Submitted:
23 January 2026
Posted:
26 January 2026
You are already at the latest version
Abstract
This study reports on the presence of two uncommon tetraodontiform fishes and reviews the presence of species of this order in Spanish marine waters. Two tetraodontid specimens (Family Tetraodontidae) were caught in Atlantic waters of the coast of Galicia, northwestern Spain. A specimen of Sphoeroides pachygaster was collected in 2021 off the Costa da Vela, and a specimen of Ephippion guttifer was captured in 2025 within the Ría de Pontevedra, both locations situated in southern Galicia. Morphological analyses, supported by photographic evidence and DNA barcoding, confirmed the preliminary taxonomic identification of both species. The histological reproductive analysis of the Ephippion guttifer specimen showed a female in spawning capable phase. These findings constitute the first verified record of S. pachygaster and the second of E. guttifer in Galician waters. A comprehensive list of tetraodontiform species found in Spanish waters across five geographical demarcations has been compiled and updated. Historically, a total of 26 species across five families have been reported in Spanish waters, counting 22 in the Canary Islands and 15 in the Spanish Iberian Peninsula and Balearic Islands. Additionally, a review of the presence of the neurotoxic tetrodotoxins (TTXs) or paralytic shellfish toxins (PSTs) in each species is also included, bringing an up-to-date overview of a largely unexplored field in European waters. The increasing occurrence of tetraodontiform fishes in Spanish waters provides further evidence of the progressive tropicalization of the Spanish marine environment.
Keywords:
Galicia
; pufferfishes
; barcoding
; morphology
; distribution
; tropicalization
; tetrodotoxins
; paralytic shellfish toxins
1. Introduction
Although the order Tetraodontiformes has long been considered a valid category, recent phylogenetic studies on ray-finned fishes have proposed a reorganization of the group. Under this new classification, Tetraodontiformes would be considered a suborder, Tetraodontoidei, within the order Acanthuriformes [1]. However, this proposal has not yet been fully accepted by the scientific community. Therefore, until this new classification is confirmed or refuted, the traditional classification is considered herein for sake of clarity.
There are 412 extant species in the 10 families of living tetraodontiform fishes distributed in tropical to temperate marine and freshwater environments worldwide, showing a remarkable diversity in shape, size, and ecological traits [2]. Tetraodontiform fishes are distinguished by their rigid bodies with diverse shapes, some of which are highly inflatable; small mouths equipped with relatively few modified, enlarged teeth or substantial, beak-like tooth plates; reduced gill openings located near the pectoral-fin base; and scales typically modified into spines, ossicles, or fused bony plates, with pelvic fins either reduced or absent.
The family Tetraodontidae is the most speciose within the order Tetraodontiformes, comprising 193 valid species across 27 genera of pufferfishes [3]. These species predominantly inhabit shallow, warm, tropical seas and freshwater environments globally, although certain species, such as the blunthead puffer Sphoeroides pachygaster (Müller & Troschel, 1848), has been recorded at depths exceeding 350 meters [2]. Tetraodontid species have been extensively studied in genomic research to elucidate genome architecture and evolutionary mechanisms [4], toxin ecology and physiology [5], and invasive processes impacting ecosystems and fisheries [6].
The classification and identification of tetraodontiform fishes have primarily relied on morphological characteristics such as stripes, spots, body color, and mouth shape. However, differentiating closely related species within this order is often difficult when based solely on morphology, as many species have very similar characteristics that make them difficult to distinguish [7]. Consequently, the use of molecular tools coupled with morphological taxonomy in an integrative context has been gaining importance in the last decades. Several molecular markers have been employed in Tetraodontiformes for phylogenetic [8] and taxonomic purposes [9]. Among them, the cytochrome c oxidase subunit I (COI) stands out as the most widely used due to its proven reliability for fish barcoding and also specifically for differentiation of tetraodontiform species [7,10]. The widespread use of this marker is currently facilitating further progress in clarifying the taxonomy of this order, detecting misidentifications, synonyms, and the screening for cryptic species.
Preliminary identification of the studied specimens indicated that they belonged to the genera Ephippion Bibron in Duméril 1855 and Sphoeroides Anonymous 1798. The genus Ephippion is monospecific, with the spiny pufferfish Ephippion guttifer (Bennett 1831) as the sole representative of the genus. This species is distributed in the eastern Atlantic Ocean, from Galicia (north-western Spain) to Angola, and in the Mediterranean Sea, in Algeria, Tunisia, and Italy [11,12].
Considering the growing number of reports of species beyond their known distribution range, the term neo-native has been recently defined to include species that change their range and establish themselves beyond their historical boundaries as a result of human-induced environmental changes [13]. The arrival of neo-native species can pose significant ecological risks, including displacing native species, introduction of new parasites, altering habitats, and causing local extinctions. In addition, pufferfishes are notorious for their toxicity and may also display aggressive behavior, thus posing a threat to human health [14]. However, biological information on these species is frequently scarce and rely on information obtained from their natural areas of distribution, often with very different ecological conditions.
Knowledge about reproductive ecology and potential of fishes provide key parameters to understanding population dynamics. Only information on the reproductive phase of a male specimen of E. gutiffer has been reported [11]. Among pufferfish, reproductive strategies show great diversity. Some marine species, like Torquigener sp., construct elaborate geometric nests on sandy seabeds, and males invest significant energy in building these structures, even providing paternal care in some species [15,16]. Other species, such as Sphoeroides nephelus (Goode & Bean, 1882) and Sphoeroides annulatus (Jenyns, 1842), exhibit batch spawning with asynchronous oocyte development and protracted reproductive seasons, often influenced by environmental factors like temperature, photoperiod, and lunar cycles [17,18,19]. Freshwater and estuarine species, including Carinotetraodon travancoricus (Hora & Nair, 1941) and Tetraodon schoutedeni Pellegrin, 1926, show batch spawning, low fecundity, and specific courtship behaviors [20,21,22]. In any case, studies on reproductive ecology of pufferfishes are scarce.
Tetraodontiformes, mainly pufferfish of the family Tetraodontidae, can be toxic, containing tetrodotoxins (TTXs) and/or paralytic shellfish toxins (PSTs). However, the toxin profile differs depending on the genus or species [23,24]. In marine ecosystems PSTs are mainly produced by certain dinoflagellates and thereafter transferred and bioaccumulated through the aquatic food web [25], reaching several invertebrate and vertebrate groups, including fish [26,27]. TTXs are believed to originate from bacteria belonging to several bacteria phylum, however several studies suggest their association to specific dinoflagellate blooms [28]. PSTs and TTXs are potent neurotoxins that block the sodium channels in the nerve cells and skeletal muscles and produce a blockade of ion conductance, resulting in the occurrence of typical symptoms and signs (paraesthesia of lips, tongue, and pharynx, weakness, dizziness, gastrointestinal and neurological symptoms [29,30]. Ingestion of toxic fish tissues with PSTs and/ or TTXs can lead to severe illness and death.
Considering the scenario described above, the study of two new records of tetraodontids identified through integrative taxonomy, provided the basis for undertaking a comprehensive review of the records within the entire order in Spain. The aim of this work is, therefore, to update relevant information on species of this order with a proven presence in the study area, with special attention to the two new records and their associated biological traits as well as to gather information on the potential risk posed by the presence of toxins.
2. Materials and Methods
2.1. Study Area
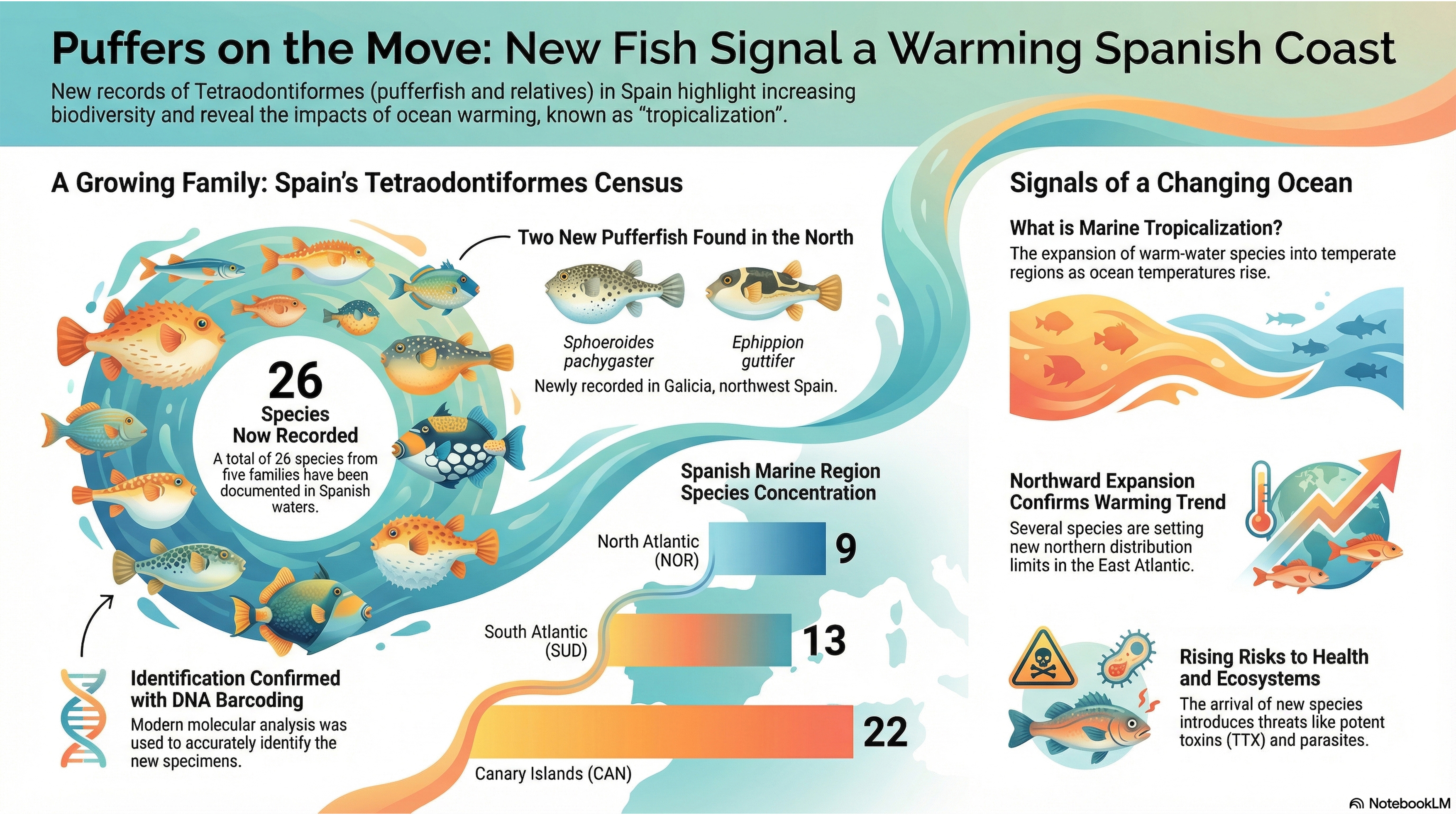
Spain, with a total coastline extending over 7879 km, represents the southernmost point of continental Europe, with most of its territory situated on the Iberian Peninsula, including the Canary and Balearic Islands and the North African cities of Ceuta and Melilla [31]. To improve the management and conservation of the species, the Spanish government established five marine regions or demarcations based on biogeographic, oceanographic, and hydrological characteristics. The Atlantic region include the North Atlantic (NOR, Spanish north coast), South Atlantic (SUD, Spanish coast of the Gulf of Cádiz), and the Canary Islands (CAN, the Canary Islands) demarcations, whereas the Mediterranean region comprises the Levantine-Balearic (LEBA, East coast of Spain and Balearic Islands) and the Strait of Gibraltar and Alboran Sea (ESAL) demarcations (Figure 1).
