Submitted:
23 January 2026
Posted:
26 January 2026
You are already at the latest version
Abstract
Aldolases are powerful biocatalysts for the stereoselective formation of carbon–carbon bonds and are widely used in the synthesis of chiral intermediates for pharmaceutical applications. Among them, 2-deoxyribose-5-phosphate aldolase (DERA) has been extensively exploited for the preparation of the conserved side chain of statins. In this work, we report a novel chemoenzymatic approach for the synthesis of nucleobase-substituted lactol products as potential precursors of new statin analogues. A C49M variant of DERA from Pectobacterium atrosepticum (PaDERA C49M) was employed to catalyze sequential aldol additions using aldehyde-functionalized nucleobases as non-natural electrophilic substrates. The formation of nucleobase-containing lactols was confirmed, demonstrating for the first time the acceptance of nucleobase-derived aldehydes in DERA-catalyzed aldol reactions. This strategy provides access to structurally novel statin side-chain precursors and expands the synthetic potential of DERA toward the generation of new classes of bioactive compounds.
Keywords:
2-deoxyribose-5-phosphate aldolase (DERA)
; Pectobacterium atrosepticum
; aldol addition
; statins
; nucleobases
1. Introduction
Cardiovascular disease remains the leading cause of mortality worldwide, with over 20 million deaths annually [1,2]. Among the established risk factors associated with its global health impact —including low physical activity, obesity, diabetes, smoking and high blood pressure—, elevated low-density lipoprotein (LDL) cholesterol represents a major determinant, implicated in approximately 3.8 million deaths [3].
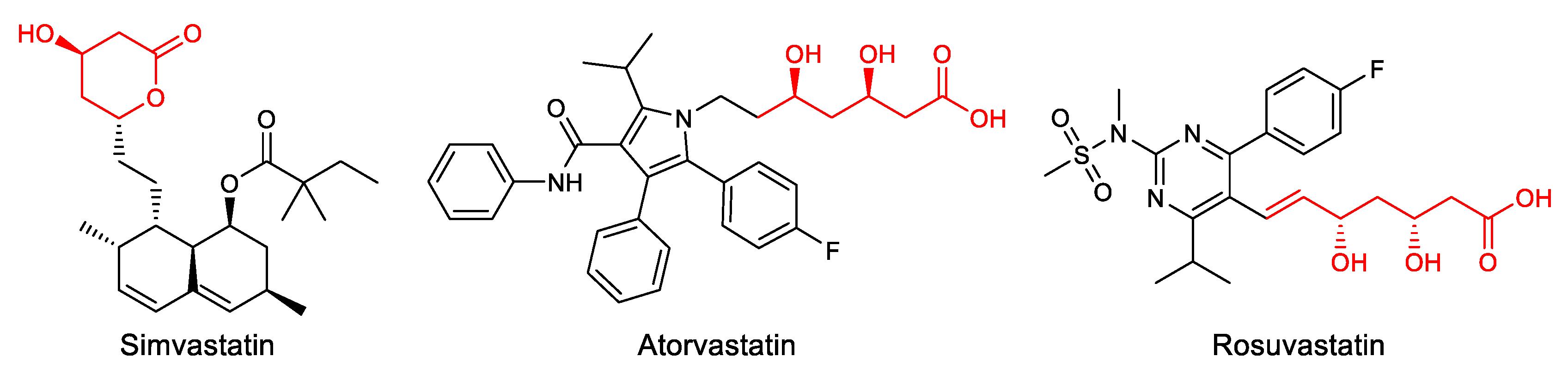
Statins are usually prescribed to reduce the levels of LDL cholesterol through inhibition of the rate limiting enzyme 3-hydroxy-3-ethylglutaryl-coenzyme A (HMG-CoA) reductase involved in the conversion of HMG-CoA to mevalonate, which is an early step in the synthesis of cholesterol [4]. Since the discovery of the first cholesterol synthesis inhibitor, Mevastatin, in the 1970s, statin families have expanded rapidly through the development of natural, semisynthetic, and fully synthetic statins (Figure 1) [5]. Although statins are generally well-tolerated and safe, adverse reactions become more significant as the number of users increases. These often include muscle weakness, hepatic damage, renal insufficiency, and the potential to induce diabetes. For this reason, there is growing interest in finding alternatives that minimize these effects [6,7].
