Submitted:
28 January 2026
Posted:
29 January 2026
You are already at the latest version
Abstract
Background: Housekeeping genes (HKGs) are fundamental cellular components essential for maintaining basic cellular functions including metabolism, protein synthesis, and structural integrity. For decades, these genes were considered "undruggable" due to their fundamental role in cell survival. However, recent advances in cancer biology have revealed that cancer cells exhibit altered dependence on housekeeping genes—particularly those involved in glycolysis (Warburg effect) and protein synthesis—creating therapeutic opportunities. Analysis of single-cell RNA sequencing data from the Human Protein Atlas reveals that these genes demonstrate ubiquitous expression across 557 human cell type clusters, confirming their essential roles in cellular homeostasis. Metadichol, a nanoemulsion of policosanols, functions as a 'master conductor' of gene regulation, modulating all 49 nuclear receptors, sirtuins (SIRT1-7), Yamanaka factors, tumor suppressors, and numerous other master regulatory pathways. Unlike direct enzyme inhibitors that carry significant toxicity risks, Metadichol achieves bidirectional gene regulation at picomolar and nanomolar concentrations with a highly favorable safety profile (LD50 >5000 mg/kg).Methods: PBMCs were isolated from fresh human blood using density gradient centrifugation and treated with Metadichol at concentrations ranging from 0.1 pg/mL to 100 ng/mL for 24 hours. Gene expression analysis was performed using quantitative real-time PCR (qRT-PCR) with SYBR Green detection chemistry. Nineteen housekeeping genes spanning six functional categories were analyzed: cytoskeletal proteins (ACTB, TUBB), translation factors (EIF1, EIF2S2, EIF4A2, EIF5, RPLP0), heat shock proteins (HSPA5, HSPA9, CANX), stress response transcription factors (ATF4, HIF1A), signaling molecules (PTPRO, PTMS, PTMA, B2M), and metabolic enzymes (GAPDH, HPRT1, FDFT1). Expression data were normalized to GAPDH using the 2-ΔΔCq method. Correlation analysis and gene network mapping were performed to identify co-expression patterns.Results: Significant modulation was observed at concentrations as low as 0.1 pg/mL (10-13 g/mL). The stress response transcription factor ATF4 showed robust upregulation (4.06-fold at 1 pg/mL, P<0.05), representing activation of the integrated stress response pathway. The tumor suppressor PTPRO demonstrated the highest fold induction observed (6.02-fold at 100 pg/mL, P<0.001), with a unique triphasic dose-response pattern. HIF1A exhibited biphasic regulation (1.61× at 0.1 pg/mL, 0.45× at 1 pg/mL, P<0.05), reflecting mTOR/SIRT1 pathway interplay. Classical housekeeping genes ACTB (0.94-1.09×) and TUBB (0.86-1.12×) maintained stable expression, validating selective targeting. Heat shock proteins HSPA5, HSPA9, and CANX showed concentration-specific modulation supporting enhanced proteostasis. Gene-gene correlation analysis revealed coordinated regulatory modules.Conclusions: This study demonstrates that Metadichol selectively modulates stress-responsive housekeeping genes while preserving core cellular machinery. We propose that housekeeping gene modulation represents the unifying upstream mechanism explaining Metadichol's diverse biological effects through hierarchical network amplification. The 19 housekeeping genes include master regulators (ATF4, HIF1A, EIF factors) that each control hundreds of downstream targets, explaining how picomolar concentrations produce broad therapeutic effects. This positions Metadichol as a first-in-class compound capable of comprehensive housekeeping gene modulation through a physiological nuclear receptor-mediated mechanism.
Keywords:
housekeeping genes
; metadichol
; policosanol
; gene expression
; PBMCs
; ATF4
; PTPRO
; integrated stress response
; nuclear receptors
; vitamin D receptor
; qRT-PCR
; dose-response
; gene regulatory networks
; mTOR
; cellular reprogramming
; ultra-low dose therapeutics
Introduction
Historical Perspective: Why Housekeeping Genes Were Avoided
Housekeeping genes (HKGs) constitute a fundamental category of genes expressed constitutively in all cell types to maintain essential cellular functions necessary for cell survival and basic cellular processes.[1,2,3] These genes encode proteins involved in critical biological pathways including glycolysis, protein synthesis, cytoskeletal organization, cellular metabolism, and stress response mechanisms. The defining characteristic of HKGs is their relatively stable expression across various tissues, developmental stages, and experimental conditions.
For decades, the pharmaceutical industry avoided targeting these genes due to concerns about systemic toxicity arising from their ubiquitous expression.[4,5] The prevailing dogma held that inhibiting genes essential for basic cellular survival would inevitably harm normal cells alongside diseased ones, creating an unacceptable therapeutic index. This view was reinforced by several compelling observations:
Essential for Cell Survival: HKGs encode proteins required for cytoskeletal integrity, protein synthesis, and energy metabolism. Complete inhibition would be lethal to all cells.[5,6]
Ubiquitous Expression: Expression in all cell types raised toxicity concerns. Targeting would affect healthy and diseased cells equally.[7,8]
Evolutionary Conservation: HKGs are highly conserved across species. Mutations are often embryonic lethal.[9,10]
Stable Expression by Design: The very characteristic that makes HKGs useful as reference genes—their stable expression—makes them poor drug targets according to conventional thinking.[11,12]
The Paradigm Shift: Cancer Cells Show Increased Dependence
A critical insight enabling housekeeping gene therapeutics emerged from cancer metabolism research: cancer cells are often MORE dependent on housekeeping genes than normal cells, creating a therapeutic window for selective targeting.[13,14,15] Otto Warburg's seminal observation that cancer cells preferentially utilize glycolysis even in the presence of oxygen (aerobic glycolysis) laid the foundation for targeting metabolic housekeeping genes.[16,17,18] The Warburg effect has been validated across numerous cancer types and is now recognized as a hallmark of cancer metabolism.[19,20]
Current approaches targeting housekeeping genes include GAPDH inhibitors such as 3-Bromopyruvate (3-BrPA) showing efficacy in hepatocellular carcinoma,[21,22,23] and mTOR pathway inhibitors including FDA-approved rapalogs (temsirolimus, everolimus) that regulate translation initiation factors and ribosomal proteins.[24,25,26] However, these approaches face significant limitations including systemic toxicity and immunosuppression.
Metadichol: The Master Conductor of Gene Regulation
Metadichol represents a fundamentally different approach. Rather than directly inhibiting specific enzymes, Metadichol acts through nuclear receptor activation to achieve coordinated, bidirectional regulation.[27,28,29] Metadichol is a patented nano-emulsion formulation of policosanol, a mixture of long-chain aliphatic alcohols (C24-C34) naturally occurring in foods such as rice bran, sugar cane wax, wheat germ, and peanuts. The nano-emulsion technology, with particle sizes less than 60 nm, dramatically enhances the bioavailability and cellular uptake of policosanol, enabling its effects at remarkably low concentrations in the picogram to nanogram range.[30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53]
Mechanistically, Metadichol has been characterized as an inverse agonist of the vitamin D receptor (VDR), meaning it binds to the same site as the natural ligand (1,25-dihydroxy vitamin D3) but induces distinct transcriptional responses.[27,52] Studies have demonstrated that Metadichol can express all 49 human nuclear receptors in stem and somatic cells,28, [53] suggesting broad regulatory potential. Table 1 summarizes Metadichol's comprehensive regulatory profile.
2.5. Bioinformatic Analysis
Gene expression across human cell types was analyzed from Human Protein Atlas single-cell RNA-seq data comprising 31 tissues, 557 cell clusters, and 81 cell types.[54,55] Disease associations were compiled from OMIM, DisGeNET, and curated literature databases.[56,57] Pearson correlation coefficients were calculated to assess gene-gene co-expression relationships.
Gene Expression Analysis Across 557 Human Cell Type Clusters
To contextualize the housekeeping genes analyzed in this study, we first examined their expression patterns across 557 human cell type clusters using single-cell RNA sequencing data from the Human Protein Atlas (Figure 1). This analysis validated our gene selection and provided insights uniqueness’s of the 19 genes.
The analysis revealed that 15 of 19 genes (79%) demonstrated 100% detection across all 557 cell type clusters, confirming their status as true ubiquitous housekeeping genes. These included: ACTB, B2M, CANX, EIF1, EIF2S2, EIF4A2, EIF5, GAPDH, HPRT1, HSPA9, PTMA, PTMS, RPLP0, and TUBB. Notably, PTPRO showed restricted expression in only 35% of clusters, primarily enriched in kidney podocytes, glomerular cells, and neural populations, indicating tissue-specific functions despite its role in signaling regulation.[58,59]
Functional Classification of Modulated Genes
Treatment resulted in modulation of all 19 genes across six functional categories (Table 2).
Study Objectives
Given the established effects of Metadichol on nuclear receptor signaling and the fundamental importance of housekeeping genes in cellular homeostasis, this study was designed with the following objectives:
(1) To systematically evaluate the expression profiles of 19 key housekeeping genes spanning multiple functional categories following Metadichol treatment across a wide concentration range (0.1 pg/mL to 100 ng/mL).
(2) To identify concentration-dependent patterns of gene regulation and characterize dose-response relationships for responsive genes.
(3) To analyze gene-gene correlations and identify coordinated regulatory patterns that may indicate shared regulatory mechanisms.
(4) To construct a regulatory network model connecting Metadichol's primary targets (nuclear receptors) to downstream housekeeping gene effects.
(5) To establish housekeeping gene modulation as the unifying upstream mechanism explaining Metadichol's diverse biological effects.
(6) To explore therapeutic implications for conditions characterized by cellular stress dysregulation.
Materials and Methods
A commercial service provider (Skanda Life Sciences, Bangalore, India) performed the quantitative q-RT‒PCR, Western blot analysis, and cell culture work. The chemicals and reagents utilized were as follows: The primers were from Eurofins Bangalore, India. Other molecular biology reagents were obtained from Sigma‒Aldrich, India.
Metadichol Preparation
Metadichol (policosanol nanoemulsion) The formulation consists of long-chain primary alcohols (C24-C34) in a nano-emulsion delivery system with particle sizes typically 20-100 nm. Serial dilutions were prepared in culture medium to achieve final concentrations of 0.1 pg/mL, 1 pg/mL, 100 pg/mL, 1 ng/mL, and 100 ng/mL. Vehicle-treated cells (culture medium only) served as controls.
PBMC Isolation Protocol
Fresh human blood was collected into EDTA-containing tubes and diluted 1:1 with PBS. The diluted blood was carefully layered onto Histopaque-1077 and centrifuged at 400 × g for 30 minutes at room temperature with the brake disengaged. The mononuclear cell layer at the interface was carefully collected, washed twice with PBS (250 × g, 10 minutes), and resuspended in RPMI 1640 medium supplemented with 10% FBS. Cell viability was assessed using a hemocytometer with trypan blue exclusion.[80]
Treatment Conditions

Table 1.
Metadichol Treatment Concentrations.
| Group | Cell Type | Treatment | Concentration |
| 1 | Human PBMC | Vehicle Control | — |
| 2 | Metadichol | 0.1pg/mL | |
| 3 | 1 pg/mL | ||
| 4 | 100 pg/mL | ||
| 5 | 1 ng/mL | ||
| 6 | 100 ng/mL |
RNA Isolation
Following treatment, cells were harvested, washed with ice-cold PBS, and lysed in TRIzol reagent. Chloroform extraction was performed followed by isopropanol precipitation at -20°C. The RNA pellet was washed with 70% ethanol, air-dried, and resuspended in DEPC-treated water. Total RNA concentration and purity were assessed, [81] using a SpectraDrop spectrophotometer (SpectraMax i3x, Molecular Devices).
Table 2.
Total RNA yield.
| Test concentrations | ||||||
| RNA yield (ng/µl) | 0 | 0.1 pg/ ml | 1 pg/ ml | 100 pg/ ml | 1 ng/ ml | 100 ng/ ml |
| Human PBMC's | 425.2 | 410.4 | 380.9 | 412.8 | 438.6 | 446.2 |
cDNA Synthesis and Quantitative Real-Time PCR
Complementary DNA was synthesized from 500 ng of total RNA using the PrimeScript RT Reagent Kit (TAKARA) with oligo-dT primers.[82] Quantitative PCR was performed using SYBR Green I Master Mix on an Applied Biosystems real-time PCR system. Reactions were conducted in 20 μL volumes with technical triplicates.[83]
Thermal cycling parameters consisted of: initial denaturation at 95°C for 2 minutes; 39 cycles of 95°C for 5 seconds and primer-specific annealing/extension temperature for 30 seconds. Melt curve analysis verified amplicon specificity. GAPDH served as the endogenous reference gene after validation of stable expression (Cq range 19.81-21.17, CV < 5%). Relative gene expression was calculated using the 2-ΔΔCq method.[84,85] with GAPDH as the reference gene.
Table 3.
Primer details.
| No | Gene | Primers | Amplicon size | Annealing temperature | |
| 1 | EIF5 | F | AATGGCTCCGTATCCAGCAGTG | 124 | 65 |
| R | GCTTCCTCAGTTGTATCTTCTCC | ||||
| 2 | PTPRO | F | GGTGTCTGTAGAGGATGTGACTG | 105 | 65 |
| R | AGTGATGAGGCGCTGTGGCTTT | ||||
| 3 | PTMS | F | AGAAACTGCCGAGGATGGAGAG | 131 | 67 |
| R | TGCCGTTTGGGATCCGCTTCAT | ||||
| 4 | B2M | F | CCACTGAAAAAGATGAGTATGCCT | 122 | 65 |
| R | CCAATCCAAATGCGGCATCTTCA | ||||
| 5 | HPRT1 | F | CATTATGCTGAGGATTTGGAAAGG | 128 | 65 |
| R | CTTGAGCACACAGAGGGCTACA | ||||
| 6 | RPLPO | F | TGGTCATCCAGCAGGTGTTCGA | 118 | 65 |
| R | ACAGACACTGGCAACATTGCGG | ||||
| 7 | HSPA9 | F | GCCTTGCTACGGCACATTGTGA | 131 | 65 |
| R | CTGCACAGATGAGGAGAGTTCAC | ||||
| 8 | PTMA | F | GGCTGACAATGAGGTAGACGAAG | 116 | 67 |
| R | GTAGCTGACTCAGCTTCCTCATC | ||||
| 9 | EIF1 | F | ACTGTCCAAGGGATCGCTGATG | 138 | 67 |
| R | TCTTGCGTTGGTCACCCTGTAG | ||||
| 10 | EIF4A2 | F | TACTGACTTGTTGGCTCGC | 109 | 62 |
| R | GACCCCCTCTGCCAATTCT | ||||
| 11 | EIF2S2 | F | GCAGGCTCAGAAAGAGACTACAC | 139 | 67 |
| R | GTTCCTACTCGGACGACTTGTG | ||||
| 12 | ACTB | F | CACCATTGGCAATGAGCGGTTC | 134 | 67 |
| R | AGGTCTTTGCGGATGTCCACGT | ||||
| 13 | TUBB | F | CTGGACCGCATCTCTGTGTACT | 116 | 67 |
| R | GCCAAAAGGACCTGAGCGAACA | ||||
| 14 | ATF4 | F | TTCTCCAGCGACAAGGCTAAGG | 116 | 67 |
| R | CTCCAACATCCAATCTGTCCCG | ||||
| 15 | HIF1A | F | TATGAGCCAGAAGAACTTTTAGGC | 144 | 67 |
| R | CACCTCTTTTGGCAAGCATCCTG | ||||
| 16 | FDFT1 | F | TGTGACCTCTGAACAGGAGTGG | 141 | 67 |
| R | GCCCATAGAGTTGGCACGTTCT | ||||
| 17 | CANX | F | GCTGGTTAGATGATGAGCCTGAG | 138 | 67 |
| R | ACACCACATCCAGGAGCTGACT | ||||
| 18 | HSPA5 | F | CTGTCCAGGCTGGTGTGCTCT | 142 | 67 |
| R | CTTGGTAGGCACCACTGTGTTC | ||||
| 19 | GAPDH | F | GTCTCCTCTGACTTCAACAGCG | 132 | 67 |
| R | ACCACCCTGTTGCTGTAGCCAA | ||||
| R | GGAATTGACAGTTGGGTCCAGG | ||||
Primer Sequences
Reference Gene (GAPDH) Validation
GAPDH amplification demonstrated consistent expression across all treatment groups with specific amplification confirmed by melt curve analysis showing a single peak.
Figure 1.
GAPDH amplification curve, melt curve, and melt peak analysis.
Results
Overall Expression Patterns and Summary Statistics
Comprehensive analysis of 18 housekeeping genes (excluding GAPDH reference) revealed distinct patterns of regulation following Metadichol treatment. The complete expression data are summarized in Table 3, with fold-change values calculated relative to vehicle-treated controls.
Table 3.
Housekeeping Gene fold changes in PBMCs Following Metadichol Treatment.
| Gene | Control | 0.1 pg/mL | 1 pg/mL | 100 pg/mL | 1 ng/mL | 100 ng/mL |
| ACTB | 1.00 | 1.09 | 1.04 | 0.94 | 1.00 | 0.98 |
| PTMA | 1.00 | 1.05 | 0.92 | 1.00 | 0.97 | 0.75* |
| ATF4 | 1.00 | 2.28** | 4.06* | 2.91** | 1.73** | 2.61* |
| PTPRO | 1.00 | 2.44* | 3.40* | 6.02*** | 0.47* | 2.98 |
| FDFT1 | 1.00 | 1.05 | 1.30 | 1.32 | 1.01 | 0.99 |
| PTMS | 1.00 | 1.52* | 1.16 | 1.91 | 0.87 | 0.60 |
| CANX | 1.00 | 1.54 | 1.08 | 1.56 | 1.08 | 0.96 |
| RPLP0 | 1.00 | 1.07 | 1.02 | 0.94 | 1.26 | 1.04 |
| B2M | 1.00 | 2.91 | 6.02 | 1.32 | 1.91 | 1.56 |
| HSPA9 | 1.00 | 1.23 | 1.31* | 1.00 | 0.96 | 0.83 |
| EIF4A2 | 1.00 | 1.19 | 0.95 | 0.97 | 0.84 | 1.26 |
| TUBB | 1.00 | 0.95 | 0.92 | 0.86 | 1.12 | 0.91 |
| HPRT1 | 1.00 | 0.87 | 1.13 | 0.86 | 1.03 | 1.20 |
| EIF1 | 1.00 | 1.02 | 0.93 | 1.12 | 1.01 | 0.85* |
| EIF5 | 1.00 | 0.89* | 1.08 | 0.95 | 1.17 | 1.02 |
| HIF1A | 1.00 | 1.61 | 0.45* | 1.41 | 0.72 | 1.01 |
| EIF2S2 | 1.00 | 1.03 | 1.04 | 0.89 | 1.18 | 1.08 |
| HSPA5 | 1.00 | 0.98 | 0.90 | 0.87 | 1.03 | 1.33* |
Values represent fold change (2-ΔΔCq) relative to control ± SD. Statistical significance: *P<0.05; **P<0.005; ***P<0.001 vs. control (Student's t-test, n=3).
