
Submitted:
28 January 2026
Posted:
29 January 2026
You are already at the latest version
Abstract
The protozoan pathogen Toxoplasma gondii is responsible for toxoplasmosis a disease that can be deadly in immunocompromised patients and the developing fetus during pregnancy. Current treatments are widely considered to be suboptimal. We have recently reported that 5-fluoropyrimidines have highly promising anti-toxoplasmosis effects and are taken up by the parasite by a high affinity uracil/uridine transporter, TgUUT1. Here, we attempt to identify the gene encoding this transport protein. The only nucleoside or nucleobase family identified in the T. gondii genome was the Equilibrative Nucleoside Transporter (ENT) family, with four members. Of these, TgAT1 is known to be purine-specific, and deletion of the TgENT2 and TgENT3 genes, either separately or jointly, did not affect uridine transport or sensitivity to 5-fluoropyrimidines. In contrast, depletion of TgENT1, an essential gene, resulted in significant reduction in the uptake of both uracil and uridine but not of the amino acid tryptophan. Moreover, expression of TgENT1 in a Leishmania mexicana cell line with low endogenous uracil uptake rates significantly increased uracil uptake for these cells. We conclude that it is highly probable that TgENT1 encodes the T. gondii uracil/uridine transporter.
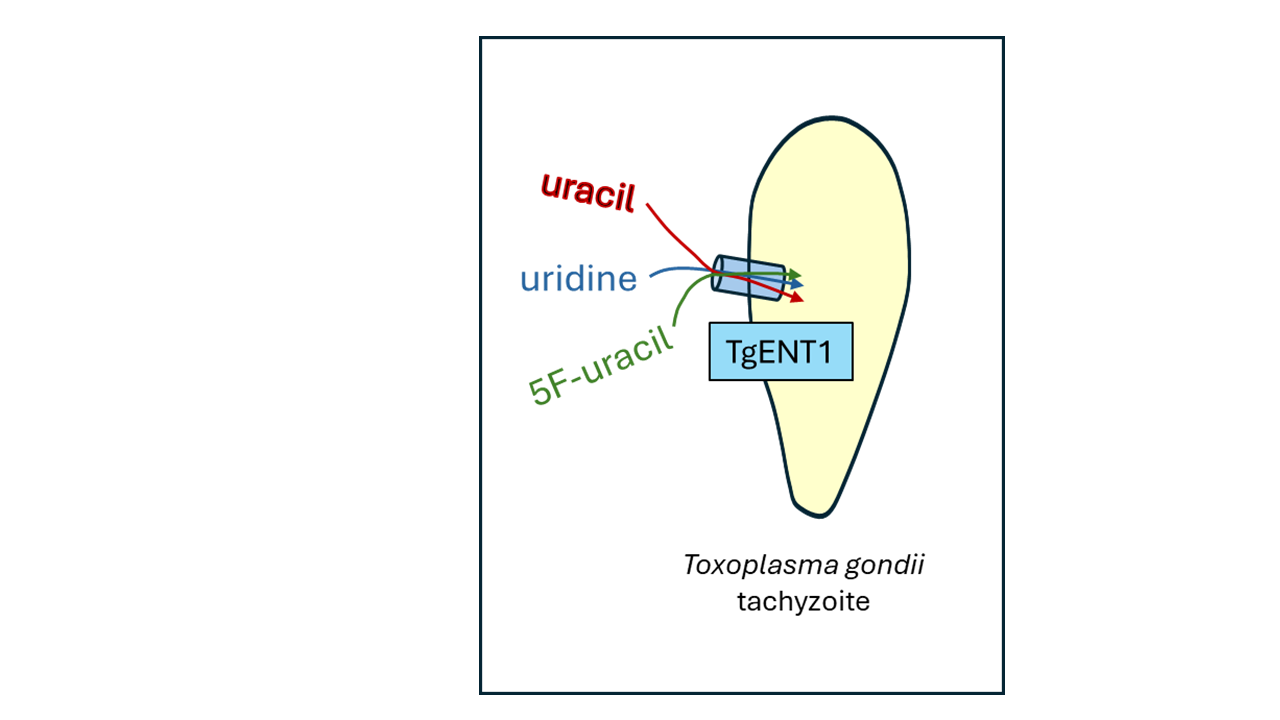
Keywords:
Toxoplasma gondii
; nucleoside transporter
; uridine transport
; 5-fluorouracil
; pyrimidine antimetabolite
1. Introduction
Toxoplasmosis is a common infection of humans and mammals that can be dangerous in its acute form, especially if contracted during pregnancy, as the pathology to the unborn baby can be severe and even fatal [1]. Moreover, chronic infection is thought to persist for life and poses a severe risk to immunocompromised patients [2]. Despite the clinical importance of toxoplasmosis there are still numerous gaps in our knowledge of the biology of its etiological agent, Toxoplasma gondii.
T. gondii is an apicomplexan parasite and, like other Apicomplexa, including the Plasmodium parasites that cause malaria, it is an obligate intracellular parasite [3]. In its tachyzoite form, T. gondii is able to invade a broad range of nucleated cell types of almost any warm-blooded animal [4]. Despite its critical dependency on the host for survival our understanding of how the parasite obtains essential nutrients such as nucleotides remains poor.
Like all other parasites, T. gondii express highly efficient salvage systems to obtain nutrients from the host environment. This involves the manipulation of nutrient uptake by the host cell [5,6]. Pores in the parasitophorous vacuole membranes allow the diffusion of these nutrients directly to the parasite [7,8], where they are internalized by an array of highly effective nutrient transporters that typically outcompete host transporters in substrate affinity and translocation efficiency.
Toxoplasma tachyzoites, like many other parasites, also have high replication rates, requiring large amounts of nucleotides for DNA and RNA synthesis, among other functions. Like all other protozoan parasites where this has been investigated, Toxoplasma lack the biosynthesis pathway for purines [9] and thus rely fully on transporters and salvage enzymes for these critical biochemicals. They cannot take up nucleotides like ATP directly from the host cytosol [10] and thus need particularly high affinity transporters to salvage free nucleosides and nucleobases, which are present in low concentrations in the host environment. High affinity transporters for purine nucleosides and oxopurine nucleobases have been described [11].
In contrast, T. gondii express the entire pathway for de novo pyrimidine synthesis and although this is important for virulence, it is not essential for cellular invasion in vitro if the media is supplemented with >20 µM uracil [12]. This constitutes functional evidence that T. gondii expresses a transporter capable of uracil salvage, and we recently confirmed that uracil and also uridine are indeed readily taken up by T. gondii tachyzoites [13], but that its capacity is insufficient to sustain in vivo replication.
In recent years, numerous nucleoside and nucleobase transporters of protozoan parasites have been cloned and characterized [14,15,16,17] and all were found to be members of the Equilibrative Nucleoside Transporter (ENT) family. Whereas members of the Concentrative Nucleoside Transporter (CNT) family are commonly found in mammalian hosts and many other species [18], none have been found in protozoan genomes. Despite this progress in the characterization of nucleoside and nucleobase transporters, particularly from kinetoplastids and Plasmodium species, our understanding of purine and pyrimidine salvage in Toxoplasma remains very incomplete.
Apart from their importance in the physiology of protozoan pathogens, transporters have potential as drug targets [19,20]. For Toxoplasma, this potential is strongly supported by our recent report showing that the 5-fluoropyrimidines 5-fluorouracil (5-FU), 5-fluorouridine (5F-Urd) and 5-fluoro,2’deoxyuridine (5F,2’dUrd) are efficiently taken up by T. gondii tachyzoites and display 10-fold higher activity against the intracellular parasites than the antifolate sulfadiazine, the ‘current gold standard treatment’ when combined with pyrimethamine [21]. In a first pilot experiment, 5-FU was at least as effective as sulfadiazine in a mouse model of acute toxoplasmosis [13]. Moreover, the 5-fluoropyrimidines have long been used as anticancer chemotherapy [22] and, according to the one report of their use against acute toxoplasmosis in AIDS patients in combination with the second-line toxoplasmosis drug clindamycin [23], would be safe to use as they are effective at one tenth the dose used against cancer.
Although our previous report identified a new Uracil/Uridine Transporter (TgUUT1), as responsible for uptake of the 5-fluoropyrimidines, the gene encoding this carrier has not yet been identified. Indeed, although the T. gondii genome project has long since identified four likely nucleoside or nucleobase transporters of the ENT family (Table 1), only one of the four genes and its product has been characterized: Tg_244440, which was first identified from a strain made resistant to the adenosine analogue adenine arabinoside (Ara-A), as a low affinity adenosine transporter, and the corresponding protein was therefore named TgAT1 [24]. Recently, we confirmed, by heterologous expression in Trypanosoma brucei, that TgAT1 is indeed a low affinity transporter for adenosine but, more surprisingly, it also displayed much higher affinity for the oxopurine nucleobases hypoxanthine and guanine, and their nucleosides inosine and guanosine [25]. Of the three remaining genes encoding putative ENT family members (here collectively called TgENTs), Tg_288540 is reportedly the only essential gene for T. gondii in culture [26] and Tg_233130 is upregulated in bradyzoites [27].
Here, we start to address the issues raised above, including the role of the TgENTs in pyrimidine uptake, to try to identify the gene encoding TgUUT1, being the carrier for the 5-fluoropyrimidine antimetabolites.
2. Materials and Methods
2.1. In Vitro Culture of Host Cells and Parasites
2.1.1. T. gondii in Human Fibroblasts
The F3 tomato strain (RH Δku80 TATi) [13,28], herein referred to as RH, was used as the primary cell line for generation of all the mutant cell lines and also used as control for all the biochemical experiments such as transport assays and drug screening assays, qRT-PCR and plaque assays.
T. gondii tachyzoites of the RH and mutant lines were cultured in human foreskin fibroblasts (HFF), sourced from ATCC (SCRC-1041). Parasites were passaged routinely as confluent monolayer of HFF in Dulbecco’s Modified Eagle’s Medium (DMEM) containing 4.5 gL-1 glucose, supplemented with 10% (v/v) fetal bovine serum, 4 mM L-glutamine and penicillin/streptomycin antibiotics and grown at 37 °C with 5% CO2. Where mentioned anhydrotetracycline (ATc) was added to the medium at a final concertation of 0.5 µM.
2.1.2. Leishmania Promastigotes
Three strains of L. mexicana promastigote forms were mainly used in this part of the project: 1) L. mexicana-Cas9 T7 strain (derived from L. mexicana WT promastigotes by expression of the Streptococcus pyogenes Cas9 nuclease gene ‘Cas9’ and maintained on 32 µg/mL hygromycin [29], generously donated by Prof. Eva Gluenz (University of Bern, Switzerland). 2) The Lmex-NT3-KO strain (∆NT3) was generated from the L. mexicana Cas9 strain by CRISPR-mediated deletion of the NT3 transporter gene [30,31]. 3) The Lmex-NT3-KO strain was used as expression system to study TgENT1. All Leishmania strains were grown as promastigotes, in standard HOMEM (GIBCO, Life Technologies, Paisley, UK) supplemented with 10% heat-inactivated fetal bovine serum (FBS; PAA Laboratories, Linz, Austria) and 1% of a penicillin–streptomycin solution (Life Technologies) at 25 °C, as described by Al-Salabi, et al. [32].
2.2. Chemicals and Radiochemicals
Uridine, uracil, thymidine, cytidine, adenosine, Inosine, 5-floururacil (5-FU), 5-flourouridine (5-FUrd), sulfadiazine, resazurin sodium salt and phenylarsine oxide (PAO) were sourced from Sigma-Aldrich (Poole, UK). 5-Flouro 2’deoxyuridine (5-F-2’dUrd) was from Avantor (VWR). [2,8-3H]-Adenine (40.3 Ci/mmol) was obtained from PerkinElmer (Waltham, MA, USA). [5,6-3H]-Uracil (40 Ci/mmol), [5,6-3H]-Uridine (60 Ci/mmol), [2,8-3H]-Adenosine (40 Ci/mmol) and [3H]-Tryptophan (25 Ci/mmol) were sourced from American Radiolabeled Chemicals Incorporated (St Louis, MO, USA).
2.3. Drug Sensitivity Assay for T. gondii Tachyzoites
Drug sensitivity assays for Toxoplasma gondii tachyzoites were performed as previously described [13]. Briefly, HFF cells were seeded in 96-well black plates and grown to confluence. Test compounds and sulfadiazine (positive control) were prepared in DMEM and serially diluted across the plate, leaving the last column as a drug-free control. Freshly egressed tachyzoites (~1000/well) were added to each well and incubated for 6 days at 37 °C with 5% CO₂. Fluorescence was measured using a PHERAstar plate reader (excitation: 540 nm; emission: 590 nm). EC₅₀ values were calculated using GraphPad Prism 10.0 using a 4-parameter sigmoid curve (variable slope). All assays were performed in triplicate and repeated independently 3–5 times.
2.4. Drug Cytotoxicity Assay for HFF Cells Using Alamar Blue Dye
Cytotoxicity was as previously described [13], using PAO as positive control. Plates were incubated for 6 days at 37 °C with 5% CO₂. On day 6, 10 µL resazurin solution (12.5 mg/100 mL ddH₂O) was added to each well, including media-only wells for background fluorescence. After 3–4 h incubation, fluorescence was measured (excitation: 540 nm; emission: 590 nm) using a PHERAstar plate reader. Data were analyzed in GraphPad Prism 10.0 using a four-parameter sigmoid curve to calculate EC₅₀ values. Experiments were performed in triplicate and repeated 3–5 times.
2.5. Transport Assays
Transport of radiolabeled uridine, uracil and tryptophan into extracellular T. gondii tachyzoites and L. mexicana promastigotes was assayed following previously published protocols [11,13,16,31,33]. Briefly, L. mexicana promastigotes were grown at 27 °C for 40–48 h to mid-log phase, while T. gondii tachyzoites were harvested from confluent HFF cultures maintained at 37 °C and 5% CO₂. Parasites were washed twice in assay buffer (AB: 33 mM HEPES, 98 mM NaCl, 4.6 mM KCl, 0.5 mM CaCl₂, 0.07 mM MgSO₄, 5.8 mM NaH₂PO₄, 0.03 mM MgCl₂, 23 mM NaHCO₃, 14 mM D-glucose; pH 7.3), counted, and resuspended at 1 × 10⁸ cells/mL (Leishmania) or 2 × 10⁸ cells/mL (T. gondii). After 30 min of recovery at room temperature, 100 µL of cell suspension was layered over an oil mixture (1:7 mineral oil:di-n-butyl phthalate for L. mexicana, 1:5 for T. gondii) containing radiolabeled substrate. Incubations were performed for predetermined times and stopped by adding 750 µL of ice-cold stop solution (0.5–2 mM unlabeled substrate). Tubes were centrifuged at 14,800 × g for 1 min, flash-frozen, and cell pellets were processed for scintillation counting after SDS lysis. All assays were done in triplicate and repeated in three independent experiments.
