
Submitted:
27 January 2026
Posted:
30 January 2026
You are already at the latest version
Abstract
Increased left ventricular mechanical dispersion is an important echocardiographic indicator of ventricular dyssynchrony and enhanced arrhythmogenesis, particularly in patients with heart failure. Precise structural and molecular aberrations explaining increased mechanical dispersion remain incompletely understood, warranting further investigation. Various ion channel, gap junction and calcium handling protein remodeling mechanisms have all been studied in connection with increased mechanical dispersion, offering however only partial solutions. This author debates that microtubular dysfunction may play distinctive role in left ventricular dyssynchrony and enhanced arrhythmogenesis observed in patients with heart failure, highlighted by increased mechanical dispersion. Testing this hypothesis may be technically challenging, however not impossible and therefore should be encouraged. In case of success potential therapeutic benefits could be diverse, including prevention of malignant arrhythmia, delay of heart failure progression, and promotion of LV reverse remodeling.
Keywords:
ventricular arrhythmogenesis
; mechanical dispersion
; heart failure with preserved ejection fraction (HFpEF)
; microtubular hypothesis
Introduction
Mechanical Dispersion (MD) – the echocardiographic indicator of left ventricular (LV) dyssynchrony – represents a standard deviation of segmental time-to-peak strain measurements. The latter strain is further defined as an interval between the onset of electrocardiographic QRS complex and maximal fractional shortening of contracting ventricular segment (measured in milliseconds). [1] Increased LV MD is associated with higher odds of ventricular arrythmia as well as sudden cardiac death (SCD) in patients with heart failure with reduced ejection fraction (HFrEF), defined as left ventricular ejection fraction (LVEF) < 40%, [2] and in patients with commonly preceding heart failure with preserved ejection fraction (HFpEF), defined as LVEF ≥50%. [3] Two-dimensional (2D) LV global longitudinal strain (GLS) obtained with the use of 2D transthoracic speckle-tracking echocardiography is another sensitive parameter capable of detecting early or subclinical LV systolic dysfunction (peak GLS depicts the relative length change of the LV myocardium between end-diastole and end-systole). [4,5,6] Importantly, hypertrophic cardiomyopathy patients with low-normal LVEF and reduced GLS were shown to have worse cardiac outcomes. [7] However, utilized echocardiographic parameters tend to differ in their ability to predict arrhythmia under various circumstances. For example, despite LVEF, GLS and MD all being reflective of the LV function, MD was shown to be superior to the GLS in predicting LV remodeling (reversal of abnormal left ventricular shape and volume) and seems to have an edge in predicting the ventricular arrhythmogenesis. [8,9] Notably, GLS was shown to be predictive of cardiac arrest in patients with LVEF >50%, whereas MD was associated with cardiac arrest in individuals with both LVEF >50 and variable grade LV diastolic dysfunction. [10] Male sex, LV-GLS, and MD were shown to predict sudden arrhythmic death, (or SAD – a subset of SCD), but not fibrosis (known arrhythmia risk-factor). However, after adjustment for age, sex, and LVEF, MD was the only significant predictor of SAD. Remarkably, despite the association between MD and fibrosis, MD was a better predictor of SAD compared to total fibrosis on histology. [11] Notwithstanding, MD measurements may also be predictive of heart failure or arrhythmogenesis in patients with Fabry’s disease (a rare X-linked storage disorder caused by deficiency of the lysosomal enzyme α-galactosidase A) and non-ischemic (not related to coronary arterial disease, or CAD) cardiomyopathy, respectively. [12,13] Remarkably, MD was shown to have high predictive value independent of the concomitant EKG reads. In fact, data obtained from over 1400 people with no overt cardiac disease showed that longer corrected QT interval was related to increased MD and better longitudinal strain; in a population-based environment, QRS was not associated with MD, implying that echocardiography-based dyssynchrony did not largely overlap with ECG-based dyssynchrony. [14]
Reflective of this discussion, it feels only fair to state that at least in some patients with heart failure increased MD exhibits significant predictive strength independent of other major proarrhythmogenic factors, such as fibrosis, abnormal longitudinal strain or EKG-based dyssynchrony. This author suggests there might be an additional and previously not explicitly described molecular link marrying structural and electrical aspects of increased MD in various scenarios of heart failure. If so, what could be that missing link?
Understanding the Relationship Between Electrical Excitation and Mechanical Force.
