Submitted:
29 January 2026
Posted:
03 February 2026
You are already at the latest version
Abstract
Abstract Cockroaches are a promising biological tool for processing biodegradable waste due to their resilience and ability to consume a wide range of organic material. Through their metabolism, they convert organic material into a humus-like substrate known as frass or blatticompost, which can potentially be used as an agricultural fertiliser. This study investigates the ability of the cockroach species Blaptica dubia, Blatta lateralis and Blaberus giganteus to process slaughterhouse by-products and compares their biocon-version efficiency, nutrient composition, chemical properties and microbiological pro-file of the resulting frass as a compost with that of cockroaches fed on conventional biowaste. By comparing these properties with conventional composting methods and the relevant literature, this research aims to provide new insights into the efficiency of blatticomposting as an alternative organic waste management strategy. The results suggest that cockroaches can effectively break down organic material, significantly reducing the volume of waste while producing compost with favorable chemical and microbiological properties. Furthermore, the inclusion of slaughterhouse waste in the process extends the potential application of blatticomposting to protein-rich organic residues that are otherwise difficult to manage sustainably. This study contributes to this topic by exploring the feasibility of integrating blatticomposting into existing waste management systems, highlighting both the benefits and the necessary precautions in terms of biosecurity and public health risks. Proper containment and monitoring of cockroach populations and risk assessment for pathogen transmission are key consid-erations for the practical implementation of this approach. The absence of hazardous levels of heavy metals and pathogenic microorganisms indicates that cock-roach-mediated bioconversion can produce a chemically stable and microbiologically safe organic amendment.
Keywords:
insects
; waste
; processing
; frass
; blatticomposting
1. Introduction
Conventional composting is an aerobic biological process that converts organic material into nutrient rich fertiliser or mulch through natural decomposition. The resulting compost is a dark, crumbly material with a distinctive earthy odour. Compost is typically characterised by a stabilised organic matter fraction rich in humic substances and essential plant nutrients, including nitrogen, phosphorus and potassium [1,2]. Microorganisms break down the organic substrate, utilising carbon and nitrogen for growth and reproduction, water as a medium for metabolism, and oxygen for aerobic respiration [3]. During composting, carbon is progressively mineralised while nutrients such as N, P and K are concentrated in the remaining solid phase, determining the final chemical composition of compost [4,5]. However, not all organic materials are suitable for conventional composting. Weed seeds, severely diseased plants and certain slaughterhouse waste, such as meat products contaminated with pathogens or toxins, are not suitable due to the risk of microbial contamination and environmental toxicity. Animal derived waste streams often exhibit high nitrogen and fat contents, which can negatively affect compost stability, pH and nutrient balance if not properly managed [1,6]. These materials can release noxious odours, attract rodents and other pests, and pose a threat to human health and ecological integrity [7]. Alternative composting methods, such as entomocomposting, which refers to the use of insects for the biological digestion of organic substrates, represent an advanced stage in the evolution of composting technologies. Initially developed to produce organic fertilisers, entomocomposting has gained increasing relevance as a strategy for the safe management of rapidly growing organic waste streams, while simultaneously enabling insect biomass production for food and feed. The efficiency and quality of the resulting frass depend on the insect species and substrate interaction and composting conditions, which influence nutrient composition and potential contamination risks [8]. Consequently, appropriate species selection, process control, and regulatory frameworks are essential to ensure the safe and effective application of entomocomposting systems, which have recently transitioned from small-scale applications to industrial implementation. Well-established examples of such entomocomposting systems include vermicomposting and black soldier fly (Hermetia illucens) larvae composting, which utilise the biological capabilities of certain organisms to accelerate the decomposition of organic waste, offering sustainable and efficient solutions for waste management and resource recovery. Vermicomposting uses earthworms such as Lumbricus rubellus, Eisenia andrei and Eisenia fetida to efficiently decompose organic waste into nutrient-rich compost, improving soil fertility and structure faster than conventional composting methods [9,10,11,12]. The larvae of the black soldier fly efficiently decompose various organic wastes, such as food waste, by-products of the agricultural industry, and poultry or dairy manure, and reduce the original waste mass by around 50% in less time than conventional composting [13,14]. The agronomic value of compost is primarily determined by its chemical composition, including total nutrient concentrations, organic carbon content and C:N ratio [2,5]. Vermicompost and black soldier fly (BSF) compost, also known as frass, are high-quality products that are rich in nutrients and beneficial microorganisms. They significantly improve soil fertility and promote plant growth. Vermicompost contains between 1.5% and 3% total nitrogen, 1% to 2% phosphorus and 1% to 2.5% potassium [10]. Such nutrient ranges are commonly used as reference benchmarks for evaluating the fertiliser potential of novel compost materials [2,6]. In addition to the macronutrients, it is rich in micronutrients such as zinc, iron and manganese, which are important for plant growth. It also harbours beneficial microorganisms, including bacteria and fungi, which promote the decomposition of organic matter and improve the biological activity of the soil. According to [15], the application of vermicompost can increase plant growth by 20% to 40% compared to non-composted material. On the other hand, BSF frass as a compost material is characterised by rapid decomposition of organic waste and a high nutrient content, containing between 1% and 2% nitrogen, 0.6% to 1.2% phosphorus and 0.8% to 2% potassium [16]. Insect derived composts generally exhibit comparable macronutrient concentrations to conventional composts, although their chemical composition may vary depending on substrate and processing conditions [17,18]. BSF composting not only improves soil biological activity and has a high-water retention capacity, making it particularly suitable for dry and degraded soils. BSF composting also produces protein-rich larvae suitable for animal feed, making this method both economically viable and environmentally sustainable [19]. Similar to earthworms and BSF, cockroaches can convert organic waste into valuable products while contributing to nutrient cycling. Their biological traits make them particularly effective for bioconversion, forming the basis of blatticomposting systems. Owing to their minimal trophic specificity, adaptability to different environmental conditions, rapid reproduction, voracious feeding behaviour, tolerance of high population densities and independence from light, cockroaches are proving to be new and effective means of bioremediating organic waste. Blatticomposting, also known as cockroach composting, is an organic waste management process that utilises certain cockroach species to break down organic waste and turn it into nutrient-rich compost (frass). This innovative waste management strategy offers a sustainable alternative to conventional composting, vermicomposting (e.g., using earthworms) or black soldier fly (BSF) composting (using Hermetia illucens larvae) and is attracting increasing scientific interest due to its ecological efficiency and scalability [20,21,22].
Cockroaches (Blattodea) are globally distributed insects with a crucial ecological role as decomposers of organic material. Although they are often perceived as pests, they thrive in a variety of habitats, particularly in tropical and subtropical regions, but are also found in other climates. Of the approximately 4,000 species described, most live in forest ecosystems, where they contribute significantly to litter decomposition, while some live in caves, semi-aquatic environments or in association with other insects. Cockroaches are primarily nocturnal and herbivorous but can adapt their feeding habits to environmental conditions [23,24]. Morphologically, cockroaches are oval-shaped, dorsoventrally flattened insects with a length of 10 to 50 mm. They typically have large eyes, long filiform antennae and chewing mouthparts that enable them to consume a wide range of organic material, including food waste, paper and even other insects [25]. Their mouthparts, labrum, mandibles, maxillae and labium are extremely flexible and adapted to different diets. Most importantly, cockroaches can regenerate damaged mouthparts during moulting, which increases their resilience and adaptability. Most adult cockroaches have wings; the forewings are usually hardened and overlap when closed. Their legs are spiny and have a five limbed tarsus, which contributes to their agility [26]. Complementing these physical adaptations, cockroaches harbor diverse microbial communities in their gastrointestinal tracts that enhance digestion of these varied substrates. These gut microbes facilitate food breakdown and potentially offer protection against pathogenic organisms, while producing key digestive enzymes such as cellulase, proteases, amylases, phosphatases, urease, glutamate dehydrogenase, and lipases. These enzymes enable the degradation of resilient components like lignocellulosic materials and proteinaceous compounds, such as slaughterhouse waste, relevant to bioconversion applications. Cockroaches acquire these bacteria from their surroundings through feeding on diverse organic matter, supporting efficient waste processing [27]. Ecologically, cockroaches play a key role in the nutrient cycle by decomposing dead plant and animal matter. However, urban species such as Blatta orientalis or Blattella germanica can act as disease vectors and transmit pathogens to food. They are also an important food source for birds, reptiles and spiders, which adds to their importance in food webs. In terms of feeding behaviour, cockroaches are classified as detritivores or omnivores. Most species feed on decomposing organic matter, facilitating nutrient recycling, while others eat both plant and animal material. An important behavioural adaptation is their nocturnal activity, which helps them evade predators and avoid dehydration. They typically seek refuge in dark, moist crevices, optimising their survival in different environments [23]. Although commonly viewed as pests, the vast majority of cockroach species are completely harmless to humans. They live in the wild and feed mostly on leaf litter and various other dead organic materials. In tropical and Mediterranean regions, cockroaches also play an immense role in the nutrient cycle as they eat leaves and wood and convert them into soil. Furthermore, [21] and [22] summarise the most suitable cockroach species that are suitable for composting: (1) Ivory-headed cockroach (Eublaberus sp. Ivory), which is characterised by its size and distinctive markings. These cockroaches are gregarious, reproduce quickly and are effective at processing organic waste; (2) Six-spotted cockroach (Eublaberus distanti), which is typically found in South American caves. It thrives in crowded conditions and consumes a wide range of organic materials, including decaying organic matter; (3) Pantanal cockroach (Eublaberus serranus), which is native to the Pantanal wetland region of Brazil. These cockroaches favour humid environments and contribute effectively to the composting process; (4) Orange head cockroach (Nauphoeta cinerea) known for its adaptability, this cockroach species efficiently digests a variety of organic waste, making it a popular choice for composting; (5) Surinam cockroach (Pycnoscelus surinamensis) is a well-known species that can process various organic materials, and its gut microbiota helps to break down tough organic material; (6) Dubia cockroach (Blaptica dubia), which is often used as a model species. It is easy to grow and has a high reproduction rate, which makes it favourable for composting plants. These species are known for their ability to digest a variety of organic wastes, making them valuable helpers in the composting process. While the ecological role of cockroaches in organic matter decomposition and nutrient cycling is well established, available evidence suggests that associated greenhouse gas emissions, particularly methane, are minimal and have received considerably less attention in the literature. [28] and [29] established that methane emissions from insects are not universal but depend on taxonomic affiliation, gut morphology, microbial symbiosis, and diet. Methane production is mediated by anaerobic hindgut microorganisms rather than by the insect host itself. In cockroaches, these methanogenic microbes are part of a broader symbiotic community (e.g., Blattabacterium spp.) that also contributes to efficient nitrogen recycling by converting uric acid into amino acids, thereby improving dietary protein utilisation [29]. As a result, methane emissions from cockroaches are species-specific and influenced by feeding strategy, diet composition, and gut microbial structure; protein-rich or omnivorous diets, such as those in slaughterhouse waste bioconversion, may further suppress methanogenesis. For instance, Blaptica dubia has been reported to emit approximately 1.4 g CH₄ per kg body