2.6. Plasmid Construction and Expression of TgENT1 in L. mexicana NT3-KO
The TgENT1 gene was expressed in L. mexicana NT3-KO promastigotes (Aldfer et al., 2022b) using the pNUS-HcN plasmid, which carries a neomycin resistance marker (G-418) [34]. The pNUS-HcN vector was digested with NdeI and XhoI according to the manufacturer’s protocol. TgENT1 was amplified by PCR using Phusion High-Fidelity DNA Polymerase (supplemental Tables S1 and S2). PCR products and digested plasmids were run on 1% agarose gels, visualized under UV light, and purified using the NucleoSpin PCR and Gel Extraction Kit (Macherey-Nagel).
Ligation was performed using the NEBuilder HiFi DNA Assembly Kit (New England Biolabs). The reaction (10 µL total volume) contained 60 ng of digested pNUS-HcN vector and 180 ng of TgENT1 insert (vector : insert ratio 1:3), combined with 5 µL of NEBuilder HiFi Master Mix and nuclease-free water. The mixture was incubated at 50 °C for 3–4 h. The assembled plasmid was transformed into NEB 5-alpha competent E. coli by heat shock. Colonies were selected on LB agar containing 100 µg/mL ampicillin, cultured in LB broth, and plasmid DNA was extracted using the NucleoSpin Plasmid Purification Kit.
Positive clones were screened by Phusion PCR using a TgENT1-specific forward primer (HDK-1739) and a pNUS-HcN reverse primer (HDK-340) (listed in Table S3). Confirmation was also achieved by restriction digestion with NdeI and XhoI, followed by Sanger sequencing (Source Bioscience, Livingston, UK) using (M13F) and gene-specific reverse primers (HDK-1740) (listed in Table S3). Sequence data were analyzed using CLC Genomics Workbench v7.0 (Qiagen). Verified plasmids (pHDK295 ‘TgENT1’) were ethanol-precipitated, and 25 µg of DNA was resuspended in 15 µL sterile water.
Transfection was performed as described [31]. Briefly, 5 × 107 L. mexicana NT3-KO promastigotes were harvested, washed twice in PBS, and resuspended in 100 µL transfection buffer. Cells were mixed with 10 µg of circular pHDK295 plasmid DNA (episomal expression) or RNase-free water (negative control), transferred to a 0.2 cm cuvette, and electroporated using the Amaxa Nucleofector (Program U-033). Cells were recovered overnight in HOMEM medium supplemented with 10% FBS at 25 °C, followed by selection with 50 µg/mL G-418. Cultures were plated by limiting dilution (1:10, 1:25, 1:100) in 96-well plates and incubated at 25 °C for 10–14 days to obtain individual clones. Positive clones were expanded in HOMEM medium containing G-418. Genomic DNA was extracted using the NucleoSpin Tissue Kit (Macherey-Nagel), and integration was confirmed by PCR using TgENT1-specific forward (HDK-1739) and pNUS-HcN reverse primers (HDK-340).
2.7. CRISPR-Mediated Gene Disruption in T. gondii Tachyzoites
2.7.1. Direct Gene Knockout
Plasmids for gene knockout and knockdown were constructed using CRISPR-guided promoter replacement in T. gondii RH tomato ΔKu80 TATi cells (herein called RH cell line) as described previously [28,35]. Three plasmids were used for these strategies: (i) pDTS4-DHFR for non-essential gene replacement or promoter replacement for essential genes [28]; (ii) an mNeonGreen cassette plasmid for homologous recombination knockout [36]; and (iii) pg474 (Tub-Cas9-YFP-pU6-ccdB-tracrRNA) for delivery of sgRNA/Cas9 [35]. The pDTS4-DHFR plasmid was used to generate two single knockout strains, of TgENT2 and TgENT3, and knockdown of TgENT1, while the mNeonGreen cassette was used to generate a double knockout (targeting TgENT2 in a ΔTgENT3 background) and ΔTgAT1 in the RH strain.
Single guide RNAs targeting the start codon region of TgAT1, TgENT1, TgENT2, and TgENT3 were designed using the ChopChop online website tool (https://chopchop.cbu.uib.no/) and are listed in (Supplemental Table S4). Each sgRNA was cloned into pg474 via BsaI restriction sites. Plasmids were purified using Qiagen Miniprep and Midiprep kits according to the manufacturer’s instructions. The DHFR cassette allowed selection for pyrimethamine resistance [28], while mNeonGreen-positive parasites were selected by fluorescence-activated cell sorting (FACS) . PCR primers for cassette amplification and diagnostic PCR are listed in (Supplemental Table S5). Homology regions for DHFR cassettes were 50 bp at the 5′ and 3′ UTRs of the target gene, while mNeonGreen cassettes used 40 bp homology regions. PCR products were confirmed by agarose gel electrophoresis and purified using QIAquick PCR Purification kit (Qiagen) prior to transfection.
2.7.2. Gene Knockdown (Tetracycline-Inducible Transactivator System)
Gene knockdown constructs were generated using the same CRISPR/Cas9 workflow described above for Direct Gene Knockout, with modifications to incorporate the ATc-repressible T7S4 promoter. Single guide RNAs targeting the start codon region of TgENT1 were designed using ChopChop online website tool and are listed in (Supplemental Table S1). Each sgRNA was cloned into pg474 via BsaI restriction sites as described for knockout. The DHFR/T7S4 cassette was amplified from pDTS4-DHFR using primers bearing 50 bp homology to the TgENT1 locus (Supplemental Table S5). PCR reactions were performed with Platinum SuperFi II DNA Polymerase (Supplemental Tables S6&S7), products were verified by agarose gel electrophoresis and purified using the QIAquick PCR Purification kit (Qiagen). Plasmids were prepared using Qiagen miniprep/midiprep kits according to the manufacturer’s instructions.
2.7.3. Transfection and Selection
Transfections were carried out using freshly egressed tachyzoites. Briefly, 1–5 × 106 parasites were resuspended in Cytomix (120 mM KCl, 0.15 mM CaCl₂, 25 mM HEPES, 5 mM MgCl₂, 10 mM K₂HPO₄/KH₂PO₄, pH 7.6) supplemented immediately before use with 65 µl of 3 mM ATP and 65 µl of 3 mM glutathione. Approximately ~50 µL of purified PCR cassette and 50–70 µg of sgRNA/Cas9 plasmid were added (total electroporation volume ~0.8 mL in a 4 mm cuvette). Electroporation was performed using a square-wave protocol (1700 V, 0.2 ms pulse length, two pulses separated by 5 s). Parasites were transferred to confluent HFF monolayers and incubated at 37 °C with 5% CO₂. Integration events using DHFR were selected with pyrimethamine (1 μM) within 24 h for 5–8 days. Once the drug-selected parasites grow out (5 –7 days), a 96-well plate of HFF cells was used for cloning the pool by serial dilution. Correct integration and promoter replacement were verified by diagnostic PCR (Supplemental S8 and S9) using primers listed in Table S5; knockdown efficiency was assessed by qRT-PCR where indicated using primers listed in Table S10.
2.7.4. Fluorescence-Activated Cell Sorting
For constructs incorporating mNeonGreen (e.g., enrichment of double knockout pools and ΔTgAT1 pool), transfected parasites were allowed to recover for ~48 h, then released, passed through a 26 G needle, and a 3 μm filter. The parasites were collected and centrifuged for 10 min at RT. The supernatant was discarded and resuspended in FACS buffer (1% Fetal calf serum, 1 mM EDTA in PBS) at roughly 107/mL and 1–2 mL transferred to FACS tubes. The transfected samples, including positive control (green), negative control sample (red tomato), sorted on a high-speed cell sorter (BD FACSAria IIu or BD FACSAria III high-speed cell sorter). Gating was established using non-transfected (RH) and mNeonGreen-positive controls. Single fluorescent parasites were sorted directly into 96-well plates pre-seeded with HFFs and cultured at 37 °C/5% CO₂ for 5–7 days to obtain single clonal plaques. Single clones were confirmed by PCR and qRT-PCR using primers listed in Tables S5 and S10.
2.8. Plaque Assay
Plaque assays were performed to assess parasite growth as described previously [37]. Confluent HFF monolayers in 6-well plates were infected with freshly egressed tachyzoites (100 parasites per well). Parasites were grown in the presence or absence of anhydrotetracycline (ATc, 0.5 µM) and incubated at 37 °C with 5% CO₂ for 7 days without agitation. After incubation, wells were examined by light microscopy to confirm plaque formation. Cells were fixed by adding 300 μL of ice-cold 100% methanol per well and incubating for 20 min at room temperature. Fixed monolayers were washed three times with PBS and stained with 2 mL of 0.4% crystal violet solution for ~2 h at room temperature. Wells were washed three times with PBS, air-dried, and plaques were imaged.
2.9. Quantitative Real Time PCR (qRT-PCR) in T. gondii Tachyzoites
qRT-PCR was carried out as previously described [38,39], to assess gene expression changes. Specifically, qRT-PCR was used to measure the expression level of mRNA for TgENT1 downregulation upon ATc treatment, to confirm the deletion of TgAT1, TgENT2, TgENT3, and the double knockout (DK) lines, and to evaluate potential upregulation or downregulation of TgENT genes in each knockout line. qRT-PCR primers were designed using NCBI Primer-BLAST website Primer designing tool (Supplemental Table S10). For knockdown experiments, parasites were grown in the presence or absence of ATc for 24, 48, or 72 h; for knockout lines, parasites were grown without ATc. Parasites were collected, passed through a 3 µm polycarbonate filter, and pelleted by centrifugation at 1500 x g for 10 min at RT. Total RNA was extracted using the RNeasy Mini Kit (Qiagen) with on-column DNase I treatment (Thermo Fisher) according to the manufacturer’s instructions. cDNA was synthesized using the High-Capacity RNA-to-cDNA Kit (Applied Biosystems). qRT-PCR reactions were set up with Power SYBR Green PCR Master Mix (Applied Biosystems) using 10 ng of cDNA as template and gene-specific primers (listed in Table S10). Reactions were run on a 7500 Real-Time PCR System (Applied Biosystems) under standard cycling conditions. Relative expression was calculated using the 2-ΔΔCt method [40], using catalase mRNA as the internal control. Three independent biological replicates were performed for each condition, and data were analyzed and plotted using GraphPad Prism 10.0.
2.10. qRT-PCR for L. mexicana Promastigotes
qRT-PCR was performed as previously described [31,41] to determine the expression levels of TgENT1 in Lmex-NT3-KO compared to the control line and to select the optimal clone. RNA was extracted from L. mexicana promastigotes (2 × 106 cells/mL) using the NucleoSpin RNA kit (Macherey-Nagel, Düren, Germany) according to the manufacturer’s instructions and quantified using a NanoDrop ND-1000 spectrophotometer. Samples were stored at −80 °C until further use. qRT-PCR primers were designed using NCBI Primer-BLAST website Primer designing tool (Supplemental Table S10). cDNA was synthesized from 2 µg of RNA using the Precision nanoScript II Reverse Transcription kit (PrimerDesign Ltd., Eastleigh, UK) following the manufacturer’s protocol and stored at −20 °C. Primer efficiency was determined using the Pfaffl method [42]. Amplification was performed using the PrecisionPLUS OneStep RT-qPCR Master Mix kit (PrimerDesign Ltd., Eastleigh, UK) with SYBR Green on a 7500 Real-Time PCR System (Applied Biosystems, Thermo Fisher Scientific, Oxford, UK). Cycling conditions were: 50 °C for 2 min, 95 °C for 10 min, followed by 45 cycles of 95 °C for 15 s and 60 °C for 1 min. A dissociation curve was included to confirm single-product amplification. Negative controls included samples without reverse transcriptase or cDNA. Gene expression was normalized to GPI8, a constitutively expressed housekeeping gene in L. mexicana [43]. Relative quantification was calculated using the 2-ΔΔCt method, and data analysis was performed using Applied Biosystems 7500 Fast Real-Time PCR System Software. Each experiment was carried out in triplicate with three independent biological replicates, and data were analyzed and plotted using GraphPad Prism 10.0.
3. Results
3.1. Creation of TgENT Knockout (KO) and Knockdown (KD) Strains
3.1.1. Identification of Toxoplasma ENTs
We performed protein-protein BLAST or position-specific iterated BLAST on ToxoDB (https://toxodb.org/toxo/app) using human (ENT1-4, CNT1-3) or Plasmodium falciparum (ENT1-4) nucleoside transporter genes as query sequences to search for Toxoplasma homologs. Consistent with similar queries of other protozoan genomes [14,44], no homologs of human CNTs were identified in any of the genomes accessible through ToxoDB (https://toxodb.org/toxo/app). However, the searches identified four genes in the Toxoplasma genome with ENT homology, that are numbered as follows in the ME49 genome: TGME49_233130, TGME49_244440, TGME49_288540, and TGME49_500147. Of these, gene TGME49_244440 was previously proposed to be an Adenosine Transporter (TgAT1) [24] and we propose to keep that name so as not to confuse the literature although we have shown that this gene encodes a transporter with much higher affinity for oxopurines than for adenosine [25]. The other genes were given an NT1 – NT3 numbering in the PhD thesis by Kshitiz Chaudhary of the research group of David Roos [45] and we propose, again, to keep the same numbering, but naming the genes TgENT1 – 3 (Table 1), as the function of these genes has not been established yet. While they are likely members of the ENT gene family, nucleoside transporter activity was not demonstrated at this point. This naming is also consistent with a recent paper by Messina, Goerner, Bennett, Brennan, Carruthers and Martorelli Di Genova [26] and while a paper describing the cellular localization of some of the T. gondii nucleoside transporters [46] provided slightly different names, we propose to consolidate according to the current majority here (Table 1).