The underlying relationship between cardiac force build-up and ventricular synchrony (reflected by change in MD) has not been completely understood. Various ion channels, gap junction and calcium handling protein remodeling mechanisms have been studied with regards to increased MD. [15,16]
The onset of local myofiber shortening in viable ventricular wall follows the local electrical depolarization by as much as tens of milliseconds. This electromechanical delay, or EMD, is comprised of the intrinsic latent period (spreading from membrane depolarization to myofilament engagement during myocyte excitation-contraction coupling) and an additional EMD part reflective of the myofiber mechanical load variations in the intact heart (e.g., transient over-stretch, volume overload). Notably, during sinus rhythm, the EMD distribution is longer at the base and epicardium compared to apex and endocardium, respectively. [17] Conversely, cardiac mechanoelectrical feedback mechanism can alter the EMD duration: abnormal hemodynamic chamber loading, or regional changes in ventricular muscle synchrony affect the pressure and stretch before or during electrical activation, in some cases naturally leading to arrhythmia. [18] In theory, virtually any structural or functional aberrations occurring within failing cardiomyocytes can potentially affect EMD duration thus MD measurements. Furthermore, mavacamten (a recently developed cardiac myosin inhibitor) was shown to decrease LVEF and LV deformation but improve LV MD as well as symptoms in patients with hypertrophic obstructive cardiomyopathy; none of the patients experienced life-threatening arrhythmias. [19]
Effects of Impaired Coronary Circulation on Mechanical Dispersion. Focus on HFpEF.
HFpEF is a unique case of heart failure and can be divided in several phenotypes, depending on predominant underlying pathophysiological mechanism (e.g., metabo-inflammatory, microvascular, ventricular stiffening, and cardiac predominant). [20] Interestingly, more recent evidence also links the epicardial adipose tissue inflammation to HFpEF onset. [21] With that being said, epidemiological data describes cardiometabolic HFpEF as the most prevalent HFpEF phenotype. Furthermore, cardiometabolic alterations were shown to be more tightly associated with the development of HFpEF than HFrEF. [22] While CAD and cardiac metabolism are immensely complex and are beyond the scope of this article, it is useful to briefly describe the sequence of related proarrhythmic events occuring within metabolically failing hearts to underline their connection to structural cell aberrations (e.g., microtubular dysfunction).
Briefly, CAD, either epicardial or microvascular, is a known prerequisite for metabolic aberrations occurring in failing cardiomyocytes including mitochondrial dysfunction, poor metabolic flexibility (ability to readily switch the fuel sources such as carbohydrates and fats) and energy reserve (heart’s capacity to respond to increased workload). [23,24,25,26,27] Resultant deficit of intracellular ATP (adenosine triphosphate) and creatine pool [28] in combination with toxic reactive oxygen species disrupt normal functionality of organelles and various ATP-dependent channels. [29] Chronic myocardial ischemia triggers a sophisticated electro-mechanical remodeling cascade pleiotropically affecting the action potential duration (APD) of failing cardiomyocytes. [30] At earlier stages, ischemia-related decrease in ATP / ADP (adenosine diphosphate) ratio tends to prolong opening of sarcolemmal K+ channels and promote K+ efflux thus shortening of the APD. [31] However, the same chronic ischemia also impairs functionality of sarcoplasmic reticulum (SR) – bound phospholamban and sarcoplasmic/endoplasmic reticulum Ca2+-ATPase, disrupting normal intracellular calcium flows and promoting cytoplasmic calcium build-up. Resultant calcium build-up can delay normal relaxation (and repolarization) of cardiac muscle effectively enhancing arrhythmogenesis; to counteract the detrimental effects of Ca2+ build-up the repolarization reserve is mobilized (e.g., upregulation of reverse mode Na+/Ca+ exchange and suppression of L-type Ca2+ channels, respectively). [32,33,34] Unfortunately, earlier or later ischemia overwhelms compensatory mechanisms leading to inward ion current dominance, increase in calcium transients (rapid increase and following decrease in the concentration of cytoplasmic calcium ions) often leading to APD prolongation. As a result, highly proarrhythmogenic early after depolarizations (EADs) frequently emerge within failing cardiomyocytes, heralding the possibility of malignant ventricular arrhythmia. [35,36,37] Notwithstanding, other less common proarrhythmogenic events can also occur in failing cardiomyocyte (e.g., EADs with shortened APD in atrial fibrillation, hybrid EADs with delayed afterdepolarizations, etc). [38]
Microtubules.