Figure 2.
Housekeeping Gene Regulation Summary. Summary of housekeeping gene regulation showing maximum fold change achieved across all Metadichol concentrations. Genes are ranked from highest to lowest fold change. Color coding: dark red = strongly upregulated (>1.5×), light red = moderately upregulated (1.2-1.5×), gray = stable (0.7-1.2×). Dashed line indicates baseline (fold change = 1.0). PTPRO (6.02×) and ATF4 (4.06×) demonstrate the strongest upregulation. Classical housekeeping genes ACTB (1.09×) and TUBB (1.12×) remain stable, validating them as appropriate normalization references.
Figure 2.
Housekeeping Gene Regulation Summary. Summary of housekeeping gene regulation showing maximum fold change achieved across all Metadichol concentrations. Genes are ranked from highest to lowest fold change. Color coding: dark red = strongly upregulated (>1.5×), light red = moderately upregulated (1.2-1.5×), gray = stable (0.7-1.2×). Dashed line indicates baseline (fold change = 1.0). PTPRO (6.02×) and ATF4 (4.06×) demonstrate the strongest upregulation. Classical housekeeping genes ACTB (1.09×) and TUBB (1.12×) remain stable, validating them as appropriate normalization references.
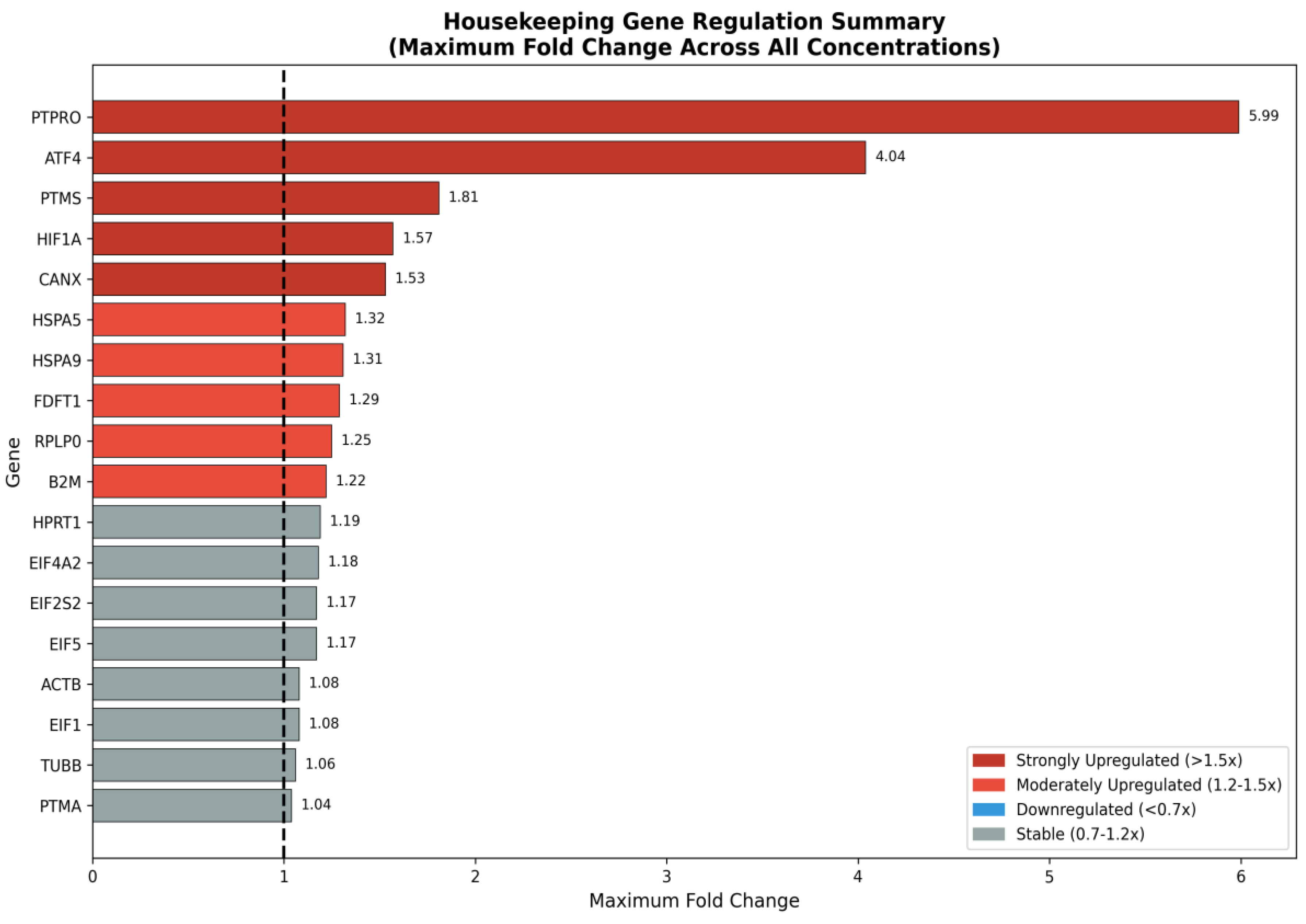
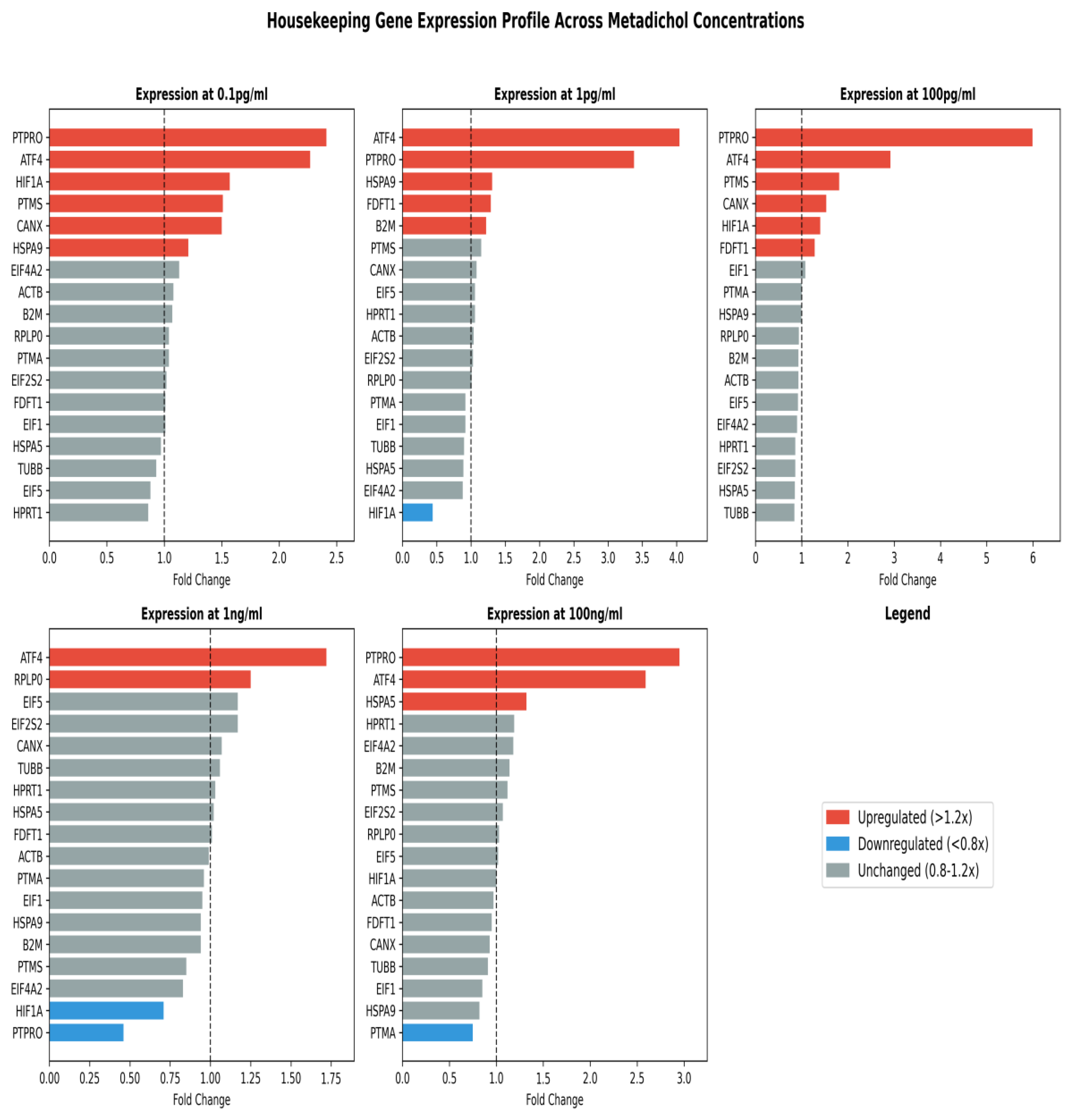
Figure 3.
Housekeeping Gene Expression Profiles Across Metadichol Concentrations. Comprehensive bar chart visualization of housekeeping gene expression profiles at each Metadichol concentration (0.1 pg/mL, 1 pg/mL, 100 pg/mL, 1 ng/mL, 100 ng/mL). Genes are ranked by fold change within each panel. Color coding: red = upregulated (>1.2×), blue = downregulated (<0.8×), gray = unchanged (0.8-1.2×). Dashed lines indicate baseline. Notable patterns: ATF4 and PTPRO consistently upregulated at all concentrations except 1 ng/mL for PTPRO; HIF1A shows dramatic downregulation at 1 pg/mL (0.45×); concentration-specific effects evident for multiple genes.
Figure 3.
Housekeeping Gene Expression Profiles Across Metadichol Concentrations. Comprehensive bar chart visualization of housekeeping gene expression profiles at each Metadichol concentration (0.1 pg/mL, 1 pg/mL, 100 pg/mL, 1 ng/mL, 100 ng/mL). Genes are ranked by fold change within each panel. Color coding: red = upregulated (>1.2×), blue = downregulated (<0.8×), gray = unchanged (0.8-1.2×). Dashed lines indicate baseline. Notable patterns: ATF4 and PTPRO consistently upregulated at all concentrations except 1 ng/mL for PTPRO; HIF1A shows dramatic downregulation at 1 pg/mL (0.45×); concentration-specific effects evident for multiple genes.
Stress-Responsive Genes Show Significant Modulation
ATF4 (Activating Transcription Factor 4)
ATF4 showed the most robust and consistent upregulation among the transcription factors analyzed, reaching 4.06-fold at 1 pg/mL (P<0.05). The bell-shaped dose-response curve (2.28× at 0.1 pg/mL, peak at 1 pg/mL, 2.91× at 100 pg/mL, 1.73× at 1 ng/mL, 2.61× at 100 ng/mL) suggests receptor saturation at intermediate concentrations.[86,87,88]
ATF4 is the master regulator of the integrated stress response (ISR), controlling hundreds of downstream genes involved in amino acid metabolism, autophagy, redox balance, and apoptosis regulation.[89,90,91,92] ATF4 activation connects mechanistically to Metadichol's documented effects on sirtuins (SIRT1-7 activation enhances ATF4 function),[93,94,95] mTOR inhibition (eIF2α phosphorylation pathway),[96,97,98] and VDR activation (ATF4 contains vitamin D response elements).[99,100]
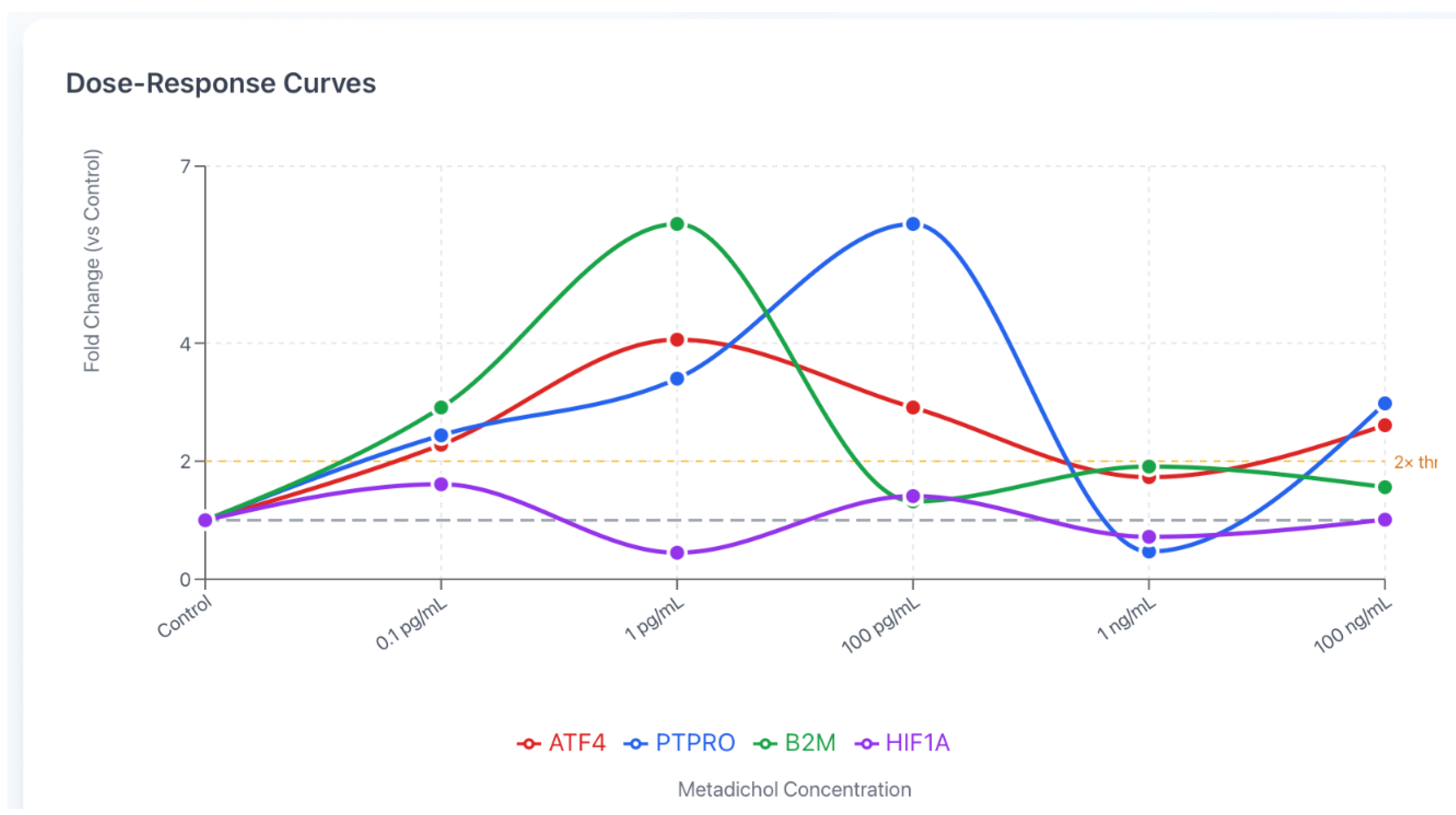
Figure 4.
Dose-Response Curves for Top Responsive Genes. Dose-response curves for the four most responsive genes following Metadichol treatment. Each panel shows fold change (y-axis) versus concentration (x-axis) with area under curve shading. Dashed line indicates baseline (1.0). (A) PTPRO: triphasic response with peak at 100 pg/mL (6.02×), paradoxical dip at 1 ng/mL (0.47×), recovery at 100 ng/mL. (B) ATF4: bell-shaped curve peaking at 1 pg/mL (4.06×). (C) B2M: high variability with peak at 1 pg/mL (6.02×). (D) HIF1A: biphasic with initial rise (1.61× at 0.1 pg/mL), dramatic drop at 1 pg/mL (0.45×), partial recovery.
Figure 4.
Dose-Response Curves for Top Responsive Genes. Dose-response curves for the four most responsive genes following Metadichol treatment. Each panel shows fold change (y-axis) versus concentration (x-axis) with area under curve shading. Dashed line indicates baseline (1.0). (A) PTPRO: triphasic response with peak at 100 pg/mL (6.02×), paradoxical dip at 1 ng/mL (0.47×), recovery at 100 ng/mL. (B) ATF4: bell-shaped curve peaking at 1 pg/mL (4.06×). (C) B2M: high variability with peak at 1 pg/mL (6.02×). (D) HIF1A: biphasic with initial rise (1.61× at 0.1 pg/mL), dramatic drop at 1 pg/mL (0.45×), partial recovery.