3.1.2. Construction of TgENT2 and TgENT3 Knockouts in T. gondii RH Cell Line
In order to generate the tools to identify which one of the TgENT genes may encode the previously described Uracil/Uridine Transporter TgUUT [13], we opted to use genetic deletion of TgENT2 and TgENT3 in the T. gondii RH background. Likewise, we targeted TgENT1 where all attempt at creating a direct knockout were futile, as predicted in the CRISPR/Cas9 screen [47] indicating that TgENT1 is essential for growth in culture, and a conditional knockdown was thus attempted instead; the other three TgENTs were found to be non-essential in the same screen and thus direct knock-out (KO) was attempted. Finally, although we have previously characterized TgAT1 in detail and showed that it is not fully inhibited by even 1 mM uracil or uridine [25], the gene encoding this transporter was also targeted for deletion: (1) in order to confirm the previous observations regarding uracil and uridine, (2) the transporter displayed higher affinity for thymidine than for uracil and 5F-uracil is really a thymine analogue, i.e. it is just possibly that TgAT1 could take up 5-fluorinated uracil and uridine even though it does not recognize uracil and uridine itself, and (3) to examine the possibility that even if TgAT1 only displays low affinity and a slow translocation rate for 5-fluoropurimidines, this still could be sufficient for a parasiticidal effect if the parasite is exposed to a drug for a prolonged period [6].
TgENT2-KO and TgENT3-KO were generated using CRISPR/Cas9 by transfecting RH parasites with the cassette amplified from pDT7S4 template [28] and with Tub-Cas9-YFP-pU6-ccdB-tracrRNA plasmid [35] containing the specific sgRNAs to cut the gene near the start or stop codon, respectively, together facilitating the replacement of the gene of interest (GoI) with the DHFR gene that confers pyrimethamine resistance for selection (Figure 1A,C). Correct integration was confirmed by PCR showing the presence of the ~3.8 kb DHFR amplicon and the absence of the targeted GoI in the transfectants (Figure 1B,D).
The absence of each targeted GoI was further confirmed at the mRNA level using qRT-PCR; gene expression was normalized to the catalase gene expression (Figure 1E,F). In both knockout strains, mRNA of the targeted gene could not be detected but the mRNA level of the other three TgENTs was unchanged relative to the parental RH cells (except a minor, just-significant (P < 0.05, t-test) reduction in TgENT1 mRNA in the TgENT2-KO), suggesting that there was no compensatory upregulation of other transporters after the deletion of either TgENT2 or TgENT3, at least in culture and at the mRNA level.
3.1.3. Creation of a TgENT2/3 Double Knockout Cell Line (ENT2/3dKO)
An TgENT2/3dKO strain was generated using TgENT3-KO as the parental line; the mNeonGreen (mNG) cassette [36] was used as a selectable marker for this construct (Figure 2A). A cassette whereby mNG is flanked by the 5’- and 3’-UTRs of TgENT2 was amplified and transfected into TgENT3-KO cells. Transfectants expressing mNG were isolated using flow cytometry. Successful integration in the TgENT2 locus, was confirmed by PCR reactions (Figure 2B) and the absence of the TgENT2 and TgENT3 corresponding mRNAs was confirmed by qRT-PCR (Figure 2C). As in the single gene deletions, there was no significant upregulation of the two remaining TgENTs relative to their expression level in RH control cells.
3.1.4. Deletion of TgAT1 in the RH Cell Line
The knockout of TgAT1 by CRISPR-assisted homologous recombination and replacement with a DHFR cassette, as for TgENT2 and TgENT3 was attempted multiple times but did not yield viable cells upon selection pressure with pyrimethamine, indicating that, at least in our RH strain and under the in vitro conditions used, TgAT1 may essential unlike the other two TgENTs. We therefore created a new construct for replacement with mNG, flanked by the TgAT1 UTRs (Figure 3A). Replacement with this cassette was followed by flow cytometry to isolate positive green fluorescent clones. Correct integration and TgAT1-KO were confirmed by PCR reactions (Figure 3B) and mRNA depletion was confirmed by qRT-PCR. Once again, the depletion of TgAT1 had no significant effect on the mRNA levels of the other TgENT genes in support for an absence of compensatory mechanism between those genes in T. gondii (Figure 3C).
3.1.5. Creation of a Conditional Knockout of TgENT1
As mentioned, TgENT1 is predicted to be essential to tachyzoites in vitro [47]. We therefore used promotor replacement (PR) to engineer a conditional depletion line for this gene under control of a tetracycline-inducible transactivator system, illustrated in Figure 4A. This technique relies on replacing the endogenous promotor with the tetracycline responsive promoter T7S4 [28,48]. Primers containing homology to 50 bp stretches of the 5’ UTR and the first exon promoted the insertion of the pyrimethamine-resistance (DHFR) cassette for selection followed by the T7S4 promoter that then controls transcription of the native copy of TgENT1 (Figure 4A) (as described in Lacombe, et al. [49]). In this system the addition of anhydrous tetracycline (ATc) binds to the transactivator and prevents transcription, resulting in gene depletion. The resulting manipulation was verified by PCR and after transfection clones with the correct integration of the construct were selected on pyrimethamine and verified by PCR (Figure 4B); the new cell line was named TgENT1-KD. Depletion of the corresponding mRNA was tested by qRT-PCR which showed a strong and stable knockdown of mRNA levels as early as 24 h (Figure 4C). Using a plaque assay, we showed that the TgENT1-KD cells did not replicate in the presence of ATc, whereas the parental RH cells did (Figure 4D), confirming that TgENT1 is essential for growth in culture.
3.1.6. Expression of TgENTs in a Leishmania Mexicana Cell Line Deficient in Nucleobase Transport
We have previously shown the utility of expressing protozoan ENT-family transporters in L. mexicana cell lines that lack either nucleoside transport (ΔLmexNT1.1/1.2/2, “SUPKO”) or nucleobase transport (ΔLmexNT3, “LmexNT3-KO”) [31]. Here, we used that same procedure to express TgENT1 in the LmexNT3-KO strain. Th TgENT1 minigene was amplified by PCR from cDNA of the T. gondii RH strain and ligated into the NdeI and XhoI-digested plasmid pNUS-HcN [34] (Figure 5A). TgENT1 was then ligated into the digested pNUS-HcN using the NEBuilder HiFi DNA Assembly Cloning Kit, creating plasmid pHDK295, and used to transform E. coli. Six positive clones were picked and the correct plasmid assembly verified by a diagnostic digestion with NdeI and XhoI and by PCR for TgENT1 integration into pHDK295, followed by Sanger sequencing of the amplicon. Upon confirmation of the correct sequence, pHDK295 was transfected into LmexNT3-KO and selected on 50 µg/mL of G-418. Clones were obtained by limiting dilution and the presence of the plasmid in these clones was confirmed by PCR (Figure 5B). The relative expression level of TgENT1 in these clones was assessed by qRT-PCR, showing highest expression in clone 2 (Figure 5C), which was therefore used for further analysis of the transporter function (see below).
3.2. Investigation into the Physiological Role of the TgENT Transporters
3.2.1. Is TgENT2 or TgENT3 Involved in Pyrimidine Uptake in T. gondii?
Messina et al. [26] reported that TgAT1 and TgENT3 gene products are important in bradyzoite formation and thus for persistence of T. gondii in the host. Several authors also demonstrated that TgENT1 is essential in tachyzoites [26,46,47]. However, the role of the individual transport proteins in T. gondii physiology is poorly defined. We have recently reported the identification of a uracil/uridine transporter, TgUUT, in T. gondii tachyzoites [13] that is potentially important for anti-toxoplasma chemotherapy development. Our starting point was therefore to try to establish which of the four TgENT genes encodes this pyrimidine transporter, using the knockout and knockdown lines we generated.
As the TgAT1 gene product has already been characterized as a purine-specific transporter [24,25] and TgENT1 is essential and thus harder to manipulate, we first performed uridine transport assays with the knockout strains of TgENT2 and TgENT3 (Section 1.2) in parallel with the parental strain as a control. Figure 6A shows that the two knockout strains displayed identical rates of uridine uptake, although both showed an apparently lower rate than the control and requiring us to investigate uridine uptake in both strains in detail.
The knockout strain TgENT2-KO displayed robust uptake of 0.1 µM [3H]-uridine, with an average Km of 5.29 ± 0.58 µM and a Vmax of 0.025 ± 0.02 pmol(107 cells)-1min-1 (Figure 6B), which is not significantly different from the corresponding values in the parental cell line (Table 2). Indeed, the inhibition constants for uracil, inosine and adenosine were also highly similar to the parental line (Figure 6C, Table2; P >0.05). The same pattern was observed with the TgENT3-KO (Figure 6D,E) and Table 2 lists the three sets of parameters side-by-side. The data allowed for two possible hypotheses: (a) neither gene encodes the TgUUT1 uridine/uracil transporter, or (b) they both express a highly similar uridine/uracil transporter and only the knockout of both genes simultaneously would create a cell line that was deficient in the uptake of these pyrimidines. The TgENT2/3dKO strain (Section 1.3) allowed us to distinguish between these possibilities. Figure 6F shows that uridine uptake was identical in TgENT2/3dKO and parental cells, and Figure 6G shows that uracil uptake was somewhat higher in the dKO cells – certainly not lower. Overall, these data show that neither TgENT2 nor TgENT3 encode the TgUUT1 pyrimidine transport activity.
3.2.2. Investigating the Role of TgENT1 in Pyrimidine Transport
As the TgENT1 gene could not be deleted, its role in pyrimidine salvage by tachyzoites was investigated using the promotor replacement line (section 1.4), with expression suppressed upon addition of ATc. As the level of mRNA depletion was almost maximal at 24 h (Figure 4D) and there is a risk of secondary effects on other cellular processes the longer the essential gene is suppressed, transport was assessed at 24 h and at 48 h, and the rates in the presence and absence of ATc compared.
The uptake of 0.1 µM [3H]-uridine was highly significantly reduced in TgENT1-KD cells incubated for either 24 h or 48 h with ATc, to 62.7% of control (P < 0.0001, F-test) or 66.5% of control (P = 0.0004), respectively; the linear regression slopes of the 24 h and 48 h knockdown lines were not significantly different from each other (P > 0.05) (Figure 7A). Similarly, uptake of 0.1 µM [3H]-uracil was significantly lower after incubation with ATc for 24 h (63.8% of control, P = 0.0063) or 48 h (47.4% of control, P = 0.0048) (Figure 7B). As a control, we conducted the same experiment also with 0.1 µM [3H]-tryptophan, and in this case the incubation with ATc for either 24 h or 48 h did not affect the rate of uptake (Figure 7C). We thus conclude that the suppression of TgENT1 mRNA production specifically diminishes the transport of uridine and uracil in tachyzoites.
For further verification we next expressed TgENT1 in our recently described Leishmania mexicana NT3-KO strain (Section 1.5), which displays a null background for purine nucleobase transport and a very low background for uracil uptake, exactly as described for the Trypanosoma vivax NT3 nucleobase transporter [31] and the T. cruzi NB1 adenine transporter [33]. Expressed from an episome in this system, the TgENT1 gene product was able to significantly increase 0.1 µM [3H]-uracil transport over the background, in a time-dependent fashion, in all three independently generated clones expressing the transporter (Figure 7D). As a further control we assessed the uptake of [3H]-adenine in the same TgENT1-expressing clones, for which the LmexNT3-KO cells give an exceptionally sensitive system, as L. mexicana adenine uptake in those cells was only 1.9% and 1.6% of control at 30 s and 60 s incubation time, respectively, in the L. mexicana-Cas9 T7 strain [29] that is the parent line of LmexNT3-KO [31] (Figure 7E). [3H]-adenine was just-detectable and very slightly elevated in two out of the three TgENT1-expressing clones tested at each timepoint but remained <3% of the rate in cas9 cells (Figure 7E). However, the data indicate that TgENT1 has a much higher capacity for uracil uptake than for adenine uptake. The combined data with the TgENT1-KD tachyzoites and the expression of TgENT1 in L. mexicana NT3-KO cells strongly suggest that TgENT1 encodes TgUUT1.
3.3. Effect of Transporter Knockouts on Sensitivity to 5-Fluorinated Pyrimidines
If the above conclusion, that TgENT1 is likely to be the gene encoding the TgUUT1 uracil/uridine uptake activity, is correct, then sensitivity to pyrimidine antimetabolites such as 5F-uracil, 5F-uridine and 5F-2’deoxyuridine, all potent agents against T. gondii in vitro [13], should not vary with the expression levels of the other three TgENTs.
All three pyrimidine analogues that were screened in our previous study [13] showed good activity against T. gondii RH with a cytostatic effect at lower concentrations and a cytocidal effect at high concentrations against HFF monolayer cells. The best activity was displayed by 5-fluorouridine, with an EC50 of 0.45 µM, 27.6-fold lower than for the standard drug sulfadiazine (Table 3).
The pyrimidine analogues displayed essentially the same activity against the knockout strains of TgENT2, TgENT3 and TgAT1, with the sole exception of a just-significant sensitization by the TgENT2-KO strain to 5F-uridine (EC50 = 0.33 µM; P = 0.02) and with a similar trend in TgENT3 cells that did not reach significance (EC50 = 0.36 µM; P = 0.07). In the double knockout strain that EC50 value further decreased to 0.29 µM (P = 0.003) and the values for 5F-uracil and 5F-2’deoxyuridine were also significantly lower. This trend could be due to a possible upregulation in the pyrimidine salvage pathways or perhaps the internal distribution of pyrimidine metabolites in the cell. The observations with the 5F-pyrimidine antimetabolites are fully consistent with TgAT1, TgENT2 and TgENT3 having no significant role in uracil and uridine transport. Unfortunately, it was not possible to test 5F-pyrimidine sensitivity on a TgENT1-KO, which we could not isolate as it is lethal, nor with the knockdown line which shows an early growth arrest, as the assay requires a healthy, growing cell population.