Microtubules are cytoskeletal structures, made of α-tubulin and β-tubulin heterodimers with an outer diameter of ≈25 nm and a length from <1 and to >100 micrometers. In adult cardiomyocytes, cortical, interfibrillar and perinuclear microtubules can be defined. [39] Importantly, microtubular assembly and microtubule-dependent kinesin transport are all energy-depending processes (relying on GTP (guanosine triphosphate) and ATP availability, respectively). Microtubules are ubiquitous structural components which also play a pivotal role in electromechanical properties of cardiac tissues such as signal transduction and proteostasis (dynamic regulation of various protein gradients), [40] stabilization of cytosolic vibrations and calcium waves, [41,42,43] regulation of tissue viscoelasticity (or rigidity), [44] vesicle and organelle trafficking, as well as autophagy (critical processes for ion channel/pump functionality). [45,46,47] In addition, microtubules synchronize mitochondrial transport with myofibril formation during muscle development, define proper organization of the SR and intercalated disks as they interact with myofibrils, and exhibit characteristic microtubule buckling properties (achieved through rapid contraction and posterior relaxation) – an important mechanism shown to be responsible for ~ 20% of the cardiomyocyte’s contractile force. [40,48,49,50]
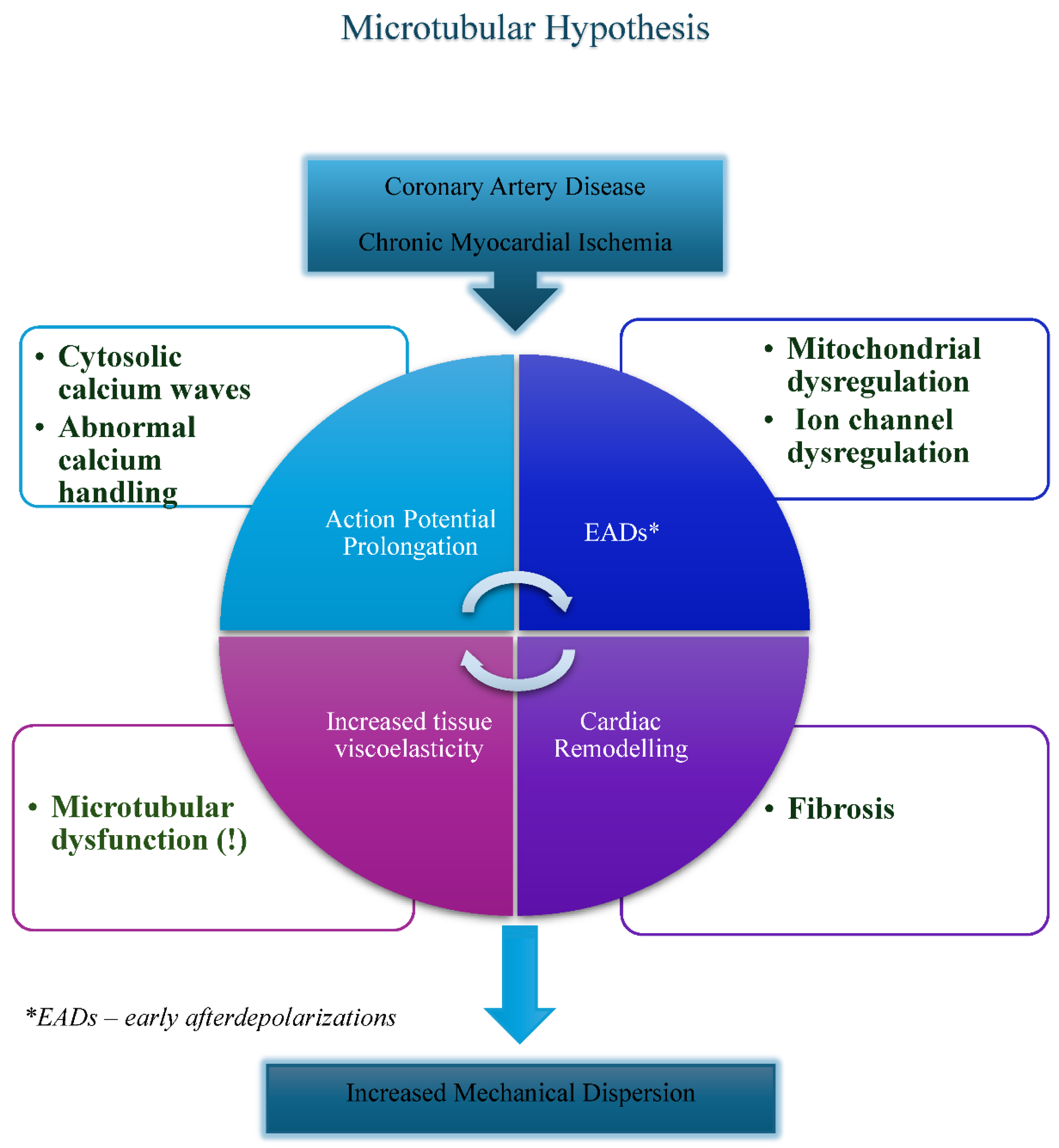
Relevantly, drastic increase in density and stability of polymerized microtubules (largely related to post-translational modifications) has been described in patients with ischemic heart disease, cardiac hypertrophy, and dilated cardiomyopathy. These changes facilitate interaction between microtubules, microtubule-associated proteins and intermediate filaments thus often translating into increased tissue rigidity, disruption of normal excitation-contraction coupling and altered mitochondrial functionality (Figure 1). [51,52] Despite available data, precise manipulation of microtubular function has been historically very challenging task. Structural and functional regulation of microtubules is highly complex; important post-translational modifications defining microtubular polarity, flexibility and longevity include mechanical stress-induced a-tubulin tyrosination / detyrosination (regulation through C-terminal tyrosine) and acetylation / deacetylation (often involving Lysine 40 on the inner surface of α-tubulin). Interestingly, cardiac gap junction microtubule-Connexin-43 regulatory link was shown to suppress arrhythmias and cardiac fibrosis in mice with Duchenne muscular dystrophy mice. [53] Nevertheless, full therapeutic potential of the aforementioned regulatory mechanisms is yet to be understood. [54,55] To date, clinical applications of microtubule-targeting agents in cardiovascular field have been limited, often focusing on the use of colchicine in atrial fibrillation and CAD. [56] To author’s best knowledge, microtubular studies rigorously investigating microtubular effects on MD have not been widely published.