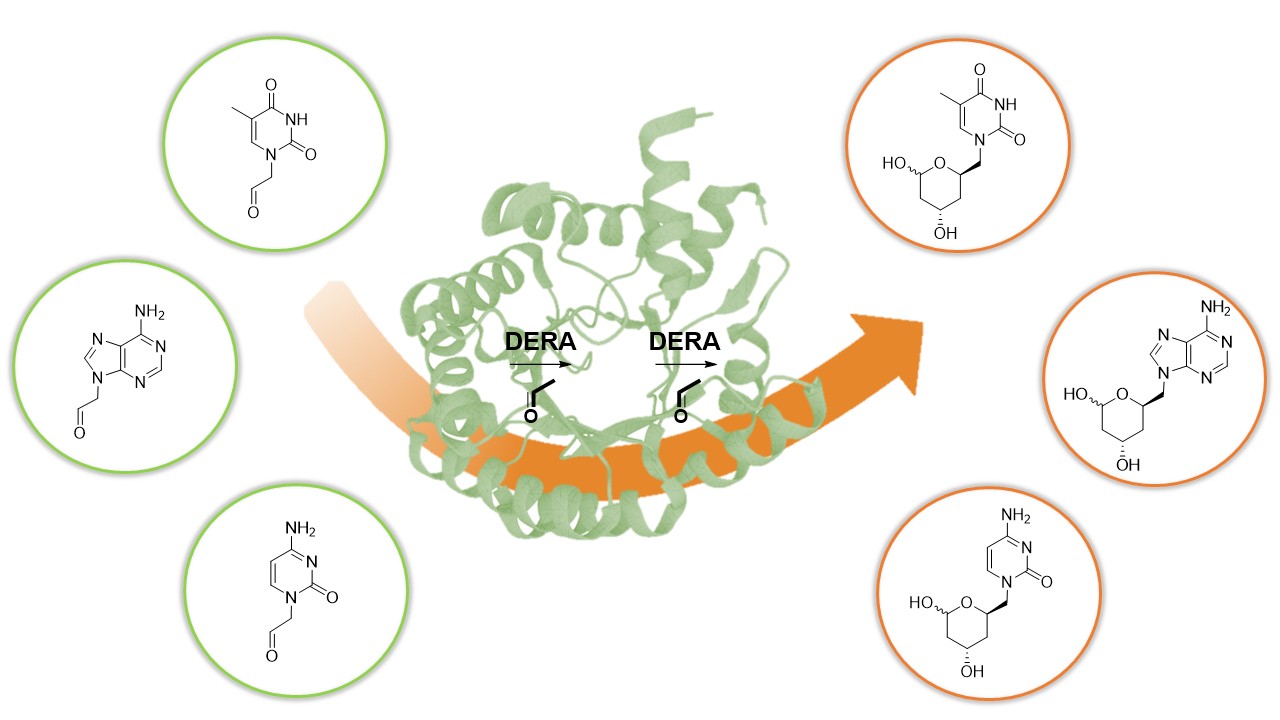
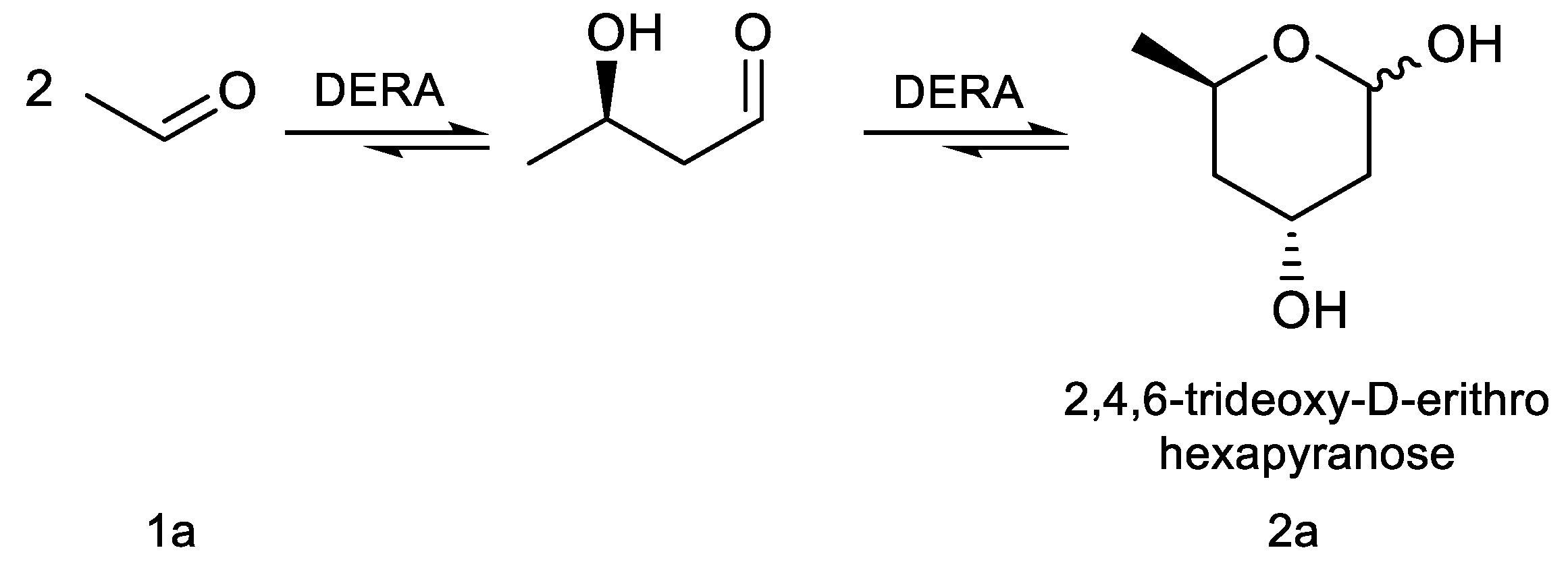
The aldol reaction is an important tool in biological systems and in organic chemistry to produce carbon skeletons, being aldolases the most suitable enzymes to catalyze this reaction in an asymmetric way [8,9]. In particular, 2-deoxyribose-5-phosphate aldolase (DERA) catalyzes the stereoselective aldol addition between two aldehydes via an enamine intermediate, typical mechanism of Class I aldolases [10]. This enzyme is strictly dependent on acetaldehyde as a nucleophile but can accept a wide range of aldehydes as electrophiles, producing 2,4,6-trideoxyhexoses (Scheme 1). This sequential aldol addition is thermodynamically controlled and stops when a stable intramolecular hemiacetal is formed [11]. The two chiral centers generated, controlled by the enzyme [12], have the proper stereochemistry for said cyclic hemiacetal (lactol) to be used as intermediates in the production of Atorvastatin and other related cholesterol-lowering drugs [13,14,15]. However, the application of DERA in large-scale production of these pharmaceutical compounds is limited by some drawbacks, such as low catalytic activity and low tolerance to aldehydes [16]. To overcome this disadvantage, different technologies such as directed evolution, protein engineering, computational design, and immobilization have been applied to improve DERA’s properties [17]. In particular, a number of recombinant DERA variants with increased productivity, from mesophilic [18], psychrophilic [19], and thermophilic organisms [20], have been reported.
