
Submitted:
27 January 2026
Posted:
27 January 2026
You are already at the latest version
Abstract
Liver fibrosis can develop after sustained oxidative stress and inflammatory signaling and transforming growth factor-β (TGF-β) is a key profibrotic mediator. Rutin (Ru) is a plant-derived flavonoid with antioxidant and anti-inflammatory effects, but with limited bioavailability, which impedes therapeutic efficacy. This study investigated the hepatoprotective potential of rutin-phytoreduced gold nanoparticles (RuAuNPs) in a thioacetamide (TAA)-induced rat model of liver fibrosis. RuAuNPs were spherical, 18nm diameter (transmission electron microscopy -TEM) and exhibited SPR peak at 523nm (UV–Vis spectroscopy). Following liver fibrosis induction, animals were treated with free rutin or RuAuNPs. Oxidative stress (spectrophotometry, fluorimetry), inflammation (ELISA) and liver morphology (histopathology, TEM) were evaluated. TAA induced hepatic injury and fibrotic remodeling, associated with elevated TGF-β expression, oxidative stress, and inflammation. Free rutin showed limited recovery of liver parameters. RuAuNP treatment significantly improved hepatic histology and ultrastructure, reduced TGF-β and pro-inflammatory cytokine levels (TNFα, IL1β, IL6), decreased oxidative stress damage (lipid peroxidation), restored glutathione redox balance, and enhanced superoxide dismutase activity. Improvement in the AST/ALT ratio suggested early functional recovery, despite persistent aminotransferase elevation. Overall, RuAuNPs enhanced rutin’s antifibrotic and hepatoprotective effects by targeting oxidative and inflammatory pathways and can represent an effective delivery system for rutin in liver fibrosis therapy.
Keywords:
gold nanoparticles
; rutin
; thioacetamide
; liver fibrosis
; TGF-β
; oxidative stress
; glutathione redox balance
; pro-inflammatory cytokines
1. Introduction
Chronic liver disease involves a progressive loss of hepatic function, impairing essential processes such as protein and coagulation factor synthesis, detoxification, and bile production [1] It arises from ongoing cycles of inflammation, hepatocellular injury, and regeneration, which eventually overwhelm the liver’s regenerative capacity. This results in excessive extracellular matrix deposition, leading to liver fibrosis and, in advanced stages, cirrhosis [2,3]. Liver fibrogenesis is a wound-healing response to chronic injury aimed at maintaining tissue integrity and involves coordinated interactions among multiple cell types and molecular mediators [4,5]. Hepatic stellate cells (HSCs) are key drivers of this process. Normally quiescent, HSCs become activated following liver injury and transition into proliferative, myofibroblast-like cells that produce extracellular matrix components [5,6].
Reactive oxygen species (ROS) act as important pro-fibrotic signals by activating pathways such as transforming growth factor-β (TGF-β), which promotes collagen synthesis and inhibits matrix degradation, and accelerate fibrosis progression [7,8,9].
Rutin is a naturally occurring flavonoid found in plants such as buckwheat, tea, and apples. It consists of quercetin linked to the disaccharide rutinose and exhibits notable antioxidant and anti-inflammatory activities. These effects are mediated through regulation of intracellular redox balance and inhibition of pro-inflammatory mediators, including TNF-α, IL-6, IL-1β, and COX-2, leading to a hepatoprotective potential [10,11].
Gold nanoparticles (AuNPs) offer a promising strategy for antifibrotic therapy due to their favorable biodistribution and liver-targeting properties. Their nanoscale size promotes hepatic accumulation via the enhanced permeability and retention effect, with uptake primarily by Kupffer cells and hepatic stellate cells—key mediators of liver fibrosis [12]. Functionalized AuNPs have been shown to reduce hepatic stellate cell activation, including decreased α-smooth muscle actin expression, and to exert antioxidant and anti-inflammatory effects by suppressing cytokine production and oxidative stress [13].
Emerging evidence from in vivo and in vitro studies indicates that flavonoid-based nanomedicines, including quercetin-derived formulations, can attenuate liver fibrosis by improving redox balance and inhibition of profibrogenic signaling [14].
Based on these, the present study aimed to develop a novel therapeutic strategy for hepatic fibrosis using rutin-mediated gold nanoparticles (AuNPs) and to evaluate their antifibrotic efficacy in a validated experimental liver injury model. In this model, previously established by our group, there is a reliable induction of oxidative stress, liver inflammation, and fibrosis [15]. The model enables the assessment of the studied compound’s ability to modulate redox imbalance and TGF-β–driven profibrotic pathways, done by histopathological and biochemical analyses.
2. Materials and Methods
2.1. Preparation and Characterization of Rutin-Reduced Gold Nanoparticles
Rutin-reduced gold nanoparticles were synthesized through a green chemical reduction approach, using rutin as both a reducing and stabilizing agent. Briefly, 61 mg of rutin was dispersed in 100 mL of ultrapure deionized water, and approximately 4 mL of a 2 M aqueous sodium hydroxide solution was gradually added under continuous stirring until complete dissolution of rutin was achieved and the solution exhibited a yellow–orange coloration. To this basic rutin solution, 100 mL of a 1 mM aqueous solution of chloroauric acid (HAuCl₄) was added, and the reaction mixture was stirred for 2 h at room temperature to ensure complete reduction of Au³⁺ ions, allowing the formation of gold nanoparticles. The reaction mixture gradually changed color from pale yellow to ruby red, indicating the formation of gold nanoparticles due to the specific surface plasmon resonance of colloidal gold.
The synthesized gold nanoparticles were purified by centrifugation to remove unreacted rutin and soluble byproducts. The colloidal suspension was centrifuged at 10,000 rpm for 20 minutes, and the supernatant was discarded. The resulting pellet was dispersed in deionized water using mild sonication. The washing process was repeated three times to ensure adequate purification. The final purified gold nanoparticles were stored in deionized water at 4 °C for further characterization and application studies -biological experiments.
Formation of colloidal gold nanoparticles was monitored by UV–Vis spectroscopy, with absorption spectra recorded in the 300–800 nm range, using a PerkinElmer Lambda 25 UV–Vis spectrophotometer (PerkinElmer, Shelton, CT, USA). The formation of AuNPs was confirmed by the appearance of a characteristic surface plasmon resonance (SPR) band typically around 520–550 nm.
Transmission electron microscopy was employed to assess nanoparticles’ morphology and size, using a Hitachi H-7650 transmission electron microscope operating at 120 kV (Hitachi, Tokyo, Japan). TEM samples were prepared by placing a drop of the AuNP suspension onto a carbon-coated copper grid and allowing it to dry under ambient conditions.
Rutin-reduced gold nanoparticles were synthesized through a green chemical reduction approach, using rutin as both a reducing and stabilizing agent. Briefly, 61 mg of rutin was dispersed in 100 mL of distilled water, and approximately 4 mL of a 2 M aqueous sodium hydroxide solution was gradually added under continuous stirring until complete dissolution of rutin was achieved and the solution exhibited a yellow–orange coloration. Subsequently, 100 mL of a 1 mM chloroauric acid (HAuCl₄) solution was added to the rutin solution, and the reaction mixture was stirred for an additional 1 h at room temperature, allowing for the formation of gold nanoparticles.
Purification of the obtained gold nanoparticle suspension was carried out by centrifugation at 10,000 rpm. The resulting pellet was washed twice with distilled water to remove unreacted compounds and residual reagents. After washing, the pellet was resuspended and further used for biological experiments.
2.2. Biological Experiments
2.2.1. Experimental Design
The study was conducted on 42 adult female Wistar albino rats, aged 4 months and weighing 280 ± 20 g. Animals were housed under controlled environmental conditions (12h light/ 12h dark cycle, 35% relative humidity, constant temperature), with free access to water and a standard normo-caloric diet. All experimental procedures involving animals were conducted in accordance with the ARRIVE guidelines and complied with national and international regulations for the care and use of laboratory animals. The study protocol was reviewed and approved by the Institutional Animal Care and Use Committee of “Iuliu Hațieganu” University of Medicine and Pharmacy and by the Veterinary Public Health Directorate of Cluj-Napoca (approval no. AVZ56/26.03.2024), and all efforts were made to minimize animal suffering and reduce the number of animals used.
Animals were randomly allocated into six experimental groups (n = 7/ group). Hepatic fibrosis was induced by oral administration of thioacetamide (TAA, 150 mg/kg body weight) for six consecutive weeks. Groups I–III received physiological saline, while groups IV–VI received TAA according to the same schedule. Starting from the 7th week, the animals received the following treatments: group I (Control) -vehicle (30 mg/kg NaCl 0.9% solution); group II (Ru) - rutin (30 mg/kg body weight); and group III (RuAuNP) - rutin-mediated gold nanoparticles (0.3 mg/kg body weight), group IV (TAA) - vehicle; group V (TAARu) - rutin (30 mg/kg body weight); and group VI (TAARuAuNP) - rutin-mediated gold nanoparticles (0.3 mg/kg body weight). All treatments were administered for four weeks of the experimental protocol (Figure 1).
The rutin dose (30 mg/kg b.w.) was selected as a moderate, literature-supported oral exposure. The used dose was within the range commonly used in rat models to achieve antioxidant/hepatoprotective effects while avoiding excessive dosing. Rutin exhibited antioxidant and hepatoprotective effects, when administered orally at a dose of 25 mg/kg/day in a liver-toxicity setting [16] and respectively at 50–70 mg/kg in experimental hepatotoxicity models [17]. The 0.3 mg/kg dose corresponds to the administered AuNP (nanoparticle mass), rather than an equimass amount of free rutin. It was selected as a low, biologically active exposure based on our group’s previously validated TAA hepatopathy protocol using oral TAA (150 mg/kg for 6 weeks) and AuNPs (0.3 mg/kg), where antioxidant and anti-inflammatory effects were consistently observed [18].
At the end of the 10-week experimental period, animals were humanely euthanized by overdose of ketamine and xylazine, in accordance with accepted ethical standards for laboratory animal care. Blood samples were collected from the retro-orbital sinus into heparinized tubes and centrifuged at 3500 rpm for 10 minutes for plasma separation and further processed for biochemical analyses. Liver tissue samples were collected and used for the evaluation of inflammatory markers, oxidative stress parameters, morphology examination by histopathology and transmission electron microscopy (TEM) analyses.
2.2.2. Histopathology Evaluation
Liver tissue samples were collected at necropsy and immediately fixed in neutral buffered 5% formalin. After adequate fixation, specimens were processed through graded ethanol dehydration and xylene clearing and were subsequently embedded in paraffin. Paraffin blocks were sectioned on a rotary microtome at a thickness of 5 µm. The sections were mounted on glass slides, dried, and subjected to routine hematoxylin–eosin (HE) staining following standard histological protocols. Histological evaluation was performed by light microscopy. Necro-inflammatory activity and architectural alterations were assessed using the Knodell scoring system [19]. The components of the score, including periportal and bridging necrosis, intralobular degeneration and focal necrosis, portal inflammation, and fibrosis, were individually evaluated according to predefined criteria. For each sample, partial scores were summed to generate the total Knodell histological activity index. For each experimental group, individual scores were recorded and subsequently used to calculate a group index. Data were expressed as the distribution of Knodell scores within each group, and comparative analysis between experimental groups was based on these index values. All scoring was performed in a blinded manner to minimize observer bias.
2.2.3. Transmission Electron Microscopy
The liver samples were fixed with glutharaldehyde (Agar Scientific Ltd., Stansted, UK) at 4 °C in), washed 4 times with 0.1 M phosphate buffer (pH = 7.4), and postfixed with osmium tetroxide (Sigma-Aldrich, St. Louis, MO, USA). They were then dehydrated in a series of acetone solutions of increasing concentrations (30-100%) and infiltrated with EMbed 812 (Electron Microscopy Sciences, Hatfield, PA, USA). From the blocks obtained, 70 nm thick sections were cut on an LKB Ultrotome III Bromma 8800 ultramicrotome (LKB Produckter AB, Stockholm-Bromma, Sweden) using a DiATOME diamond knife (DiATOME, Hatfield, PA, USA). Following their collection on 300 mesh copper grids (Agar Scientific Ltd., Stansted, UK), the sections were double contrasted with uranyl acetate and lead citrate. The samples were examined on a JEOL JEM 100CX II microscope (Jeol Ltd., Tokyo, Japan), at 80 kV, equipped with a MegaView G3 camera, operating with the Radius 2.1 software (both from Emsis, Münster, Germany).
2.2.4. Tissue Processing for Biochemical Analyses
Liver tissue samples were homogenized using a POLYTRON PT 1200 E (Kinematica AG Luzemerstrasse 147a CH-6014 Lithau-Lucerne Switzerland) homogenizer in TRIS buffer (50 mM) containing EDTA (10 mM), pH 7.5. Homogenates were centrifuged for 30 min at 10000rot/min. Supernatant was collected and used for analyses. Protein concentration was determined by the Bradford method [20].
2.2.5. Liver Function, Inflammatory and Fibrosis Markers
Aspartate aminotransferase (AST), alanine aminotransferase (ALT), cholesterol, glucose, and triglycerides, were determined from serum by using spectrophotometric methods with commercially available diagnostic kits (Hospitex Diagnostic, Sesto F.no-Firenze-Italy), following the manufacturer’s instructions.
The levels of proinflammatory cytokines interleukin 6 (IL-6), interleukin 1β (IL-1β), and tumor necrosis factor α (TNF-α) and the fibrosis marker transforming growth factor beta (TGF-β) were measured from the liver lysates by using enzyme-linked immunosorbent assay (ELISA) kits, according to the manufacturers’ protocols (Elabscience, Wuhan ,China). Results were expressed as pg/mg protein.
2.2.6. Oxidative Stress Markers
Lipid peroxidation was evaluated by measuring malondialdehyde (MDA) levels using a thiobarbituric acid–based fluorimetric assay. Plasma samples and tissue homogenates were incubated with 2-thiobarbituric acid (10 mM) in K₂HPO₄ buffer (75 mM, pH 3), followed by extraction in n-butanol. Fluorescence was measured by synchronous spectrofluorimetry (Δλ = 14 nm) at 534 nm nm using Perkin Elmer LS 45 Fluorescence Spectrometer U.S, and MDA concentrations were expressed as nM/mg protein [21].
