
Submitted:
26 January 2026
Posted:
27 January 2026
You are already at the latest version
Abstract
The medical mushroom Ophiocordyceps sinensis (Caterpillar Fungus) is one of the most popular medicines in Asian traditional medical systems for its pleiotropic indications, including an ability to enhance “vitality”. However, the polyvalent adaptogenic potential of O. sinensis has not been systematically proven, and a consolidated mechanistic synthesis integrating both network predictions and experimental validation is lacking. The primary aim of this narrative review was to provide a rationale for the pleiotropic therapeutic effects of two traditionally used botanicals from the Fungi Kingdom (O.sinensis and the related fungus Cordyceps militaris (Orange Cordyceps) and their primary metabolite, cordycepin, using a network pharmacology approach to support or refute their adaptogenic potential. The secondary aim was to identify the source of temporary failures in marketing authorization for TCM products in Europe, arising from an ambiguous fundamental approach to the action of the same natural product across Western and Oriental medical systems. and provide a possible, reasonable solution. This review, for the first time show that: • O. sinensis, C. militaris, and 3-deoxyadenosine share a common adaptogenic mechanism for maintaining cellular and integrated biological system functions homeostasis. • The systems-level adaptogenic mechanism of these fungi is characterized by their ability to modulate multiple interconnected biological networks rather than acting on a single target. • This is in line with holistic TCM and Ayurvedic concepts and the modern concept of the pleiotropic therapeutic activity of adaptogens, particularly of O. sinensis, including cordycepin. • Their polyvalent pharmacological activity is related to the high content of nucleotides adenosine and cordycepin in these fungi, and to their involvement in metabolism and in signaling via protein kinases (PKB/Akt, PI3K, AMPK, and MAPK), which regulate diverse cellular processes and physiological functions • Network pharmacology studies identify multi-target pathways, including convergent hubs, such as PI3K-Akt, AMPK–mTOR, MAPK, apoptosis, and Nrf2. • By influencing pathways associated with immune regulation, mitochondrial function, and metabolic adaptation, they orchestrate a holistic response that enhances the organism's resilience to various stressors. • Validation studies confirm predictions across chronic obstructive pulmonary disease, pulmonary arterial hypertension, cancer, obesity, influenza, and immunogenicity. • This integrative effect is especially significant in the context of complex disorders, where multifactorial interventions are necessary for effective therapeutic outcomes. • The review reveals controversy regarding the bioavailability of cordycepin in vivo and its concentration in vitro studies, raising the hypothesis that cordycepin may act as a driver, triggering the organism's adaptive stress response in stress-induced and aging-related diseases. This review clarifies conceptual and regulatory barriers to recognizing resilience-supporting interventions and informs future regulatory innovation. The review for the first time suggests establishing a regulatory category for resilience-supporting physiological modulators that could align food and drug regulation in the EU with contemporary systems biology, thereby complementing EFSA, EMA, FDA, and Asian authorities. Overall, from a scientific point of view, both reductionistic and holistic integrative approaches are complementary and essential for understanding the mechanism of action of adaptogens and for the discovery of new effective therapeutics. Future research should focus on rigorous quality control and regulatory alignment to support evidence-based clinical use.
Keywords:
1. Introduction
2. Results
2.1. Bioactive Compounds of O. sinensis and C. militaris Fungi and Their Pharmacological Activity
2.2. Nucleosides Adenosine and Cordycepin as Mediators of Adaptive Stress Response Andanti-Fatigue Activity of O. sinensis, and C. militaris
2.3. Efficacy and Safety of Ophiocordyceps Sinensis and Cordyceps Militaris in Human Subjects
2.3.1. Traditional Use
2.3.2. Clinical Studies
Systematic Reviews and Meta-Analyses of Randomized Clinical Trials

| References | Main preparations studied | Indications (systematic reviews) | No. of SRs / Meta- analyses | Approx. no. of RCTs (patients) | Consistency of clinical effects | Methodological quality Certainty of evidence | Key limitations | EMA WEU relevance |
|---|---|---|---|---|---|---|---|---|
| Liu et al.,2024; Wu et al., 2025 Pu et al., 2024; Tao et al., 2024; Xue et al., 2024; [137,138,139,140,141] |
Fermented mycelium (Bailing, Jinshuibao, Zhiling, others) | Dialysis (HD/PD), DKD, CKD, CA-AKI prevention, renal dysfunction | 5 | >210 RCTs (>16,000 pts, overlapping); | Consistent improvement in renal and inflammatory surrogate markers (Scr, BUN, CRP, proteinuria); reduced CA-AKI incidence when used preventively | Low to very low (GRADE); majority of SRs critically low (AMSTAR-2) | Open-label RCTs; short follow-up; surrogate endpoints; heterogeneous preparations | Does not meet WEU (efficacy signal present, but insufficient robustness and preparation definition) |
| Wang et al, 2024; Yu et al., 2019; Ma & Jin, 2024 [142,143,144] |
Bailing capsule, cultivated mycelia, mixed CS preparations | COPD, lung cancer (adjunctive), | 3 | ~80–100 RCTs (>5,000 pts) | Consistent adjunctive benefits (lung function, QoL, immune markers, tumour response rate) | Low to moderate | Lack of blinding; all trials conducted in China; unclear allocation concealment; adjunctive designs only | Does not meet WEU (methodological and EU-use limitations) |
| Welch et al.,2023; Dewi & Khemtong, 2025* [145,146] | Supplements (varied) | Healthy volunteers/exercise performance | 2 | 7 RCTs (286 pts) | Inconsistent, small effects | Low | Small samples; heterogeneous outcomes; limited clinical relevance | Not relevant for WEU |
- Trials were almost exclusively conducted in China
- Most studies were small and short-term
- Blinding and placebo control were largely absent
- Outcomes were frequently surrogate or supportive rather than definitive clinical endpoints
- Insufficient methodological robustness
- Absence of EU-based medicinal use documentation
- Lack of a single, standardized herbal substance or preparation
- Predominant reliance on surrogate outcomes
Clinical Evidence for Anti-Fatigue Effects of Ophiocordyceps Sinensis and Cordyceps Sinensis: Implications for EMA Well-Established Use
2.4. Systems-Level Mechanisms of Ophiocordyceps sinensis, Cordyceps militaris, and Cordycepin: An Integrative Network-Pharmacology and Experimental Evidence Review
- Immune modulation via TLR4/TNF-α [184]
- Cancer apoptosis via PI3K–Akt and caspase regulation [189]
- Cytokine normalization and lung restoration in COPD [176]
- PAH through apoptosis and vascular remodeling repair [193]
- Obesity/metabolic syndrome via AKT1/MAPK14/GSK3B [194]
- HBV vaccine response enhancement [179]
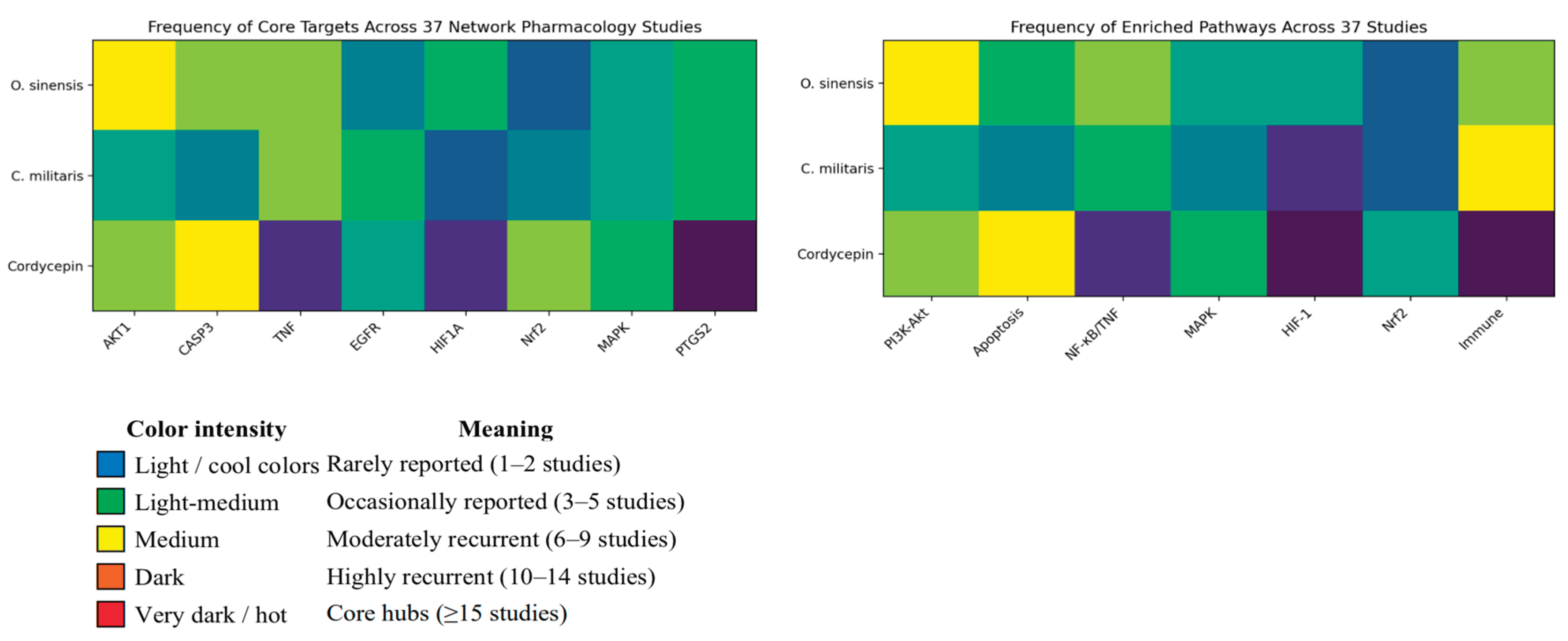
- O. sinensis exhibits broad-spectrum organ support and multi-pathway modulation, especially in chronic diseases and viral infections. The most defensible, network-anchored claims are around kidney support/adjunct in chronic kidney diseases, inflammation/oxidative stress, and respiratory immune modulation—with some clinical meta-analytic support but still needing higher-quality trials [137,140] .
- C. militaris shows targeted cytotoxicity and immune activation, making it more suitable for oncology and immunotherapy applications. C. militaris preparations, explicitly characterized by high content of cordycepin, have the strongest network-mechanistic case for anticancer mechanisms, immune modulation (TLR4–TNF, macrophage polarization), and metabolic/urate axes. The evidence is growing, but disease-specific clinical endpoints remain limited [184,185,187].
| Feature | O. sinensis | C. militaris | Cordycepin |
|---|---|---|---|
| Main bioactive constituents | Adenosine / 2′-deoxyadenosine, ergosterol, adenosine, glutamine, diverse peptides | Cordycepin (3’-deoxyadenosine), adenosine, polysaccharides, ergosterol | Cordycepin |
| Dominant validated indications | COPD, lung cancer, CKD, ischemic stroke, diabetic nephropathy, UV injury, vaccine adjuvant | Immune modulation, lung cancer, COPD, gouty nephropathy, SARS-CoV-2 | Cancer, obesity, depression, PAH, immunosuppression, glioblastoma |
| Main predicted indications (*) | Liver cancer, DN, hypoxia, depression, PCOS, influenza | COVID-19 inflammation | Cancer, Alzheimer’s, kinase inhibition |
| Signature pathways | PI3K-Akt, TNF, AGE-RAGE, IL-17, chemokine, apoptosis, lipid metabolism | TLR, inflammatory lipid pathways, apoptosis, oxidative stress, metabolic regulation | Apoptosis, AMPK-mTOR, PI3K-Akt, Nrf2, Wnt/β-catenin, metabolic reprogramming |
| Predicted (*) pathways | AGE-RAGE, VEGF, PI3K-Akt, FOXO, neurotrophic, inflammatory | A3AR, JNK, AMPK, AKT, MAPKs, FOXO3 | Apoptosis, neurodegeneration, and kinase inhibition |
| Core hubs repeatedly validated | AKT1, CASP3, PTGS2, HIF1A, NFKB1, VEGFA, PTEN, STING, CXCR4, | TLR4, TNF, EGFR, IDH1, PTGS2, ABCG2, p53 | AKT1, CASP3, EGFR, Nrf2, GSK3β, β-catenin, FOXO, MAPKs |
| Core predicted hubs (*) | TNF, MAPK1, EGFR, IL6, VEGFA, ACE, CASP3 | NR3C1, thrombin, inflammatory kinases | A3AR, JNK, AMPK, AKT, MAPKs, FOXO3 |
| Key validated differences | immune-pulmonary-renal axis, lipid/steroid mediation, inflammatory microenvironment remodeling | immune activation, metabolic enzymes, antiviral/anticancer lipid-nucleoside networks | intracellular stress signaling, kinase control, apoptosis, metabolic rewiring |
| Signaling Pathway | Key Genes Involved | Physiological Role |
|---|---|---|
| AMPK pathway | AMPK, SIRT1, MTOR | Energy sensing, lifespan extension |
| Sirtuin signaling | SIRT1, FOXO3, PARP1 | Regulates stress response, aging, |
| mTOR signaling | MTOR, AMPK, SIRT1, TP53 | Nutrient sensing, autophagy, and protein synthesis |
| DNA damage response | TP53, PARP1 | Genomic stability, cancer prevention |
2.5. Food and Drug Regulation of Ophiocordyceps sinensis and Cordyceps militaris in Western and Oriental Worlds
Resilience Biology, Inflammation, and Adaptogens: An Umbrella Review of Biological Evidence and Regulatory Frameworks Across Europe, the United States, and Asia
| Dimension | European Union | United States | Asia (China, Japan, Korea, India) |
|---|---|---|---|
| Adaptogen term | Not recognized | Informally tolerated | Explicitly recognized |
| Regulatory model | Disease- & nutrient-centric | Risk-based consumer access | Systems & functional medicine |
| Acceptable claim style | Narrow, mechanistic | Structure–function | Functional, pattern-based |
| Stress-related claims | Rejected | Allowed with disclaimer | Allowed |
| Traditional use | Secondary | Optional | Central |
| Dimension | Western regulatory paradigm (EFSA / EMA) | Traditional medical paradigms (TCM, Ayurveda, etc.) |
|---|---|---|
| Primary goal | Treat or prevent a defined disease | Restore balance and resilience |
| Model | Reductionist, mechanistic | Systems-based, integrative |
| Body concept | Sum of organs and pathways | Dynamic, interconnected whole |
| Health | Absence of pathology | Capacity to adapt and recover |
| Evidence | Isolated endpoints, biomarkers | Pattern recognition, outcomes over time |
| Disease vs dysfunction | Requires: o A defined pathological condition o Clear causal pathway o Measurable, reproducible endpoints Works well for: o Inflammation o Deficiency diseases o Infections |
Focuses on: o Functional imbalance before disease o Fatigue, low vitality, stress intolerance o Recovery capacity |
| Evidence hierarchy mismatch | • Standardized extracts • Dose–response relationships • Single or narrow endpoints |
• Multi-target effects • Long-term use • Context-dependent outcomes • Synergistic formulations |
3. Discussion
3.1. An Integrative Network-Pharmacology and Systems-Level Mechanisms of Ophiocordyceps sinensis, Cordyceps militaris, and Cordycepin
3.1.1. Convergence on a Conserved Stress-Adaptive Signaling Core
3.1.2. Species-Level Pharmacology Reflects Network Breadth, Not Redundancy
3.1.3. Cordycepin Acts as a Dominant Molecular Effector Within Broader Fungal Networks
3.1.4. Validated Versus Predicted Networks Define Evidence Tiers
3.1.5. Implications for Natural-Product Systems Pharmacology
3.1.6. Limitations
3.1.7. Future Perspectives
- Disease-targeted bioinformatic-based network analysis reveals the molecular mechanisms of action, common molecular targets of active compounds, and stimulus-response coupling signaling pathways [57,60,103,141,145,176,177,179,180,181,182,183,184,185,186,187,188,189,190,191,192,193,194,195,196,197,198,199,200,201,202,203,204,205,206,207,208,209,210,211]. Such study design and methodology do not account for synergistic and antagonistic interactions in networks induced by various constituents of the complex, multi-component extracts, which can lead to unexpected outcomes, e.g., [249,250]
- Non-targeted transcriptome-wide microarray profiling of gene expression-based experiments following integrated metabolomics and network analysis, revealing all molecular targets of active compounds and their response, coupling signaling pathways (xx). The results of these studies can predict unknown physiological functions, health consequences, and therapeutic indications due to synergistic and antagonistic interactions [173,175,249,250]
- Currently, all conducted Network pharmacology studies on Cordiceps are limited to disease-targeted investigations of Chinese TCM prescriptions for the treatment of kidney diseases and related disorders. Non-targeted transcriptome-wide microarray profiling of gene expression-based experiments can lead to the discovery of new therapeutic indications and functional claims.