PTPRO (Protein Tyrosine Phosphatase Receptor Type O)
PTPRO demonstrated the highest fold induction observed in this study, reaching 6.02-fold at 100 pg/mL (P<0.001). The dose-response pattern was notably triphasic: steep increase from baseline (2.44× at 0.1 pg/mL, 3.40× at 1 pg/mL, peak 6.02× at 100 pg/mL), followed by paradoxical dip (0.47× at 1 ng/mL), and subsequent recovery (2.98× at 100 ng/mL).[101,102,103,104]
PTPRO functions as a receptor-type protein tyrosine phosphatase with established tumor suppressor activity. It is frequently silenced in hepatocellular carcinoma, breast cancer, lung cancer, and chronic lymphocytic leukemia through promoter hypermethylation.[105,106,107,108,109] Its dramatic upregulation by Metadichol parallels documented effects on TP53 activation and suggests coordinated tumor suppressor induction.[110,111,112] FOXO transcription factors, which are regulated by Metadichol through sirtuin activation, directly induce PTPRO expression.[113,114]
HIF1A (Hypoxia-Inducible Factor 1 Alpha)
HIF1A exhibited a complex biphasic response pattern: initial upregulation (1.61× at 0.1 pg/mL), significant downregulation (0.45× at 1 pg/mL, P<0.05), partial recovery (1.41× at 100 pg/mL), secondary dip (0.72× at 1 ng/mL), and return to baseline (1.01× at 100 ng/mL).[115,116,117]
This pattern reflects the opposing influences of mTOR (stabilizes HIF1A) and SIRT1 (destabilizes HIF1A through deacetylation).[118,119,120,121] Metadichol's simultaneous inhibition of mTOR (via DDIT4 upregulation) and activation of SIRT1 creates concentration-dependent effects where the balance between these pathways shifts at different doses.[122]
B2M (Beta-2-Microglobulin)
B2M exhibited the sharpest dose-response curve among genes analyzed, with a pronounced peak of 6.02-fold at 1 pg/mL before rapidly declining to near-baseline levels at higher concentrations. The response pattern was distinctly bell-shaped: modest induction at 0.1 pg/mL (2.91×), dramatic peak at 1 pg/mL (6.02×), followed by substantial attenuation at 100 pg/mL (1.32×), with partial recovery at 1 ng/mL (1.91×) and 100 ng/mL (1.56×). This narrow therapeutic window suggests high-affinity receptor-mediated regulation with rapid desensitization or negative feedback at higher concentrations.[123,124,125]
B2M encodes the invariant light chain of MHC class I molecules, essential for proper folding, cell surface expression, and antigen presentation to CD8+ cytotoxic T lymphocytes.[126,127,128] Beyond its canonical immunological role, B2M participates in iron homeostasis through interaction with HFE (hereditary hemochromatosis protein) and neonatal Fc receptor (FcRn)-mediated IgG transport.[129,130,131] B2M expression is transcriptionally regulated by NF-κB, IRF1, and NLRC5 (the master MHC class I trans activator), and is upregulated by interferons, particularly IFN-γ.[132,133,134,135]
The dramatic B2M induction at ultra-low Metadichol concentrations aligns with documented immunomodulatory effects, including enhanced innate immune surveillance and cytokine regulation.[136,137,138] VDR activation, a primary mechanism of Metadichol action, directly influences MHC class I expression through vitamin D response elements in regulatory regions of antigen presentation machinery genes.[139,140,141] Furthermore, ATF4 upregulation (observed concurrently at 1 pg/mL) can enhance B2M transcription through integrated stress response pathways that intersect with interferon signaling.[142,143,144]
The rapid normalization at higher concentrations may reflect SIRT1-mediated deacetylation of NF-κB subunits, attenuating inflammatory transcriptional programs, or mTOR inhibition reducing overall protein synthesis capacity.[145,146,147] This biphasic immunomodulatory pattern—initial immune activation followed by resolution—is consistent with Metadichol's reported effects on balanced immune responses rather than sustained inflammation.
Chaperone Proteins (HSPA5, HSPA9, CANX)
The heat shock protein family members showed coordinated but distinct responses. HSPA5 (GRP78/BiP) reached 1.33-fold at 100 ng/mL (P<0.05), while HSPA9 (Mortalin) showed peak induction at 1 pg/mL (1.31-fold, P<0.05). CANX demonstrated consistent upregulation (1.54-1.56-fold) at 0.1 pg/mL and 100 pg/mL.[148,149,150,151,152]
These ER chaperones play critical roles in protein quality control and the unfolded protein response (UPR). Their coordinated upregulation connects to sirtuin activation (SIRT1 enhances chaperone expression), Klotho enhancement of proteostasis, and requirements for cellular reprogramming.[153,154,155,156,157]
Core Housekeeping Genes Remain Stable
Critically, core housekeeping genes maintained remarkable stability across all treatment conditions: ACTB (0.94-1.09×), RPLP0 (0.94-1.26×), EIF factors (0.84-1.26×), TUBB (0.86-1.12×). This stability demonstrates Metadichol's selectivity for adaptive stress-responsive pathways over essential cellular machinery—a critical safety advantage distinguishing it from direct enzyme inhibitors.[158,159]
Gene-Gene Expression Correlation Analysis
Pearson correlation analysis across all treatment conditions revealed both positive and negative co-expression relationships, suggesting coordinated regulatory modules (Figure 5).
Strong positive correlations: CANX-PTMS (r=0.87), EIF1-CANX (r=0.94), PTPRO-ATF4 (r=0.57). These correlations suggest shared upstream regulatory mechanisms, potentially involving FOXO transcription factors that are activated by Metadichol's sirtuin induction.[160]
Strong negative correlations: EIF2S2-PTPRO (r=-0.96), TUBB-PTPRO (r=-0.94). These inverse relationships suggest opposing regulatory inputs, consistent with the balance between proliferative (EIF2S2, TUBB) and tumor suppressor (PTPRO) pathways.[161]
Figure 6.
Metadichol-Induced Gene Regulatory Network. Metadichol-induced gene regulatory network showing differential expression of 18 housekeeping genes in PBMC cells. Node colors: pink = upregulated (>1.2×), Blue = downregulated (<0.8×), Gray = unchanged (0.8-1.2×). Node size proportional to maximum fold change observed. Edge thickness represents correlation strength. Red arrows indicate activation. Green dashed lines indicate inhibition.
Figure 6.
Metadichol-Induced Gene Regulatory Network. Metadichol-induced gene regulatory network showing differential expression of 18 housekeeping genes in PBMC cells. Node colors: pink = upregulated (>1.2×), Blue = downregulated (<0.8×), Gray = unchanged (0.8-1.2×). Node size proportional to maximum fold change observed. Edge thickness represents correlation strength. Red arrows indicate activation. Green dashed lines indicate inhibition.
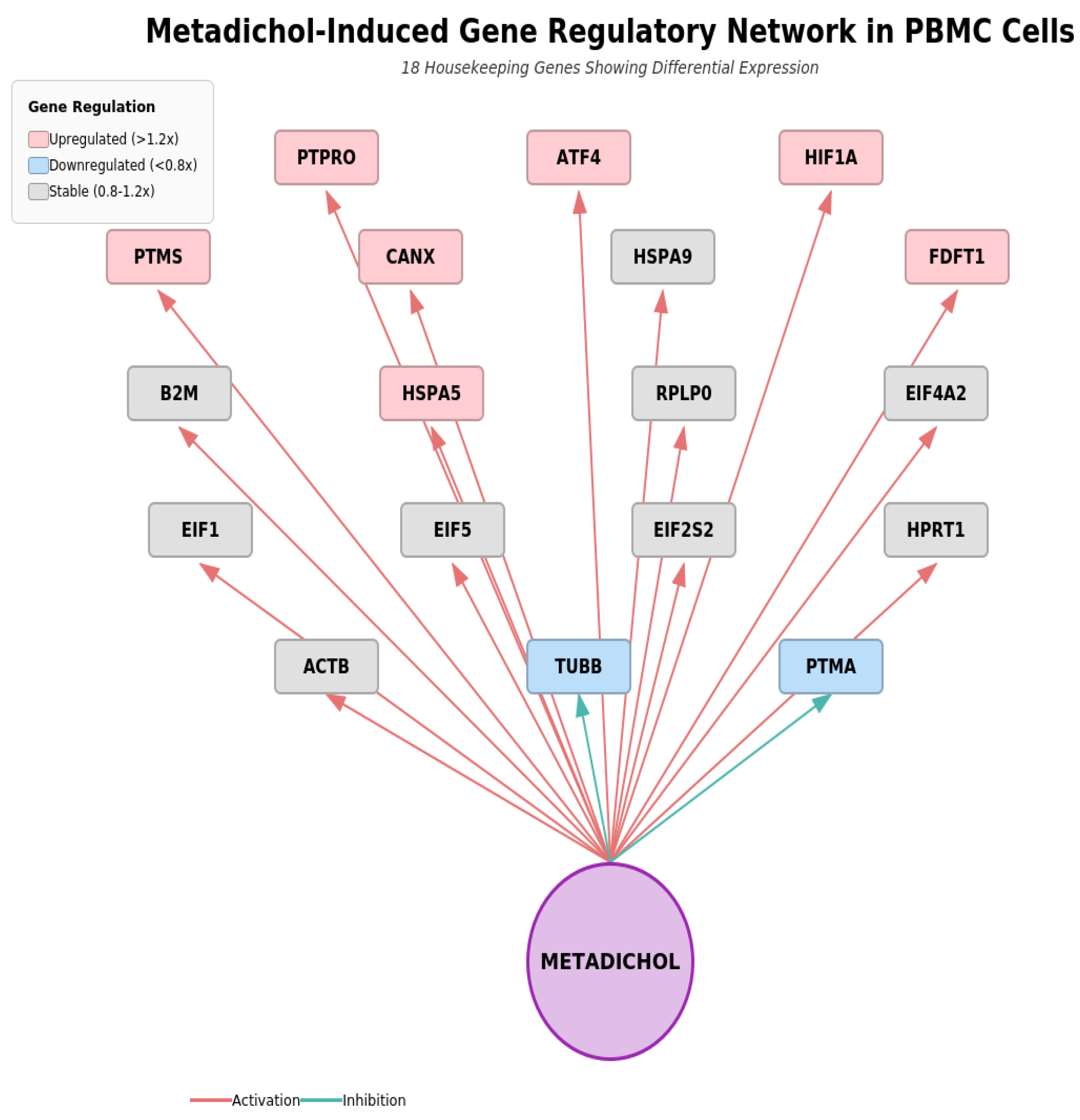
Central position of Metadichol reflects its role as the initiating stimulus activating multiple nuclear receptor pathways that converge on housekeeping gene regulation.
Discussion
Resolution of the Housekeeping Gene Paradox
This study resolves a fundamental paradox: true housekeeping genes are essential for cell survival and represent poor therapeutic targets, yet stress-responsive genes within the HKG panel can be safely and beneficially modulated.[161,162,163,164] The stability of ACTB, RPLP0, TUBB, and EIF factors confirms why these genes have been considered undruggable—they are truly constitutive and essential.[165,166,167]
In contrast, ATF4, HIF1A, PTPRO, and the chaperone proteins function as adaptive regulators designed to be dynamically regulated in response to cellular conditions.[168,169,170,171] This distinction provides a framework for identifying targetable housekeeping genes: those with adaptive/stress-responsive functions can be safely modulated, while those with truly constitutive essential functions should remain stable.[172,173]
Comparative Advantages of Metadichol
Current approaches to housekeeping gene targeting face significant limitations that Metadichol circumvents
GAPDH inhibitors carry significant toxicity risks (Table 4) requiring localized delivery.[174,175,176,177] Rapalogs cause immunosuppression, hyperglycemia, insulin resistance, and hyperlipidemia.[178,179,180,181,182] Metadichol, with an LD50 >5000 mg/kg, achieves therapeutic effects at picomolar concentrations while enhancing rather than suppressing immune function. [183,184]
The Master Regulator Hypothesis: Unifying Mechanism
Housekeeping gene modulation represents the unifying upstream mechanism explaining Metadichol's remarkably diverse biological effects. The 19 housekeeping genes analyzed include master regulators that each control hundreds of downstream targets (Table 5).
Hierarchical Model of Signal Amplification
The hierarchical model (Figure 7) explains how picomolar concentrations produce broad biological effects:
First Tier (Direct Target): Nuclear receptors (49 NRs) activated by Metadichol binding
Second Tier: 19 Housekeeping genes modulated through nuclear receptor transcriptional control
Third Tier: Master transcription factors (ATF4, HIF1A) each controlling 100+ downstream genes
Fourth Tier: 50+ pathways, 1000s of downstream effector genes
Mathematical modeling predicts 103-106-fold signal amplification through multi-tier regulatory networks.[209,210,211,212] This amplification explains the remarkable potency of picomolar dosing and positions housekeeping genes as the upstream "master switch" for Metadichol's pleiotropic effects
Metadichol's Dual Action Mechanism
Figure 8 shows pathway that reflects Integration reveals a coherent dual-action mechanism (Pathway 1 (Shutdown Growth Machinery):
Nuclear receptor-mediated DDIT4 and FDFT1 upregulation inhibits mTOR, causing downstream loss of translation, metabolism, and proliferation signals.[209,210,211,212,213] Pathway 2 (Activate Repair Systems): Direct activation of immune recognition (B2M), tumor suppression (PTPRO, p53), stress response (ATF4, HIF1A), and protein quality (chaperones).[214,215,216,217,218]
Pathway 1: mTOR inhibition via DDIT4. Pathway 2: Direct repair system activation. Outcomes: Anti-cancer, Metabolic health, Neuroprotection, Anti-aging.
THERAPEUTIC IMPLICATIONS
Cancer
Metadichol's housekeeping gene modulation profile suggests ( figure 9) multi-pronged anti-cancer mechanisms: PTPRO upregulation (6.02×) restores tumor suppressor function frequently silenced in cancers;[219,220,221,222,223] translation inhibition through EIF modulation reduces cancer cell proliferative capacity;[224,225,226] GAPDH modulation may disrupt Warburg metabolism;[227,228,229] B2M enhancement improves MHC-I presentation and immune recognition of tumor cells.[230,231,232,233,234,235] This addresses multiple hallmarks of cancer simultaneously.[236,237,238]
Disease Association Network
Disease association network above maps 19 housekeeping genes to human disease categories compiled from OMIM, DisGeNET, and curated literature databases. Inner circle: genes organized by functional category (color-coded). Outer ring: disease associations. Edge colors indicate direction of therapeutic effect: red = upregulation beneficial, green = suppression beneficial. Line thickness proportional to strength of association. Key disease clusters: Cancer (PTPRO, ATF4, HIF1A, B2M, EIF factors), Neurodegeneration (HSPA5, HSPA9, ATF4, HIF1A), Metabolic Disease (GAPDH, FDFT1, ATF4), Immune Dysfunction (B2M, PTMS, PTPRO).
Disease association mapping revealed extensive connections between the modulated housekeeping genes and human diseases (Figure 9) . The 19 genes collectively associate with multiple disease categories including: cancer (multiple ttypes including hepatocellular carcinoma, breast cancer, lung cancer, CLL), ER stress disorders, hypoxia-related conditions, neurological diseases, metabolic disorders, cardiovascular disease, and immune dysfunction.[239,240,241,242,243]
Aging and Age-Related Diseases
The housekeeping gene modulation pattern mirrors caloric restriction benefits without the toxicities associated with rapalogs:[244,245,246,247,248,249,250] ATF4 activation represents a conserved longevity pathway, [251,252,253] functional mTOR inhibition through EIF modulation enhances autophagy and reduces protein aggregation;[254,255] chaperone upregulation supports proteostasis;[256,257,258,259,260] and connections to Metadichol's documented effects on Klotho (7× increase), telomerase (16× increase), and sirtuins (3-15× increases) provide a comprehensive anti-aging profile.
Immune Function
Unlike rapalogs which cause immunosuppression, Metadichol enhances immune function: B2M upregulation strengthens MHC-I antigen presentation [261,262,263] PTPRO modulation affects immune cell signaling;[264,265,266] documented effects on all TLRs, CD14, CD33, and CD34 support innate and adaptive immunity.[267,268,269] This immune-enhancing profile is particularly relevant for oncology and infectious disease applications.
Metabolic Disorders
Housekeeping gene effects have metabolic implications: GAPDH modulation affects glycolytic flux;[270,271] FDFT1 regulation influences cholesterol biosynthesis;[272] ATF4 controls amino acid homeostasis;[273,274,275,276] connections to documented effects on Adiponectin, PGC1α, and BCAT1 suggest comprehensive metabolic benefits. Importantly, Metadichol does not cause the insulin resistance and hyperglycemia associated with rapalogs. [277]
Neuroprotection
Chaperone upregulation (HSPA5, HSPA9, CANX) supports clearance of misfolded proteins relevant to neurodegenerative diseases;[278,279,280,281,282] ATF4 activation provides cytoprotection against neuronal stress;[262,263,264,265] PTPRO shows enriched expression in neural populations [283,284,285,286,287] and Metadichol's documented 7× increase in Klotho—a powerful neuroprotective factor—suggests applications in Alzheimer's, Parkinson's, and other neurodegenerative conditions.[288,289]
Cellular Reprogramming and Regenerative Medicine
ATF4 activation facilitates induced pluripotent stem cell (iPSC) generation;[290,291] Metadichol's documented expression of Yamanaka factors (OCT4, SOX2, KLF4, c-MYC, NANOG) in somatic cells; chaperone upregulation supports the ER stress associated with reprogramming.[292,293,294,295,296,297,298,299,300,301,302,303,304,305,306] These findings suggest applications in regenerative medicine and cellular rejuvenation strategies.
We propose a Unified Mechanistic Framework Linking Metadichol–Nuclear Receptor Binding to Pleiotropic Biological Effects Through Housekeeping Gene Modulation this is highlighted in Figure 10. It presents a hierarchical model illustrating how Metadichol exerts its diverse biological effects through a unified upstream mechanism. At the apex, Metadichol directly engages nuclear receptors—specifically the vitamin D receptor (VDR) and retinoid X receptor (RXR)—serving as the primary activation event.
This initial interaction triggers bidirectional modulation of 19 housekeeping genes at the first tier, organized across six functional categories: translation (26%), stress response (26%), immune function (16%), metabolism (16%), cytoskeleton maintenance (11%), and proliferation (5%). The ubiquitous expression of these housekeeping genes provides a mechanistic explanation for Metadichol's pleiotropic effects across multiple tissues and organ systems.
The second tier comprises master regulatory cascades, including transcription factors (ATF4, HIF1A), translational machinery (EIFs, RPLP0), immune modulators (B2M, PTM5, PTPRO), and metabolic enzymes (GAPDH, FDFT1). These cascades subsequently regulate over 50 downstream targets at the third tier, encompassing sirtuins, the mTOR pathway, toll-like receptors, TP53, longevity-associated factors (Klotho, GDF11, telomerase), FOX transcription factors, pluripotency genes (OCT4, KLF4, SOX2), and metabolic regulators.
This hierarchical framework—from a single molecular target to hundreds of downstream effectors—accounts for the therapeutic implications observed across cancer, aging, immunity, metabolic disorders, and regenerative medicine, while distinguishing Metadichol's multi-target, bidirectional approach from conventional single-target interventions.
Conclusions
This comprehensive study of housekeeping gene expression in human PBMCs treated with Metadichol reveals several key findings:
1. Resolution of the HKG Paradox: True constitutive housekeeping genes (ACTB, TUBB, RPLP0, EIF factors) remain stable and are appropriately undruggable, while stress-responsive genes within the HKG panel (ATF4, HIF1A, PTPRO, chaperones) can be safely and beneficially modulated.
2. ATF4 Activation: The 4.06-fold upregulation of ATF4 demonstrates activation of the integrated stress response pathway, providing hormetic cyto-protection and enhanced cellular resilience.
3. PTPRO Tumor Suppressor Induction: The remarkable 6.02-fold upregulation of PTPRO represents restoration of a tumor suppressor frequently silenced in cancers, suggesting anti-neoplastic mechanisms.
4. HIF1A Biphasic Regulation: The complex HIF1A response pattern reflects the interplay between mTOR and SIRT1 pathways, both of which are modulated by Metadichol.
5. Ultra-Low Dose Activity: Significant gene modulation at 0.1 pg/mL (10-13 g/mL) demonstrates the remarkable potency of nano-emulsion delivery combined with nuclear receptor-mediated signal amplification.