mass gain, which is substantially lower than values reported for other methanogenic detritivores and far below those of ruminant livestock [30]. Environmental factors, including temperature and developmental stage, further contribute to emission variability. Although methane emissions have been detected in some cockroach species (e.g., Blaptica dubia), available data indicate that their overall contribution to greenhouse gas emissions is low when compared with conventional livestock systems, especially ruminants, suggesting that blatticomposting may offer a comparatively climate-efficient option for the bioconversion of slaughterhouse waste [28]. According to the literature, the blatticomposting process consists of several important steps that begin with the careful selection of suitable cockroach species capable of efficient organic waste bioconversion. Certain species are characterised by a remarkable composting ability, as they are able to efficiently digest a wide range of organic waste materials. Among these, Blaptica dubia stands out, having demonstrated high efficiency in processing leaf litter into nutrient-rich frass across different developmental instars, with larger individuals showing consumption rates up to 30 mg dry weight per day and assimilation efficiencies of 10-21%, highlighting their potential beyond plant-based wastes to protein-rich substrates like slaughterhouse by-products [24]. This proven capacity with plant-based lignocellulosic materials establishes Blaptica dubia as an excellent candidate for extending blatticomposting to more challenging, protein-rich substrates like slaughterhouse by-products, as investigated in this study. This species is an ideal model organism for experimental set-ups due to its ease of cultivation, high reproduction rate and low maintenance requirements under laboratory conditions. Furthermore, next crucial aspect of blatticomposting is the preparation of the bedding material, i.e., the material that forms the basis of the composting environment. The bedding material not only harbours the cockroaches, but also facilitates the burial of the organic waste, thus reducing the risk of attracting flies and other pests. Suitable substrates for bedding include vegetable peelings, agricultural residues, coconut fibres, egg cartons and shredded paper. To optimise the decomposition process, it is important to maintain an appropriate moisture content and a balanced carbon-nitrogen ratio. Cockroaches contribute to the decomposition of organic waste through mechanical decomposition and enzymatic digestion. Their highly efficient digestive system allows them to process various organic materials, including cellulose-rich substrates that are challenging for other decomposers. The microbiota of the scrape gut plays a crucial role in cellulolytic digestion. For example, Pycnoscelus cockroaches harbour the cellulolytic strain Bacillus pumilus IB 320, which promotes the bioconversion of various paper-based waste materials. As a result of this digestion process, the cockroaches excrete nutrient-rich faeces containing significant concentrations of nitrogen, phosphorus and potassium, important elements for soil fertility. Over time, the compost matures and turns into a stable, dark-coloured material. Once the compost has fully decomposed, it can be harvested and used in agriculture, where it improves soil quality, retains moisture and promotes plant growth [21,22]. In addition to its role in composting, research has shown that the American cockroach (Periplaneta americana) has significant nutritional value. Dried and processed cockroach protein meal has a long shelf life and retains high-quality nutritional properties. Their biochemical composition confirms that they are a rich source of proteins, lipids and essential micro- and macronutrients, supporting their potential use as food and feed [31,32]. According to research by [33], cockroaches of the genuses Blatta and Blaptica, as well as other species, have proven to be extremely useful for the biodegradation of organic waste. Authors have shown that cockroaches can effectively reduce the amount of waste and produce valuable compost that is useful for agriculture or other industrial processes. However, when using cockroaches for waste disposal, it is necessary to carefully manage cockroach populations to prevent the spread of these organisms outside of controlled environments. It is also necessary to understand the potential risks to human health and the environment and take appropriate safety measures [34]. Research by [21] shows that cockroaches efficiently decompose a wide range of organic waste, including food scraps, paper, cardboard and other materials. This approach is not only environmentally sustainable, as it reduces the amount of waste going to landfill, but also economically beneficial, as it lowers waste processing costs. Blatticomposting can be used in various settings, in households, restaurants, farms and industrial plants. In addition, this method has potential for the production of biogas or other renewable energy sources from organic waste. Cockroaches like to eat materials that are traditionally considered difficult or unsuitable for composting, such as cellulose-rich plant components or complex organic compounds. According to [21], the main advantages of blatticomposting include efficient decomposition, as cockroaches accelerate the decomposition of organic waste and thus significantly reduce the time required for compost formation. In addition, the use of cockroaches in the composting process reduces the amount of waste destined for landfill and the resulting compost is enriched with essential nutrients that make it an excellent fertiliser or substrate for growing plants. This means that this technique minimises reliance on synthetic fertilisers and artificial substrates and promotes environmental sustainability. The low trophic specificity of cockroaches, their adaptability to different conditions, high reproduction rate and tolerance to overpopulation make them an extremely effective bioremedium for various organic wastes. The effectiveness of blatticomposting depends on factors such as the choice of cockroach species, composting substrate, environmental conditions (temperature, moisture and pH) and management practises. Unlike pest species, many cockroach species are not invasive and thrive in natural outdoor environments where they feed on leaf litter and other organic waste. In tropical ecosystems, cockroaches contribute to nutrient cycling by feeding on leaves and wood, promoting humification processes [22]. These attributes position blatticomposting as a promising, scalable solution for sustainable waste management and resource recovery.
The aim of this paper was to investigate and describe a new method for the disposal of biodegradable waste using cockroaches of the species Blaptica dubia, Blatta lateralis and Blaberus giganteus and to determine their potential in the disposal of slaughterhouse waste in the production of blatticompost. In order to achieve the stated aim, we analysed the amount and potential for decomposition of slaughterhouse waste using these three cockroach species. We compared the microbiological and chemical properties of the frass as a resulting substrate, i.e., the results of their activity, with literature sources and other types of compost.
2. Materials and Methods
2.1. General Characteristics of the Cockroach Species Used in the Study
Three cockroach species were used in this study: Blaptica dubia [35], Blatta lateralis [36], and Blaberus giganteus [37]. These species are commonly used in breeding and experimental studies due to their high reproductive capacity, tolerance to overcrowding, and ability to thrive under controlled laboratory conditions. Unlike synanthropic pest species such as the German cockroach (Blattella germanica), they are unable to permanently infest human dwellings. All three species exhibit hemimetabolic metamorphosis, i.e., the transition from juvenile to adult is gradual and characterised by three life stages: egg, nymph, and adult [38]. Only adult individuals are capable of reproduction and possess wings, while nymphal development proceeds gradually through successive moults. Prolonged nymphal stages with additional moults may occur due to factors such as lack of stimulation of the colony or unfavourable environmental conditions, such as lower temperatures and humidity. The studied species display pronounced sexual dimorphism, with males generally being more capable of flight than females, and differ in their reproductive strategies, ranging from ovoviviparity in B. dubia to ootheca formation in B. giganteus. In addition to their suitability for laboratory rearing, these cockroaches fulfil an important ecological function as decomposers, contributing to the breakdown of organic matter and nutrient recycling. These species are commonly bred under controlled laboratory conditions due to their high reproductive capacity, tolerance of high population densities, and adaptability to artificial habitats. Importantly, none of the studied species can establish permanent populations under European climatic conditions, as their survival depends on specific temperature and humidity regimes that are not naturally present in most regions of Europe. This characteristic is an important biosafety aspect in experimental rearing. Therefore, they will not become invasive and safe for the environment.
2.1.1. Blaptica dubia
B. dubia is a medium-sized cockroach species, reaching a body length of approximately 4.0–4.5 cm. Adult individuals are dark brown to black in colour, with occasional orange striping visible under strong light conditions [39]. The species exhibits pronounced sexual dimorphism: adult males possess fully developed wings covering the entire body, whereas females have reduced wing stubs, with the forewings extending to approximately one quarter of the body length (Figure 1). In its natural habitat, B. dubia inhabits the leaf litter of tropical forests in northern South America, where high temperature, humidity and predation pressure have favoured adaptations such as rapid reproduction, early larval development and relatively large nymphs at birth compared to other cockroach species. The species exhibits exceptional tolerance to high population densities, making it particularly suitable for intensive and industrial scale rearing. Colonies are self-sustaining and do not require separation of nymphs, allowing regeneration within approximately six months [40]. B. dubia does not require a substrate and thrives in plastic containers with adequate air circulation, enabling efficient space-efficient cultivation under controlled conditions. B. dubia is flightless and unable to climb smooth surfaces, which facilitates handling and minimises the risk of escape. These traits, combined with its tolerance to overcrowding, further contribute to its suitability for large-scale rearing systems. The species is ovoviviparous, with females producing approximately 20 to 40 live nymphs per month under optimal conditions. The ideal breeding temperature ranges between 24 and 35 °C, while reproductive activity ceases below 20 °C [39,41]. Nymphs of B. dubia are susceptible to dehydration, making access to moisture-rich food a critical factor for successful development. Although the species tolerates a relatively wide range of ambient humidity, insufficient moisture can impair moulting success. In captive breeding, dietary water availability is therefore more important than environmental humidification. When adequately fed with fresh fruits or vegetables, individuals are able to store water in their bodies, which may be released under stress conditions. Due to its relatively soft exoskeleton with low chitin content, B. dubia exhibits a higher protein concentration compared to many other cockroach species, further enhancing its value for breeding and experimental applications.
2.1.2. Blatta lateralis
B. lateralis is a relatively small cockroach species, reaching approximately 3 cm in body length, and is smaller than B. dubia and B. giganteus. The species exhibits pronounced sexual dimorphism. Adult males are slender and reddish-brown to orange, with elongated yellow wings used in courtship displays, while females are broader-bodied, dark brown to black, and characterised by a light marking on the pronotum and a pale stripe along the wing margins. Females are wider, possess short, rudimentary wings and are flightless (Figure 2). Nymphs are wingless and display a characteristic colour pattern, being brown on the anterior body and black on the posterior segments [42]. In its natural habitat, B. lateralis inhabits arid and semi-arid regions of the Arabian Peninsula, characterised by high temperatures and low humidity. It is typically found under rocks, within damp crevices or on desert farms, where it exhibits primarily nocturnal foraging behaviour [43]. B. lateralis is characterised by an exceptionally high reproductive capacity, allowing colonies to expand rapidly under favourable conditions [44]. The species tolerates overcrowding well and does not require a substrate, which facilitates large-scale breeding in plastic rearing containers [26]. Due to its ecological similarities with B. orientalis Linnaeus, a species widely distributed in Europe, B. lateralis is considered well suited for breeding under European climatic conditions [44]. Adult males possess fully developed wings and are capable of flight, which requires additional care during handling. Females are flightless, reducing escape risk in mixed colonies. In certain tropical regions, male flight activity combined with attraction to light can result in nuisance behaviour and they can often be found near human dwellings; however, B. lateralis is not generally regarded as a household pest, unlike synanthropic species such as the German cockroach. The species is highly active and exhibits rapid population growth, which, together with its tolerance to high stocking densities, makes B. lateralis particularly suitable for experimental and production-oriented rearing systems under controlled conditions.