2.2. Sampling Data and Morphological Analysis
The two specimens described were caught as by-catch by professional artisanal fishers working in Galician waters, northwest of Spain. Specimens were first preserved frozen and later thawed and measured to the nearest mm. Morphometric and meristic characters were determined according to Shipp [32]. The specimens were preserved frozen and deposited in the fish collection of the Museo de Historia Natural Luis Iglesias, in Santiago de Compostela, reference code MHN USC 25103-1 for E. guttifer and MHN USC 25234 for S. pachygaster. Total length (TL) and Standard length (SL) were used throughout.
2.3. Molecular Analysis
To generate the DNA barcodes for both tetraodontiforms, the partial sequence of the 5’ end of the mitochondrial genome locus COI was first obtained. DNA was extracted from a muscle sample from each of the specimens caught using the E.Z.N.A. Tissue DNA Kit, following the manufacturer’s instructions. Amplification was performed by PCR, using the universal fish primer sets C_FishF1t1-C_FishR1t1, and the recommended reaction conditions [33], employing the Horse-Power Taq DNA polymerase MasterMix from Canvax Reagents (Valladolid, Spain). In both cases, amplicons of the expected size were obtained and sequenced using the Sanger method [34] at the Centro de Apoyo Científico y Tecnológico a la Investigación of the University of Vigo (https://cactiweb.uvigo.es/). The nucleotide sequences and their corresponding trace files, together with images of the specimens and the geographical locations of the catches, were deposited in the Marine Fishes from Galicia project created in the BOLD database (https://boldsystems.org/), from where they were also submitted to the GenBank database (https://www.ncbi.nlm.nih.gov/genbank/). Sequence analysis was performed in MEGA11 [35]. To this end, an alignment was performed with all publicly available E. guttifer sequences and a selection of S. pachygaster sequences, provided that their geographical origin was known. Next, a clustering analysis was done using the Neighbour-Joining algorithm in pairwise comparison mode with 1,000 iterations. The resulting dendrogram was edited using the open-source programme Inkscape (https://inkscape.app/). The proportion of nucleotide differences between sequences was measured as uncorrected p-distance.
2.4. Reproductive Analysis
Total weight of thawed ovary was recorded. One lobe was fixed in 4% buffered formaldehyde. One sub-sample from central part of the ovary was histologically processed (embedded in paraffin, sectioned and stained with Haematoxylin-Eosin) for maturity stage determination. Additionally, four sub-samples (20-46 mg in weight) from anterior, central and posterior part of the lobe were taken to estimate fecundity by applying the gravimetric method [36] combined with the whole-mounts method [37]. Oocytes were separated from connective tissue using a washing process (modified from Lowerre-Barbieri and Barbieri [38]) and by size with sieve of 500 µm. All sub-samples were photographed using a stereo microscope Nikon SMZ18 connected to a video camera Nikon Fi13. Oocytes were counted and measured using the software Fiji [39]. Oocyte density per sub-sample (oocytes per gram of ovary weight, OD), as well as potential (PF) and relative (PFrel) fecundity (oocytes and oocytes per gram of female weight, respectively) were estimated with the package stats of the software R [40] and RStudio console [41], results were plotted with the package ggplot2 [42].
2.5. Spanish Tetraodontiform Species and Vulnerability
A list of tetraodontiform species has been compiled for each geographical area based on exhaustive bibliographic research and complemented with GBIF records containing relevant information to permit the validation of the original identification [43]. The most recent previous list [44] was used to determine temporal and spatial changes. Records found outside the five demarcations, in the Spanish African territory, were included, by proximity in the nearest demarcation. Thus, the records of Aluterus monoceros (Linnaeus, 1758) from the Chafarinas Islands [45], and Lagocephalus sceleratus (Gmelin, 1789) from Ceuta [46], were included in the ESAL demarcation. The vulnerability status of species recorded in Spanish waters has been analysed following the International Union for Conservation of Nature (IUCN) Red List [47]. The IUCN assesses species for their vulnerability status, which indicates the risk of extinction. According to IUCN criteria, species are consider threatened if they are categorized as critically endangered (CR), endangered (EN), or vulnerable (VU), and non-threatened if categorized as near threatened (NT), least concern (LC), or data deficient (DD). Species without evaluation was categorized as not evaluated (NE), and not applicable (NA) category was employed for species of marginal occurrence in Europe (i.e. species whose population in Europe represents less than 1% of the total population) [48].
2.6. Marine Biotoxins
The presence of two groups of marine biotoxins normally associated with certain tetraodontiform fishes, TTXs and PSTs, on the Spanish species was explored in literature. For that purpose, a comprehensive literature search was conducted. The keywords combined the name of each of the species reported in Spain, entered individually, with these others: intoxications, tetrodotoxins, TTXs, paralytic shellfish poisoning, PSP, paralytic shellfish toxins, PSTs, saxitoxins and STX.
The presence of any of those toxin groups in a fish species, independently of the fish capture location and toxicities levels, is considered. Presence/absence were categorized as detected (D, when they were detected and described as detected or quantified in a manuscript), not detected (ND, the species has been examined and no toxin was detected) and not evaluated (NE, when there is not information).
3. Results
3.1. New Tetraodontidae Records
Two tetraodontid specimens were examined (Figure 2): Sphoeroides pachygaster MHN USC-25234, 220 mm LT, 26 October 2021, 42.306, -8.890, 15 m depth; Ephippion guttifer MHN USC-25103-1, 567 mm LT, 14 October 2025, 42.409, -8.688, 6 m depth. The main meristic and morphometric characters of both specimens are summarized in Table 1.
3.1.1. Molecular Analysis
The molecular dendrogram (Figure 3) shows how the new sequences grouped unequivocally with those corresponding to their respective species in separate clades, with a minimum distance between them of 17.64%. The mean intraspecific distance was 0.44% (0-0.83%) in the case of E. guttifer, and 0.64% (0-2.23%) in the case of S. pachygaster. The wide range of distances between sequences of the latter, compared to its relatively low average value, is due to the values between the sequences from the United States and Uruguay in relation to the others, whose subclade can be seen in the dendrogram, and which corresponds to values ranging from 1.99% to 2.23%.
3.1.2. Reproductive Analysis
Oocytes in advanced vitellogenesis predominated in the histological section of E. guttifer, whereas oocytes in early vitellogenesis, cortical alveoli, and atretic oocytes were present but in low numbers. Some melanomacrophage centers are also observed, likely linked to the atretic process. Apparently, no postovulatory follicles were detected in the sample; however, because the gonad was frozen and thawed prior to fixation, the structures were not preserved with sufficient quality to confirm this with certainty (Figure 4).
Oocyte diameter ranged between 214 and 1134 µm (692.4±192.3 µm, mean±s.d.). Sub-sample 1 (sub1) showed bi-modal distribution (371 and 851 µm) while the other ones presented a single mode that were 407, 793 and 772 µm for the sub-samples 2, 3 and 4 respectively. When all diameter data were pooled together, bi-modal distribution was detected with a main mode equal to 782 µm and a secondary mode of 383 µm (Figure 5).
The number of oocytes in each sub-sample ranged between 85 and 197 oocytes (mean 141 oocytes), what corresponds to an OD between 1891 and 9850 oocytes per ovary gram (Table 2).
Given the large difference observed in OD among subsamples -up to five times higher in sub3 compared to sub2- the potential fecundity was calculated by pooling data from all subsamples. This resulted in a PF of 1,335,584 oocytes and a relative PF of 533.4 oocytes per gram of female gutted weight. If fecundity is estimated as the average fecundity from the 4 sub-samples, it results in a PF of 1,661,950 oocytes and PFrel of 663.7 oocytes per gram of female body weight.
3.2. Spanish Tetraodontiform Species
A list of Tetraodontiformes recorded by Spanish marine demarcations is shown in Table 3, including a total of 26 species belonging to five families.
The most represented family is Tetraodontidae, with seven species, while the least represented is Molidae, with four species. Following the latest review [44], five new species were added to the list: Cantherhines pullus (Ranzani, 1842) and Cantherhines macrocerus (Hollard, 1853) (Monacanthidae), Chilomycterus mauretanicus (Le Danois, 1954) (Diodontidae), Mola alexandrini (Ranzani 1834) (Molidae) and Melichthys niger (Bloch, 1786) (Balistidae).
In terms of spatial distribution, CAN, with 22 species (85% of the total), is the most speciose region, followed by SUD (13 species, 50%), ESAL (12 species, 46%), NOR (9 species, 35%) and LEBA (8 species, 31%).
3.2.1. Vulnerability Status
Table 4 shown the IUCN vulnerability status of Spanish species of Tetraodontiformes at the global and European levels.
The global assessment covers 25 of the 26 species (96.2%) and most of them (84.6%) were assessed as LC with only three species (11.5%) was vulnerable. At the European level, the assessment is reduced to 14 species (53.8%), seven (26.9%) assessed as LC and none into a vulnerable category.
3.2.2. Report of TTXs and/or PSTs in the Spanish Tetraodontiform Species
4. Discussion
Biometrics measurements and meristic counts reported in this study agree in general terms with previous descriptions of E. guttifer and S. pachygaster [2,12,32]. The molecular analysis supports the reliability of the morphological identification, showing the newly generated sequences clustered in the same clades as those retrieved from public databases and assigned to the same species, although obtained from different geographical areas.