The most efficient modification to provide acetaldehyde resistance involves the mutation of a cysteine, non-essential for catalysis (C47 in DERA from E. coli), by a non-nucleophilic amino acid such as methionine, which avoids the formation of a bridge via a Michael addition of crotonaldehyde, resulting from the condensation of 2 acetaldehyde molecules with the lysine K167 of the active site [16]. In this regard, EcDERA C47M showed no loss of activity after incubation in 300 mM of acetaldehyde for 16 h, in contrast to wild-type EcDERA which was completely inactivated after 3 h [21]. Similar behavior was observed in DERAs from Lactobacillus brevis [22], Rhodococcus erythropolis strain DSM 311 [23], Thermotoga maritima [24], archaeon Pyrobaculum aerophilum [25] and Staphylococcus aureus [26], among others, after cloning, modification and recombinant expression in E. coli. Previous work in our lab involved DERA from Pectobacterium atrosepticum ATCC 33260 (PaDERA) [27], which was genetically modified, cloned and expressed in E. coli. In comparison to the wild-type enzyme, PaDERA C49M (which corresponds to C47 in EcDERA) retained 90% of its activity after incubation for 90 min in 300 mM of acetaldehyde [28].
Different 6-substituted 2,4,6-trideoxy-D-erithro hexapyranoses (6-R-TDHP) were reported over time as precursors of the side chain of statins. (3R,5S)-6-chloro-2,4,6-trideoxyhexapyranoside (ClTDHP) was the first biocatalytically synthesized intermediate using diverse DERA variants. This approach represented a great improvement, in chemical and stereochemical purity and cheap achiral starting materials, over its chemical synthesis [29,30,31,32,33]. This biotransformation has been successfully used to prepare the statin side-chain of Rosuvastatin on manufacturing scale [34]. Besides chlorine, acetoxy [35], azido [36] and N-protected amino [37] groups were reported as TDHP 6-substituent in order to improve productivity. Švarc et al. [34] developed an alternative strategy in which aminophenylacetamide-protected propanal is used as the starting aldehyde of the aminolactol precursor of Atorvastatin. Lactols can then be oxidized to the corresponding lactones after which further chemical steps attach them to the heterocycle moiety to obtain the different statins [5,38,39].
The conserved 3,5-dihydroxyheptanoic acid moiety present in all statins is responsible for HMG-CoA reductase inhibition through competitive and reversible binding due to its structural similarity to its natural substrate, HMG-CoA. In addition, interactions between the enzyme and the remaining portions of the molecule contribute to binding affinity [40]. While natural statins contain a hexahydronaphthalene core, synthetic statins incorporate diverse heterocyclic structures, such as pyrrole in Atorvastatin, pyrimidine in Rosuvastatin, or quinoline in Pitavastatin [41]. Accordingly, drug development efforts aimed at improving efficacy and tolerability, have primarily targeted the heterocyclic scaffold [42].
Unlike statins, eritadenine, an acyclic adenosine analog with hypocholesterolemic activity, does not affect cholesterol production but accelerates cholesterol metabolism in the liver, modifying phospholipid metabolism and promoting the removal of cholesterol from the blood through mechanisms that increase LDL cholesterol uptake [43,44]. This led us to consider the possibility of preparing statins containing a nucleoside base such as the heterocycle. Therefore, the objective of this work was to generate the side chain precursors for new statin analogues from aldehyde derivatives of nitrogen bases, using PaDERA C49M, recently cloned and over expressed by us, and whose synthetic activity towards the formation of TDHP has already been reported [45].
2. Materials and Methods
2.1. Chemicals and Microorganisms
Reagents and substrates were purchased from Sigma-Aldrich. TLC analysis was performed using silica gel 60 F254 aluminum plates from Merck (Darmstadt, Germany). The culture media components were obtained from Anedra, Britania and Sigma-Aldrich. Solvents were from Sintorgan and Biopack. Escherichia coli DH5α (Invitrogen) and E. coli BL21 (DE3) (ATCC 47092, acid phosphatase deficient) strains were used in cloning and expression experiments.