4. Discussion
In this study we aimed to identify the gene encoding the T. gondii uracil/uridine transporter TgUUT1 that we recently described in isolated tachyzoites [13]. Tachyzoites with a disrupted pyrimidine biosynthesis pathway (i.e. auxotrophs) are able to grow in vitro on uracil or uridine as the sole pyrimidine source [12] but not on thymidine, as the species lack thymidine kinase [50] and is unable to take up [3H]-thymidine [13]. The lack of thymidine salvage makes Toxoplasma, like Plasmodium species, particularly sensitive to antifolates, which are the first-line treatment for toxoplasmosis, as the parasite relies completely on the synthesis of thymidine from 2’deoxyuridine by folate-dependent thymidylate synthase. [12]. The lack of thymidine kinase similarly underpins the remarkable sensitivity to 5F-uracil, 5F-uridine and 5F-2’deoxyuridine as their mode of action as the metabolite 5F-2’deoxyuridine monophosphate (5F-dUMP) inhibits thymidylate synthase, and 5F-2’dUTP is incorporated in place of TTP into nucleic acids, leading to growth arrest and fatal DNA damage, respectively [51]. The reliance on thymidylate synthase is shared with the related Plasmodium species as these are also unable to take up thymidine [44], yet Plasmodium are very much less sensitive to 5F-uracil, 5F-uridine and 5F-2’deoxyuridine but highly sensitive to 5F-orotate [52] as, unlike T. gondii [13], orotate is the only pyrimidine they are able to take up or incorporate into their nucleic acids [53,54]. This provides a demonstration for the importance of transporters in the effectiveness of fluorinated pyrimidines and other antimetabolites.
In agreement with other recent reports [26,46] we identified four potential nucleoside transporter genes in the Toxoplasma genomes available on toxoDB.org, all of the ENT family. Of these, we have previously characterized TgAT1 in great detail and shown that it encodes a high affinity purine transporter with no measurable affinity for uracil and uridine [25]. As TgUUT1 displays high affinity for both uracil and uridine, and low affinity at best for purine nucleosides [13], TgAT1 does not encode the uracil/uridine transporter. In the current study we made gene deletion strains for TgENT2 and TgENT3 as well as the double knockout TgENT2/3dKO and showed that these deletions did not materially change [3H]-uridine uptake parameters or reduce sensitivity to 5F-pyrimidines leading us to conclude that neither of these genes encode the TgUUT1 activity either, leaving TgENT1 as the most likely candidate.
As the gene could not be deleted [26] we created a cell line where its expression is controlled by the tetracycline responsive promoter T7S4 and thus suppressed in the presence of ATc. As expected, the abundance of the corresponding mRNA was reduced by >80% at 24 h, 48 h and 72 h. In order to minimize any secondary effects from the knockdown we assessed the uptake of 0.1 µM [3H]-uridine or 0.1 µM uracil at 24 h and 48 h and compared the rate in the presence and absence of ATc. The uptake rate for both substrates, and at both timepoints, was highly and significantly reduced, while the rate of uptake of 0.1 µM [3H]-tryptophan was unaffected. Moreover, the expression of TgENT1 in a Leishmania cell line with a low endogenous rate of uracil uptake increased uptake of 0.1 µM [3H]-uracil significantly in all three independent clones assayed while barely affecting [3H]-adenine uptake rates despite the near-zero uptake level for the purine nucleobase.
The combined data presented strongly suggest that TgENT1 encodes the TgUUT1 uracil/uridine transport activity. Certainly, we have conclusively shown, in this and our previous paper [13], that none of the other TgENTs are involved in uracil or uridine uptake. Logically, however, that leaves the possibility of a non-ENT family transporter to be also involved in this process. Such a hypothetical transporter is unlikely to be of the CNT family either, nor of any of the other known nucleobase or nucleoside transporter families [14,55] as, to the best of our knowledge, no members of any of these gene families have to date been identified in protozoan genomes. However, it should be noted here that the genes encoding the high affinity uracil transporters of Leishmania [56] and Trypanosoma species [57,58] have not yet been identified and we have speculated that they may be encoded by a different, as yet unknown transporter family [59]. However, these kinetoplastid transporters are extremely selective for uracil only whereas TgUUT1 displays identical affinity for uracil and uridine [13]. Substantial efforts from our group [60,61] and the group of Marc Ouellette [62] have not identified any such novel transporter in kinetoplastids.
In conclusion, we believe that TgENT1 encodes the T. gondii uracil/uridine transporter TgUUT1, responsible for the uptake of 5F-pyrimidines with high activity against this parasite. Since the TgENT1 transport activity is essential, as is the thymidine synthase pathway that mediates the action of these compounds, its use would not be expected to lead to rapid onset of resistance.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: PCR master mix for Phusion High-Fidelity DNA Polymerase; Table S2: The PCR condition for Phusion High-Fidelity DNA Polymerase; Table S3: Primers for TgENT1 ligation into pNUS-HcN and post-transfection integration confirmation in L. mex NT3-KO; Table S4: sgRNA primers for generating knockouts (TgENT2, TgENT3, TgAT1, Knockdown of TgENT1, and DK in (TgENT3) in T. gondii; Table S5: List of primers designed and used for generation and confirmation of TgAT1-KO, TgENT1, TgENT2, TgENT3 and DK (ΔTgENT2/3); Table S6: Platinum SuperFI™ Green PCR Master Mix; Table S7: PCR (SuperFi™) program; Table S8: GoTaq G2 Hot Start Green Master Mix DNA Polymerase; Table S9: PCR conditions for GoTaq DNA Polymerase; Table S10: List of primers designed and used in qRT-PCR.
Author Contributions
Conceptualization, H.P.dK.; methodology: L.S., M.F.S.; investigation, H.A.A.E., M.F.S.; formal analysis, H.A.A.E., L.S., H.P.dK.; resources: L.S., H.P.dK.; writing—original draft preparation, H.A.A.E., H.P.dK.; writing—review and editing, , H.A.A.E., L.S., H.P.dK.; supervision, L.S., H.P.dK.; funding acquisition, L.S., H.P.dK.
Funding
This research was funded by a Wellcome Investigator Award (217173/Z/19/Z) and a Wellcome Discovery Award (310879/Z/24/Z)(to L.S.). HE was supported by a studentship from the government of Libya.
Data Availability Statement
All relevant data are contained in the manuscript and the supplementary materials.
Acknowledgments
We thank the Cellular Analysis Facility from MVLS SRF for their support and assistance with the flow cytometry work. We thank the ToxoDB team (https://toxodb.org) and the community contributors for enabling genome mining for Toxoplasma gondii.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ENT | Equilibrative Nucleoside Transporter |
| TgUUT1 | Toxoplasma gondii Uracil/Uridine Transporter 1 |
| TgAT1 | Toxoplasma gondii Adenosine Transporter 1 |
References
- Dubey, J.; Murata, F.; Cerqueira-Cézar, C.; Kwok, O.; Villena, I. Congenital toxoplasmosis in humans: an update of worldwide rate of congenital infections. Parasitology 2021, 148, 1406–1416. [Google Scholar] [CrossRef] [PubMed]
- Montoya, J.G.; Liesenfeld, O. Toxoplasmosis. Lancet 2004, 363, 1965–1976. [Google Scholar] [CrossRef]
- Kim, K.; Weiss, L.M. Toxoplasma gondii: the model apicomplexan. Int J Parasitol 2004, 34, 423–432. [Google Scholar] [CrossRef]
- Boothroyd, J.C. Expansion of host range as a driving force in the evolution of Toxoplasma. Mem Inst Oswaldo Cruz 2009, 104, 179–184. [Google Scholar] [CrossRef]
- El Kouni, M.H.; Guarcello, V.; Al Safarjalani, O.N.; Naguib, F.N. Metabolism and selective toxicity of 6-nitrobenzylthioinosine in Toxoplasma gondii. Antimicrob Agents Chemother 1999, 43, 2437–2443. [Google Scholar] [CrossRef]
- Counihan, N.A.; Modak, J.K.; de Koning-Ward, T.F. How malaria parasites acquire nutrients from their host. Front Cell Dev Biol 2021, 9, 649184. [Google Scholar] [CrossRef]
- Schwab, J.; Beckers, C.; Joiner, K. The parasitophorous vacuole membrane surrounding intracellular Toxoplasma gondii functions as a molecular sieve. Proc Natl Acad Sci USA 1994, 91, 509–513. [Google Scholar] [CrossRef]
- Bitew, M.A.; Gaete, P.S.; Swale, C.; Maru, P.; Contreras, J.E.; Saeij, J.P.J. Two Toxoplasma gondii putative pore-forming proteins, GRA47 and GRA72, influence small molecule permeability of the parasitophorous vacuole. mBio 2024, 15, e0308123. [Google Scholar] [CrossRef]
- Berens, R.L.; Krug, E.C.; Marr, J.J. Purine and pyrimidine metabolism. In Biochemistry and molecular biology of parasites; Elsevier, 1995; pp. 89–117. [Google Scholar]
- Ngo, H.M.; Ngo, E.O.; Bzik, D.J.; Joiner, K.A. Toxoplasma gondii: are host cell adenosine nucleotides a direct source for purine salvage? Exp Parasitol 2000, 95, 148–153. [Google Scholar] [CrossRef]
- De Koning, H.P.; Al-Salabi, M.I.; Cohen, A.M.; Coombs, G.H.; Wastling, J.M. Identification and characterisation of high affinity nucleoside and nucleobase transporters in Toxoplasma gondii. Int J Parasitol 2003, 33, 821–831. [Google Scholar] [CrossRef]
- Fox, B.A.; Bzik, D.J. De novo pyrimidine biosynthesis is required for virulence of Toxoplasma gondii. Nature 2002, 415, 926–929. [Google Scholar] [CrossRef]
- Elati, H.A.; Goerner, A.L.; Martorelli Di Genova, B.; Sheiner, L.; De Koning, H.P. Pyrimidine salvage in Toxoplasma gondii as a target for new treatment. Frontiers in Cellular and Infection Microbiology 2023, 13, 1320160. [Google Scholar] [CrossRef]
- Campagnaro, G.D.; De Koning, H.P. Purine and pyrimidine transporters of pathogenic protozoa–conduits for therapeutic agents. Medicinal Research Reviews 2020, 40, 1679–1714. [Google Scholar] [CrossRef]
- Natto, M.J.; Miyamoto, Y.; Munday, J.C.; AlSiari, T.A.; Al-Salabi, M.I.; Quashie, N.B.; Eze, A.A.; Eckmann, L.; De Koning, H.P. Comprehensive characterization of purine and pyrimidine transport activities in Trichomonas vaginalis and functional cloning of a trichomonad nucleoside transporter. Mol Microbiol 2021, 116, 1489–1511. [Google Scholar] [CrossRef]
- Aldfer, M.M.; Alfayez, I.A.; Elati, H.A.; Gayen, N.; Elmahallawy, E.K.; Milena Murillo, A.; Marsiccobetre, S.; Van Calenbergh, S.; Silber, A.M.; De Koning, H.P. The Trypanosoma cruzi TcrNT2 Nucleoside Transporter Is a Conduit for the Uptake of 5-F-2′-Deoxyuridine and Tubercidin Analogues. Molecules 2022, 27. [Google Scholar] [CrossRef]
- Ungogo, M.A.; Aldfer, M.M.; Natto, M.J.; Zhuang, H.; Chisholm, R.; Walsh, K.; McGee, M.; Ilbeigi, K.; Asseri, J.I.; Burchmore, R.J.S.; et al. Cloning and Characterization of Trypanosoma congolense and T. vivax Nucleoside Transporters Reveal the Potential of P1-Type Carriers for the Discovery of Broad-Spectrum Nucleoside-Based Therapeutics against Animal African Trypanosomiasis. Int J Mol Sci 2023, 24, 3144. [Google Scholar] [CrossRef]
- Young, J.D. The SLC28 (CNT) and SLC29 (ENT) nucleoside transporter families: a 30-year collaborative odyssey. Biochem Soc Trans 2016, 44, 869–876. [Google Scholar] [CrossRef]
- Delespaux, V.; de Koning, H.P. Transporters in Anti-Parasitic Drug Development and Resistance. In Trypanosomatid diseases: molecular routes to drug discovery; Jäger, T., Koch, O., Flohe, L., Eds.; 2013; pp. 335–349. [Google Scholar]
- Munday, J.C.; Settimo, L.; de Koning, H.P. Transport proteins determine drug sensitivity and resistance in a protozoan parasite, Trypanosoma brucei. Front Pharmacol 2015, 6, 32. [Google Scholar] [CrossRef] [PubMed]
- Dunay, I.R.; Gajurel, K.; Dhakal, R.; Liesenfeld, O.; Montoya, J.G. Treatment of Toxoplasmosis: Historical Perspective, Animal Models, and Current Clinical Practice. Clin Microbiol Rev 2018, 31. [Google Scholar] [CrossRef]
- Vodenkova, S.; Buchler, T.; Cervena, K.; Veskrnova, V.; Vodicka, P.; Vymetalkova, V. 5-fluorouracil and other fluoropyrimidines in colorectal cancer: Past, present and future. Pharmacol Ther 2020, 206, 107447. [Google Scholar] [CrossRef]
- Dhiver, C.; Milandre, C.; Poizot-Martin, I.; Drogoul, M.P.; Gastaut, J.L.; Gastaut, J.A. 5-Fluoro-uracil-clindamycin for treatment of cerebral toxoplasmosis. AIDS 1993, 7, 143–144. [Google Scholar] [CrossRef] [PubMed]