Reduced and oxidized glutathione (GSH and GSSG) levels were determined using a fluorimetric method based on o-phthalaldehyde derivatization. Fluorescence was recorded at an excitation wavelength of 350 nm and an emission wavelength of 420 nm, using Perkin Elmer LS 45 Fluorescence Spectrometer U.S [22].
Glutathione peroxidase (GPx) activity was spectrophotometrically assessed by monitoring NADPH consumption at 340 nm according to the method of Flohé et al. [23]. Superoxide dismutase (SOD) activity was measured in liver lysates using a commercial assay kit (Sigma-Aldrich), based on the inhibition of nitro blue tetrazolium reduction. Enzyme activities were expressed as enzymatic units/ gram protein [24].
2.2.7. Statistical Analysis
Data were analyzed using GraphPad Prism (v8.2; GraphPad Software, San Diego, CA, USA; downloaded on 15 March 2019) Normality was evaluated using the Shapiro–Wilk test. For normally distributed data, group differences were assessed by one-way ANOVA followed by Fisher’s LSD multiple-comparisons test. For non-normally distributed data, the Kruskal–Wallis test was applied, followed by Dunn’s multiple-comparisons test. Results are reported as mean ± SD. A two-sided p-value < 0.05 was considered statistically significant. Graphs were generated in GraphPad Prism (v8.2).
3. Results
3.1. Characterization of Rutin-Phytoreduced Gold Nanoparticles
Gold nanoparticles were synthesized via a green and facile reduction method using rutin as a natural reducing and capping agent. Upon mixing the rutin basic solution with the Au³⁺ solution, a rapid color change from pale yellow to intense purple red was observed, indicating that the reduction process had occurred. In addition to reducing Au³⁺ ions, rutin also acted as a stabilizing agent by binding to the nanoparticle surface through its hydroxyl functional groups, thereby preventing aggregation and yielding stable, well-dispersed gold nanoparticles [25].
The size and shape of gold nanoparticles (AuNPs) play a critical role in determining their biological activity, including cellular uptake, biodistribution, toxicity and therapeutic efficacy.
Characterization of the nanoparticles was further done by UV–Vis spectroscopy and transmission electron microscopy (TEM). The UV–Vis spectrum of rutin-reduced gold nanoparticles exhibited a characteristic absorption maximum at 523 nm, confirming the formation of gold nanoparticles (Figure 2).
Figure 2.
UV-VIS spectrum of the rutin- phytoreduced nanoparticles.
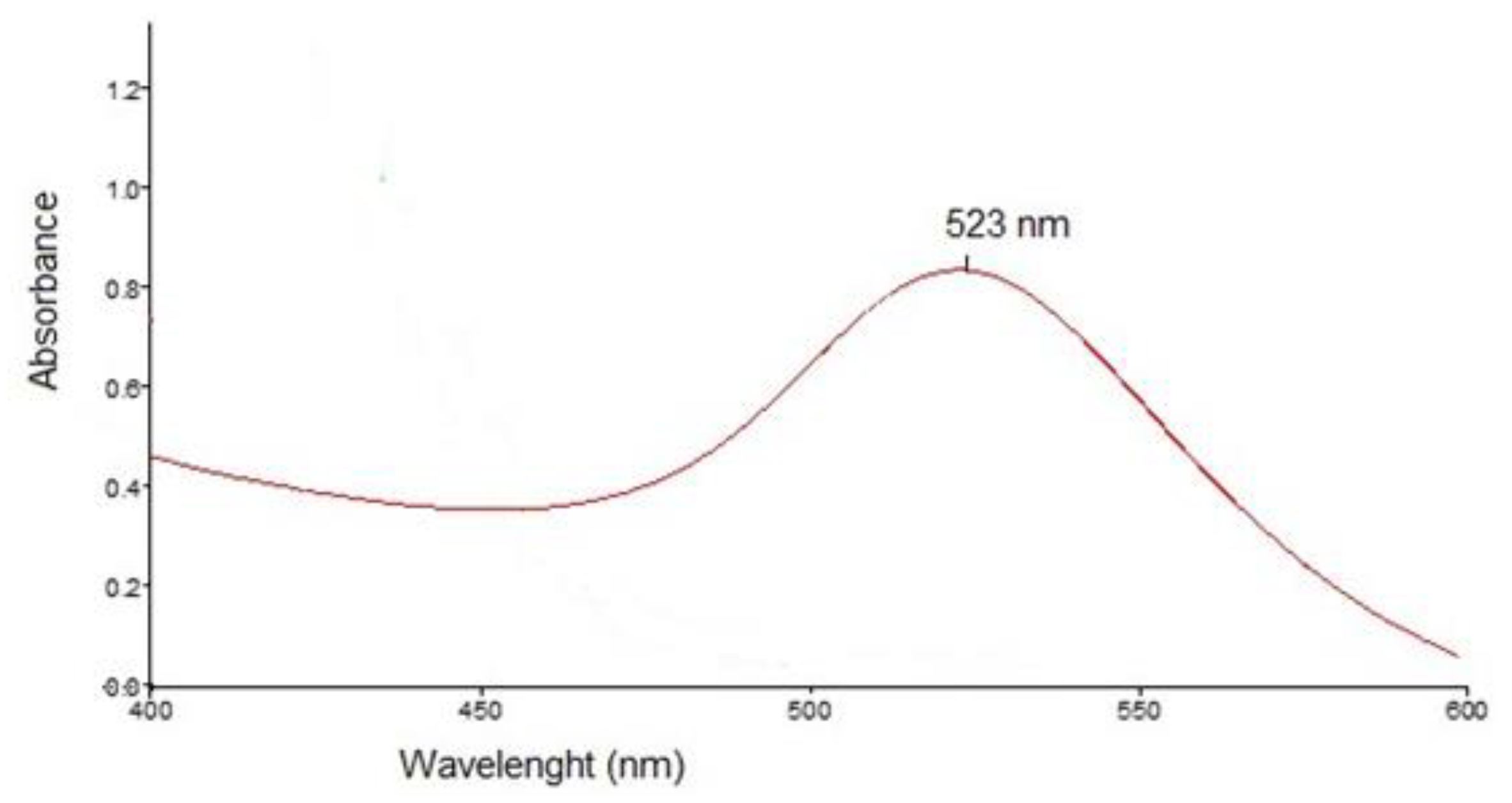
Figure 3.
Transmission electron microscopy of the rutin- reduced gold nanoparticles.
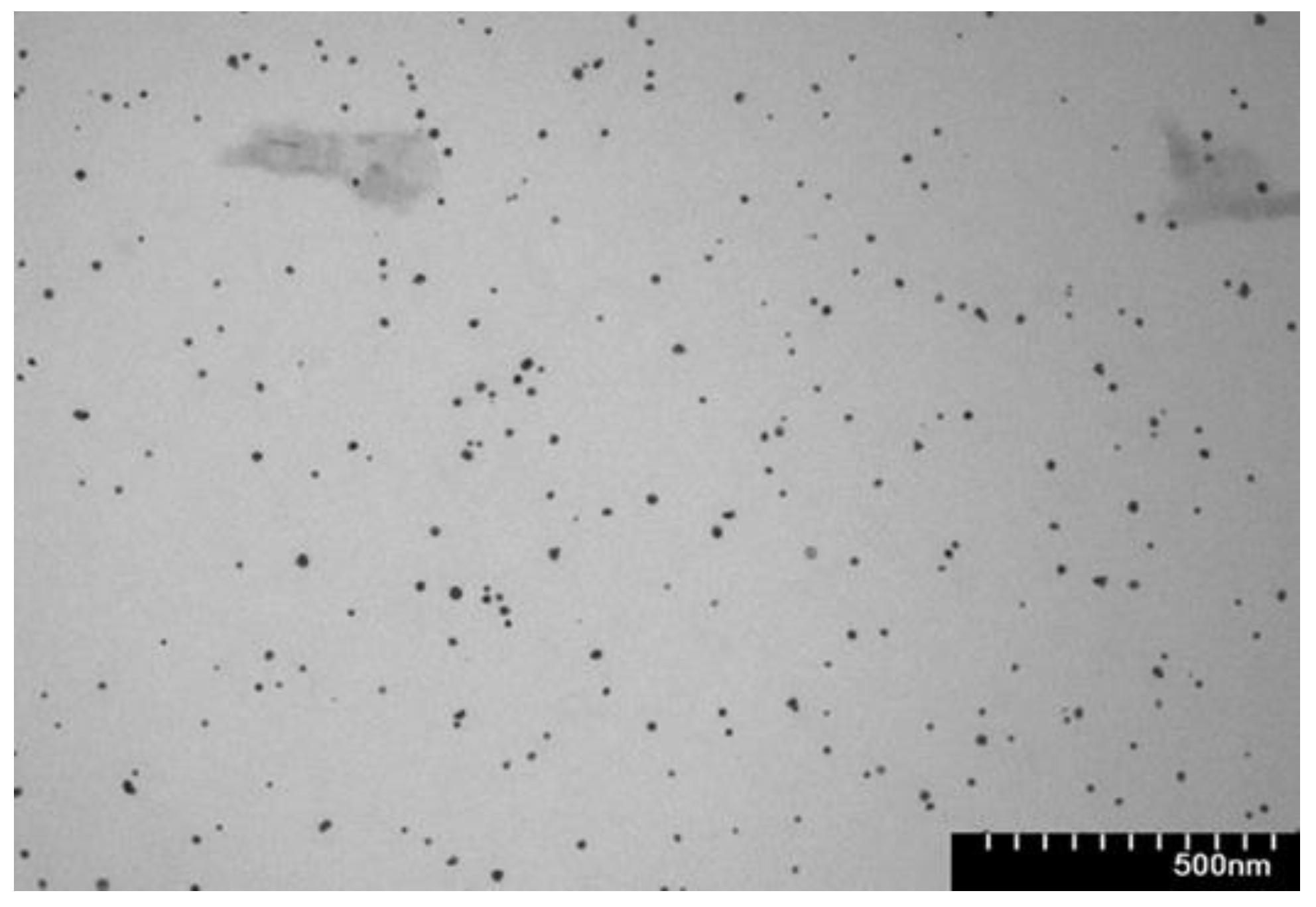
The size, shape, and morphology of the synthesized gold nanoparticles were examined using transmission electron microscopy. TEM imaging confirmed the formation of nearly spherical gold nanoparticles, with a narrow distribution and an average diameter of 18 nm.
3.2. Morphological Liver Alterations
Histological examination using hematoxylin–eosin staining (Figure 4) showed that, compared with the control group, TAA increased the vascular wall thickness, a marker of the fibrogenic hepatic response. Rutin alone caused mild histological changes, mainly characterized by mixed dystrophy, predominantly vacuolar and pigment accumulation in hepatocytes. Combined treatment of TAA and rutin did not improve the TAA and rutin induced alterations. In contrast, rutin-phytoreduced AuNPs induced dystrophic yet regenerative changes. These lesions are suggestive of osmotic-type hepatic alterations induced by the nanoparticles at the administered dose. Similar features were observed in the TAA + rutin-phytoreduced AuNP group. The liver alterations were less severe than in the TAA + rutin group, which indicates a hepatoprotective effect of the nanoformulation. Images are representative of each group.
Evaluation using the Knodell histological activity index showed varying degrees of liver injury among groups. The control group had minimal alterations (score = 1), while the positive control group (TAA) exhibited moderate injury (animals from the group scores ranged between 6-8). The Ru group showed mild lesions (score = 3), lower than TAA. The TAARu group had the highest score = 10, with severe necroinflammatory damage. Nanoparticle administration exhibited a similar score compared to TAA (scores ranged 6-8), yet lower when compared to the TAARu group. TAARuAuNP showed a similar score ranging of 6 - 8 within the samples, with no further worsening in the presence of TAA as compared to RuAuNP.
In the control group, the liver presented a normal ultrastructure. The hepatocytes had large, rounded nuclei and abundant cytoplasm (Figure 5 a-c). The numerous mitochondria were round or oval, with small cristae (Figure 5 a-d). Additionally, rough endoplasmic reticulum profiles and glycogen granules were observed (Figure 5 a-d), alongside lysosomes (not shown) or occasional small lipid droplets (Figure 5 c), some with dense inclusions. Bile canaliculi with normal aspect were noted between adjacent cells (Figure 5 d).
In the TAA group, many of the hepatocytes had irregular, indented nuclear contour (Figure 6a). Long mitochondria were observed in the cytoplasm (Figure 6b), as well as regions of expanded endoplasmic reticulum (Figure 6a, d) and occasional autophagosomes (Figure 6e). Large lipid droplets with an electron-dense outer rim were also noted (Figure 6c). Additionally, glycogen and lipid accumulations were present in some of the nuclei of the hepatocytes (Figure 6d). No changes were observed at the level of the bile canaliculi (Figure 6e).
In some areas, abundant collagen fibers were observed in the intercellular space (Figure 7 f). The hepatocytes of the TAARu group had areas with expanded profiles of endoplasmic reticulum (Figure 7 a). The lipid droplets were large and numerous, and some contained electron-dense inclusions (Figure 7 b, c). No changes were observed at the level of the bile canaliculi (Figure 7 c). Significant amounts of collagen fibers were also present between the cells in some regions (Figure 7 d).
In the TAARuAuNP group, the hepatocytes had nuclei with slightly irregular contours (Figure 8a, c-e). Some of the mitochondria were long or polymorphic (Figure 8 b, c), and expanded profiles of endoplasmic reticulum were observed (Figure 8 a-e). Large lipid droplets were also noted, some with a heterogenous aspect (Figure 8c). No changes were observed at the level of the bile canaliculi (Figure 8e). Between the hepatocytes, there were collagen-abundant regions (Figure 8f).