3.2. Nucleosides Adenosine and Cordycepin as Mediators of Adaptive Stress Response and Anti-Fatigue Activity of O. sinensis, and C. militaris
3.3. Western Regulatory vs Traditional Medical Paradigms
3.4. Resilience Biology as the Missing Regulatory Construct
3.4.1. Why a Hybrid EU–Asia Model Is Scientifically Justified
Resilience-Supporting Physiological Modulators as a Regulatory Bridge
Regulatory Language as a Determinant of Scientific Visibility
3.4.2. Outlook and Future Directions
From Disease Regulation to Health Maintenance
Implications for Regulatory Science
- Formal recognition of homeostasis and recovery as legitimate physiological outcomes
- Acceptance of systems-level functional endpoints alongside classical biomarkers
- Development of claim guidance specific to resilience-supporting functions
Implications for Research and Industry
Concluding Perspective
4. Materials and Methods
4.1. Literature Search
4.2. Data Extraction
4.3. Data Integration
5. Conclusions
- O. sinensis, C.militaris, and cordycepin share a common adaptogenic mechanism of maintenance of cellular and integrated biology system functions homeostasis.
- The systems-level adaptogenic mechanism of these fungi is characterized by their ability to modulate multiple interconnected biological networks rather than acting on a single target.
- This is in line with TCM and Ayurveda holistic concepts and the modern concept of pleiotropic therapeutic activity of adaptogens and particularly of O. sinensis.
- The review reveals controversy regarding the bioavailability of cordycepin in vivo and its concentration in vitro studies, raising the hypothesis that cordycepin may act as a driver, triggering the organism's adaptive stress response in stress-induced and aging-related diseases.
- Nucleosides, adenosine and cordycepin, along with other adaptogenic botanical metabolites (steroids and phenolics), contribute to the maintenance of cellular and integrated biology system functions homeostasis.
- Network pharmacology studies identify multi-target pathways, including convergent hubs, such as PI3K-Akt, AMPK–mTOR, MAPK, apoptosis, Nrf2 and AMPK–SIRT1–PGC-1α pathways.
- By influencing pathways associated with immune regulation, mitochondrial function, and metabolic adaptation, they orchestrate a holistic response that enhances the organism's resilience to various stressors.
- This integrative effect is especially significant in the context of complex disorders, where multifactorial interventions are necessary for effective therapeutic outcomes.
- This is the first comparative meta-analysis of validated vs predicted effects of O. sinensis, C.militaris and species-level versus cordycepin-driven mechanisms.
- Validation studies confirm predictions across chronic obstructive pulmonary disease, pulmonary arterial hypertension, cancer, obesity, influenza, and immunogenicity.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Acknowledgments
AI Assistance Statement
References
- Taxonomy Browser: Ophiocordyceps sinensis; NCBI, National Library of Medicine: Bethesda, MD, USA. Available online: https://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi?mode=Info&id=72228 (accessed on 21 January 2026).
- Taxonomy Browser: Cordyceps militaris.; NCBI, National Library of Medicine: Bethesda, MD, USA. Available online: https://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi?command=show&mode=node&id=73501&lvl= (accessed on 21 January 2026).
- Botanical Drug Development. Guidance for Industry. FDA: Silver Spring, MD, USA, 2016. Available online: https://www.fda.gov/files/drugs/published/Botanical-Drug-Development--Guidance-for-Industry.pdf (accessed on 21 January 2026).
- Ahmadjian, V.; Alexopoulos, C.J.; Moore, D. Fungus. Encyclopaedia Britannica. 2025. Available online: https://www.britannica.com/science/fungus (accessed on 21 January 2026).
- Moore, D. Outline of classification of fungi. Encyclopaedia Britannica. 2025. Available online: https://www.britannica.com/science/fungus/Outline-of-classification-of-fungi (accessed on 21 January 2026).
- Petruzzello, M. Cordyceps. Encyclopaedia Britannica. 2025. Available online: https://www.britannica.com/science/cordyceps (accessed on 21 January 2026).
- Winkler, D. Cordyceps sinensis (Berk.) Sacc.: Economy, Ecology, and Ethno-Mycology of Yartsa Gunbu, a Medicinal Fungus Endemic for the Tibetan Plateau. Int. J. Med. Mashrooms 2005, 7, 481–482. [Google Scholar] [CrossRef]
- Winkler, D. Caterpillar Fungus Production and Sustainability on the Tibetan Plateau and in the Himalayas. Asian Med. 2009, 5, 291–316. Available online: https://www.researchgate.net/publication/233695593_Caterpillar_Fungus_Ophiocordyceps_sinensis_Production_and_Sustainability_on_the_Tibetan_Plateau_and_in_the_Himalayas. [CrossRef]
- Dong, C.; Yao, Y.-J. Comparison of Some Metabolites Among Cultured Mycelia of Medicinal Fungus, Ophiocordyceps sinensis (Ascomycetes) from Different Geographical Regions. International Journal of Medicinal Mushrooms 2010, 12, 287–297. Available online: https://www.researchgate.net/publication/270471851_Comparison_of_Some_Metabolites_Among_Cultured_Mycelia_of_Medicinal_Fungus_Ophiocordyceps_sinensis_Ascomycetes_from_Different_Geographical_Regions. [CrossRef]
- Kunwar, R.M.; Mahat, L.; Acharya, R.P.; Bussmann, R.W. Medicinal plants, traditional medicine, markets and management in far-west Nepal. J. Ethnobiol. Ethnomed. 2013, 9, 24. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Cheng, H.; Bussmann, R.W.; Guo, Z.; Liu, B.; Long, C. An ethnobotanical survey of edible fungi in Chuxiong City, Yunnan, China. J. Ethnobiol. Ethnomed. 2018, 14, 42. [Google Scholar] [CrossRef]
- Wei, Y.; Zhang, L.; Wang, J.; Wang, W.; Niyati, N.; Guo, Y.; Wang, X. Chinese caterpillar fungus (Ophiocordyceps sinensis) in China: Current distribution, trading, and futures under climate change and overexploitation. The Sci. Total Environ. 2021, 755, 142548. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.S.; Halpern, G.M.; Jones, K. The scientific rediscovery of an ancient Chinese herbal medicine: Cordyceps sinensis: Part I. J. Altern. Complement. Med. 1998, 4, 289–303. [Google Scholar] [CrossRef]
- Zhu, J.S.; Halpern, G.M.; Jones, K. The scientific rediscovery of a precious ancient Chinese herbal regimen: Cordyceps sinensis: part II. Part II. J. Altern. Complement. Med. 1998, 4, 429–457. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Y.; Shiao, M.S. Pharmacological functions of Chinese medicinal fungus Cordyceps sinensis and related species. Food Drug Anal. 2000, 8, 15. [Google Scholar] [CrossRef]
- Chen, P.X.; Wang, S.; Nie, S.; Marcone, M. Properties of Cordyceps sinensis: A review. J. Funct. Foods 2013, 5, 550–569. [Google Scholar] [CrossRef]
- Yue, K.; Ye, M.; Zhou, Z.; Sun, W.; Lin, X. The genus Cordyceps: A chemical and pharmacological review. J. Pharm. Pharmacol. 2013, 65, 474–493. [Google Scholar] [CrossRef]
- Li, S.P.; Yang, F.Q.; Tsim, K.W.K. Quality control of Cordyceps sinensis. J. Pharm. Biomed. Anal. 2006, 41, 1571–1584. [Google Scholar] [CrossRef]
- Olatunji, O.J.; Tang, J.; Tola, A.; Auberon, F.; Oluwaniyi, O.; Ouyang, Z. The genus Cordyceps: An extensive review. Fitoterapia 2018, 129, 293–316. [Google Scholar] [CrossRef]
- Das, S.K.; Masuda, M.; Sakurai, A.; Sakakibara, M. Medicinal uses of Cordyceps militaris. Fitoterapia 2010, 81, 961–968. [Google Scholar] [CrossRef]
- Panda, A.K.; Swain, K.C. Traditional uses and medicinal potential of Cordyceps sinensis. J. Ayurveda Integr. Med. 2011, 2, 9–13. [Google Scholar] [CrossRef]
- Jędrejko, K.J.; Lazur, J.; Muszyńska, B. Cordyceps militaris: Chemical constituents and biological activity. Foods 2021, 10, 2634. [Google Scholar] [CrossRef]
- Phull, A.R.; Ahmed, M.; Park, H.J. Cordyceps militaris as a biofunctional food source. Microorganisms 2022, 10, 405. [Google Scholar] [CrossRef]
- Abdullah, S.; Kumar, A. A brief review on the medicinal uses of Cordyceps militaris. Pharmacol. Res. Mod. Chin. Med. 2023, 7, 100228. [Google Scholar] [CrossRef]
- Hasnain, M.; Jamsheed, R.A.; Hussain, Z.; Latif, R. Comparative study on Cordyceps militaris and Ophiocordyceps sinensis. Int. J. Nat. Med. Health Sci. 2023, 2, 1–6. Available online: https://journals.iub.edu.pk/index.php/ijnms.
- Panossian, A.; Lemerond, T. Adaptogenic botanicals as nutraceuticals. Pharmaceuticals 2025, 18, 1346. [Google Scholar] [CrossRef] [PubMed]
- Brekhman, I.I.; Dardymov, I.V. New substances of plant origin. Annu. Rev. Pharmacol. 1969, 9, 419–430. [Google Scholar] [CrossRef]
- Panossian, A.; Lemerond, T.; Efferth, T. Adaptogens in brain fatigue. Pharmaceuticals 2025, 18, 261. [Google Scholar] [CrossRef]
- Shashidhar, M.G.; Giridhar, P.; Sankar, K.U.; Manohar, B. Bioactive principles from Cordyceps sinensis. J. Funct. Foods 2013, 5, 1013–1030. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Tulsawani, R. Efficacy of aqueous extract of Ophiocordyceps sinensis. Int. J. Med. Mushrooms 2022, 24, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Rajput, R.; Sharma, P.; Mishra, J.; Bhardwaj, A.; Sharma, R.K.; Singh, K.; Bansal, A.; Misra, K. Bioactive Fractions from the Chinese Caterpillar Mushroom, Ophiocordyceps sinensis (Ascomycetes), Elucidate Adaptogenic Role against Hypoxia Stress. Int. J. Med. Mushrooms 2020, 22, 1121–1133. [Google Scholar] [CrossRef]
- Di Lorenzo, R.; Falanga, D.; Ricci, L.; Colantuono, A.; Greco, G.; Angelillo, M.; Nugnes, F.; Di Serio, T.; Costa, D.; Tito, A.; Laneri, S. NAD-Driven Sirtuin Activation by Cordyceps sinensis Extract: Exploring the Adaptogenic Potential to Promote Skin Longevity. Int. J. Mol. Sci. 2024, 25, 4282. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, X.; Qu, K.; Zhu, P.; Guo, N.; Zhang, R.; Abliz, Z.; Yu, H.; Zhu, H. Binding of cordycepin monophosphate to AMP-activated protein kinase and its effect on AMP-activated protein kinase activation. Chem. Biol. Drug Des. 2010, 76, 340–344. [Google Scholar] [CrossRef]
- Wang, M.; Han, Z.; Fan, B.; Qu, K.; Zhang, W.; Li, W.; Li, J.; Li, L.; Li, J.; Li, H.; Wu, S.; Wang, D.; Zhu, H. Discovery of Oral AMP-Activated Protein Kinase Activators for Treating Hyperlipidemia. J. Med. Chem. 2024, 67, 7870–7890. [Google Scholar] [CrossRef]
- Wu, C.; Guo, Y.; Su, Y.; Zhang, X.; Luan, H.; Zhang, X.; Zhu, H.; He, H.; Wang, X.; Sun, G.; Sun, X.; Guo, P.; Zhu, P. Cordycepin activates AMP-activated protein kinase (AMPK) via interaction with the γ1 subunit. J. Cell. Mol. Med. 2014, 18, 293–304. [Google Scholar] [CrossRef]
- Hawley, S.A.; Ross, F.A.; Russell, F.M.; Atrih, A.; Lamont, D.J.; Hardie, D.G. Mechanism of Activation of AMPK by Cordycepin. Cell Chem. Biol. 2020, 27, 214–222. [Google Scholar] [CrossRef]
- Radhi, M.; Ashraf, S.; Lawrence, S.; Tranholm, A.A.; Wellham, P.A.D.; Hafeez, A.; Khamis, A.S.; Thomas, R.; McWilliams, D.; de Moor, C.H. A Systematic Review of the Biological Effects of Cordycepin. Molecules 2021, 26, 5886. [Google Scholar] [CrossRef]
- Chueaphromsri, P.; Kunhorm, P.; Phonchai, R.; Chaicharoenaudomrung, N.; Noisa, P. Cordycepin Enhances SIRT1 Expression and Maintains Stemness of Human Mesenchymal Stem Cells. In vivo 2023, 37, 596–610. [Google Scholar] [CrossRef]
- Hobbs, C.; Upton, R.; Gafner, S. Cordyceps botanical adulterants prevention bulletin. Botanical Adulterants Prevention Bulletin. Austin, TX: ABC-AHP-NCNPR Botanical Adulterants Prevention Program. Austin, TX, USA, 2024. Available online: https://www.herbalgram.org/resources/botanical-adulterants-prevention-program/adulterants-bulletins/new-cordyceps-bulletin-october-2024/.
