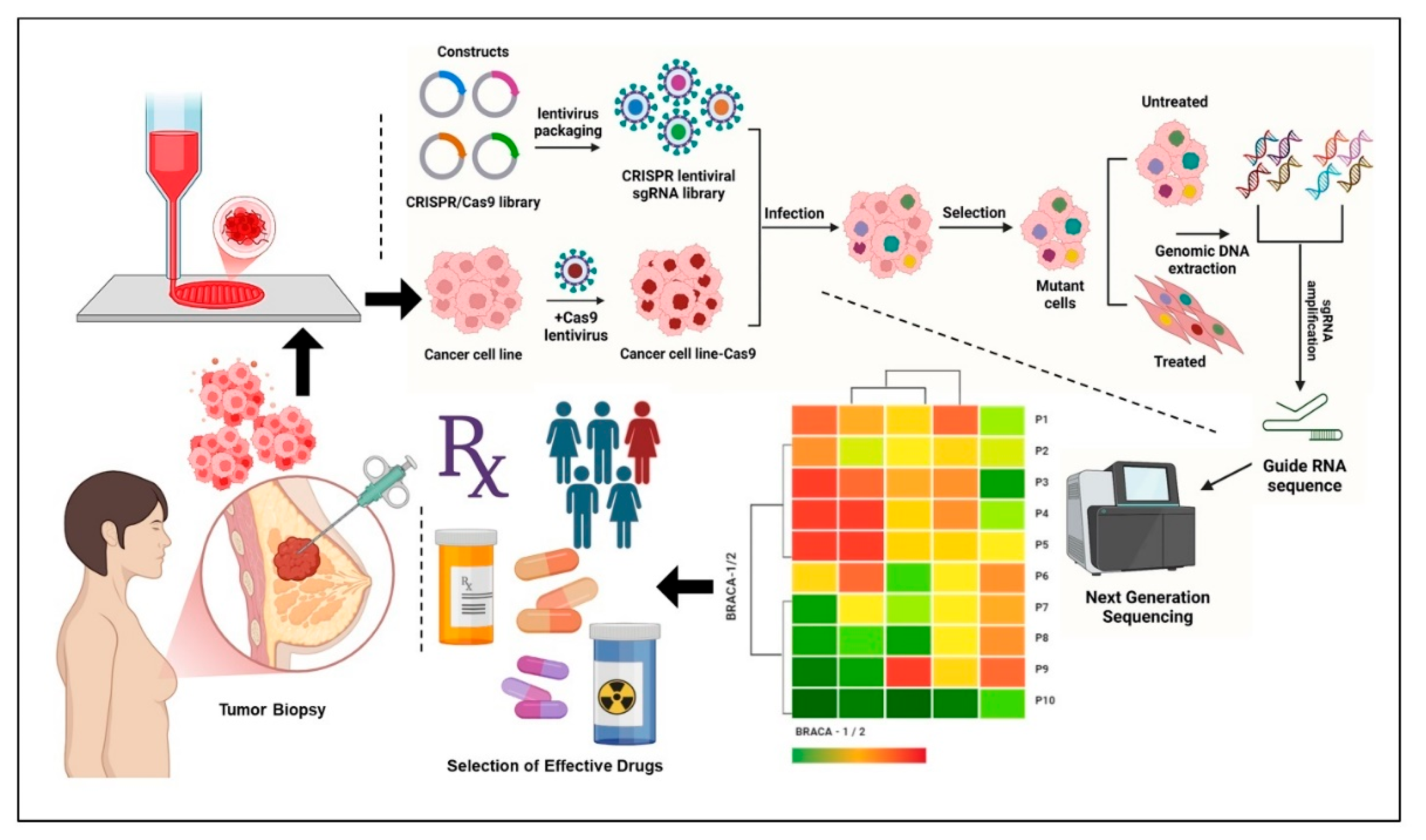
Table 1.
Strategies to improve CRISPR/Cas9 specificity and reduce off-target effects.
| Strategy | Mechanism/Approach | Advatages |
|---|---|---|
| In silico sgRNA design | Bioinformatics tools to predict and avoid off-target sequences | Reduces the likelihood of off-target cleavage |
| High-fidelity Cas9 variants (e.g., eSpCas9) | Engineered Cas9 with reduced non-specific binding | Minimizes unintended genome editing |
| Paired Cas9 nickases | Two Cas9 nickases targeting opposite strands, creating DSB only at the intended site | Decreases off-target DSBs and indels |
| Truncated sgRNAs | Shortened guide RNAs to reduce off-target binding | Enhances specificity without sacrificing on-target efficiency |
| Genome-wide off-target screening | Techniques like GUIDE-seq, CIRCLE-seq, and Digenome-seq | Validates safety before clinical application |
Table 2.
Functional modifications of the Cas9 protein enabling targeted DNA sequence manipulation without inducing double-strand breaks (DSBs) [33,36,37,38,39,40].
| Cas9 Modification | Application |
|---|---|
| Cas9 nickase | Introduces a single-strand break in DNA; using a pair of nickases to target opposite strands enhances specificity and reduces off-target mutations. |
| deadCas9 (dCas9) | Lacks nuclease activity; functions as a programmable DNA-binding scaffold to recruit regulatory proteins or effector molecules. |
| dCas9 with KRAB domain | Inhibits gene expression by recruiting transcriptional repressors, silencing downstream gene activity. |
| dCas9 with VP64 | Activates transcription of downstream genes by recruiting transcriptional activators. |
| dCas9 with methylases/demethylases or histone deacetylases | Modulates epigenetic marks at target loci, enabling control of gene expression through epigenetic regulation. |
|
Cas9 nickase with adenine or cytosine deaminase (Base editors; BEs) |
Allows precise base transitions (e.g., A→G, C→T) without introducing DSBs, offering efficient and accurate gene correction. |
|
Cas9 nickase with a reverse transcription domain (Prime editors; PEs) |
Permits the insertion, deletion, or substitution of specific DNA sequences with high precision and without generating DSBs. |
| Delivery Type | Method | Advantages | Disadvantages |
|---|---|---|---|
| Capsule | Viral vector | - High delivery efficiency - Broad range of available viral systems | - Immunogenic response - Potential pathogenicity - Genome integration risk |
| AAV (Adeno-associated virus) | - Low immunogenicity - Non-integrating nature - Tissue-specific tropism via serotypes | - Low packaging capacity | |
| Lipid-based vector | - Customizable surface modifications for targeted delivery | - Low transfection efficiency - Possible cytotoxicity | |
| Cas9 Form | Plasmid DNA | - Stable and long-term expression - Easy to produce | - Slow onset of gene editing - Requires nuclear entry |
| mRNA | - Rapid onset of editing - Transient expression reduces off-target effects | - Unstable and rapidly degraded - Requires cold-chain storage | |
| Protein (Cas9 RNP complex) | - Immediate gene editing upon delivery - Lower risk of off-target effects due to short activity window | - Difficult intracellular delivery - Risk of bacterial contaminants |
Table 5.
Ethical and Regulatory Considerations in CRISPR-Cas9 and 3D Bioprinting Integration.
| Consideration | Description |
|---|---|
| Germline Editing | Potential for heritable genetic modifications in bioprinted reproductive tissues |
| Human Enhancement | Ethical debate over non-therapeutic gene edits for performance or appearance |
| Chimeric Constructs | Concerns over creating human-animal hybrids with cognitive functions |
| Informed Consent and Data Privacy | Challenges in obtaining consent for the use of edited tissues and genomic data |
| Regulatory Oversight | Lack of clear, unified regulations for combined bioprinting and gene editing |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