Consistent with these findings, TGF-β levels (Figure 9) were strongly increased in the TAA group compared to control (p < 0.0001), confirming activation of profibrotic pathways. TGF-β was also significantly higher than Control in both treated TAA groups (p ≤ 0.0034), indicating that fibrogenic signaling remained elevated despite treatment. TAARuAuNP significantly reduced TGF-β compared with the TAA fibrotic group (p = 0.0002), while TAARu exhibited no significance. Overall, AuNP-mediated delivery of rutin down-modulated TGF-β in the fibrotic setting, even though levels did not fully return to Control values within the experimental timeframe.
3.3. Liver Function and Metabolic Results
The TAA group showed marked elevations of AST compared to control (p < 0.0001), consistent with hepatocellular injury. AST levels remained similarly increased in the treated TAA groups, with no significant differences versus TAA (p ≤ 0.8129) (Table 1). In contrast, the AST/ALT ratio differed between treatments, being significantly lower in TAARuAuNP compared with TAARu (p = 0.0215), leading to partial normalization of the enzymatic profile with nanoparticle-mediated rutin delivery (Figure 10).
Metabolic readouts showed limited changes attributable to TAA exposure (Figure 11). Blood glucose and cholesterol showed no significantly different levels between Control and TAA (p ≥ 0.7684). TAARuAuNP displayed significantly lower glycemia than TAARu (p = 0.0307), and Ru (p = 0.0118). For cholesterol, a significant difference was observed between TAARu and TAARuAuNP (p = 0.0268). Triglycerides were significantly lower in TAARuAuNP compared to Control (p = 0.0155) and Ru (p = 0.0045).
3.4. Oxidative Stress
Hepatic malondialdehyde (MDA), a marker of lipid peroxidation, was significantly increased after TAA exposure, compared to control (p < 0.0001), confirming pronounced oxidative damage in liver tissue (Figure 12). In contrast, TAARuAuNP significantly lowered hepatic MDA compared with both the untreated fibrotic group and the free rutin group (p ≤ 0.0038), supporting a stronger antioxidant effect when rutin was delivered via AuNPs. Nevertheless, hepatic MDA in TAARuAuNP remained higher than Control (p = 0.0079), suggesting partial but incomplete recovery within the experimental timeframe.
Serum MDA did not show a consistent treatment- or TAA-related shift, with no significant differences between control and any TAA group (p > 0.05), indicating that lipid peroxidation changes were primarily detected at the hepatic level in this model (Figure 12).
The GSH/GSSG ratio, a key indicator of glutathione-dependent redox balance, was significantly altered by TAA exposure versus control (p = 0.0098), supporting the presence of oxidative stress in the fibrotic setting (Figure 13). Importantly, TAARuAuNP showed a significant improvement in this parameter compared with both the untreated fibrotic group and free rutin (p ≤ 0.0006), consistent with a stronger antioxidant effect when rutin was delivered via AuNPs. Moreover, the GSH/GSSG ratio in TAARuAuNP was not significantly different from Control (p = 0.2657), suggesting a shift towards redox normalization.
GPx activity showed limited modulation in this model (Figure 9). A significant difference was detected only under non-fibrotic conditions, Control versus Ru groups (p = 0.0144), while no significant differences were observed among TAA-exposed groups (p > 0.05). Overall, these results indicate that the nanoparticle induced antioxidant benefit was better captured by restoration of the glutathione redox system rather than by consistent changes in GPx activity.
Superoxide dismutase (SOD), a key enzymatic antioxidant involved in superoxide radical scavenging, showed a significant treatment effect in the fibrotic setting (Figure 13). TAARuAuNP exhibited higher SOD activity than the TAA untreated fibrotic group (p = 0.0029). TAARu exhibited no statistical significance as compared to TAA. Moreover, SOD activity was significantly higher in TAARuAuNP compared with TAARu (p = 0.0479), supporting an AuNP-dependent enhancement of enzymatic antioxidant defense.
Overall, these findings indicate that TAA-induced liver fibrosis was accompanied by oxidative stress and dysregulated antioxidant responses. Consistent with the MDA and glutathione data, rutin-phytoreduced AuNPs provided a more robust antioxidant signature than free rutin, including reduced lipid peroxidation, improved glutathione redox balance, and increased SOD activity, suggesting that nanoparticle-mediated delivery strengthens rutin’s cytoprotective effects on hepatic oxidative stress pathways.
3.5. Inflammatory Response
IL-1β and IL-6 are key pro-inflammatory cytokines. They are mainly produced by macrophages during oxidative stress exposure and hepatocellular injury. Therefore, their levels reflect the severity of inflammation in our model of the thioacetamide-induced liver fibrosis and the anti-inflammatory potential of treatments. TNF-α is another central mediator of hepatic inflammation, whose elevation indicates activation of inflammatory pathways in this model.
TAA exposure induced a pronounced inflammatory response, as reflected by significantly higher IL-1β (p≤0.0288), IL-6 (p≤0.0006), and TNF-α (p≤0.0201) levels in all TAA-treated groups compared with non-fibrotic controls (Figure 14). For IL-1β, no significant differences were observed among the TAA groups. Neither rutin nor rutin-phytoreduced AuNPs significantly modulated this cytokine relative to the untreated fibrotic group within the experimental timeframe. In contrast, both IL-6 and TNF-α showed significant differences between TAARu and TAARuNP (p≤0.008). This suggests that rutin delivered via AuNPs induced a distinct improved inflammatory profile compared with free rutin, although neither treatment reached statistical significance versus TAA for these cytokines.
4. Discussion
This study evaluated whether delivering rutin via phytoreduced gold nanoparticles (RuAuNP) enhances the hepatic antifibrotic efficacy and biochemical profile in a thioacetamide-driven model of chronic liver injury. The TAA model is widely used because it generates sustained hepatocellular stress with intertwined oxidative and inflammatory responses that drive fibrogenic signaling [26,27]. In line with this framework, the TAA group showed clear histological injury and a marked increase of the hepatic TGF-β, supporting robust activation of profibrotic signaling. Importantly, TAARuAuNP significantly reduced TGF-β compared with TAA, whereas free rutin (TAARu) did not produce a comparable effect compared to the TAA group. This pattern is consistent with the central role of TGF-β in fibrogenesis and with prior TAA induced fibrosis studies where attenuation of oxidative stress and inflammatory cascades converges on reduced TGF-β/Smad signaling and improved tissue architecture [28].
H&E sections showed prominent vascular and parenchymal alterations following TAA exposure, indicative of the induced liver damage. The nanoformulation yielded a qualitatively more favorable pattern than free rutin under TAA exposure. The mild structural changes observed in RuAuNP alone can be interpreted in light of the known hepatic handling of AuNPs—preferential liver accumulation and predominant uptake by Kupffer cells—which may produce subtle exposure-related parenchymal alterations even in the absence of a co-administered hepatotoxic [29,30]. The presence of intranuclear glycogen, shown by TEM analysis has been correlated with numerous pathologies, such as diabetes, obesity, glycogen storage disorders or Wilson’s disease. The intranuclear lipids are also markers of hepatocyte damage [31].
The dystrophic pattern described as “osmotic-type” is consistent with vacuolar/hydropic change, a form of acute cell swelling that is widely regarded as a potentially reversible, sublethal response to cellular stress. [32]. This lesion aspect, named as “regenerative” reflects a pattern suggestive for adaptation/repair rather than progressive destruction. This is in line with prior AuNP liver reports, describing vacuolar-to-hydropic degeneration together with features such as binucleation, often discussed as a marker of hepatocyte turnover after injury [33] .
Although H&E does not directly quantify collagen deposition, the parallel reduction in TGF-β supports the interpretation that TAARuAuNP dampened profibrotic signaling and partially preserved liver structure. The combined evaluation of histology and TGF-β/Smad axis was previously used as supportive evidence of antifibrotic pathway modulation in TAA models. Dendropanax morbifera reduced TGF-β1/p-Smad2/3 alongside with improved histopathology and fibrosis-related readouts [28]. Dendropanoxide attenuated TAA induced fibrosis with decreased TGF-β1/p-Smad2/3 and reduced collagen/α-SMA signals [34]. A comparable pattern was also reported for hesperidin in TAA-induced fibrosis, where antifibrotic effects were linked to modulation of TGF-β/α-SMA with supportive tissue-level improvement [35].
Oxidative stress modulation was one of the clearest differentiators between formulations. Lipid peroxidation, reflected by hepatic MDA, is a well-established feature of TAA-driven liver injury and fibrosis, typically accompanied by depletion of antioxidant defenses such as GSH and reduced enzymatic activity (SOD) [34,36,37]. In our study, hepatic MDA was strongly increased after TAA and was significantly reduced by TAARuAuNP versus both TAA and TAARu. This indicates that nanoparticle-mediated delivery enhanced rutin’s antioxidant impact at tissue level. This direction is consistent with multiple TAA based fibrosis studies in which improved outcomes are closely linked to restoration of redox homeostasis, [34,36] proven by the reduced lipid peroxidation (lower MDA) together with recovery of glutathione-dependent and enzymatic antioxidant systems. Serum MDA did not show significant group differences in our experimental setting. This reflects localized intrahepatic oxidative damage and/or lower sensitivity of systemic MDA to the hepatic alterations.
The glutathione system provided a complementary readout of intracellular redox state. The GSH/GSSG ratio is widely used as an integrated indicator of glutathione-dependent antioxidant capacity; a lower ratio reflects a shift toward a more oxidized intracellular environment and reduced ability to buffer ROS [38,39]. In our data, TAARuAuNP differed significantly from TAA and TAARu, and it was not statistically different from Control, supporting a “toward-normalization” shift in glutathione redox balance with the nanoparticles. This aligns with TAA- induced fibrosis studies showing that effective interventions often improve oxidative injury by restoring glutathione-related defenses and antioxidant enzyme activity (e.g., GSH and SOD recovery with reduced lipid peroxidation) [34,40]. Consistent with this, SOD activity was higher in TAARuAuNP than in TAA, and respectively TAARu. This indicated improved capacity to detoxify superoxide radicals. Finally, reviews on liver fibrosis nanomedicine and flavonoid-based nanoformulations emphasize that successful systems frequently lead to improved antioxidant status (lower lipid peroxidation and improved glutathione/antioxidant metrics) as part of the antifibrotic efficacy [14,41].
Beyond rutin, several plant-derived treatments in TAA models report a similar antifibrotic pattern of histological improvement together with reduced profibrotic/oxidative signaling. Silymarin attenuated chronic TAA injury in mice and downregulated key fibrosis markers, including TGF-β1 and α-SMA, alongside with tissue-level improvement [42]. Dihydromyricetin administered in TAA-induced liver fibrosis in mice improved the histopathology aspect, and reversed oxidative damage leading to decreased hepatic MDA and increased antioxidant defenses such as SOD and GSH, together with dampening of inflammatory signaling [43]. A similar pattern co-modulation was reported for Gardeniae Fructus, which attenuated hepatic oxidation and inflammation in TAA-injected mice and was mechanistically linked to pathways including AMPK/SIRT1/NF-κB and Nrf2 signaling [44]. Comparable results have also been described for other dietary phytochemicals with antifibrotic profiles in TAA injury. In a rat TAA fibrosis setting, the flavone apigenin exhibited antifibrotic effects correlated with downregulation of TGF-β1 (along with other profibrotic signaling nodes), and simultaneous effects on oxidative stress and inflammation [45]. In addition, combining natural antioxidants has shown pathway-level convergence: pomegranate extract + curcumin in TAA-induced fibrosis in rats was reported to impact TGF-β/Smad3 and NF-κB signaling.
This reinforces the broader theme that plant-derived antioxidants often translate benefit through coordinated suppression of the inflammatory–oxidative loops that feed profibrotic signaling [46].
The effects induced by the RuAuNP are consistent with earlier AuNP-based strategies in liver injury. For example, silymarin-coated gold nanoparticles improved CCl₄-induced hepatic injury in Wistar rats, with reductions in liver serum enzymes and fibrosis-related readouts, as indicated by histopathology and immunohistochemistry [47]. This type of AuNP–polyphenol combination is also discussed in liver-fibrosis nanotherapy reviews as a way to influence key fibrogenic compartments, including hepatic stellate cells and Kupffer cells [3]. As such we used a novel formulation of an AuNP platform delivery for an antioxidant - rutin with difficult oral absorption and liver accumulation to strengthen the tissue-level benefit compared with the free rutin alone in a TAA live fibrosis model. In a complementary toxicity-oriented setting, quercetin and arginine mitigated AuNP-induced hepatotoxicity in rats, improving hepatic oxidative injury, supporting the broader idea that flavonoids can counter oxidative damage in AuNP contexts [48]. Finally, recent reviews on liver fibrosis nanomedicine and flavonoid-based nanomedicines highlight that nanocarriers can enhance local exposure and cellular uptake, often translating into stronger antioxidant/anti-inflammatory effects with antifibrotic relevance [14,49,50].
Persistent AST/ALT elevation despite improvements in histology and mechanistic markers can be interpreted as an endpoint–timing mismatch rather than a contradiction. ALT and AST primarily reflect ongoing hepatocyte injury/necrosis and inflammation, not fibrosis burden per se. Therefore, they may remain elevated even when profibrotic signaling (e.g., TGF-β) and redox status begin to improve. This is particularly plausible in chronic toxin models such as TAA, which produce a predictable, sustained injury pattern with concurrent fibrosis development [51].
In our study, AST and ALT remained elevated at the end of the post-induction treatment phase despite clearer improvements in tissue-level readouts (histological directionality, reduced TGF-β, and improved redox markers). This pattern is plausible in the TAA model because aminotransferases primarily reflect ongoing hepatocyte membrane leakage/necrosis, while fibrosis regression and remodeling can persist for weeks after stopping the toxin and may not translate immediately into “normalized” enzymes. In support of this, TAA/TA (thioacetamide/thioacetamide analog) models explicitly describe that biochemical peaks can improve after drug withdrawal, while some liver enzymes can remain elevated, and that fibrotic injury is maintained for several weeks after discontinuation [52]. Therefore, in our dataset, the most conservative interpretation is that the RuAuNP produced a more robust shift in profibrotic/oxidative pathways, but the four-week treatment window after a six-week TAA induction likely captured a phase in which hepatocellular injury markers can lag behind pathway and structural improvements.