- Sung, G.H.; Hywel-Jones, N.L.; Sung, J.M.; Luangsa-Ard, J.J.; Shrestha, B.; Spatafora, J.W. Phylogenetic classification of Cordyceps and the clavicipitaceous fungi. Stud. Mycol. 2007, 57, 5–59. [Google Scholar] [CrossRef] [PubMed]
- Islam, W.; Adnan, M.; Shabbir, A.; Naveed, H.; Abubakar, Y.S.; Qasim, M.; Tayyab, M.; Noman, A.; Nisar, M.S.; Khan, K.A.; Ali, H. Insect-fungal interactions: A detailed review of entomopathogenic fungi's pathogenicity to combat insect pests. Microb. Pathog. 2021, 159, 105122. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, S.A.; Elkhalifa, A.E.O.; Siddiqui, A.J.; Patel, M.; Awadelkareem, A.M.; Snoussi, M.; Ashraf, M.S.; Adnan, M.; Hadi, S. Cordycepin for Health and Wellbeing: A Potent Bioactive Metabolite of an Entomopathogenic Cordyceps Medicinal Fungus and Its Nutraceutical and Therapeutic Potential. Molecules 2020, 25, 2735. [Google Scholar] [CrossRef] [PubMed]
- Boddy, L. Chapter 9. Interactions with humans and other animals. In The Fungi, 3rd ed.; Elsevier: London, UK, 2016; pp. 293–336. Available online: https://www.sciencedirect.com/science/article/pii/B9780123820341000098.
- Lin, W.J.; Lee, Y.I.; Liu, S.L.; Lin, C.C.; Chung, T.Y.; Chou, J.Y. Evaluating the tradeoffs of a generalist parasitoid fungus, Ophiocordyceps unilateralis, on different sympatric ant hosts. Sci. Rep. 2020, 10, 6428. [Google Scholar] [CrossRef]
- Qiu, X.; Cao, L.; Han, R. Analysis of Volatile Components in Different Ophiocordyceps sinensis and Insect Host Products. Molecules 2020, 25, 1603. [Google Scholar] [CrossRef]
- Sen, S.; Karati, D.; Priyadarshini, R.; Dua, T.K.; Paul, P.; Sahu, S.; Nandi, G. Cordyceps sinensis (yarsagumba): Pharmacological properties of a mushroom. Pharmacol. Res. Mod. Chin. Med. 2023, 8, 100294. [Google Scholar] [CrossRef]
- Watanabe, T.; Rajbhandari, K.K.; Malla, K.J.; Yahara, S. Cordyceps synensis (Berk.) Sacc. In A Handbook of Medicinal Plants of Nepal; Kobfa Publishing Project: Bangkok, Thailand 2005; pp. 72–73. Available online: https://www.researchgate.net/publication/259973143_A_Hand_Book_of_Medicinal_Plants_of_Nepal#fullTextFileContent (accessed on 24 April 2025).
- Wei, P.; Wang, K.; Luo, C.; Huang, Y.; Misilimu, D.; Wen, H.; Jin, P.; Li, C.; Gong, Y.; Gao, Y. Cordycepin confers long-term neuroprotection via inhibiting neutrophil infiltration and neuroinflammation after traumatic brain injury. J. Neuroinflamm. 2021, 18, 137. [Google Scholar] [CrossRef]
- Lin, B.; Li, S. Cordyceps as an Herbal Drug. In Herbal Medicine: Biomolecular and Clinical Aspects, 2nd ed.; Benzie, I.F.F., Wachtel-Galor, S., Eds.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2011; Chapter 5; Available online: https://www.ncbi.nlm.nih.gov/books/NBK92758/.
- Cordyceps (Dongchongxiacao), Chinese Caterpillar Fungus, in: Pharmacopoeia of the People's Republic of China, English ed.; National Pharmacopoeia Committee: Beijing, China; p. p. 129. Available online: https://books.google.se/books/about/Pharmacopoeia_of_the_People_s_Republic_o.html?id=Z49IXwAACAAJ&redir_esc=y (accessed on 3 September 2025).
- Ma, Z.; Liu, Q.; Hong, Y.; Chen, J.; Tang, J.; Tang, Y.; Wang, L. Comparative analysis of bioactive ingredients of natural and cultivated Ophiocordyceps sinensis. Future Integr. Med. 2024, 3, 251–261. Available online: https://www.xiahepublishing.com/2835-6357/FIM-2024-00047/pdf.
- Wang, M.; Meng, X.Y.; Yang, R.L.; Qin, T.; Wang, X.Y.; Zhang, K.Y.; Fei, C.Z.; Li, Y.; Hu, Y.L.; Xue, F.Q. Cordyceps militaris polysaccharides enhance immunity. Carbohydr. Polym. 2012, 89, 461–466. [Google Scholar] [CrossRef]
- Jędrejko, K.; Kała, K.; Sułkowska-Ziaja, K.; Krakowska, A.; Zięba, P.; Marzec, K.; Szewczyk, A.; Sękara, A.; Pytko-Polończyk, J.; Muszyńska, B. Cordyceps militaris-Fruiting Bodies, Mycelium, and Supplements: Valuable Component of Daily Diet. Antioxidants 2022, 11, 1861. [Google Scholar] [CrossRef]
- Jędrejko, K.; Kała, K.; Sułkowska-Ziaja, K.; Krakowska, A.; Szewczyk, A.; Guśpiel, K.; Muszyńska, B. Analysis of bioactive compounds in Cordyceps militaris fruiting bodies and dietary supplements: In vitro bioaccessibility determination in artificial digestive juices. Int. J. Food Sci. Technol. 2024, 59, 4011–4019. [Google Scholar] [CrossRef]
- Sun, H.; Wei, S.; Gong, Y.; Ding, K.; Tang, S.; Sun, W.; Yuan, C.; Huang, L.; Liu, Z.; Chen, C.; Yao, L. Neuroprotective effects of cordycepin inhibit glutamate-induced apoptosis in hippocampal neurons. Cell Stress Chaperones 2024, 29, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Chai, X.; Pan, M.; Wang, J.; Feng, M.; Wang, Y.; Zhang, Q.; Sun, Y. Cordycepin exhibits anti-fatigue effect via activating TIGAR/SIRT1/PGC-1α signaling pathway. Biochem. Biophys. Res. Commun. 2022, 637, 127–135. [Google Scholar] [CrossRef]
- Chen, L.; Wei, W.; Sun, J.; Sun, B.; Deng, R. Cordycepin enhances anti-tumor immunity in breast cancer by enhanceing ALB expression. Heliyon 2024, 10, e29903. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; He, W.; Zhou, X.; Lv, Q.; Xu, X.; Yang, S.; Zhao, C.; Guo, L. Cordycepin protects against cerebral ischemia/reperfusion injury in vivo and in vitro. Eur. J. Pharmacol. 2011, 664, 20–28. [Google Scholar] [CrossRef]
- Govindula, A.; Pai, A.; Baghel, S.; Mudgal, J. Molecular mechanisms of cordycepin emphasizing its potential against neuroinflammation: An update. Eur. J. Pharmacol. 2021, 908, 174364. [Google Scholar] [CrossRef]
- Ma, X.; Zhao, Y.; Yang, T.; Gong, N.; Chen, X.; Liu, G.; Xiao, J. Integration of network pharmacology and molecular docking to explore the molecular mechanism of Cordycepin in the treatment of Alzheimer’s disease. Front. Aging Neurosci. 2022, 14, 1058780. Available online: https://www.frontiersin.org/journals/aging-neuroscience/articles/10.3389/fnagi.2022.1058780/full. [CrossRef]
- Ramesh, T.; Yoo, S.K.; Kim, S.W.; Hwang, S.Y.; Sohn, S.H.; Kim, I.W.; Kim, S.K. Cordycepin (3'-deoxyadenosine) attenuates age-related oxidative stress and ameliorates antioxidant capacity in rats. Exp. Gerontol. 2012, 47, 979–987. [Google Scholar] [CrossRef]
- Tuli, H.S.; Sandhu, S.S.; Sharma, A.K. Pharmacological and therapeutic potential of Cordyceps with special reference to Cordycepin. 3 Biotech 2014, 4, 1–12. [Google Scholar] [CrossRef]
- Cunningham, K.G.; Manson, W.; Spring, F.S.; Hutchinson, S.A. Cordycepin, a metabolic product isolated from cultures of Cordyceps militaris (Linn.) link. Nature 1950, 166, 949–54. [Google Scholar] [CrossRef]
- Radwan, M.A.; Willson, H.R. The Structure of Cordyeepin. Acta Crystallogr. B 1980, 36, 2185–2187. Available online: https://journals.iucr.org/paper?S056774088000831X. [CrossRef]
- Maľučká, L.U.; Uhrinová, A.; Lysinová, P. Medicinal mushrooms Ophiocordyceps sinensis and Cordyceps militaris. Liečivé huby Ophiocordyceps sinensis a Cordyceps militaris. Ceska Slov. Farm. 2022, 71, 259–265. [Google Scholar] [CrossRef]
- Lee, C.T.; Huang, K.S.; Shaw, J.F.; Chen, J.R.; Kuo, W.S.; Shen, G.; Grumezescu, A.M.; Holban, A.M.; Wang, Y.T.; Wang, J.S.; Hsiang, Y.P.; Lin, Y.M.; Hsu, H.H.; Yang, C.H. Trends in the Immunomodulatory Effects of Cordyceps militaris: Total Extracts, Polysaccharides and Cordycepin. Front. Pharmacol. 2020, 11, 575704. [Google Scholar] [CrossRef] [PubMed]
- Kała, K.; Cicha-Jeleń, M.; Hnatyk, K.; Krakowska, A.; Sułkowska-Ziaja, K.; Szewczyk, A.; Lazur, J.; Muszyńska, B. Coffee with Cordyceps militaris and Hericium erinaceus Fruiting Bodies as a Source of Essential Bioactive Substances. Pharmaceuticals 2024, 17, 955. [Google Scholar] [CrossRef]
- Xiaoying, M.; Peng, Z.; Hong, W.; Na, G.; Jun, X.; Ying, Z.; Xun, C.; Guoli, L. From functional foods to immunotherapeutic agents: mechanistic insights into medicinal mushroom bioactives in chronic inflammation management. Front. Nutr. 2025, 12, 1725297. [Google Scholar] [CrossRef]
- Chen, Y.; Guo, H.; Du, Z.; Liu, X.Z.; Che, Y.; Ye, X. Ecology-based screen identifies new metabolites from a Cordyceps-colonizing fungus as cancer cell proliferation inhibitors and apoptosis inducers. Cell Prolif. 2009, 42, 838–847. [Google Scholar] [CrossRef] [PubMed]
- Tshikhudo, P.P.; Mabhaudhi, T.; Koorbanally, N.A.; Mudau, F.N.; Avendaño Caceres, E.O.; Popa, D.; Calina, D.; Sharifi-Rad, J. Anticancer Potential of β-Carboline Alkaloids: An Updated Mechanistic Overview. Chem. Biodivers. 2024, 21, e202301263. [Google Scholar] [CrossRef]
- Yang, M.L.; Kuo, P.C.; Hwang, T.L.; Wu, T.S. Anti-inflammatory principles from Cordyceps sinensis. J. Nat. Prod. 2011, 74, 1996–2000. [Google Scholar] [CrossRef]
- Corradetti, R.; Lo Conte, G.; Moroni, F.; Passani, M.B.; Pepeu, G. Adenosine decreases aspartate and glutamate release from rat hippocampal slices. Eur. J. Pharmacol. 1984, 104, 19–26. [Google Scholar] [CrossRef]
- Schmidt, C.; Bellingham, M.C.; Richter, D.W. Adenosinergic modulation of respiratory neurones and hypoxic responses in the anaesthetized cat. J. Physiol. 1995, 483, 769–781. [Google Scholar] [CrossRef]
- Ballarin, M.; Herrera-Marschitz, M.; Casas, M.; Ungerstedt, U. Striatal adenosine levels measured 'in vivo' by microdialysis in rats with unilateral dopamine denervation. Neurosci. Lett. 1987, 83, 338–344. [Google Scholar] [CrossRef]
- Benowitz, L.I.; Goldberg, D.E.; Irwin, N. Inosine stimulates axon growth in vitro and in the adult CNS. Prog. Brain Res. 2002, 137, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Li, S.P.; Su, Z.R.; Dong, T.T.; Tsim, K.W. The fruiting body and its caterpillar host of Cordyceps sinensis show close resemblance in main constituents and anti-oxidation activity. Phytomedicine 2002, 9, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Kagota, S.; Nakamura, K.; Shinozuka, K.; Kunitomo, M. Antioxidant activity of the extracts from fruiting bodies of cultured Cordyceps sinensis. Phytother. Res. 2000, 14, 647–649. [Google Scholar] [CrossRef] [PubMed]
- Kiho, T.; Ookubo, K.; Usui, S.; Ukai, S.; Hirano, K. Structural features and hypoglycemic activity of a polysaccharide (CS-F10) from the cultured mycelium of Cordyceps sinensis. Biol. Pharm. Bull. 1999, 22, 966–970. [Google Scholar] [CrossRef]
- Li, S.P.; Zhao, K.J.; Ji, Z.N.; Song, Z.H.; Dong, T.T.; Lo, C.K.; Cheung, J.K.; Zhu, S.Q.; Tsim, K.W. A polysaccharide isolated from Cordyceps sinensis, a traditional Chinese medicine, protects PC12 cells against hydrogen peroxide-induced injury. Life Sci. 2003, 73, 2503–2513. [Google Scholar] [CrossRef]
- Yu, R.; Song, L.; Zhao, Y.; Bin, W.; Wang, L.; Zhang, H.; Wu, Y.; Ye, W.; Yao, X. Isolation and biological properties of polysaccharide CPS-1 from cultured Cordyceps militaris. Fitoterapia 2004, 75, 465–472. [Google Scholar] [CrossRef]
- Bok, J.W.; Lermer, L.; Chilton, J.; Klingeman, H.G.; Towers, G.H. Antitumor sterols from the mycelia of Cordyceps sinensis. Phytochemistry 1999, 51, 891–898. [Google Scholar] [CrossRef]
- Lindequist, U.; Lesnau, A.; Teuscher, E.; Pilgrim, H. Untersuchungen zur antiviralen Wirksamkeit von Ergosterol-peroxid [The antiviral action of ergosterol peroxide]. Pharmazie 1989, 44, 579–580. [Google Scholar] [PubMed]
- Lin, C.Y.; Ku, F.M.; Kuo, Y.C.; Chen, C.F.; Chen, W.P.; Chen, A.; Shiao, M.S. Inhibition of activated human mesangial cell proliferation by the natural product of Cordyceps sinensis (H1-A): an implication for treatment of IgA mesangial nephropathy. J. Lab. Clin. Med. 1999, 133, 55–63. [Google Scholar] [CrossRef]
- Arts, I.C.; Coolen, E.J.; Bours, M.J.; Huyghebaert, N.; Stuart, M.A.; Bast, A.; Dagnelie, P.C. Adenosine 5'-triphosphate (ATP) supplements are not orally bioavailable: a randomized, placebo-controlled cross-over trial in healthy humans. J. Int. Soc. Sports Nutr. 2012, 9, 16. [Google Scholar] [CrossRef]
- Coolen, E.J.; Arts, I.C.; Bekers, O.; Vervaet, C.; Bast, A.; Dagnelie, P.C. Oral bioavailability of ATP after prolonged administration. Br. J. Nutr. 2011, 105, 357–366. [Google Scholar] [CrossRef]
- Chatnarin, S.; Thirabunyanon, M. Potential bioactivities via anticancer, antioxidant, and immunomodulatory properties of cultured mycelial enriched β-D-glucan polysaccharides from a novel fungus Ophiocordyceps sinensis OS8. Front. Immunol. 2023, 14, 1150287. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Li, Q.; Chen, Y.; Wang, X.; Zhou, X. Determination and analysis of cordycepin and adenosine in the products of Cordyceps spp. Afr. J. Microbiol. Res. 2009, 3, 953–961. Available online: https://www.researchgate.net/publication/228479929_Determination_and_analysis_of_cordycepin_and_adenosine_in_the_products_of_Cordyceps_spp.
