
Submitted:
28 January 2026
Posted:
29 January 2026
You are already at the latest version
Abstract
Median overall survival (OS) is a useful summary statistic in oncology, but it can obscure a clini-cally important and recurring phenomenon: a non-negligible fraction of patients with advanced (stage IV) solid tumors survive far beyond the median, forming a long tail on the survival curve. Accumulating mechanistic evidence indicates that tumor microenvironment (TME) acido-sis—driven by aerobic glycolysis, hypoxia, and proton-export systems (e.g., NHE1, monocarbox-ylate transporters, carbonic anhydrase IX, and V-ATPase)—promotes invasion, therapy resistance, and immune suppression. Buffering and other alkalization strategies can partially neutralize acidic TME in preclinical models and may enhance antitumor immunity, suggesting that modulating extracellular pH (pHe) is a plausible system-level lever. Science-based medicine (SBM) provides a complementary framework to evidence-based medicine (EBM) by integrating population-level evidence with causal mechanisms and longitudinal patient data, viewing cancer as a complex, non-equilibrium system with multiple attractor states. From this perspective, long-tail survival is not merely an outlier but may represent a qualitative state transition of the tumor–host system. We summarize key biology of TME acidosis, review alkalization-oriented interventions and their translational challenges, and discuss implications for trial design, including endpoints and analyses that capture delayed separation of survival curves and tail effects.
Keywords:
tumor microenvironment
; acidosis
; extracellular pH
; immunotherapy
; long-tail survival
; science-based medicine
1. Introduction
Despite remarkable advances in oncology over the past several decades, advanced cancers—particularly stage IV solid tumors—have long been regarded as “incurable” within the framework of evidence-based medicine (EBM) grounded in randomized controlled trials (RCTs). A representative metric supporting this view is the median overall survival (median OS), which has functioned as an indispensable summary statistic for drug approval and the development of clinical guidelines (hereafter, overall survival is abbreviated as OS). However, survival curves in advanced cancer often show a steep early decline followed by a more gradual tail, and a reproducible clinical phenomenon is observed whereby a non-negligible fraction of patients survive far beyond the median. This “long tail” in the survival curve is not merely a statistical outlier; rather, it may represent a clinically meaningful signal suggestive of prolonged disease stability, a state approaching functional remission, or the absence of irreversible breakdown in treatment resistance. By definition, median OS is determined solely by the time point at which 50% of a cohort has died, and therefore it structurally fails to capture information about tail behavior. As a result, the clinical meaning implied by long-tail survival is addressed only inadequately within a median OS–centered framework.
Metabolic reprogramming in cancer (the Warburg effect) together with hypoxia promotes the production and export of lactate and protons, thereby lowering extracellular pH (pHe). An acidic tumor microenvironment (TME) has been shown to simultaneously drive invasion/metastasis, therapy resistance, and immune suppression, functioning as a hub for malignant phenotypes. Accordingly, interventions that shift extracellular pH toward a more physiological range may be positioned as a “system-level lever,” distinct from reductionist approaches that target individual molecules. In addition, science-based medicine (SBM) offers a framework that integrates causal mechanisms grounded in basic science/biomedicine with longitudinal, patient-level data, complementing population-average inference in EBM and viewing cancer as a complex non-equilibrium open system. In this review, SBM is not proposed as a replacement for EBM; rather, it is positioned as an operational framework for generating testable clinical hypotheses by integrating mechanistic causality, longitudinal patient-level data, and system-level dynamics. From an SBM perspective, long-tail survival is not an exception but may indicate that the tumor–host system has transitioned to another stable state (an attractor state). [1,2,3,4,5,6,7]
In this narrative review, we aim to make visible the information obscured by median OS and to focus on TME acidosis and its neutralization (“alkalization”) as a biological framework that could help explain long-tail survival. We summarize the mechanisms underlying TME acidosis, synthesize the preclinical and clinical evidence and challenges for alkalization-oriented interventions, and discuss endpoints and trial-design considerations to capture long-term survival (tail effects). We also position urinary pH as a clinically feasible longitudinal marker while explicitly stating its limitations. In this manuscript, “alkalization” does not refer to an attempt to directly raise blood pH; rather, it denotes a framework that, based on evidence that TME acidosis may contribute to invasion, immune suppression, and therapy resistance, examines the potential for buffering and diet-based interventions to influence TME acidosis and pH-related outcomes (including urinary pH).
2. Limitations of Median Overall Survival in Advanced Cancer
Median OS is robust and intuitive; however, it can average out the heterogeneity of advanced cancer and thereby obscure clinically decisive differences. Patients with advanced cancer are not biologically uniform: tumor burden, host inflammation, nutritional status, treatment responsiveness, and immune competence jointly shape distinct subgroups within the same clinical stage. When Kaplan–Meier curves exhibit a biphasic pattern, the steep early decline may reflect high tumor burden and/or unfavorable host conditions, whereas the later, more gradual tail may reflect a subgroup that achieves durable disease control. Nevertheless, the median is heavily influenced by the time window in which events are concentrated and does not adequately reflect changes in the proportion of long-term survivors that emerge in the tail.