Microtubular Hypothesis of Increased Mechanical Dispersion.
As described above, cardiac microtubules are directly involved in several paramount intracellular processes such as cellular growth, metabolism and aging. Microtubules are expected to affect the structural and electromechanical properties of left ventricle, namely its stiffness, contractility and propensity to arrhythmia. According to this author’s hypothesis, microtubular dysfunction induced by ischemic (but also possibly non-ischemic) causes may enhance ventricular arrhythmogenesis in patients with cardiometabolic HFpEF, reflected by the increased MD. [57,58,59] Microtubules seem to satisfy the functional criteria of universal “carabiners” capable of tying characteristic molecular, structural and electromechanical processes occurring within metabolically compromised cardiomyocytes, therefore regulating the ventricular wall rigidity, anatomy of electromechanical coupling and MD. [60,61] Co-existent factors such as fibrosis commonly present in failing hearts, tend to independently affect ventricular mechanics and action potential duration. As a result, oftentimes, the metabolically remodeled, stiff and therefore highly arrhythmogenic ventricular wall gets formed (Figure 1). [62]
Conclusion
Increased MD is a powerful indicator of enhanced arrhythmogenesis in patients with heart failure therefore it is highly relevant that we understand molecular mechanisms of its regulation. Presently, causative data pertaining to microtubular dysfunction’s role in abnormal MD remains at best tangential. Developing cardiotropic microtubular therapy is anticipatorily challenging due to their ubiquitous cellular distribution, highly dynamic regulatory mechanisms and limited understanding of therapeutic or toxic effects. On the other hand, in case of experimental success achieved therapeutic benefits may be quite remarkable, encompassing prevention of malignant arrhythmia, slow down of heart failure progression, and promotion of LV reverse remodeling. Consequently, further investigation of the MD-directed effects of microtubular inhibitors (e.g., colchicine) or experimental post-translational modifying agents could be the reasonable next step in the proposed research.
Statements and Declarations.
The author declares no conflict of interest.
The author declares no acknowledgements.
The author received no external funding.
The authors did not use generative AI or AI-assisted technologies in the development of this manuscript.
Human Ethics and Consent to Participate declarations: not applicable.
References
- Mechanical Dispersion Discriminates between Arrhythmic and Non-Arrhythmic Sudden Death: From the POST SCD Study. Lionel Tastet, Satvik Ramakrishna, Lisa J. Lim, Dwight Bibby, Jeffrey E. Olgin, Andrew J. Connolly, Ellen Moffatt, Zian H. Tseng, Francesca N. Delling. 4, April 2024, JACC: Clinical Electrophysiology, Vol. 10, pp. 771-773.