Molecular techniques based on voucher specimens identified by expert taxonomists are highly recommended for accurately identifying fish. This is particularly useful in the case of potentially toxic species, as it provides the necessary certainty in identification when assessing and managing the risks associated with their accidental capture and subsequent entry into the food chain, thus ensuring public health [61]. DNA barcoding is a powerful technique used for food safety and prevent commercial fraud. It has been used successfully to detect fillets of E. guttifer imported from African countries into the European Union (Germany and Italy) labelled as African monkfish (Lophius spp.) [62] and also a toxic puffer fish illegally imported which was mislabeled as headless monkfish in Chicago market [63].
According to the inspection of the histological section and the frequency distribution of oocyte diameters, the species could exhibit either asynchronous or group-synchronous oocyte development. The fact that most of the observed oocytes were in advanced vitellogenesis and that the cohort of oocytes smaller than 500 microns was very small leads us to suspect group-synchronous development; however, because hydrated oocytes were not observed and the sample quality was insufficient to detect post-ovulatory follicles, we cannot rule out that eggs have already been released and that, therefore, the species has asynchronous development. For the same reason, it is not possible to determine whether spawning is partial or total. Previous studies in other genera of the family Tetraodontidae report that pufferfishes have asynchronous oocyte development with partial spawning [19,20,22], so it is expected that E. gutiffer follows the same reproductive strategy.
The female analyzed was in the spawning-capable phase according to Lowerre-Barbieri et al. [64]. Although there is no information on the spawning season of this species, reproductive studies of other genera within the family Tetraodontidae show a wide variability in patterns depending on species and location, ranging from a protracted spawning season in S. nephelus [19] and Lagocephalus lunaris (Bloch & Schneider, 1801) [65], to distinct seasonal peaks, e.g., spring for Marilyna pleurosticta (Günther, 1872) [66], spring/summer for C. travancoricus [20], summer for Torquigener flavimaculosus Hardy & Randall, 1983 [67] or autumn/winter for Tetractenos hamiltoni (Richardson, 1846) [66] and Sphoeroides testudineus (Linnaeus, 1758) [68].
The distribution of oocytes was not homogeneous within the ovary, which may be due either to a progressive development of oocytes along the ovary or to the fact that oocytes do not hydrate or are not released homogeneously throughout the ovary, but rather by zones. This second hypothesis would imply partial spawning. In any case, it is recommended that several subsamples be taken from different regions of the ovary in future fecundity studies. Likewise, to properly assign maturity status based on histology, it is recommended to analyze sections from the anterior, central, and posterior regions of the ovary.
The potential fecundity estimated in the present study was 1,335,584 oocytes, and relative fecundity reached 533.4 oocytes per gram of female gutted weight, although it could be higher, since we were unable to confirm whether the female had already released at least one batch. No previous references on fecundity were found for this species; however, there are data on partial fecundity of S. nephelus ranging between 59,087 and 367,022 hydrated oocytes (mean 176,456) [19], and fecundity data (without specification of type) for L. lunaris ranging between 103,355 and 298,795 oocytes [65]. In both cases, the values are one order of magnitude lower than those estimated in the present study, likely because both refer to partial fecundity.
The previous record of E. gutttifer in Galician waters, a male specimen caught in January, was also in spawning capable phase [11]. The occurrence of specimens both in maturation and/or mature would testify the presence of a stable population with active reproductive individuals [69].
Due to their mainly tropical distribution and non-commercial nature of, the species composition of tetraodontiform fishes has historically been poorly understood in Spain. Only three species were initially recorded by Cornide (1734-1803) and Cabrera (1763-1827). Mola mola (Linnaeus, 1758) was recorded by Cornide [70] and Cabrera [71] in the NOR and SUD demarcations respectively, while Diodon hystrix Linnaeus, 1758 and Balistes capriscus Gmelin, 1789 were only recorded in the SUD [71]. Subsequently, E. guttifer [72], Lagocephalus lagocephalus (Linnaeus, 1758) [73] and Ranzania laevis (Pennant, 1776) [74] were added to the list, bringing the total number of species to six by the mid-twentieth century. Since then, this figure has increased considerably, reaching 21 species in the latest update [44].
Our review increases the total number of species to 26 (an 18% increase) and the total number of new demarcation records to 17 (a 36% increase) compared to Báez et al. [44]. Five new species were recorded: C. pullus [75], M. alexandrini [76], C. mauretanicus, C. macrocerus and M. niger [77].
CAN have the highest number of species of any demarcation (22 species), but it also experienced the greatest increase, with five new species being recorded. The Canary Islands’ geographical location — at a lower latitude than the Iberian Peninsula and close to the African coast, where a similar number of tetraodontiform species are found — is probably the reason for their greater species richness. Along the Iberian coast, there are more species per demarcation in the south (SUD and ESAL, with 13 and 12 species respectively) than in the north (NOR and LEBA, with 9 and 8 species respectively). This suggests a general gradient in species abundance from south to north, with fewer species present at higher latitudes and lower temperatures. This is in accordance with the tropical nature of this order.
Although an increase in the number of known species is a normal process in ichthyological knowledge, climate change and warming waters also appear to have played an important role in increasing Spanish fish biodiversity in recent decades [44,78,79]. Global warming can cause fish to adapt or migrate in search of optimal thermal conditions, resulting in changes to their distribution over time and space, typically northward [80]. Consequently, the tropicalization of temperate marine ecosystems is evident, a phenomenon that has been well documented in different Atlantic and Mediterranean Spanish regions [77,78,81,82].
Tetraodontiform species occur mostly in tropical and subtropical waters. Therefore, the new occurrence of species of this order in higher latitudes provides further evidence on the evolution of this process. In the pioneering works about the tropicalization of the north Atlantic waters it is mentioned S. pachygaster as one of the species which has spread more rapidly northward [83,84]. The evolution of the Atlantic French fauna of Tetraodontiformes was also related to the global warming [85]. In Spanish waters, records of Lagocephalus laevigatus (Linnaeus, 1766) [86] and E. guttifer ([11]; this study) in NOR, and C. pullus [75] and C. macrocerus [77] in CAN, represent a new northern limit of distribution for these species in the East Atlantic, which also supports the tropicalization hypothesis.
Another record of biogeographical interest is the westernmost occurrence of the Lessepsian L. sceleratus in the Mediterranean Sea, at the Strait of Gibraltar [46]. This Indo-Pacific species was introduced to the Mediterranean via the Suez Canal in 2003, and has since spread across the entire basin. As climate models predict, global warming is weakening the climatic barriers that have historically prevented Lessepsian species from expanding further and moving into the Atlantic Ocean in the coming years [88].
Only two species of the list, Balistes punctatus Gmelin, 1789 and C. pullus from CAN, have been included in the national reference inventories of non-native marine species [89]. Both species have been classified as “crypto-expansive species” by these authors, defined as species with some evidence of their non-native status, but uncertain due to an unclear mode of introduction from their native range (i.e., natural range expansion versus human-mediated expansion). However, following these criteria, the authors omit many other species, such as the aforementioned E. guttifer and L. laevigatus from the NOR demarcation. In fact, according to this criterion, many tetraodontiforms recorded in Spain should be added to this list.
In terms of vulnerability status, only C. reticulatus is listed as vulnerable in the Spanish Catalogue of Threatened Species [90]. However, the European IUCN Red List assessment does not include any species in the Vulnerable category, which is consistent with a previous report [48]. These authors only assessed tetraodontiform species in the LC or DD categories, the latter often due to a small number of known specimens and taxonomic uncertainty [48]. The last IUCN assessment of the species was conducted in the 2010s. Since then, new taxonomic data and regional changes in abundance have prompted a re-evaluation of the IUCN assessment. In fact, although LC remains the dominant category, an increase in the percentage of threatened species has been observed in recent reviews [91,92]. About 8% of tetraodontids are considered threatened or Near Threatened [91] and three of the five species in the Molidae family changed its status from LC to VU [93].
There is clear evidence that many Tetraodontiformes will benefit from global warming and other human-induced environmental changes, expanding their distribution range, which could have severe socio-economic and ecological impacts. Of the species listed, L. sceleratus has been considered the worst invasive species in the Mediterranean Sea due to its potent neurotoxin (TTXs), its impact on marine biodiversity, and the increased costs and labor it entails for fishermen [94].
Several studies worldwide have reported the presence of either TTXs or PSTs in certain tetraodontiform species that inhabit Spanish waters. However, the information available on the potential presence of toxins in the species included here is still scarce, with some fish species only evaluated for either TTXs or PSTs. Moreover, for 16 of the species evaluated we could not find any study on the analyses of TTXs or PSTs. While Rambla-Alegre et al. [51] reported the first evidence of high amounts of TTXs in a L. sceleratus specimens caught in the Spanish Mediterranean coast, they did not find TTXs in S. pachygaster or L. lagocephalus specimens from the same area. However, Alkassar et al. [53], detected PSTs in L. lagocephalus from the same coast. With regard to Portugal, which shares waters with Spain, TTXs have already been reported in S. marmoratus [49,50], and PSTs were found in L. lagocephalus [50]. The presence of some strong neurotoxins as TTXs or PSTs in pufferfish from the Spanish coast combined with the scarcity of data found point to the need to extend the studies to more Spanish tetraodontiform species.
The arrival of E. guttifer to Atlantic European waters also showed a high new parasitic nematode load [95]. The introduction of parasite species into a new marine environment could negatively affect native fauna because these have not been evolutionary time to reach an equilibrium relationship [96].
Species belonging to the order Tetraodontiformes can be toxic if consumed. However, the number and abundance of toxic species in Spanish waters is relatively low and there is neither a target fishery nor a traditional culinary history, and the current European food legislation prohibits their commercialization [97]. Therefore, the probability of potential intoxications is currently low. However, under the present scenario of climate change and tropicalization process, this risk is growing. Knowing the composition, geographical distribution and potential toxicity of species is particularly important for early detection of toxic fish species and alerting the population on potential risks. This way, food safety issues with potential severe consequences resulting from accidental consumption, can be prevented.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1.
Author Contributions
Conceptualization, R.B.; methodology, R.B., B.A., R.D.P, B.B.G. and A.D.C.; formal analysis, R.B., B.A., R.D.P, B.B.G. and A.D.C.; investigation, R.B., A.V., M.G.C., A.D.C.; resources, R.B., C.G.S, R.D.P and A.D.C; writing—original draft preparation, R.B., B.A., R.D.P, B.B.G. and A.D.C.; writing—review and editing, R.B., B.A., R.D.P, B.B.G. and A.D.C.; visualization, R.B.; supervision, R.B. and B.A.; project administration, R.B.; funding acquisition, R.D.P. and A.D.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The sequences generated and used in the current study are available in the BOLD systems (https://www.boldsystems.org/, accessed on 25 November 2025) and GenBank (https://www.ncbi.nlm.nih.gov/genbank/, accessed on 25 November 2025) repositories. The specimens used in this study for a taxonomic purpose is deposited in the fish collection of the Museo de Historia Natural, Universidade de Santiago de Compostela (MHNUSC) in Santiago de Compostela, Spain.