- Chiang, C.W.; Carter, N.; Sullivan, W.J., Jr.; Donald, R.G.; Roos, D.S.; Naguib, F.N.; el Kouni, M.H.; Ullman, B.; Wilson, C.M. The adenosine transporter of Toxoplasma gondii. Identification by insertional mutagenesis, cloning, and recombinant expression. J Biol Chem 1999, 274, 35255–35261. [Google Scholar] [CrossRef]
- Campagnaro, G.D.; Elati, H.A.A.; Balaska, S.; Martin Abril, M.E.; Natto, M.J.; Hulpia, F.; Lee, K.; Sheiner, L.; Van Calenbergh, S.; de Koning, H.P. A Toxoplasma gondii Oxopurine Transporter Binds Nucleobases and Nucleosides Using Different Binding Modes. Int J Mol Sci 2022, 23, 710. [Google Scholar] [CrossRef]
- Messina, G.; Goerner, A.; Bennett, C.; Brennan, E.; Carruthers, V.B.; Martorelli Di Genova, B. Impact of equilibrative nucleoside transporters on Toxoplasma gondii infection and differentiation. mBio 2025, 16, e0220725. [Google Scholar] [CrossRef]
- Pittman, K.J.; Aliota, M.T.; Knoll, L.J. Dual transcriptional profiling of mice and Toxoplasma gondii during acute and chronic infection. BMC Genomics 2014, 15, 806. [Google Scholar] [CrossRef]
- Sheiner, L.; Demerly, J.L.; Poulsen, N.; Beatty, W.L.; Lucas, O.; Behnke, M.S.; White, M.W.; Striepen, B. A systematic screen to discover and analyze apicoplast proteins identifies a conserved and essential protein import factor. PLoS Pathog 2011, 7, e1002392. [Google Scholar] [CrossRef]
- Beneke, T.; Madden, R.; Makin, L.; Valli, J.; Sunter, J.; Gluenz, E. A CRISPR Cas9 high-throughput genome editing toolkit for kinetoplastids. R Soc Open Sci 2017, 4, 170095. [Google Scholar] [CrossRef]
- Kabli, A.M.M. Polyomic analyses for rational antileishmanial vaccine development: a role for membrane transporters? PhD thesis, University of Glasgow, 2021. [Google Scholar]
- Aldfer, M.M.; AlSiari, T.A.; Elati, H.A.A.; Natto, M.J.; Alfayez, I.A.; Campagnaro, G.D.; Sani, B.; Burchmore, R.J.S.; Diallinas, G.; De Koning, H.P. Nucleoside Transport and Nucleobase Uptake Null Mutants in Leishmania mexicana for the Routine Expression and Characterization of Purine and Pyrimidine Transporters. Int J Mol Sci 2022, 23, 8139. [Google Scholar] [CrossRef] [PubMed]
- Al-Salabi, M.I.; Wallace, L.J.; De Koning, H.P. A Leishmania major nucleobase transporter responsible for allopurinol uptake is a functional homolog of the Trypanosoma brucei H2 transporter. Mol Pharmacol 2003, 63, 814–820. [Google Scholar] [CrossRef]
- Aldfer, M.M.; Hulpia, F.; van Calenbergh, S.; De Koning, H.P. Mapping the transporter-substrate interactions of the Trypanosoma cruzi NB1 nucleobase transporter reveals the basis for its high affinity and selectivity for hypoxanthine and guanine and lack of nucleoside uptake. Mol Biochem Parasitol 2024, 258, 111616. [Google Scholar] [CrossRef] [PubMed]
- Tetaud, E.; Lecuix, I.; Sheldrake, T.; Baltz, T.; Fairlamb, A.H. A new expression vector for Crithidia fasciculata and Leishmania. Mol Biochem Parasitol 2002, 120, 195–204. [Google Scholar] [CrossRef]
- Curt-Varesano, A.; Braun, L.; Ranquet, C.; Hakimi, M.A.; Bougdour, A. The aspartyl protease TgASP5 mediates the export of the Toxoplasma GRA16 and GRA24 effectors into host cells. Cell Microbiol 2016, 18, 151–167. [Google Scholar] [CrossRef]
- Aghabi, D.; Sloan, M.; Gill, G.; Hartmann, E.; Antipova, O.; Dou, Z.; Guerra, A.J.; Carruthers, V.B.; Harding, C.R. The vacuolar iron transporter mediates iron detoxification in Toxoplasma gondii. Nat Commun 2023, 14, 3659. [Google Scholar] [CrossRef]
- Ufermann, C.M.; Muller, F.; Frohnecke, N.; Laue, M.; Seeber, F. Toxoplasma gondii plaque assays revisited: Improvements for ultrastructural and quantitative evaluation of lytic parasite growth. Exp Parasitol 2017, 180, 19–26. [Google Scholar] [CrossRef]
- Ovciarikova, J.; Shikha, S.; Lacombe, A.; Courjol, F.; McCrone, R.; Hussain, W.; Maclean, A.; Lemgruber, L.; Martins-Duarte, E.S.; Gissot, M.; et al. Two ancient membrane pores mediate mitochondrial-nucleus membrane contact sites. J Cell Biol 2024, 223, e202304075. [Google Scholar] [CrossRef] [PubMed]
- Maclean, A.E.; Sloan, M.A.; Renaud, E.A.; Argyle, B.E.; Lewis, W.H.; Ovciarikova, J.; Demolombe, V.; Waller, R.F.; Besteiro, S.; Sheiner, L. The Toxoplasma gondii mitochondrial transporter ABCB7L is essential for the biogenesis of cytosolic and nuclear iron-sulfur cluster proteins and cytosolic translation. MBio 2024, 15, e00872-00824. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ali, J.A.; Tagoe, D.N.; Munday, J.C.; Donachie, A.; Morrison, L.J.; De Koning, H.P. Pyrimidine biosynthesis is not an essential function for Trypanosoma brucei bloodstream forms. Plos one 2013, 8, e58034. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Kang, X.; Szallies, A.; Rawer, M.; Echner, H.; Duszenko, M. GPI anchor transamidase of Trypanosoma brucei: in vitro assay of the recombinant protein and VSG anchor exchange. J Cell Sci 2002, 115, 2529–2539. [Google Scholar] [CrossRef]
- De Koning, H.P.; Bridges, D.J.; Burchmore, R.J. Purine and pyrimidine transport in pathogenic protozoa: from biology to therapy. FEMS Microbiol Rev 2005, 29, 987–1020. [Google Scholar] [CrossRef]
- Chaudhary, K. Purine transport and salvage in apicomplexan parasites. Ph.D thesis, University of Pennsylvania, 2005. [Google Scholar]
- Qian, J.; Guo, L.; Yang, Y.; He, Z.; He, M.; Chen, C.; Luo, Y.; Xu, J.; Li, S.; Fang, R. A nucleoside transporter on the mitochondria of T. gondii is essential for maintaining normal growth of the parasite. Parasites & Vectors 2025, 18, 418. [Google Scholar] [CrossRef]
- Sidik, S.M.; Huet, D.; Ganesan, S.M.; Huynh, M.H.; Wang, T.; Nasamu, A.S.; Thiru, P.; Saeij, J.P.J.; Carruthers, V.B.; Niles, J.C.; et al. A Genome-wide CRISPR Screen in Toxoplasma Identifies Essential Apicomplexan Genes. Cell 2016, 166, 1423–1435 e1412. [Google Scholar] [CrossRef]
- Meissner, M.; Schluter, D.; Soldati, D. Role of Toxoplasma gondii myosin A in powering parasite gliding and host cell invasion. Science 2002, 298, 837–840. [Google Scholar] [CrossRef]
- Lacombe, A.; Maclean, A.E.; Ovciarikova, J.; Tottey, J.; Muhleip, A.; Fernandes, P.; Sheiner, L. Identification of the Toxoplasma gondii mitochondrial ribosome, and characterisation of a protein essential for mitochondrial translation. Mol Microbiol 2019, 112, 1235–1252. [Google Scholar] [CrossRef] [PubMed]
- Fox, B.A.; Belperron, A.A.; Bzik, D.J. Negative selection of Herpes simplex virus thymidine kinase in Toxoplasma gondii. Mol Biochem Parasitol 2001, 116, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Longley, D.B.; Harkin, D.P.; Johnston, P.G. 5-fluorouracil: mechanisms of action and clinical strategies. Nat Rev Cancer 2003, 3, 330–338. [Google Scholar] [CrossRef]
- Rathod, P.K.; Khatri, A.; Hubbert, T.; Milhous, W.K. Selective activity of 5-fluoroorotic acid against Plasmodium falciparum in vitro. Antimicrob Agents Chemother 1989, 33, 1090–1094. [Google Scholar] [CrossRef]
- Gutteridge, W.; Trigg, P. Incorporation of radioactive precursors into DNA and RNA of Plasmodium knowlesi in vitro. The J Protozool 1970, 17, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Van Dyke, K.; Tremblay, G.C.; Lantz, C.H.; Szustkiewicz, C. The source of purines and pyrimidines in Plasmodium berghei. Am J Trop Med Hyg 1970, 19, 202–208. [Google Scholar] [CrossRef]
- De Koning, H.; Diallinas, G. Nucleobase transporters. Mol Membr Biol 2000, 17, 75–94. [Google Scholar] [CrossRef] [PubMed]
- Papageorgiou, I.G.; Yakob, L.; Al Salabi, M.I.; Diallinas, G.; Soteriadou, K.P.; De Koning, H.P. Identification of the first pyrimidine nucleobase transporter in Leishmania: similarities with the Trypanosoma brucei U1 transporter and antileishmanial activity of uracil analogues. Parasitology 2005, 130, 275–283. [Google Scholar] [CrossRef]
- De Koning, H.P.; Jarvis, S.M. A highly selective, high-affinity transporter for uracil in Trypanosoma brucei brucei: evidence for proton-dependent transport. Biochem Cell Biol 1998, 76, 853–858. [Google Scholar] [CrossRef]
- Gudin, S.; Quashie, N.B.; Candlish, D.; Al-Salabi, M.I.; Jarvis, S.M.; Ranford-Cartwright, L.C.; de Koning, H.P. Trypanosoma brucei: a survey of pyrimidine transport activities. Exp Parasitol 2006, 114, 118–125. [Google Scholar] [CrossRef]
- De Koning, H.P. Pyrimidine transporters of trypanosomes–a class apart? Trends in Parasitology 2007, 23, 190. [Google Scholar] [CrossRef]
- Ali, J.A.M. Pyrimidine salvage and metabolism in kinetoplastid parasites; University of Glasgow, 2013. [Google Scholar]
- Alzahrani, K.J.H. Strategies for the identification and cloning of genes encoding pyrimidine transporters of Leishmania and Trypanosoma species. Ph.D thesis, University of Glasgow, 2017. [Google Scholar]
- Ritt, J.F.; Raymond, F.; Leprohon, P.; Legare, D.; Corbeil, J.; Ouellette, M. Gene amplification and point mutations in pyrimidine metabolic genes in 5-fluorouracil resistant Leishmania infantum. PLoS Negl Trop Dis 2013, 7, e2564. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Verification of TgENT3 and TgENT2 knockout in RH cells. A. Schematic depicting the genetic manipulation used to replace TgENT3 with the DHFR cassette. Primers used in (B) are shown. B. PCR confirmation of TgENT3-KO of DHFR integration after transfection of the RH cells. (L) 1 kb DNA Ladder; Using the TgENT3 -KO: (1) LSD-2071-(UTR-F of TgENT3) and LSD-1125-DHFR-R; ~1.3 kb; (2) LSD-1286 (DHFR-F) and LSD-2072 (UTR-R of TgENT3), 236 bp; (3) LSD-2071-(UTR-F of TgENT3) and LSD-2072-(UTR-R of TgENT3); ~3.8 kb; (4) LSD-2935 and LSD2936 (open reading frame primers of TgENT3); no band. Negative control, RH line: (5) LSD-2071-(UTR-F of TgENT3) and LSD-1125-DHFR-R; no band; (6) LSD-2072-(UTR-R of TgENT3 and LSD-1286-DHFR-F; no band (7) LSD-2071 (UTR-F of TgENT3) and LSD-2072 (UTR-R of TgENT3); 2.2 kb); (8) LSD-2935 and LSD-2936 ~2 kb. C. Schematic depicting the genetic manipulation used to replace TgENT2 with the DHFR cassette. D. PCR Confirmation of TgENT2-KO in RH cell line after transfection of the RH cells. (L) 1kb DNA Ladder (Promega); (1) LSD-2083 (UTR-F of TgENT2) and LSD-1125 (DHFR-R), ~1.3 kb; (2) LSD-2084 (UTR-R of TgENT2) and LSD-1286 (DHFR-F), 244 bp; (3) LSD-2083 (UTR-F of TgENT2 and LSD-2084 (UTR-R- of TgENT2), ~3.8 kb; (4) LSD-2939 and LSD-2940 (ORF primers of TgENT2), no band. RH cell line as negative control: (5) LSD-2083 (UTR-F of TgENT2) and LSD-1125 (DHFR-R); (6) LSD-2084 (UTR-R of TgENT2) and LSD-1286 (DHFR-F); no band (7) LSD-2083 (UTR-F of TgENT2) and LSD-2084 (UTR-R of TgENT2), ~2.7 kb; (8) LSD-2939-F and LSD-2940-R, ~2.5 kb. E. qRT-PCR analysis of TgENTs gene expression in the TgENT2-KO cell line. F. qRT-PCR analysis of TgENTs gene expression in the TgENT3-KO cell line.
Figure 1.
Verification of TgENT3 and TgENT2 knockout in RH cells. A. Schematic depicting the genetic manipulation used to replace TgENT3 with the DHFR cassette. Primers used in (B) are shown. B. PCR confirmation of TgENT3-KO of DHFR integration after transfection of the RH cells. (L) 1 kb DNA Ladder; Using the TgENT3 -KO: (1) LSD-2071-(UTR-F of TgENT3) and LSD-1125-DHFR-R; ~1.3 kb; (2) LSD-1286 (DHFR-F) and LSD-2072 (UTR-R of TgENT3), 236 bp; (3) LSD-2071-(UTR-F of TgENT3) and LSD-2072-(UTR-R of TgENT3); ~3.8 kb; (4) LSD-2935 and LSD2936 (open reading frame primers of TgENT3); no band. Negative control, RH line: (5) LSD-2071-(UTR-F of TgENT3) and LSD-1125-DHFR-R; no band; (6) LSD-2072-(UTR-R of TgENT3 and LSD-1286-DHFR-F; no band (7) LSD-2071 (UTR-F of TgENT3) and LSD-2072 (UTR-R of TgENT3); 2.2 kb); (8) LSD-2935 and LSD-2936 ~2 kb. C. Schematic depicting the genetic manipulation used to replace TgENT2 with the DHFR cassette. D. PCR Confirmation of TgENT2-KO in RH cell line after transfection of the RH cells. (L) 1kb DNA Ladder (Promega); (1) LSD-2083 (UTR-F of TgENT2) and LSD-1125 (DHFR-R), ~1.3 kb; (2) LSD-2084 (UTR-R of TgENT2) and LSD-1286 (DHFR-F), 244 bp; (3) LSD-2083 (UTR-F of TgENT2 and LSD-2084 (UTR-R- of TgENT2), ~3.8 kb; (4) LSD-2939 and LSD-2940 (ORF primers of TgENT2), no band. RH cell line as negative control: (5) LSD-2083 (UTR-F of TgENT2) and LSD-1125 (DHFR-R); (6) LSD-2084 (UTR-R of TgENT2) and LSD-1286 (DHFR-F); no band (7) LSD-2083 (UTR-F of TgENT2) and LSD-2084 (UTR-R of TgENT2), ~2.7 kb; (8) LSD-2939-F and LSD-2940-R, ~2.5 kb. E. qRT-PCR analysis of TgENTs gene expression in the TgENT2-KO cell line. F. qRT-PCR analysis of TgENTs gene expression in the TgENT3-KO cell line.