- LV mechanical dispersion as a predictor of ventricular arrhythmia in patients with advanced systolic heart failure. . Banasik, G., Segiet, O., Elwart, M. et al. 2016, Herz, Vol. 41, pp. 599–604.
- Sudden cardiac death in heart failure with preserved ejection fraction: an updated review. . Wu, SJ., Hsieh, YC. 2022, Int J Arrhythm, Vols. 23, 7. [CrossRef]
- Recommendations for Cardiac Chamber Quantification by Echocardiography in Adults: An Update from the American Society of Echocardiography and the European Association of Cardiovascular Imaging. Lang, Roberto M. et al. 1, Journal of the American Society of Echocardiography., Vol. 28.
- Prognostic Relevance and Lower Limit of the Reference Range of Left Ventricular Global Longitudinal Strain: A Clinical Validation Study. . Morris, D, Hung, C, Biering-Sørensen, T. et al. s.l. : 18, 2025 May, J Am Coll Cardiol Img., Vol. 5, pp. 525–536.
- Myocardial Strain Imaging: Theory, Current Practice, and the Future. . Smiseth, O, Rider, O, Cvijic, M. et al. 3, 2025 Mar, J Am Coll Cardiol Img., Vol. 18, pp. 340–381. [CrossRef]
- PROGNOSTIC VALUE OF LEFT VENTRICULAR GLOBAL LONGITUDINAL STRAIN IN PATIENTS WITH HYPERTROPHIC CARDIOMYOPATHY AND LOW-NORMAL LEFT VENTRICULAR EJECTION FRACTION. . Laenens, D, Kamperidis, V, Tjahjadi, C. et al. 2024 Apr,, JACC., Vol. 83 (13_Supplement) 620. [CrossRef]
- Left Ventricular Mechanical Dispersion and Its Value in Combination With Global Longitudinal Strain to Predict Reverse Remodeling in Patients With Heart Failure With Reduced Ejection Fraction. . Liu F, Li R, Liu Y, Ma F, Yang H, Yang Q, Yang X, Yu T, Zeng H, Tang J, Wang H. 2025 Jul 15, J Am Heart Assoc., Vol. 14(14):e040652. . [CrossRef]
- Prediction of Ventricular Arrhythmias With Left Ventricular Mechanical Dispersion: A Systematic Review and Meta-Analysis. . Kawakami H, Nerlekar N, Haugaa KH, Edvardsen T, Marwick TH. (2 Pt 2), 2020 Feb, JACC Cardiovasc Imaging., Vol. 13.
- DIASTOLIC DYSFUNCTION AND MECHANICAL DISPERSION BY SPECKLE TRACKING ECHOCARDIOGRAPHY IN PREDICTING CARDIAC ARREST RISK. Ozbay, B, Saba, S, Villanueva, F. et al. 2024 Apr, JACC., Vol. 83 (13_Supplement) 1443. [CrossRef]
- Mechanical Dispersion Discriminates Between Arrhythmic and Nonarrhythmic Sudden Death: From the POST SCD Study. . Tastet, L, Ramakrishna, S, Lim, L. et al. 4, 2024 Apr, J Am Coll Cardiol EP. , Vol. 10, pp. 771–773.
- Mechanical dispersion in Fabry disease assessed with speckle tracking echocardiography. . Cianciulli TF, Saccheri MC, Rísolo MA, Lax JA, Méndez RJ, Morita LA, Beck MA, Kazelián LR. s.l. : 37, Feb 2020, Echocardiography, Vol. 2, pp. 293-301. [CrossRef]
- Association between ventricular arrhythmias and myocardial mechanical dispersion assessed by strain analysis in patients with nonischemic cardiomyopathy. Kosiuk J, Dinov B, Bollmann A, Koutalas E, Mussigbrodt A, Sommer P, Arya A, Richter S, Hindricks G, Breithardt OA. 12, Dec 2015, lin Res Cardiol., Vol. 104, pp. 1072-7. [CrossRef]
- Left ventricular myocardial deformation pattern, mechanical dispersion, and their relation with electrocardiogram markers in the large population-based STANISLAS cohort: insights into electromechanical coupling. Mario Verdugo-Marchese, Stefano Coiro, Christine Selton-Suty, Masatake Kobayashi, Erwan Bozec, Zohra Lamiral, Clément Venner, Faiez Zannad, Patrick Rossignol, Nicolas Girerd, Olivier Huttin. s.l. : 21, Nov 2020, European Heart Journal - Cardiovascular Imaging., Vol. 11, pp. 1237–1245. [CrossRef]
- Left ventricular repolarization heterogeneity as an arrhythmic substrate in heart failure. . FG., Akar. 2, 2010 Apr, Minerva Cardioangiol., Vol. 58, pp. :205-12.