Similarly, commonly used hypothesis tests such as the log-rank test and the Gehan–Breslow–Wilcoxon test may undervalue delayed and tail effects because their weighting is biased toward periods with a higher event density. In the delayed separation or biphasic survival curves typically observed with immunotherapy, the tail can be lifted even when the median changes only modestly; thus, interpretations centered on the median risk missing the therapy’s most consequential clinical value. To address this blind spot, it is important to complement the median with milestone survival pre-specified at clinically meaningful time points (e.g., 3- or 5-year survival), restricted mean survival time (RMST; with τ pre-defined according to feasible follow-up, such as 3 or 5 years), conditional survival probabilities focused on the tail, and modeling approaches that incorporate a cure fraction, such as mixture cure models. These frameworks help characterize the overall shape of the survival curve and its terminal behavior. In particular, when discussing delayed separation and tail effects observed with immunotherapy, this family of metrics provides an essential interpretive foundation. [8,9]
3. Science-Based Medicine as a Framework
Science-based medicine (SBM) does not reject the population-average inference provided by evidence-based medicine (EBM); rather, it complements and extends EBM by clarifying its scope and limitations and by incorporating causal mechanisms and longitudinal data from individual patients. In this review, SBM refers to an integrative framework that starts from causal mechanisms grounded in basic science and biomedicine to reinterpret and redesign clinical phenomena, and then integrates population-level evidence with patient-level longitudinal data to generate and refine testable hypotheses.
Cancer is not merely a collection of genetic mutations but a dynamic system governed by multilayered feedback loops involving metabolism, perfusion, immunity, inflammation, nutrition, and acid–base regulation. Thermodynamically, such a system can be viewed as a non-equilibrium open system that, even under perturbations, tends to converge toward specific stable states (attractor states). From the standpoint of non-equilibrium thermodynamics, the entropy balance of such systems can be expressed as follows:
ΔSsystem = Sin − Sout + Sgen.
Here, Sin and Sout denote entropy inflow and outflow across the system boundary, respectively, and Sgen is the entropy generated by irreversible processes within the system. The second law requires Sgen ≥ 0. In an open system, however, net entropy export can exceed internal entropy generation, so ΔSsystem is not constrained to be positive. Accordingly, living systems—including the tumor–host system—can maintain their organization by exchanging heat, matter, and information with the environment, and under changing conditions may transition to alternative stable states
An acidic TME represents one such prototypical attractor. Tumor cells produce lactate and protons through aerobic glycolysis and rewire transport and buffering programs—such as the Na+/H+ exchanger (NHE1), monocarboxylate transporters (MCT1/4), carbonic anhydrase IX (CAIX), and V-ATPase—to preserve intracellular pH (pHi) while imposing an acid load on the extracellular space. In addition, abnormal vasculature and hypoxia impair acid washout, thereby sustaining an acidic milieu with pHe typically around 6.5–6.9. This process is better understood not merely as the accumulation of metabolic byproducts, but as a self-maintaining “operating mode” in which metabolic reactions (lactate/proton production) are coupled to membrane transport and buffering (e.g., NHE1, MCT1/4, CAIX, and V-ATPase).
In other words, tumor cells dissipate free energy derived from nutrients and oxygen through irreversible processes while exporting acid to maintain intracellular pH homeostasis, thereby establishing and sustaining an extracellular pH gradient (acidic TME). A state in which sustained gradients and fluxes are maintained—under continuous inflow of matter and energy—through internal rewiring of reactions and transport can be positioned as a form of dissipative structure in a non-equilibrium open system.
This acidic state can self-reinforce invasion/metastasis, therapy resistance, and immune suppression, thereby stabilizing the tumor–host system toward a malignant regime. From the perspective of a non-equilibrium open system, interactions among metabolism, perfusion, immunity, and acid–base conditions can generate stable states that tend to preserve characteristic behaviors (attractor states). Acidic TME can be understood as an attractor maintained by mutually reinforcing coupling among glycolytic dominance, hypoxia, acid export, immune suppression, and invasion-promoting programs. When interventions or host-condition changes weaken these couplings, a transition to another stable state (state transition) may occur. Therefore, from an SBM perspective, long-tail survival is not merely a statistical outlier; it can be hypothesized as indicating that the tumor–host system has shifted into a different state region, a proposition that should be evaluated through longitudinal data integrated with mechanistic reasoning (Figure 1). [6,10,11,12,13,14,15,16,17,18]
4. Tumor Microenvironment Acidosis
TME acidosis is not merely a byproduct of metabolism; it can be a functional adaptation that is selectable as part of cancer’s survival strategy [20]. From the non-equilibrium open-system perspective described above, TME acidosis can be understood as a self-sustaining flux state in which metabolic production is coupled to membrane transport and perfusion. In this section, we organize the major components of this process based on mechanistic evidence.