- Zhou, Y.; Wang, M.; Zhang, H.; Huang, Z.; Ma, J. Comparative study of the composition of cultivated, naturally grown Cordyceps sinensis, and stiff worms across different sampling years. PLoS ONE 2019, 14, e0225750. [Google Scholar] [CrossRef]
- Li, Q.; Liao, H.; Zhang, C. Comparative HPLC Determination of Cordycepin in Cordyceps militaris from Different Locations. Food Sci. 2014. Available online: https://www.spkx.net.cn/EN/Y2014/V35/I14/118?utm_source=chatgpt.com. [CrossRef]
- Su, Y.; Li, P.; Zhang, H.; Lin, M.; Liu, W.; Xu, R.; Hu, H.; Liu, Y.M. Simultaneous Quantification of Adenosine and Deoxyadenosine Isomers in Foods with High Sensitivity. Anal. Methods 2019, 11, 4946–4950. [Google Scholar] [CrossRef]
- Gu, C.; Zhang, D.; Zhai, W.; Zhang, H.; Wang, S.; Lv, S.; Bao, Y.; Zhu, D.; Feng, S.; Guo, S.; Wang, Z. Research progress on Cordyceps militaris polysaccharides. Food Biosci. 2022, 45, 101503. [Google Scholar] [CrossRef]
- Chen, L.H.; Wu, Y.; Guan, Y.M.; Jin, C.; Zhu, W.F.; Yang, M. Analysis of the High-Performance Liquid Chromatography Fingerprints and Quantitative Analysis of Multicomponents by Single Marker of Products of Fermented Cordyceps sinensis. J. Anal. Methods Chem. 2018, 2018, 5943914. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, W.; Wu, X.; Fu, W.; Yang, C.; Long, N. Insights into the Mycosphere Fungal Community and Its Association with Nucleoside Accumulation in Ophiocordyceps sinensis. J. Fungi 2025, 11, 696. [Google Scholar] [CrossRef]
- Yuan, J.P.; Wang, J.H.; Liu, X.; Kuang, H.C.; Zhao, S.Y. Simultaneous determination of free ergosterol and ergosteryl esters in Cordyceps sinensis by HPLC. Food Chem. 2007, 105, 1755–1759. [Google Scholar] [CrossRef]
- Hu, D.; Yang, X.; Hu, C.; Feng, Z.; Chen, W.; Shi, H. Comparison of Ergosterol and Vitamin D2 in Mushrooms Agaricus bisporus and Cordyceps militaris Using Ultraviolet Irradiation Directly on Dry Powder or in Ethanol Suspension. ACS Omega 2021, 6, 29506–2951. [Google Scholar] [CrossRef]
- Zhao, J.; Shi, T.; Zhu, W.; Chen, L.; Guan, Y.; Jin, C. Quality control method of sterols in fermented Cordyceps sinensis based on combined fingerprint and quantitative analysis of multicomponents by single marker. J. Food Sci. 2020, 85, 2994–3002. [Google Scholar] [CrossRef] [PubMed]
- Lo, H.C.; Hsieh, C.; Lin, F.Y.; Hsu, T.H. A Systematic Review of the Mysterious Caterpillar Fungus Ophiocordyceps sinensis in Dong-ChongXiaCao ( Dōng Chóng Xià Cǎo) and Related Bioactive Ingredients. J. Tradit. Complement. Med. 2013, 3, 16–32, https://pmc.ncbi.nlm.nih.gov/articles/PMC3924981/. [Google Scholar] [CrossRef] [PubMed]
- Suhadolnik, R.J. Naturally occurring nucleoside and nucleotide antibiotics. Prog. Nucleic Acid Res. Mol. Biol. 1979, 22, 193–291. [Google Scholar] [CrossRef]
- Hueng, D.Y.; Hsieh, C.H.; Cheng, Y.C.; Tsai, W.C.; Chen, Y. Cordycepin inhibits migration of human glioblastoma cells by affecting lysosomal degradation and protein phosphatase activation. J. Nutr. Biochem. 2017, 41, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wu, X.; Liang, Y.N.; Wang, L.; Song, Z.X.; Liu, J.L.; Tang, Z.S. Cordycepin Induces Apoptosis and Inhibits Proliferation of Human Lung Cancer Cell Line H1975 via Inhibiting the Phosphorylation of EGFR. Molecules 2016, 21, 1267. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, Z.; Zhao, Z.; Zhang, N. Structural and pharmacological insights into cordycepin for neoplasms and metabolic disorders. Front. Pharmacol. 2024, 15, 1367820. [Google Scholar] [CrossRef]
- Wang, Y.; Deng, Y.; Feng, M.; Chen, J.; Zhong, M.; Han, Z.; Zhang, Q.; Sun, Y. Cordycepin extracted from Cordyceps militaris mitigated CUMS-induced depression of rats via targeting GSK3β/β-catenin signaling pathway. J. Ethnopharmacol. 2025, 340, 119249. [Google Scholar] [CrossRef]
- Kopalli, S.R.; Cha, K.M.; Lee, S.H.; Hwang, S.Y.; Lee, Y.J.; Koppula, S.; Kim, S.K. Cordycepin, an Active Constituent of Nutrient Powerhouse and Potential Medicinal Mushroom Cordyceps militaris Linn., Ameliorates Age-Related Testicular Dysfunction in Rats. Nutrients 2019, 11, 906. [Google Scholar] [CrossRef]
- Takahashi, S.; Tamai, M.; Nakajima, S.; Kato, H.; Johno, H.; Nakamura, T.; Kitamura, M. Blockade of adipocyte differentiation by cordycepin. Br. J. Pharmacol. 2012, 167, 561–575. [Google Scholar] [CrossRef] [PubMed]
- Jiaonang, Bailing. Bailing capsule, in: Pharmacopoeia of the People's Republic of China, English ed.; National Pharmacopoeia Committee: Beijing, China, 2010; pp. 540–541. Available online: https://books.google.se/books/about/Pharmacopoeia_of_the_People_s_Republic_o.html?id=Z49IXwAACAAJ&redir_esc=y (accessed on 3 September 2025).
- Latini, S.; Pedata, F. Adenosine in the central nervous system: release mechanisms and extracellular concentrations. J. Neurochem. 2001, 79, 463–484. [Google Scholar] [CrossRef] [PubMed]
- Porkka-Heiskanen, T.; Kalinchuk, A.V. Adenosine, energy metabolism, and sleep homeostasis. Sleep Med. Rev. 2011, 15, 123–135. [Google Scholar] [CrossRef]
- Eltzschig, H.K.; Thompson, L.F.; Karhausen, J.; Cotta, R.J.; Ibla, J.C.; Robson, S.C.; Colgan, S.P. Endogenous adenosine produced during hypoxia attenuates neutrophil accumulation: coordination by extracellular nucleotide metabolism. Blood 2004, 104, 3986–3992. [Google Scholar] [CrossRef]
- Eltzschig, H.K.; Carmeliet, P. Hypoxia and inflammation. N. Engl. J. Med. 2011, 364, 656–665. [Google Scholar] [CrossRef]
- Rau, A.R.; Ariwodola, O.J.; Weiner, J.L. Presynaptic adenosine A₁ receptors modulate excitatory transmission in the rat basolateral amygdala. Neuropharmacology 2014, 77, 465–474. [Google Scholar] [CrossRef]
- Brambilla, D.; Chapman, D.; Greene, R. Adenosine mediation of presynaptic feedback inhibition of glutamate release. Neuron 2005, 46, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Dobson, G.P.; Arsyad, A.; Letson, H.L. The Adenosine Hypothesis Revisited: Modulation of Coupling between Myocardial Perfusion and Arterial Compliance. Front. Physiol. 2017, 8, 824. [Google Scholar] [CrossRef]
- Görlach, A. Control of adenosine transport by hypoxia. Circ. Res. 2005, 97, 1–3. [Google Scholar] [CrossRef]
- Tune, J.D.; Gorman, M.W.; Feigl, E.O. Matching coronary blood flow to myocardial oxygen consumption. J. Appl. Physiol. 2004, 97, 404–415. [Google Scholar] [CrossRef] [PubMed]
- Hori, M.; Kitakaze, M. Adenosine, the heart, and coronary circulation. Hypertension 1991, 18, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Casey, D.P.; Madery, B.D.; Pike, T.L.; Eisenach, J.H.; Dietz, N.M.; Joyner, M.J.; Wilkins, B.W. Adenosine receptor antagonist and augmented vasodilation during hypoxic exercise. J. Appl. Physiol. 2009, 107, 1128–1137. [Google Scholar] [CrossRef]
- Dengler, F. Activation of AMPK under Hypoxia: Many Roads Leading to Rome. Int. J. Mol. Sci. 2020, 21, 2428. [Google Scholar] [CrossRef]
- Bowser, J.L.; Phan, L.H.; Eltzschig, H.K. The Hypoxia-Adenosine Link during Intestinal Inflammation. J. Immunol. 2018, 200, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Decking, U.K.; Schlieper, G.; Kroll, K.; Schrader, J. Hypoxia-induced inhibition of adenosine kinase potentiates cardiac adenosine release. Circ. Res. 1997, 81, 154–164. [Google Scholar] [CrossRef]
- Hardie, D.G. AMP-activated protein kinase: an energy sensor that regulates all aspects of cell function. Genes Dev. 2011, 25, 1895–1908. [Google Scholar] [CrossRef]
- Jeon, S.M. Regulation and function of AMPK in physiology and diseases. Exp. Mol. Med. 2016, 48, e245. [Google Scholar] [CrossRef]
- Melani, A.; Pugliese, A.M.; Pedata, F. Chapter Thirteen - Adenosine Receptors in Cerebral Ischemia. In Int. Rev. Neurobiol.; Mori, Akihisa, Ed.; Academic Press: London, UK, 2014; Vol. 119, pp. 309–348. [Google Scholar] [CrossRef]
- Jacobson, K.A.; Gao, Z.G. Adenosine receptors as therapeutic targets. Nat. Rev. Drug Discov. 2006, 5, 247–264. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, Z.; Zhao, Z.; Zhang, N. Structural and pharmacological insights into cordycepin for neoplasms and metabolic disorders. Front. Pharmacol. 2024, 15, 1367820. [Google Scholar] [CrossRef]
- Willson, C. The clinical toxicology of caffeine: A review and case study. Toxicol. Rep. 2018, 5, 1140–1152. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Dueñas, V.; Gómez-Soler, M.; López-Cano, M.; Taura, J.J.; Ledent, C.; Watanabe, M.; Jacobson, K.A.; Vilardaga, J.P.; Ciruela, F. Uncovering caffeine's adenosine A2A receptor inverse agonism in experimental Parkinsonism. ACS Chem. Biol. 2014, 9, 2496–2501. [Google Scholar] [CrossRef]
- Ishibashi, K.; Miura, Y.; Wagatsuma, K.; Toyohara, J.; Ishiwata, K.; Ishii, K. Adenosine A2A Receptor Occupancy by Caffeine After Coffee Intake in Parkinson's Disease. Mov. Disord. 2022, 37, 853–857. [Google Scholar] [CrossRef]
- Qi, S.; Guan, H.; Wang, Y.; Fang, Q.; Cheng, X.; Liu, P.; Wei, H.; Liu, W.; Wang, C. Simultaneous determination of cordycepin and its metabolite 3'-deoxyinosine in rat whole blood by ultra-high-performance liquid chromatography coupled with Q Exactive hybrid quadrupole orbitrap high-resolution accurate mass spectrometry and its application to accurate pharmacokinetic studies. J. Sep. Sci. 2023, 46, e2200602. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Liu, F.; Guo, L.; Fan, W.; Wang, J.; Lu, B.; Hong, G.; Zhang, W.; Tian, S.; Mao, J.; Xie, J. Distribution and Pharmacokinetic Characteristics of Cordycepin in Rat: Investigated by UHPLC-HRMS/MS and Blood-Brain Synchronous Microdialysis. Biomed. Chromatogr. 2025, 39, e70038. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Shinozuka, K.; Yoshikawa, N. Anticancer and antimetastatic effects of cordycepin, an active component of Cordyceps sinensis. J. Pharmacol. Sci. 2015, 127, 53–56. [Google Scholar] [CrossRef]
- Chang, W.K.; Chen, Y.T.; Lin, C.P.; Wang, C.J.; Shieh, H.R.; Chi, C.W.; Tsai, T.H.; Chen, Y.J. Cordycepin Augments the Efficacy of Anti-PD1 against Colon Cancer. Biomedicines 2024, 12, 1568. [Google Scholar] [CrossRef]
- Lee, J.B.; Radhi, M.; Cipolla, E.; Gandhi, R.D.; Sarmad, S.; Zgair, A.; Kim, T.H.; Feng, W.; Qin, C.; Adrower, C.; Ortori, C.A.; Barrett, D.A.; Kagan, L.; Fischer, P.M.; de Moor, C.H.; Gershkovich, P. A novel nucleoside rescue metabolic pathway may be responsible for therapeutic effect of orally administered cordycepin. Sci. Rep. 2019, 9, 15760. [Google Scholar] [CrossRef]
- Tsai, Y.J.; Lin, L.C.; Tsai, T.H. Pharmacokinetics of adenosine and cordycepin, a bioactive constituent of Cordyceps sinensis in rats. J. Agric. Food Chem. 2010, 58, 4638–4643. [Google Scholar] [CrossRef]
- Kaptchuk, T.J. The Web That Has No Weaver; Congdon and Weed: New York, NY, USA, 1983; pp. 55–65. Available online: https://books.google.se/books/about/The_Web_That_Has_No_Weaver.html?id=6BiGl562OgEC&redir_esc=y.