- Molecular determinants of cardiac transient outward potassium current (I(to)) expression and regulation. Niwa N, Nerbonne JM. 1, 2010 Jan, J Mol Cell Cardiol. , Vol. 48, pp. 12-25. [CrossRef]
- Distribution of electromechanical delay in the heart: insights from a three-dimensional electromechanical model. . Gurev V, Constantino J, Rice JJ, Trayanova NA. 3, 2010 Aug 4, Biophys J., Vol. 99, pp. 745-54. [CrossRef]
- Biomechanics of cardiac electromechanical coupling and mechanoelectric feedback. . Pfeiffer ER, Tangney JR, Omens JH, McCulloch AD. (2):021007, 2014 Feb, J Biomech Eng., Vol. 136. [CrossRef]
- Mavacamten reduces left ventricular mechanical dispersion in hypertrophic obstructive cardiomyopathy, , . B Ozbay, N Venkatesan, T C Wong, L E Sade. 1, January 2025, jeae333.192, European Heart Journal - Cardiovascular Imaging, Vol. 26. [CrossRef]
- Heart Failure With Preserved Ejection Fraction: JACC Scientific Statement. Borlaug, B, Sharma, K, Shah, S. et al. 18, 2023 May, JACC., Vol. 81, pp. 1810–1834. [CrossRef]
- The epicardial adipose inflammatory triad: coronary atherosclerosis, atrial fibrillation, and heart failure with a preserved ejection fraction. M., Packer. Nov 2018, Eur J Heart Fail. , Vol. 20(11), pp. 1567-1569. [CrossRef]
- Cardiometabolic heart failure with preserved ejection fraction: from molecular signatures to personalized treatment. Gorica, E., Geiger, M.A., Di Venanzio, L. et al. 265, 2025, Cardiovasc Diabetol., Vols. 24,. [CrossRef]
- Microvascular and lymphatic dysfunction in HFpEF and its associated comorbidities. . Cuijpers I, Simmonds SJ, van Bilsen M, Czarnowska E, González Miqueo A, Heymans S, Kuhn AR, Mulder P, Ratajska A, Jones EAV, Brakenhielm E. May 25 2020 , Basic Res Cardiol, Vol. 115(4):39. [CrossRef]
- Chronic Myocardial Ischemia Leads to Loss of Maximal Oxygen Consumption and Complex I Dysfunction. Schipper DA, Palsma R, Marsh KM, O’Hare C, Dicken DS, Lick S, Kazui T, Johnson K, Smolenski RT, Duncker DJ, Khalpey Z. Oct 2017, Ann Thorac Surg, Vol. 104(4), pp. 1298-1304. [CrossRef]
- Metabolic Flexibility of the Heart: The Role of Fatty Acid Metabolism in Health, Heart Failure, and Cardiometabolic Diseases. Actis Dato V, Lange S, Cho Y. Jan 19 2024 , Int J Mol Sci., Vol. 25(2), p. 1211. [CrossRef]
- Epicardial and microvascular angina or atypical chest pain: differential diagnoses with cardiovascular magnetic resonance. Baggiano A, Guglielmo M, Muscogiuri G, Guaricci AI, Del Torto A, Pontone G. Jun 2020, Eur Heart J Suppl., Vol. 22(Suppl E), pp. E116-E120. [CrossRef]
- The microtubule cytoskeleton in cardiac mechanics and heart failure. . Caporizzo, M.A., Prosser, B.L. 2022, Nat Rev Cardiol, Vol. 19, pp. 364–378. [CrossRef]
- Impaired Myocardial Energetics Causes Mechanical Dysfunction in Decompensated Failing Hearts. Lopez R, Marzban B, Gao X, Lauinger E, Van den Bergh F, Whitesall SE, Converso-Baran K, Burant CF, Michele DE, Beard DA. 2020, Function (Oxf)., Vol. 1(2):zqaa018. [CrossRef]
- Mechanisms of Chronic Metabolic Stress in Arrhythmias. Gowen BH, Reyes MV, Joseph LC, Morrow JP. 2020 Oct 19, Antioxidants (Basel)., Vol. 9(10):1012. [CrossRef]
- Repolarization Reserve and Action Potential Dynamics in Failing Myocytes. Fu JD, Laurita KR. 2018 Feb, Circ Arrhythm Electrophysiol. , Vol. 1(2):e006137. [CrossRef]
- Mitochondrial Dysfunction as Substrate for Arrhythmogenic Cardiomyopathy: A Search for New Disease Mechanisms. van Opbergen CJM, den Braven L, Delmar M, van Veen TAB. Dec 10 2019 , Front Physiol. , Vol. 10:1496. [CrossRef]
- Modulation of cardiac contractility by the phospholamban/SERCA2a regulatome. . Kranias EG, Hajjar RJ. s.l. : 110, 2012 Jun 8, Circ Res., Vol. 12, pp. 1646-60.