2.1.3. Blaberus giganteus
B. giganteus is one of the largest cockroach species in the world. Adult females reach up to 10 cm in body length, while males attain lengths of approximately 7.5 cm; females also exhibit a wingspan of up to 15 cm. Adults are brown in colour with distinct black markings, and their dorsoventrally flattened bodies facilitate concealment within narrow spaces [45]. Both sexes possess two pairs of wings that can be folded over the abdomen; however, females are less inclined to fly due to their larger body mass (Figure 3). Paired appendages (cerci) are present on the terminal abdominal segment in both sexes, while males additionally possess small hair like appendages (styli). The species is endemic to the Neotropical region and occurs primarily in caves and tropical forests of South and Central America and Brazil. It favours environments with high humidity and minimal light, such as caves, hollow trees and rock crevices, and is frequently associated with bat habitats [46]. These ecological conditions have shaped its preference for humid environments and its predominantly nocturnal activity. Compared to B. dubia and B. lateralis, B. giganteus exhibits a slower reproductive rate and requires lower stocking densities during rearing due to its large body size. Under controlled conditions, colonies are therefore maintained at reduced population density per container to ensure adequate space and resource availability. Both sexes possess wings, but female flight activity is limited by body mass, reducing escape risk during handling. The species prefers higher ambient humidity, and prolonged nymphal development may occur under suboptimal temperature or humidity conditions. These physiological sensitivities were considered when maintaining experimental colonies. B. giganteus undergoes hemimetabolic development consisting of egg, nymph and adult stages [47]. After mating, females carry the fertilised ootheca for approximately 60 days before nymphs emerge [48]. The lifespan of the species can extend up to 20 months, depending on environmental conditions and diet. The species is a nocturnal omnivore and scavenger, feeding primarily on decaying plant material, but also consuming bat guano, fruits, seeds and carrion [45]. In addition, B. giganteus exhibits advanced physiological adaptations, including an efficient tracheal respiratory system with activity-dependent oxygen consumption and a symbiotic association with Blattabacterium spp., which convert nitrogenous wastes into amino acids, compensating for the low nitrogen content of its natural diet [45].
2.2. General Rearing Conditions
Cockroaches were housed in plastic containers with a volume of 25 dm³. For each species, four containers were prepared, each containing 100 individuals composed of males, females and nymphs, maintaining an approximate female-to-male ratio of 5:1. Cardboard egg cartons were vertically stacked inside each container to provide shelter, increase usable surface area and facilitate movement and moulting (Figure 4). The containers were sealed with perforated lids equipped with ventilation holes (1 mm diameter) to ensure sufficient air circulation while preventing escape and contamination by other insects. All colonies were maintained under controlled environmental conditions corresponding to the eco-physiological requirements of the species: temperature 25±2 °C and relative humidity 70±5%. The rearing environment was kept dry and clean to promote desiccation of faeces and reduce microbial growth. The total duration of the experiment was two months.
2.3. Experimental Design and Feeding Regimes
The experimental setup consisted of test and control groups for each cockroach species. In the test groups, cockroaches were fed Category III slaughterhouse waste, while control groups received a standard cockroach rearing diet. Category III slaughterhouse waste includes animal by-products derived from animals fit for human consumption but not intended for human use, such as meat trimmings and selected offal. These materials are considered low-risk and are permitted for use in animal feed and other non-food applications under EU Regulation [49]. For each species, three containers were assigned to the test group and supplied once weekly with an average of 20 g of slaughterhouse waste. The waste consisted of chopped or ground animal material, including chicken, beef, pork, veal and wild boar, in raw or thermally processed form. Control groups, housed in separate containers (one per species), were fed a conventional diet consisting of fruits (bananas, apples), vegetables (carrots) and pelleted cat food commonly used in cockroach rearing. All samples were analysed in triplicate.
Figure 5.
Feeding of B. lateralis cockroaches with raw slaughterhouse waste and B. dubia with carrot and banana.
Figure 5.
Feeding of B. lateralis cockroaches with raw slaughterhouse waste and B. dubia with carrot and banana.

2.4. Frass Collection and Sample Preparation
Cockroach faecal material (frass) was collected once per week over a period of 10 weeks. Frass was removed from the bottom of each container and placed into sterile 100 mL plastic containers. All samples were labelled and stored prior to microbiological and chemical analysis. Before chemical analysis, frass samples were dried at 105 °C, ground and homogenised to ensure sample uniformity.
2.4.1. Physicochemical Analysis of Frass
Water content and dry matter content were determined gravimetrically at 105 °C according to [50]. Ash content was determined following [51]. pH (10% w/v in water) was measured electrometrically according to [52], and electrical conductivity (10%) was determined conductometrically. Organic carbon content was analysed using the dichromate method, and organic matter content was calculated accordingly. Total nitrogen was determined using the Kjeldahl method [53]. Total phosphorus (P₂O₅), potassium (K₂O), calcium (Ca), magnesium (Mg), manganese (Mn) and iron (Fe) were determined after aqua regia digestion following [54], using spectrophotometry, flame photometry and atomic absorption spectroscopy, respectively.
2.4.2. Microbiological Analysis of Frass
For microbiological analysis, 10 g of each frass sample was analysed according to [55]. The presence of Salmonella spp., Shigella spp., Escherichia coli and Campylobacter spp. was determined using standard microbiological detection methods.
2.5. Statistical Analysis
The results are presented as mean value (arithmetic mean) and standard deviation. An analysis of variance (ANOVA) was performed to determine the difference between the variables observed in the study. The study was conducted in three replicates. Principal component analysis (PCA) was used to reduce the multidimensionality of the data set. The interpretation of the numerical designations used in the PCA representation can be found in Table 1. The number in the column “PCA No.” corresponds to the identifier of each analysed sample as shown in the PCA diagram below. This identifier represents the interaction between two categories: type and parameter.
3. Results and Discussion
To assess the quality of the produced blatticompost, basic chemical analyses were carried out on frass material derived from three cockroach species (B. dubia, B. lateralis and B. giganteus). Two feeding treatments were used: a control diet and diet based on slaughterhouse waste. The results obtained are shown in Table 2.
The results presented in Table 2 show significant differences between the frass samples produced by the three cockroach species tested for all parameters analysed (p < 0.01). Each species produced compost with different physical and chemical properties, illustrating the influence of species-specific digestion and metabolism on the resulting compost-like material. Recent studies on insect frass and organic composts confirm that such interspecific differences are a dominant driver of frass chemical composition, often exceeding the influence of substrate type alone [17,18]. B. giganteus frass had the lowest moisture content and produced the driest compost samples, while B. dubia produced compost with the highest moisture content, regardless of the type of feed. According to [56], low-moisture frass materials exhibit enhanced storage stability and reduced microbial degradation during post-processing, making them more suitable for commercial handling and long-term storage. The pH values were generally close to neutral (around 7.0), except in the control group of B. dubia, where the pH dropped significantly to 5.04. This acidification is possibly due to incomplete decomposition or microbial activity. Comparable pH shifts have been reported in recent frass-based compost studies, where acidic conditions were linked to incomplete stabilisation of labile organic fractions and accumulation of short-chain organic acids [18]. Neutral pH values are widely recognised as optimal for soil application, enhancing nutrient availability and microbial activity while minimising phytotoxic risks [56]. Electrical conductivity (EC), a measure of soluble salt content, varied considerably: the lowest EC was measured in B. giganteus, indicating lower ion accumulation, while the highest values were found in B. lateralis, possibly related to higher nitrogen or salt content due to metabolic processes or feed composition. Recent comparative analyses of insect derived composts indicate that EC is highly species dependent and should be considered a key parameter when evaluating frass suitability for salt-sensitive crops [17]. Ash content, which represents the inorganic mineral fraction, showed great variability, ranging from 13.93% to 34.69%, with no consistent pattern between species or feed types. This irregularity is probably due to uncontrolled factors such as feed composition, gut residence time or environmental factors such as moisture and microbial colonisation during rearing. [57] similarly reported high variability in ash content of insect frass, attributing it to selective mineral assimilation by insects and post-excretion microbial transformations. The loss on ignition (LOI), which estimates the organic matter content, also showed no clear trends and ranged between 65.31% and 86.06%. As with ash, this inconsistency could reflect differences in feed digestibility, microbial biomass or the stability of organic compounds in the blatticompost matrix. High LOI values are increasingly interpreted as indicators of organic matter preservation and potential benefits for soil structure and carbon sequestration rather than as a limitation of compost maturity [17]. The moisture content and dry matter values observed in this study fall within the broad ranges reported by [21] but also highlight pronounced species-specific differences. Recent meta-analyses of insect-based composts emphasise that species-specific digestive physiology plays a decisive role in determining frass physical properties, even under identical feeding regimes [18]. The notably low moisture content and high dry matter observed in B. giganteus frass confirm earlier findings that larger-bodied cockroach species produce structurally denser and drier residues, potentially improving the storage stability and handling properties of the final product. In contrast, B. dubia consistently produced the wettest frass, particularly under the control diet, which aligns with its higher organic matter and carbon content. These findings suggest that frass physical properties are not solely determined by substrate but are strongly influenced by species-specific digestion efficiency and water balance regulation. The pH values clustered around neutrality in most treatments, corroborating previous reports that frass generally exhibits favourable pH conditions for soil application. The markedly acidic pH observed in the B. dubia control group is an exception and may reflect diet composition and microbial activity rather than an inherent limitation of the species. Electrical conductivity values were significantly lower in B. giganteus frass compared to B. lateralis and B. dubia, suggesting reduced soluble salt accumulation. [56] highlight low-EC frass materials as particularly suitable for repeated application in sustainable and low-input agricultural systems. This is particularly relevant from an agronomic perspective, as lower EC values reduce the risk of salinity stress in sensitive crops. Ash content and loss on ignition (LOI) displayed high variability across species and treatments, with no clear monotonic trend between control and slaughterhouse-fed groups. Similar inconsistencies have been reported by [21], indicating that mineralisation processes in cockroach systems are likely influenced by a combination of diet composition, gut retention time, and microbial transformations, rather than substrate type alone. Overall, the physicochemical diversity of the frass as a blatticomposts emphasises the complexity of bioconversion processes in cockroach-based systems and suggests that each species may offer unique advantages depending on the desired properties of the final product, such as moisture, nutrient richness or mineral content.
Figure 6.
Physical and chemical analysis of frass sample.