2.2. Culture Conditions
Plasmids encoding P. atrosepticum DERA including the mutation C49M (PaDERA C49M), as described in ref. [28], were transformed into chemically competent E. coli BL21 (DE3) for expression. Each recombinant E. coli strain was grown in LB medium supplemented with kanamycin 30 μg/μL at 37 °C and 200 rpm overnight. The pre-cultures (25 mL) were then used to inoculate a fresh LB medium containing kanamycin 30 μg/μL (250 mL) and grown at 37 °C, 200 rpm until OD600 reached 0.6–0.8, at which point the protein expression was induced with isopropyl-β-thiogalactoside (IPTG) at a final concentration of 0.1 mM. The cultures continued growing at 28 °C and 180 rpm overnight. Subsequently, the cells were harvested by centrifugation (5000 rpm, 10 min, 4 °C), washed with 0.1 M potassium phosphate buffer pH 7.0, recentrifuged and used as whole cell biocatalysts (2.5x109 cells/mg wet weight) or to continue towards protein purification.
2.3. Enzyme Purification
E. coli (PaDERA C49M) cells were resuspended in 0.1 M potassium phosphate buffer pH 7 to achieve a final concentration of 30% w/v. The suspension was sonicated at 50% amplitude for four cycles at 4 °C using the Vibra Cell disruptor (VCX130, Sonics, USA) or incubated with lysozyme (1 mg/g wet weight of cells) for 24 h at 4 °C and 200 rpm. The suspension was centrifuged at 4 °C and 9000 rpm for 15 min, and the supernatant was conserved as Cells Free Extract (CFE).
For the enzyme purification by batch-mode affinity chromatography, 20 mL of Ni- Sepharose TM 6 Fast Flow resine (obtained from GE Life Science) and 20 mL of CFE were incubated for 1 h at 4 °C. After that, the solid phase (resin with bound enzyme) was separated from the supernatant by centrifugation, and the unbound CFE was discarded. The resin was then subjected to sequential washing-centrifugation steps with imidazole solutions at increasing concentrations (5 mM–500 mM). Relevant fractions of the eluted proteins were combined, desalted and concentrated to a final volume of 1 mL by centrifuging at 5700 rpm at 4 °C in a Vivaspin 20 Centrifugal Concentrator (Sartorius).
2.4. Activity Assay and Determination of Protein Concentration
The activity of PaDERA C49M was determined by the coupled assay described previously [46], measuring the oxidation of NADH. The assay mixture contained 0.1 M potassium phosphate buffer pH 7.0, 0.2 mM NADH, 0.4 mM 2-deoxyribose-5-phosphate (DR5P), 4 μL of the mixture α-glycerophosphate dehydrogenase/triosephosphate isomerase (α-GDH-TPI) and 5μL of purified protein. The reaction was initiated by the addiction of purified protein and the subsequent decrease of NADH concentration was monitored at 340 nm. 1 U of protein activity was defined as the amount of enzyme required for cleavage of 1 μmol of DR5P per minute at 28 °C. The protein concentration was determined by the Bradford assay [47]. PaDERA C49M protein concentration: 0.98 mg/mL and PaDERA C49M activity: 28.9 U/mg.
2.5. Substrate Synthesis
N-1-(2,2-dimethoxyethyl) thymine (4i), N-1-(2,2-dimethoxyethyl) cytosine (4ii), and N-9-(2,2-dimethoxyethyl) adenine (4iii), were prepared according to our previous report [48]. Mixtures of thymine (3i), cytosine (3ii) or adenine (3iii) (1 equiv.) and K2CO3 (2 equiv.) in DMF (10 mL) were stirred at 90 °C in the presence of 2-bromo-1,1-dimethoxy ethane (2 equiv.). After 24 h, the reactions were filtered and the solvent was removed under reduced pressure. The crude mixtures were purified by column chromatography on silica gel (230 mesh), using approximately 40% w/w of crude material relative to the stationary phase. Elution was performed using dichloromethane/methanol as the mobile phase, gradually increasing the polarity. Column progress was monitored by TLC, using dichloromethane/methanol (9:1) as the eluent. Aldehydes (N-1-(2-oxoethyl) thymine (5i), N-1-(2-oxoethyl) cytosine (5ii) or N-9-(2-oxoethyl) adenine (5iii) were obtained after hydrolysis of the corresponding acetals (4i-iii) in HCl (1 N) at 90 °C for 1 h and neutralization with NaOH (2 N). NMR and ESI-MS spectral characteristics are in accordance with the literature [49].
2.6. Whole Cell Biotransformations
2.6.1. Synthesis of 2,4,6-Trideoxy- D-Erythro-Hexapyranose (TDHP, 2a)
The reactions performed in 4 mL final volume 100 mM potassium phosphate buffer pH 7 carrying 1 M of acetaldehyde and 2.5x109 E. coli (PaDERA C49M) cells/mg wet weight as biocatalyst. The reaction mixture was shaken at 28 °C and 200 rpm for 24 h and then centrifuged for 10 min at 5000 rpm. The supernatant was extracted with 2 volumes of ethyl acetate and purified by silica gel column chromatography using ethyl acetate as eluent. The NMR data are in accordance with the literature [45].