- Panossian, A.G.; Efferth, T.; Shikov, A.N.; Pozharitskaya, O.N.; Kuchta, K.; Mukherjee, P.K.; Banerjee, S.; Heinrich, M.; Wu, W.; Guo, D.A.; Wagner, H. Evolution of the adaptogenic concept from traditional use to medical systems: Pharmacology of stress- and aging-related diseases. Med. Res. Rev. 2021, 41, 630–703. [Google Scholar] [CrossRef]
- Jiang, S.J. A tonic application of Cordyceps sinensis in medicinal diets. J. Chin. Med. Mater. 1994, 17, 47–48. [Google Scholar]
- Liu, M.; Cui, C.; Chang, T.; Zhou, Q.; Cui, Y.; Zhang, S.; Liao, X. Effects and safety of Ophiocordyceps sinensis preparation in the adjuvant treatment for dialysis patients: a systematic review and meta-analysis. Front. Pharmacol. 2024, 15, 1360997. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Xu, C.; Si, X.; He, F.; Xu, K.; Zhang, Y.; Lin, S. Efficacy of traditional Chinese medicine Cordyceps sinensis as an adjunctive treatment in patients with renal dysfunction: a systematic-review and meta-analysis. Front. Med. 2025, 11, 1477569. [Google Scholar] [CrossRef] [PubMed]
- Pu, F.; Li, T.; Shen, C.; Wang, Y.; Tang, C.; Zhang, X.; Yan, L.; Xu, Q.; Liu, J. Fermented Ophiocordyceps sinensis mycelium products for preventing contrast-associated acute kidney injury: a systematic review of randomized controlled trials. Ren. Fail. 2024, 46, 2300302. [Google Scholar] [CrossRef]
- Tao, Y.; Luo, R.; Xiang, Y.; Lei, M.; Peng, X.; Hu, Y. Use of bailing capsules (cordyceps sinensis) in the treatment of chronic kidney disease: a meta-analysis and network pharmacology. Front. Pharmacol. 2024, 15, 1342831. [Google Scholar] [CrossRef] [PubMed]
- Xue, X.; Jin, X.Y.; Ye, X.L.; Li, K.Y.; Li, J.X.; Liu, X.H.; Bai, J.; Liu, Q.; Zhang, B.R.; Zou, X.R.; Yuan, J.; Lu, C.L.; Zhao, F.F.; Liu, J.P.; Wang, X.Q. Ophiocordyceps sinensis preparations combined with the renin-angiotensin system inhibitor for diabetic kidney disease treatment: an umbrella review of systematic reviews and network meta-analysis. Front. Pharmacol. 2024, 15, 1360633. [Google Scholar] [CrossRef]
- Wang, C.; Wang, J.; Qi, Y. Adjuvant treatment with Cordyceps sinensis for lung cancer: A systematic review and meta-analysis of randomized controlled trials. J. Ethnopharmacol. 2024, 327, 118044. [Google Scholar] [CrossRef]
- Yu, X.; Mao, Y.; Shergis, J.L.; Coyle, M.E.; Wu, L.; Chen, Y.; Zhang, A.L.; Lin, L.; Xue, C.C.; Xu, Y. Effectiveness and Safety of Oral Cordyceps sinensis on Stable COPD of GOLD Stages 2-3: Systematic Review and Meta-Analysis. Evid Based Complement Alternat Med. 2019, 2019, 4903671. [Google Scholar] [CrossRef]
- Ma, G.; Jin, Y. Therapeutic efficacy and pharmacological mechanism of Bailing capsule on chronic obstructive pulmonary disease: a meta-analysis and network pharmacology. Pharm. Biol. 2024, 62, 803–817. [Google Scholar] [CrossRef]
- Welch, J.; Kole, R.; Daniels, S. The effectiveness and efficacy of Ophiocordyceps sinensis supplementation: A systematic review of randomized clinical trials on healthy humans. Int. J. Herb. Med. 2023, 11, 15–22. [Google Scholar] [CrossRef]
- Dewi, L.; Khemtong, C. Ergogenic Aid by Cordyceps: Does It Work?? Curr. Nutr. Rep. 2025, 14, 97. [Google Scholar] [CrossRef]
- European Medicines Agency. Guideline on the assessment of clinical safety and efficacy in the preparation of EU herbal monographs for well-established and traditional herbal medicinal products EMA/HMPC/104613/2005 – Rev. 1; EMA: London, UK, 2017; Available online: https://www.ema.europa.eu/en/documents/scientific-guideline/guideline-assessment-clinical-safety-and-efficacy-preparation-eu-herbal-monographs-well-established-and-traditional-herbal-medicinal-products_en.pdf.
- Brennan, S.E.; Johnston, R.V. Research Note: Interpreting findings of a systematic review using GRADE methods. J. Physiother. 2023, 69, 198–202. [Google Scholar] [CrossRef]
- Cao, Z.; Wen, Y. Therapeutic effect analysis of JinShuiBao capsule in treatment of 33 elderly senescent XuZheng patients. J. Appl. Tradit. Chin. Med. 1993, 1, 32–33. [Google Scholar]
- Zhang, Z.; Huang, W.; Liao, S.; Li, J.; Lei, L.; Lui, J.; Leng, F.; Gong, W.; Zhang, H.; Wan, L.; Wu, R.; Li, S.; Luo, H.; Zhu, F. Clinical and laboratory studies of JinShuiBao in scavenging oxygen free radicals in elderly senescent XuZheng patients. J. Admin. Tradit. Chin. Med. 1995, 5, 14–18. [Google Scholar]
- Chen, S.; Li, Z.; Krochmal, R.; Abrazado, M.; Kim, W.; Cooper, C.B. Effect of Cs-4® (Cordyceps sinensis) on exercise performance in healthy older subjects: A double-blind, placebo-controlled trial. J. Altern. Complement. Med. 2010, 16, 585–590. [Google Scholar] [CrossRef] [PubMed]
- Nagata, A.; Tajima, T.; Moriyasu, S. Cordyceps drinks and running performance. J. Exerc. Sports Physiol. 2002, 9, 85–92. [Google Scholar]
- Nagata, A.; Tajima, T.; Uchida, M. Supplemental anti-fatigue effects of Cordyceps sinensis (Tochu-Kaso) extract powder during three stepwise exercise of human. Jpn. J. Phys. Fitness Sports Med. 2006, 55 Suppl., S145–S152. Available online: https://www.jstage.jst.go.jp/article/jspfsm/55/Supplement/55_S145/_article/-char/ja/. [CrossRef]
- Hirsch, K.R.; Smith-Ryan, A.E.; Roelofs, E.J.; Trexler, E.T.; Mock, M.G. Cordyceps militaris Improves Tolerance to High-Intensity Exercise After Acute and Chronic Supplementation. J Diet Suppl. 2017, 14, 42–53. [Google Scholar] [CrossRef]
- Nakamura, A.; Shinozaki, E.; Suzuki, Y.; Santa, K.; Kumazawa, Y.; Kobayashi, F.; Nagaoka, I.; Koikawa, N. Effect of the Administration of Cordyceps militaris Mycelium Extract on Blood Markers for Anemia in Long-Distance Runners. Nutrients 2024, 16, 1835. [Google Scholar] [CrossRef]
- Hsu, C.C.; Lin, Y.A.; Su, B.; Li, J.H.; Huang, H.Y.; Hsu, M.C.P. No effect of Cordyceps sinensis supplementation on testosterone level and muscle strength in healthy young adults for resistance training. Biol Sport. 2011, 28, 107–110. Available online: https://www.webofscience.com/wos/woscc/full-record/WOS:000293048800005. [CrossRef]
- Parcell, A.C.; Smith, J.M.; Schulthies, S.S.; Myrer, J.W.; Fellingham, G. Cordyceps Sinensis (CordyMax Cs-4) supplementation does not improve endurance exercise performance. Int. J. Sport Nutr. Exerc. Metab. 2004, 14, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Nicodemus, J.K.; Hagan, R.D.; Zue, J.S.; Baker, C. Supplementation with cordyceps cs-4 fermentation product promotes fat metabolism during prolonged exercise. Med. Sci. Sports Exerc. 2001, 33, S164. [Google Scholar] [CrossRef]
- Xiao, Y.; Huang, X.; Zhu, J. Randomized double-blind placebo-controlled clinical trial and assessment of fermentation product of Cordyceps sinensis (Cs-4) in enhancing aerobic capacity and respiratory function of the healthy elderly volunteers. Chin. J. Integr. Med. 2004, 10, 187–192. [Google Scholar]
- Savioli, F.P.; Zogaib, P.; Franco, E.; Alves De Salles, F.C.; Giorelli, G.V.; et al. Effects of Cordyceps sinensis supplementation during 12 weeks in amateur marathoners: A randomized, double-blind placebo-controlled trial. J. Herb. Med. 2022, 34, 100570. [Google Scholar] [CrossRef]
- Zhou, X.; Gong, Z.; Su, Y.; Lin, J.; Tang, K. Cordyceps fungi: natural products, pharmacological functions and developmental products. J. Pharm. Pharmacol. 2009, 61, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.; Xi-zhen, H.; Jia-shi, Z. Randomized double-blind placebo-controlled clinical trial and assessment of fermentation product of Cordyceps sinensis (Cs-4) in enhancing aerobic capacity and respiratory function of the healthy elderly volunteers. Chin. J. Integr. Med. 2004, 10, 187–192. Available online: https://link.springer.com/article/10.1007/BF02836405. [CrossRef]
- Dudgeon, W.D.; Thomas, D.D.; Dauch, W.; Scheett, T.P.; Webster, M.J. The effects of high and low-dose Cordyceps Militaris-containing mushroom blend supplementation after seven and twenty-eight days. Am. J. Sports Sci. 2018, 6, 1–7. Available online: https://sciencepublishinggroup.com/article/10.11648/j.ajss.20180601.11. [CrossRef]
- Ontawong, A.; Pengnet, S.; Thim-Uam, A.; Munkong, N.; Narkprasom, N.; Narkprasom, K.; Kuntakhut, K.; Kamkeaw, N.; Amornlerdpison, D. A randomized controlled clinical trial examining the effects of Cordyceps militaris beverage on the immune response in healthy adults. Sci. Rep. 2024, 14, 7994. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, G.; Zhang, C.; Tang, G.; Chan, Y.; Wang, N.; Feng, Y. Randomized, waitlist-controlled trial of Cordyceps sinensis mycelium culture extract (Cs4) for long COVID patients in Hong Kong. Acta Mater. Med. 2025, 4, 250–261. Available online: https://www.scienceopen.com/hosted-document?doi=10.15212/AMM-2024-0089. [CrossRef]
- ANM Health. A randomized, double-blind, placebo-controlled study to evaluate the efficacy and safety of Cordyceps capsules as an add-on therapy in patients with mild to moderate COVID-19 infection. Available online: https://anm.health/wp-content/uploads/2023/08/Annexure-i-Clinical-Trials-Research-Summry.pdf.