To further strengthen these findings, future work could complement the current H&E-based assessment and mechanistic readouts with quantitative collagen-specific endpoints (e.g., Sirius Red morphometry or hydroxyproline content) and biodistribution analyses to directly confirm hepatic uptake and cell-compartment targeting of the RuAuNP. Extending the follow-up period would also help clarify the durability of the antifibrotic response and whether longer treatment windows translate into more complete biochemical recovery.
5. Conclusions
TAA induced consistent experimental liver fibrosis, supported by the histological injury, TEM morphology, increased hepatic TGF-β and oxidative and inflammatory changes. Compared to free rutin, the rutin-phytoreduced gold nanoparticles exhibited a significantly better antifibrotic profile, by decreasing TGF-β and the inflammatory changes and led to a more favorable tissue appearance on H&E and TEM sections. It also provided stronger antioxidant protection, with reduced hepatic oxidative damage and improved both the antioxidants balance, and antioxidant defense. Although AST and ALT remained elevated, the combined tissue and mechanistic data suggest that pathway and redox improvements may come before full biochemical recovery.
Overall, nanoparticle-mediated hepatic delivery of rutin significantly potentiated its therapeutic efficacy in the TAA-induced fibrosis model, supporting its potential use as an adjuvant strategy in the management of liver fibrotic diseases.
Author Contributions
Conceptualization, R.M.D., G.A.F. and S.V.C.; methodology, G.A.F., S.V.C.; software, R.M.D.; validation, I.B., L.D. and S.V.C.; formal analysis, V.T., M.M.; investigation, C.A.M., B.E.M.; resources, R.M.D., I.B.; data curation, R.M.D., L.D; writing—original draft preparation, R.M.D.; writing—review and editing, I.B.; visualization, R.M.D.; supervision, S.V.C.; project administration, R.M.D.; funding acquisition, R.M.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
All experimental data, supporting the results is presented in the current article.
Acknowledgments
During the preparation of this manuscript, the authors used ChatGPT (OpenAI, GPT-5.2) to assist with language refinement. The authors critically reviewed, edited, and verified all content and take full responsibility for the final version of the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
TGF-β Transforming growth factor beta
Ru Rutin
RuAuNPs Rutin-mediated (rutin-capped) gold nanoparticles
UV–Vis Ultraviolet–visible spectroscopy
TAA Thioacetamide
ROS Reactive oxygen species
TNF-α Tumor necrosis factor alpha
IL-6 Interleukin-6
IL-1β Interleukin-1 beta
COX-2 Cyclooxygenase-2
AuNPs Gold nanoparticles
HSCs Hepatic stellate cells
α-SMA Alpha-smooth muscle actin
SPR Surface plasmon resonance
TEM Transmission electron microscopy
ARRIVE Animal Research Reporting of In Vivo Experiments
B.w. Body weight
AST Aspartate aminotransferase
ALT Alanine aminotransferase
ELISA Enzyme-linked immunosorbent assay
MDA Malondialdehyde
GSH Reduced glutathione
GSSG Oxidized glutathione
GPx Glutathione peroxidase
SOD Superoxide dismutase
SD Standard deviation
ANOVA Analysis of variance
HE / H&E Hematoxylin and eosin
AMPK AMP-activated protein kinase
SIRT1 Sirtuin 1
NF-κB Nuclear factor kappa-B
Nrf2 Nuclear factor erythroid 2–related factor 2
References
- Sharma, A.; Nagalli, S. Chronic Liver Disease; StatPearls Publishing, 2023. [Google Scholar]
- Khanam, A.; Saleeb, P.G.; Kottilil, S. Pathophysiology and Treatment Options for Hepatic Fibrosis: Can It Be Completely Cured? Cells 2021, 10. [Google Scholar] [CrossRef]
- Poilil Surendran, S.; George Thomas, R.; Moon, M.J.; Jeong, Y.Y. Nanoparticles for the Treatment of Liver Fibrosis. Int. J. Nanomedicine 2017, 12, 6997–7006. [Google Scholar] [CrossRef]
- Friedman, S.L. Mechanisms of Hepatic Fibrogenesis. Gastroenterology 2008, 134, 1655–1669. [Google Scholar] [CrossRef] [PubMed]
- Roehlen, N.; Crouchet, E.; Baumert, T.F. Liver Fibrosis: Mechanistic Concepts and Therapeutic Perspectives. Cells 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Berumen, J.; Baglieri, J.; Kisseleva, T.; Mekeel, K. Liver Fibrosis: Pathophysiology and Clinical Implications. WIREs Mech Dis 2021, 13, e1499. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Valle, V.; C. Chavez-Tapia, N.; Uribe, M.; Mendez-Sanchez, N. Role of Oxidative Stress and Molecular Changes in Liver Fibrosis: A Review. Curr. Med. Chem. 2012, 19, 4850–4860. [Google Scholar] [CrossRef]
- Seki, E.; Schwabe, R.F. Hepatic Inflammation and Fibrosis: Functional Links and Key Pathways. Hepatology 2015, 61, 1066–1079. [Google Scholar] [CrossRef]
- Dhar, D.; Baglieri, J.; Kisseleva, T.; Brenner, D.A. Mechanisms of Liver Fibrosis and Its Role in Liver Cancer. Exp. Biol. Med. 2020, 245, 96–108. [Google Scholar] [CrossRef]
- Ganeshpurkar, A.; Saluja, A.K. The Pharmacological Potential of Rutin. Saudi Pharm. J. 2017, 25, 149–164. [Google Scholar] [CrossRef]
- Muvhulawa, N.; Dludla, P.V.; Ziqubu, K.; Mthembu, S.X.H.; Mthiyane, F.; Nkambule, B.B.; Mazibuko-Mbeje, S.E. Rutin Ameliorates Inflammation and Improves Metabolic Function: A Comprehensive Analysis of Scientific Literature. Pharmacol. Res. 2022, 178, 106163. [Google Scholar] [CrossRef]
- Hirn, S.; Semmler-Behnke, M.; Schleh, C.; Wenk, A.; Lipka, J.; Schäffler, M.; Takenaka, S.; Möller, W.; Schmid, G.; Simon, U.; et al. Particle Size-Dependent and Surface Charge-Dependent Biodistribution of Gold Nanoparticles after Intravenous Administration. Eur. J. Pharm. Biopharm. 2011, 77, 407–416. [Google Scholar] [CrossRef]
- Tee, J.K.; Peng, F.; Ho, H.K. Effects of Inorganic Nanoparticles on Liver Fibrosis: Optimizing a Double-Edged Sword for Therapeutics. Biochem. Pharmacol. 2019, 160, 24–33. [Google Scholar] [CrossRef]
- Shree Harini, K.; Ezhilarasan, D. Flavonoids-Based Nanomedicines for the Treatment of Liver Fibrosis: A Recent Progress. J. Drug Deliv. Sci. Technol. 2024, 93, 105467. [Google Scholar] [CrossRef]
- Clichici, S.; Catoi, C.; Mocan, T.; Filip, A.; Login, C.; Nagy, A.; Daicoviciu, D.; Decea, N.; Gherman, C.; Moldovan, R.; et al. Non-Invasive Oxidative Stress Markers for Liver Fibrosis Development in the Evolution of Toxic Hepatitis. Acta Physiol. Hung. 2011, 98, 195–204. [Google Scholar] [CrossRef]
- Anwar, M.M.; Laila, I.M.I. The Ameliorating Effect of Rutin on Hepatotoxicity and Inflammation Induced by the Daily Administration of Vortioxetine in Rats. BMC Complement. Med. Ther. 2024, 24, 153. [Google Scholar] [CrossRef] [PubMed]
- Hafez, M.M.; Al-Harbi, N.O.; Al-Hoshani, A.R.; Al-Hosaini, K.A.; Al Shrari, S.D.; Al Rejaie, S.S.; Sayed-Ahmed, M.M.; Al-Shabanah, O.A. Hepato-Protective Effect of Rutin via IL-6/STAT3 Pathway in CCl4-Induced Hepatotoxicity in Rats. Biol. Res. 2015, 48, 30. [Google Scholar] [CrossRef] [PubMed]
- Filip, M.; Clichici, S.V.; Muntean, M.; David, L.; Moldovan, B.; Toma, V.A.; Login, C.; Suciu, Ş.M. The Impact of Hybrid Bionanomaterials Based on Gold Nanoparticles on Liver Injury in an Experimental Model of Thioacetamide-Induced Hepatopathy. Biomolecules 2025, 15, 1068. [Google Scholar] [CrossRef] [PubMed]
- Knodell, R.G.; Ishak, K.G.; Black, W.C.; Chen, T.S.; Craig, R.; Kaplowitz, N.; Kiernan, T.W.; Wollman, J. Formulation and Application of a Numerical Scoring System for Assessing Histological Activity in Asymptomatic Chronic Active Hepatitis. Hepatology 1981, 1, 431–435. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Conti, M.; Morand, P.C.; Levillain, P.; Lemonnier, A. Improved Fluorometric Determination of Malonaldehyde. Clin. Chem. 1991, 37, 1273–1275. [Google Scholar] [CrossRef]
- Vats, P.; Singh, V.K.; Singh, S.N.; Singh, S.B. Glutathione Metabolism under High-Altitude Stress and Effect of Antioxidant Supplementation. Aviat. Space Environ. Med. 2008, 79, 1106–1111. [Google Scholar] [CrossRef]
- Flohé, L.; Günzler, W.A. Assays of Glutathione Peroxidase. Methods Enzymol. 1984, 105, 114–121. [Google Scholar] [CrossRef]
- Fridovich, I. Superoxide Dismutases. J. Biol. Chem. 1989, 264, 7761–7764. [Google Scholar] [CrossRef]
- Wu, H.; Tang, L.; Dong, H.; Zhi, M.; Guo, L.; Hong, X.; Liu, M.; Xiao, Y.; Zeng, X. Shape and Size Dependence of Pharmacokinetics, Biodistribution, and Toxicity of Gold Nanoparticles. Mol. Pharm. 2025, 22, 196–208. [Google Scholar] [CrossRef] [PubMed]
- Liedtke, C.; Luedde, T.; Sauerbruch, T.; Scholten, D.; Streetz, K.; Tacke, F.; Tolba, R.; Trautwein, C.; Trebicka, J.; Weiskirchen, R. Experimental Liver Fibrosis Research: Update on Animal Models, Legal Issues and Translational Aspects. Fibrogenesis Tissue Repair 2013, 6, 19. [Google Scholar] [CrossRef] [PubMed]