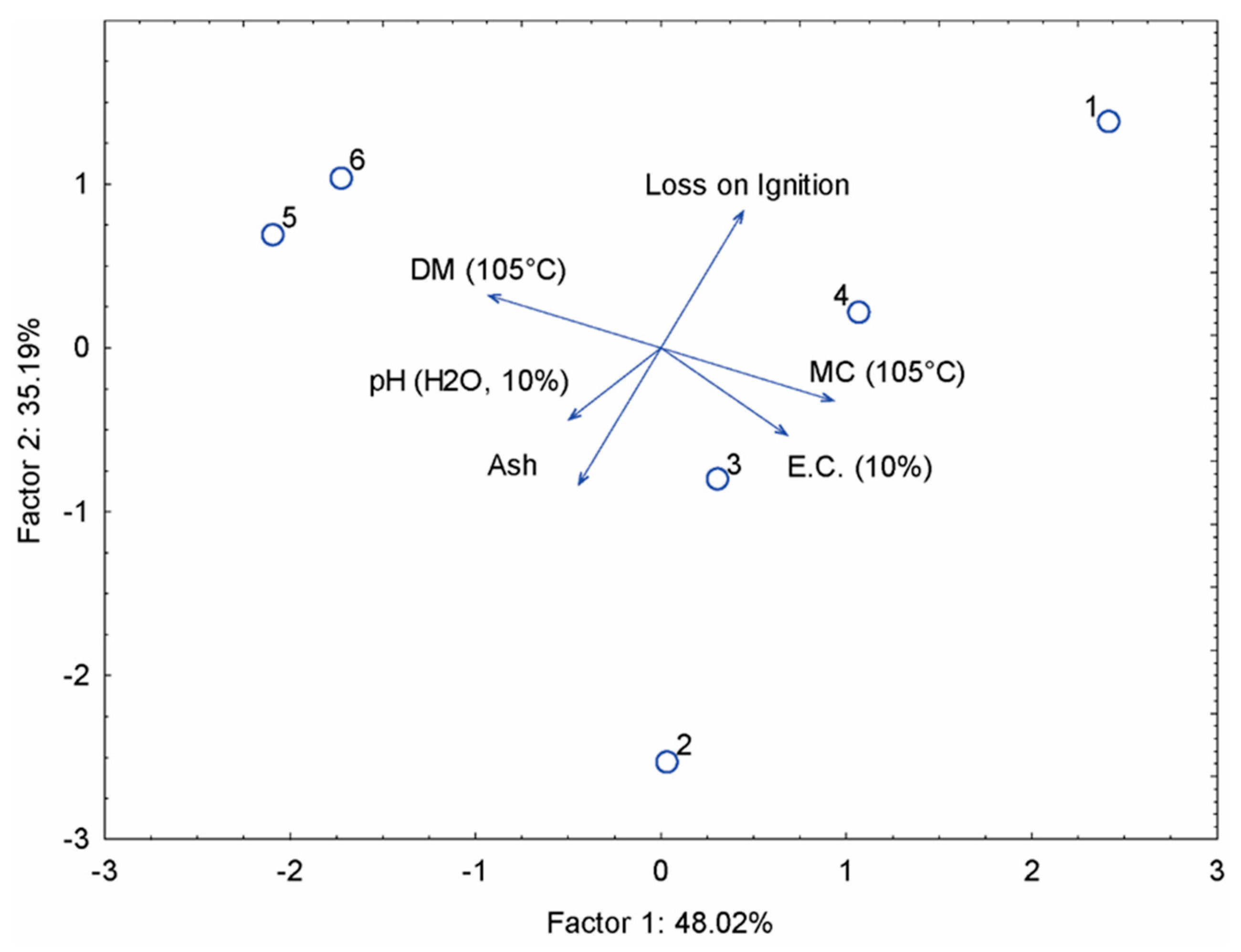
The first principal component (horizontal axis) explains 48.02% of the total variance and clearly separates samples 1, 2, 3 and 4 on the right side of the graph. These samples are associated with higher values of electrical conductivity (EC) and moisture content (MC) at 105 °C. Conversely, samples 5 and 6 are on the left-hand side of the graph and are characterised by higher pH values and dry matter (DM) contents. The second principal component (vertical axis) accounts for 35.19% of the variance and distinguishes the samples based on their ash content and loss on ignition (LOI). Sample 2 has the lowest ash content, while sample 6 has the highest LOI, indicating a significant amount of volatile organic matter. The elemental composition of the frass was evaluated to assess their agronomic potential and nutritional value as soil conditioners. Table 3 shows the concentrations of the main macroelements, including essential nutrients such as total nitrogen, phosphorus and potassium, as well as other elements important for plant growth and soil health. The organic matter and organic carbon content was also determined to assess the stability and quality of the compost. In addition, the presence of secondary macronutrients (e.g., calcium and magnesium) and traces of important micronutrients (e.g., iron and manganese) were measured to provide a comprehensive overview of the fertilising potential of the material. The composition of the frass is significantly influenced by the type of substrate provided to the cockroaches. Different substrates lead to fluctuations in the main nutrients: The nitrogen, phosphorus and potassium content varies depending on the food source, with the diet of B. giganteus having a low nitrogen, phosphorus, potassium, calcium, magnesium and iron content. Similar substrate- and species-dependent nutrient variability in blatticompost has been reported by [21] and [22]. C organic, organic matter and manganese content is found in B. dubia as a highest in all investigation, but other components showed in Table 3 has higher level in frass made from B. lateralis. Elevated organic matter and carbon contents in insect-derived composts are commonly associated with incomplete mineralisation and enhanced microbial activity [1,2].
The macronutrient composition of frass as potential organic fertiliser in this study is broadly comparable to values reported in the literature but again shows strong species-dependent nutrient partitioning. Comparable nutrient ranges have been reported for vermicompost and other insect-based composting systems [10,16]. B. lateralis consistently exhibited the highest concentrations of total nitrogen, phosphorus and potassium, making it the most nutritionally balanced species for fertiliser applications. Similar findings regarding enhanced nutrient mineralisation in fast-metabolising detritivorous insects were reported by [5] and [6]. In contrast, B. giganteus frass contained lower concentrations of nitrogen, phosphorus and potassium, but relatively stable calcium and magnesium levels, suggesting potential suitability for soil conditioning rather than intensive fertilisation. Materials with lower nutrient density but higher chemical stability are commonly classified as soil conditioners rather than fertilisers [5,58]. Organic carbon and organic matter contents were highest in B. dubia, regardless of feed type, indicating enhanced carbon retention and incomplete mineralisation. High organic carbon retention is considered beneficial for soil structure improvement and long-term carbon sequestration [2,59]. This may be advantageous for soil organic carbon sequestration but could limit immediate nutrient availability.Slow nutrient release from organic amendments with high C content is a well-documented phenomenon [2,6]. This result confirms earlier findings that cockroaches can buffer substrate variability and produce chemically stable frass even when fed nutritionally complex inputs. Similar buffering effects of cockroach digestion have been highlighted by [21]. Compared to black soldier fly compost reported by [19], the nutrient composition of blatticompost in some parameters closely matches that of BSF-derived materials, particularly in nitrogen and potassium content. Comparable overlaps between BSF frass and other insect-derived composts were also discussed by [16]. However, blatticompost shows greater variability, reflecting the less industrialised and more species-sensitive nature of cockroach systems. Species sensitivity and system variability are recognised limitations but also functional advantages of emerging insect-based waste treatment technologies [19,21]. This variability, while challenging for standardisation, may also offer a functional advantage by allowing targeted substrate specialisation and adaptation to waste streams unsuitable for BSF larvae, such as slaughterhouse residues or potentially contaminated organic materials. Compared to the macronutrient composition of BSF compost reported by [19], the total nitrogen and potassium content in the blatticompost obtained in this study falls within a similar range to that of Hermetia illucens compost, while the phosphorus content is somewhat lower. In comparison with garden compost, which according to [60] and [61] typically contains lower concentrations of nitrogen and potassium, the analysed blatticompost shows a more favourable N status and especially a higher K content, although with a lower phosphorus content than values reported in some vermicomposts. Vermicompost, according to the same authors, often exhibits higher phosphorus but lower potassium levels compared to the blatticompost from this study, positioning blatticompost as a potassium-rich amendment with moderate nitrogen and relatively modest phosphorus levels. This macronutrient profile, although somewhat more variable due to the sensitivity of cockroach species to substrate composition, allows for the targeted application of blatticompost as a complementary amendment to BSF compost, vermicompost and conventional compost in production systems where additional potassium input is desirable, while phosphorus requirements can be adjusted through supplementary sources.
To ensure that all data in the graph fit on the same axis (in this case, the y-axis), a standard normalisation method was applied, whereby numerical values were rescaled to the range 0–1. The data normalisation method used for the graph 3 is calculated using the following equation (1):
where “x” is the original data value, “xₘᵢₙ” is the minimum value in the entire dataset, “xₘₐₓ” is the maximum value in the entire dataset, and “xₙₒᵣₘ” is the normalized value.
Figure 7 presents the min–max normalised values (0–1) of selected macronutrient and elemental parameters in frass produced by three cockroach species under control conditions (0) and feeding regimes supplemented with slaughterhouse waste (1). Normalisation was applied to enable direct comparison of parameters with different absolute ranges and units. Overall, clear species- and substrate-dependent patterns are observed. Samples from B. lateralis consistently show higher normalised values for total nitrogen, phosphorus (P₂O₅), potassium (K₂O), and calcium, indicating a nutrient-richer frass compared to the other species. In contrast, B. giganteus generally exhibits lower relative values across most nutrients, particularly for nitrogen, potassium, and micronutrient-associated elements, suggesting reduced nutrient accumulation or altered assimilation efficiency. Organic carbon and organic matter display relatively stable and comparable normalised values across species, indicating that carbon stabilisation during digestion is less sensitive to feeding regime than mineral nutrient dynamics. Feeding with slaughterhouse waste moderately shifts nutrient profiles within species; however, no uniform directional trend is evident across all parameters, suggesting complex interactions between substrate composition, species-specific digestive physiology, and nutrient partitioning. Manganese and iron display distinct patterns compared to major nutrients, with higher variability and weaker coupling to nitrogen- and phosphorus-related parameters, supporting the hypothesis that trace element dynamics are influenced by additional biological and environmental factors rather than substrate composition alone.
The composition of frass as a blatticompost is significantly influenced by the type of substrate provided to the cockroaches, as well as by species-specific physiological and metabolic characteristics. Nutrient concentrations varied between cockroach species, particularly in response to different feeding regimes. For example, B. giganteus consistently produced compost with the lowest levels of nitrogen (N), phosphorus (P), potassium (K), calcium (Ca), magnesium (Mg), and iron (Fe), regardless of whether it was fed a control diet or slaughterhouse waste. This may be related to its lower digestive efficiency or a slower rate of nutrient assimilation, especially with nutrient-rich substrates such as animal by-products. Conversely, B. dubia had the highest concentrations of organic carbon (C), organic matter, and manganese (Mn), suggesting a greater microbial contribution to substrate degradation and possibly a more active gut microbiome. In contrast, B. lateralis produced compost with the highest concentrations of most other elements measured, indicating increased nutrient mineralisation, possibly due to its higher metabolic rate and shorter digestion time. When comparing the nutrient profiles of frass from control feed and slaughterhouse waste, no consistent trend was observed across all species. While certain elements increased in some groups fed slaughterhouse waste, others showed no significant changes or even lower concentrations. This inconsistency suggests that the response to nutrient-rich substrates such as offal is highly species-dependent and may be influenced by factors such as the adaptability of the gut microbiota, nutrient intake thresholds, and stress responses to protein- or lipid-rich diets.
The heatmap (Figure 8) shows Pearson correlation coefficients among major nutrients, organic matter, and selected microelements in frass samples produced by different cockroach species and feeding regimes. Colour intensity represents the strength and direction of correlations, with warmer colours (red to orange) indicating strong positive correlations and cooler colours (blue to green) indicating weak or negative correlations. Strong positive associations are observed among macronutrients, particularly between total nitrogen, phosphorus (P₂O₅), potassium (K₂O), calcium, and magnesium, suggesting coupled nutrient dynamics during bioconversion. In contrast, manganese shows predominantly negative correlations with organic matter, iron, and macronutrients, indicating distinct behaviour compared to other elements within the system.