Increased lactate production via aerobic glycolysis, hypoxia-driven metabolic shifts, and impaired acid clearance due to reduced perfusion jointly lead to the accumulation of lactate and protons in the extracellular space. The Na+/H+ exchanger 1 (NHE1) exports intracellular protons to maintain intracellular pH on the relatively alkaline side while imposing an extracellular acid load. Monocarboxylate transporters (MCTs) co-transport lactate and protons, carbonic anhydrase IX (CAIX) promotes extracellular acidification through the CO2 hydration reaction, and V-ATPase also contributes to proton export. As a result, tumors develop a pH gradient characterized by relatively elevated intracellular pH and decreased extracellular pH, creating conditions favorable for proliferation and invasion.
An acidic TME contributes to local invasion and metastasis. Acidic conditions can promote local invasion through activation of matrix-degrading enzymes and enhanced cell motility, and may increase metastatic potential by selecting for acid-resistant phenotypes. Moreover, acidity is implicated in therapy resistance. Uptake of weakly basic drugs into cells can be reduced, intratumoral drug distribution may become unfavorable, and adaptive responses to therapeutic stress can be promoted. From an immunological perspective, lactate and low pHe have been shown to suppress effector functions of T cells and NK cells, thereby blunting tumor immune surveillance [21,22]. Collectively, TME acidosis serves as an integrative hub linking “invasion/metastasis,” “therapy resistance,” and “immune suppression,” providing a rational target for system-level interventions. [3,4,5,6,7,10,11,12,21,23,24]
5. Alkalization Approaches
If an acidic TME self-reinforces malignant phenotypes, then partial neutralization of this acidity is a rational therapeutic strategy. Preclinical studies have shown that buffering approaches using bicarbonate and related agents can increase extratumoral pHe and may suppress metastasis. Moreover, neutralizing an acidic environment has been reported to restore aspects of immune function and potentially enhance the efficacy of immunotherapy. From a translational standpoint, interventions can be broadly organized into (i) orally administered alkalizing agents that increase systemic buffering capacity (e.g., bicarbonate and citrate salts) and (ii) lifestyle interventions that include dietary modification. However, intratumoral pHi is spatially and temporally heterogeneous and does not map simply onto systemic acid–base balance. In addition, dose optimization, safety (e.g., electrolyte disturbances, volume and sodium load, renal function), drug–drug interactions, and objective biomarkers demonstrating whether “local tumor pHe actually changed” remain key translational challenges.
From the same perspective, buffering and dietary interventions may be positioned as system-level “levers” that could weaken the couplings sustaining the acidic TME attractor (e.g., hypoxia, acid export, and immune suppression). Nevertheless, repeatedly measuring local tumor pHe in humans is difficult, and this proposed chain should be translated into testable predictions that can be empirically evaluated. In addition, urinary pH is one of the most practical longitudinal markers in current clinical practice because it can be measured repeatedly; it may serve as a proximal indicator of systemic acid–base status and/or adherence to the intervention. However, physiological validation is currently insufficient to assert that changes in urinary pH directly represent changes in human tumor extracellular pH (TME pH). Accordingly, urinary pH should be interpreted longitudinally not so much as a surrogate of local tumor pH, but rather as a pH-related outcome/proximal marker. SBM provides a framework to integrate such proximal markers as patient-level time series and to evaluate the hypothesized chain—intervention → acid–base environmental change → immune/metabolic change → clinical outcomes—based on testable predictions (Figure 1). [7,23,25,26,27]
We previously reported that increases in urinary pH during alkalization therapy were associated with improved survival and more favorable responses to chemotherapy in patients with various cancers [28]. Furthermore, higher urinary pH has been linked to greater fruit and vegetable intake, suggesting that urinary pH may serve as a useful biomarker reflecting changes in dietary acid load (proton excretion) [29].
6. Long-Tail Survival as a State Transition
A recurring feature among long-term survivors is not necessarily an early dramatic response, but rather durable stabilization that emerges over time. Even in the absence of radiographic complete response, some patients appear to shift toward a near-dormant state in which the tumor can coexist with the host over prolonged periods while maintaining quality of life [30]. Discordance between early response and long-term outcome is also not uncommon, and long-term prognosis cannot be readily explained by short-term tumor shrinkage alone. These clinical patterns suggest that outcomes may be determined less by tumor volume per se than by the conditions of the TME and the host’s immune and metabolic milieu.