- Dashti, H.; Jeppsson, B.; Hägerstrand, I.; Hultberg, B.; Srinivas, U.; Abdulla, M.; Bengmark, S. Thioacetamide- and Carbon Tetrachloride-Induced Liver Cirrhosis. Eur. Surg. Res. 1989, 21, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.Y.; Kim, K.S.; Lee, Y.H.; Park, J.H.; Kim, J.-H.; Lee, S.-Y.; Kim, Y.-M.; Kim, I.S.; Kacew, S.; Lee, B.M.; et al. Dendropanax Morbifera Ameliorates Thioacetamide-Induced Hepatic Fibrosis via TGF-β1/Smads Pathways. Int. J. Biol. Sci. 2019, 15, 800–811. [Google Scholar] [CrossRef]
- Bailly, A.-L.; Correard, F.; Popov, A.; Tselikov, G.; Chaspoul, F.; Appay, R.; Al-Kattan, A.; Kabashin, A.V.; Braguer, D.; Esteve, M.-A. In Vivo Evaluation of Safety, Biodistribution and Pharmacokinetics of Laser-Synthesized Gold Nanoparticles. Sci. Rep. 2019, 9, 12890. [Google Scholar] [CrossRef]
- Sadauskas, E.; Wallin, H.; Stoltenberg, M.; Vogel, U.; Doering, P.; Larsen, A.; Danscher, G. Kupffer Cells Are Central in the Removal of Nanoparticles from the Organism. Part. Fibre Toxicol. 2007, 4, 10. [Google Scholar] [CrossRef]
- Imai, N.; Ohsaki, Y.; Cheng, J.; Kawecka, H.; Zhang, J.; Mizuno, F.; Tanaka, T.; Yokoyama, S.; Yamamoto, K.; Ito, T.; et al. Pathological Significance of Intranuclear Structures in Liver Biopsy Samples. Hepatol. Res. 2025, 55, 1065–1074. [Google Scholar] [CrossRef]
- Miller, M.A.; Zachary, J.F. Mechanisms and Morphology of Cellular Injury, Adaptation, and Death. In Pathologic Basis of Veterinary Disease; Elsevier, 2017; pp. 2–43.e19. ISBN 9780323357753. [Google Scholar]
- Abdelhalim, M.A.K.; Jarrar, B.M. Gold Nanoparticles Induced Cloudy Swelling to Hydropic Degeneration, Cytoplasmic Hyaline Vacuolation, Polymorphism, Binucleation, Karyopyknosis, Karyolysis, Karyorrhexis and Necrosis in the Liver. Lipids Health Dis. 2011, 10, 166. [Google Scholar] [CrossRef]
- Kundu, A.; Gali, S.; Sharma, S.; Kacew, S.; Yoon, S.; Jeong, H.G.; Kwak, J.H.; Kim, H.S. Dendropanoxide Alleviates Thioacetamide-Induced Hepatic Fibrosis via Inhibition of ROS Production and Inflammation in BALB/C Mice. Int. J. Biol. Sci. 2023, 19, 2630–2647. [Google Scholar] [CrossRef]
- Megahed, A.; Gadalla, H.; Filimban, W.A.; Albukhari, T.A.; Sembawa, H.; Bagadood, R.M.; Sindi, G.; Abdelhamid, F.M.; El-Boshy, M.E.; Risha, E.F. Hesperidin Ameliorates Thioacetamide-Induced Liver Fibrosis via Antioxidative and Anti-Inflammatory Mechanisms Targeting TGF-β/α-SMA Pathways in Rats. Int. J. Immunopathol. Pharmacol. 2024, 38, 3946320241309004. [Google Scholar] [CrossRef]
- Kozman, M.R.; Ewees, M.G.E.-D.; Ahmed, Y.M.; El-Shoura, E.A.M.; Alqahtani, S.M.; Ahmed, L.K.; Farghaly, O.A.; Sabry, M.; Ahmed, Y.H.; Abdel Rahman, F.E.-Z.S.; et al. Nicorandil Attenuates Thioacetamide Induced Liver Fibrosis via AMPK, SIRT1 and HIF1α Mediated Cellular Energy Homeostasis. Sci. Rep. 2025, 15, 43477. [Google Scholar] [CrossRef]
- Alamri, Z.Z. Protective and Therapeutic Effects of Apigenin on Thioacetamide-Induced Hepatotoxicity in Male Rats: Physiological and Morphological Study. Egypt. Liver J. 2024, 14, 15. [Google Scholar] [CrossRef]
- Georgiou-Siafis, S.K.; Tsiftsoglou, A.S. The Key Role of GSH in Keeping the Redox Balance in Mammalian Cells: Mechanisms and Significance of GSH in Detoxification via Formation of Conjugates. Antioxidants (Basel) 2023, 12, 1953. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Rodríguez, M.A.; Mendoza-Núñez, V.M. Oxidative Stress Indexes for Diagnosis of Health or Disease in Humans. Oxid. Med. Cell. Longev. 2019, 2019, 4128152. [Google Scholar] [CrossRef] [PubMed]
- Alkreathy, H.M.; Esmat, A. Lycorine Ameliorates Thioacetamide-Induced Hepatic Fibrosis in Rats: Emphasis on Antioxidant, Anti-Inflammatory, and STAT3 Inhibition Effects. Pharmaceuticals (Basel) 2022, 15, 369. [Google Scholar] [CrossRef]
- Gu, L.; Zhang, F.; Wu, J.; Zhuge, Y. Nanotechnology in Drug Delivery for Liver Fibrosis. Front. Mol. Biosci. 2021, 8, 804396. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.-S.; Chen, Y.-C.; Chou, C.-H.; Chuang, R.-F.; Sheen, L.-Y.; Chiu, C.-H. Hepatoprotection of Silymarin against Thioacetamide-Induced Chronic Liver Fibrosis: Hepatoprotective Mechanism of Silymarin against TAA. J. Sci. Food Agric. 2012, 92, 1441–1447. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, X.; Ding, C.; Gu, Y.; Liu, W. Dihydromyricetin Reverses Thioacetamide-Induced Liver Fibrosis through Inhibiting NF-κB-Mediated Inflammation and TGF-β1-Regulated of PI3K/akt Signaling Pathway. Front. Pharmacol. 2021, 12, 783886. [Google Scholar] [CrossRef]
- Shin, M.-R.; Lee, J.A.; Kim, M.; Lee, S.; Oh, M.; Moon, J.; Nam, J.-W.; Choi, H.; Mun, Y.-J.; Roh, S.-S. Gardeniae Fructus Attenuates Thioacetamide-Induced Liver Fibrosis in Mice via Both AMPK/SIRT1/NF-κB Pathway and Nrf2 Signaling. Antioxidants (Basel) 2021, 10, 1837. [Google Scholar] [CrossRef]
- Abdel-Rahman, R.F.; Fayed, H.M.; Mohamed, M.A.; Hessin, A.F.; Asaad, G.F.; AbdelRahman, S.S.; Salama, A.A.; Arbid, M.S.; Ogaly, H.A. Apigenin Role against Thioacetamide-Triggered Liver Fibrosis: Deciphering the PPARγ/TGF-β1/NF-κB and the HIF/FAK/AKT Pathways. J. Herbmed Pharmacol. 2023, 12, 202–213. [Google Scholar] [CrossRef]
- Gowifel, A.M.H.; Khalil, M.G.; Nada, S.A.; Kenawy, S.A.; Ahmed, K.A.; Salama, M.M.; Safar, M.M. Combination of Pomegranate Extract and Curcumin Ameliorates Thioacetamide-Induced Liver Fibrosis in Rats: Impact on TGF-β/Smad3 and NF-κB Signaling Pathways. Toxicol. Mech. Methods 2020, 30, 620–633. [Google Scholar] [CrossRef]
- Kabir, Nurul; Ali, Hamid; Ateeq, Muhammad; Bertino, Massimo F.; Shah, Muhammad Raza; Franzelc, Louis. Silymarin Coated Gold Nanoparticles Ameliorates CCl4-Induced Hepatic Injury and Cirrhosis through down Regulation of Hepatic Stellate Cells and Attenuation of Kupffer Cells. RSC Adv. 2014, 4, 9012–9020. [Google Scholar] [CrossRef]
- Abdelhalim, M.A.K.; Moussa, S.A.A.; Qaid, H.A.Y. The Protective Role of Quercetin and Arginine on Gold Nanoparticles Induced Hepatotoxicity in Rats. Int. J. Nanomedicine 2018, 13, 2821–2825. [Google Scholar] [CrossRef]
- Du, X.; Niu, R.; Liu, X.; Wu, F.; Yang, X.; Ma, X.; Zhang, J.; Zhou, H.; Shao, L.; Wang, S. Nanomedicines in the Treatment of Liver Fibrosis: A Review. Int. J. Nanomedicine 2025, 20, 9641–9665. [Google Scholar] [CrossRef] [PubMed]
- Padmanaban, S.; Baek, J.-W.; Chamarthy, S.S.; Chandrasekaran, S.; Samrot, A.V.; Gosu, V.; Park, I.-K.; Radhakrishnan, K.; Kim, D.-K. Nanoparticle-Based Therapeutic Strategies for Chronic Liver Diseases: Advances and Insights. Liver Res. 2025, 9, 104–117. [Google Scholar] [CrossRef]
- Enciso, N.; Amiel, J.; Fabián-Domínguez, F.; Pando, J.; Rojas, N.; Cisneros-Huamaní, C.; Nava, E.; Enciso, J. Model of Liver Fibrosis Induction by Thioacetamide in Rats for Regenerative Therapy Studies. Anal. Cell. Pathol. (Amst.) 2022, 2022, 2841894. [Google Scholar] [CrossRef] [PubMed]
- Nishi, K.; Yagi, H.; Ohtomo, M.; Nagata, S.; Udagawa, D.; Tsuchida, T.; Morisaku, T.; Kitagawa, Y. A Thioacetamide-Induced Liver Fibrosis Model for Pre-Clinical Studies in Microminipig. Sci. Rep. 2023, 13, 14996. [Google Scholar] [CrossRef]
Figure 1.
Experimental timeline of the fibrosis induction and therapeutical intervention.
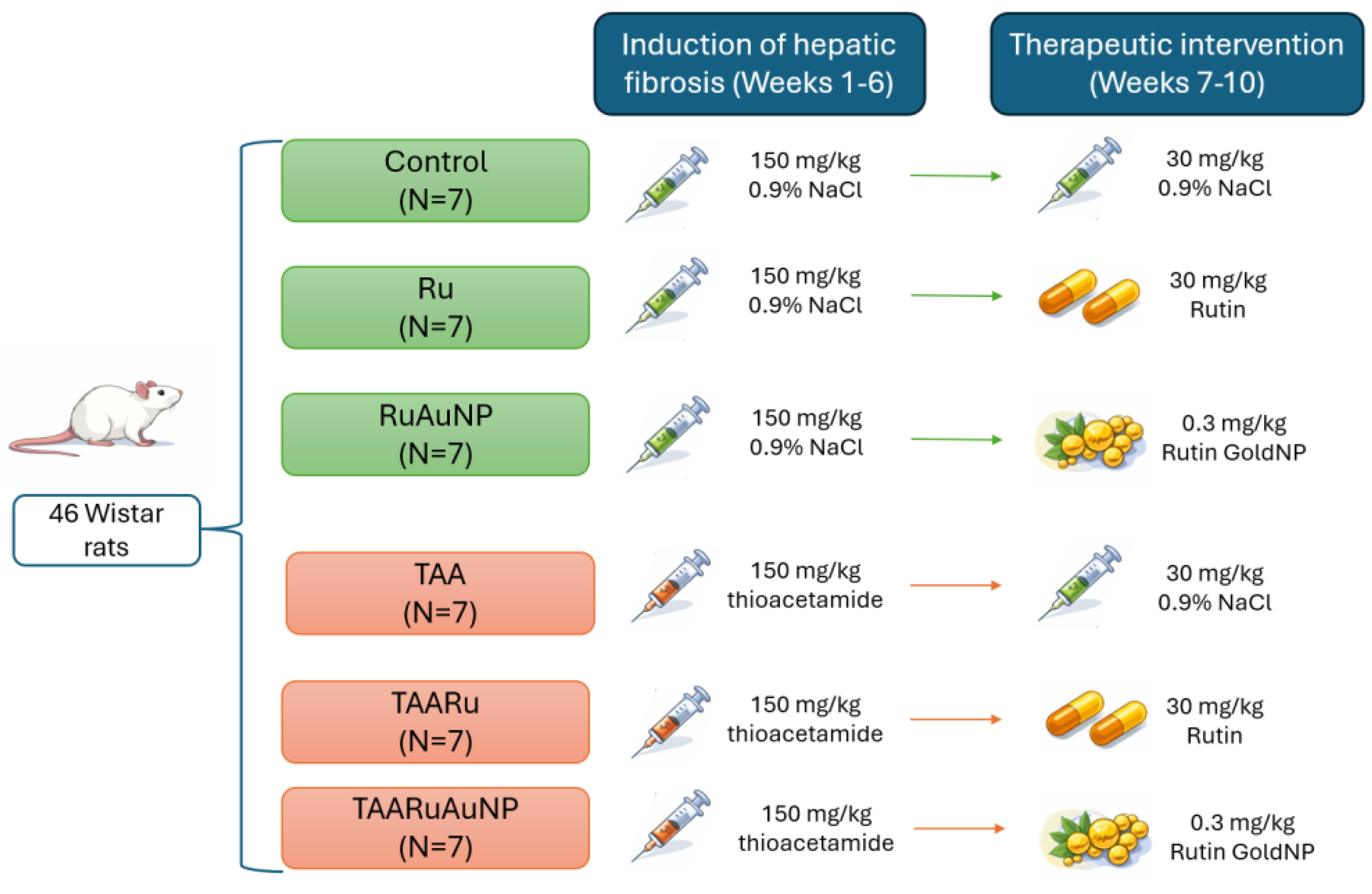
Figure 4.
Representative H&E-stained liver sections from the experimental groups. (a) Control: preserved hepatic architecture. (b) TAA: increased vascular wall thickness; the measured segment is indicated by black line. (c) Ru: vacuolar degeneration (arrowheads) and hepatocellular pigment accumulation (black arrow). (d) TAARu: persistence of vacuolar degeneration and pigment deposits after TAA exposure. (e) RuAuNP: dystrophic changes with regenerative features (arrowheads). (f) TAARuAuNP: similar dystrophic/regenerative changes (arrow), with additional areas of interest highlighted (circles). Images are representative of each group. Scale bar: 100 μm.
Figure 4.
Representative H&E-stained liver sections from the experimental groups. (a) Control: preserved hepatic architecture. (b) TAA: increased vascular wall thickness; the measured segment is indicated by black line. (c) Ru: vacuolar degeneration (arrowheads) and hepatocellular pigment accumulation (black arrow). (d) TAARu: persistence of vacuolar degeneration and pigment deposits after TAA exposure. (e) RuAuNP: dystrophic changes with regenerative features (arrowheads). (f) TAARuAuNP: similar dystrophic/regenerative changes (arrow), with additional areas of interest highlighted (circles). Images are representative of each group. Scale bar: 100 μm.
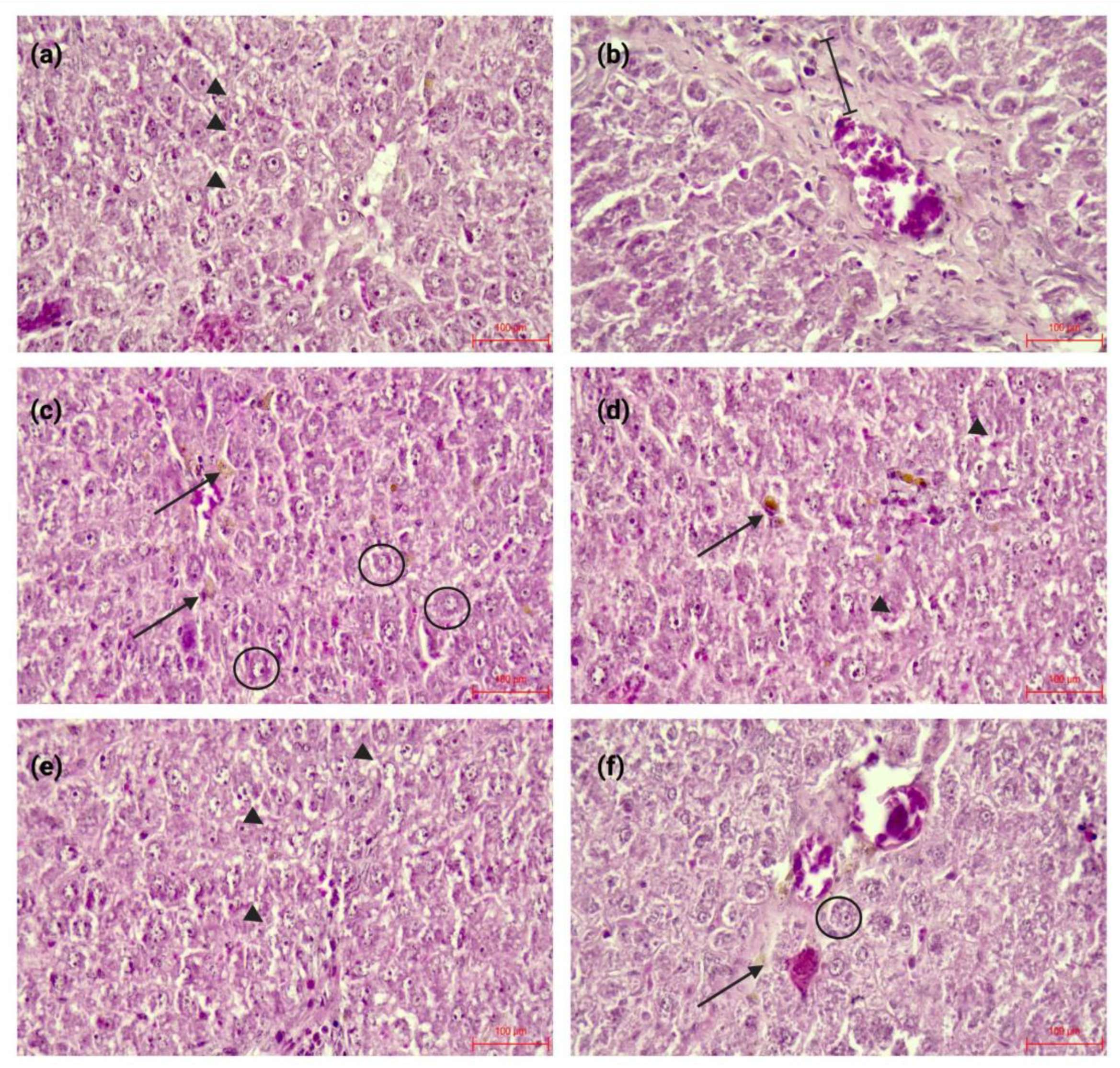
Figure 5.