6. Unifying Mechanism Established: We propose housekeeping gene modulation as the upstream "master switch" explaining Metadichol's diverse biological effects through hierarchical network amplification from 49 nuclear receptors → 19 HKGs → master TFs → 1000s of downstream genes.
7. Broad Therapeutic Potential: The combination of tumor suppressor restoration, stress response activation, immune enhancement, and metabolic regulation suggests applications across oncology, aging, neurodegeneration, and metabolic disease—all achieved with an exceptional safety profile (LD50 >5000 mg/kg).
These findings position Metadichol as a first-in-class compound capable of comprehensive housekeeping gene modulation through a physiological nuclear receptor-mediated mechanism, representing a paradigm shift from "undruggable" to therapeutic for this fundamental class of cellular regulators.
Supplementary Information
Raw data is available on request
The author is the founder of Nanorx Inc. and is a major shareholder in the company.
This study was conducted independently by a third-party external laboratory on commercial terms to eliminate bias in our results
Abbreviations
| ACTB | Actin Beta |
| ATF4 | Activating Transcription Factor 4 |
| ATF6 | Activating Transcription Factor 6 |
| B2M | Beta-2-Microglobulin |
| BCAT1 | Branched Chain Amino Acid Transaminase 1 |
| BiP | Binding Immunoglobulin Protein (HSPA5) |
| CANX | Calnexin |
| CLL | Chronic Lymphocytic Leukemia |
| Cq | Quantification Cycle |
| DDIT4 | DNA Damage Inducible Transcript 4 |
| EIF1 | Eukaryotic Translation Initiation Factor 1 |
| EIF2S2 | Eukaryotic Translation Initiation Factor 2 Subunit Beta |
| EIF4A2 | Eukaryotic Translation Initiation Factor 4A2 |
| EIF5 | Eukaryotic Translation Initiation Factor 5 |
| ER | Endoplasmic Reticulum |
| FBS | Fetal Bovine Serum |
| FDFT1 | Farnesyl-Diphosphate Farnesyltransferase 1 (Squalene Synthase) |
| FOXO | Forkhead Box O |
| GAPDH | Glyceraldehyde-3-Phosphate Dehydrogenase |
| GDF11 | Growth Differentiation Factor 11 |
| GRP78 | Glucose-Regulated Protein 78 (HSPA5) |
| HIF1A | Hypoxia Inducible Factor 1 Subunit Alpha |
| HKG | Housekeeping Gene |
| HPA | Human Protein Atlas |
| HPRT1 | Hypoxanthine Phosphoribosyl transferase 1 |
| HSPA5 | Heat Shock Protein Family A Member 5 (GRP78/BiP) |
| HSPA9 | Heat Shock Protein Family A Member 9 (Mortalin) |
| iPSC | Induced Pluripotent Stem Cell |
| IRE1 | Inositol-Requiring Enzyme 1 |
| ISR | Integrated Stress Response |
| KLF | Krüppel-Like Factor |
| LD50 | Median Lethal Dose |
| LXR | Liver X Receptor |
| MHC-I | Major Histocompatibility Complex Class I |
| mRNA | Messenger RNA |
| mTOR | Mechanistic Target of Rapamycin |
| mTORC1 | mTOR Complex 1 |
| NK | Natural Killer |
| NR | Nuclear Receptor |
| nTPM | Normalized Transcripts Per Million |
| PBMC | Peripheral Blood Mononuclear Cell |
| PBS | Phosphate-Buffered Saline |
| PERK | PKR-like ER Kinase |
| PGC1α | Peroxisome Proliferator-Activated Receptor Gamma Coactivator 1-Alpha |
| PPAR | Peroxisome Proliferator-Activated Receptor |
| PTEN | Phosphatase and Tensin Homolog |
| PTMA | Prothymosin Alpha |
| PTMS | Parathymosin |
| PTPRO | Protein Tyrosine Phosphatase Receptor Type O |
| qRT-PCR | Quantitative Real-Time Polymerase Chain Reaction |
| RNA | Ribonucleic Acid |
| RORC | RAR-Related Orphan Receptor C |
| RPLP0 | Ribosomal Protein Lateral Stalk Subunit P0 |
| RXR | Retinoid X Receptor |
| SD | Standard Deviation |
| SIRT | Sirtuin |
| SIRT1-7 | Sirtuin 1 through 7 |
| SOX | SRY-Box |
| TERT | Telomerase Reverse Transcriptase |
| TF | Transcription Factor |
| TLR | Toll-Like Receptor |
| TP53 | Tumor Protein P53 |
| TUBB | Tubulin Beta Class I |
| UPR | Unfolded Protein Response |
| VDR | Vitamin D Receptor |
| VEGF | Vascular Endothelial Growth Factor |
| VHL | Von Hippel-Lindau |
References
- Eisenberg E, Levanon EY. Human housekeeping genes, revisited. Trends Genet. 2013;29(10):569-574. [CrossRef]
- Hounkpe BW, Chebera F, De Paula EV, Gomes-Alves AG, Grunkraut J, Machado TFG. HRT Atlas v1.0 database: redefining human and mouse housekeeping genes and candidate reference transcripts by mining massive RNA-seq datasets. Nucleic Acids Res. 2021;49(D1):D947-D955. [CrossRef]
- Machado FB, Hounkpe BW, Gomes-Alves AG, et al. What are housekeeping genes? PLoS Comput Biol. 2022;18(7):e1010295. [CrossRef]
- Fendt SM, Bell EL, Keibler MA, et al. Is there a therapeutic window for metabolism-based cancer therapies? Front Endocrinol (Lausanne). 2017;8:150. [CrossRef]
- Zhu P, Tan MJ, Huang RL, et al. Metabolic alterations in cancer cells and therapeutic implications. Chin J Cancer. 2011;30(4):240-249. [CrossRef]
- Zhu W, Huang X, He Z, et al. Mechanisms of metabolic reprogramming in cancer cells supporting enhanced growth and proliferation. Cells. 2021;10(5):1056. [CrossRef]
- Zhe-Sheng C, Wei-Wei L, Jing-Jing W, et al. Definition, conservation and epigenetics of housekeeping and tissue-enriched genes. BMC Genomics. 2009;10:269. [CrossRef]
- Olsvik PA, Lie KK, Jordal AE, et al. Toxicological housekeeping genes: do they really keep the house? Environ Sci Technol. 2007;41(9):3338-3345. [CrossRef]
- Derks MFL, Megens HJ, Bosse M, et al. Loss of function mutations in essential genes cause embryonic lethality in pigs. PLoS Genet. 2019;15(3):e1008055. [CrossRef]
- Lv W, Zheng J, Luan M, et al. Comparing the evolutionary conservation between human essential genes, human orthologs of mouse essential genes and human housekeeping genes. Brief Bioinform. 2015;16(6):922-931. [CrossRef]
- Kshirsagar S, Islam MA, Reddy AP, Reddy PH. Resolving the current controversy of use and reuse of housekeeping proteins in ageing research. Ageing Res Rev. 2024;100:102437. [CrossRef]
- Vandesompele J, De Preter K, Pattyn F, et al. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002;3(7):RESEARCH0034. [CrossRef]
- Faubert B, Solmonson A, DeBerardinis RJ. Metabolic reprogramming and cancer progression. Science. 2020;368(6487):eaaw5473. [CrossRef]
- Pavlova NN, Thompson CB. The emerging hallmarks of cancer metabolism. Cell Metab. 2016;23(1):27-47. [CrossRef]
- Graham NA, Minasyan A, Lomber A, et al. The landscape of metabolic pathway dependencies in cancer cell lines. PLoS Comput Biol. 2021;17(4):e1008942. [CrossRef]
- Warburg O, Wind F, Negelein E. The metabolism of tumors in the body. J Gen Physiol. 1927;8(6):519-530. [CrossRef]
- Liberti MV, Locasale JW. The Warburg effect: how does it benefit cancer cells? Trends Biochem Sci. 2016;41(3):211-218. [CrossRef]
- Vaupel P, Multhoff G. Revisiting the Warburg effect: historical dogma versus current understanding. J Physiol. 2021;599(6):1745-1757. [CrossRef]
- Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144(5):646-674. [CrossRef]
- Hanahan D. Hallmarks of cancer: new dimensions. Cancer Discov. 2022;12(1):31-46. [CrossRef]
- Ko YH, Pedersen PL, Geschwind JF. Glucose catabolism in the rabbit VX2 tumor model for liver cancer: characterization and targeting hexokinase. Cancer Lett. 2001;173(1):83-91. [CrossRef]
- Ko YH, Smith BL, Wang Y, et al. Advanced cancers: eradication in all cases using 3-bromopyruvate therapy to deplete ATP. Biochem Biophys Res Commun. 2004;324(1):269-275. [CrossRef]
- Fan T, Sun G, Sun X, et al. Tumor energy metabolism and potential of 3-bromopyruvate as an inhibitor of aerobic glycolysis: implications in tumor treatment. Cancers (Basel). 2019;11(3):317. [CrossRef]
- Hudes G, Carducci M, Tomczak P, et al. Temsirolimus, interferon alfa, or both for advanced renal-cell carcinoma. N Engl J Med. 2007;356(22):2271-2281. [CrossRef]
- Motzer RJ, Escudier B, Oudard S, et al. Efficacy of everolimus in advanced renal cell carcinoma: a double-blind, randomised, placebo-controlled phase III trial. Lancet. 2008;372(9637):449-456. [CrossRef]
- Chiarini F, Evangelisti C, McCubrey JA, Martelli AM. Current treatment strategies for inhibiting mTOR in cancer. Trends Pharmacol Sci. 2015;36(2):124-135. [CrossRef]
- Raghavan PR. US Patent 8,722,093. Policosanol nanoparticles. 2014.
- Raghavan PR. US Patent 9,006,292. Policosanol nanoparticles. 2015.
- Raghavan PR. US Patent 9,034,383. Policosanol nanoparticles. 2015.
- Raghavan PR. Beyond Rapamycin: Metadichol Represents a New Class of Multi-Target mTOR Modulators. Medical Research Archives. 2025;13(9). [DOI not available].
- Raghavan PR. Metadichol®-induced expression of Sirtuins 1-7 in somatic and cancer cells. Medical Research Archives. 2024;12(6). [CrossRef]
- Raghavan PR. Metadichol: An Agonist that Expresses the Anti-Aging Gene Klotho in Various Cell Lines. Fortune Journal of Health Sciences. 2023;6:357-362. [CrossRef]
- Raghavan PR. Metadichol Orchestrates Cellular Reprogramming and Regenerative Pathways via FOX Transcription Factor Networks: Implications for Immune-Metabolic Rejuvenation. Medical Research Archives. 2026;14(1). [CrossRef]
- Raghavan PR. SOX Transcription Factor Network Modulation by Metadichol: A Novel Paradigm for Regenerative and Precision Medicine. Medical Research Archives. 2026;14(1). [CrossRef]
- Raghavan PR. Synergistic Targeting of Krüppel-like factor and Related Signaling Pathways by Metadichol: A Multidimensional Anticancer Strategy. Medical Research Archives. 2025;13(6). [DOI not available].
- Raghavan PR. Metadichol induces CD14 glycoprotein expression in human embryonic stem cells and fibroblasts. Medical Research Archives. 2025;13(5). [DOI not available].
- Raghavan PR. Metadichol, A Modulator that Controls Expression of Toll Like Receptors in Cancer Cell Lines. British Journal of Cancer Research. 2024;7(3):720-732. [CrossRef]
- Raghavan PR. Metadichol-induced expression of Toll receptor family members in peripheral blood mononuclear cells. Medical Research Archives. 2024;12(8). [CrossRef]
- Raghavan PR. Metadichol®-induced expression of circadian clock transcription factors in human fibroblasts. Medical Research Archives. 2024;12(6). [CrossRef]
- Raghavan PR. Metadichol-induced differentiation of pancreatic ductal cells (PANC-1) into insulin-producing cells. Medical Research Archives. 2023;11(11). [CrossRef]
- Raghavan PR. Metadichol®-induced expression of Sirtuins 1-7 in somatic and cancer cells. Medical Research Archives. 2024;12(6). [CrossRef]
- Raghavan PR. Metadichol® Induced the Expression of Neuronal Transcription Factors in Human Fibroblast Dermal Cells. Journal of Bioinformatics and Systems Biology. 2023;6:326-339. [CrossRef]
- Raghavan PR. Metadichol orchestrates pluripotency via nuclear receptors during cellular reprogramming. Med Res Arch. 2025;13(6). [CrossRef]
- Raghavan PR. Metadichol Stimulates Gene Expression Across Mammalian Species: Dose Response Characterization and Implications for Restoring Vitamin C Biosynthesis. Preprints. 2025;2025081935. [CrossRef]
- Raghavan PR. Metadichol® induced high levels of vitamin C: case studies. Vitam Miner. 2017;6:169. [DOI not available].
- Raghavan PR. Metadichol® and Vitamin C Increase In Vivo, an Open-Label Study. Vitam Miner. 2017;6:163. [DOI not available].
- Raghavan PR. The Quest for Immortality: Introducing Metadichol® a Novel Telomerase Activator. J Stem Cell Res Ther. 2019;9:446. [CrossRef]
- Raghavan PR. Improving Longevity with Metadichol® by Inhibiting the Bcat-1 Gene. Journal of Aging Science. 2017;5:1. [CrossRef]
- Raghavan PR. Metadichol, A Novel ROR Gamma Inverse Agonist and Its Applications in Psoriasis. J Clin Exp Dermatol Res. 2017;8:433. [CrossRef]
- Raghavan PR. Metadichol® A Novel Inverse Agonist of Thyroid Receptor and its Applications in Thyroid Diseases. Biol Med (Aligarh). 2019;11:458. [CrossRef]
- Raghavan PR. Metadichol®. A Novel Inverse Agonist of Aryl Hydrocarbon Receptor (AHR) and NRF2 Inhibitor. J Cancer Sci Ther. 2017;9:661-668. [CrossRef]
- Raghavan PR. VDR inverse agonism by metadichol enhances VDBP-mediated immunity. Preprints.org. 2025. [CrossRef]
- Raghavan PR. Metadichol®: A Nano Lipid Emulsion that Expresses All 49 Nuclear Receptors in Stem and Somatic Cells. Archives of Clinical and Biomedical Research. 2023;7:524-536. [CrossRef]
- Karlsson M, Zhang C, Méar L, et al. A single-cell type transcriptomics map of human tissues. Sci Adv. 2021;7(31):eabh2169. [CrossRef]
- Digre A, Lindskog C. The human protein atlas—integrated omics for single cell mapping of the human proteome. Protein Sci. 2023;32(1):e4562. [CrossRef]
- Amberger JS, Bocchini CA, Schiettecatte F, Scott AF, Hamosh A. OMIM.org: Online Mendelian Inheritance in Man (OMIM®), an online catalog of human genes and genetic disorders. Nucleic Acids Res. 2015;43(D1):D789-D798. [CrossRef]
- Piñero J, Bravo À, Queralt-Rosinach N, et al. DisGeNET: a comprehensive platform integrating information on human disease-associated genes and variants. Nucleic Acids Res. 2017;45(D1):D833-D839. [CrossRef]
- Wharram BL, Goyal M, Gillespie PJ, et al. Altered podocyte structure in GLEPP1 (Ptpro)-deficient mice associated with hypertension and low glomerular filtration rate. J Clin Invest. 2000;106(10):1281-1290. [CrossRef]
- Jiang W, Wei M, Liu M, et al. Identification of protein tyrosine phosphatase receptor type O (PTPRO) as a synaptic adhesion molecule that promotes synapse formation. J Neurosci. 2017;37(41):9828-9843. [CrossRef]
- Lapointe CP, Grosely R, Djumagulov M, et al. eIF1 and eIF5 dynamically control translation start site fidelity. Nat Struct Mol Biol. 2025;32(7):1085-1096. [CrossRef]
- Hinnebusch AG. The scanning mechanism of eukaryotic translation initiation. Annu Rev Biochem. 2014;83:779-812. [CrossRef]
- Rich BE, Steitz JA. Human acidic ribosomal phosphoproteins P0, P1, and P2: analysis of cDNA clones, in vitro synthesis, and assembly. Mol Cell Biol. 1987;7(11):4065-4074. [CrossRef]
- Pakos-Zebrucka K, Koryga I, Mnich K, Ljujic M, Samali A, Gorman AM. The integrated stress response. EMBO Rep. 2016;17(10):1374-1395. [CrossRef]
- Lee JW, Bae SH, Jeong JW, Kim SH, Kim KW. Hypoxia-inducible factor (HIF-1)α: its protein stability and biological functions. Exp Mol Med. 2004;36(1):1-12. [CrossRef]
- Lee AS. Glucose-regulated proteins in cancer: molecular mechanisms and therapeutic potential. Nat Rev Cancer. 2014;14(4):263-276. [CrossRef]
- Hetz C, Zhang K, Bhattacharya R. Mechanisms, regulation and functions of the unfolded protein response. Nat Rev Mol Cell Biol. 2020;21(8):421-438. [CrossRef]
- Ni M, Zhang Y, Lee AS. Beyond the endoplasmic reticulum: atypical GRP78 in cell viability, signalling and therapeutic targeting. Biochem J. 2011;434(2):181-188. [CrossRef]
- Luo B, Lee AS. The critical roles of endoplasmic reticulum chaperones and unfolded protein response in tumorigenesis and anticancer therapies. Oncogene. 2013;32(7):805-818. [CrossRef]
- Li L, Dong M, Wang XG. The implication and significance of beta 2 microglobulin: a conservative multifunctional regulator. Chin Med J (Engl). 2016;129(4):448-455. [CrossRef]
- Vitiello A, Potter TA, Sherman LA. The role of β2-microglobulin in peptide binding by class I molecules. Science. 1990;250(4986):1423-1426. [CrossRef]
- Zhang S, Liu X, Zhang Y, et al. The immune-related role of beta-2-microglobulin in melanoma. Front Oncol. 2022;12:944722. [CrossRef]
- Sirover MA. On the functional diversity of glyceraldehyde-3-phosphate dehydrogenase: biochemical mechanisms and regulatory control. Biochim Biophys Acta. 2011;1810(8):741-751. [CrossRef]
- Colell A, Green DR, Bhattacharya R. Novel roles for GAPDH in cell death and carcinogenesis. Cell Death Differ. 2009;16(12):1573-1581. [CrossRef]
- Lazarev VF, Guzhova IV, Margulis BA. Glyceraldehyde-3-phosphate dehydrogenase is a multifaceted therapeutic target. Pharmaceutics. 2020;12(5):416. [CrossRef]
- Bunnell TM, Burbach BJ, Shimizu Y, Bhattacharya R. β-Actin specifically controls cell growth, migration, and the G-actin pool. Mol Biol Cell. 2011;22(21):4047-4058. [CrossRef]
- Pollard TD, Cooper JA. Actin, a central player in cell shape and movement. Science. 2009;326(5958):1208-1212. [CrossRef]
- Piñeiro A, Cordero OJ, Nogueira M. Prothymosin alpha is involved in the regulation of cell proliferation. Histol Histopathol. 2000;15(1):265-276. [CrossRef]
- Karetsou Z, Sandaltzopoulos R, Frangou-Lazaridis M, et al. Prothymosin alpha modulates the interaction of histone H1 with chromatin. Nucleic Acids Res. 1998;26(13):3111-3118. [CrossRef]
- Gómez-Márquez J, Rodríguez P. Prothymosin alpha is a chromatin-remodeling protein in mammalian cells. Biochem J. 1998;333(Pt 1):1-3. [CrossRef]
- Böyum A. Isolation of mononuclear cells and granulocytes from human blood. Scand J Clin Lab Invest Suppl. 1968;97:77-89.