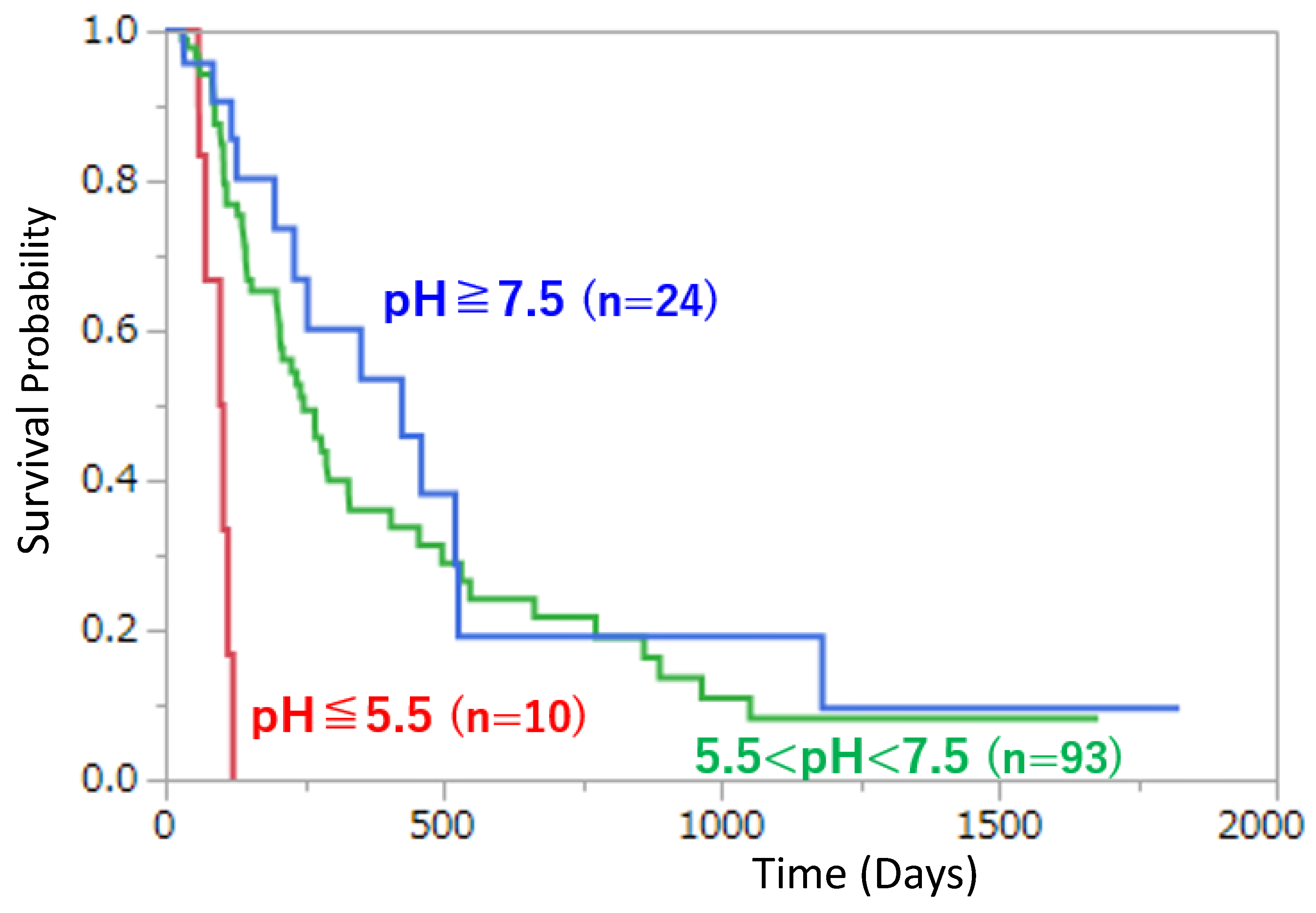
Within the SBM framework, we hypothesize that a long tail on the survival curve can arise when the tumor–host system transitions from an acidified, glycolysis-dominant attractor to another attractor in which extracellular pHe is closer to neutral and immune function is more effectively maintained. As a concrete clinical illustration, the Kaplan–Meier curves reported for stage IV pancreatic cancer in Oncologie, stratified by urinary pH, show an early truncation of the survival curve in the low–urinary pH group (pH ≤ 5.5), whereas a longer survival tail persists in the other groups [31]. Between-group differences are statistically clear (log-rank test) and the long-tail pattern is visually apparent (Figure 2).
However, this analysis is observational and may be influenced by confounding factors, including disease burden, treatment intensity, nutritional status, renal function, and measurement frequency. Therefore, urinary pH should be treated not as a single (one-time) value but as a longitudinal trajectory, and its fluctuations accompanying treatment changes or shifts in disease status should be modeled. Combining robust summary measures (e.g., trimmed means) with time-varying covariate models can help prioritize the “qualitative differences” that emerge in the tail as signals for hypothesis generation, which can then be tested by integrating longitudinal metabolic, inflammatory, and immunologic markers—an approach that captures the core operational intent of SBM.
If increases in urinary pH show no association with immune or inflammatory markers, support for the model would be weakened. Moreover, if tail formation is not reproducible under pH-related interventions, the attractor hypothesis would need to be reconsidered. Nevertheless, as suggested by the clear association observed in Figure 2, the tail structure provides a clinically compelling signal for further mechanistically informed, longitudinal validation. [31,32,33,34]
7. Implications for Clinical Trial Design
To detect and interpret long-tail effects, endpoints and analysis plans that can evaluate delayed separation and tail behavior are needed in addition to median OS and hazard ratios. Milestone survival at 3 and 5 years, RMST (with τ pre-specified), and approaches that explicitly consider a cure fraction provide a useful reference framework to avoid missing delayed separation/tail effects often seen with immunotherapy. Candidate approaches include pre-specified landmark survival rates, RMST, conditional survival probabilities focused on the tail, and mixture models incorporating a cure concept.