Acknowledgments
We would like to thank the crew of the vessel “A Entallada” and the Combarro fishermen’s association for providing the specimen and data of E. guttifer. We are also grateful to the crew of vessel “Dous mil” and the Bueu fishermen’s association for providing the specimen and data of S. pachygaster. We also want to thank to the assistance of the Aquarium Finisterrae staff that helped us manage some of the specimens examined.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Near, T.J.; Thacker, C.E. Phylogenetic classification of living and fossil ray-finned fishes (Actinopterygii). Bull. Peabody Mus. Nat. Hist. 2024, 65, 3–302. [Google Scholar] [CrossRef]
- Matsuura, K. Taxonomy and systematics of tetraodontiform fishes: a review focusing primarily on progress in the period from 1980 to 2014. Ichthyol. Res. 2015, 62, 72–113. [Google Scholar] [CrossRef]
- Fricke, R.; Eschmeyer, W.N.; Fong, J.D. Eschmeyer’s catalog of fishes: genera/species by family/subfamily. Electronic version. http://researcharchive.calacademy.org/research/ichthyology/catalog/SpeciesByFamily.asp (accessed on 12 December 2025).
- Neafsey, D.E.; Palumbi, S.R. Genome size evolution in pufferfish: a comparative analysis of diodontid and tetraodontid pufferfish genomes. Genome Res. 2003, 13, 821–830. [Google Scholar] [CrossRef]
- Noguchi, T.; Ebesu, J.S.M. Puffer poisoning: epidemiology and treatment. J. Toxicol. Toxin Rev. 2001, 20, 1–10. [Google Scholar] [CrossRef]
- Sabatino, L.; Esposito, G.; Meloni, D. Occurrence of silver-cheeked toadfish Lagocephalus sceleratus (Tetraodontiformes: Tetraodontidae) and tetrodotoxins in the Mediterranean Sea: Implications for the Italian fishing sector? Reg. Stud. Mar. Sci. 2024, 75, 103562. [Google Scholar] [CrossRef]
- Liu, K.; Sun, H.; Zhao, X.; Wang, C.; An, C.; Li, A.; Liu, S.; Zhuang, Z. DNA barcoding, identification, and validation of the pufferfish (Order: Tetraodontiformes) in China coastal waters. Ecol. Evol. 2024, 14, e10944. [Google Scholar] [CrossRef]
- Santini, F.; Sorenson, L.; Alfaro, M.E. A new phylogeny of tetraodontiform fishes (Tetraodontiformes, Acanthomorpha) based on 22 loci. Mol. Phylogenet. Evol. 2013, 69, 177–187. [Google Scholar] [CrossRef]
- Masuda, Y.; Takeoka, S.; Tabeta, O.; Dotsa, Y. Genetic differences between two newly described swellfishes of the genus Lagocephalus from southern Kyushu, Japan. Nippon Suisan Gakk 1987, 53, 767–772. [Google Scholar] [CrossRef]
- Turan, C.; Gürlek, M.; Ergüden, D.; Uyan, A.; Karan, S.; Doğdu, S.A. Assessing DNA barcodes for identification of pufferfish species (Tetraodontidae) in Turkish marine waters. Nat. Eng. Sci. 2017, 2, 48–59. [Google Scholar] [CrossRef]
- Bañón, R.; Alonso-Fernández, A.; Barros-García, D.; Rios, M.; de Carlos, A. Geographic range expansion of Ephippion guttifer (Tetraodontidae) in the northeastern Atlantic. J. Fish Biol. 2018, 93, 733–737. [Google Scholar] [CrossRef]
- Virgili, R.; Fasciglione, P.; Tiralongo, F.; Pannone, R.; Palomba, M.; Santoro, M.; De Luca, P.; Crocetta, F. Interdisciplinary approaches at early invasion stages maximise the evidence on human-induced and climate-driven fish dispersal. Hydrobiologia 2025, 852, 2241–2261. [Google Scholar] [CrossRef]
- Essl, F.; Dullinger, S.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Katsanevakis, S.; Kühn, I.; Lenzner, B.; Pauchard, A.; Pysek, P.; Rabitsch, W.; Richardson, D.M.; Seebens, H.; Van Kleunen, M.; Van Der Putten, W.H.; Vila, M.; Bacher, S. A conceptual framework for range-expanding species that track human-induced environmental change. BioScience 2019, 69, 908–919. [Google Scholar] [CrossRef]
- Ulman, A.; Abd Rabou, A.F.N.; Al Mabruk, S.; Bariche, M.; Bilecenoğlu, M.; Demirel, N.; Galil, B.S.; Hüseyinoglu, M.F.; Jimenez, C.; Hadjioannou, L.; Kosker, A.R.; Peristeraki, P.; Saad, A.; Samaha, Z.; Stoumboudi, M.Th.; Temraz, T.A.; Karachle, P.K. Assessment of human health impacts from invasive pufferfish (attacks, poisonings and fatalities) across the Eastern Mediterranean. Biology 2024, 13, 208. [Google Scholar] [CrossRef] [PubMed]
- Kawase, H.; Okata, Y.; Ito, K. Role of huge geometric circular structures in the reproduction of a marine pufferfish. Sci. Rep. 2013, 3, 2106. [Google Scholar] [CrossRef]
- Kawase, H.; Okata, Y.; Ito, K. Spawning behavior and paternal egg care in a circular structure constructed by pufferfish, Torquigener albomaculosus (Pisces: Tetraodontidae). Bull. Mar. Sci. 2014, 91, 33–43. [Google Scholar] [CrossRef]
- Sánchez-Cárdenas, R.; Arellano-Martínez, M.; Valdez-Pineda, M.C.; Morán-Angulo, R.E.; Ceballos-Vázquez, B.P. Reproductive cycle and sexual maturity of Sphoeroides annulatus (Jenyns, 1842) (Tetraodontiformes, Tetraodontidae) from the coast of Mazatlan, Sinaloa, Mexico. J. Appl. Ichthyol. 2011, 27, 1190–1196. [Google Scholar] [CrossRef]
- Valdez-Pineda, M.; Morán-Angulo, R.; Voltolina, D.; Castillo-Vargasmachuca, S. Population structure and reproductive aspects of puffer fish Sphoeroides annulatus (Jenyns, 1842) (Osteichthyes: Tetraodontidae), landed in Teacapán, Sinaloa, Mexico. Lat. Am. J. Aquat. Res. 2017, 42, 121–126. [Google Scholar] [CrossRef]
- Peniche-Pérez, J.; González-Salas, C.; Villegas-Hernández, H.; Díaz-Gamboa, R.; Aguilar-Perera, A.; Guillen-Hernández, S.; Poot-López, G. Reproductive biology of the southern pufferfish, Sphoeroides nephelus (Actinopterygii: Tetraodontiformes: Tetraodontidae), in the northern coast off the Yucatan Peninsula, Mexico. Acta Ichthyol. Pisc 2019, 49, 133–146. [Google Scholar] [CrossRef]
- Anupama, K.M.; Hari Sankar, H.S.; Rithin Raj, M.; Harikrishnan, M. Reproductive biology of malabar pufferfish Carinotetraodon travancoricus (Tetraodontidae). J. Ichthyol. 2019, 59, 545–554. [Google Scholar] [CrossRef]
- Momota, K.; Doi, H.; Obata, H.; Sakai, H. Tandem spawning and development in the African freshwater pufferfish Tetraodon schoutedeni. Zoo Biol. 2023, 42, 357–363. [Google Scholar] [CrossRef]
- Chandana, B.; Sanal, A.; Raghavan, R.; Varghese, B. Embryonic development of the world’s smallest puffer fish, Carinotetraodon travancoricus – a threatened freshwater fish of the Western Ghats Biodiversity Hotspot. Zygote 2024, 32, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Sonoyama, T.; Yamada, M.; Gao, W.; Tatsuno, R.; Takatani, T.; Arakawa, O. Co-occurrence of tetrodotoxin and saxitoxins and their intra-body distribution in the pufferfish Canthigaster valentini. Toxins 2020, 12, 436. [Google Scholar] [CrossRef]
- Barrientos, R.G.; Hernández-Mora, G.; Alegre, F.; Field, T.; Flewelling, L.; McGrath, S.; Deeds, J.; Chacón, Y.S.; Rojas Arrieta, K.; Vargas, E.C.; Artavia, K.B.; Stacy, B.A. Saxitoxin poisoning in green turtles (Chelonia mydas) linked to scavenging on mass mortality of Caribbean sharpnose puffer fish (Canthigaster rostrata-Tetraodontidae). Front. Vet. Sci. 2019, 6, 466. [Google Scholar] [CrossRef]
- Rey, V.; Rossignoli, A. E.; Rodríguez, F.; Blanco, J.; Garrido, S.; Ben-Gigirey, B. Evaluation of the prevalence of Paralytic Shellfish Toxins in non-traditional vectors and potential health risks associated to their consumption. Food Control 2025, 176, 111351. [Google Scholar] [CrossRef]
- Ben-Gigirey, B.; Rossignoli, A.E.; Riobó, P.; Rodríguez, F. First report of paralytic shellfish toxins in marine invertebrates and fish in Spain. Toxins 2020, 12, 723. [Google Scholar] [CrossRef]
- Ben-Gigirey, B.; Soliño, L.; Bravo, I.; Rodríguez, F.; Casero, M.V. Paralytic and amnesic shellfish toxins impacts on seabirds, analyses and management. Toxins 2021, 13, 454. [Google Scholar] [CrossRef]
- Katikou, P.; Gokbulut, C.; Kosker, A.R.; Campàs, M.; Ozogul, F. An updated review of tetrodotoxin and its peculiarities. Mar. Drugs 2022, 20, 47. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.; Onuki, K.; Arakawa, O. Tetrodotoxin poisoning due to pufferfish and gastropods, and their intoxication mechanism. Int. Sch. Res. Notices 2011, 2011, 276939. [Google Scholar] [CrossRef] [PubMed]
- Martínez, A.; Garrido-Maestu, A.; Ben-Gigirey, B.; Chapela, M.J.; González, V.; Vieites, J.M.; Cabado, A.G. Marine Biotoxins. In Emerging Marine Biotoxins; Kim, S.K., Ed.; Springer Handbook of Marine Biotechnology; Springer Handbooks. Springer: Berlin, Heidelberg, Germany, 2015; pp. 869–904. [Google Scholar] [CrossRef]
- Almón, B.; Bañón, R. Research trends and state of knowledge of decapod crustaceans in Spain: A bibliometric analysis. Ecologies 2025, 6, 63. [Google Scholar] [CrossRef]
- Shipp, R.L. The pufferfishes (Tetraodontidae) of the Atlantic Ocean. GCRL Publications 1974, 41, 1–162. [Google Scholar]
- Ivanova, N.V.; Zemlak, T.S.; Hanner, R.H.; Hebert, P.D.N. Universal primer cocktails for fish DNA barcoding. Mol. Ecol. Notes 2007, 7, 544–548. [Google Scholar] [CrossRef]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. PNAS 1977, 74, 5463–5467. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Bagenal, T.B.; Braum, E. Eggs and early life history. In Methods for Assessment of Fish Production in Fresh Waters; Bagenal, T.B., Ed.; Blackwell Scientific: Oxford, 1978; pp. 165–201. [Google Scholar]
- Witthames, P.R.; Thorsen, A.; Murua, H.; Saborido-Rey, F.; Greenwood, L.N.; Dominguez, R.; Korta, M.; Kjesbu, O.S. Advances in methods for determining fecundity: application of the new methods to some marine fishes. Fish. Bull. 2009, 107, 148–164. [Google Scholar]
- Lowerre-Barbieri, S.K.; Barbieri, L.R. A new method of oocyte separation and preservation for fish reproduction studies. Fish. Bull. 1993, 91, 159–164. [Google Scholar]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; Tinevez, J.Y.; White, D.J.; Hartenstein, V.; Eliceiri, K.; Tomancak, P.; Cardona, A. Fiji: an open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A language and environment for statistical computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; https://www.R-project.org/.