2.6.2. Synthesis of 6-Substituted-2,4,6-Trideoxy-D-Erythro-Hexapyranoses (2b-e)
The reactions performed in 4 mL final volume 100 mM potassium phosphate buffer pH 7 carrying 100 mM of chloroacetaldehyde (1b), propanaldehyde (1c), phenylacetaldehyde (1d) or benzaldehyde (1e) as acceptors, acetaldehyde 200 mM as donor, and 2.5x109 E. coli (PaDERA C49M) cells/mg wet weight as biocatalyst. The reaction mixtures were shaken at 28 °C and 200 rpm for 24 h and then centrifuged for 10 min at 5000 rpm. The supernatants were extracted with 2 volumes of ethyl acetate, purified by silica gel column chromatography using ethyl acetate as eluent and analyzed by GC.
2.7. Free-Enzyme Biotransformation
2.7.1. Synthesis of 6-Substituted Lactols (6i-iii)
The reactions were carried out in a final volume of 1 mL of 0.1 M potassium phosphate buffer (pH 7), containing 100 mM (5i, 5ii, 5iii) as the acceptors, 100 mM acetaldehyde as the donor, and 1 mg/mL PaDERA C49M. The reaction mixtures were shaken at 28 °C and 200 rpm for 24 h. Subsequently, the reactions were quenched with acetonitrile and centrifuged at 11000 rpm for 3 min. The resulting supernatants were extracted three times with one volume of ethyl acetate. Compounds 6i-iii were analyzed by HPLC-UV/MS. Compound 6i was isolated from 8 mL reaction mixture through extensive extraction with ethyl acetate and subsequently purified by silica gel (230 mesh) column chromatography employing dichloromethane/methanol (9:1) as the mobile phase.
2.8. Analytical Methods
2.8.1. Instrumental
HPLC-UV analyses were carried out with a Gilson chromatograph (321 Pump, 156UV/VIS detector, and 234 Autoinjector Series; Middleton, WI, USA). GC analyses were performed with Thermo Scientific Trace 1300 using a Thermo Scientific TR-5 (30 m× 0.25 mm×0.25 μm) column using He as gas carrier (1 mL/min) and injector temperature 200 °C. The oven temperature was set at 50 °C (5 min), increased to 250 °C (10 °C/min) and maintained at the final temperature for 5 min. 1H and 13C NMR spectra were recorded on a Bruker Avance II 500 spectrometer (Madison, WI, USA) at 500 MHz and 125 MHz, respectively, or on a Bruker Avance III 600 spectrometer (Madison, WI, USA) at 600 MHz, using CDCl3 as solvent. HPLC-MS spectra were recorded on an ACQUITY UPLC system (Waters, UK) coupled online to a high-resolution Orbitrap mass spectrometer (Q-Exactive Focus, Thermo Fisher Scientific, Germany). Chromatographic separation was achieved on a reverse-phase ACQUITY UPLC BEH C18 column (1.0 × 100 mm, 1.7 μm; Waters, UK) operated at room temperature. The mobile phases consisted of water containing 0.1% formic acid (A) and acetonitrile containing 0.1% formic acid (B). A gradient was maintained at 50 μL/min at 5% B over 15 minutes. Solvent B was then increased to 75% over 22 minutes, before equilibrating back to the starting conditions. The mass spectrometer was operated alternating in full scan and PRM mode. Full scan was acquired from 50–750 m/z in ESI positive mode at a resolution of 70 K. Raw data were analyzed using XCalibur 4.1 (Thermo Fisher Scientific, Germany). The mass spectrometer was calibrated using the Pierce™ LTQ ESI positive ion calibration solution (Thermo Fisher Scientific, Germany). Spectrophotometric assays were performed at 340 nm using a UV-160A-UV-Visible recording Shimadzu spectrophotometer (Kyoto, Japan).
2.9. 1H and 13C NMR Product Data
All chromatographic and spectra data are available in the Supplementary Materials (SM) file.
2,4,6-trideoxy-d-erythro-hexapyranose (2a)
1H NMR (600 MHz, CDCl3) δ (ppm) 5.34 (t, J = 4.7 Hz, 1H, α), 5.22 – 5.12 (m, 1H, β), 4.44 (dtd, J = 12.4, 6.3, 2.4 Hz, 1H, α/β), 4.35 (q, J = 3.1 Hz, 1H, β), 4.24 (dt, J = 6.4, 3.1 Hz, 1H, α), 4.10 (qd, J = 5.7, 5.2, 2.0 Hz, 1H, β), 4.07 (d, J = 6.5 Hz, 1H, α), 3.17 (d, J = 6.1 Hz, 1H, β), 3.01 (d, J = 6.5 Hz, 1H, α), 2.05 – 1.95 (m, 1H, α/β), 1.88 – 1.76 (m, 2H, α/β), 1.56 (ddd, J = 14.2, 11.7, 2.7 Hz, 1H, α), 1.48 (ddd, J = 14.2, 11.5, 2.9 Hz, 1H, β), 1.26 (d, J = 6.3 Hz, 3H, β), 1.24 (d, J = 6.3 Hz, 3H, α).
13C NMR (126 MHz, CDCl3) δ (ppm): 94.32 (CHOH, C-1), 93.04 (CHOH, C-1 ́), 68.35 (CH, C-5), 66.84 (CH, C-5 ́), 65.11 (CHOH, C-3), 64.17 (CHOH, C-3 ́), 42.85 (CH2, C-2), 42.31 (CH2, C-2 ́), 39.92 (CH2, C-4), 39.11 (CH2, C-4 ́), 21.48 (CH3, C-6), 21.26 (CH3, C-6 ́).