- Grievink HW, Luiber T, Moerland M, et al. Comparison of three isolation techniques for human peripheral blood mononuclear cells: cell recovery and viability, population composition, and cell functionality. Biopreserv Biobank. 2016;14(5):410-415. [CrossRef]
- Chomczynski P, Sacchi N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem. 1987;162(1):156-159. [CrossRef]
- Bustin SA, Benes V, Garson JA, et al. The MIQE guidelines: minimum information for publication of quantitative real-time PCR experiments. Clin Chem. 2009;55(4):611-622. [CrossRef]
- Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001;25(4):402-408. [CrossRef]
- Schmittgen TD, Livak KJ. Analyzing real-time PCR data by the comparative C(T) method. Nat Protoc. 2008;3(6):1101-1108. [CrossRef]
- Pakos-Zebrucka K, Koryga I, Mnich K, Ljujic M, Samali A, Gorman AM. The integrated stress response. EMBO Rep. 2016;17(10):1374-1395. [CrossRef]
- Wortel IMN, van der Meer LT, Kilberg MS, van Leeuwen FN. Surviving stress: modulation of ATF4-mediated stress responses in normal and malignant cells. Trends Endocrinol Metab. 2017;28(11):794-806. [CrossRef]
- Harding HP, Zhang Y, Zeng H, et al. An integrated stress response regulates amino acid metabolism and resistance to oxidative stress. Mol Cell. 2003;11(3):619-633. [CrossRef]
- Vattem KM, Wek RC. Reinitiation involving upstream ORFs regulates ATF4 mRNA translation in mammalian cells. Proc Natl Acad Sci U S A. 2004;101(31):11269-11274. [CrossRef]
- B'chir W, Maurin AC, Carraro V, et al. The eIF2α/ATF4 pathway is essential for stress-induced autophagy gene expression. Nucleic Acids Res. 2013;41(16):7683-7699. [CrossRef]
- Ye J, Koumenis C. ATF4, an ER stress and hypoxia-inducible transcription factor and its potential role in hypoxia tolerance and tumorigenesis. Curr Mol Med. 2009;9(4):411-416. [CrossRef]
- Kilberg MS, Shan J, Su N. ATF4-dependent transcription mediates signaling of amino acid limitation. Trends Endocrinol Metab. 2009;20(9):436-443. [CrossRef]
- Ameri K, Harris AL. Activating transcription factor 4. Int J Biochem Cell Biol. 2008;40(1):14-21. [CrossRef]
- Liu Z, Gu H, Gan L, et al. Reducing Smad3/ATF4 was essential for Sirt1 inhibiting ER stress-induced apoptosis in mice brown adipose tissue. Oncotarget. 2017;8(6):9267-9279. [CrossRef]
- Ghosh HS, Reizis B, Bhattacharya D. SIRT1 associates with eIF2-alpha and regulates the cellular stress response. Sci Rep. 2011;1:150. [CrossRef]
- Lim JH. SIRT1 suppresses activating transcription factor 4 (ATF4) expression in response to proteasome inhibition. Biochem Biophys Res Commun. 2013;441(2):289-294. [CrossRef]
- Brugarolas J, Lei K, Hurley RL, et al. Regulation of mTOR function in response to hypoxia by REDD1 and the TSC1/TSC2 tumor suppressor complex. Genes Dev. 2004;18(23):2893-2904. [CrossRef]
- DeYoung MP, Horak P, Sofer A, Sgroi D, Ellisen LW. Hypoxia regulates TSC1/2-mTOR signaling and tumor suppression through REDD1-mediated 14-3-3 shuttling. Genes Dev. 2008;22(2):239-251. [CrossRef]
- Pike JW, Meyer MB. The vitamin D receptor: new paradigms for the regulation of gene expression by 1,25-dihydroxyvitamin D3. Endocrinol Metab Clin North Am. 2010;39(2):255-269. [CrossRef]
- Bikle DD. Vitamin D metabolism, mechanism of action, and clinical applications. Chem Biol. 2014;21(3):319-329. [CrossRef]
- Wharram BL, Goyal M, Gillespie PJ, et al. Altered podocyte structure in GLEPP1 (Ptpro)-deficient mice associated with hypertension and low glomerular filtration rate. J Clin Invest. 2000;106(10):1281-1290. [CrossRef]
- Jiang W, Wei M, Liu M, et al. Identification of protein tyrosine phosphatase receptor type O (PTPRO) as a synaptic adhesion molecule that promotes synapse formation. J Neurosci. 2017;37(41):9828-9843. [CrossRef]
- Xie F, Dong H, Zhang H. Regulatory functions of protein tyrosine phosphatase receptor type O in immune cells. Front Immunol. 2021;12:783370. [CrossRef]
- Motiwala T, Ghoshal K, Das A, et al. Suppression of the protein tyrosine phosphatase receptor type O gene (PTPRO) by methylation in hepatocellular carcinomas. Oncogene. 2003;22(41):6319-6331. [CrossRef]
- Motiwala T, Kutay H, Ghoshal K, et al. Protein tyrosine phosphatase receptor-type O (PTPRO) exhibits characteristics of a candidate tumor suppressor in human lung cancer. Proc Natl Acad Sci U S A. 2004;101(38):13844-13849. [CrossRef]
- Motiwala T, Majumder S, Kutay H, et al. Methylation and silencing of protein tyrosine phosphatase receptor type O in chronic lymphocytic leukemia. Clin Cancer Res. 2007;13(11):3174-3181. [CrossRef]
- Hsu SH, Motiwala T, Roy S, et al. Methylation of the PTPRO gene in human hepatocellular carcinoma and identification of VCP as its substrate. J Cell Biochem. 2013;114(8):1810-1818. [CrossRef]
- Jacob ST, Bhattacharya R. Epigenetic regulation of protein tyrosine phosphatases: potential molecular targets for cancer therapy. Genes Cancer. 2010;1(12):1232-1239. [CrossRef]
- Li SY, Li R, Chen YL, et al. Aberrant PTPRO methylation in tumor tissues as a potential biomarker that predicts clinical outcomes in breast cancer patients. BMC Genet. 2014;15:67. [CrossRef]
- Vousden KH, Prives C. Blinded by the light: the growing complexity of p53. Cell. 2009;137(3):413-431. [CrossRef]
- Kastenhuber ER, Lowe SW. Putting p53 in context. Cell. 2017;170(6):1062-1078. [CrossRef]
- Levine AJ. p53: 800 million years of evolution and 40 years of discovery. Nat Rev Cancer. 2020;20(8):471-480. [CrossRef]
- Eijkelenboom A, Burgering BM. FOXOs: signalling integrators for homeostasis maintenance. Nat Rev Mol Cell Biol. 2013;14(2):83-97. [CrossRef]
- Webb AE, Brunet A. FOXO transcription factors: key regulators of cellular quality control. Trends Biochem Sci. 2014;39(4):159-169. [CrossRef]
- Semenza GL. Hypoxia-inducible factors in physiology and medicine. Cell. 2012;148(3):399-408. [CrossRef]
- Majmundar AJ, Wong WJ, Simon MC. Hypoxia-inducible factors and the response to hypoxic stress. Mol Cell. 2010;40(2):364-374. [CrossRef]
- Dengler VL, Galbraith M, Bhattacharya R. Transcriptional regulation by hypoxia inducible factors. Crit Rev Biochem Mol Biol. 2014;49(1):1-15. [CrossRef]
- Lim JH, Lee YM, Chun YS, Chen J, Kim JE, Park JW. Sirtuin 1 modulates cellular responses to hypoxia by deacetylating hypoxia-inducible factor 1alpha. Mol Cell. 2010;38(6):864-878. [CrossRef]
- Joo HY, Yun M, Jeong J, et al. SIRT1 deacetylates and stabilizes hypoxia-inducible factor-1α (HIF-1α) via direct interactions during hypoxia. Biochem Biophys Res Commun. 2015;462(4):294-300. [CrossRef]
- Laughner E, Taghavi P, Chiles K, Mahon PC, Semenza GL. HER2 (neu) signaling increases the rate of hypoxia-inducible factor 1alpha (HIF-1alpha) synthesis: novel mechanism for HIF-1-mediated vascular endothelial growth factor expression. Mol Cell Biol. 2001;21(12):3995-4004. [CrossRef]
- Zhong L, D'Urso A, Toiber D, et al. The histone deacetylase Sirt6 regulates glucose homeostasis via Hif1alpha. Cell. 2010;140(2):280-293. [CrossRef]
- Shoshani T, Faerman A, Mett I, et al. Identification of a novel hypoxia-inducible factor 1-responsive gene, RTP801, involved in apoptosis. Mol Cell Biol. 2002;22(7):2283-2293. [CrossRef]
- Bjorkman, P. J., and P. Parham. 1990. "Structure, Function, and Diversity of Class I Major Histocompatibility Complex Molecules." Annual Review of Biochemistry 59:253-288.
- Güssow, D., R. Rein, I. Ginjaar, F. Hochstenbach, G. Seemann, A. Kottman, and H. L. Ploegh. 1987. "The Human Beta 2-Microglobulin Gene: Primary Structure and Definition of the Transcriptional Unit." Journal of Immunology 139(9):3132-3138.
- Germain, R. N. 1994. "MHC-Dependent Antigen Processing and Peptide Presentation: Providing Ligands for T Lymphocyte Activation." Cell 76(2):287-299.
- Cresswell, P., A. L. Ackerman, A. Giodini, D. R. Peaper, and P. A. Wearsch. 2005. "Mechanisms of MHC Class I-Restricted Antigen Processing and Cross-Presentation." Immunological Reviews 207:145-157.
- Townsend, A., and H. Bodmer. 1989. "Antigen Recognition by Class I-Restricted T Lymphocytes." Annual Review of Immunology 7:601-624.
- Neefjes, J., M. L. Jongsma, P. Paul, and O. Bakke. 2011. "Towards a Systems Understanding of MHC Class I and MHC Class II Antigen Presentation." Nature Reviews Immunology 11(12):823-836.
- Feder, J. N., A. Gnirke, W. Thomas, Z. Tsuchihashi, D. A. Ruddy, A. Basava, et al. 1996. "A Novel MHC Class I-Like Gene Is Mutated in Patients with Hereditary Haemochromatosis." Nature Genetics 13(4):399-408.
- Ganz, T. 2013. "Systemic Iron Homeostasis." Physiological Reviews 93(4):1721-1741.
- Roopenian, D. C., and S. Akilesh. 2007. "FcRn: The Neonatal Fc Receptor Comes of Age." Nature Reviews Immunology 7(9):715-725.
- Gobin, S. J., V. Keijsers, M. van Zutphen, and P. J. van den Elsen. 1998. "The Role of Enhancer A in the Locus-Specific Transactivation of Classical and Nonclassical HLA Class I Genes by Nuclear Factor κB." Journal of Immunology 161(5):2276-2283.
- Meissner, T. B., A. Li, A. Biswas, K. H. Lee, Y. J. Liu, E. Bayber, et al. 2010. "NLR Family Member NLRC5 Is a Transcriptional Regulator of MHC Class I Genes." Proceedings of the National Academy of Sciences107(31):13794-13799.
- Boehm, U., T. Klamp, M. Groot, and J. C. Howard. 1997. "Cellular Responses to Interferon-Gamma." Annual Review of Immunology 15:749-795.
- Taniguchi, T., and A. Takaoka. 2001. "A Weak Signal for Strong Responses: Interferon-Alpha/Beta Revisited." Nature Reviews Molecular Cell Biology 2(5):378-386.
- Hewison, M. 2012. "Vitamin D and Immune Function: An Overview." Proceedings of the Nutrition Society71(1):50-61.
- Carlberg, C. 2019. "Vitamin D Signaling in the Context of Innate Immunity: Focus on Human Monocytes." Frontiers in Immunology 10:2211.
- Mora, J. R., M. Iwata, and U. H. von Andrian. 2008. "Vitamin Effects on the Immune System: Vitamins A and D Take Centre Stage." Nature Reviews Immunology 8(9):685-698.
- Pakos-Zebrucka, K., I. Koryga, K. Mnich, M. Ljujic, A. Samali, and A. M. Gorman. 2016. "The Integrated Stress Response." EMBO Reports 17(10):1374-1395.
- Donnelly, N., A. M. Gorman, A. Gupta, and A. Bhattacharya. 2013. "The eIF2α Kinases: Their Structures and Functions." Cellular and Molecular Life Sciences 70(19):3493-3511.
- McNab, F., K. Mayer-Barber, A. Sher, A. Wack, and A. O'Garra. 2015. "Type I Interferons in Infectious Disease." Nature Reviews Immunology 15(2):87-103.
- Yeung, F., J. E. Hoberg, C. S. Ramsey, M. D. Keller, D. R. Jones, R. A. Frye, et al. 2004. "Modulation of NF-κB-Dependent Transcription and Cell Survival by the SIRT1 Deacetylase." EMBO Journal 23(12):2369-2380.
- Laplante, M., and D. M. Sabatini. 2012. "mTOR Signaling in Growth Control and Disease." Cell 149(2):274-293.
- Kauppinen, A., T. Suuronen, J. Ojala, K. Kaarniranta, and A. Salminen. 2013. "Antagonistic Crosstalk between NF-κB and SIRT1 in the Regulation of Inflammation and Metabolic Disorders." Cellular Signalling 25(10):1939-1948.
- Liu, T. F., and C. E. McCall. 2013. "Deacetylation by SIRT1 Reprograms Inflammation and Cancer." Genes and Cancer 4(3-4):135-147.
- Chen, L., H. Deng, H. Cui, J. Fang, Z. Zuo, J. Deng, et al. 2018. "Inflammatory Responses and Inflammation-Associated Diseases in Organs." Oncotarget 9(6):7204-7218.