- Posit; team. RStudio: Integrated Development Environment for R. Posit Software, PBC; Boston, MA, 2025; http://www.posit.co/.
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer-Verlag New York, 2016; https://ggplot2.tidyverse.org[R1ISBN 978-3-319-24277-4.
- GBIF Occurrence Download. Available online: https://www.gbif.org/es/ (accessed on 12 November 2025). [CrossRef]
- Báez, C.; Rodríguez-Cabello, C.; Bañón, R.; Brito, A.; Falcón, J.M.; Maño, T.; Baro, J.; Macías, D.; Meléndez, M.J.; Camiñas, J.A.; Arias-García, A.; Gil, J.; Farias, C.; Artexe, I.; Sánchez, F. Updating the national checklist of marine fishes in Spanish waters: An approach to priority hotspots and lessons for conservation. Mediterr. Mar. Sci. 2019, 20, 260–270. [Google Scholar] [CrossRef]
- Guallart, J.; Vicent, J.J. First record of the unicorn leatherjacket Aluterus monoceros (Pisces: Monacanthidae) from the Mediterranean Sea. Mar. Biodivers. Rec. 2009, 2, e103. [Google Scholar] [CrossRef]
- Azzurro, E.; Bariche, M.; Cerri, J.; Garrabou, J. The long reach of the Suez Canal: Lagocephalus sceleratus (Gmelin, 1789) an unwanted Indo-Pacific pest at the Atlantic gates. BioInvasions Rec. 2020, 9, 204–208. [Google Scholar] [CrossRef]
- The IUCN Red List of Threatened Species. 1964 - onwards. Version 2025-1. Available online: https://www.iucnredlist.org (accessed on 23 November 2025).
- Nieto, A.; Ralph, G.M.; Comeros-Raynal, M.T.; Kemp, J.; García-Criado, M.; Allen, D.J.; Dulvy, N.K.; Walls, R.H.L.; Russell, B.; Pollard, D.; et al. European Red List of marine fishes; Publications Office of the European Union: Luxembourg, 2015; pp. 1–81. [Google Scholar]
- Silva, M.; Rodríguez, I.; Barreiro, A.; Kaufmann, M.; Neto, A.I.; Hassouani, M.; Sabour, B.; Alfonso, A.; Botana, L.M.; Vasconcelos, V. Tetrodotoxins occurrence in non-traditional vectors of the north atlantic waters (Portuguese maritime territory, and morocco coast). Toxins 2019, 11, 306. [Google Scholar] [CrossRef]
- Pinto, E.P.; Rodrigues, S.M.; Gouveia, N.; Timóteo, V.; Costa, P.R. Tetrodotoxin and saxitoxin in two native species of puffer fish, Sphoeroides marmoratus and Lagocephalus lagocephalus, from NE Atlantic Ocean (Madeira Island, Portugal). Mar. Environ. Res. 2019, 151, 104780. [Google Scholar] [CrossRef]
- Rambla-Alegre, M.; Reverté, L.; Del Río, V.; de la Iglesia, P.; Palacios, O.; Flores, C.; Caixach, J.; Campbell, K.; Elliott, C.T.; Izquierdo-Muñoz, A.; Campàsa, M.; Diogène, J. Evaluation of tetrodotoxins in puffer fish caught along the Mediterranean coast of Spain. Toxin profile of Lagocephalus sceleratus. Environ. Res. 2017, 158, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Malloggi, C.; Rizzo, B.; Giusti, A.; Guardone, L.; Gasperetti, L.; Dall’Ara, S.; Armani, A. First toxicological analysis of the pufferfish Sphoeroides pachygaster collected in Italian waters (Strait of Sicily): Role of citizens science in monitoring toxic marine species. Animals 2023, 13, 1873. [Google Scholar] [CrossRef] [PubMed]
- Alkassar, M.; Tudó, À.; Rambla-Alegre, M.; Ferreres, L.; Diogène, J.; Sureda, F.X.; Campàs, M. First record of paralytic shellfish toxins in marine pufferfish from the Spanish Mediterranean coast using cell-based assay, automated patch clamp and HPLC-FLD. Chemosphere 2024, 364, 143053. [Google Scholar] [CrossRef]
- Oliveira, J.S.; Pires Junior, O.R.; Morales, R.A.V. Toxicity of Puffer fish-two species (Lagocephalus laevigatus, Linaeus 1766 and Sphoeroides spengleri, Bloch 1785) from the southeren Brazilian coast. J. Venom. Anim. Toxins Incl. Trop. 2003, 9, 76–82. [Google Scholar] [CrossRef]
- Rodríguez, P.; Alfonso, A.; Otero, P.; Katikou, P.; Georgantelis, D.; Botana, L.M. Liquid chromatography–mass spectrometry method to detect Tetrodotoxin and Its analogues in the puffer fish Lagocephalus sceleratus (Gmelin, 1789) from European waters. Food Chem. 2012, 132, 1103–1111. [Google Scholar] [CrossRef]
- Saito, T.; Noguchi, T.; Shida, Y.; Abe, T.; Hashimoto, K. Screening of Tetrodotoxin and Its Derivatives in Puffer-related Species. Nippon Suisan Gakk 1991, 57, 1573–1577. [Google Scholar] [CrossRef]
- Yu, C.; Yu, P.H. A preliminary study of puffer fishes and their toxins found in Hong Kong waters. J. Food Hyg. Soc. Japan 1997, 7(38), 460–463. [Google Scholar] [CrossRef]
- Tamele, I.J.; Timba, I.; Costa, P.R.; Vasconcelos, V. Tetrodotoxin and analogues in two local pufferfish species from Inhaca Island–South of Mozambique: First report in the Mozambican coast. Toxicon 2022, 216, 88–91. [Google Scholar] [CrossRef]
- Elshaer, F.M. Comparative histopathological studies on kidney and liver of rat treated by tetrodotoxin (TTX) extracted from gonads and muscles of porcupine fish species. Int. J. Fish. Aquat. Stud. 2016, 4, 355–360. [Google Scholar]
- Baptista, M.; Braga, A.C.; Rosa, R.; Costa, P.R. Does ocean sunfish Mola spp. (Tetraodontiformes: Molidae) represent a risk for tetrodotoxin poisoning in the Portuguese coast? Mar. Drugs 2022, 20, 594. [Google Scholar] [CrossRef]
- Giusti, A.; Guarducci, M.; Stern, N.; Davidovich, N.; Golani, D.; Armani, A. The importance of distinguishing pufferfish species (Lagocephalus spp.) in the Mediterranean Sea for ensuring public health: Evaluation of the genetic databases reliability in supporting species identification. Fish. Res. 2019, 210, 14–21. [Google Scholar] [CrossRef]
- ICES. Report of the Working Group on the Application of Genetics in Fisheries and Mariculture (WGAGFM). ICES WGAGFM Report, Reykjavik, 7–9 May 2013, Iceland. ICES, Copenhagen, Denmark, 2013; pp. 1–52. [Google Scholar]
- Muhammad Tahir, H.; Akhtar, S. Services of DNA barcoding in different fields. Mitochondrial DNA A 2016, 27, 4463–4474. [Google Scholar] [CrossRef]
- Lowerre-Barbieri, S.K.; Brown-Peterson, N.J.; Wyanski, D.M.; Moncrief-Cox, H.E.; Kolmos, K.J.; Menendez, H.S.; Barnett, B.K.; Friess, C. A unified framework and terminology for reproductive traits integral to understanding fish population productivity. Mar. coast. fish. 2023, 15, e210276. [Google Scholar] [CrossRef]
- Sirisha, R.; Imandi; Rao, P.Y. Maturation and spawning of green – rough – back pufferfish, Lagocephalus lunaris. NVBS 2021, 4, 8–18. [Google Scholar] [CrossRef]
- Mat Piah, R.; Bucher, D.J. Reproductive Biology of Estuarine Pufferfish, Marilyna pleurosticta and Tetractenos hamiltoni (Teleostei: Tetraodontidae) in Northern New South Wales: Implications for Biomonitoring. Proc. Linn. Soc. N.S.W 2014, 136, 219–229. [Google Scholar]
- Ramadan, A.M.; El-Halfawy, M.M. Reproductive biology of the yellow-spotted puffer Torquigener flavimaculosus (Osteichthyes: Tetraodontidae) from Gulf of Suez, Egypt. EJABF 2019, 23, 503–511. [Google Scholar] [CrossRef]
- Rocha, C.; Favaro, L.F.; Spach, H.L. Reproductive biology of Sphoeroides testudineus (Linneus) (Pisces, Osteichthyes, Tetraodontidae) of the gamboa do Baguaçu, bay of Paranaguá, State of Paraná, Brazil. Rev. Bras. Zool 2002, 19, 57–63. [Google Scholar] [CrossRef]
- Carbonara, P.; Kolitari, J.; Đurović, M.; Gaudio, P.; Ikica, Z.; Kroqi, G.; Milonbe, N.; Spedicato, M.T. The presence of Tetraodontidae species in the Central Mediterranean: an update from the southern Adriatic Sea. Acta Adriat. 2017, 58, 325–338. [Google Scholar] [CrossRef]
- Cornide, J. Ensayo de una historia de los peces y otras producciones marinas de la costa de Galicia. Edición facsmil; Edicións do Castro: O Castro-Sada, Spain, 1788; pp. 1–263. [Google Scholar]