- The Control of Diastolic Calcium in the Heart: Basic Mechanisms and Functional Implications. Eisner DA, Caldwell JL, Trafford AW, Hutchings DC. 3, 2020 Jan 31, Circ Res., Vol. 126, pp. 395-412. [CrossRef]
- Physiology of intracellular calcium buffering. ;):. . Eisner D, Neher E, Taschenberger H, Smith G. 4, 2023 Oct 1, Physiol Rev., Vol. 103, pp. 2767-2845. [CrossRef]
- Plasticity of sarcolemmal KATP channel surface expression: relevance during ischemia and ischemic preconditioning. Yang HQ, Foster MN, Jana K, Ho J, Rindler MJ, Coetzee WA. 2016 Jun 1, Am J Physiol Heart Circ Physiol., Vols. 310(11):H1558-66. [CrossRef]
- Early afterdepolarizations and cardiac arrhythmias. Weiss JN, Garfinkel A, Karagueuzian HS, Chen PS, Qu Z. s.l. : 7(12), Dec 2010, Heart Rhythm., pp. 1891-9. [CrossRef]
- Arrhythmogenic consequences of intracellular calcium waves. Xie LH, Weiss JN. Sep 2009 , Am J Physiol Heart Circ Physiol., Vol. 297(3), pp. H997-H1002. [CrossRef]
- Calcium Handling Defects and Cardiac Arrhythmia Syndromes. . Kistamás K, Veress R, Horváth B, Bányász T, Nánási PP, Eisner DA. 72, Feb 25 2020, Front Pharmacol, Vol. 11. [CrossRef]
- Targeting Microtubules for the Treatment of Heart Disease. . Warner EF, Li Y, Li X. 11, 2022 May 27, Circ Res, Vol. 130, pp. 1723-1741. [CrossRef]
- Cardiac microtubules in health and heart disease. Caporizzo MA, Chen CY, Prosser BL. Nov 2019, Exp Biol Med (Maywood), Vol. 244(15), pp. 1255-1272. [CrossRef]
- Quantitative tests reveal that microtubules tune the healthy heart but underlie arrhythmias in pathology. Joca HC, Coleman AK, Ward CW, Williams GSB. Apr 2020 , J Physiol. , Vol. 598(7), pp. 1327-1338. [CrossRef]
- Microtubule-Dependent Mitochondria Alignment Regulates Calcium Release in Response to Nanomechanical Stimulus in Heart Myocytes. Miragoli, Michele et al. Issue 1, 2016, Cell Reports, Vol. Volume 14, pp. 140 - 151.