6-chloro-2,4,6-trideoxy-d-erythro-hexapyranose (2b)
1H RMN (500 MHz, CDCl3): δ (ppm) 5.51 (d, J = 3.5 Hz, 1H, β), 5.41 (d, J = 3.6 Hz, 1H, α), 4.53 (dd, J = 9.0, 4.2 Hz, 1H, H-5), 4.32 (m, 1H, H-5’), 4.28 – 4.18 (m, 1H, H-3’), 3.91 (dq, J = 11.7, 6.2 Hz, 1H, H-3), 3.63 – 3.55 (m, 4H, H’s-6), 2.22 – 1.74 (H-4, H-4’, H-2b, H-2’b), 1.55 (td, J = 12.0, 3.6 Hz, 1H, H-2a), 1.47 – 1.28 (m, 1H, H-2’a).
13C NMR (126 MHz, CDCl3) δ (ppm): 93.16 (CHOH, C-1), 92.92 (CHOH, C-1’), 68.00 (CH, C-5), 64.60 (CH, C5’), 63.16 (CHOH, C-3), 63.12 (CHOH, C-3’), 47.75 (CH2, C-6), 47.28 (CH2, C-6’), 39.07 (CH2, C-2), 38.34 (CH2, C-2’), 35.49 (CH2, C-4), 34.84 (CH2, C-4’).
6-thyminyl-2,4,6-trideoxy-d-erythro-hexapyranose (6i)
1H NMR (500 MHz, DMSO-d6): δ (ppm) 11.25 (d, J = 12.5 Hz, 1H, NH), 7.44 (s, 1H, H-7), 4.85 (d, J = 9.1 Hz, 1H, H-1), 4.08 (s, 1H, H-3), 4.03 (d, J = 8.6 Hz, 1H, H-5), 3.74 (d, J = 3.5 Hz, 1H, H-6a), 3.58 (d, J = 8.4 Hz, 1H, H-6b), 1.75 (s, 3H, H-9), 1.68 (d, J = 14.0 Hz, 1H, H-2b), 1.47 (d, J = 12.7 Hz, 1H, H-4a), 1.34 (t, J = 11.6 Hz, 1H, H-2a), 1.26 (d, J = 11.6 Hz, 1H, H-4b).
13C NMR (126 MHz, DMSO-d6): δ (ppm): 164.81 (CO, C-11), 151.43 (CO, C-10), 143.03 (CH, C-7), 108.17 (CH, C-8), 92.17 (CHOH, C-1), 68.33 (CH, C-5), 63.70 (CHOH, C-3), 52.15 (CH2, C-6), 40.21 (CH2, C-2), 35.45 (CH2, C-4), 12.42 (CH3, C-9).
m/z found: 257.1124, estimated [C11H17N2O5]+: 257.2634.
3. Results
The chemical synthesis of statins presents several drawbacks including modest overall yields, the use of large amounts of hazardous reagents, and the generation of considerable chemical waste [50]. Furthermore, the stereoselective construction of the chiral centers in the side chain, the main pharmacophoric motif, is particularly challenging. In contrast, the use of biocatalysts offers advantages such as high regio and stereo selectivity, use of non-polluting raw materials and solvents, and mild reaction conditions, which helps to reduce costs and improve sustainability [51]. Among the different biocatalysts used to synthesize the conserved chiral side chain of statins [52], DERA is one of the most studied because it has the ability to simultaneously generate both stereocenters from simple aldehydes [28].
3.1. Expression and Purification of PaDERA C-His AA C49M
Pectobacterium atrosepticum ATCC 33260 DERA (PaDERA) [27] was previously cloned and expressed in E. coli BL21 (DE3) and the variant that includes a 6 ×His C terminal tag, a 5 amino acid spacer and the C49M mutation (PaDERA C49M) was selected due to its high activity in the presence of up to 300 mM acetaldehyde [28]. The synthetic activity of E. coli (PaDERA C49M) as whole cells biocatalyst was first studied in the aldol addition of glyceraldehyde-3-phosphate and acetaldehyde to synthesize 2-deoxyribose-5-phosphate in almost quantitative yield [45].
To enable the use of free PaDERA C49M as a biocatalyst, the enzyme was purified from a crude cell-free extract (CFE) obtained via two cell disruption methods, namely enzymatic and mechanical lysis. Both approaches yielded comparable enzyme recovery and overall catalytic activity. However, a significant difference emerged in terms of stereoselectivity. While the enzyme purified from enzymatically disrupted cells retained the expected stereoselectivity, the enzyme obtained following mechanical disruption exhibited a marked reduction in stereoselective performance. This effect is likely attributable to structural perturbations induced by the physically aggressive nature of mechanical lysis. Although similar losses of stereocontrol have been reported for other aldolases [53], the underlying causes of these alterations deserve further investigation.