Interventions aimed at neutralizing acidic TME may manifest not as immediate maximization of tumor shrinkage, but rather as delayed separation by alleviating metabolic and immune stress and thereby enhancing the effectiveness of standard therapies. Accordingly, it is important to ensure sufficiently long follow-up and to incorporate analyses that anticipate delayed effects. From a stratification standpoint, beyond conventional histology and molecular markers, it is desirable to incorporate system-level indicators such as acid–base balance, lactate metabolism, inflammation, and immune competence. When urinary pH is used, it should be handled not as a single measurement but as a longitudinal trajectory (e.g., using robust summaries such as trimmed means), and variability associated with treatment changes or shifts in disease status should be modeled. Furthermore, because alkalization interventions may be more likely to expand the therapeutic window when combined with immunotherapy or chemotherapy rather than acting as monotherapy, trial designs that prespecify combination strategies are rational. Trial design informed by SBM calls for evaluation not only of statistical significance, but also of mechanistic readouts—namely, the extent to which the intervention alters TME, metabolic, and immune states—so that the overall effect can be assessed as a plausible causal chain.
8. Conclusions
The long tail that repeatedly appears in survival curves of advanced cancer contains clinical and biological information that cannot be fully captured by median OS alone. Acidic TME is a central factor integrating invasion/metastasis, therapy resistance, and immune suppression, and interventions aimed at its partial neutralization merit consideration as system-level therapeutic strategies. SBM provides a complementary framework that preserves the rigor of EBM while integrating mechanisms and longitudinal data to explain and test long-tail survival. Future research should integrate methods to assess local tumor pHe, safely optimize interventions, develop combination strategies, and adopt analytic approaches capable of capturing tail effects, thereby enabling study designs that evaluate long-tail formation as a reproducible phenomenon.
Author Contributions
Conceptualization, H.Wada. and K.Suzuki.; Methodology, K. Suzuki., H. Wada., and S.Kachi; Writing—original draft, K.Suzuki.; Writing—review and editing, H. Wada., S. Kachi and K.Suzuki; Supervision, H. Wada. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable.
Acknowledgments
The authors would like to express their sincere gratitude to Dr. Reo Hamaguchi and Dr. Hiromasa Morikawa for their valuable discussions and insightful comments that contributed to the development of the concepts and perspectives presented in this narrative review.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg Effect: The Metabolic Requirements of Cell Proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [PubMed]
- Corbet, C.; Feron, O. Tumour Acidosis: From the Passenger to the Driver’s Seat. Nat. Rev. Cancer 2017, 17, 577–593. [Google Scholar] [CrossRef]
- Gatenby, R.A.; Gillies, R.J. Why Do Cancers Have High Aerobic Glycolysis? Nat. Rev. Cancer 2004, 4, 891–899. [Google Scholar] [CrossRef]
- Parks, S.K.; Chiche, J.; Pouysségur, J. pH Control Mechanisms of Tumor Survival and Growth. J. Cell. Physiol. 2011, 226, 299–308. [Google Scholar] [CrossRef]
- Ibrahim-Hashim, A.; Estrella, V. Acidosis and Cancer: From Mechanism to Neutralization. Cancer Metastasis Rev. 2019, 38, 149–155. [Google Scholar] [CrossRef]
- Schadendorf, D.; Hodi, F.S.; Robert, C.; Weber, J.S.; Margolin, K.; Hamid, O.; Patt, D.; Chen, T.-T.; Berman, D.M.; Wolchok, J.D. Pooled Analysis of Long-Term Survival Data from Phase II and Phase III Trials of Ipilimumab in Unresectable or Metastatic Melanoma. J. Clin. Oncol. 2015, 33, 1889–1894. [Google Scholar] [CrossRef]
- Daud, A.I.; Wolchok, J.D.; Robert, C.; Hwu, W.-J.; Weber, J.S.; Ribas, A.; Hodi, F.S.; Joshua, A.M.; Kefford, R.; Hersey, P.; et al. Programmed Death-Ligand 1 Expression and Response to the Anti–PD-1 Antibody Pembrolizumab in Melanoma. J. Clin. Oncol. 2016, 34, 4102–4109. [Google Scholar] [CrossRef]