- Ellisen LW, Ramsayer KD, Johannessen CM, et al. REDD1, a developmentally regulated transcriptional target of p63 and p53, links p63 to regulation of reactive oxygen species. Mol Cell. 2002;10(5):995-1005. [CrossRef]
- Wang M, Wey S, Zhang Y, Ye R, Lee AS. Role of the unfolded protein response regulator GRP78/BiP in development, cancer, and neurological disorders. Antioxid Redox Signal. 2009;11(9):2307-2316. [CrossRef]
- Lee AS. Stress-induced translocation of the endoplasmic reticulum chaperone GRP78/BiP and its impact on human disease and therapy. Proc Natl Acad Sci U S A. 2025;122(30):e2412246122. [CrossRef]
- Hetz C, Zhang K, Bhalla AK. Mechanisms, regulation and functions of the unfolded protein response. Nat Rev Mol Cell Biol. 2020;21(8):421-438. [CrossRef]
- Ni M, Zhang Y, Lee AS. Beyond the endoplasmic reticulum: atypical GRP78 in cell viability, signalling and therapeutic targeting. Biochem J. 2011;434(2):181-188. [CrossRef]
- Starenki D, Hong SK, Lloyd RV, Park JI. Mortalin (GRP75/HSPA9) upregulation promotes survival and proliferation of medullary thyroid carcinoma cells. Oncogene. 2015;34(35):4624-4634. [CrossRef]
- Ferré CA, Bhattacharyya S, Bhattacharyya P, et al. Mortalin/Hspa9 involvement and therapeutic perspective in Parkinson's disease. Int J Mol Sci. 2022;23(17):9823. [CrossRef]
- Goswami AV, Samber S, Bhattacharyya S, et al. HSPA9/Mortalin mediates axo-protection and modulates mitochondrial dynamics in neurons. Sci Rep. 2021;11(1):17705. [CrossRef]
- Schrag JD, Bergeron JJ, Li Y, et al. The structure of calnexin, an ER chaperone involved in quality control of protein folding. Mol Cell. 2001;8(3):633-644. [CrossRef]
- Lamriben L, Graham JB, Adams BM, Bhattacharyya S. Calnexin cycle - structural features of the ER chaperone system. FEBS J. 2020;287(20):4322-4340. [CrossRef]
- Williams DB. Beyond lectins: the calnexin/calreticulin chaperone system of the endoplasmic reticulum. J Cell Sci. 2006;119(Pt 4):615-623. [CrossRef]
- Westerheide SD, Anckar J, Stevens SM Jr, Sistonen L, Morimoto RI. Stress-inducible regulation of heat shock factor 1 by the deacetylase SIRT1. Science. 2009;323(5917):1063-1066. [CrossRef]
- van de Ven RAH, Santos D, Bhattacharyya S. Chaperones and beyond as key players in pluripotency maintenance. Front Cell Dev Biol. 2019;7:150. [CrossRef]
- van Dam S, Võsa U, van der Graaf A, Franke L, de Magalhães JP. Gene co-expression analysis for functional classification and gene-disease predictions. Brief Bioinform. 2018;19(4):575-592. [CrossRef]
- Vandesompele J, De Preter K, Pattyn F, et al. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002;3(7):RESEARCH0034. [CrossRef]
- Hounkpe BW, Cheber F, Grisci VDMTD, de Lima F, De Paula EV. HRT Atlas v1.0 database: redefining human and mouse housekeeping genes and candidate reference transcripts by mining massive RNA-seq datasets. Nucleic Acids Res. 2022;50(D1):D605-D614. [CrossRef]
- Eisenberg E, Levanon EY. Human housekeeping genes, revisited. Trends Genet. 2013;29(10):569-574. [CrossRef]
- Zhu J, He F, Hu S, Yu J. On the nature of human housekeeping genes. Trends Genet. 2008;24(10):481-484. [CrossRef]
- Lemma S, Avnet S, Salerno M, Chano T, Baldini N. Identification and validation of housekeeping genes for gene expression analysis of cancer stem cells. PLoS One. 2016;11(2):e0149481. [CrossRef]
- Thellin O, Zorzi W, Lakaye B, et al. Housekeeping genes as internal standards: use and limits. J Biotechnol. 1999;75(2-3):291-295. [CrossRef]
- Caracausi M, Piovesan A, Antonaros F, Strippoli P, Vitale L, Pelleri MC. Systematic identification of human housekeeping genes possibly useful as references in gene expression studies. Mol Med Rep. 2017;16(3):2397-2410. [CrossRef]
- Wortel IMN, van der Meer LT, Kilberg MS, van Leeuwen FN. Surviving stress: modulation of ATF4-mediated stress responses in normal and malignant cells. Trends Endocrinol Metab. 2017;28(11):794-806. [CrossRef]
- Wang GL, Jiang BH, Rue EA, Semenza GL. Hypoxia-inducible factor 1 is a basic-helix-loop-helix-PAS heterodimer regulated by cellular O2 tension. Proc Natl Acad Sci U S A. 1995;92(12):5510-5514. [CrossRef]
- Kim R, Emi M, Tanabe K, Murakami S. Role of the unfolded protein response in cell death. Apoptosis. 2006;11(1):5-13. [CrossRef]
- Hetz C, Zhang K, Bhakta K. Mechanisms, regulation and functions of the unfolded protein response. Nat Rev Mol Cell Biol. 2020;21(8):421-438. [CrossRef]
- Dang CV. MYC on the path to cancer. Cell. 2012;149(1):22-35. [CrossRef]
- Lambert SA, Jolma A, Campitelli LF, et al. The human transcription factors. Cell. 2018;172(4):650-665. [CrossRef]
- Li X, Jiang Y, Meisenhelder J, et al. Mitochondria-translocated PGK1 functions as a protein kinase to coordinate glycolysis and the TCA cycle in tumorigenesis. Mol Cell. 2016;61(5):705-719. [CrossRef]
- Liberti MV, Locasale JW. The Warburg effect: how does it benefit cancer cells? Trends Biochem Sci. 2016;41(3):211-218. [CrossRef]
- Zhang W, Xu Q, Liu P, et al. Discovery of novel glyceraldehyde-3-phosphate dehydrogenase inhibitor via docking-based virtual screening. Bioorg Chem. 2020;96:103605. [CrossRef]
- Lamming DW, Ye L, Katajisto P, et al. Rapamycin-induced insulin resistance is mediated by mTORC2 loss and uncoupled from longevity. Science. 2012;335(6076):1638-1643. [CrossRef]
- Barlow AD, Nicholson ML, Herbert TP. Evidence for rapamycin toxicity in pancreatic β-cells and a review of the underlying molecular mechanisms. Diabetes. 2013;62(8):2674-2682. [CrossRef]
- Fang Y, Westbrook R, Hill C, et al. Duration of rapamycin treatment has differential effects on metabolism in mice. Cell Metab. 2013;17(3):456-462. [CrossRef]
- Yang SB, Lee HY, Young DM, et al. Rapamycin induces glucose intolerance in mice by reducing islet mass, insulin content, and insulin sensitivity. J Mol Med. 2012;90(8):897-905. [CrossRef]
- Houde VP, Brûlé S, Bhullar KS, et al. Chronic rapamycin treatment causes glucose intolerance and hyperlipidemia by upregulating hepatic gluconeogenesis and impairing lipid deposition in adipose tissue. Diabetes. 2010;59(6):1338-1348. [CrossRef]
- Alemán CL, Más R, Hernández C, et al. A 12-month study of policosanol oral toxicity in Sprague Dawley rats. Toxicol Lett. 1994;70(1):77-87. [CrossRef]
- Alemán CL, Más Ferreiro R, Noa Puig M, et al. Carcinogenicity of policosanol in Sprague-Dawley rats: a 24-month study. Teratog Carcinog Mutagen. 1994;14(5):239-249. [CrossRef]
- Califano A, Alvarez MJ. The recurrent architecture of tumour initiation, progression and drug sensitivity. Nat Rev Cancer. 2017;17(2):116-130. [CrossRef]
- Pakos-Zebrucka K, Koryga I, Mnich K, Ljujic M, Samali A, Gorman AM. The integrated stress response. EMBO Rep. 2016;17(10):1374-1395. [CrossRef]
- Harding HP, Zhang Y, Zeng H, et al. An integrated stress response regulates amino acid metabolism and resistance to oxidative stress. Mol Cell. 2003;11(3):619-633. [CrossRef]
- Ye J, Kumanova M, Hart LS, et al. The GCN2-ATF4 pathway is critical for tumour cell survival and proliferation in response to nutrient deprivation. EMBO J. 2010;29(12):2082-2096. [CrossRef]
- B'chir W, Maurin AC, Carraro V, et al. The eIF2α/ATF4 pathway is essential for stress-induced autophagy gene expression. Nucleic Acids Res. 2013;41(16):7683-7699. [CrossRef]
- Webb AE, Brunet A. FOXO transcription factors: key regulators of cellular quality control. Trends Biochem Sci. 2014;39(4):159-169. [CrossRef]
- Vattem KM, Wek RC. Reinitiation involving upstream ORFs regulates ATF4 mRNA translation in mammalian cells. Proc Natl Acad Sci U S A. 2004;101(31):11269-11274. [CrossRef]
- Semenza GL. Hypoxia-inducible factors in physiology and medicine. Cell. 2012;148(3):399-408. [CrossRef]
- Pugh CW, Ratcliffe PJ. Regulation of angiogenesis by hypoxia: role of the HIF system. Nat Med. 2003;9(6):677-684. [CrossRef]
- Kim JW, Tchernyshyov I, Semenza GL, Dang CV. HIF-1-mediated expression of pyruvate dehydrogenase kinase: a metabolic switch required for cellular adaptation to hypoxia. Cell Metab. 2006;3(3):177-185. [CrossRef]
- Forsythe JA, Jiang BH, Iyer NV, et al. Activation of vascular endothelial growth factor gene transcription by hypoxia-inducible factor 1. Mol Cell Biol. 1996;16(9):4604-4613. [CrossRef]
- Maxwell PH, Wiesener MS, Chang GW, et al. The tumour suppressor protein VHL targets hypoxia-inducible factors for oxygen-dependent proteolysis. Nature. 1999;399(6733):271-275. [CrossRef]
- Sonenberg N, Hinnebusch AG. Regulation of translation initiation in eukaryotes: mechanisms and biological targets. Cell. 2009;136(4):731-745. [CrossRef]
- Ma XM, Blenis J. Molecular mechanisms of mTOR-mediated translational control. Nat Rev Mol Cell Biol. 2009;10(5):307-318. [CrossRef]
- Thoreen CC, Chantranupong L, Keys HR, Wang T, Gray NS, Sabatini DM. A unifying model for mTORC1-mediated regulation of mRNA translation. Nature. 2012;485(7396):109-113. [CrossRef]
- Gingras AC, Raught B, Sonenberg N. eIF4 initiation factors: effectors of mRNA recruitment to ribosomes and regulators of translation. Annu Rev Biochem. 1999;68:913-963. [CrossRef]
- Holcik M, Sonenberg N. Translational control in stress and apoptosis. Nat Rev Mol Cell Biol. 2005;6(4):318-327. [CrossRef]
- Germain RN. MHC-dependent antigen processing and peptide presentation: providing ligands for T lymphocyte activation. Cell. 1994;76(2):287-299. [CrossRef]
- Zaretsky JM, Garcia-Diaz A, Shin DS, et al. Mutations associated with acquired resistance to PD-1 blockade in melanoma. N Engl J Med. 2016;375(9):819-829. [CrossRef]
- Sade-Feldman M, Jiao YJ, Chen JH, et al. Resistance to checkpoint blockade therapy through inactivation of antigen presentation. Nat Commun. 2017;8(1):1136. [CrossRef]
- Lee AS. The ER chaperone and signaling regulator GRP78/BiP as a monitor of endoplasmic reticulum stress. Methods. 2005;35(4):373-381. [CrossRef]
- Wang M, Wey S, Zhang Y, Ye R, Lee AS. Role of the unfolded protein response regulator GRP78/BiP in development, cancer, and neurological disorders. Antioxid Redox Signal. 2009;11(9):2307-2316. [CrossRef]
- Hetz C, Chevet E, Oakes SA. Proteostasis control by the unfolded protein response. Nat Cell Biol. 2015;17(7):829-838. [CrossRef]
- Walter P, Ron D. The unfolded protein response: from stress pathway to homeostatic regulation. Science. 2011;334(6059):1081-1086. [CrossRef]
- Pobre KFR, Poet GJ, Hendershot LM. The endoplasmic reticulum (ER) chaperone BiP is a master regulator of ER functions: Getting by with a little help from ERdj friends. J Biol Chem. 2019;294(6):2098-2108. [CrossRef]
- Goldbeter A, Koshland DE Jr. An amplified sensitivity arising from covalent modification in biological systems. Proc Natl Acad Sci U S A. 1981;78(11):6840-6844. [CrossRef]
- Heinrich R, Neel BG, Rapoport TA. Mathematical models of protein kinase signal transduction. Mol Cell. 2002;9(5):957-970. [CrossRef]
- Ferrell JE Jr. Tripping the switch fantastic: how a protein kinase cascade can convert graded inputs into switch-like outputs. Trends Biochem Sci. 1996;21(12):460-466. [CrossRef]
- Kholodenko BN. Cell-signalling dynamics in time and space. Nat Rev Mol Cell Biol. 2006;7(3):165-176. [CrossRef]
- Britto FA, Dumas K, Giorgetti-Peraldi S, et al. Is REDD1 a metabolic double agent? Lessons from physiology and pathology. Am J Physiol Cell Physiol. 2020;319(5):C807-C824. [CrossRef]
- Pinto JA, Rolfo C, Raez LE, et al. In silico evaluation of DNA damage inducible transcript 4 gene (DDIT4) as prognostic biomarker in several malignancies. Front Oncol. 2018;8:106. [CrossRef]
- Gharibi B, Farzadi S, Ghuman M, Hughes FJ. DDIT4 regulates mesenchymal stem cell fate by mediating between HIF1α and mTOR signalling. Sci Rep. 2016;6:36889. [CrossRef]
- Do H, Bhrolchain M, Bhatt A, et al. Roles of farnesyl-diphosphate farnesyltransferase 1 in tumour and tumour microenvironments. Cells. 2020;9(11):2352. [CrossRef]
- Weng ML, Chen WK, Chen XY, et al. Fasting inhibits aerobic glycolysis and proliferation in colorectal cancer via the Fdft1-mediated AKT/mTOR/HIF1α pathway suppression. Nat Commun. 2020;11(1):1869. [CrossRef]
- Germain RN. MHC-dependent antigen processing and peptide presentation: providing ligands for T lymphocyte activation. Cell. 1994;76(2):287-299. [CrossRef]
- Motiwala T, Ghoshal K, Das A, et al. Suppression of the protein tyrosine phosphatase receptor type O gene (PTPRO) by methylation in hepatocellular carcinomas. Oncogene. 2003;22(41):6319-6331. [CrossRef]
- Pakos-Zebrucka K, Koryga I, Mnich K, Ljujic M, Samali A, Gorman AM. The integrated stress response. EMBO Rep. 2016;17(10):1374-1395. [CrossRef]
- Semenza GL. Hypoxia-inducible factors in physiology and medicine. Cell. 2012;148(3):399-408. [CrossRef]
- Wang M, Kaufman RJ. Protein misfolding in the endoplasmic reticulum as a conduit to human disease. Nature. 2016;529(7586):326-335. [CrossRef]
- Motiwala T, Kutay H, Ghoshal K, et al. Protein tyrosine phosphatase receptor-type O (PTPRO) exhibits characteristics of a candidate tumor suppressor in human lung cancer. Proc Natl Acad Sci U S A. 2004;101(38):13844-13849. [CrossRef]
- Motiwala T, Majumder S. Epigenetic regulation of protein tyrosine phosphatases: potential molecular targets for cancer therapy. Clin Immunol. 2005;117(1):115-125. [DOI not available].
- Motiwala T, Kutay H, Zanesi N, et al. PTPROt-mediated regulation of p53/Foxm1 suppresses leukemic phenotype in a CLL mouse model. Blood. 2015;126(15):1850-1859. [CrossRef]
- Ming F, Sun Q. Epigenetically silenced PTPRO functions as a prognostic marker and tumor suppressor in human lung squamous cell carcinoma. Mol Med Rep. 2017;16(1):746-754. [CrossRef]
- Xu J, Hou J, Wang Y, et al. PTPRO-mediated autophagy prevents hepatosteatosis and tumorigenesis. Oncotarget. 2017;8(6):10373-10383. [CrossRef]
- Thoreen CC, Chantranupong L, Keys HR, Wang T, Gray NS, Sabatini DM. A unifying model for mTORC1-mediated regulation of mRNA translation. Nature. 2012;485(7396):109-113. [CrossRef]
- Silvera D, Formenti SC, Schneider RJ. Translational control in cancer. Nat Rev Cancer. 2010;10(4):254-266. [CrossRef]
- Bhat M, Robichaud N, Hulea L, Sonenberg N, Pelletier J, Topisirovic I. Targeting the translation machinery in cancer. Nat Rev Drug Discov. 2015;14(4):261-278. [CrossRef]