- Arias, A.M.; Romero, P.; De la Torre, M.; Bañón, R. La ictiología de Andalucía. Catálogo crítico e ilustrado de las especies marinas estudiadas por Antonio Cabrera y Corro (1762-1827), canónigo magistral de la catedral de Cádiz; Editorial UCA, Servicio de Publicaciones de la Universidad de Cádiz: Cádiz, Spain, 2025; pp. 1–636. [Google Scholar]
- Pérez-Arcas, L. Sobre T. guttifer en la costa de Málaga. Anales Soc. Esp. Hist. Nat. 1872, 1, 4–7. [Google Scholar]
- De Buen, F. Fauna ictiológica. Catálogo de los peces ibéricos de la planicie continental, aguas dulces, pelágicos y de los abismos próximos. II parte. Not. y Res. Inst. Esp. Ocean. 1935, 82, 89–148. [Google Scholar]
- Lozano-Rey, L. Peces Fisoclistos: O. Bericiformes, Zeiformes, Perciformes, Escorpeniformes y Balistiformes; Memorias de la Real Academia de Ciencias Exactas, Fisicas y Naturales de Madrid: Madrid, España, 1952; Primera Parte; pp. 1–378. [Google Scholar]
- Triay-Portella, R.; Pajuelo, J.G.; Manen, T.P.; Espino, F.; Ruiz-Díaz, R.; Lorenzo, J.M.; González-Pérez, J.A. New records of non-indigenous fishes (Perciformes and Tetraodontiformes) from the Canary Islands (north-eastern Atlantic). Cybium 2015, 39, 163–174. [Google Scholar] [CrossRef]
- Wirtz, P.; Biscoito, M. The distribution of Mola alexandrini in the subtropical eastern Atlantic, with a note on Mola mola. Bocagiana 2019, 245, 1–6. [Google Scholar]
- Falcón, J.M.; Brito, A.; Herrera, R.; Ayza, O.; Moro, L; Caro, M.B. Peces marinos tropicales exóticos de Canarias; Consejería de Transición Ecológica, Lucha contra el Cambio Climático y Planificación Territorial del Gobierno de Canarias, 2023; pp. 1–247. [Google Scholar]
- Bañón, R.; Conde-Pardo, P.; Álvarez-Salgado, X.A.; de Carlos, A.; Arronte, J.C.; Piedracoba, S. Tropicalization of fish fauna of Galician coastal waters, in the NW Iberian upwelling system. Reg. Stud. Mar. Sci. 2024, 70, 103369. [Google Scholar] [CrossRef]
- González, J.A.; Espino, F.; González-Lorenzo, J.G. Changes in biogeographic patterns of coastal fishes: Indicators of tropicalization in the Canary Islands over the last 40 years. Mar. Environ. Res. 2025, 205, 107002. [Google Scholar] [CrossRef] [PubMed]
- Booth, D.J.; Bond, N.; Macreadie, P. Detecting range shifts among Australian fishes in response to climate change. Mar. Freshw. Res. 2011, 62, 1027–1042. [Google Scholar] [CrossRef]
- de Carvalho-Souza, G.F.; Lobato, C.; Mesa, D.A.; González-Ortegón, E. Ongoing tropicalization of marine fishes: is range expansion and establishment occurring in the Gulf of Cadiz (southernmost Europe)? J. Mar. Biol. Assoc. U.K. 2025, 105, e61. [Google Scholar] [CrossRef]
- Torreblanca, D.; Báez, J.C. Tropicalization of the Mediterranean Sea Reflected in Fish Diversity Changes: A Case Study from Spanish Waters. J. Mar. Sci. Eng. 2025, 13, 1119. [Google Scholar] [CrossRef]
- Quéro, J.C.; Du Buit, M.H.; Vayne, J.J. Les captures de poissons à affinités tropicales le long des côtes Atlantiques Européennes. Ann. Soc. Sci. Nat. Charente-Marit. 1996, 8, 651–673. [Google Scholar]
- Quéro, J.C. Changes in the Euro-Atlantic fish species composition resulting from fishing and ocean warming. Ital. J. Zool. 1998, 65, 493–499. [Google Scholar] [CrossRef]
- Quéro, J.C.; Spitz, J.; Vayne, J.J. Faune française de l’Atlantique. Poissons Tetraodontiformes. Ann. Soc. Sci. Nat. Charente-Marit. 2008, 9, 815–832. [Google Scholar]
- Bañón, R.; Santás, V. First record of Lagocephalus laevigatus (Tetraodontiformes, Tetraodontidae) from Galician waters (northwest Spain), a northernmost occurrence in the north-east Atlantic. J. Fish Biol. 2011, 78, 1574–1578. [Google Scholar] [CrossRef] [PubMed]
- Azzurro, E.; Bariche, M.; Cerri, J.; Garrabou, J. The long reach of the Suez Canal: Lagocephalus sceleratus (Gmelin, 1789) an unwanted Indo-Pacific pest at the Atlantic gates. Bioinvasions Rec. 2020, 9, 204–208. [Google Scholar] [CrossRef]
- Azzurro, E.; D’Amen, M. Climate change paves the way for a new inter-ocean fish interchange. Front. Ecol. Environ 2022, 20, 558–563. [Google Scholar] [CrossRef]
- Png-Gonzalez, L.; Comas-González, R.; Calvo-Manazza, M.; Follana-Berná, G.; Ballesteros, E.; Díaz-Tapia, P.; Falcón, J.M.; García Raso, J.E.; Gofas, S.; González-Porto, M.; López, E.; Ramos-Esplá, A.A.; Velasco, E.; Carbonell, A. Updating the national baseline of non-indigenous species in Spanish marine waters. Diversity 2023, 15, 630. [Google Scholar] [CrossRef]
- Espino, F.; Tuya, F.; del Rosario, A.; Bosch, N.E.; Coca, J.; González-Ramos, A.J.; del Rosario, F.; Otero-Ferrer, F.J.; Moreno, A.C.; Haroun, R. Geographical range extension of the spotfin burrfish, Chilomycterus reticulatus (L. 1758), in the Canary Islands: a response to ocean warming? Diversity 2019, 11, 230. [Google Scholar] [CrossRef]
- Stump, E.; Ralph, G.M.; Comeros-Raynal, M.T.; Matsuura, K.; Carpenter, K.E. Global conservation status of marine pufferfishes (Tetraodontiformes: Tetraodontidae). Glob. Ecol. Conserv. 2018, 14, e00388. [Google Scholar] [CrossRef]
- Eduardo, L.N.; Bertrand, A.; Frédou, T.; Lira, A.S.; Lima, R.S.; Ferreira, B.P.; Menard, F.; Lucena-Frédou, F. Biodiversity, ecology, fisheries, and use and trade of Tetraodontiformes fishes reveal their socio-ecological significance along the tropical Brazilian continental shelf. Aquat. Conserv.: Mar. Freshw. Ecosyst. 2020, 30, 761–774. [Google Scholar] [CrossRef]
- Phillips, N.; Nyegaard, M.; Sawai, E.; Chang, C.T.; Baptista, M.; Thys, T. The ocean sunfishes (family Molidae): Recommendations from the IUCN molidae review panel. Mar. Pol. 2023, 155, 105760. [Google Scholar] [CrossRef]
- Ulman, A.; Harris, H.E.; Doumpas, N.; Deniz Akbora, H.; Al Mabruk, S.A.A.; Azzurro, E.; Bariche, M.; Çiçek, B.A.; Deidun, A.; Demirel, N.; Fogg, A.Q.; Katsavenakis, S.; Kletou, D.; Kleitou, P.; Papadopoulou, A.; Ben Souissi, J.; Hall-Spencer, J.M.; Tiralongo, F.; Yildiz, T. Low pufferfish and lionfish predation in their native and invaded ranges suggests human control mechanisms may be necessary to control their Mediterranean abundances. Front. Mar. Sci. 2021, 8, 670413. [Google Scholar] [CrossRef]
- Rodríguez, H.; Bañón, R.; Ramilo, A. The hidden companion of non-native fishes in north-east Atlantic waters. J. Fish Dis. 2019, 42, 1013–1021. [Google Scholar] [CrossRef] [PubMed]
- Azzurro, E. Unusual occurrences of fish in the Mediterranean Sea: an insight into early detection. In Fish Invasions of the Mediterranean Sea: Change and Renewal; Golani, D., Appelbaum-Golani, B., Eds.; Pensoft Publishers: Sofia-Moscow, 2010; pp. 99–126. [Google Scholar]
- Reglamento (CE) Nº 853/2004 del parlamento europeo y del consejo de 29 de abril de 2004 por el que se establecen normas específicas de higiene de los alimentos de origen animal. Diario Oficial de la Unión Europea L 139/55.
- Nakatani, T.; Shimizu, M.; Yamano, T. The contents and composition of tetrodotoxin and paralytic shellfish poisoning toxins in marine pufferfish Canthigaster rivulata. J. Food Hyg. Soc. Japan 2016, 57, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Guardone, L.; Maneschi, A.; Meucci, V.; Gasperetti, L.; Nucera, D.; Armani, A. A global retrospective study on human cases of tetrodotoxin (TTX) poisoning after seafood consumption. Food Rev. Int. 2020, 36, 645–667. [Google Scholar] [CrossRef]
Figure 1.
Map of the study area showing the different geographical demarcations and the number of species recorded in each of them. Close-up views in red show the detailed location of the new records.
Figure 1.
Map of the study area showing the different geographical demarcations and the number of species recorded in each of them. Close-up views in red show the detailed location of the new records.