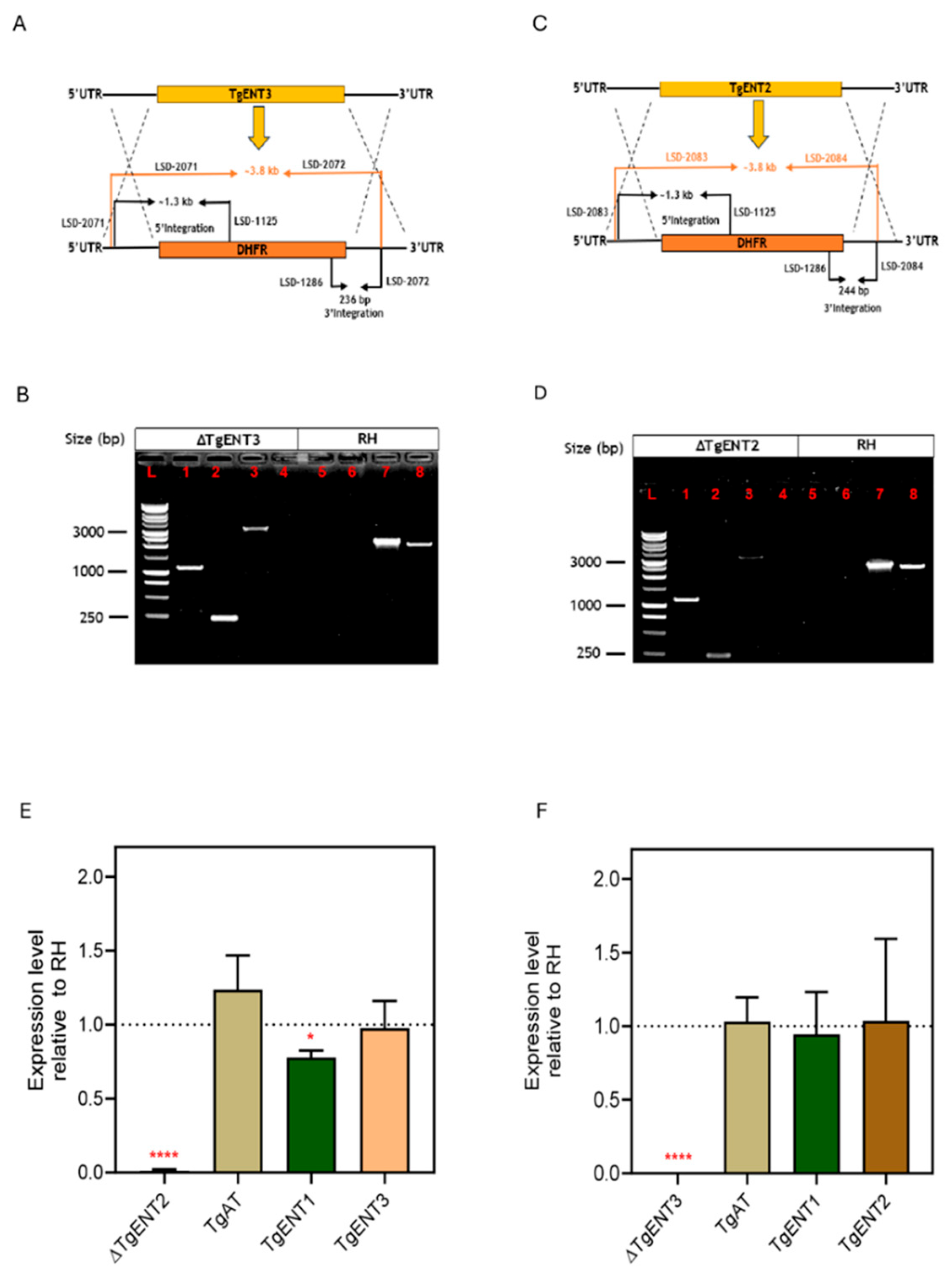
Figure 2.
Construction and verification of TgENT2/3dKO in RH cells. A. Schematic for the strategy of making a double knockout of TgENT2 and TgENT3, indicating primer positions. B. PCR confirmation of the TgENT2/3dKO in RH tachyzoites. L) 1kb DNA Ladder (Promega); (1) LSD-3004-F (5’ UTR of TgENT2) and LSD-3003-R (downstream start of TgENT2), no band; (2) LSD-3002-F (ORF of TgENT2) and LSD-3001-R (3’ UTR-R of TgENT2), no band; (3) LSD-3004 (5’ UTR of TgENT2) and LSD-2667 (mNG reverse), ~1.3 kb; (4) LSD-2666-F (mNG forward) and LSD-3001-R (3’ UTR-R of TgENT2), ~800 bp; (5) LSD-2935-F and LSD-2936-R (ORF of TgENT2), no band; (6) LSD-2939-F and LSD-2940-R (ORF of TgENT3), no band. RH cell line as a negative control: (7) LSD-3004 (5’ UTR of TgENT2) and LSD-3003-R (downstream start of TgENT2), ~750 bp. (8) LSD-3002-F (ORF of TgENT2) and LSD-3001-R (3’ UTR-R of TgENT2), 900 bp. (9) LSD-3004-F (5’ UTR of TgENT2) and LSD-2667-R (mNG ORF reverse); (10) LSD-2666-F (mNG ORF forward) and LSD-3001 (3’ UTR-R of TgENT2), ~800 bp; (11) LSD-2935-F and LSD-2936-R (ORF of TgENT2), ~2 kb; (12) LSD-2939-F and LSD-2940-R (ORF of TgENT3), ~2.5 kb. C. qRT-PCR assessment of mRNA levels in TgENT2/3dKO.
Figure 2.
Construction and verification of TgENT2/3dKO in RH cells. A. Schematic for the strategy of making a double knockout of TgENT2 and TgENT3, indicating primer positions. B. PCR confirmation of the TgENT2/3dKO in RH tachyzoites. L) 1kb DNA Ladder (Promega); (1) LSD-3004-F (5’ UTR of TgENT2) and LSD-3003-R (downstream start of TgENT2), no band; (2) LSD-3002-F (ORF of TgENT2) and LSD-3001-R (3’ UTR-R of TgENT2), no band; (3) LSD-3004 (5’ UTR of TgENT2) and LSD-2667 (mNG reverse), ~1.3 kb; (4) LSD-2666-F (mNG forward) and LSD-3001-R (3’ UTR-R of TgENT2), ~800 bp; (5) LSD-2935-F and LSD-2936-R (ORF of TgENT2), no band; (6) LSD-2939-F and LSD-2940-R (ORF of TgENT3), no band. RH cell line as a negative control: (7) LSD-3004 (5’ UTR of TgENT2) and LSD-3003-R (downstream start of TgENT2), ~750 bp. (8) LSD-3002-F (ORF of TgENT2) and LSD-3001-R (3’ UTR-R of TgENT2), 900 bp. (9) LSD-3004-F (5’ UTR of TgENT2) and LSD-2667-R (mNG ORF reverse); (10) LSD-2666-F (mNG ORF forward) and LSD-3001 (3’ UTR-R of TgENT2), ~800 bp; (11) LSD-2935-F and LSD-2936-R (ORF of TgENT2), ~2 kb; (12) LSD-2939-F and LSD-2940-R (ORF of TgENT3), ~2.5 kb. C. qRT-PCR assessment of mRNA levels in TgENT2/3dKO.
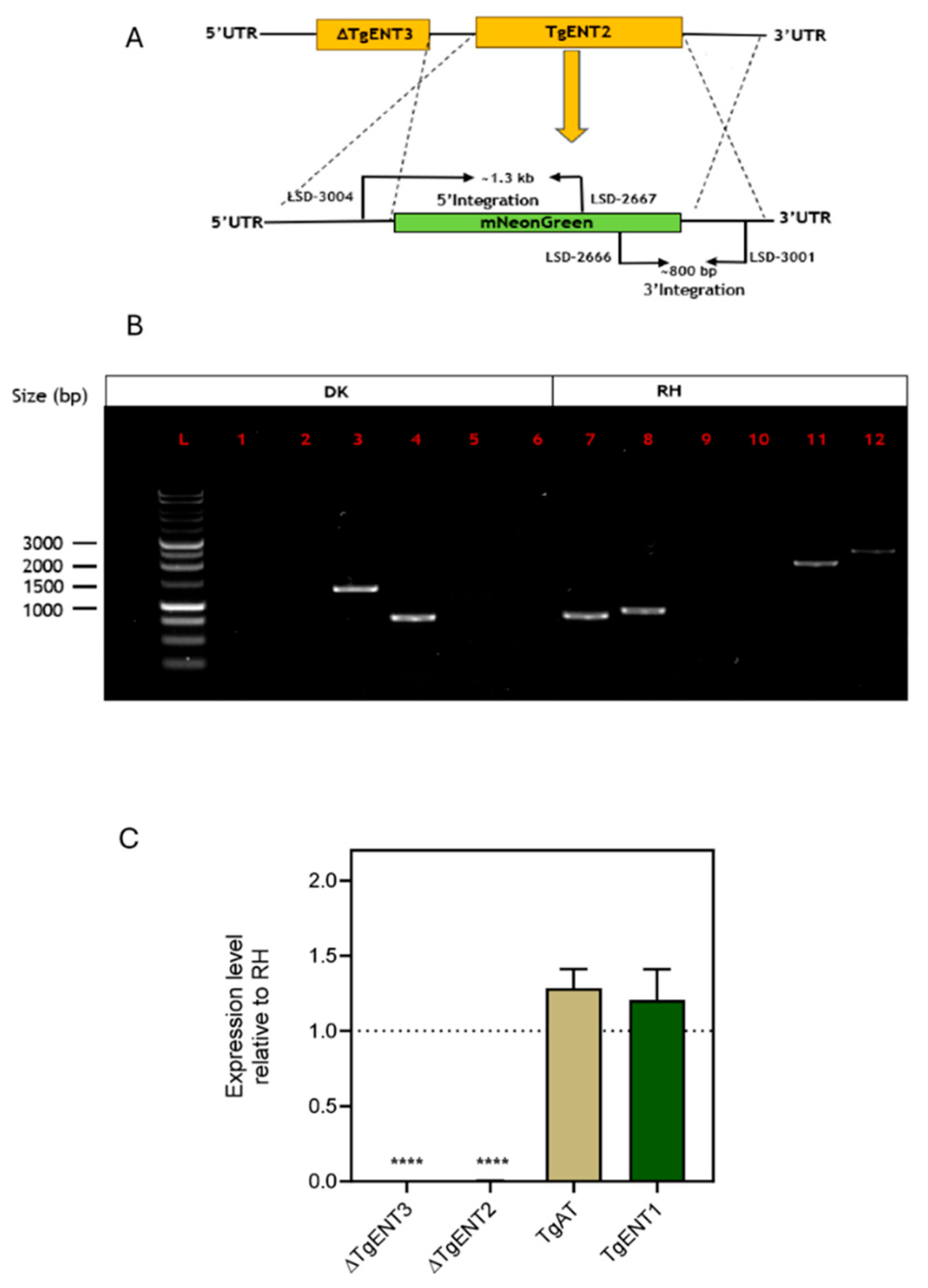
Figure 3.
Construction and verification of TgAT1-KO in RH cells. A. Schematic representing the genetic manipulation used to replace TgAT1 with the mNeonGreen cassette. Primers used in (B) are shown. B. PCR confirmation of TgAT1-KO using primers of TgAT1 and mNeonGreen cassette; (L) 1 kb DNA Ladder (Promega); (1) LSD-2669-F (upstream of the start codon of UTR of TgAT1) and LSD-2668-R (downstream start codon of TgAT1), no band; (2) LSD-2665-F (Upstream of the stop codon of TgAT1) and LSD-2664-R (downstream of stop UTR-R of TgAT1), no band; (3) LSD-2669-F (upstream of the start codon of UTR of TgAT1) and LSD-2667-R (mNeonGreen upstream reverse); band size (~1.4 kb); (4) LSD-2666-F (mNeonGreen downstream forward) and LSD-2664 (downstream of stop codon UTR-R of TgAT1); (~760 bp); (5) LSD-2937 and LSD-2938 (ORF of TgAT1), no band. RH cell line as a negative control: (6) LSD-2669-F (upstream of the start codon of UTR of TgAT1) and LSD-2668-R (downstream start codon of TgAT1); Expected band (~860 bp); (7) LSD-2665-F (Upstream of the stop codon of TgAT1) and LSD-2664-R (downstream of stop UTR-R of TgAT1); Expected band (~796 bp); (8) LSD-2669-F (upstream of the start codon of UTR of TgAT1) and LSD-2667-R (mNeonGreen upstream reverse); No band expected in RH control; (9) LSD-2666-F (mNeonGreen downstream forward and LSD-26664 (downstream of stop codon UTR-R of TgAT1); No band expected in RH control; (10) LSD-2937+LSD-2938 (open reading frame of TgAT1); Expected band (~ 1.5 kb). C. qRT-PCR analysis of TgENTs gene expression in TgAT1-KO cells.
Figure 3.
Construction and verification of TgAT1-KO in RH cells. A. Schematic representing the genetic manipulation used to replace TgAT1 with the mNeonGreen cassette. Primers used in (B) are shown. B. PCR confirmation of TgAT1-KO using primers of TgAT1 and mNeonGreen cassette; (L) 1 kb DNA Ladder (Promega); (1) LSD-2669-F (upstream of the start codon of UTR of TgAT1) and LSD-2668-R (downstream start codon of TgAT1), no band; (2) LSD-2665-F (Upstream of the stop codon of TgAT1) and LSD-2664-R (downstream of stop UTR-R of TgAT1), no band; (3) LSD-2669-F (upstream of the start codon of UTR of TgAT1) and LSD-2667-R (mNeonGreen upstream reverse); band size (~1.4 kb); (4) LSD-2666-F (mNeonGreen downstream forward) and LSD-2664 (downstream of stop codon UTR-R of TgAT1); (~760 bp); (5) LSD-2937 and LSD-2938 (ORF of TgAT1), no band. RH cell line as a negative control: (6) LSD-2669-F (upstream of the start codon of UTR of TgAT1) and LSD-2668-R (downstream start codon of TgAT1); Expected band (~860 bp); (7) LSD-2665-F (Upstream of the stop codon of TgAT1) and LSD-2664-R (downstream of stop UTR-R of TgAT1); Expected band (~796 bp); (8) LSD-2669-F (upstream of the start codon of UTR of TgAT1) and LSD-2667-R (mNeonGreen upstream reverse); No band expected in RH control; (9) LSD-2666-F (mNeonGreen downstream forward and LSD-26664 (downstream of stop codon UTR-R of TgAT1); No band expected in RH control; (10) LSD-2937+LSD-2938 (open reading frame of TgAT1); Expected band (~ 1.5 kb). C. qRT-PCR analysis of TgENTs gene expression in TgAT1-KO cells.