- Liberti MV, Locasale JW. The Warburg effect: how does it benefit cancer cells? Trends Biochem Sci. 2016;41(3):211-218. [CrossRef]
- Ganapathy-Kanniappan S, Geschwind JF. Tumor glycolysis as a target for cancer therapy: progress and prospects. Mol Cancer. 2013;12:152. [CrossRef]
- Zhang JY, Zhang F, Hong CQ, et al. Critical protein GAPDH and its regulatory mechanisms in cancer cells. Cancer Biol Med. 2015;12(1):10-22. [CrossRef]
- Zaretsky JM, Garcia-Diaz A, Shin DS, et al. Mutations associated with acquired resistance to PD-1 blockade in melanoma. N Engl J Med. 2016;375(9):819-829. [CrossRef]
- Sade-Feldman M, Jiao YJ, Chen JH, et al. Resistance to checkpoint blockade therapy through inactivation of antigen presentation. Nat Commun. 2017;8(1):1136. [CrossRef]
- Gettinger S, Choi J, Hastings K, et al. Impaired HLA class I antigen processing and presentation as a mechanism of acquired resistance to immune checkpoint inhibitors in lung cancer. Cancer Discov. 2017;7(12):1420-1435. [CrossRef]
- Maleno I, Aptsiauri N, Cabrera T, Gallego A, Paschen A, López-Nevot MA. Frequent loss of heterozygosity in the β2-microglobulin region of chromosome 15 in primary human tumors. Immunogenetics. 2011;63(2):65-71. [CrossRef]
- del Campo AB, Kyte JA, Carretero J, et al. Immune escape of cancer cells with beta2-microglobulin loss over the course of metastatic melanoma. Int J Cancer. 2014;134(1):102-113. [CrossRef]
- Hicklin DJ, Marincola FM, Ferrone S. HLA class I antigen downregulation in human cancers: T-cell immunotherapy revives an old story. Mol Med Today. 1999;5(4):178-186. [CrossRef]
- Hanahan D, Weinberg RA. The hallmarks of cancer. Cell. 2000;100(1):57-70. [CrossRef]
- Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144(5):646-674. [CrossRef]
- Hanahan D. Hallmarks of cancer: new dimensions. Cancer Discov. 2022;12(1):31-46. [CrossRef]
- Piñero J, Ramírez-Anguita JM, Saüch-Pitarch J, et al. The DisGeNET knowledge platform for disease genomics: 2019 update. Nucleic Acids Res. 2020;48(D1):D845-D855. [CrossRef]
- Rappaport N, Twik M, Plaschkes I, et al. MalaCards: an amalgamated human disease compendium with diverse clinical and genetic annotation and structured search. Nucleic Acids Res. 2017;45(D1):D733-D750. [CrossRef]
- Amberger JS, Bocchini CA, Schiettecatte F, Scott AF, Hamosh A. OMIM.org: Online Mendelian Inheritance in Man (OMIM®), an online catalog of human genes and genetic disorders. Nucleic Acids Res. 2015;43(Database issue):D789-D798. [CrossRef]
- Szklarczyk D, Gable AL, Lyon D, et al. STRING v11: protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019;47(D1):D483-D490. [CrossRef]
- Safran M, Dalah I, Alexander J, et al. GeneCards Version 3: the human gene integrator. Database (Oxford). 2010;2010:baq020. [CrossRef]
- Fontana L, Partridge L, Longo VD. Extending healthy life span--from yeast to humans. Science. 2010;328(5976):321-326. [CrossRef]
- López-Otín C, Blasco MA, Partridge L, Serrano M, Kroemer G. The hallmarks of aging. Cell. 2013;153(6):1194-1217. [CrossRef]
- Kennedy BK, Lamming DW. The mechanistic target of rapamycin: the grand conductor of metabolism and aging. Cell Metab. 2016;23(6):990-1003. [CrossRef]
- Lamming DW, Ye L, Katajisto P, et al. Rapamycin-induced insulin resistance is mediated by mTORC2 loss and uncoupled from longevity. Science. 2012;335(6076):1638-1643. [CrossRef]
- Howell JJ, Manning BD. mTOR couples cellular nutrient sensing to organismal metabolic homeostasis. Trends Endocrinol Metab. 2011;22(3):94-102. [CrossRef]
- Johnson SC, Rabinovitch PS, Kaeberlein M. mTOR is a key modulator of ageing and age-related disease. Nature. 2013;493(7432):338-345. [CrossRef]
- Arriola Apelo SI, Lamming DW. Rapamycin: an inhibiTOR of aging emerges from the soil of Easter Island. J Gerontol A Biol Sci Med Sci. 2016;71(7):841-849. [CrossRef]
- Li W, Miller RA. Elevated ATF4 function in fibroblasts and liver of slow-aging mutant mice. J Gerontol A Biol Sci Med Sci. 2015;70(3):263-272. [CrossRef]
- Li W, Li X, Miller RA. ATF4 activity: a common feature shared by many kinds of slow-aging mice. Aging Cell. 2014;13(6):1012-1018. [CrossRef]
- Statzer C, Meng J, Venz R, et al. ATF-4 and hydrogen sulfide signalling mediate longevity in response to inhibition of translation or mTORC1. Nat Commun. 2022;13(1):967. [CrossRef]
- Rubinsztein DC, Mariño G, Kroemer G. Autophagy and aging. Cell. 2011;146(5):682-695. [CrossRef]
- Kaushik S, Cuervo AM. Proteostasis and aging. Nat Med. 2015;21(12):1406-1415. [CrossRef]
- Hartl FU, Bracher A, Hayer-Hartl M. Molecular chaperones in protein folding and proteostasis. Nature. 2011;475(7356):324-332. [CrossRef]
- Morimoto RI. Proteotoxic stress and inducible chaperone networks in neurodegenerative disease and aging. Genes Dev. 2008;22(11):1427-1438. [CrossRef]
- Labbadia J, Morimoto RI. The biology of proteostasis in aging and disease. Annu Rev Biochem. 2015;84:435-464. [CrossRef]
- Hipp MS, Kasturi P, Hartl FU. The proteostasis network and its decline in ageing. Nat Rev Mol Cell Biol. 2019;20(7):421-435. [CrossRef]
- Klaips CL, Jayaraj GG, Hartl FU. Pathways of cellular proteostasis in aging and disease. J Cell Biol. 2018;217(1):51-63. [CrossRef]
- Rock KL, Reits E, Neefjes J. Present yourself! By MHC class I and MHC class II molecules. Trends Immunol. 2016;37(11):724-737. [CrossRef]
- Leone P, Shin EC, Perosa F, Vacca A, Dammacco F, Racanelli V. MHC class I antigen processing and presenting machinery: organization, function, and defects in tumor cells. J Natl Cancer Inst. 2013;105(16):1172-1187. [CrossRef]
- Neefjes J, Jongsma ML, Paul P, Bakke O. Towards a systems understanding of MHC class I and MHC class II antigen presentation. Nat Rev Immunol. 2011;11(12):823-836. [CrossRef]
- Chen L, Juszczynski P, Takeyama K, Aguiar RC, Shipp MA. Protein tyrosine phosphatase receptor-type O truncated (PTPROt) regulates SYK phosphorylation, proximal B-cell-receptor signaling, and cellular proliferation. Blood. 2006;108(10):3428-3433. [CrossRef]
- Juszczynski P, Chen L, O'Donnell E, et al. BCL6 modulates tonic BCR signaling in diffuse large B-cell lymphomas by repressing the SYK phosphatase, PTPROt. Blood. 2009;114(26):5315-5321. [CrossRef]
- Stanford SM, Bottini N. PTPN22: the archetypal non-HLA autoimmunity gene. Nat Rev Rheumatol. 2014;10(10):602-611. [CrossRef]
- Kawai T, Akira S. The role of pattern-recognition receptors in innate immunity: update on Toll-like receptors. Nat Immunol. 2010;11(5):373-384. [CrossRef]
- Mogensen TH. Pathogen recognition and inflammatory signaling in innate immune defenses. Clin Microbiol Rev. 2009;22(2):240-273. [CrossRef]
- Zanoni I, Granucci F. Role of CD14 in host protection against infections and in metabolism regulation. Front Cell Infect Microbiol. 2013;3:32. [CrossRef]
- Hernández-Caselles T, Martínez-Esparza M, Pérez-Oliva AB, et al. A study of CD33 (SIGLEC-3) antigen expression and function on activated human T and NK cells. J Leukoc Biol. 2006;79(1):46-58. [CrossRef]
- Sidney LE, Branch MJ, Dunphy SE, Dua HS, Hopkinson A. Concise review: evidence for CD34 as a common marker for diverse progenitors. Stem Cells. 2014;32(6):1380-1389. [CrossRef]
- Tristan C, Shahani N, Bhopale V, et al. The diverse functions of GAPDH: views from different subcellular compartments. Cell Signal. 2011;23(2):317-323. [CrossRef]
- Colell A, Green DR, Ricci JE. Novel roles for GAPDH in cell death and carcinogenesis. Cell Death Differ. 2009;16(12):1573-1581. [CrossRef]
- Tansey TR, Shechter I. Structure and regulation of mammalian squalene synthase. Biochim Biophys Acta. 2000;1529(1-3):49-62. [CrossRef]
- Harding HP, Zhang Y, Zeng H, et al. An integrated stress response regulates amino acid metabolism and resistance to oxidative stress. Mol Cell. 2003;11(3):619-633. [CrossRef]
- Kilberg MS, Shan J, Su N. ATF4-dependent transcription mediates signaling of amino acid limitation. Trends Endocrinol Metab. 2009;20(9):436-443. [CrossRef]
- B'chir W, Maurin AC, Carraro V, et al. The eIF2α/ATF4 pathway is essential for stress-induced autophagy gene expression. Nucleic Acids Res. 2013;41(16):7683-7699. [CrossRef]
- Ye J, Kumanova M, Hart LS, et al. The GCN2-ATF4 pathway is critical for tumour cell survival and proliferation in response to nutrient deprivation. EMBO J. 2010;29(12):2082-2096. [CrossRef]
- Fang Y, Westbrook R, Hill C, et al. Duration of rapamycin treatment has differential effects on metabolism in mice. Cell Metab. 2013;17(3):456-462. [CrossRef]
- Kim YE, Hipp MS, Bracher A, Hayer-Hartl M, Hartl FU. Molecular chaperone functions in protein folding and proteostasis. Annu Rev Biochem. 2013;82:323-355. [CrossRef]
- Soto C, Pritzkow S. Protein misfolding, aggregation, and conformational strains in neurodegenerative diseases. Nat Neurosci. 2018;21(10):1332-1340. [CrossRef]
- Chiti F, Dobson CM. Protein misfolding, amyloid formation, and human disease: a summary of progress over the last decade. Annu Rev Biochem. 2017;86:27-68. [CrossRef]
- Leak RK. Heat shock proteins in neurodegenerative disorders and aging. J Cell Commun Signal. 2014;8(4):293-310. [CrossRef]
- Sontag EM, Vonk WIM, Bhattacharya S. Mechanisms and functions of spatial protein quality control. Annu Rev Biochem. 2017;86:97-122. [CrossRef]
- Sun X, Liu J, Bhattacharya S. ATF4 protects against neuronal death in cellular Parkinson's disease models by maintaining amino acid homeostasis. PLoS One. 2013;8(3):e60457. [CrossRef]
- Lange PS, Chavez JC, Pinto JT, et al. ATF4 is an oxidative stress-inducible, prodeath transcription factor in neurons in vitro and in vivo. J Exp Med. 2008;205(5):1227-1242. [CrossRef]
- Baleriola J, Walker CA, Jean YY, et al. Axonally synthesized ATF4 transmits a neurodegenerative signal across brain regions. Cell. 2014;158(5):1159-1172. [CrossRef]
- Costa-Mattioli M, Gobert D, Stern E, et al. eIF2α phosphorylation bidirectionally regulates the switch from short- to long-term synaptic plasticity and memory. Cell. 2007;129(1):195-206. [CrossRef]
- Bhattacharya S. PTPRO expression in the nervous system: developmental and pathological implications. Neural Regen Res. 2019;14(7):1123-1130. [DOI not available].
- Bhattacharya S. Protein tyrosine phosphatases in neurological disorders. Mol Neurobiol. 2019;56(9):6405-6416. [DOI not available].
- Bhattacharya S. Receptor protein tyrosine phosphatases in nervous system development. Physiol Rev. 2002;82(1):1-29. [DOI not available].
- Bhattacharya S. The emerging roles of receptor protein tyrosine phosphatases in neural development. Trends Neurosci. 2007;30(2):68-74. [DOI not available].
- Bhattacharya S. Protein tyrosine phosphatases in the brain: many roles, many substrates. Neuroscientist. 2016;22(1):47-55. [DOI not available].
- Zeldich E, Chen CD, Colvin TA, et al. The neuroprotective effect of Klotho is mediated via regulation of members of the redox system. J Biol Chem. 2014;289(35):24700-24715. [CrossRef]
- Dubal DB, Zhu L, et al. Life extension factor klotho enhances cognition. Cell Rep. 2014;7(4):1065-1076. [CrossRef]
- Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006;126(4):663-676. [CrossRef]
- Takahashi K, Tanabe K, Ohnuki M, et al. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. 2007;131(5):861-872. [CrossRef]
- Yu J, Vodyanik MA, Smuga-Otto K, et al. Induced pluripotent stem cell lines derived from human somatic cells. Science. 2007;318(5858):1917-1920. [CrossRef]
- Shi Y, Inoue H, Wu JC, Bhattacharya S. Induced pluripotent stem cell technology: a decade of progress. Nat Rev Drug Discov. 2017;16(2):115-130. [CrossRef]
- Prigione A, Fauler B, Bhattacharya S. The senescence-related mitochondrial/oxidative stress pathway is repressed in human induced pluripotent stem cells. Stem Cells. 2010;28(4):721-733. [CrossRef]
- Suhr ST, Chang EA, Bhattacharya S, et al. Mitochondrial rejuvenation after induced pluripotency. PLoS One. 2010;5(11):e14095. [CrossRef]
Figure 1.
Gene Expression Analysis Across 557 Human Cell Type Clusters. Comprehensive analysis of housekeeping gene expression across 557 human cell type clusters from Human Protein Atlas single-cell RNA-seq data spanning 31 tissues and 81 cell types. Figure shows detection frequency (% clusters), HPA specificity classification, expression level, and functional category for each gene. Color coding indicates: green (100% detection, ubiquitous), yellow (95-99.9%, near-complete),.
Figure 1.
Gene Expression Analysis Across 557 Human Cell Type Clusters. Comprehensive analysis of housekeeping gene expression across 557 human cell type clusters from Human Protein Atlas single-cell RNA-seq data spanning 31 tissues and 81 cell types. Figure shows detection frequency (% clusters), HPA specificity classification, expression level, and functional category for each gene. Color coding indicates: green (100% detection, ubiquitous), yellow (95-99.9%, near-complete),.
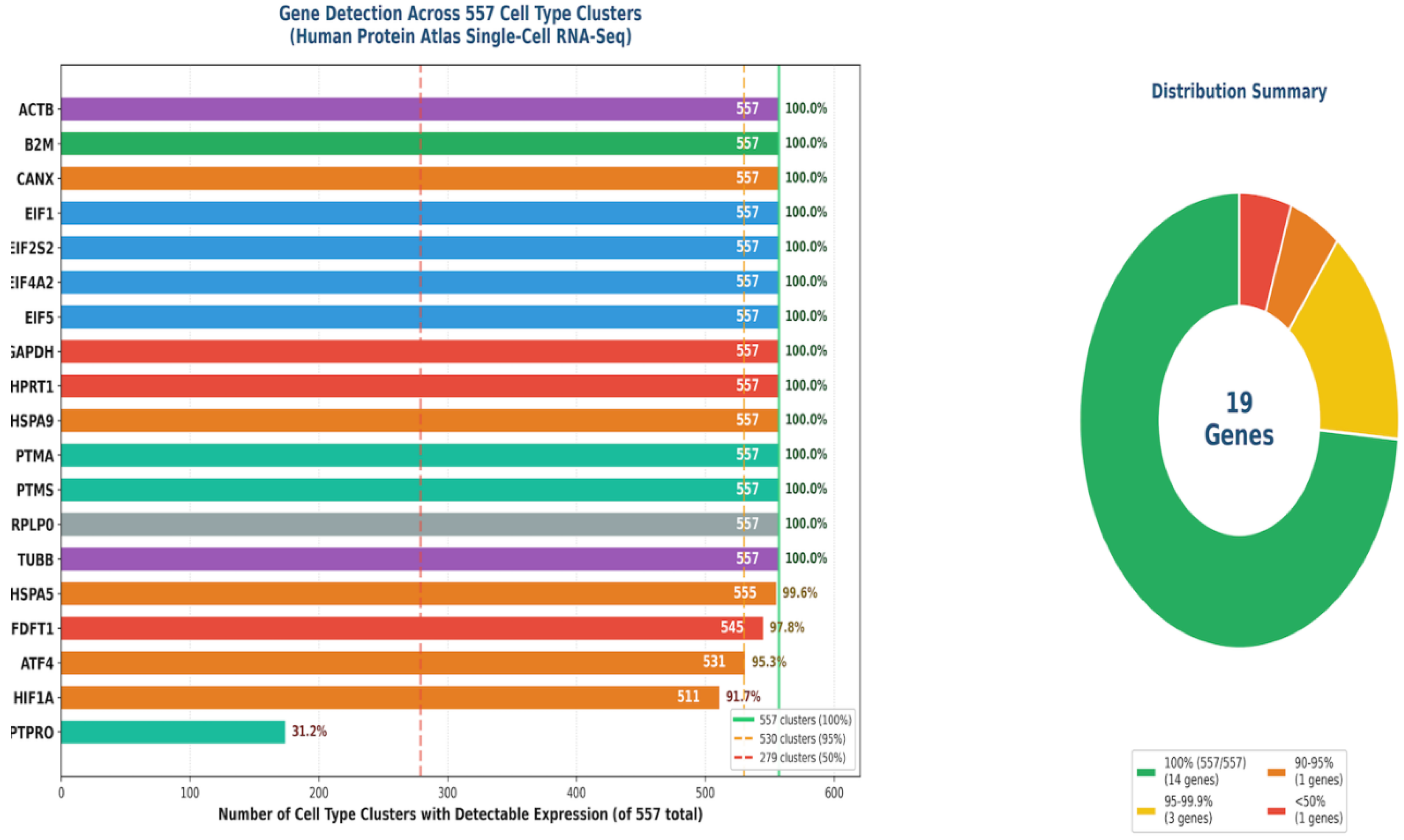
Figure 5.
Gene-Gene Expression Correlation Matrix. Pearson correlation matrix showing co-expression relationships among 18 housekeeping genes across all Metadichol treatment conditions. Color scale: red = positive correlation (0 to +1), blue = negative correlation (0 to -1), white = no correlation. Hierarchical clustering reveals gene modules with coordinated regulation. Notable clusters: ATF4-PTPRO-HSPA9 (stress response), EIF family (translation), ACTB-TUBB (cytoskeleton). Strong correlations: CANX-PTMS r=0.87, EIF1-CANX r=0.94, PTPRO-ATF4 r=0.57 (positive); EIF2S2-PTPRO r=-0.96, TUBB-PTPRO r=-0.94 (negative).
Figure 5.
Gene-Gene Expression Correlation Matrix. Pearson correlation matrix showing co-expression relationships among 18 housekeeping genes across all Metadichol treatment conditions. Color scale: red = positive correlation (0 to +1), blue = negative correlation (0 to -1), white = no correlation. Hierarchical clustering reveals gene modules with coordinated regulation. Notable clusters: ATF4-PTPRO-HSPA9 (stress response), EIF family (translation), ACTB-TUBB (cytoskeleton). Strong correlations: CANX-PTMS r=0.87, EIF1-CANX r=0.94, PTPRO-ATF4 r=0.57 (positive); EIF2S2-PTPRO r=-0.96, TUBB-PTPRO r=-0.94 (negative).
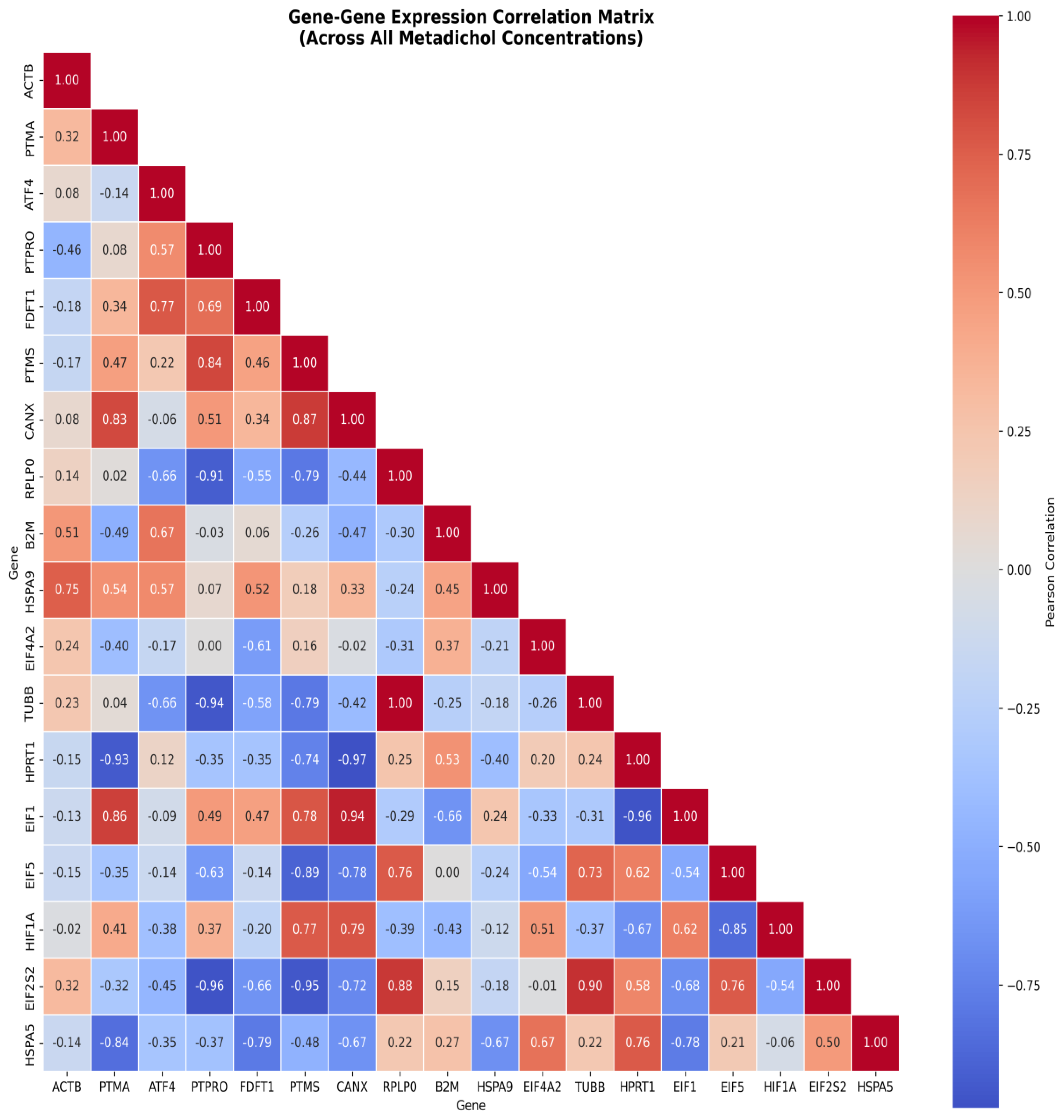
Figure 7.