- Swietach, P.; Vaughan-Jones, R.D.; Harris, A.L.; Hulikova, A. The Chemistry, Physiology and Pathology of pH in Cancer. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130099. [Google Scholar] [CrossRef]
- Cardone, R.A.; Casavola, V.; Reshkin, S.J. The Role of Disturbed pH Dynamics and the Na+/H+ Exchanger in Metastasis. Nat. Rev. Cancer 2005, 5, 786–795. [Google Scholar] [CrossRef] [PubMed]
- Halestrap, A.P.; Price, N.T. The Proton-Linked Monocarboxylate Transporter (MCT) Family: Structure, Function and Regulation. Biochem. J. 1999, 343, 281–299. [Google Scholar] [CrossRef] [PubMed]
- Prigogine, I. From Being to Becoming: Time and Complexity in the Physical Sciences; W.H. Freeman: San Francisco, CA, USA, 1980. [Google Scholar]
- Nicolis, G.; Prigogine, I. Self-Organization in Nonequilibrium Systems: From Dissipative Structures to Order through Fluctuations; Wiley: New York, NY, USA, 1977. [Google Scholar]
- Waddington, C.H. The Strategy of the Genes; Allen & Unwin: London, UK, 1957. [Google Scholar]
- Huang, S. Cell Lineage Determination in State Space: A Systems View Brings Flexibility to Dogmatic Canonical Rules. PLoS Biol. 2010, 8, e1000380. [Google Scholar] [CrossRef] [PubMed]
- Kitano, H. Systems Biology: A Brief Overview. Science 2002, 295, 1662–1664. [Google Scholar] [CrossRef]
- Anderson, A.R.A.; Quaranta, V. Integrative Mathematical Oncology. Nat. Rev. Cancer 2008, 8, 227–234. [Google Scholar] [CrossRef]
- Gatenby, R.A.; Brown, J.S. Integrating Evolutionary Dynamics into Cancer Therapy. Nat. Rev. Clin. Oncol. 2020, 17, 675–686. [Google Scholar] [CrossRef]
- Faubert, B.; Solmonson, A.; DeBerardinis, R.J. Metabolic Reprogramming and Cancer Progression. Science 2020, 368, eaaw5473. [Google Scholar] [CrossRef]
- Brand, A.; Singer, K.; Koehl, G.E.; Kolitzus, M.; Schoenhammer, G.; Thiel, A.; Matos, C.; Bruss, C.; Klobuch, S.; Peter, K.; et al. LDHA-Associated Lactic Acid Production Blunts Tumor Immunosurveillance by T and NK Cells. Cell Metab. 2016, 24, 657–671. [Google Scholar] [CrossRef]
- Fischer, K.; Hoffmann, P.; Voelkl, S.; Meidenbauer, N.; Ammer, J.; Edinger, M.; Gottfried, E.; Schwarz, S.; Rothe, G.; Hoves, S.; et al. Inhibitory Effect of Tumor Cell–Derived Lactic Acid on Human T Cells. Blood 2007, 109, 3812–3819. [Google Scholar] [CrossRef]
- Gillies, R.J.; Pilot, C.; Marunaka, Y.; Fais, S. Targeting Acidity in Cancer and Diabetes. Biochim. Biophys. Acta Rev. Cancer 2019, 1871, 273–280. [Google Scholar] [CrossRef]
- Estrella, V.; Chen, T.; Lloyd, M.; Wojtkowiak, J.; Cornnell, H.H.; Ibrahim-Hashim, A.; Bailey, K.; Balagurunathan, Y.; Rothberg, J.M.; Sloane, B.F.; et al. Acidity Generated by the Tumor Microenvironment Drives Local Invasion. Cancer Res. 2013, 73, 1524–1535. [Google Scholar] [CrossRef] [PubMed]
- Calcinotto, A.; Filipazzi, P.; Grioni, M.; Iero, M.; De Milito, A.; Ricupito, A.; Cova, A.; Canese, R.; Jachetti, E.; Rossetti, M.; et al. Modulation of Microenvironment Acidity Reverses Anergy in Human and Murine Tumor-Infiltrating T Lymphocytes. Cancer Res. 2012, 72, 2746–2756. [Google Scholar] [CrossRef] [PubMed]
- Pilon-Thomas, S.; Kodumudi, K.N.; El-Kenawi, A.E.; Russell, S.; Weber, A.M.; Luddy, K.; Damaghi, M.; Wojtkowiak, J.W.; Mulé, J.J.; Ibrahim-Hashim, A.; et al. Neutralization of Tumor Acidity Improves Antitumor Responses to Immunotherapy. Cancer Res. 2016, 76, 1381–1390. [Google Scholar] [CrossRef] [PubMed]
- Robey, I.F.; Baggett, B.K.; Kirkpatrick, N.D.; Roe, D.J.; Dosescu, J.; Sloane, B.F.; Hashim, A.I.; Morse, D.L.; Raghunand, N.; Gatenby, R.A.; et al. Bicarbonate Increases Tumor pH and Inhibits Spontaneous Metastases. Cancer Res. 2009, 69, 2260–2268. [Google Scholar] [CrossRef]
- Isowa, M.; Hamaguchi, R.; Narui, R.; Morikawa, H.; Okamoto, T.; Wada, H. Potential of Alkalization Therapy for the Management of Metastatic Pancreatic Cancer: A Retrospective Study. Cancers (Basel) 2024, 16(1), 61. [Google Scholar] [CrossRef]