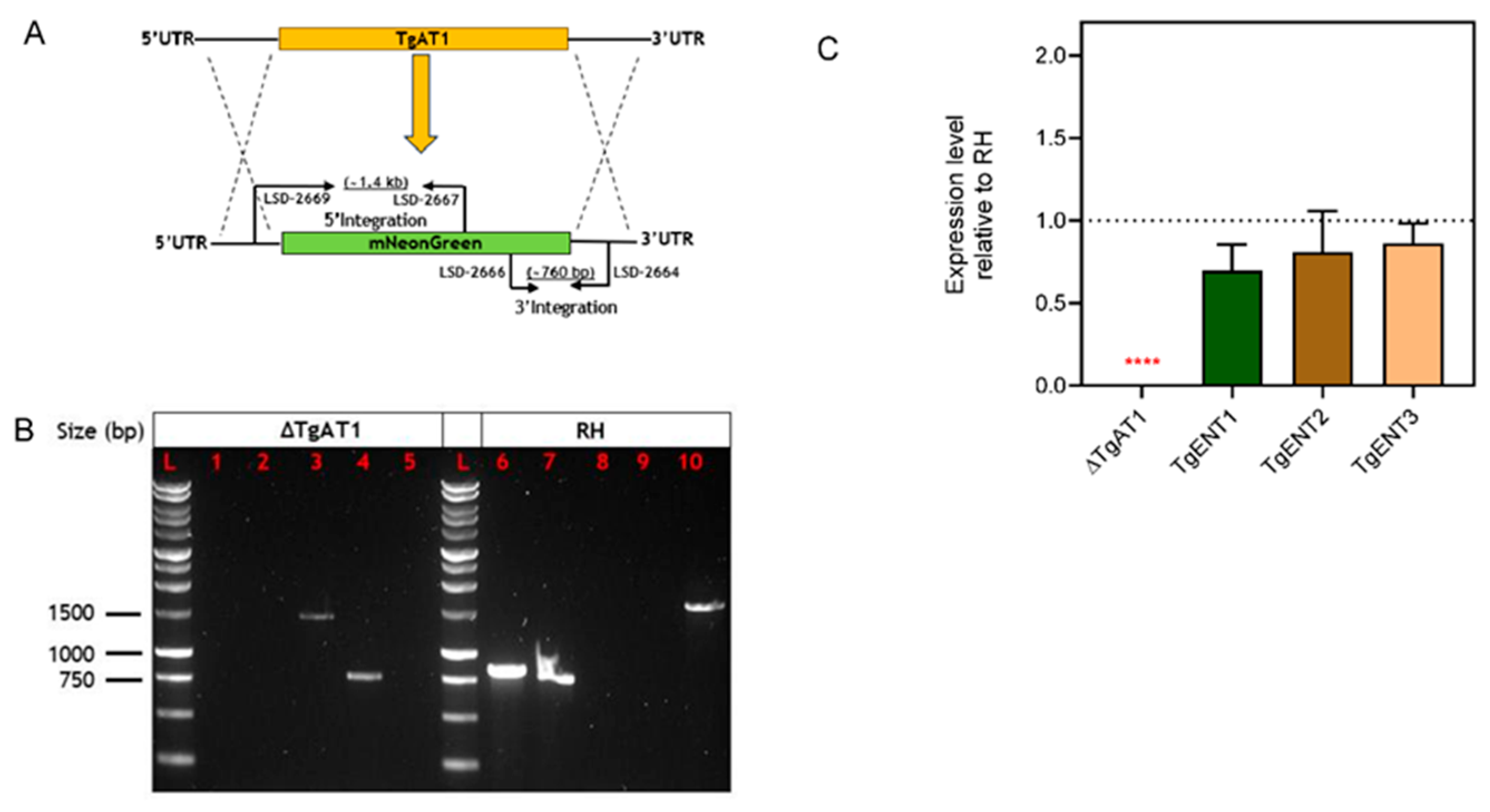
Figure 4.
Construction and verification of the promotor replacement for TgENT1. A. Schematic of the CRISPR/cas9-aided strategy for the introduction of the DHFR-TS74 construct upstream of the TgENT1 start codon. B. PCR validation of the promotor replacement for TgENT1 in RH cells. (L) 1 kb DNA Ladder; 1) LSD-1752 (5’ UTR-F of TgENT1) and LSD-37 (DHFR ORF reverse), ~ 1 kb; (2) LSD-38-F (T7S4-F) and LSD-1753 (ORF-R of TgENT1), ~380; (3) LSD-1752 (5’ UTR of TgNET1) and LSD-1753 (ORF-R of TgENT1), ~4.5 kb. (4) LSD-1752 (5’ UTR-F of TgENT1) and LSD-37 (DHFR cassette reverse), no band (5) LSD-38 (T7S4-F) and LSD-1753 (ORF-R of TgENT1), no band; (6) LSD-1752F and LSD-1753; band size ~280 bp. C. qRT-PCR of TgENT1 mRNA after addition of ATc, normalized to the expression of catalase. The amount of cDNA template used was 10 ng. The presented results are the mean and ± SEM of 3 independent biological replicates, each performed in triplicate. **P < 0.01 by one sample t-test. D. Plaque assay to assess in vitro growth of the TgENT1-KD strain (left) along with parental control (right) in HFF cells, in the presence and absence of ATc.
Figure 4.
Construction and verification of the promotor replacement for TgENT1. A. Schematic of the CRISPR/cas9-aided strategy for the introduction of the DHFR-TS74 construct upstream of the TgENT1 start codon. B. PCR validation of the promotor replacement for TgENT1 in RH cells. (L) 1 kb DNA Ladder; 1) LSD-1752 (5’ UTR-F of TgENT1) and LSD-37 (DHFR ORF reverse), ~ 1 kb; (2) LSD-38-F (T7S4-F) and LSD-1753 (ORF-R of TgENT1), ~380; (3) LSD-1752 (5’ UTR of TgNET1) and LSD-1753 (ORF-R of TgENT1), ~4.5 kb. (4) LSD-1752 (5’ UTR-F of TgENT1) and LSD-37 (DHFR cassette reverse), no band (5) LSD-38 (T7S4-F) and LSD-1753 (ORF-R of TgENT1), no band; (6) LSD-1752F and LSD-1753; band size ~280 bp. C. qRT-PCR of TgENT1 mRNA after addition of ATc, normalized to the expression of catalase. The amount of cDNA template used was 10 ng. The presented results are the mean and ± SEM of 3 independent biological replicates, each performed in triplicate. **P < 0.01 by one sample t-test. D. Plaque assay to assess in vitro growth of the TgENT1-KD strain (left) along with parental control (right) in HFF cells, in the presence and absence of ATc.
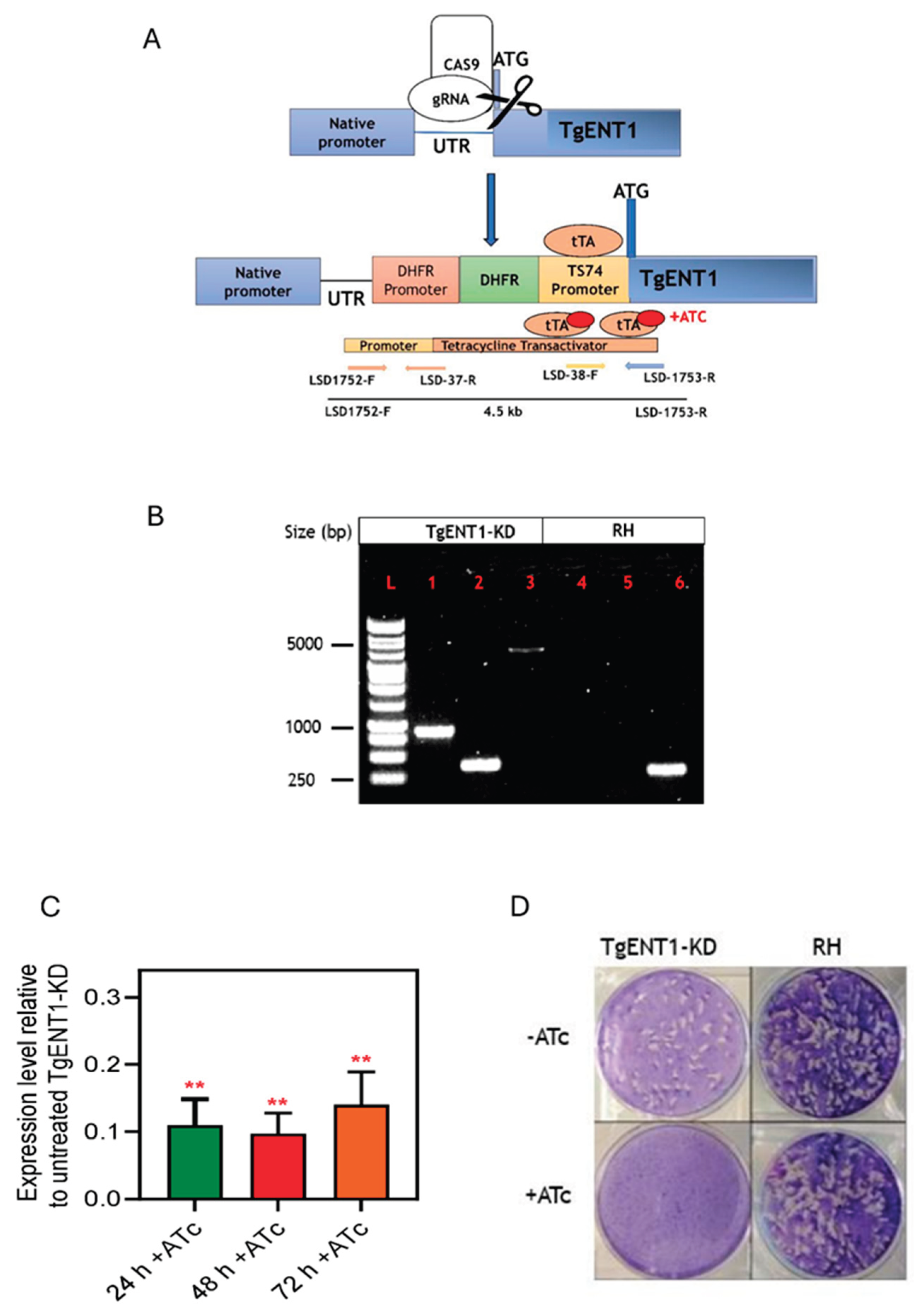
Figure 5.
Construction of Leishmania mexicana cell lines expressing the TgENT1 minigene. A. pNUS-HcN vector digestion by NdeI and XhoI restriction enzymes run on 1% agarose gel; band size was 6.3 kb. (L) 1 kb DNA Ladder (Promega); (1): Negative control water; (2): digested pNUS-HcN with the size band (6.3 kb). B. PCR confirmation of the presence of TgENT1 minigene in LmexNT3-KO clones grown out after transfection with pNUS/TgENT1 and selection on G-418; TgENT1-specifc forward primer HDK-1739 and reverse primer (3’ vector sequence) HDK-340. (L) 1 kb DNA Ladder (Promega); (1): Clone1; (2): clone2; (3): clone3; (4): clone 4 (~2.2 kb). C. The expression levels of TgENT1 in Lmex-NT3-KO and compared to the control (LmexNT3-KO) determined by qRT-PCR; expression levels in three independent clones is shown.
Figure 5.
Construction of Leishmania mexicana cell lines expressing the TgENT1 minigene. A. pNUS-HcN vector digestion by NdeI and XhoI restriction enzymes run on 1% agarose gel; band size was 6.3 kb. (L) 1 kb DNA Ladder (Promega); (1): Negative control water; (2): digested pNUS-HcN with the size band (6.3 kb). B. PCR confirmation of the presence of TgENT1 minigene in LmexNT3-KO clones grown out after transfection with pNUS/TgENT1 and selection on G-418; TgENT1-specifc forward primer HDK-1739 and reverse primer (3’ vector sequence) HDK-340. (L) 1 kb DNA Ladder (Promega); (1): Clone1; (2): clone2; (3): clone3; (4): clone 4 (~2.2 kb). C. The expression levels of TgENT1 in Lmex-NT3-KO and compared to the control (LmexNT3-KO) determined by qRT-PCR; expression levels in three independent clones is shown.