Purification was performed by batch-mode affinity chromatography, in which the CFE was incubated with Ni-Sepharose resin in centrifuge tubes, followed by washing and centrifugation steps, since column-based purification resulted in complete loss of enzymatic activity. This effect is likely related to intrinsic structural features of the enzyme, particularly the high flexibility of the last eight C-terminal residues reported for the wild-type protein [54]. The absence of electron density in this region suggests a functional role in nucleophile activation via proton abstraction, which may explain the poor compatibility of C-terminal His-tagged constructs with Ni–NTA purification [55]. Prolonged residence time on the affinity column, along with extended exposure to imidazole and binding/elution pH conditions, may therefore promote enzyme inactivation. In contrast, affinity purification in batch mode involves shorter and less uniform contact with the resin, likely reducing these destabilizing effects and preserving catalytic activity. After the purification, the enzyme displayed an initial specific activity of 28.9 U/mg; however, activity declined to 17.19 U/mg after 24 h at 4 °C and was completely lost after 48 h. Limited stability of isolated DERAs has been widely reported, and although approaches such as protein engineering and enzyme immobilization have been investigated, improving their stability continues to be challenging [32,45,56].
3.2. PaDERA C-His AA C49M and Chiral Lactols
The use of whole-cell biocatalysts represents an attractive alternative for industrial processes, not only because of their low production cost, but also because they help mitigate stability issues and circumvent the need for laborious enzyme purification. E. coli (PaDERA C49M) was employed as whole cell biocatalyst to prepare 2,4,6-trideoxy-d-erythro-hexapyranose (TDHP, 2a) in 99% yield by tandem aldol addition of acetaldehyde even at concentration of 500 mM [57].
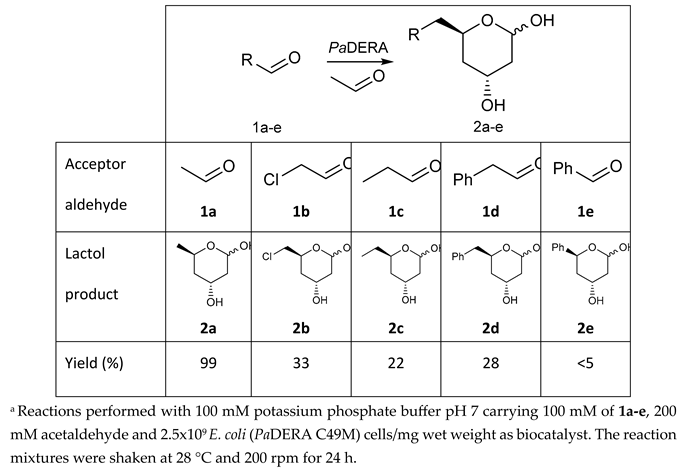
To validate the potential of the new mutant as versatile whole cell biocatalyst, we performed a DERA-catalyzed aldol addition using acetaldehyde as donor and different acceptor aldehydes as shown in Table 1. Chloroacetaldehyde (1b) was the first substrate studied and used in DERA-catalyzed reactions. Moreover, it is the most widely industrially employed due to the versatility of chlorine to participate in subsequent reactions to introduce the heterocycle moiety to statins preparation [52,54]. Additionally, propanaldehyde (1c), phenylacetaldehyde (1d) and benzaldehyde (1e) allowed us to explore the acceptance of non-phosphorylated electrophilic substrates by DERA, non-polar, bulky and aromatic respectively. None of the substrates were as good as 1a for PaDERA C49M. As usual, aromatic aldehydes were not substrates for the enzyme [54]. Recently, Rizzo et al. [53], using metagenomic technology for enzyme discovery, reported for the first time a DERA capable of accepting furfural and benzaldehyde with higher activity and stereoselectivity than those achieved with other well-known DERAs with non-natural substrates.
All biotransformations were carried out using experimental conditions previously optimized for the synthesis of 2a [57]. After biotransformations, lactols 2b-e were extracted with ethyl acetate and further purified by silica column chromatography. Due to their instability and the presence of byproducts, the purification was quite problematic. Indeed, given the difficulty of purifying these compounds, most reports that employ them as intermediates utilize the crude product [56]. All compounds were quantified by GC, observing the presence of TDHP as a secondary product, a pattern that was maintained in all aldol biotransformations. Only 2b was isolated and analyzed by 1H and 13C NMR which were consistent with previous reports [33,57]. The presence of both anomers was corroborated by the duplication of signals in both spectra and, in particular, by the signals corresponding to the hydrogens bonded to the anomeric carbons of the two anomers at 5.41 ppm and 5.51 ppm of the 1H NMR spectrum with a 7:3 (α/β) ratio (Figures S5 and S6).
3.3. Novel Statin Precursors
As previously mentioned, traditional statin synthesis involves the construction of the functionalized chiral side-chain with a reactive group (e.g., chloro, cyano, or acetoxy), which then serves as building block for introducing the corresponding heterocycle [54]. For over two decades, our laboratory has focused on the synthesis of nucleoside analogues using biocatalytic approaches. In the last years we have explored the use of different aldolases as biocatalysts for the synthesis of acyclic nucleoside analogues employing aldehyde-functionalized nucleobases as key precursors [48,58,59]. In line with these efforts, we implemented a chemoenzymatic strategy aimed at generating novel nucleobase-containing statin candidates. This approach comprises the synthesis of suitably functionalized aldehyde nucleobases, their conversion into the corresponding substituted lactols through PaDERA C49M catalysis, and a final oxidation step affording the target lactone (Scheme 2).