Ultrastructural aspects of the liver in the control group. b: bile canaliculus; er: endoplasmic reticulum; g: glycogen; l: lipid droplet; m: mitochondria; n: nucleus.
Figure 5.
Ultrastructural aspects of the liver in the control group. b: bile canaliculus; er: endoplasmic reticulum; g: glycogen; l: lipid droplet; m: mitochondria; n: nucleus.

Figure 6.
Ultrastructural aspects of the liver in the TAA group. ap: autophagosome; b: bile canaliculus; c: collagen; er: endoplasmic reticulum; g: glycogen; l: lipid droplet; m: mitochondria; n: nucleus.
Figure 6.
Ultrastructural aspects of the liver in the TAA group. ap: autophagosome; b: bile canaliculus; c: collagen; er: endoplasmic reticulum; g: glycogen; l: lipid droplet; m: mitochondria; n: nucleus.
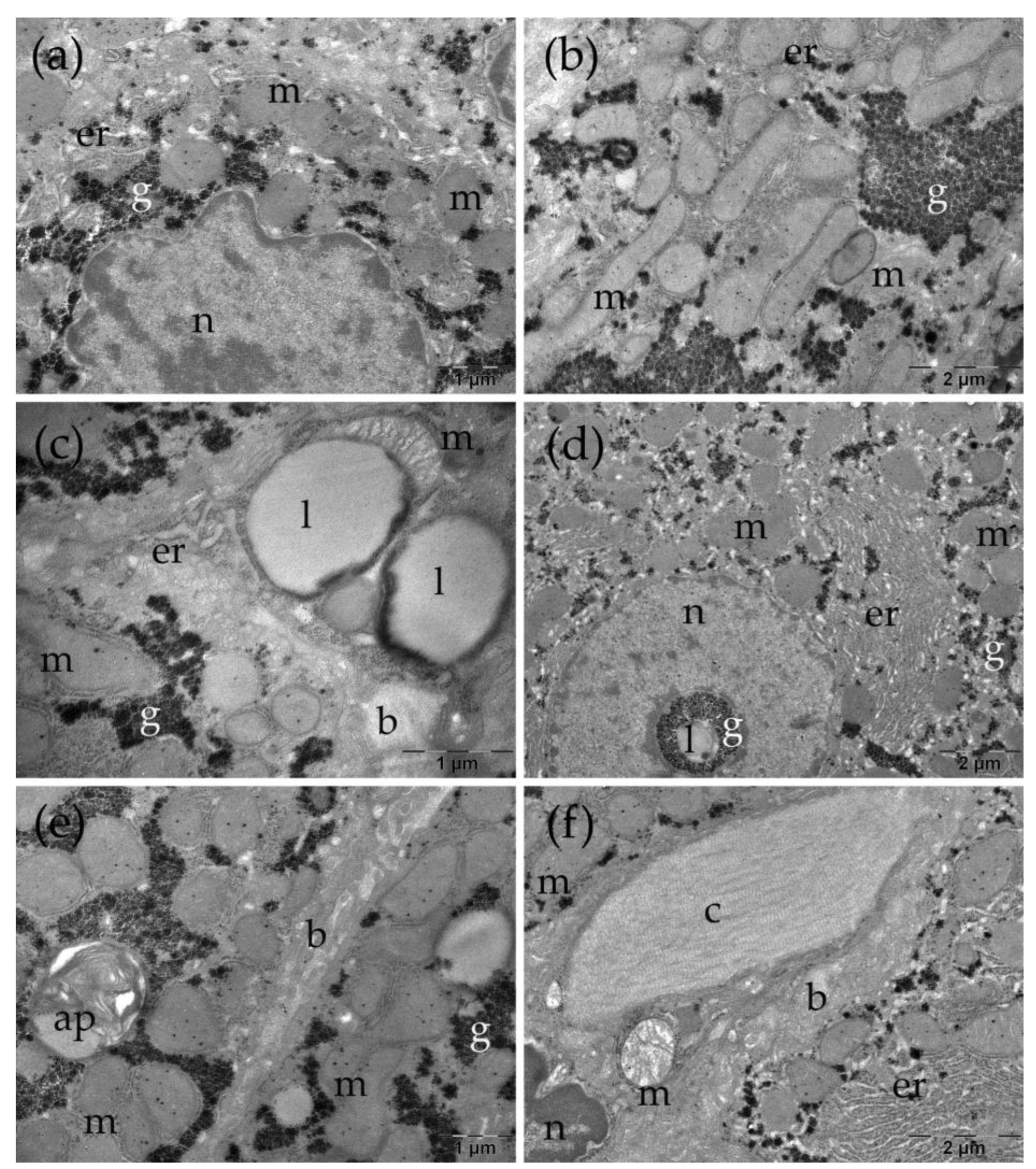
Figure 7.
Ultrastructural aspects of the liver in the TAARu group. b: bile canaliculus; c: collagen; er: endoplasmic reticulum; g: glycogen; l: lipid droplet; ly: lysosomes; m: mitochondria; n: nucleus.
Figure 7.
Ultrastructural aspects of the liver in the TAARu group. b: bile canaliculus; c: collagen; er: endoplasmic reticulum; g: glycogen; l: lipid droplet; ly: lysosomes; m: mitochondria; n: nucleus.
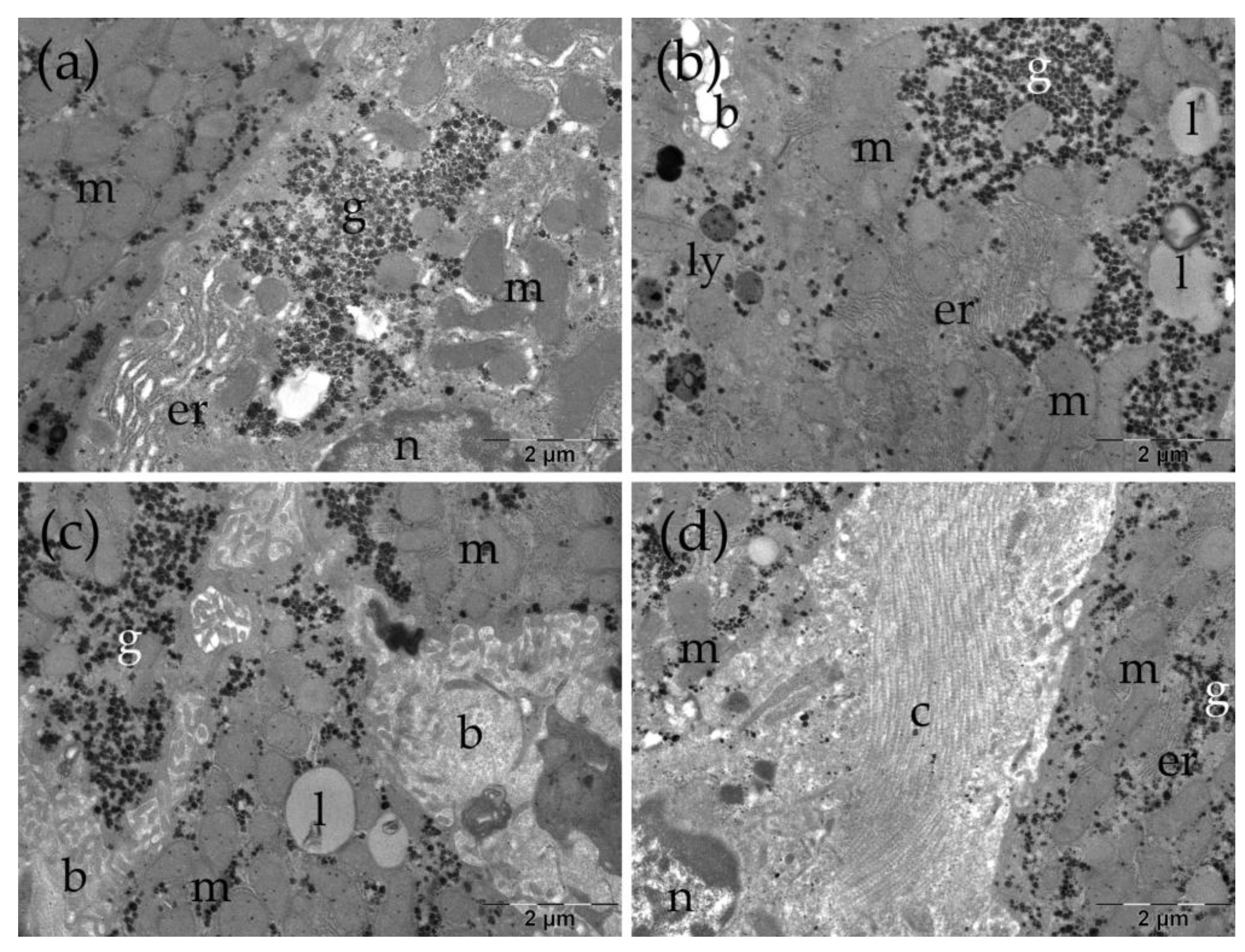
Figure 8.
Ultrastructural aspects of the liver in the TAARuAuNP group. b: bile canaliculus; c: collagen; er: endoplasmic reticulum; g: glycogen; l: lipid droplet; ly: lysosomes; m: mitochondria; n: nucleus.
Figure 8.
Ultrastructural aspects of the liver in the TAARuAuNP group. b: bile canaliculus; c: collagen; er: endoplasmic reticulum; g: glycogen; l: lipid droplet; ly: lysosomes; m: mitochondria; n: nucleus.
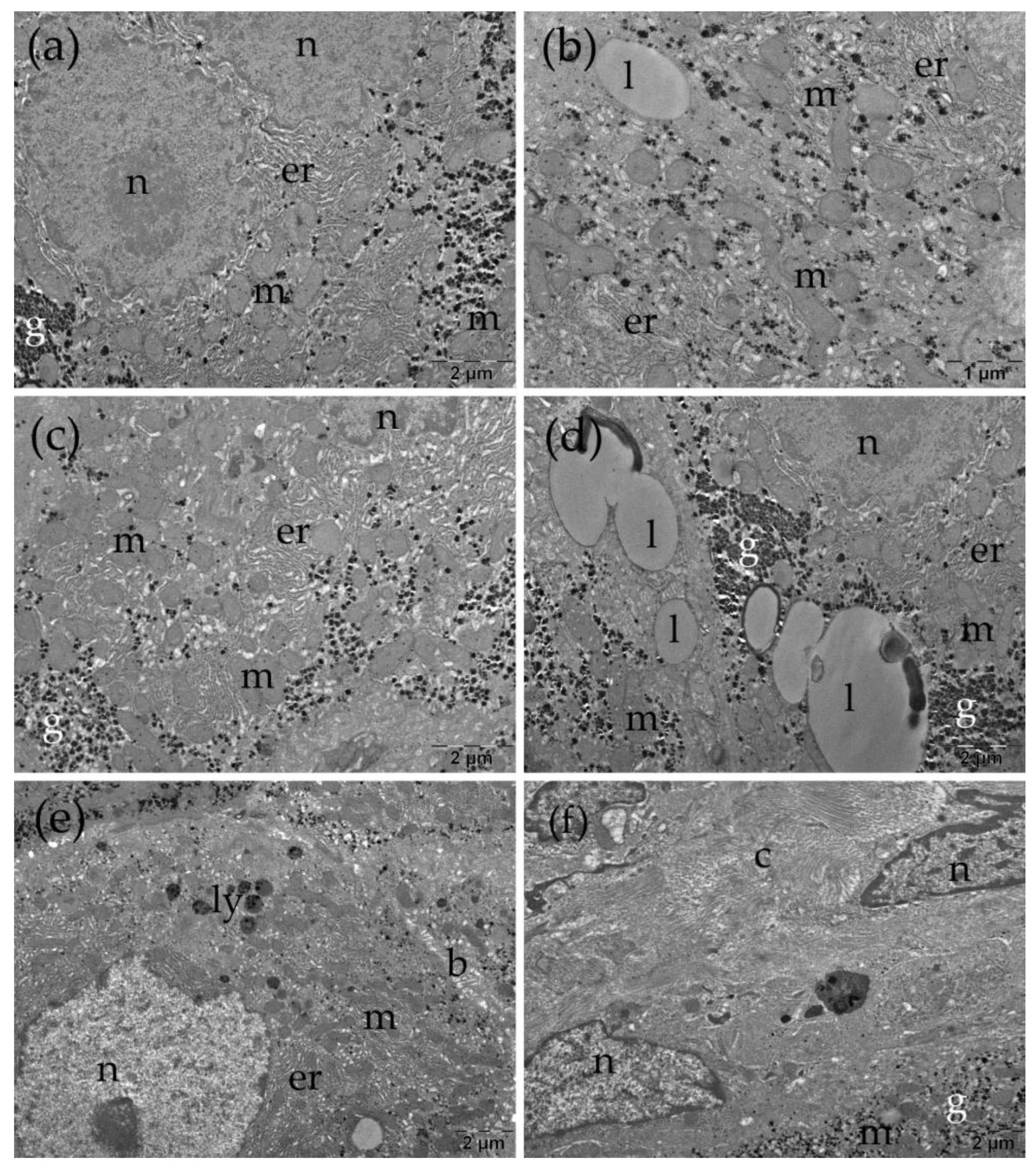
Figure 9.