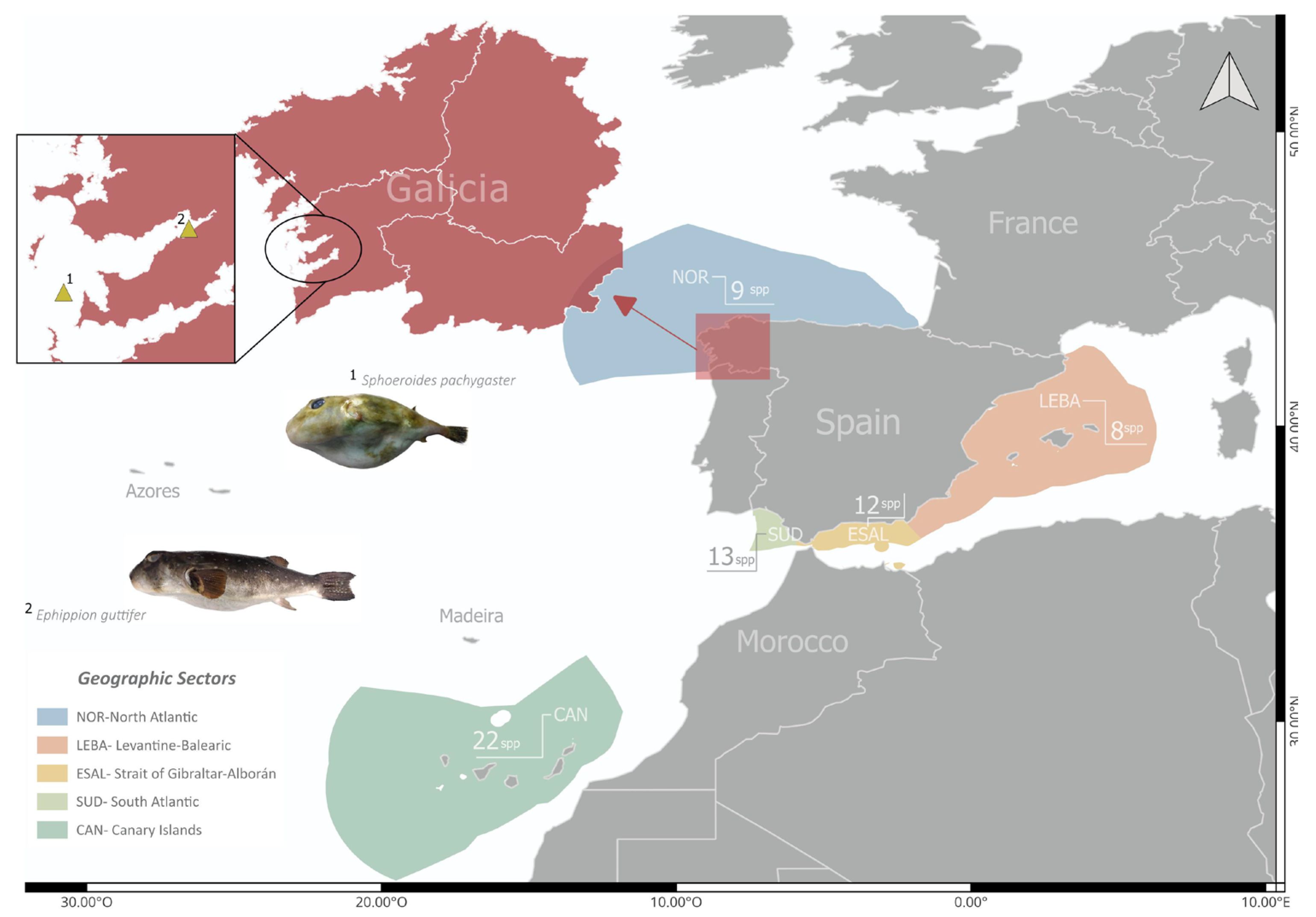
Figure 2.
Lateral (left) and dorsal (middle) views of Sphoeroides pachygaster (A) and Ephippion guttifer (B).
Figure 2.
Lateral (left) and dorsal (middle) views of Sphoeroides pachygaster (A) and Ephippion guttifer (B).

Figure 3.
Neighbor-Joining dendrogram representation of uncorrected p-distances based on the analysis of 652 nucleotides of the cytochrome c oxidase subunit I (COI) gene in two taxa of tetraodontidae. The sequences of E. guttifer and S. pachygaster are accompanied by their respective GenBank accession numbers, with the exception of the specimens covered by this study (in bold), which are listed with their BOLD process IDs. Geographical locations are also included, when available. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test is shown below the branches. The bar indicates the number of nucleotide differences per site.
Figure 3.
Neighbor-Joining dendrogram representation of uncorrected p-distances based on the analysis of 652 nucleotides of the cytochrome c oxidase subunit I (COI) gene in two taxa of tetraodontidae. The sequences of E. guttifer and S. pachygaster are accompanied by their respective GenBank accession numbers, with the exception of the specimens covered by this study (in bold), which are listed with their BOLD process IDs. Geographical locations are also included, when available. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test is shown below the branches. The bar indicates the number of nucleotide differences per site.

Figure 4.
Histological section of the ovary of Ephippion gutiffer. A) General view (2x) of the ovary, B) Detailed view (4x) of oocytes developmental stages and C) Detailed view (10x) of atretic oocyte. D) Melanomacrophage centre. PG: primary growth, CA: cortical alveoli, AV: advanced vitelogenic, AT: atretic, MC: melanomacrophage center.
Figure 4.
Histological section of the ovary of Ephippion gutiffer. A) General view (2x) of the ovary, B) Detailed view (4x) of oocytes developmental stages and C) Detailed view (10x) of atretic oocyte. D) Melanomacrophage centre. PG: primary growth, CA: cortical alveoli, AV: advanced vitelogenic, AT: atretic, MC: melanomacrophage center.
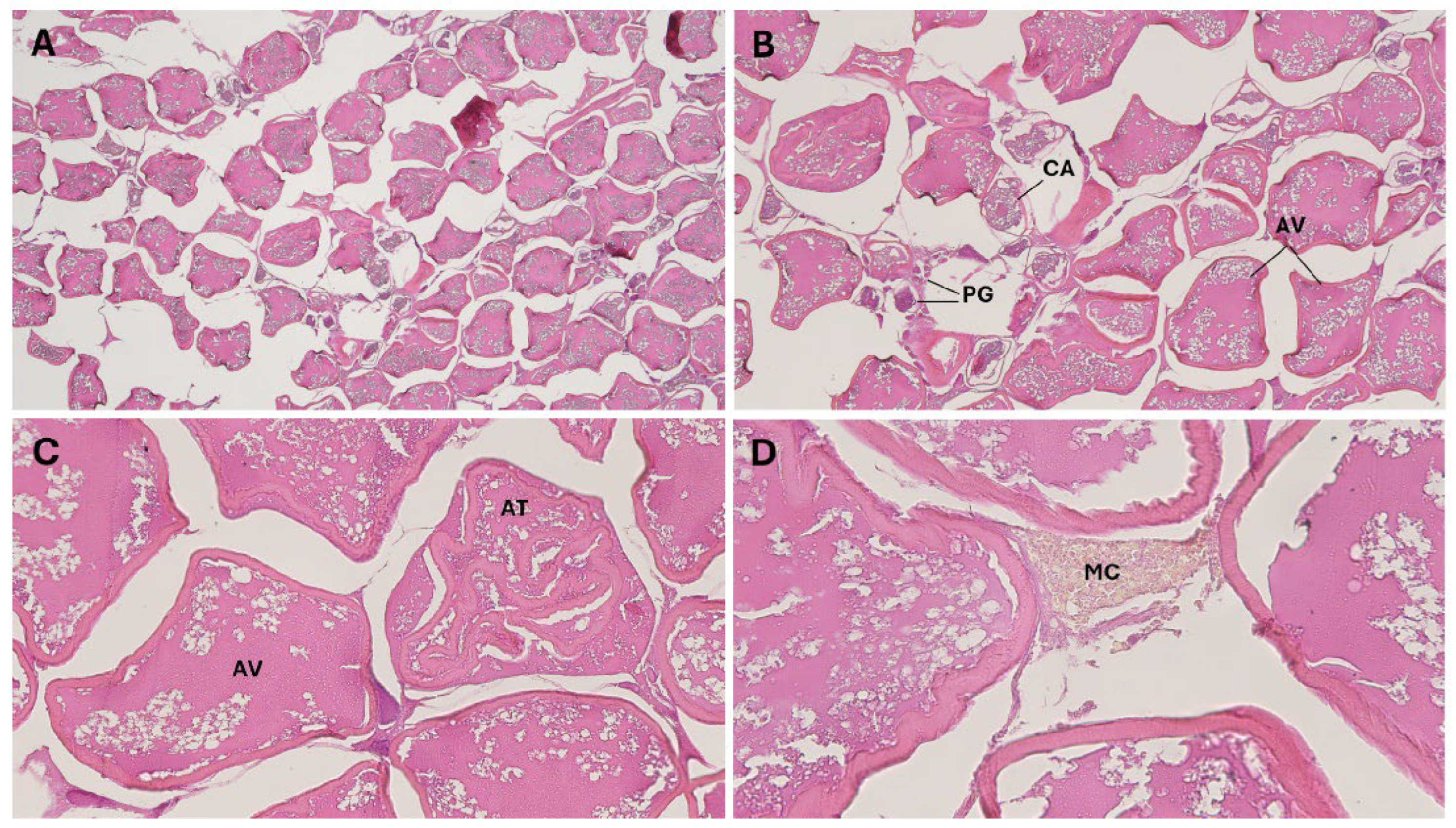
Figure 5.
Density plot of oocyte diameter frequency in each sub-sample: sub1 in red, sub2 in green, sub3 in blue and sub4 in purple (left) and pooling all sub-samples together (right).
Figure 5.
Density plot of oocyte diameter frequency in each sub-sample: sub1 in red, sub2 in green, sub3 in blue and sub4 in purple (left) and pooling all sub-samples together (right).