- Microtubule disruption by colchicine reversibly enhances calcium signaling in intact rat cardiac myocytes. Kerfant BG, Vassort G, Gómez AM. Apr 13 2001, Circ Res., Vol. 88(7), pp. E59-65. [CrossRef]
- Cardiomyocyte Microtubules: Control of Mechanics, Transport, and Remodeling. Uchida K, Scarborough EA, Prosser BL. Feb 10 2022, Annu Rev Physiol., Vol. 84, pp. 257-283. [CrossRef]
- Microtubule plus-end tracking proteins: novel modulators of cardiac sodium channels and arrhythmogenesis. Marchal GA, Galjart N, Portero V, Remme CA. 2023 Jul 4, Cardiovasc Res., Vol. 119(7), pp. 1461-1479. [CrossRef]
- Microtubules: From understanding their dynamics to using them as potential therapeutic targets. Y., Ilan. 2019 Jun, J Cell Physiol., Vol. 234(6), pp. 7923-7937. [CrossRef]
- Microtubules: Evolving roles and critical cellular interactions. Logan CM, Menko AS. Nov 2019 , Exp Biol Med (Maywood), Vol. 244(15, pp. 1240-1254. [CrossRef]
- Characterization of microtubule buckling in living cells. . Pallavicini C, Monastra A, Bardeci NG, Wetzler D, Levi V, Bruno L. 6, 2017 Sep, Eur Biophys J., Vol. 46, pp. 581-594. [CrossRef]
- Detyrosinated microtubules buckle and bear load in contracting cardiomyocytes. al., Patrick Robison et. 2016, Science, Vols. 352,aaf0659. [CrossRef]
- Microtubules coordinate mitochondria transport with myofibril morphogenesis during muscle development. Avellaneda J, Candeias D, da Rosa Soares A, Gomes ER, Luis NM, Schnorrer F. 2025 Jul 15, Dev Cell., Vols. S1534-5807(25)00411-3. [CrossRef]
- Suppression of detyrosinated microtubules improves cardiomyocyte function in human heart failure. . Chen, C.Y., Caporizzo, M.A., Bedi, K. et al. 2018, Nat Med , Vol. 24, pp. 1225–1233 . [CrossRef]
- Cardiac microtubules in health and heart disease. Caporizzo MA, Chen CY, Prosser BL. 15, 2019 Nov, Exp Biol Med (Maywood)., Vol. 244.
- A microtubule-connexin-43 regulatory link suppresses arrhythmias and cardiac fibrosis in Duchenne muscular dystrophy mice. . Himelman E, Nouet J, Lillo MA, Chong A, Zhou D, Wehrens XHT, Rodney GG, Xie LH, Shirokova N, Contreras JE, Fraidenraich D. 5, 2022 Nov 1, Am J Physiol Heart Circ Physiol. , Vol. 323, pp. H983-H995. . [CrossRef]
- Cardiomyocyte Microtubules: Control of Mechanics, Transport, and Remodeling. . Uchida K, Scarborough EA, Prosser BL. 84, 2022 Feb, Annu Rev Physiol. , Vol. 10, pp. 257-283. [CrossRef]
- Rethinking tubulin acetylation: From regulation to cellular adaptation. . Donker L, Godinho SA. 2025 Jun, Curr Opin Cell Biol., Vol. 94:102512. [CrossRef]
- Consensus Statement Regarding the Efficacy and Safety of Long-Term Low-Dose Colchicine in Gout and Cardiovascular Disease. Robinson, Philip C. et al. 2022 Feb 10, Annu Rev Physiol. , Vol. 84, pp. 257-283. [CrossRef]
- Dysfunction of microtubules induces cardiac dysfunction. . M., Kitakaze. Nov 2018, EBioMedicine, Vols. 37:3-4. [CrossRef]
- The interconnections between the microtubules and mitochondrial networks in cardiocerebrovascular diseases: Implications for therapy. Shi X, Jiang X, Chen C, Zhang Y, Sun X. 2022 Oct, Pharmacol Res. , Vol. 184:106452. [CrossRef]
- Mitochondria are transported along microtubules in membrane nanotubes to rescue distressed cardiomyocytes from apoptosis. Shen, J., Zhang, JH., Xiao, H. et al. 2018, Cell Death Dis, Vols. 9, 81. [CrossRef]
- Ion Channel Impairment and Myofilament Ca2+ Sensitization: Two Parallel Mechanisms Underlying Arrhythmogenesis in Hypertrophic Cardiomyopathy. Santini L, Coppini R, Cerbai E. 2021 Oct 18, Cells., Vol. 10(10):2789. [CrossRef]
- How calcium causes microtubule depolymerization. O’Brien ET, Salmon ED, Erickson HP. 1997, Cell Motil Cytoskeleton., Vol. 36(2), pp. 125-35. [CrossRef]
- Arrhythmogenic Mechanisms in Heart Failure: Linking β-Adrenergic Stimulation, Stretch, and Calcium. Johnson DM, Antoons G. Oct 16 2018 , Front Physiol., p. 9:1453.
Figure 1.
Proposed multi-factorial model of increased mechanical dispersion in cardiometabolic HFpEF. Emphasizing the role of microtubular dysfunction.
Figure 1.
Proposed multi-factorial model of increased mechanical dispersion in cardiometabolic HFpEF. Emphasizing the role of microtubular dysfunction.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