Hepatic TGF-β levels across experimental groups. Groups: Control, TAAControl, Ru, TAARu, RuAuNP, TAARuAuNP. Data are presented as mean ± SD (n = 7). *** p < 0.001.
Figure 9.
Hepatic TGF-β levels across experimental groups. Groups: Control, TAAControl, Ru, TAARu, RuAuNP, TAARuAuNP. Data are presented as mean ± SD (n = 7). *** p < 0.001.
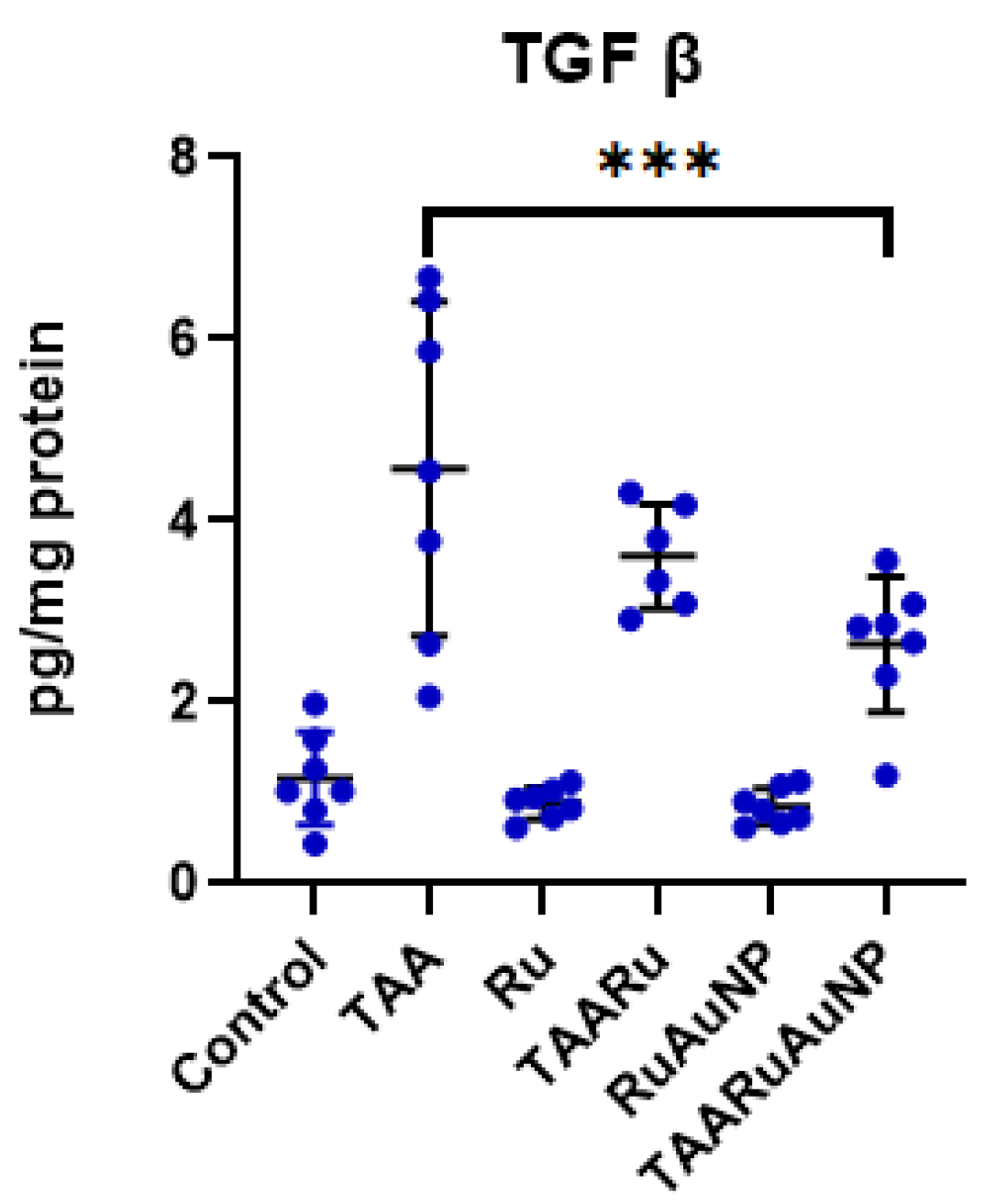
Figure 10.
Liver transaminase (AST/ALT) ratios across experimental groups. Groups: Control, TAA, Ru, TAARu, RuAuNP, TAARuAuNP. Data are presented as mean ± SD (n = 7). * p < 0.05; **** p < 0.0001.
Figure 10.
Liver transaminase (AST/ALT) ratios across experimental groups. Groups: Control, TAA, Ru, TAARu, RuAuNP, TAARuAuNP. Data are presented as mean ± SD (n = 7). * p < 0.05; **** p < 0.0001.
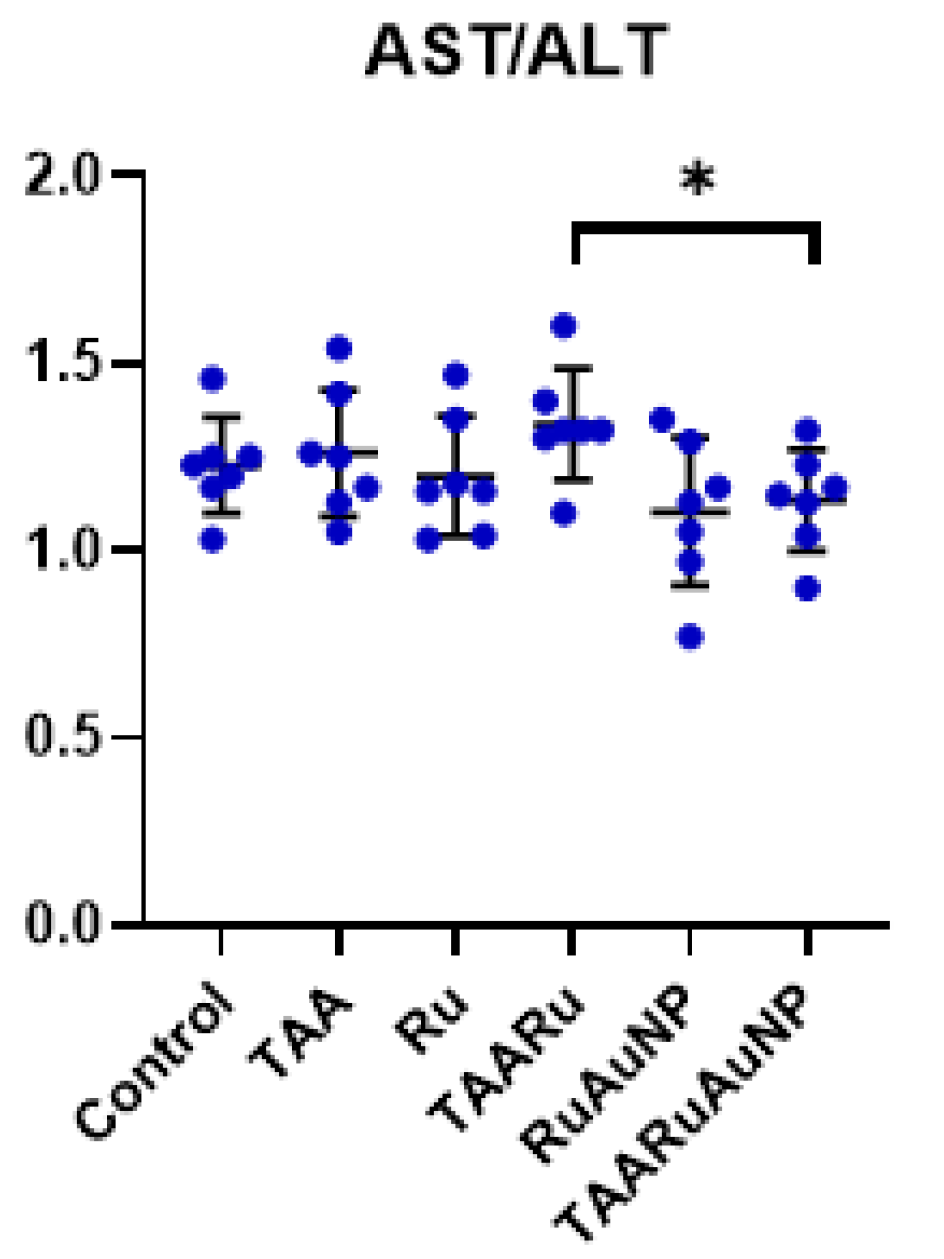
Figure 11.
Metabolic parameters across experimental groups. (a) Blood glucose; (b) total cholesterol; (c) triglycerides. Groups: Control, TAA, Ru, TAARu, RuAuNP, TAARuAuNP. Data are presented as mean ± SD (n = 7). * p < 0.05.
Figure 11.
Metabolic parameters across experimental groups. (a) Blood glucose; (b) total cholesterol; (c) triglycerides. Groups: Control, TAA, Ru, TAARu, RuAuNP, TAARuAuNP. Data are presented as mean ± SD (n = 7). * p < 0.05.
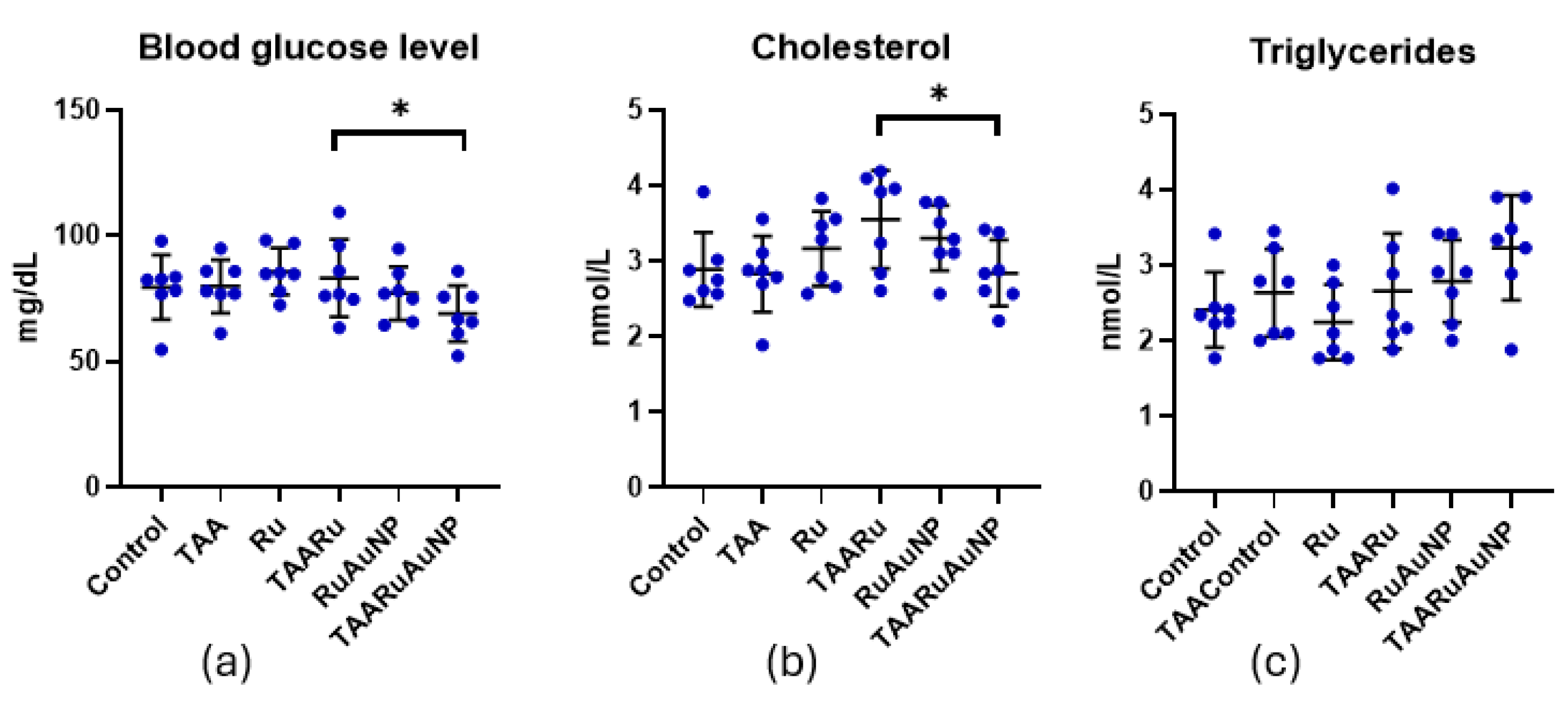
Figure 12.
Hepatic and serum malondialdehyde (MDA) levels across experimental groups. (a) Hepatic MDA; (b) serum MDA. Groups: Control, TAA, Ru, TAARu, RuAuNP, TAARuAuNP. Data are presented as mean ± SD (n = 7). ** p < 0.01; *** p < 0.001.
Figure 12.