- Welch, A.A.; Mulligan, A.; Bingham, S.A.; Khaw, K.-T. Urine pH is an indicator of dietary acid-base load, fruit and vegetables and meat intakes: results from the European Prospective Investigation into Cancer and Nutrition (EPIC)-Norfolk population study. Br. J. Nutr. 2008, 99, 1335–1343. [Google Scholar] [CrossRef]
- Mollica, M.A.; Nekhlyudov, L.; et al. Survivorship for Individuals Living with Advanced and Metastatic Cancer: A Synopsis and Call for Research. J. Natl. Cancer Inst. 2022, 114, 489–496. [Google Scholar] [CrossRef]
- Kachi, S.; Hamaguchi, R.; Narui, R.; Morikawa, H.; Okamoto, T.; Wada, H. Cancer Can Be Suppressed by Alkalizing the Tumor Microenvironment: The Effectiveness of “Alkalization Therapy” in Cancer Treatment. Oncologie 2025, 27, 393–401. [Google Scholar] [CrossRef]
- Hamaguchi, R.; Narui, R.; Morikawa, H.; Okamoto, T.; Uemoto, S.; Wada, H. Meaning and Significance of “Alkalization Therapy for Cancer”. Front. Oncol. 2022, 12, 920843. [Google Scholar] [CrossRef]
- Wada, H.; Hamaguchi, R.; Uemoto, S. How Does Cancer Occur? How Should It Be Treated? Treatment from the Perspective of Alkalization Therapy Based on Science-Based Medicine. Biomedicines 2024, 12, 2197. [Google Scholar] [CrossRef]
- Hamaguchi, R.; Uemoto, S.; Wada, H. Editorial: The Impact of Alkalizing the Acidic Tumor Microenvironment to Improve Efficacy of Cancer Treatment, Volume II. Front. Oncol. 2024, 14, 1542787. [Google Scholar] [CrossRef]
Figure 1.
Acidic tumor microenvironment as a self-sustaining vicious cycle and an SBM-informed attractor-state model for long-tail survival. (A) Panel A illustrates a vicious cycle in which enhanced glycolysis (Warburg effect) with lactate/proton export, together with acid retention driven by abnormal vasculature and hypoxia, promotes formation and maintenance of an acidic TME (pHe ~6.5–6.9). An acidic TME simultaneously facilitates invasion/metastasis, drug resistance, and immunosuppression via impaired T-cell and NK-cell effector functions, thereby contributing to early poor outcomes in advanced cancer. (B) Panel B is a conceptual schematic that views the tumor–host system as a non-equilibrium open system and hypothesizes that an acidic TME can constitute a self-sustaining malignant regime (an attractor), while interventions and/or changes in host conditions may enable transitions toward an alternative stable state (an alternative attractor) characterized by lower acidity and more permissive immune function. Arrows indicate proposed relationships and may include non-causal links; they should not be interpreted as established causality. “Perturbations/levers” denote examples such as standard therapy, immunotherapy, buffering/dietary interventions, changes in perfusion, host factors (e.g., nutrition, inflammation, renal function), and treatment timing or adaptive modifications [19].
Figure 1.
Acidic tumor microenvironment as a self-sustaining vicious cycle and an SBM-informed attractor-state model for long-tail survival. (A) Panel A illustrates a vicious cycle in which enhanced glycolysis (Warburg effect) with lactate/proton export, together with acid retention driven by abnormal vasculature and hypoxia, promotes formation and maintenance of an acidic TME (pHe ~6.5–6.9). An acidic TME simultaneously facilitates invasion/metastasis, drug resistance, and immunosuppression via impaired T-cell and NK-cell effector functions, thereby contributing to early poor outcomes in advanced cancer. (B) Panel B is a conceptual schematic that views the tumor–host system as a non-equilibrium open system and hypothesizes that an acidic TME can constitute a self-sustaining malignant regime (an attractor), while interventions and/or changes in host conditions may enable transitions toward an alternative stable state (an alternative attractor) characterized by lower acidity and more permissive immune function. Arrows indicate proposed relationships and may include non-causal links; they should not be interpreted as established causality. “Perturbations/levers” denote examples such as standard therapy, immunotherapy, buffering/dietary interventions, changes in perfusion, host factors (e.g., nutrition, inflammation, renal function), and treatment timing or adaptive modifications [19].