- Wang, N.; Li, J.; Huang, X.; Chen, W.; Chen, Y. Herbal Medicine Cordyceps sinensis Improves Health-Related Quality of Life in Moderate-to-Severe Asthma. Evid.-Based Complement. Altern. Med. 2016, 2016, 6134593. [Google Scholar] [CrossRef]
- Heo, J.Y.; et al. Jin Young Heo, Hyun Wook Baik, Hyuk Jung Kim, Jae Min Lee, Hyung Woo Kim, Yong Sun Choi, Jung Ho Won, Hyun Mi Kim, Won Il Park, Chul Young Kim. Efficacy and Safety of Cordyceps militaris in Korean adults with mild liver dysfunction. J. Clin. Nutr. 2015, 7, 81–86. [CrossRef]
- Zhou, J.; Chen, X.; Xiao, L.; Zhou, J.; Feng, L.; Wang, G. Efficacy and Safety of Cordyceps militaris as an Adjuvant to Duloxetine in the Treatment of Insomnia in Patients With Depression: A 6-Week Double- Blind, Randomized, Placebo-Controlled Trial. Front. Psychiatry 2021, 12, 754921. [Google Scholar] [CrossRef]
- Ru, J.; Li, P.; Wang, J.; Zhou, W.; Li, B.; Huang, C.; Li, P.; Guo, Z.; Tao, W.; Yang, Y.; Xu, X.; Li, Y.; Wang, Y.; Yang, L. TCMSP: a database of systems pharmacology for drug discovery from herbal medicines. J. Cheminform. 2014, 6, 13. [Google Scholar] [CrossRef]
- Cui, G.; Li, M.; Guo, W.; Gao, M.; Zhu, Q.; Liao, J. AI-driven network pharmacology: Multi-scale mechanisms of traditional Chinese medicine from molecular to patient analysis. Comput. Struct. Biotechnol. J. 2025, 27, 5087–5104. [Google Scholar] [CrossRef]
- Barth, S.W.; Efferth, T. Network pharmacology to elucidate the role of phytotherapy in neurocognitive disorders. Phytomedicine 2025, 148, 157031. [Google Scholar] [CrossRef] [PubMed]
- Panossian, A.; Efferth, T. Network Pharmacology of Adaptogens in the Assessment of Their Pleiotropic Therapeutic Activity. Pharmaceuticals 2022, 15, 1051. [Google Scholar] [CrossRef]
- Panossian, A. Trends and Pitfalls in the Progress of Network Pharmacology Research on Natural Products. Pharmaceuticals 2025, 18, 538. [Google Scholar] [CrossRef]
- Panossian, A.; Abdelfatah, S.; Efferth, T. Network Pharmacology of Red Ginseng (Part I): Effects of Ginsenoside Rg5 at Physiological and Sub-Physiological Concentrations. Pharmaceuticals 2021, 14, 999. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Liu, Y.; Gao, Y.; Suonanlamao Ma, Y.; Xiao, Y.; Wei, L. Integrating proteomics, metabolomics, and network pharmacology to investigate the mechanism of Cordyceps sinensis in the treatment of COPD rats. J. Pharm. Biomed. Anal. 2025, 266, 117076. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Liu, Z.; Hu, J.; Sun, S.; Xie, X.; Kong, X.; Tang, Y. Pharmacodynamic evaluation of Cordyceps sinensis (Berk.) Sacc. for ischemic stroke in rats and potential mechanism through network pharmacology and molecular docking. J. Tradit. Chin. Med. Sci. 2023, 10, 196–207. [Google Scholar] [CrossRef]
- Butterweck, V.; Nahrstedt, A. What is the best strategy for preclinical testing of botanicals? A critical perspective. Planta medica 2012, 78, 747–754. [Google Scholar] [CrossRef]
- Wang, J.; Liu, R.; Liu, B.; Yang, Y.; Xie, J.; Zhu, N. Systems Pharmacology-based strategy to screen new adjuvant for hepatitis B vaccine from Traditional Chinese Medicine Ophiocordyceps sinensis. Sci. Rep. 2017, 7, 44788. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, M.; Liang, X.; Wang, J.; Zhang, M.; Li, C.; Wei, L.; Bi, H. Bioinformatics analysis and in vivo validation study of Ophiocordyceps sinensis (Berk.)G.H.Sungetal against lung adenocarcinoma. J. Ethnopharmacol. 2023, 317, 116739. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, L.; Lu, Y.; Zhang, J.; Yang, M.; Tian, Y.; Dong, J.; Liao, L. Protective effect of Cordyceps sinensis against diabetic kidney disease through promoting proliferation and inhibiting apoptosis of renal proximal tubular cells. BMC Complement. Med. Ther. 2023, 23, 109. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Pang, W.; Wang, Y.; Zhang, Y. Cordyceps sinensis extract protects against acute kidney injury by inhibiting perforin expression in NK cells via the STING/IRF3 pathway. Aging 2024, 16, 5887–5904. [Google Scholar] [CrossRef]
- He, H.; Tang, J.; Ru, D.; Shu, X.; Li, W.; Li, J.; Ma, L.; Hu, X.; Xiong, L.; Li, L. Protective effects of Cordyceps extract against UVB-induced damage and prediction of application prospects in the topical administration: An experimental validation and network pharmacology study. Biomed. Pharmacother. 2020, 121, 109600. [Google Scholar] [CrossRef]
- Pei, H.; He, Z.; Chen, W.; Zhao, Y.; Li, J.; Wang, R.; Zong, Y.; Du. RNetwork pharmacology molecular docking analysis on the mechanism of Cordyceps militaris polysaccharide regulating immunity through TLR4/TNF-α pathways. J. Biochem. Mol. Toxicol. 2023, 37, e23345. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.L.; Park, J.; Kim, H.I.; Boo, M.; Sim, J.H.; Kim, H.; Choi, H.; An, S.; Yang, H.; Shin, S.; Han, Y.; Kim, H.; Park, J.; Lee, K. Integrating Network Pharmacology and Optimization of Preparation Methods to Enhance the Anticancer Effect of Cordyceps militaris on Lung Cancer. Anticancer Res. 2025, 45, 2963–2984. [Google Scholar] [CrossRef]
- Wang, P.; Zong, Y.; Huang, X.; Zhang, W.; Di, L. Study on the potential mechanism of Cordyceps militaris in chronic obstructive pulmonary disease via integrating metabolomics and network pharmacology. J. Pharm. Pharmacol. Advance online publication. 2025, 78, rgaf114. [Google Scholar] [CrossRef]
- Zhou, H.B.; Feng, L.J.; Liang, X.D.K.; Zhou, Z.X.; Li, M.; Zhang, J.L.; Su, G.H. Mechanism of Cordyceps militaris in gouty nephropathy explored using network pharmacology and molecular docking technology. Food Biosci. 2024, 60, 104299. [Google Scholar] [CrossRef]
- Gandhale, P.; Chikhale, R.; Khanal, P.; Biswa, V.; Ali, R.; Khan, M.S.; Gurav, N.; Ayyanar, M.; Das, S.; Gurav, S. Quest for Anti-SARS-CoV-2 antiviral therapeutics: in-silico and in-vitro analysis of edible mushroom- Cordyceps militaris. J. Ayurveda Integr. Med. 2024, 15, 100979. [Google Scholar] [CrossRef]
- Lee, D.; Lee, W.-Y.; Jung, K.; Kwon, Y.S.; Kim, D.; Hwang, G.S.; Kim, C.-E.; Lee, S.; Kang, K.S. The Inhibitory Effect of Cordycepin on the Proliferation of MCF-7 Breast Cancer Cells, and Its Mechanism: An Investigation Using Network Pharmacology-Based Analysis. Biomolecules 2019, 9, 414. [Google Scholar] [CrossRef]
- Qiu, X.; Li, C.; Wang, J.; Yu, H.; Wang, R.; Wang, L.; Zhang, S.; Tang, Y.; Li, Z.; Li, Q. Cordycepin Inhibits Proliferation, Migration, and Promotes Apoptosis in Fibrosarcoma HT1080 Cells by Targeting Akt1 and Kinase Activity Through Network Pharmacology Analysis. Cell Biochem. Funct. 2025, 43, e70103. [Google Scholar] [CrossRef]
- Zhong, M.; Feng, M.; Chen, X.; Zhang, Y.; Wang, Y.; Yuan, P.; Zhang, T.; Han, Z.; Tang, C.; Zhang, Q.; Sun, Y. Cordycepin attenuated cyclophosphamide (CTX)-induced immunosuppression in mice via EGFR/Nrf2 antioxidant signaling pathway. Int. Immunopharmacol. 2025, 163, 115235. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhuang, Y.-D.; Zhang, Q.; Liu, S.; Zhuang, B.-B.; Wang, C.-H.; Liang, R.-S. Exploring the mechanism of cordycepin combined with doxorubicinin treating glioblastoma based on network pharmacology and biological verification. Peer J 2022, 10, e12942. Available online: https://peerj.com/articles/12942.pdf?utm_source=chatgpt.com. [CrossRef] [PubMed]
- Lin, J.; Zhang, Y.; Lin, S.; Ding, H.; Liu, W. Integrating Network Pharmacology and Experimental Verification to Explore the Pharmacological Mechanisms of Cordycepin against Pulmonary Arterial Hypertension in Rats. Comb. Chem. High Throughput Screen. 2024, 27, 2776–2789. [Google Scholar] [CrossRef]
- Liao, Y.; Wang, M.; Qin, F.; Liu, T.; Chen, J. Integrating network pharmacology, quantitative transcriptomic analysis, and experimental validation revealed the mechanism of cordycepin in the treatment of obesity. Front. Pharmacol. 2025, 16, 1571480. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chen, Y.; Li, J.; Chen, L.; Wang, J.; Jiang, Y.; Zhang, B. Study on the Mechanism of Ophiocordyceps sinensis in Treating Liver Cirrhosis Based on Network Pharmacology. J. Dali Univ. 2021, 6, 16–21. Available online: http://journal15.magtechjournal.com/Jwk_dlxyzk/EN/Y2021/V6/I8/16.
- Mu, Y.; Zheng, D.; Peng, D.H.; Wang, E.C.; Ye, F.; Wang, Jing. Cordyceps sinensis for the Treatment of Liver Cancer: A Network Pharmacology and Molecular Docking Study. Indian J Pharm Sci 2023, 85, 119–128. Available online: https://www.ijpsonline.com/articles/cordyceps-sinensis-for-the-treatment-of-liver-cancer-a-network-pharmacology-and-molecular-docking-study-5117.html?utm_source=chatgpt.com. [CrossRef]
- Gonzalez-Llerena, J.L.; Treviño-Almaguer, D.; Leal-Mendez, J.A.; Garcia-Valdez, G.; Balderas-Moreno, A.G.; Heya, M.S.; Balderas-Renteria, I.; Camacho-Corona, M.D.R.; Espinosa-Rodriguez, B.A. The Cordyceps Genus as a Potential Source of Bioactive Compounds for Adjuvant Cancer Therapy: A Network Pharmacology Approach. Pharmaceuticals 2025, 18, 667. [Google Scholar] [CrossRef]
- Long, H.; Qiu, X.; Cao, L.; Han, R. Discovery of the signal pathways and major bioactive compounds responsible for the anti-hypoxia effect of Chinese cordyceps. J. Ethnopharmacol. 2021, 277, 114215. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, M.; Qiao, Y.; Shan, Z.; Yang, M.; Li, G.; Xiao, Y.; Wei, L.; Bi, H.; Gao, T. Exploring the mechanisms of action of Cordyceps sinensis for the treatment of depression using network pharmacology and molecular docking. Ann. Transl. Med. 2022, 282. Available online: https://atm.amegroups.org/article/view/91340/html?utm_source=chatgpt.com. [CrossRef]
- Zhou, J.; Wang, M.; Sun, T.; Zhou, X.; Wang, J.; Wang, Y.; Zhang, R.; Luo, R.; Yu, H. Uncovering anti-influenza mechanism of Ophiocordyceps sinensis using network pharmacology, molecular pharmacology, and metabolomics. Medicine 2023, 102, e34843. [Google Scholar] [CrossRef]
- Li, Y.; Wang, L.; Xu, B.; Zhao, L.; Li, L.; Xu, K.; Tang, A.; Zhou, S.; Song, L.; Zhang, X.; Zhan, H. Network pharmacology investigation of Cordyceps sinensis in diabetic nephropathy. J. Diabetes Res. 2021, 2021, 8891093. [Google Scholar] [CrossRef]
- Xi, W.; Zhao, X.; Wang, B.; Zhu, Y.; Li, H. Mechanism of Bailing for diabetic nephropathy based on network pharmacology. Int. J. Clin. Pract. 2024, 2024, 1432594. [Google Scholar] [CrossRef]
- Guan, H.R.; Li, B.; Zhang, Z.H.; Wu, H.S.; He, X.L.; Dong, Y.J.; Su, J.; Lv, G.Y.; Chen, S.H. Network pharmacology of Bailing capsule in polycystic ovary syndrome. BMC Complement. Med. Ther. 2023, 23, 458. [Google Scholar] [CrossRef]
- Singh, M.; Verma, H.; Gera, N.; Baddipadige, R.; Choudhary, S.; Bhandu, P.; Silakari, O. Evaluation of Cordyceps militaris steroids against COVID-19 cytokine storm. J. Biomol. Struct. Dyn. 2024, 42, 5159–5177. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, P.; Zhang, M.; Yang, H.; Liang, B. Cordycepin against colorectal cancer via network pharmacology. Comb. Chem. High Throughput Screen. 2025. [Google Scholar] [CrossRef]
- Li, Y.; Han, Y.; Wang, N.; Bi, Y.; Yang, W.; Yang, Y.; Ren, X. Cordycepin as an anticancer compound. Am. J. Chin. Med. 2025, 53, 2137–2165. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Tania, M. Cordycepin and kinase inhibition in cancer. Drug Discov. Today 2023, 28, 103481. [Google Scholar] [CrossRef]
- Ma, H.; Wang, G.; Guo, X.; Yao, Y.; Li, C.; Li, X.; Xin, M.; Xu, X.; Zhang, S.; Sun, Z.; Zhao, H. Network pharmacology of Cordyceps sinensis in oral lichen planus. J. Oncol. 2022, 2022, 3156785. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhou, H.; Feng, L.; Liang, X.; Zhou, Z.; Li, M.; Su, G. Network pharmacology of Cordyceps militaris in gouty nephropathy. SSRN Electron. J. 2024. [Google Scholar] [CrossRef]
- Yang, L.; Yin, T.; Li, Y. Cordyceps preparations in chronic kidney disease. Integr. Med. Nephrol. Androl. 2025, 12, e24-00030. [Google Scholar] [CrossRef]
- Liu, Y.; Guo, Z.J.; Zhou, X.W. Chinese cordyceps: bioactive components and antitumor mechanisms. Molecules 2022, 27, 6576. [Google Scholar] [CrossRef] [PubMed]
- Marcelo, A.; Brito, F.; Carmo-Silva, S.; Matos, C.A.; Alves-Cruzeiro, J.; Vasconcelos-Ferreira, A.; Koppenol, R.; Mendonça, L.; de Almeida, L.P.; Nóbrega, C. Cordycepin activates autophagy in Machado-Joseph disease. Hum. Mol. Genet. 2019, 28, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhou, Y.; Zhang, X.; Cao, X.; Wu, C.; Guo, P. Cordycepin prevents atherosclerosis via autophagy. Oncotarget 2017, 8, 94726–94737. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, Z.; Jiang, Z.; Luo, P.; Liu, L.; Huang, Y.; Wang, H.; Wang, Y.; Long, L.; Tan, X.; et al. Cordycepin prevents radiation ulcer via NRF2/AMPK. Nat. Commun. 2019, 10, 2538. [Google Scholar] [CrossRef]
- Choi, Y.H.; Kim, G.Y.; Lee, H.H. Anti-inflammatory effects of cordycepin. Drug Des. Dev. Ther. 2014, 8, 1941–1953. [Google Scholar] [CrossRef]
- Yang, J.; Li, Y.Z.; Hylemon, P.B.; Zhang, L.Y.; Zhou, H.P. Cordycepin modulates NLRP3 inflammasome. Biomed. Pharmacother. 2017, 95, 1777–1788. [Google Scholar] [CrossRef]
- Sun, Y.; Huang, W.; Tang, P.; Zhang, X.; Yu, B.; Fan, Y.; Ge, X. Neuroprotective effects of cordycepin in Parkinson’s disease. J. Funct. Foods 2020, 75, 104274. [Google Scholar] [CrossRef]
- Liu, Z.; Lv, L.; Wei, J.; Xie, Y.; Jili, M.; Huang, Y.; Yang, R.; Luo, Y. Cordycepin attenuates macrophage pyroptosis. Front. Pharmacol. 2025, 16, 1526616. [Google Scholar] [CrossRef]
- Zhang, X.L.; Huang, W.M.; Tang, P.C.; Sun, Y.; Qiu, L.; Yu, B.C.; Hong, Y.X.; He, Y.; Ge, X.Q. Cordycepin inhibits mitochondrial fission. Neurotoxicology 2021, 84, 1–13. [Google Scholar] [CrossRef]
- Li, T.; Wen, L.; Cheng, B. Cordycepin alleviates hepatic lipid accumulation. Biochem. Biophys. Res. Commun. 2019, 516, 632–638. [Google Scholar] [CrossRef]
- Hu, Y.; Wu, Y.; Song, J.; Ma, M.; Xiao, Y.; Zeng, B. Advancing Cordyceps militaris industry. Bioengineering 2024, 11, 783. [Google Scholar] [CrossRef]
- European Commission. Novel Food Status Catalogue. Available online: https://food.ec.europa.eu/food-safety/novel-food/novel-food-status-catalogue_en (accessed on 20 January 2026).
- European Commission. Food and Feed Information Portal. Available online: https://ec.europa.eu/food/food-feed-portal/screen/novel-food-catalogue/updates (accessed on 20 January 2026).
- European Commission. RASFF Notification 2024.6020. Available online: https://webgate.ec.europa.eu/rasff-window/screen/notification/703657 (accessed on 20 January 2026).
- European Commission. RASFF Notification 2022.4226. Available online: https://webgate.ec.europa.eu/rasff-window/screen/notification/559027 (accessed on 20 January 2026).