The correlation analysis revealed different patterns between the measured macro- and micronutrients, indicating possible co-accumulation processes and common physiological or chemical pathways. A strong positive correlation was observed between the main macronutrients, in particular between total nitrogen (N), phosphorus (P₂O₅), potassium (K₂O), calcium (Ca) and magnesium (Mg). Thus, total nitrogen showed an almost perfect correlation with phosphorus (r = 0.99), potassium (r = 0.95), calcium (r = 0.89) and magnesium (r = 0.85), suggesting a synchronised nutrient assimilation or concentration in the composted substrates. Similarly, phosphorus also showed high correlations with potassium (r = 0.94), calcium (r = 0.88) and magnesium (r = 0.83), while potassium also correlated strongly with calcium and magnesium (r = 0.85 and r = 0.82, respectively). These relationships indicate a probable co-dependence or mutual accumulation trend between these elements in the substrate matrix. A moderately positive correlation was also observed between total iron (Fe) and organic matter content (r = 0.69), suggesting that the availability of iron is partly determined by the organic matter pool, possibly through complexation or organic chelation mechanisms. On the other hand, manganese (Mn) showed a consistent pattern of negative correlations, particularly with organic matter (r = -0.84), total iron (r = -0.46) and various macronutrients such as nitrogen, phosphorus, potassium, calcium and magnesium (r between -0.31 and -0.18). This inverse relationship suggests that manganese behaves antagonistically with respect to both organic content and macro element profile, which could be due to its pronounced solubility dynamics or its redox-sensitive behaviour in organic-rich substrates. Taken together, these correlation patterns provide a valuable insight into the nutrient interactions in the produced frass and may help to optimise its application based on specific nutritional or agronomic needs. Overall, the heatmap clearly illustrates two contrasting correlation domains: a strongly interconnected macronutrient cluster dominated by nitrogen–phosphorus–potassium relationships, and a negatively associated manganese-driven cluster, highlighting divergent nutrient pathways within the frass as a blatticompost system. The strong positive correlations between total nitrogen, phosphorus, potassium, calcium and magnesium observed in the correlation analysis reinforce the interpretation that these nutrients are co-regulated and largely driven by organic matter decomposition dynamics. Similar nutrient coupling has been reported for food waste derived blatticompost and supports the classification of these materials as slow-release organic fertilisers.

Table 4.
Concentrations of trace elements (microelements) in frass samples.
| Cockroach species | B. dubia | B. lateralis | B. giganteus | Statistically significant |
|||
| Type of feed | Control group |
Slaughterhouse waste |
Control group |
Slaughterhouse waste |
Control group |
Slaughterhouse waste |
|
| Zn (mg/kg) | 52.05±0.07 | 36.41±0.01 | 39.52±0.03 | 40.32±0.25 | 12.70±0.14 | 12.1±0.14 | * |
| Cu (mg/kg) | 20.63±0.04 | 12.05±0.07 | 14.6±0.14 | 13.35±0.07 | 5.54±0.06 | 4.83±0.04 | * |
| Cd (mg/kg) | 0±0 | 0±0 | 0±0 | 0±0 | 0.24±0.01 | 0±0 | n.s. |
| Pb (mg/kg) | 2.73±0.04 | 2.36±0.02 | 3.03±0.03 | 2.95±0.01 | 1.99±0.01 | 0.59±0.01 | * |
| Ni (mg/kg) | 12.8±0.01 | 22.2±0.01 | 21.4±0.01 | 22.10±0.01 | 10.57±0.01 | 10.68±0.01 | * |
| Cr (mg/kg) | 21.05±0.07 | 22.3±0.01 | 31.78±0.04 | 25.73±0.04 | 14.53±0.01 | 16.21±0.01 | * |
| Hg (mg/kg) | 0±0 | 0±0 | 0±0 | 0±0 | 0±0 | 0±0 | n.s. |
| As (mg/kg) | 0±0 | 0±0 | 0±0 | 0±0 | 0±0 | 0.17±0.01 | n.s. |
| Mo (mg/kg) | 0.32±0.01 | 0.35±0.01 | 0.19±0.01 | 0.18±0.01 | 0±0 | 0±0 | * |
| Co (mg/kg) | 0.71±0.01 | 0.71±0.01 | 0.98±0.01 | 0.91±0.01 | 0±0 | 0±0 | * |
*Statistically significant at p < 0.01.
The first factor (horizontal axis) accounts for 68.85% of the variance and shows that lead (Pb), nickel (Ni), cobalt (Co), copper (Cu), molybdenum (Mo), chromium (Cr), and zinc (Zn) cluster near the centre of the diagram, indicating a similar distribution among the samples. The second factor (vertical axis) accounts for 16.41% of the variance and clearly distinguishes the samples: sample 5 displays elevated element concentrations in the upper part of the diagram, while sample 6 shows a negative correlation with the other elements, indicating a distinct chemical profile compared to the other samples. Trace element concentrations in all frass samples remained within regulatory thresholds specified in European guidelines and published compost standards. Compliance of insect frass with European compost and fertiliser regulations has been reported by [17] and [56]. Zinc, copper, nickel, and chromium concentrations varied significantly among species but showed no consistent enrichment pattern associated with slaughterhouse waste feeding. Similar interspecific variability without systematic substrate-driven accumulation of Zn, Cu, Ni and Cr was observed in insect frass across multiple rearing systems [17,62]. The absence or negligible concentrations of cadmium, mercury, and arsenic in most treatments is particularly noteworthy and supports the hypothesis that cockroaches may limit the bioavailability or accumulation of certain toxic elements. Low transfer of Cd, Hg and As into insect frass has also been reported for Hermetia illucens and other detritivorous insects, suggesting physiological or microbial exclusion mechanisms [18,57]. This observation is consistent with [21], who suggested that cockroach digestion and excretion pathways may act as a partial barrier to heavy metal transfer. Comparable barrier effects during insect-mediated bioconversion have been discussed by [62]. Recent research has increasingly recognised blatticomposting as a promising yet underexplored biological waste treatment strategy, particularly when compared with more established insect-based systems such as black soldier fly composting. [21] highlight that cockroaches display a high degree of physical and chemical substrate tolerance, enabling them to process heterogeneous organic waste streams that may be unsuitable for other insects. In this context, the present study advances existing knowledge by providing a comparative, species-specific, and substrate-dependent assessment of blatticompost quality derived from B. dubia, B. lateralis and B. giganteus, fed either a control diet or slaughterhouse waste. Unlike most previous studies, which primarily focused on food waste, the inclusion of animal by-products introduces a more complex and nutrient-dense substrate, thereby enhancing the applied relevance of the results. Microbiological analysis of compost is carried out to assess its safety, particularly regarding the presence of pathogenic bacteria such as Salmonella spp., Shigella spp., Escherichia coli, and Campylobacter spp. These microorganisms may pose a significant risk to human health if present in compost intended for agricultural or horticultural use. The results of the microbiological analysis are reported as presence or absence (isolated/not isolated) of Salmonella spp., Shigella spp., E. coli, and Campylobacter spp. in 10 g of wet sample. Evaluation of all culture media indicated the absence of the tested pathogens, except on SS agar. Typical Salmonella colonies appear as transparent colonies with a black centre, while Shigella colonies are colourless on SS agar. Although some colonies displayed Salmonella-like morphology (black centre), the presence of pink pigmentation in and around each colony prevented clear reading and unambiguous interpretation of the results (Figure 9). Due to the ambiguous results on SS agar, additional confirmation was performed using the MALDI-TOF method. MALDI-TOF (Matrix-Assisted Laser Desorption/Ionisation Time-of-Flight) mass spectrometry is a widely used analytical technique for identifying biomolecules such as proteins, peptides, lipids, and carbohydrates, as well as large organic molecules [63]. The additional MALDI-TOF analysis confirmed the absence of the investigated pathogenic bacteria; however, Pseudomonas aeruginosa was detected. According to the [55] standard, the presence of this species does not represent a hazard in environmental samples. Overall, the microbiological analysis showed that none of the targeted pathogenic bacterial strains were isolated from the compost samples. The absence of Salmonella spp., Shigella spp., E. coli, and Campylobacter spp. may indicate that these bacteria were not present in the substrate before the composting process. Alternatively, their elimination may be due to biological activity and metabolic processes associated with cockroach digestion; however, due to limited relevant literature, further targeted studies are required to confirm this hypothesis. Regardless of the underlying mechanism, the absence of pathogenic microorganisms in the analysed samples indicates the microbiological safety of the compost, supporting its suitability for agricultural and horticultural applications without posing a risk of pathogen transmission to crops and, consequently, to human consumers. In addition, the observed absence of pathogenic bacteria may indicate a potential antimicrobial effect associated with cockroach mediated bioconversion. Previous studies have shown that cockroaches possess a highly active gut microbiome and produce various antimicrobial peptides and enzymes as part of their innate immune system, which help suppress pathogenic microorganisms within the digestive tract [64,65]. These biological mechanisms may contribute to the inactivation or reduction of enteric pathogens during the digestion and transformation of organic waste. Furthermore, the rapid passage of organic material through the cockroach gut, combined with mechanical fragmentation, enzymatic degradation, and intense microbial competition, may create unfavourable conditions for the survival of pathogens. Similar reduction of pathogen has been reported in other insect-based bioconversion systems, including black soldier fly larvae, where pathogen suppression is attributed to gut-associated microbiota, antimicrobial compounds, and altered physicochemical conditions of the substrate [66].
Although the present study was not designed to directly quantify pathogen inactivation mechanisms, the consistent absence of the tested pathogens across all samples suggests that cockroach-based bioconversion may inherently contribute to microbiological stabilisation and sanitisation of organic waste. This highlights the need for future targeted research on the antimicrobial potential of cockroach digestion, including controlled inoculation experiments and metagenomic analyses of gut microbiota, to elucidate the underlying mechanisms and confirm their relevance for safe waste management applications.