Hepatic and serum malondialdehyde (MDA) levels across experimental groups. (a) Hepatic MDA; (b) serum MDA. Groups: Control, TAA, Ru, TAARu, RuAuNP, TAARuAuNP. Data are presented as mean ± SD (n = 7). ** p < 0.01; *** p < 0.001.
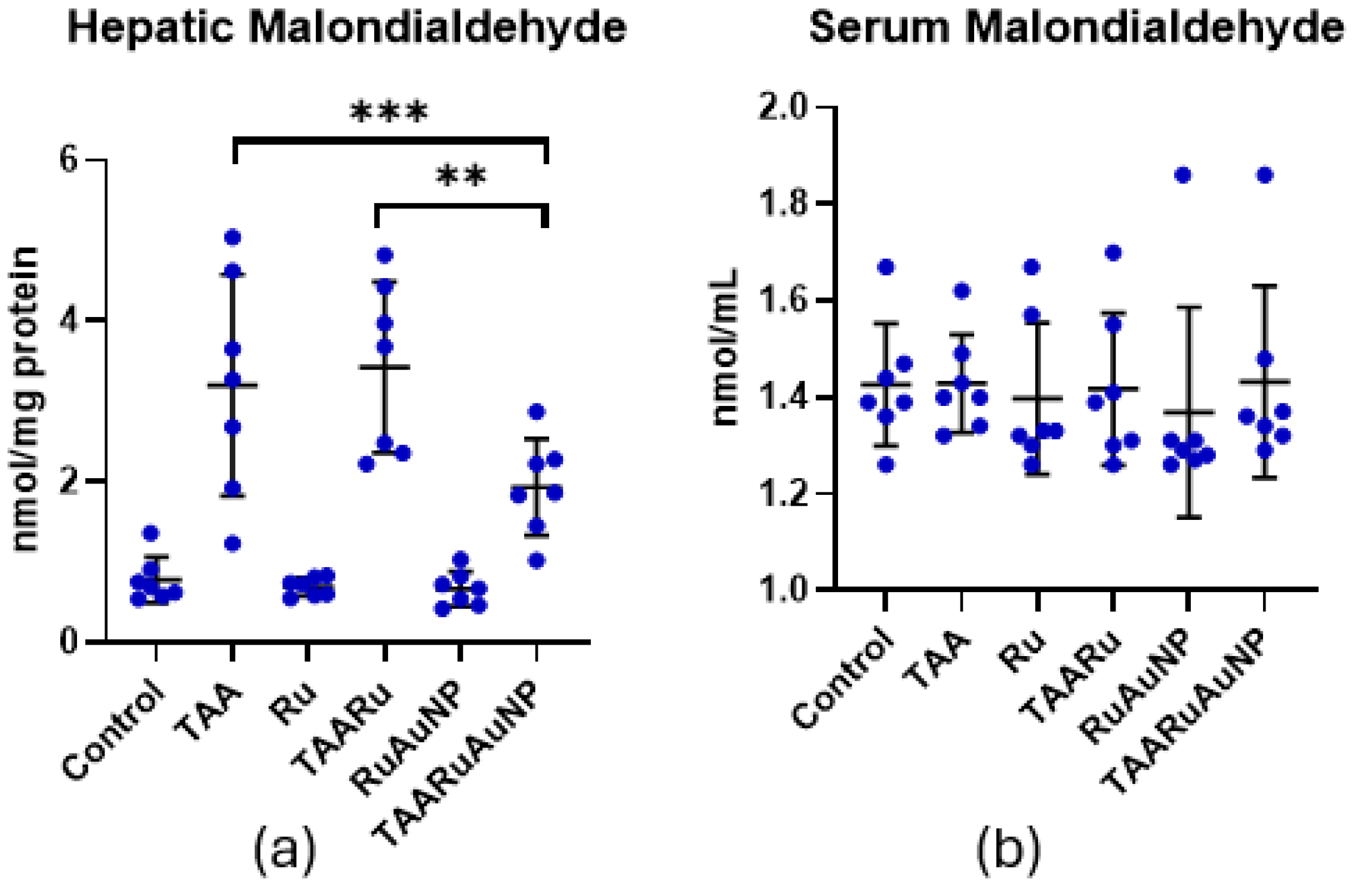
Figure 13.
Hepatic antioxidant defense markers across experimental groups. (a) GSH/GSSG ratio; (b) glutathione peroxidase (GPx) activity; (c) superoxide dismutase (SOD) activity. Groups: Control, TAA, Ru, TAARu, RuAuNP, TAARuAuNP. Data are presented as mean ± SD (n = 7). * p < 0.05; ** p < 0.01; *** p < 0.001; **** p < 0.0001.
Figure 13.
Hepatic antioxidant defense markers across experimental groups. (a) GSH/GSSG ratio; (b) glutathione peroxidase (GPx) activity; (c) superoxide dismutase (SOD) activity. Groups: Control, TAA, Ru, TAARu, RuAuNP, TAARuAuNP. Data are presented as mean ± SD (n = 7). * p < 0.05; ** p < 0.01; *** p < 0.001; **** p < 0.0001.
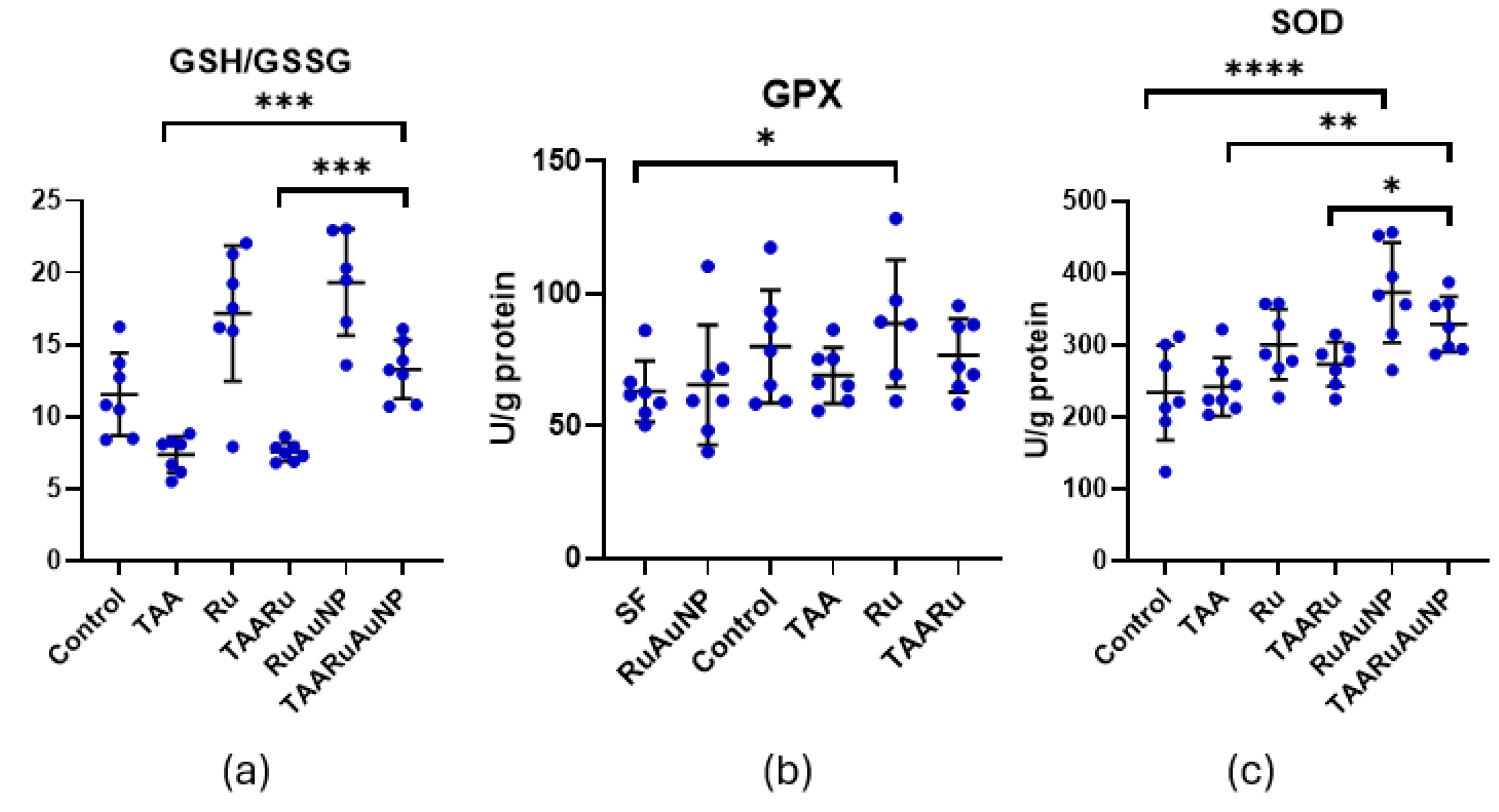
Figure 14.
Hepatic pro-inflammatory cytokine levels across experimental groups. (a) IL-1β; (b) IL-6; (c) TNF-α. Groups: Control, TAA, Ru, TAARu, RuAuNP, TAARuAuNP. Data are presented as mean ± SD (n = 7). ** p < 0.01; *** p < 0.001.
Figure 14.
Hepatic pro-inflammatory cytokine levels across experimental groups. (a) IL-1β; (b) IL-6; (c) TNF-α. Groups: Control, TAA, Ru, TAARu, RuAuNP, TAARuAuNP. Data are presented as mean ± SD (n = 7). ** p < 0.01; *** p < 0.001.
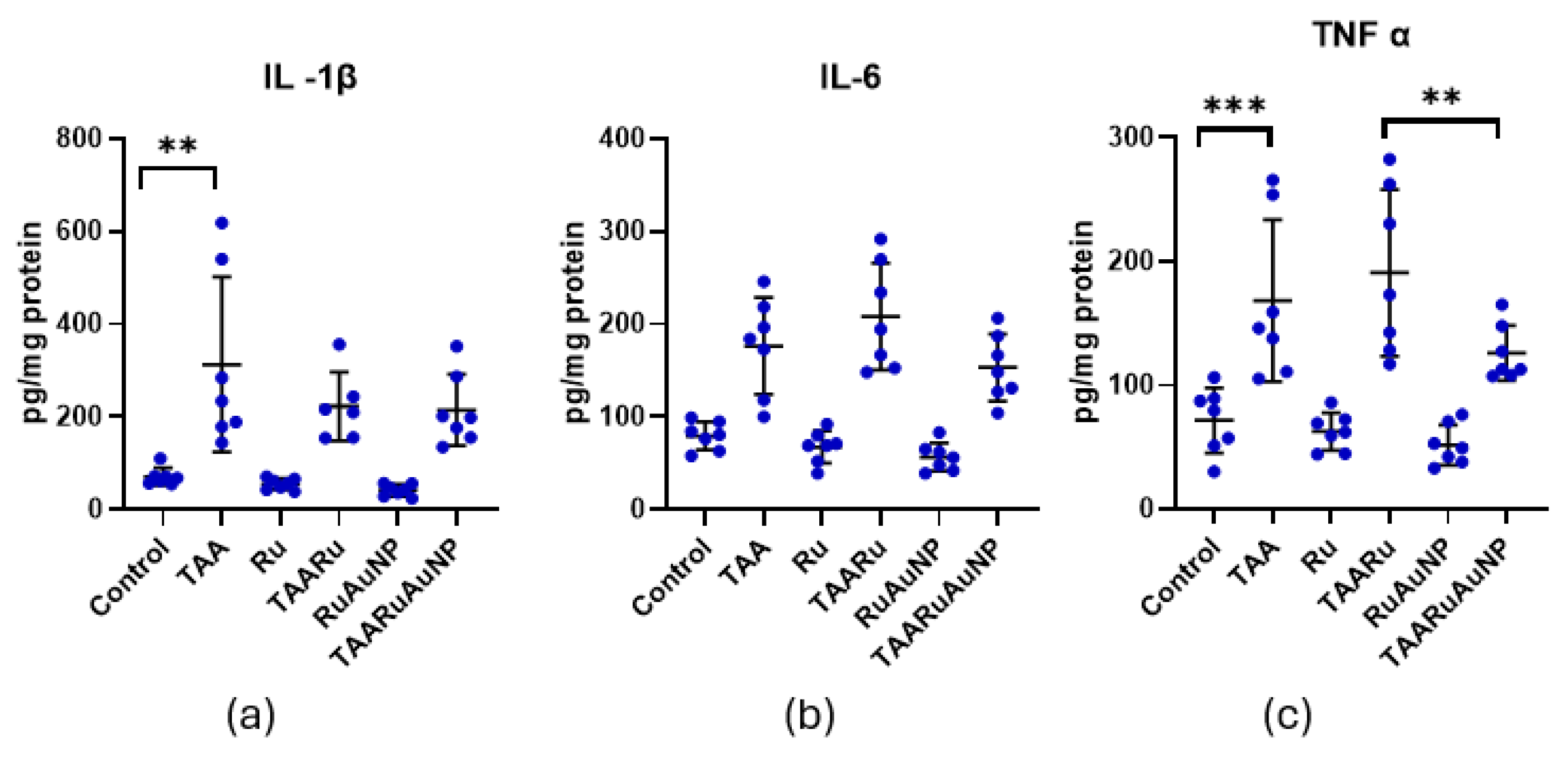

Table 1.
Level of liver transaminases across the experimental groups.
| Group | AST (U/I) | ALT (U/I) |
|---|---|---|
| Control | 76.57 +/-5.966 | 62.70+/-4.502 |
| TAA | 109.1 +/-8.977**** | 87.78+/-11.69**** |
| Ru | 80.89+/-9.961 | 67.54+/-3.228 |
| TAARu | 107.8+/-14.41 | 81.01+/-10.23 |
| RuAuNP | 73.50+/-7.652 | 67.69+/-8.469 |
| TAARuAuNP | 102.7+/-12.85 | 90.56+/-4.738 |
**** p≤ 0.0001.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