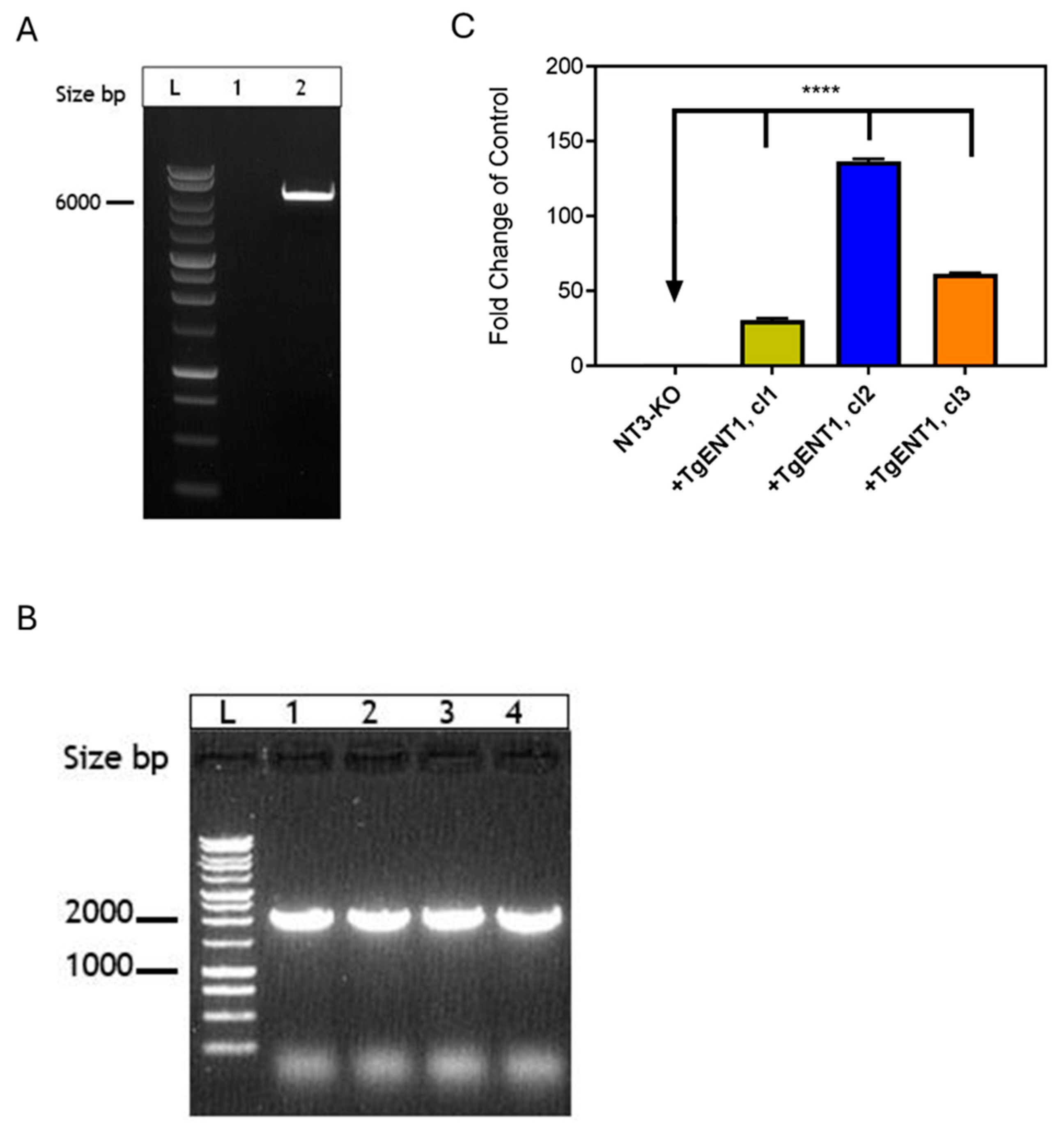
Figure 6.
Pyrimidine transport in tachyzoites of the TgENT2 and TgENT3 knockout strains. A. Time course of 0.1 µM [3H]-uridine transport by the indicated strains in the absence (0, 3, 6 and 9 min) or presence (9 min) of 1 mM unlabeled uridine. B/D. Dose-response inhibition of unlabeled uridine or of 100 µM of other purines and pyrimidines of 0.1 µM [3H]-uridine transport by TgENT2-KO and TgENT3-KO cells respectively. Inset: conversion of the uridine inhibition data to a Michaelis-Menten saturation curve. C./E. Dose-dependent inhibition of 0.1 µM [3H]-uridine transport by TgENT2-KO and TgENT3-KO cells by uracil respectively, inosine and adenosine; expressed as % of uninhibited control. F. Timecourse of 0.1 µM [3H]-uridine transport by TgENT2/3dKO and RH tachyzoites. G. As frame F but 0.1 µM [3H]-uracil transport.
Figure 6.
Pyrimidine transport in tachyzoites of the TgENT2 and TgENT3 knockout strains. A. Time course of 0.1 µM [3H]-uridine transport by the indicated strains in the absence (0, 3, 6 and 9 min) or presence (9 min) of 1 mM unlabeled uridine. B/D. Dose-response inhibition of unlabeled uridine or of 100 µM of other purines and pyrimidines of 0.1 µM [3H]-uridine transport by TgENT2-KO and TgENT3-KO cells respectively. Inset: conversion of the uridine inhibition data to a Michaelis-Menten saturation curve. C./E. Dose-dependent inhibition of 0.1 µM [3H]-uridine transport by TgENT2-KO and TgENT3-KO cells by uracil respectively, inosine and adenosine; expressed as % of uninhibited control. F. Timecourse of 0.1 µM [3H]-uridine transport by TgENT2/3dKO and RH tachyzoites. G. As frame F but 0.1 µM [3H]-uracil transport.
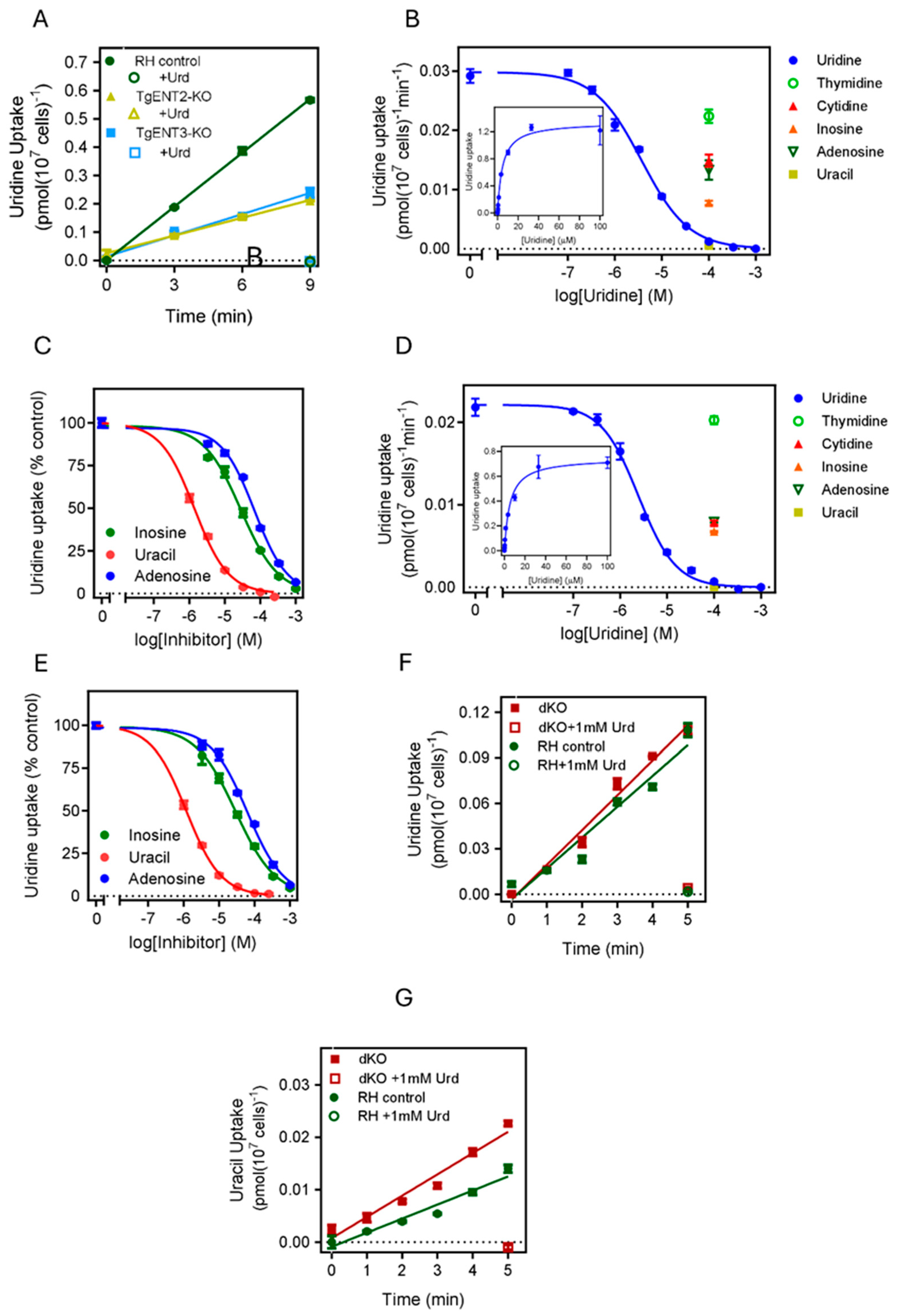
Figure 7.
TgENT1 is a pyrimidine transporter. A/B/C. Transport of 0.1 µM [3H]-uridine, 0.1 µM of [3H]-uracil or [3H]-tryptophan respectively, by T. gondii tachyzoites engineered with promotor replacement for the suppression of TgENT1 expression in the presence of anhydrous tetracycline (ATc). Cells were cultured in the presence of ATc for either 24 h or 48 h prior to the uptake experiment and then compared to cells cultured in the absence of ATc (green circles); for each category uptake was also measured at 9 min of incubation in the presence of 1 mM unlabeled substrate (uridine, Urd). D. Uptake of 0.1 µM of [3H]-uracil by L. mexicana NT3-KO promastigotes expressing the TgENT1 minigene. Cas9 is the parental strain of NT3-KO, i.e. does express the LmexNT3 nucleobase transporter. Three independent clones of NT3/TgENT1 were assessed, each in triplicate. E. As frame D but with 0.1 µM [3H]-adenine.
Figure 7.
TgENT1 is a pyrimidine transporter. A/B/C. Transport of 0.1 µM [3H]-uridine, 0.1 µM of [3H]-uracil or [3H]-tryptophan respectively, by T. gondii tachyzoites engineered with promotor replacement for the suppression of TgENT1 expression in the presence of anhydrous tetracycline (ATc). Cells were cultured in the presence of ATc for either 24 h or 48 h prior to the uptake experiment and then compared to cells cultured in the absence of ATc (green circles); for each category uptake was also measured at 9 min of incubation in the presence of 1 mM unlabeled substrate (uridine, Urd). D. Uptake of 0.1 µM of [3H]-uracil by L. mexicana NT3-KO promastigotes expressing the TgENT1 minigene. Cas9 is the parental strain of NT3-KO, i.e. does express the LmexNT3 nucleobase transporter. Three independent clones of NT3/TgENT1 were assessed, each in triplicate. E. As frame D but with 0.1 µM [3H]-adenine.

Table 1.
ENT genes.
| T gondii Gene ID | Name | bp (no introns) | a.a. | TMD* | Qian et al | |
|---|---|---|---|---|---|---|
| TGME49 | TGGT1 | |||||
| 244440 | 244440 | TgAT1 | 1389 | 462 | 10 | TgAT1 |
| 288540 | 288540 | TgENT1 | 2091 | 696 | 10 | TgNT1 |
| 500147 | 359630 | TgENT2 | 2007 | 668 | 10 | TgNT3 |
| 233130 | 233130 | TgENT3 | 1596 | 531 | 10 | TgNT2 |
Data according to ToxoDB release 68, accessed 10 December 2025. *, https://services.healthtech.dtu.dk/services/TMHMM-2.0/.
Table 2.
Kinetic parameters of uridine transport in three cell lines.
| TgENT2-KO | TgENT3-KO | RH1 | ||
|---|---|---|---|---|
| Km | 5.29 ± 0.58 | 3.78 ± 0.92 | 3.34 ± 0.82 | µM |
| Vmax | 0.025 ± 0.002 | 0.011 ± 0.002 | 0.020 ± 0.004 | pmol(107 cells)-1min-1 |
| Uracil Ki | 0.96 ± 0.17 | 1.06 ± 0.14 | 1.15 ± 0.07 | µM |
| Inosine Ki | 32.2 ± 3.3 | 37.7 ± 3.82 | 28.2 ± 4.1 | µM |
| Adenosine Ki | 77.4 ± 10.7 | 58.9 ± 1.10* | 111 ± 6 | µM |
All values are average ± SEM for n ≥ 3. *, P = 0.002 in unpaired t-test, relative to RH; all other values were not significantly different from the parental control. 1values for the control RH cell line were taken from [13] for comparison.
Table 3.
In vitro drug screening and HFF cytotoxicity using pyrimidine nucleobase and nucleoside analogous on different cell lines of T. gondii RH intracellular tachyzoites, TgAT1-KO, TgENT1-KO, TgENT2-KO and TgENT2/3dKO.
Table 3.
In vitro drug screening and HFF cytotoxicity using pyrimidine nucleobase and nucleoside analogous on different cell lines of T. gondii RH intracellular tachyzoites, TgAT1-KO, TgENT1-KO, TgENT2-KO and TgENT2/3dKO.
| ID | RH | TgAT1-KO | TgENT2-KO | TgENT3-KO | TgENT2/3dKO | HFF cytotoxicity | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EC50 ± SEMa | EC50 ± SEM | P b | EC50 ± SEM | P b | EC50 ± SEM | P b | EC50 ± SEM | P b | EC50 Cytostatica |
EC50 Cytocidala |
SI | |
| 5-FU | 1.13 ± 0.16 | 0.70 ± 0.21 | 0.17 | 1.36 ± 0.26 | 0.49 | 0.82 ± 0.24 | 0.34 | 0.51 ± 0.08 | 0.03 | 2.49 ± 0.43 | 3074 ±339 | 2720 |
| 5-FUrd | 0.45 ± 0.02 | 0.89 ± 0.42 | 0.35 | 0.33 ± 0.02 | 0.02 | 0.36 ± 0.03 | 0.07 | 0.29 ± 0.002 | 0.003 | 0.11 ± 0.10 | 20.4 ± 1 | 45 |
| 5-F-2’dUrd | 0.67 ± 0.07 | 0.41 ± 0.14 | 0.16 | 0.84 ± 0.07 | 0.14 | 0.56 ± 0.07 | 0.28 | 0.40 ± 0.06 | 0.04 | 0.19 ± 0.05 | 1478 ± 512 | 2205 |
| Sulfadiazine | 12.4 ± 1.08 | 3.95 ± 0.29 | 0.0003 | 7.15 ± 0.19 | 0.01 | 8.12 ± 0.22 | 0.02 | 5.74 ± 0.29 | 0.004 | -- | -- | -- |
| PAO | -- | -- | -- | -- | -- | -- | 0.06 ± 0.003 | |||||
All EC50 values are averages and SEM of at least three independent experiments, given in µM. SI, selectivity index (HFF cytocidal EC50/RH EC50). PAO. aValues taken from [13]. bP value is based on an unpaired Student t-test (RH EC50 versus knockout TgENT gene EC50).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