4. Conclusions
Cockroaches are a potential means of disposing of specific types of biodegradable waste, such as slaughterhouse waste, due to their resilience and broad food spectrum. Based on the research carried out, it can be concluded that cockroaches of the species Blaptica dubia, Blatta lateralis and Blaberus giganteus have significant potential for the disposal of slaughterhouse waste and the production of compost. Their ability to decompose organic material leads to a reduction in the volume of waste, while the resulting compost has favourable microbiological and chemical properties. These results suggest that the use of cockroaches for the disposal of slaughterhouse waste could provide a sustainable and environmentally friendly alternative to traditional methods of slaughterhouse waste disposal, with the added benefit of producing a high quality compost. In the future, the use of cockroaches may significantly facilitate the disposal of slaughterhouse waste from an economic and environmental perspective. Overall, the results of this study confirm and extend previous findings by demonstrating that blatticomposting is a robust, flexible and species-dependent waste valorisation strategy. The strong influence of cockroach species on frass quality, combined with the relatively minor impact of feed type, suggests that species selection is a key design parameter for future blatticomposting systems. The chemical stability, favourable nutrient profiles and low contamination levels observed in this study support the potential integration of cockroach-based bioconversion into circular bioeconomy frameworks, particularly for complex organic waste streams that challenge conventional composting technologies. The chemical composition of the produced blatticompost showed balanced macronutrient levels and stable organic matter, comparable to established composting systems. Trace element concentrations remained within regulatory thresholds, indicating chemical stability and suitability for agricultural and horticultural application. In summary, while cockroach bioconversion offers promising potential for nutrient recycling, its application requires a customised, species- and substrate-specific approach. Further research on substrate composition, interaction with gut microbiota and post-processing safety is crucial to advance cockroach composting as a sustainable waste management strategy. Despite this promising potential, it is important to acknowledge the regulatory and legal challenges associated with cockroach-based bioconversion within the European Union. Unlike black soldier fly larvae, which are already regulated under EU legislation on animal by-products, feed safety, and insect biomass processing, cockroaches are not clearly defined within the current regulatory framework. In most EU Member States, they have no harmonised classification regarding their legal status (zoo-technical organism, feed material, waste-processing insect, or pest). This lack of regulatory clarity may hinder commercial development, particularly regarding the handling of slaughterhouse waste, biosafety standards, movement of insect-derived materials across borders, and the use of frass as a fertilising product. Therefore, future implementation of cockroach-based waste management may require the establishment of clearer EU level guidance, including safety, biosanitary, and fertiliser compliance standards, before full-scale deployment can occur.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, N.V.; methodology, A.P.; K.G.; T.K.; J.Š.; and M.H.; software, I.B.; validation, N.V., T.K. and A.P.; formal analysis, I.B.; investigation, K.G.; M.H.; N.V.; A.P.; resources, T.K.; J.Š.; data curation, I.B.; writing—original draft preparation, N.V.; writing—review and editing, A.P.; T.K.; visualization, J.Š.; supervision, N.V.; funding acquisition, T.K. All authors have read and agreed to the published version of the manuscript.”.
Funding
This research received no external funding.
Data Availability Statement
The data can be made available by contacting the corresponding author.
Acknowledgments
The authors have reviewed and edited the output and take full responsibility for the content of this publication.”.
Conflicts of Interest
The authors declare no conflicts of interest.”.
References
- Haug, R.T. The Practical Handbook of Compost Engineering, 1st ed.; Lewis Publishers: Boca Raton, Florida, USA, 1993. [Google Scholar]
- Epstein, E. Industrial Composting: Environmental Engineering and Facilities Management, 1st ed.; CRC Press: Boca Raton, Florida, USA, 2011. [Google Scholar]
- Ganji, A.; Hajrajabi, A.; Babaei, A.; Aayush, K.; Sharma, S.; Shayegan, J. An overview of home composting: configurations and design of bin, process parameters, and benefits. J. Mater. Cycles Waste Manag. 2025, 1–24. [Google Scholar] [CrossRef]
- Diaz, L.F.; Savage, G.M.; Eggerth, L.L.; Golueke, C.G. Composting and Recycling Municipal Solid Waste, 2nd ed.; CRC Press: Boca Raton, Florida, USA, 2007. [Google Scholar]
- Brady, N.C.; Weil, R.R. The Nature and Properties of Soils, 15th ed; Pearson Education: Upper Saddle River, New Jersey, USA, 2016. [Google Scholar]
- Cooperband, L. The Art and Science of Composting: A Resource for Farmers and Compost Producers; University of Wisconsin Extension, Madison: Wisonsin, USA, 2002. [Google Scholar]
- Baldwin, K.R.; Greenfield, J.T. Composting on Organic Farms; Center for Environmental Farming Systems: Goldsboro, North Carolina, USA, 2009. [Google Scholar]
- Menino, R.; Murta, D. The insects as a workforce for organic fertilizers production–insect frass. In New Generation of Organic Fertilizers; IntechOpen: London, UK, 2021. [Google Scholar]
- Edwards, C.A.; Arancon, N. Vermicompost suppresses plant pests and disease attacks. Rednova News 2004. [Google Scholar]
- Edwards, C.A.; Arancon, N.Q.; Sherman, R. (Eds.) Vermiculture Technology: Earthworms, Organic Wastes, and Environmental Management; CRC Press: Boca Raton, Florida, USA, 2010. [Google Scholar]
- Lim, S.L.; Wu, T.Y.; Lim, P.N.; Shak, K.P. The use of vermicompost in organic farming: overview, effects on soil and economics. J. Sci. Food Agric. 2015, 95, 1143–1156. [Google Scholar] [CrossRef]
- Ahmad, A.; Mahmood, A.; Ahmad, S.; Li, W. Harnessing earthworms for sustainable waste management: insights into vermicomposting. Water Air Soil Pollut. 2025, 236, 840. [Google Scholar] [CrossRef]
- Barragan Fonseca, K.; Dicke, M.; van Loon, J. Nutritional value of the black soldier fly (Hermetia illucens L.) and its suitability as animal feed. J. Insects Food Feed 2017, 3, 7–14. [Google Scholar] [CrossRef]
- Fitriana, A.R.; Bagastyo, A.Y.; Febrianto, A.; Tay, C.Y.; Warmadewanthi, I.D.A.A. A systematic review of the potential of industrial organic by-products as black soldier fly larvae rearing substrates: nutrient content and environmental impact. J. Mater. Cycles Waste Manag. 2025, 1–18. [Google Scholar] [CrossRef]
- Tognetti, C.; Mazzarino, M.J.; Laos, F. Composting vs. vermicomposting: A comparison of end product quality. Compost Sci. Util. 2007, 15, 147–154. [Google Scholar] [CrossRef]
- Insam, H.; Franke-Whittle, I.; Goberna, M. (Eds.) Microbes at Work: From Wastes to Resources; Springer: Heidelberg, Germany, 2010. [Google Scholar]
- Praeg, N.; Klammsteiner, T. Primary study on frass fertilizers from mass-reared insects: Species variation, heat treatment effects, and implications for soil application at laboratory scale. J. Environ. Manage. 2024, 356, 120622. [Google Scholar] [CrossRef] [PubMed]
- Safitri, R.A. Exploring potential uses of insect frass for agricultural application. J. Insects Food Feed 2024, 11, 167–182. [Google Scholar] [CrossRef]
- Smetana, S.; Schmitt, E.; Mathys, A. Sustainable use of Hermetia illucens insect biomass for feed and food: attributional and consequential Life Cycle Assessment. Resour. Conserv. Recycl. 2019, 146, 1–15. [Google Scholar] [CrossRef]
- West, J. Getting to the Bottom of Blatticomposting: Using Cockroaches to Create Soil Amendments from Food Scraps and Other Organic Wastes. Community Dev. J. 2010, 45, 251–261. [Google Scholar]
- Patón, D.; García-Gómez, J.C. Blatticomposting of Food Waste, Production Estimates, Chemical Composition and CO2 Emissions Savings: A Case Study. Waste Biomass Valor 2023, 14, 1123–1135. [Google Scholar] [CrossRef]
- Gowrisankar, R.; Sumithramma, N.; Mulimani, V.; Pradhan, S.K. Blatticomposting: A Sustainable Approach for Organic Waste Management. Int. J. Environ. Clim. Change 2023, 13, 754–762. [Google Scholar] [CrossRef]
- Robinson, W. Urban Insects and Arachnids: A Handbook of Urban Entomology, 1st ed.; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Ardestani, M.M.; Šustr, V.; Hnilička, F.; Frouz, J. Food consumption of the cockroach species Blaptica dubia Serville (Blattodea: Blaberidae) using three leaf litter types in a microcosm design. Appl. Soil Ecol. 2020, 150, 103460. [Google Scholar] [CrossRef]
- Elzinga, R.J. Fundamentals of Entomology, 6th ed.; Pearson: London, UK, 2004. [Google Scholar]
- Bell, W.J.; Roth, L.M.; Nalepa, C.A. Cockroaches: Ecology, Behavior, and Natural History; JHU Press: Baltimore, Maryland, USA, 2007. [Google Scholar]
- Enebe, M.C.; Erasmus, M. Blatticomposting– a perspective on cockroach-mediated waste valorization to nutrient-rich organic fertilizer for enhancing soil health. Soil Adv. 2024, 2, 100012. [Google Scholar] [CrossRef]
- Salomone, R.; Saija, G.; Mondello, G.; Giannetto, A.; Fasulo, S.; Savastano, D. Environmental impact of food waste bioconversion by insects: Application of Life Cycle Assessment to process using Hermetia illucens. J. Clean. Prod. 2017, 140, 890–905. [Google Scholar] [CrossRef]
- Oonincx, D.G.A.B.; Finke, M.D. Nutritional value of insects and ways to manipulate their composition. J. Insects Food Feed 2021, 7, 639–660. [Google Scholar] [CrossRef]
- Oonincx, D.G.A.B.; van Itterbeeck, J.; Heetkamp, M.J.; van den Brand, H.; van Loon, J.J. An exploration on greenhouse gas and ammonia production by insect species suitable for animal or human consumption. PLoS ONE 2010, 5, e14445. [Google Scholar] [CrossRef] [PubMed]
- Borah, N.; Hazarika, L.K. Biology and morphometrics of Periplaneta americana. J. Entomol. Zool. Stud. 2019, 7, 1206–1210. [Google Scholar]
- Boate, U.; Suotonye, B. Cockroach (Periplaneta americana): Nutritional value as food and feed for man and livestock. Asian J. Agric. Food Sci. 2020, 15, 37–46. [Google Scholar] [CrossRef]
- Adedara, I.A.; Mohammed, K.A.; Da-Silva, O.F.; Salaudeen, F.A.; Gonçalves, F.L.S.; Rosemberg, D.B.; Aschner, M.; Rocha, J.B.T.; Farombi, E.O. Utility of cockroach as a model organism in the assessment of toxicological impacts of environmental pollutants. Environ. Adv. 2022, 10, 100245. [Google Scholar] [CrossRef]
- Wang, Y.S.; Shelomi, M. Review of Black Soldier Fly (Hermetia illucens) as Animal Feed and Human Food. Foods 2017, 6, 91. [Google Scholar] [CrossRef]
- Serville, J.G.A. Histoire naturelle des insectes. Orthoptères; Roret: Paris, France, 1838; p. 776. [Google Scholar]
- Walker, F. Catalogue of the specimens of Blattariae in the collection of the British Museum; British Museum (Natural History): London, UK, 1868; p. 239. [Google Scholar]
- Linnaeus, C. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis, 10th ed.; Tomus I; Laurentius Salvius: Holmiae, Sweden, 1758; pp. 1–824. [Google Scholar]
- Kambhampati, S. A phylogeny of cockroaches and related insects based on DNA sequence of mitochondrial ribosomal RNA genes. Proc. Natl. Acad. Sci. USA 1995, 92, 2017–2020. [Google Scholar] [CrossRef]
- Hao, W. Biology of Blaptica dubia (Blattodea: Blaberidae). M.Sc. Thesis, 2013. [Google Scholar]
- Mariod, A.; Mirghani, M.; Hussein, I. Blaptica dubia Cockroaches. In Unconventional Oilseeds and Oil Sources; 2017; pp. 337–340. [Google Scholar]
- Gullan, P.J.; Cranston, P.S. The Insects: An Outline of Entomology, 5th ed.; Wiley-Blackwell: Oxford, UK, 2014. [Google Scholar]
- Kim, T.; Rust, K.M. Life History and Biology of the Invasive Turkestan Cockroach (Dictyoptera: Blattidae). J. Econ. Entomol. 2013, 106, 1245–1252. [Google Scholar] [CrossRef] [PubMed]
- Appel, A.G. Insect Growth and Development: Physiology, Biochemistry, and Morphology; Springer: New York, New York, USA, 2004. [Google Scholar]
- Shal, C; Gautier, J.Y; Bell, W. J Behavioural Ecology of Cockroaches. Biological reviews 1983, 59(2), 209–254. [Google Scholar] [CrossRef]
- Huang, C.Y.; Sabree, Z.L.; Moran, N.A. Genome sequence of Blattabacterium sp. strain BGIGA, endosymbiont of the Blaberus giganteus cockroach. J. Bacteriol. 2012, 194, 4450–4451. [Google Scholar] [CrossRef]
- Smith-Herron, A.J. Host specificity of five species of Eugregarinida among six species of cockroaches (Insecta: Blattodea). Comp. Parasitol. 2008, 75, 288–291. [Google Scholar] [CrossRef]
- Kambhampati, S. A phylogeny of cockroaches and related insects based on DNA sequence of mitochondrial ribosomal RNA genes. Proc. Natl. Acad. Sci. USA 1995, 92, 2017–2020. [Google Scholar] [CrossRef] [PubMed]
- Sreng, L. Cockroach mating behaviours, sex-pheromones, and abdominal glands (Dictyoptera, Blaberidae). J. Insect Behav. 1993, 6, 715–735. [Google Scholar] [CrossRef]
- European Parliament and the Council of the European Union. Regulation (EC) No 1069/2009 of 21 October 2009 laying down health rules as regards animal by-products and derived products not intended for human consumption and repealing Regulation (EC) No 1774/2002 (Animal by-products Regulation). Official Journal of the European Union L300, 14.11.2009, pp. 1–33.