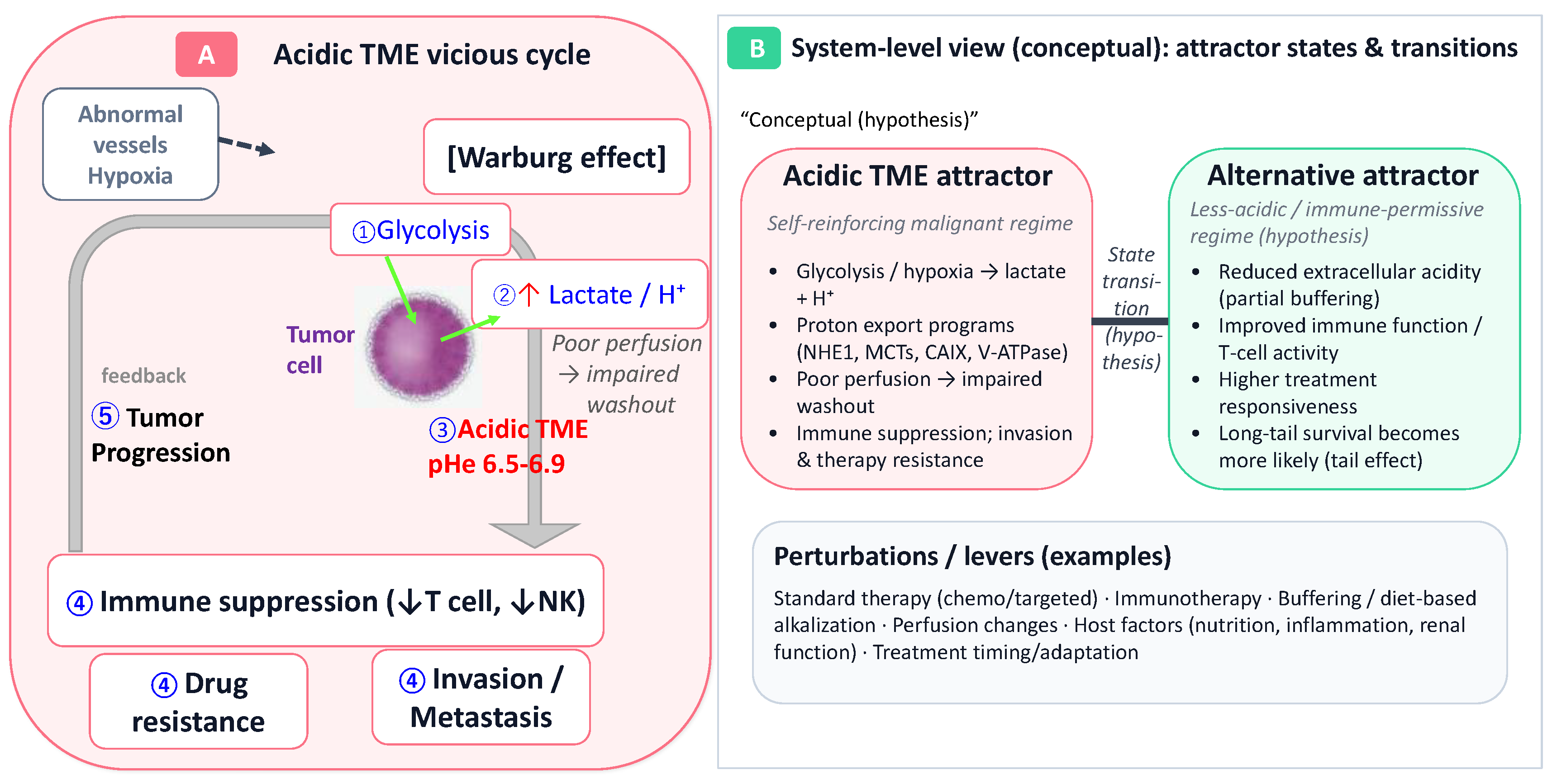
Figure 2.
Kaplan–Meier survival curves stratified by urinary pH in stage IV pancreatic cancer. Patients were stratified into three groups by urinary pH (Group 1: pH ≥ 7.5, n = 24; Group 2: 5.5 < pH < 7.5, n = 93; Group 3: pH ≤ 5.5, n = 10). Differences among groups were significant by the log-rank test (χ² = 22.6, df = 2, p < 0.0001) and also by the Wilcoxon test (χ² = 17.5, df = 2, p = 0.0002). Median survival times were 426.0 days (Group 1), 249.0 days (Group 2), and 101.5 days (Group 3). Five-year survival rates were 9.6% (Group 1), 8.1% (Group 2), and 0% (Group 3). This figure was prepared and modified based on Kachi et al. [31]; please refer to the original article for cohort definitions and the method used to summarize urinary pH.
Figure 2.
Kaplan–Meier survival curves stratified by urinary pH in stage IV pancreatic cancer. Patients were stratified into three groups by urinary pH (Group 1: pH ≥ 7.5, n = 24; Group 2: 5.5 < pH < 7.5, n = 93; Group 3: pH ≤ 5.5, n = 10). Differences among groups were significant by the log-rank test (χ² = 22.6, df = 2, p < 0.0001) and also by the Wilcoxon test (χ² = 17.5, df = 2, p = 0.0002). Median survival times were 426.0 days (Group 1), 249.0 days (Group 2), and 101.5 days (Group 3). Five-year survival rates were 9.6% (Group 1), 8.1% (Group 2), and 0% (Group 3). This figure was prepared and modified based on Kachi et al. [31]; please refer to the original article for cohort definitions and the method used to summarize urinary pH.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