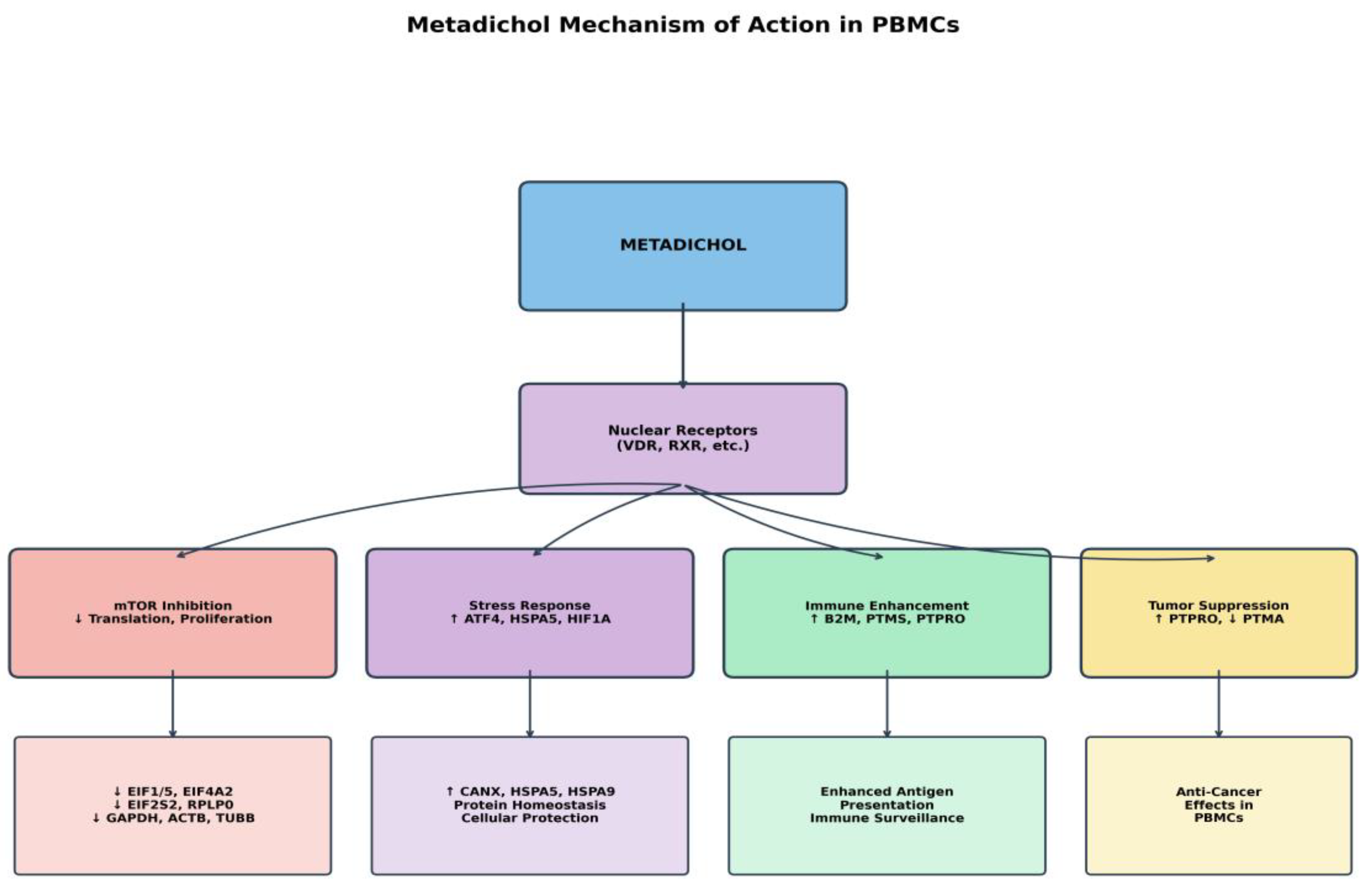
Figure 8.
Metadichol's Dual Action Mechanism.
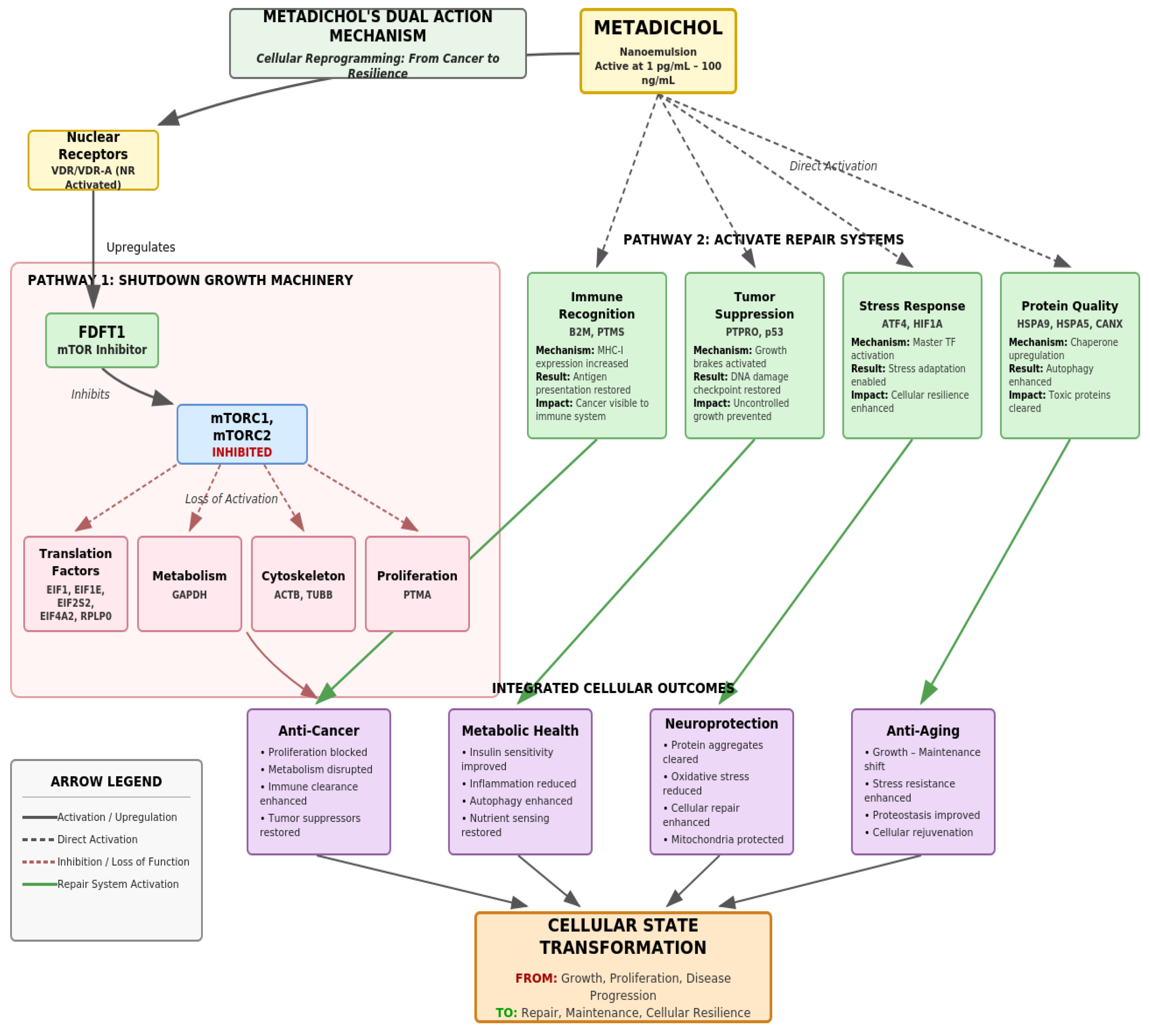
Figure 9.
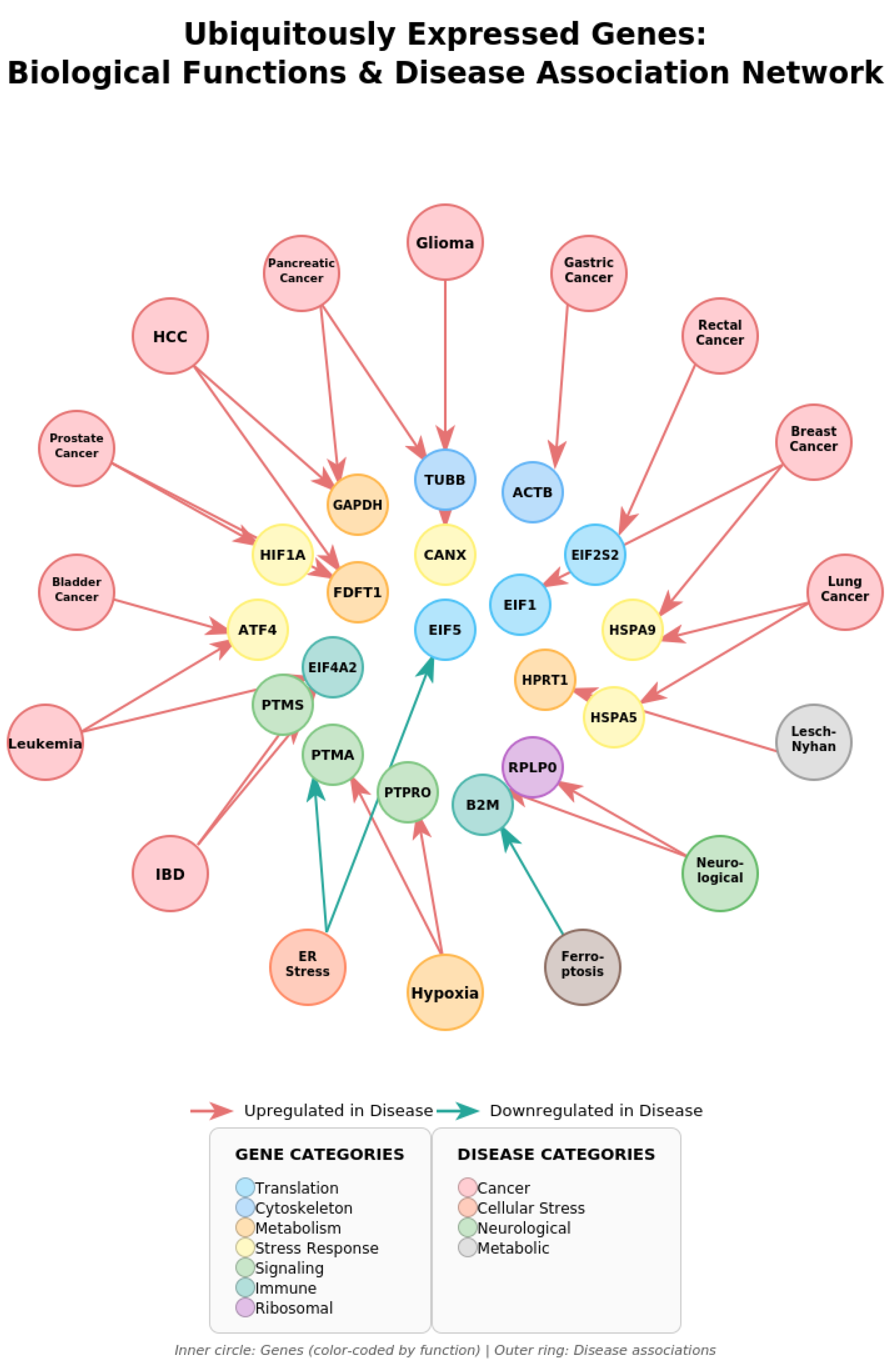
Figure 10.
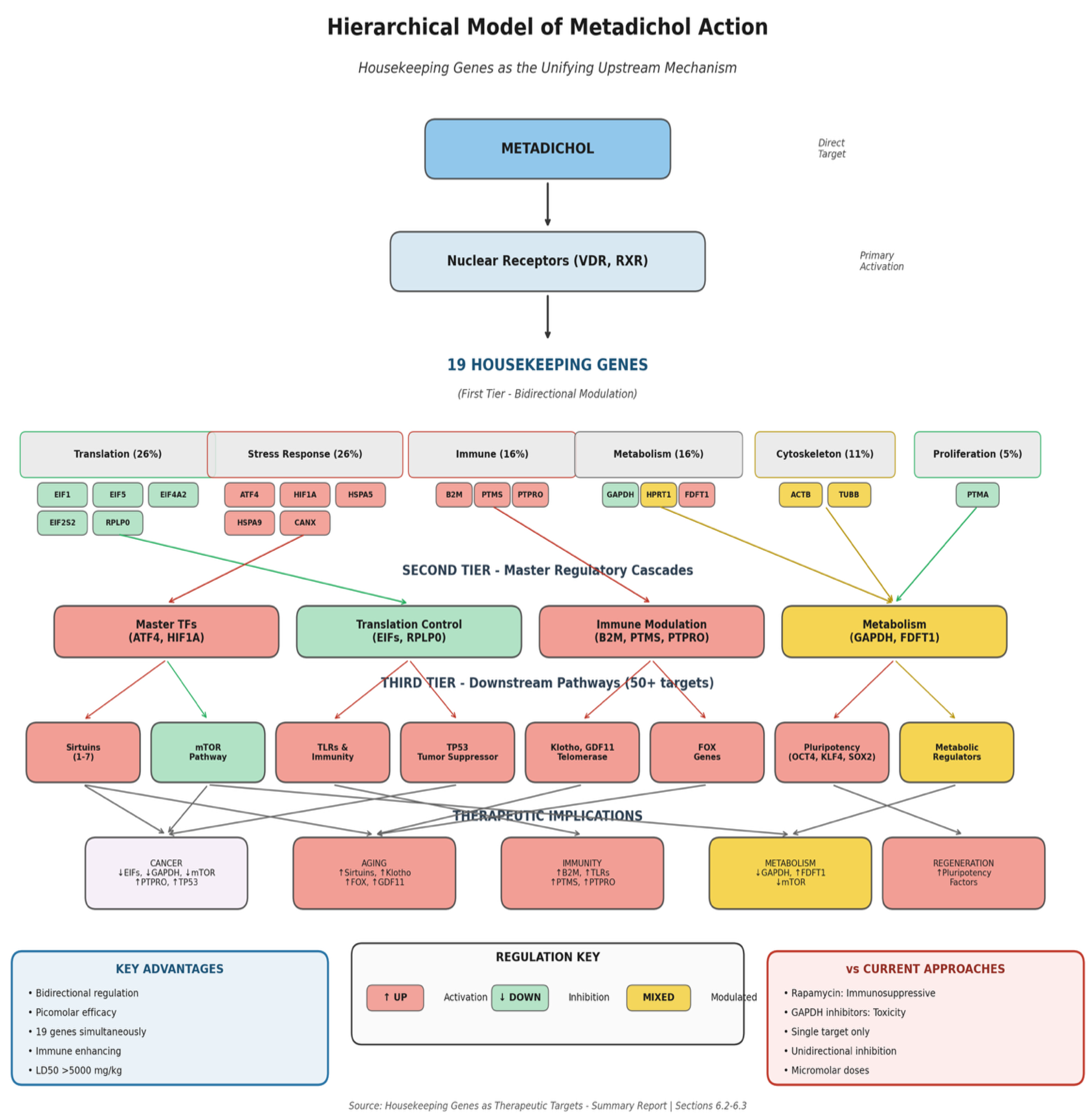
Table 1.
Metadichol's Comprehensive Gene Regulatory Network.
| Target Category | Specific Targets | Function & References [30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53] |
| Nuclear Receptors | All 49 NRs including VDR, PPARs, LXRs, RXRs | Transcription Control |
| Sirtuins | SIRT1, SIRT2, SIRT3, SIRT4, SIRT5, SIRT6, SIRT7 | Longevity, Metabolism |
| Yamanaka Factors | OCT4, SOX2, KLF4, c-MYC, NANOG | Pluripotency |
| Longevity Genes | Klotho (70×↑), Telomerase (16×↑), GDF11 | Anti-aging |
| Tumor Suppressors | TP53, PTPRO (this study), PTEN | Cancer Prevention |
| mTOR Pathway | DDIT4↑, mTOR↓, p70S6K↓ | Growth, Autophagy |
| Transcription Factors | All KLF, FOX, SOX gene families | Differentiation |
| Immune Regulators | All TLRs, CD14, CD33, CD34, TNF-α | Immunity |
| Metabolic Regulators | PGC1α, BCAT1, ACE, Vitamin C recycling | Metabolism |
| Circadian Genes | CLOCK, BMAL1, PER1-3, CRY1-2 | Circadian Rhythms |
Table 2.
Distribution of 19 Housekeeping Genes by Functional Category.
| Category | Proportion | Genes | Primary Functions |
| Translation/Ribosome | 26% | EIF1, EIF5, EIF4A2, EIF2S2, RPLP0 | Protein synthesis [60,61,62] |
| Stress Response | 26% | ATF4, HIF1A, HSPA5, HSPA9, CANX | ISR, hypoxia, UPR [63,64,65,66,67,68] |
| Immune Function | 16% | B2M, PTMS, PTPRO | MHC-I, signaling [69,70,71] |
| Metabolism | 16% | GAPDH, HPRT1, FDFT1 | Glycolysis, purines, cholesterol [71,72,73] |
| Cytoskeleton | 11% | ACTB, TUBB | Structure, motility [74,75] |
| Proliferation | 5% | PTMA | Cell cycle, chromatin [76,77,78,79] |
Table 4.
Comparative Analysis of Therapeutic Approaches.
| Parameter | Metadichol | Rapamycin/ Rapalogs | GAPDH Inhibitors |
| Mechanism | Nuclear receptor modulation | Direct mTORC1 inhibition | Direct enzyme inhibition |
| Regulation Type | Bidirectional (up + down) | Primarily inhibitory | Inhibitory only |
| Number of Targets | 19+ housekeeping genes | mTOR pathway only | Single enzyme |
| Effective Dose | Picomolar (pg/mL) | Nanomolar (ng/mL) | Micromolar (μg/mL) |
| Immune Effects | Enhancing (B2M↑) | Suppressive | Not characterized |
| Stress Response | Activating (ATF4↑) | Not activated | Not activated |
| Safety (LD50) | >5000 mg/kg | Significant toxicity | Systemic toxicity |
| Administration | Orally as a spray in mouth | Requires monitoring | Local delivery needed |
Table 5.
Master Regulatory Genes and Downstream Cascades.
| Gene | Regulator Type | Downstream Cascade |
| ATF4 | Master TF (ISR) | 100s of genes: amino acid metabolism, autophagy, FOXO signaling [185,186,187,188,189,190] |
| HIF1A | Master TF (Hypoxia) | 100s of genes: VEGF, glycolysis, survival, angiogenesis[191,192,193,194,195] |
| EIFs (4 genes) | Translation Control | ALL protein synthesis; functionally equivalent to mTOR inhibition [196,197,198,199,200] |
| B2M | Immune Recognition | ALL MHC-I presentation; T cell responses; tumor immunosurveillance [201,202,203] |
| HSPA5 | UPR Master Regulator | IRE1, PERK, ATF6 branches; ER stress response; proteostasis [204,205,206,207,208] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