- European Committee for Standardization (CEN). EN 12048:2001 – Solid fertilisers and soil improvers — Determination of water content — Gravimetric method by drying at (105 ± 2) °C; CEN: Brussels, Belgium, 2001.
- European Committee for Standardization (CEN). EN 13039:2012 – Soil improvers and growing media — Determination of organic matter content and ash; CEN: Brussels, Belgium, 2012.
- International Organization for Standardization (ISO). ISO 10390:2004 – Soil quality — Determination of pH; ISO: Geneva, Switzerland, 2004.
- European Committee for Standardization (CEN). EN 16169:2013 – Sludge, treated biowaste and soil — Determination of Kjeldahl nitrogen; CEN: Brussels, Belgium, 2013.
- International Organization for Standardization (ISO)/European Committee for Standardization (CEN). EN ISO 54321:2021; Soil, treated biowaste, sludge and waste — Fractionation of elements soluble in aqua regia. ISO/CEN: Geneva/Brussels, 2021.
- European Committee for Standardization (CEN). EN 12880:2005 – Sludge characterisation — Determination of dry residue and water content; CEN: Brussels, Belgium, 2005.
- Ashworth, A.J.; Amorim, H.C.S.; Drescher, G.L.; Moore, P.A., Jr.; Rojas, M.G.; Morales-Ramos, J.; Donoghue, A.M. Insect frass effects on soil properties and forage systems. Sci. Rep. 2025, 15, 87075. [Google Scholar]
- Lomonaco, G.; Franco, A.; De Smet, J.; Scieuzo, C.; Salvia, R.; Falabella, P. Larval frass of Hermetia illucens as organic fertilizer. Insects 2024, 15, 293. [Google Scholar] [CrossRef] [PubMed]
- Lavelle, P.; Spain, A.V. Soil Ecology; Kluwer Academic Publishers: New York, New York, USA, 2001. [Google Scholar]
- Hillel, D. Introduction to Environmental Soil Physics; Academic Press: New York, New York, USA, 2004. [Google Scholar]
- Othman, N.; Irwan, J.M.; Roslan, M. Vermicomposting of Food Waste. Int. J. Integr. Eng. 2012, 4, 39–48. [Google Scholar]
- Jadia, C.D.; Fulekar, M.H. Phytoremediation of Heavy Metals: Recent Techniques. Afr. J. Biotechnol. 2009, 8, 921–928. [Google Scholar]
- Lopes, I.G.; Brandon, M.G.; Praeg, N.; Claeys, J.; Yakti, W.; Bitterlich, M.; Jones, J.J.; Geilfus, C.M.; Klammsteiner, T. Critical considerations for evaluating insect frass composition and soil interactions. J. Insects Food Feed 2025, pp 1–28. [Google Scholar]
- Singhal, N.; Kumar, M.; Kanaujia, P.K.; Virdi, J.S. MALDI-TOF mass spectrometry: an emerging technology for microbial identification and diagnosis. Front. Microbiol. 2015, 6, 791. [Google Scholar] [CrossRef] [PubMed]
- Wada-Katsumata, A.; Zurek, L.; Nalyanya, G.; Roelofs, W.L.; Zhang, A.; Schal, C. Gut bacteria mediate aggregation in the German cockroach. Proc. Natl. Acad. Sci. USA 2015, 112, 15678–15683. [Google Scholar] [CrossRef]
- Ademolu, K.O.; Mustapha, O.Y.; Idowu, A.B. Nutritional and gut microbial analyses of adult male cockroaches (Periplaneta americana, Dictyoptera, Blattodea) from three locations in Abeokuta, Nigeria. Entomol. Romanica 2020, 24, 19–23. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiao, X.; Elhag, O.; Cai, M.; Zheng, L.; Huang, F.; Jordan, H.R.; Tomberlin, J.K.; Sze, S.H.; Yu, Z.; Zhang, J. Hermetia illucens L. larvae-associated intestinal microbes reduce the transmission risk of zoonotic pathogens in pig manure. Microb. Biotechnol. 2022, 15, 2631–2644. [Google Scholar] [CrossRef]
Figure 1.
B. dubia male and female.

Figure 2.
B. lateralis male and female.

Figure 3.
B. giganteus male and female.

Figure 4.
Example of the arrangement of a tank for the species B. dubia.

Figure 7.
Normalised macronutrient profiles of frass samples.
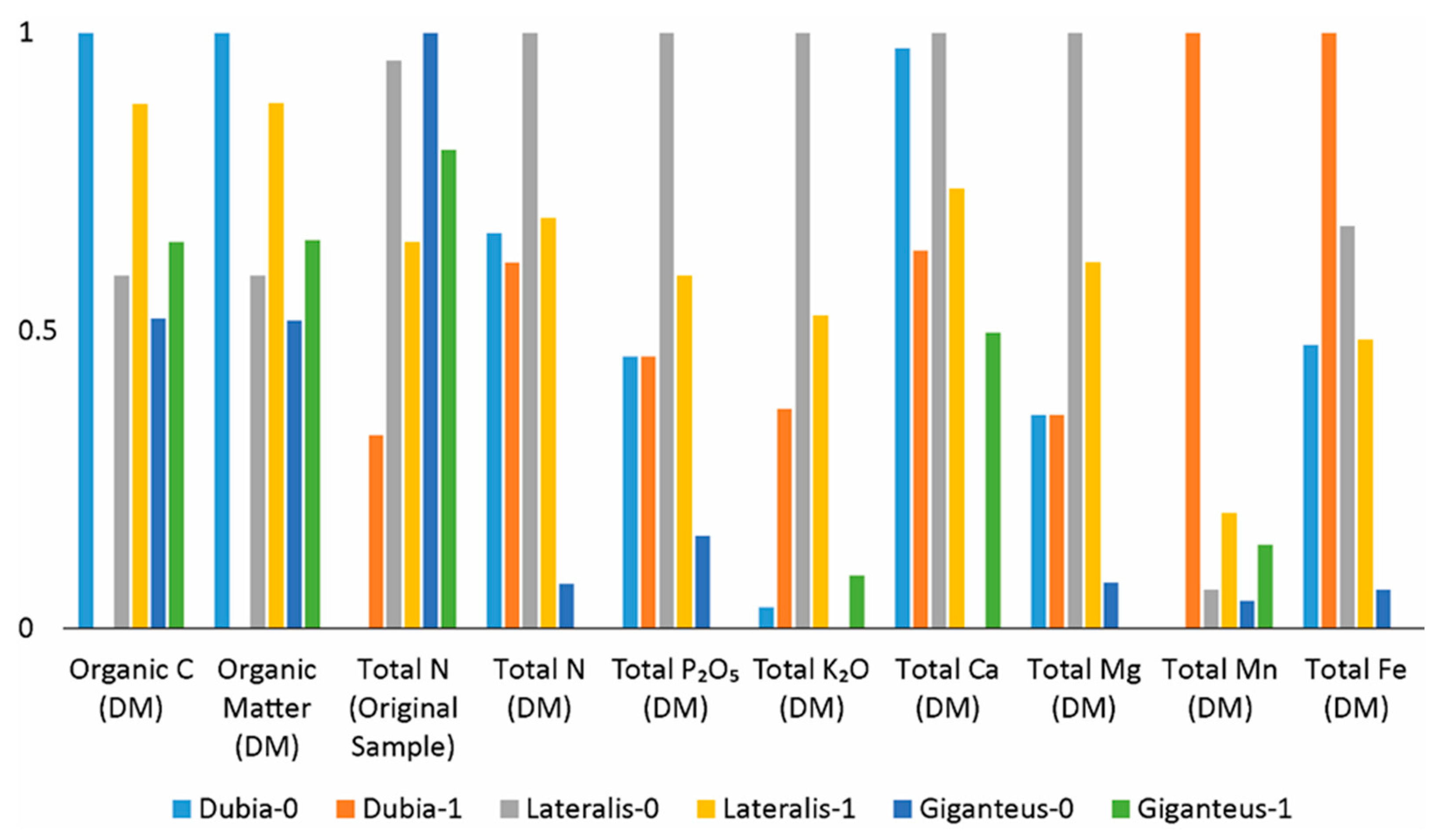
Figure 8.
Heatmap of Pearson correlation coefficients among chemical parameters of frass.