- European Commission. RASFF Notification 2024.2611. Available online: https://webgate.ec.europa.eu/rasff-window/screen/notification/674503 (accessed on 20 January 2026).
- Functional Mushrooms Simplified. EU Regulations Guide. Available online: https://functionalmushrooms.eu/novel-food-and-functional-mushrooms/ (accessed on 20 January 2026).
- Nutra Compliance. EU Novel Food Catalogue Update. Available online: https://nutracompliance.com/EU-novel-food-catalogue-updates-february-2025 (accessed on 20 January 2026).
- European Food Safety Authority. Navigating Novel Foods. Available online: https://www.efsa.europa.eu/en/news/navigating-novel-foods-what-efsas-updated-guidance-means-safety-assessments (accessed on 20 January 2026).
- US Food and Drug Administration. Mushroom Revival Warning Letter. Available online: https://www.fda.gov/inspections-compliance-enforcement-and-criminal-investigations/warning-letters/mushroom-revival-inc-610361-12012020 (accessed on 20 January 2026).
- Antion Consulting. China Novel Food Ingredients System. Available online: https://www.antion.net/En/Blog/view/id/36Ul9no000opK4Da6QEXKVkzxQO0O0OO0O0O.html (accessed on 20 January 2026).
- McEwen, B.S.; Stellar, E. Stress mechanisms. Arch. Intern. Med. 1993, 153, 2093–2101. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S. Stress mediators. N. Engl. J. Med. 1998, 338, 171–179. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S. Neurobiology of stress. Physiol. Rev. 2007, 87, 873–904. [Google Scholar] [CrossRef]
- McEwen, B.S.; Wingfield, J.C. Homeostasis and stress. Horm. Behav. 2010, 57, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. Origin of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef]
- Nathan, C.; Ding, A. Nonresolving inflammation. Cell 2010, 140, 871–882. [Google Scholar] [CrossRef]
- Serhan, C.N.; Levy, B.D. Resolvins in inflammation. J. Clin. Investig. 2018, 128, 2657–2669. [Google Scholar] [CrossRef]
- Serhan, C.N.; Gupta, S.K.; Perretti, M.; Godson, C.; Brennan, E.; Li, Y.; Soehnlein, O.; Shimizu, T.; Werz, O.; Chiurchiù, V.; Azzi, A.; Dubourdeau, M.; Gupta, S.S.; Schopohl, P.; Hoch, M.; Gjorgevikj, D.; Khan, F.M.; Brauer, D.; Tripathi, A.; Cesnulevicius, K.; Lescheid, D.; Schultz, M.; Sarndahl, E.; Repsilbers, D.; Kruse, R.; Sala, A.; Haeggstrom, J.Z.; Levy, B.D.; Janos, G.; Filepx, J.G.; Wolkenhauer, O. Atlas of inflammation resolution. Mol. Aspects Med. 2020, 74, 100894. [Google Scholar] [CrossRef]
- US Food and Drug Administration. Dietary Supplement Health and Education Act. Available online: https://ods.od.nih.gov/About/DSHEA_Wording.aspx (accessed on 20 January 2026).
- European Food Safety Authority Antioxidant health claims guidance, E.F.S.A.J. 2012, 10, 2474. Available online: https://www.efsa.europa.eu/sites/default/files/consultation/nda110426a%2C0.pdf (accessed on 20 January 2026).
- European Food Safety Authority Health claim applications guidance, E.F.S.A.J. 2011, 9, 2170. Available online: https://efsa.onlinelibrary.wiley.com/doi/pdf/10.2903/j.efsa.2016.4367 (accessed on 20 January 2026).
- European Food Safety Authority Physical performance health claims, E.F.S.A.J. 2012, 10, 2817. Available online: https://efsa.onlinelibrary.wiley.com/doi/epdf/10.2903/j.efsa.2012.2817 (accessed on 20 January 2026).
- European Medicines Agency. Reflection paper on adaptogens. Available online: https://www.ema.europa.eu/en/documents/scientific-guideline/reflection-paper-adaptogenic-concept_en.pdf (accessed on 20 January 2026).
- Panossian, A. Adaptogenic activity. Ann. N. Y. Acad. Sci. 2017, 1401, 49–64. [Google Scholar] [CrossRef]
- Cohen Healthcare. Adaptogens and FDA/FTC compliance. Available online: https://cohenhealthcarelaw.com/adaptogens-and-fda-ftc-compliance/ (accessed on 20 January 2026).
- World Health Organization. WHO Traditional Medicine Strategy 2014–2023; WHO Press: Geneva, Switzerland, 2013; Available online: https://www.who.int/publications/i/item/9789241506096 (accessed on 20 January 2026).
- European Medicines Agency. EU herbal monograph on Rhodiola rosea. Available online: https://www.ema.europa.eu/en/documents/herbal-monograph/final-european-union-herbal-monograph-rhodiola-rosea-l-rhizoma-et-radix-revision-1_en.pdf (accessed on 20 January 2026).
- Panossian, A.; Seo, E.J.; Efferth, T. Novel molecular mechanisms for the adaptogenic effects of herbal extracts on isolated brain cells using systems biology. Phytomedicine 2018, 50, 257–284. [Google Scholar] [CrossRef] [PubMed]
- Panossian, A.; Lemerond, T.; Efferth, T. Botanical hybrid preparations and stress-related diseases. Pharmaceuticals 2024, 17, 483. [Google Scholar] [CrossRef]
- Qin, P.; Li, X.; Yang, H.; Wang, Z.-Y.; Lu, D. Therapeutic Potential and Biological Applications of Cordycepin and Metabolic Mechanisms in Cordycepin-Producing Fungi. Molecules 2019, 24, 2231. [Google Scholar] [CrossRef]
- Efferth, T. Establishing traditional Chinese medicine in Europe. Chin. Herb. Med. 2025, 17, 409–413. [Google Scholar] [CrossRef] [PubMed]
- Chrousos, G.P. Stress and disorders of the stress system. Nat. Rev. Endocrinol. 2009, 5, 374–381. [Google Scholar] [CrossRef]
- Dhabhar, F.S. Effects of stress on immune function: The good, the bad, and the beautiful. Immunol. Res. 2014, 58, 193–210. [Google Scholar] [CrossRef] [PubMed]
- Segerstrom, S.C.; Miller, G.E. Psychological stress and the human immune system: A meta-analytic study. Psychol. Bull. 2004, 130, 601–630. [Google Scholar] [CrossRef]
- Lupien, S.J.; McEwen, B.S.; Gunnar, M.R.; Heim, C. Effects of stress throughout the lifespan on the brain, behaviour and cognition. Nat. Rev. Neurosci. 2009, 10, 434–445. [Google Scholar] [CrossRef]
- Arnsten, A.F.T. Stress signalling pathways that impair prefrontal cortex structure and function. Nat. Rev. Neurosci. 2009, 10, 410–422. [Google Scholar] [CrossRef]
- Ferrucci, L.; Fabbri, E. Inflammageing: Chronic inflammation in ageing, cardiovascular disease, and frailty. Nat. Rev. Cardiol. 2018, 15, 505–522. [Google Scholar] [CrossRef]
- World Health Organization. WHO Monographs on Selected Medicinal Plants; WHO Press: Geneva, Switzerland, 1999; Vol. 1, pp. 168–182. [Google Scholar]
- World Health Organization. WHO Monographs on Selected Medicinal Plants; WHO Press: Geneva, Switzerland, 2002; Vol. 2, pp. 82–96. [Google Scholar]
- World Health Organization. WHO Monographs on Selected Medicinal Plants; WHO Press: Geneva, Switzerland, 2007; Vol. 3, pp. 296–313. [Google Scholar]
- World Health Organization. WHO Monographs on Selected Medicinal Plants; WHO Press: Geneva, Switzerland, 2009; Vol. 4, pp. 373–391. [Google Scholar]
- World Health Organization. WHO Monographs on Selected Medicinal Plants in the Newly Independent States (NIS); WHO Press: Geneva, Switzerland, 2010; Vol. 5, pp. 141–159. [Google Scholar]
- Samuelsson, G.; Bohlin, L. Drugs of Natural Origin, 6th ed.; Swedish Academy of Pharmaceutical Sciences: Stockholm, Sweden, 2009; Available online: https://www.routledge.com/Drugs-of-Natural-Origin-ATreatiseof-Pharmacognosy-Seventh-Edition/Samuelsson-Bohlin/p/book/9789198094251 (accessed on 10 February 2025).
- British Herbal Medicine Association. British Herbal Pharmacopoeia; BHMA: Exeter, UK, 1996; pp. 89–90. [Google Scholar]
- British Herbal Compendium; Bradley, P.R., Ed.; BHMA: Exeter, UK, 1996; Vol. 1, pp. 89–91. [Google Scholar]
- British Herbal Compendium; Bradley, P.R., Ed.; BHMA: Exeter, UK, 1996; Vol. 1, pp. 115–118. [Google Scholar]
- Heinrich, M.; Barnes, J.; Prieto-Garcia, J.; Gibbons, S.; Williamson, E.M. Fundamentals of Pharmacognosy and Phytotherapy, 4th ed.; Elsevier: Amsterdam, The Netherlands, 2023. [Google Scholar]
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W.; Miller, A.H.; Mantovani, A.; Weyand, C.M.; Barzilai, N.; Goronzy, J.J.; Rando, T.A.; Effros, R.B.; Lucia, A.; Slavich, G.M. Chronic inflammation in the etiology of disease across the life span. Nat. Med. 2019, 25, 1822–1832. [Google Scholar] [CrossRef]
- Cheng, W.; Zhang, X.; Song, Q.; Lu, W.; Wu, T.; Zhang, Q.; Li, C. Nucleosides in Ophiocordyceps sinensis. Mycology 2017, 8, 318–326, Link not found. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Zhao, J.; Zhu, Q.; Li, S.P. Macrophage biospecific extraction. J. Pharm. Biomed. Anal. 2007, 44, 439–443. [Google Scholar] [CrossRef]