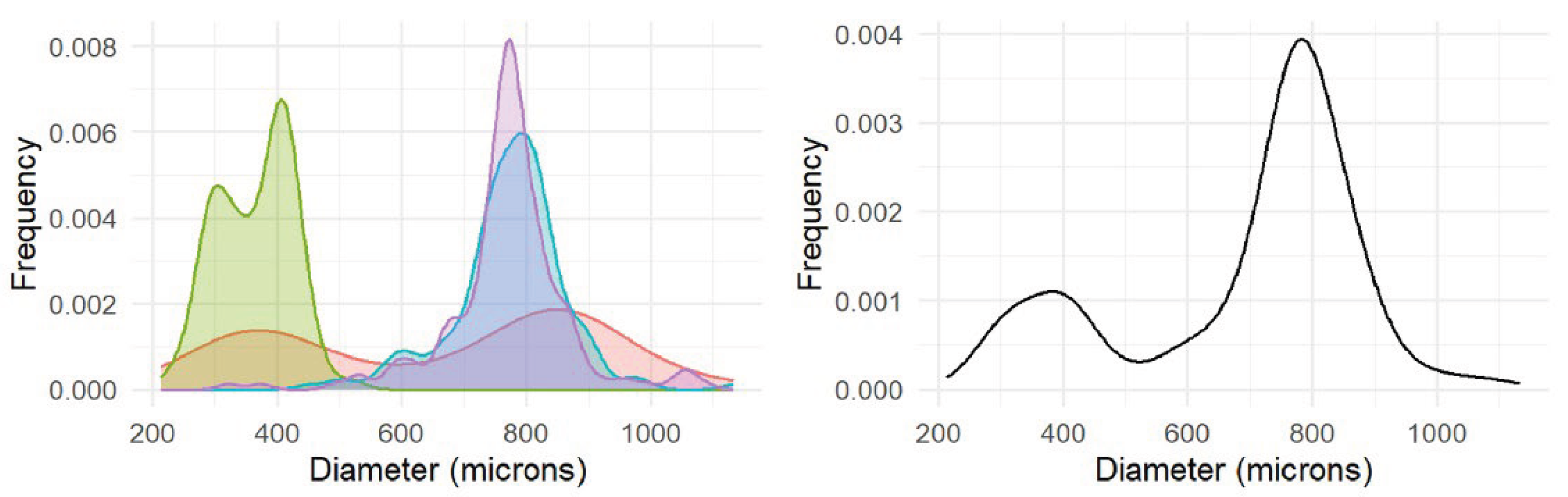

Table 1.
Morphometric and meristic data of Ephippion guttifer and Sphoeroides pachygaster caught in Galician waters.
Table 1.
Morphometric and meristic data of Ephippion guttifer and Sphoeroides pachygaster caught in Galician waters.
|
Ephippion guttifer MHN USC 25103-1 |
Sphoeroides pachygaster MHN USC 25234 |
|
|---|---|---|
| Total length (mm) | 567 | 220 |
| Standard length (mm) | 462 | 187 |
| As % Standard length | ||
| Head length | 29.6 | 28.9 |
| Pre-orbital length | 14 | 19.7 |
| Post-orbital length | 14.9 | 13.5 |
| Horizontal eye diameter | 5.8 | 8.9 |
| Interorbital width | 13.5 | 17.8 |
| Length of gill opening | 10.2 | 9.8 |
| Pre-dorsal-fin length | 68.4 | 75.5 |
| Dorsal-fin base length | 7.2 | 11.3 |
| Pre-anal-fin length | 71.2 | 81.3 |
| Pre-pectoral-fin length | 31.8 | 43.9 |
| Anal-fin base | 5.3 | 3.9 |
| Pectoral-fin length | 14.8 | 15.3 |
| Caudal peduncle depth | 10.2 | 9.8 |
| Body depth | 28.4 | 29.1 |
| Body width | 28.8 | 24.9 |
| Meristic | ||
| Dorsal fin rays | 10 | 8 |
| Anal fin rays | 9 | 8 |
| Pectoral fin rays | 19 | 14 |
| Caudal-fin rays | 10 | 10 |
Table 2.
Weight in grams (SW), number of oocytes (NO) and oocyte density (OD, oocytes/ovary g) of the 4 analyzed ovary sub-samples of Ephippion guttifer.
Table 2.
Weight in grams (SW), number of oocytes (NO) and oocyte density (OD, oocytes/ovary g) of the 4 analyzed ovary sub-samples of Ephippion guttifer.
| SW | NO | OD | |
|---|---|---|---|
| Subsample1 | 0.046 | 87 | 1891 |
| Subsample2 | 0.042 | 85 | 2024 |
| Subsample3 | 0.02 | 197 | 9850 |
| Subsample4 | 0.029 | 194 | 6690 |
Table 3.
Presence (P) of tetraodontiform fishes listed by family and geographical demarcations. The asterisk shows the new records of species and demarcations respect to Báez et al. [44]. NOR: Spanish north coast, SUD: Gulf of Cádiz, ESAL: Alboran Sea, LEBA: Levantine-Balearic, CAN: Canary Islands.
Table 3.
Presence (P) of tetraodontiform fishes listed by family and geographical demarcations. The asterisk shows the new records of species and demarcations respect to Báez et al. [44]. NOR: Spanish north coast, SUD: Gulf of Cádiz, ESAL: Alboran Sea, LEBA: Levantine-Balearic, CAN: Canary Islands.
| Family | Species | NOR | SUD | ESAL | LEBA | CAN |
|---|---|---|---|---|---|---|
| Tetraodontidae | Sphoeroides marmoratus (Lowe, 1838) | P | P | P* | P | |
| Tetraodontidae | Sphoeroides pachygaster (Müller & Troschel, 1848) | P | P | P | P* | P |
| Tetraodontidae | Canthigaster capistrata (Lowe, 1839) | P | P* | P | ||
| Tetraodontidae | Lagocephalus laevigatus (Linnaeus, 1766) | P | P* | |||
| Tetraodontidae | Lagocephalus lagocephalus (Linnaeus, 1758) | P | P | P | P | P |
| Tetraodontidae | Lagocephalus sceleratus (Gmelin, 1789) | P* | P | |||
| Tetraodontidae | Ephippion guttifer (Bennett, 1831) | P* | P* | P | ||
| Monacanthidae | Aluterus monoceros (Linnaeus, 1758) | P | P | P* | P | |
| Monacanthidae | Aluterus scriptus (Osbeck, 1765) | P | ||||
| Monacanthidae | Cantherhines pullus (Ranzani, 1842)* | P* | ||||
| Monacanthidae | Cantherhines macrocerus (Hollard, 1853)* | P* | ||||
| Monacanthidae | Stephanolepis hispida (Linnaeus, 1766) | P | ||||
| Diodontidae | Chilomycterus reticulatus (Linnaeus, 1758) | P | ||||
| Diodontidae | Chilomycterus mauretanicus (Le Danois, 1954)* | P* | P* | |||
| Diodontidae | Diodon eydouxii Brisout de Barneville, 1846 | P | P | P | ||
| Diodontidae | Diodon holocanthus Linnaeus, 1758 | P* | P | |||
| Diodontidae | Diodon hystrix Linnaeus, 1758 | P | P* | P | ||
| Molidae | Masturus lanceolatus (Liénard, 1840) | P | ||||
| Molidae | Mola mola (Linnaeus, 1758) | P | P | P | P | P |
| Molidae | Mola alexandrine (Ranzani 1834)* | P* | ||||
| Molidae | Ranzania laevis (Pennant, 1776) | P* | P | P | P | P |
| Balistidae | Canthidermis sufflamen (Mitchill, 1815) | P | ||||
| Balistidae | Canthidermis maculata (Bloch, 1786) | P | ||||
| Balistidae | Melichthys niger (Bloch, 1786)* | P* | ||||
| Balistidae | Balistes capriscus Gmelin, 1789 | P | P | P | P | P |
| Balistidae | Balistes punctatus Gmelin, 1789 | P |
Table 4.
IUCN Red List status at global and European level of Spanish tetraodontiform species. LC: least concern; DD: data deficient; VU: Vulnerable; NA: not applicable; NE: not evaluated.
Table 4.
IUCN Red List status at global and European level of Spanish tetraodontiform species. LC: least concern; DD: data deficient; VU: Vulnerable; NA: not applicable; NE: not evaluated.
| Species | IUCN Global | IUCN Europe |
|---|---|---|
| Sphoeroides marmoratus | LC | LC |
| Sphoeroides pachygaster | LC | LC |
| Canthigaster capistrata | LC | LC |
| Lagocephalus laevigatus | LC | NE |
| Lagocephalus lagocephalus | LC | LC |
| Lagocephalus sceleratus | LC | NE |
| Ephippion guttifer | LC | DD |
| Aluterus monoceros | LC | DD |
| Aluterus scriptus | LC | DD |
| Cantherhines pullus | LC | NE |
| Cantherhines macrocerus | LC | NE |
| Stephanolepis hispida | LC | LC |
| Chilomycterus reticulatus | LC | LC |
| Chilomycterus mauretanicus | LC | NA |
| Diodon eydouxii | LC | NA |
| Diodon holocanthus | LC | NE |
| Diodon hystrix | LC | DD |
| Masturus lanceolatus | LC | NA |
| Mola mola | VU | DD |
| Mola alexandrini | NE | NE |
| Ranzania laevis | LC | DD |
| Canthidermis sufflamen | LC | NA |
| Canthidermis maculata | LC | LC |
| Melichthys niger | LC | NE |
| Balistes capriscus | VU | DD |
| Balistes punctatus | VU | NE |
Table 5.
Detected (D), Not detected (ND), Not evaluated (NE) tetraodontoxins (TTXs) and Paralytic Shellfish Toxins (PSTs) in Spanish tetraodontiform fishes according to literature.
Table 5.
Detected (D), Not detected (ND), Not evaluated (NE) tetraodontoxins (TTXs) and Paralytic Shellfish Toxins (PSTs) in Spanish tetraodontiform fishes according to literature.
| Species | TTX | PST | Source |
|---|---|---|---|
| Sphoeroides marmoratus | D | NE | [49] |
| D | ND | [50] | |
| Sphoeroides pachygaster | ND | NE | [51] |
| ND | ND | [52] | |
| NE | ND | [53] | |
| Canthigaster capistrata | NE | NE | — |
| Lagocephalus laevigatus | D | NE | [54] |
| Lagocephalus lagocephalus | ND | NE | [51] |
| ND | D | [50] | |
| NE | D | [53] | |
| Lagocephalus sceleratus | D | NE | [55] |
| D | NE | [51] | |
| Ephippion guttifer | NE | NE | — |
| Aluterus monoceros | NE | NE | — |
| Aluterus scriptus | NE | NE | — |
| Cantherhines pullus | NE | NE | — |
| Cantherhines macrocerus | NE | NE | — |
| Stephanolepis hispida | NE | NE | — |
| Chilomycterus reticulatus | NE | NE | — |
| Chilomycterus mauretanicus | NE | NE | — |
| Diodon eydouxii | NE | NE | — |
| Diodon holocanthus | D | NE | [56] |
| ND | NE | [57] | |
| Diodon hystrix | D | NE | [58] |
| D | NE | [59] | |
| Masturus lanceolatus | NE | NE | — |
| Mola mola | ND | NE | [56] |
| ND | NE | [60] | |
| Mola alexandrini | ND | NE | [60] |
| Ranzania laevis | NE | NE | — |
| Canthidermis sufflamen | NE | NE | — |
| Canthidermis maculata | NE | NE | — |
| Melichthys niger | NE | NE | — |
| Balistes capriscus | NE | NE | — |
| Balistes punctatus | NE | NE | — |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