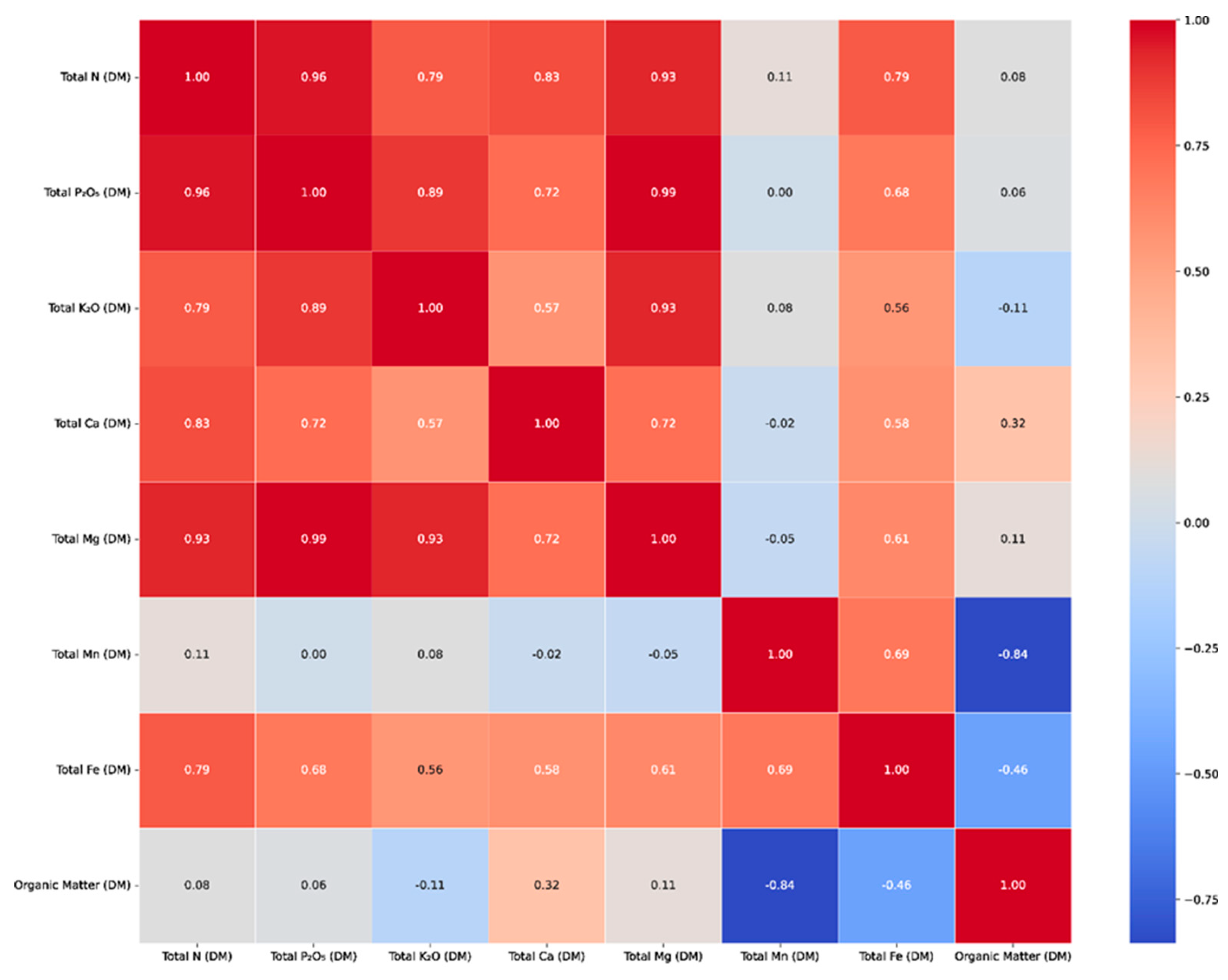
Figure 9.
Microelements of the analysed cochroach frass samples.
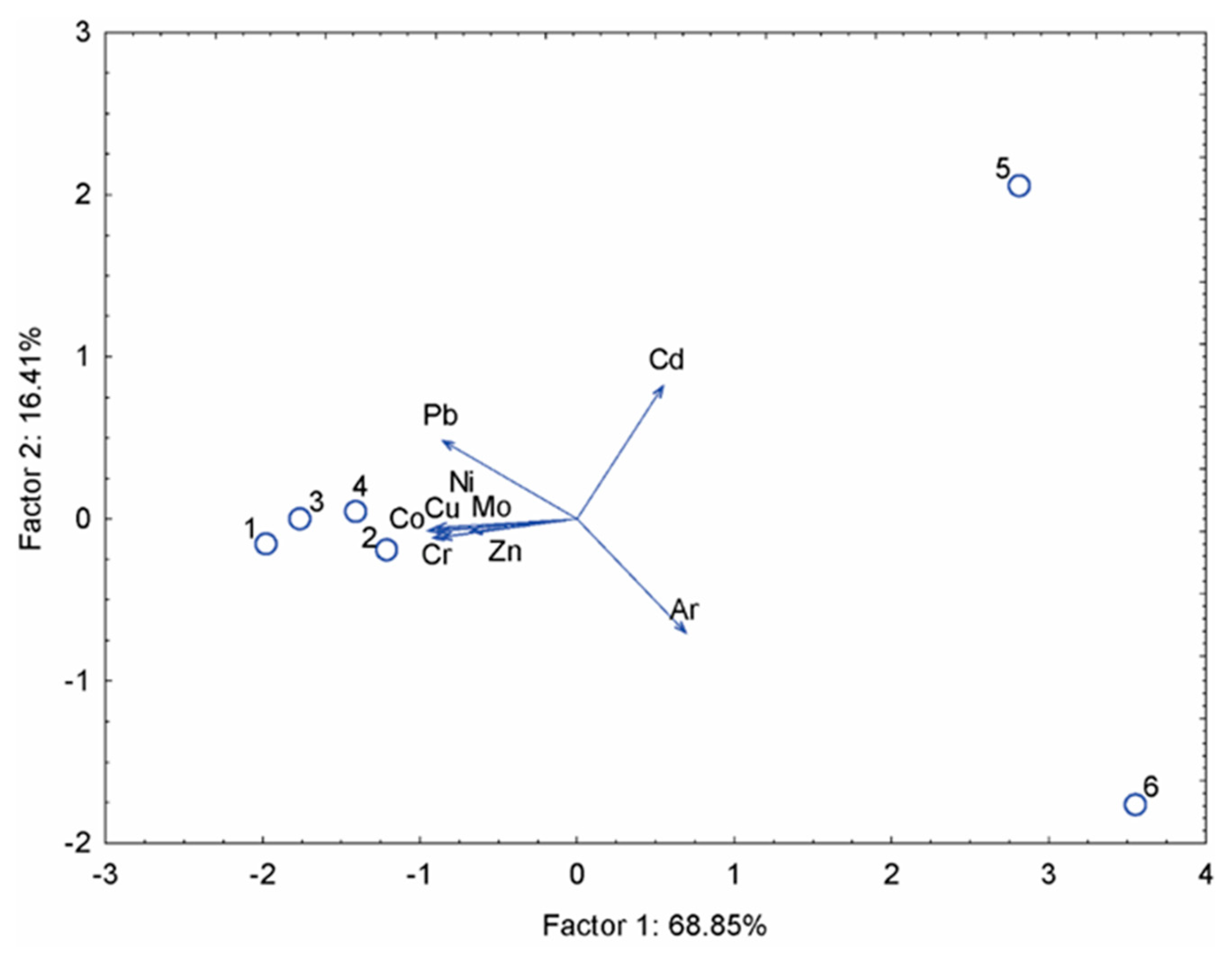
Table 1.
Explanation of the PCA sample numbers displayed in the PCA1 and PCA2 biplots.
| PCA No. | Type x Parameter | |
|---|---|---|
| 1 | B. dubia | 0 |
| 2 | B. dubia | 1 |
| 3 | B. lateralis | 0 |
| 4 | B. lateralis | 1 |
| 5 | B. giganteus | 0 |
| 6 | B. giganteus | 1 |
0 – control; 1 - Slaughterhouse waste.
Table 2.
Basic chemical parameters of frass (mean values ± SD).
| Cockroach species | B. dubia | B. lateralis | B. giganteus | Statistically significant |
|||
| Type of feed | Control group |
Slaughterhouse waste |
Control group |
Slaughterhouse waste |
Control group |
Slaughterhouse waste |
|
| MC (%) | 73.51±0.01 | 61.33±0.04 | 50.35±0.01 | 52.01±0.01 | 9.66±0.15 | 14.48±0.04 | * |
| Dry matter (%) | 26.52±0.06 | 38.65±0.01 | 49.47±0.24 | 47.89±0.13 | 90.21±0.04 | 85.4±0.14 | * |
| pH | 5.04±0.05 | 6.91±0.01 | 7.84±0.05 | 7.35±0.01 | 6.98±0.01 | 7.04±0.01 | * |
| E.C. mS/cm | 2.07±0.01 | 2.97±0.02 | 3.53±0.01 | 3.87±0.01 | 0.27±0.03 | 0.24±0.01 | * |
| Ash (%) | 13.93±0.06 | 34.69±0.06 | 22.36±0.08 | 16.39±0.05 | 23.9±0.14 | 21.1±0.14 | * |
| LOI (%) | 86.06±0.04 | 65.31±0.06 | 77.62±0.04 | 83.63±0.07 | 76.05±0.07 | 78.75±0.07 | * |
*Statistically significant at p < 0.01.
Table 3.
Macronutrient and elemental composition of frass.
| Cockroach species | B. dubia | B. lateralis | B. giganteus | Statistically significant |
|||
| Type of feed | Control group |
Slaughterhouse waste |
Control group |
Slaughterhouse waste |
Control group |
Slaughterhouse waste |
|
| C (organic) (dm) (%) | 48.24±0.08 | 36.57±0.02 | 43.46±0.01 | 46.55±0.35 | 42.55±0.07 | 44.05±0.07 | * |
| Organic matter (dm) (%) | 86.02±0.02 | 65.14±0.19 | 77.55±0.06 | 83.69±0.16 | 75.95±0.07 | 78.7±0.14 | * |
| N (total-original sample) (%) | 0.38±0.01 | 0.53±0.01 | 0.87±0.01 | 0.70±0.01 | 0.86±0.01 | 0.78±0.01 | * |
| N (total) (dm) (%) | 1.44±0.01 | 1.38±0.04 | 1.71±0.01 | 1.46±0.01 | 0.96±0.01 | 0.91±0.01 | * |
| P2O5 (total) (dm) (%) | 0.30±0.01 | 0.28±0.02 | 0.36±0.01 | 0.31±0.01 | 0.24±0.01 | 0.22±0.01 | * |
| K2O (total) (dm) (%) | 2.06±0.06 | 2.21±0.01 | 2.56±0.01 | 2.29±0.01 | 1.98±0.01 | 2.03±0.02 | * |
| Ca (total) (dm) (%) | 0.77±0.01 | 0.65±0.01 | 0.78±0.01 | 0.67±0.02 | 0.40±0.01 | 0.59±0.01 | * |
| Mg (total) (dm) (%) | 0.15±0.01 | 0.15±0.01 | 0.21±0.01 | 0.16±0.01 | 0.13±0.01 | 0.12±0.01 | * |
| Mn (total) (dm) (%) | 1.21±0.01 | 5.53±0.04 | 1.51±0.01 | 2.06±0.01 | 1.43±0.01 | 1.83±0.01 | * |
| Fe (total) (dm) (g/kg) | 56.73±0.04 | 74.43±0.04 | 63.41±0.01 | 57.05±0.07 | 42.84±0.05 | 40.63±0.04 | * |
*Statistically significant at p < 0.01.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