- Tuli, H.S.; Sharma, A.K.; Sandhu, S.S.; Kashyap, D. Cordycepin therapeutic potential. Life Sci. 2013, 93, 863–869. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Wong, J.H.; Fu, M.; Ng, T.B.; Liu, Z.K.; Wang, C.R.; et al. Antioxidants from Cordyceps militaris. Phytomedicine 2011, 18, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, H.; Akaki, J.; Nakamura, S.; Okazaki, Y.; Kojima, H.; Tamesada, M.; Yoshikawa, M. Apoptosis-inducing sterols. Chem. Pharm. Bull. 2009, 57, 411–414. [Google Scholar] [CrossRef]
- Jia, J.M.; Tao, H.H.; Feng, B.M. Cordyceamides A and B. Chem. Pharm. Bull. 2009, 57, 99–101. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, S.; Liu, H.; Liu, X.; Che, Y. Cycloaspeptides from Isaria farinosa. J. Nat. Prod. 2009, 72, 1364–1367. [Google Scholar] [CrossRef]
- Chen, P.X.; Wang, S.; Nie, S.; Marcone, M. Properties of Cordyceps sinensis. J. Funct. Foods 2013, 5, 550–569. [Google Scholar] [CrossRef]
- Tshikhudo, P.P.; Mabhaudhi, T.; Koorbanally, N.A.; et al. β-Carboline alkaloids. Chem. Biodivers. 2024, 21, e202301263. [Google Scholar] [CrossRef]
- Kim, J.H.; Park, D.K.; Lee, C.H.; Yoon, D.Y. Isoflavone from Cordyceps militaris. Phytother. Res. 2012, 26, 1807–1812. [Google Scholar] [CrossRef]
- Kneifel, H.; König, W.A.; Loeffler, W.; Müller, R. Ophiocordin antibiotic. Arch. Microbiol. 1977, 113, 121–130. [Google Scholar] [CrossRef]
- Chiu, C.P.; Liu, S.C.; Tang, C.H.; et al. Anti-inflammatory cerebrosides. J. Agric. Food Chem. 2016, 64, 1540–1548. [Google Scholar] [CrossRef] [PubMed]
- Dewi, L.; Liao, Y.C.; Jean, W.H.; et al. Cordyceps and muscle stem cells. Food Funct. 2024, 15, 4010–4020. [Google Scholar] [CrossRef]
- Hao, L.; Wang, Q.; Kobayashi, M.; Tamesada, M.; Wang, H.J. Cordyceps in lung cancer. Biotherapy 2008, 22, 345–349. [Google Scholar]
- Hao, L.; Wang, Q.; Wang, B.; Wang, H.J. Cordyceps with NP chemotherapy. J. Dalian Med. Univ. 2007, 29, 563–565. [Google Scholar]
- Jung, S.J.; Jung, E.S.; Choi, E.K.; Sin, H.S.; Ha, K.C.; Chae, S.W. Immunomodulatory effects of a Cordyceps mycelium extract (Paecilomyces hepiali; CBG-CS-2): randomized double-blind clinical trial. BMC Complement. Altern. Med. 2019, 19, 77. [Google Scholar] [CrossRef]
- Morikubo, K.; Uchida, M.; Kume, A.; Tsunoo, H.; Tajima, T.; Nagata, A. Cordyceps and mental condition. Jpn. Pharmacol. Ther. 2005, 33, 729–734. [Google Scholar]
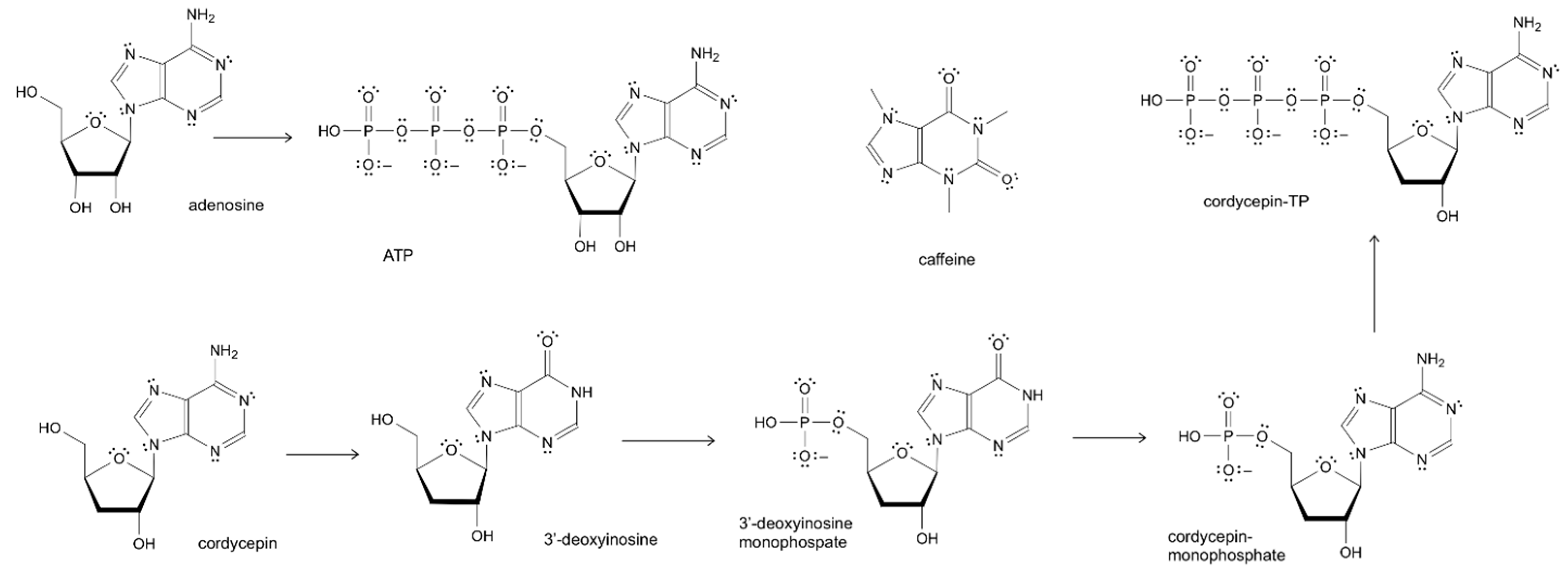
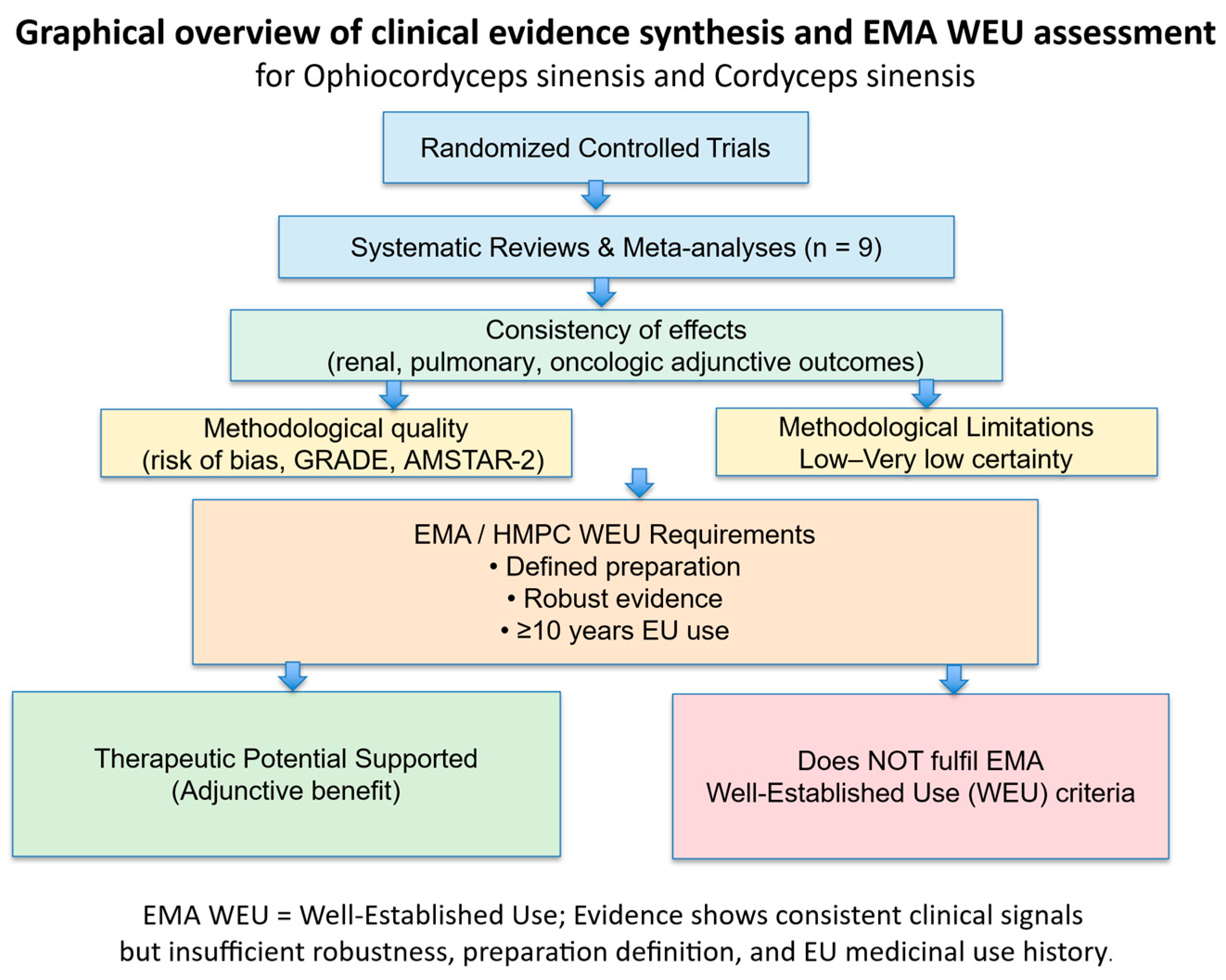
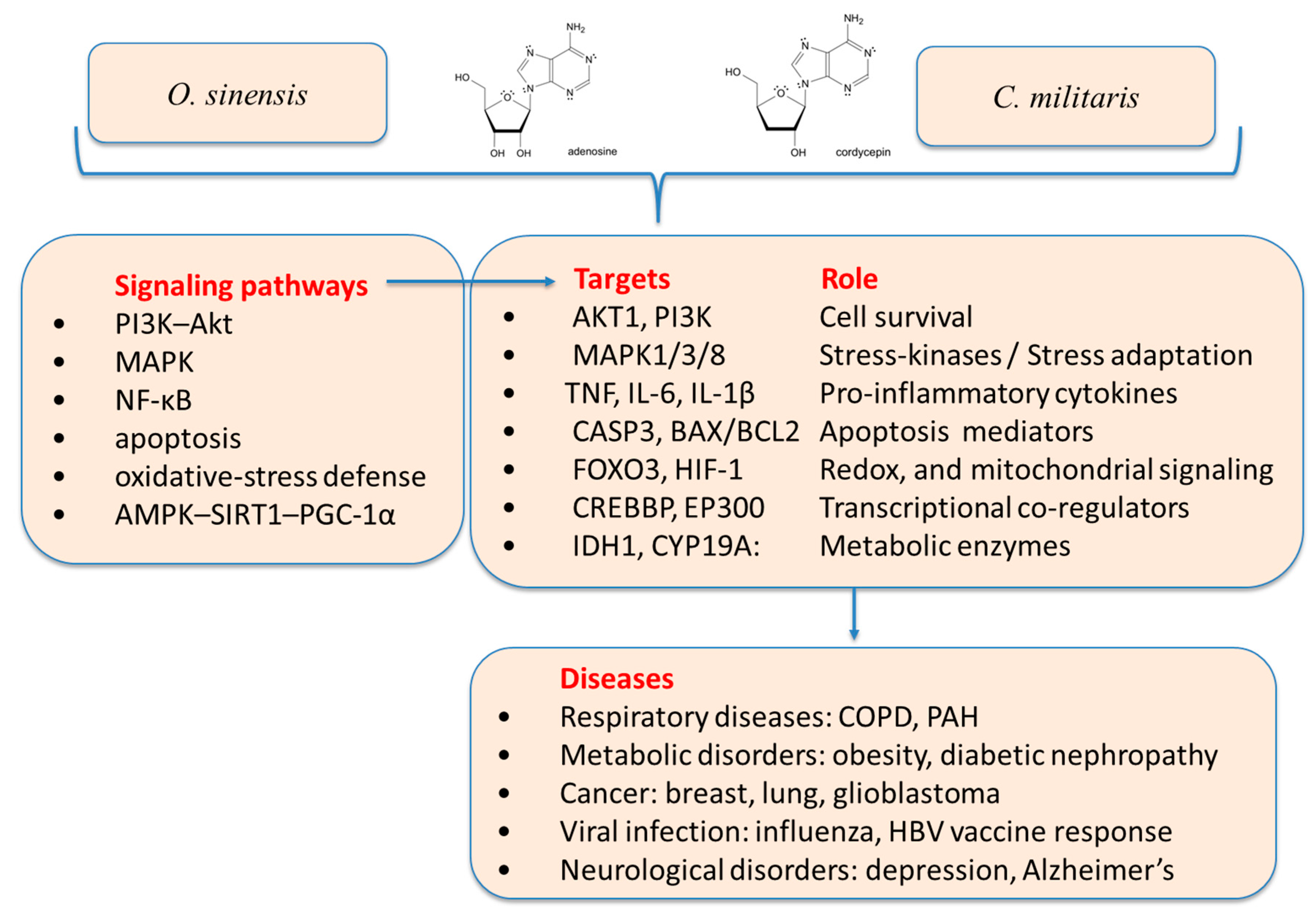
| Markers | O. sinensis, mg/g | C. militaris, mg/g | References |
|---|---|---|---|
| Adenosine | ≈ 0.2-10 | 0.3–2.5 | [86,87,88] |
| Cordycepin | <0.2 (0.006–0.075) | ~6.6 (3–26) | [53,87,88,89,90] |
| Polysaccharides | ≥60 (≈ 50–100) | ≥80 (~30–150) | [16,88,91] |
| Total nucleosides | ~2.0–3.1 | [93,94] | |
| Total Ergosterol | ~1.9- 2.7 | ~2–3 | [95,96] |
| Mattitol | 25.6-115.7 | [89,97] |
| Compound | Receptor Action | Net CNS Effect | Fatigue Impact | Mechanistic Reason |
|---|---|---|---|---|
| Adenosine | Agonist at A1, A2A | Sedative | Increases fatigue | Inhibits neuronal firing, promotes sleep pressure |
| Caffeine | Antagonist at A1, A2A | Stimulant | Reduces fatigue | Blocks adenosine, increases dopamine + cortical activity |
| Cordycepin | Minimal activity (in vivo) | Neutral | No proven effect | Rapid deamination, poor BBB penetration, weak receptor affinity |
| Pathway / Module | Key Genes / Molecules | Physiological Roles | Representative Publications | ||
|---|---|---|---|---|---|
| AMPK–mTOR–Autophagy Axis | PRKAA1/2 (AMPKα), RPTOR, RICTOR, MTOR, ULK1, BECN1, LC3B, ATG5/7, SQSTM1 | Cellular energy sensor; promotes autophagy, ATP conservation, and repair during stress. | Hawley et al. ,2020 [36] Marcelo et al.,2019 [212] Li et al. 2017.[213] |
||
| NRF2–KEAP1 Antioxidant Axis | NFE2L2 (NRF2), KEAP1, HMOX1, NQO1, GCLC, GCLM, SOD2, CAT, GPX1 | Regulates antioxidant enzymes, detoxification, redox homeostasis, and cytoprotection. | Wang Z et al., 2019 [214] |
||
| TLR4–MyD88–NF-κB / MAPK Inflammatory Axis | TLR4, MYD88, NFKB1, RELA, IKBKB, MAPK14, MAPK8, MAPK1/3, TNF, IL6, PTGS2, NOS2 | Controls innate immune activation; Cordycepin suppresses pro-inflammatory cytokines and iNOS/COX-2 expression. | Choi YH et al., 2014 [215] Yang J et al., 2017 [216] Sun Y et al., 2020 [217] |
||
| NLRP3 Inflammasome / Pyroptosis Regulation | NLRP3, CASP1, GSDMD, IL1B, IL18 | Limits pyroptotic cell death and IL-1β release; anti-inflammatory and neuroprotective effects. | Liu Z et al., 2025 [218] Zhang X et al., 2021 [219] |
||
| PI3K–AKT / FOXO–SIRT Pathway | PIK3CA/B, AKT1, FOXO3, SIRT1, TP53 | Balances survival vs repair; mediates stress resistance, longevity, and metabolic adaptation. | Li T et al., 2019 [220] |
||
| Region | O. sinensis | C. militaris | Notes |
|---|---|---|---|
| China | Traditionally medicinal, wild OS is not a general everyday food. Listed in the Chinese Pharmacopoeia as a traditional medicinal material (TCM drug), not a general food. Ophiocordyceps sinensis mycelium and fermented products (some strains) may appear as approved food ingredients if separately registered. | Approved as a “new resource food” (novel food ingredient) since 2009; widely used in foods/health foods in China. | Used in decoctions, capsules, and medicinal soups. Only specific strains of cultivated mycelium (e.g. Cs-HK1) are allowed as food supplements. Contemporary reviews of the Chinese regulatory framework note C. militaris approval as novel/new resource food (2009). [221] |
| Japan / Korea | Classified as a traditional medicinal material, used in Kampo or functional foods under regulatory oversight. | In Asian countries, the cultural and historical use supports a more permissive edible status for C. militaris | In Asian countries, the cultural and historical use supports a more permissive edible status for C. militaris |
| USA | Marketed mainly as dietary supplements (mycelium/extracts). No GRAS (Generally Recognized As Safe) listing for wild O. sinensis; marketed forms are dietary supplements under DSHEA, typically using cultivated mycelium (e.g. Cordyceps sinensis Cs-4). | Marketed as dietary supplements, enforcement actions are taken when drug-like claims are made. | The U.S. classifies many mushroom extracts as dietary supplements. If a product claims to treat a disease, the FDA may treat it as a drug. U.S. Food and Drug Administration. FDA warning letters show Cordyceps products can be challenged if marketed with disease claims. There’s no public GRAS for wild OS itself. |
| EU | Considered a novel food if intended as a food ingredient, wild O. sinensis itself is not authorized under general edible mushroom lists. Not novel in food supplements (entry in the EU Novel Food status catalogue). | “Not yet authorised – novel food.” | Authorization required under the Novel Foods Regulation. The European Commission’s Novel Food portal lists C. militaris (mycelium & fruiting body) as not yet authorised; multiple RASFF alerts flag unauthorised use of C. militaris in supplements. O. sinensis is shown as “not novel in food supplements.” |
| Sweden | Follows EU Novel Food rules. | Follows EU Novel Food rules. | In the EU, Novel Food law is harmonised; Sweden applies the EU stance. Check the EC catalogue and consult the Swedish National Food Agency if you need product-specific confirmation. |
| Aspect | Inflammation | Stress response |
|---|---|---|
| Purpose | Restore homeostasis | Restore homeostasis |
| Nature | Protective, adaptive | Protective, adaptive |
| Mediators | Cytokines, eicosanoids | Hormones, neurotransmitters |
| Resolution | Actively regulated | Actively regulated |
| Pathology arises when | Chronic, unresolved | Chronic, dysregulated |
| Localization | Primarily local | Systemic |
| Primary system | Immune system | Neuroendocrine system |
| Measurability | Clear biomarkers (CRP, IL-6) | Context-dependent biomarkers (cortisol variability) |
| Disease framing | Recognized pathological entity | Often framed as a risk factor |
| Consequences of failed resolution | Chronic inflammation:
|
Chronic stress / inflammaging:
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.