Compounds 5i–iii were obtained by N-alkylation of the corresponding bases 3i–iii, followed by acid hydrolysis of the acetal intermediates 4i–iii, as previously reported [49]. The resulting aldehydes were then added to a phosphate-buffered solution containing E. coli (PaDERA C49M) whole cells and acetaldehyde, which serves as the specific donor. Upon substrate consumption, analysis of the crude reaction mixture and the extracted supernatant revealed several low-abundance, hard-to-purify byproducts, most likely originating from biocatalyst-mediated substrate decomposition.
In light of these results, the crude cell-free extract (CFE) obtained from disrupted E. coli (PaDERA C49M) whole cells was tested as biocatalyst. This strategy did not lead to improved outcomes as the small amount of the product formed exhibited degradation after 3 h of reaction. However, the appearance of the product allowed us to determine that aldehydes 5i-iii were substrates of DERA and that, probably due to their structures being similar to natural nucleosides, they could also be substrates for other enzymes present in the CFE. Therefore, the use of free purified PaDERA C49M as a biocatalyst for this biotransformation was necessary.
The enzyme was purified as described above and used immediately to minimize storage instability. The amount of enzyme used (1 mg/mL) was equivalent to the biocatalyst loading employed in the other formats. In order to reduce the formation of TDHP as a by-product, the 1a:5i-iii ratio was adjusted to 1:1. However, the strong preference of 1a as both donor and acceptor substrate made this unavoidable, and TDHP formation was still observed. Additionally, the presence of TDHP further increased the intrinsic difficulty of product purification, as is commonly observed for nucleobase derivatives.
The biotransformation was performed in a buffered solution containing substrates and the enzyme. Reaction progress was monitored by TLC and HPLC, and upon completion, products 6i-iii formation was confirmed by HPLC–MS (Figures S15-S20).
As discussed above, the purification of lactol products from reaction mixtures is challenging due to the instability of the hemiacetal moiety, the coexistence of anomeric forms, and the poor solubility of nucleoside bases in organic solvents. In addition, when DERA is employed to catalyze sequential aldol reactions with non-natural substrates, the catalytic efficiency is quite low, which restricts its practical applicability. Consequently, scaling up the reaction was required to obtain sufficient amounts of material for successful purification [28].
Following scale-up, the lactol product 6i was isolated from 8 mL reaction mixture through an extensive work-up and chromatographic purification. Due to the high polarity of the nucleoside scaffold and the lability of the hemiacetal function, the crude reaction mixture was first subjected to multiple extractions with ethyl acetate. The combined organic phases were dried, filtered, and concentrated under reduced pressure. The resulting residue was purified by column chromatography on silica gel (230 mesh), using approximately 40% w/w of crude material relative to the stationary phase. Elution was performed using dichloromethane/methanol as the mobile phase, gradually increasing the polarity until a final ratio of 90:10 was reached. Column progress was monitored by TLC, using dichloromethane/methanol (9:1) as the eluent, where the desired lactol exhibited an Rf value of 0.20. The product was obtained as a colorless, viscous liquid in 28% yield.
1H and 13C NMR spectroscopic analyses confirmed the successful isolation of the lactol product and revealed the presence of a mixture of anomeric forms resulting from cyclization of the thymine-substituted lactol. The observed α/β anomeric ratio is consistent with that previously reported for related substituted lactols bearing different R groups [60]. As shown in Figure 2, the 1H NMR spectrum of an anomerically enriched mixture clearly displays the signal corresponding to the major anomer, appearing as a doublet with a coupling constant of 9.1 Hz, which is characteristic of the α-anomer. In the expanded region of this part of the spectrum, a significantly weaker signal attributable to the β-anomer can also be observed. Minor impurities attributable to residual TDHP were also detected in the crude material but were effectively removed during chromatographic purification.
The scale up and purification of 6ii and 6iii still remains a challenge.
4. Conclusions
In this work, a novel chemoenzymatic strategy was developed for the synthesis of nucleobase-substituted lactol products as potential precursors of new statin analogues. The C49M variant of 2-deoxyribose-5-phosphate aldolase from Pectobacterium atrosepticum (PaDERA C49M) was demonstrated to catalyze aldol reactions involving aldehyde-functionalized nucleobases as non-natural electrophilic substrates, thereby expanding the substrate scope of DERA beyond conventional electrophiles and providing access to previously unexplored statin side-chain architectures.
Notably, the methodology presented here introduces a distinct approach to statin synthesis, as the lateral chain is constructed directly from substrates already bearing the heterocyclic moiety. By incorporating the nucleobase at an early stage, this strategy avoids post-aldol coupling steps commonly required in traditional synthetic routes, which often result in longer, less efficient, and lower-yielding synthetic sequences.
While significant progress has been achieved in improving DERA stability-particularly through the use of enzymes from thermophilic organisms or via random mutagenesis-several challenges must still be addressed before DERA can be considered a robust and broadly applicable industrial biocatalyst. These include limitations in catalytic efficiency with non-natural substrates, product stability, and downstream processing.
Looking forward, constant efforts focus on enzyme optimization and reaction engineering to improve productivity and selectivity. In parallel, the lactol oxidation to lactone and the biological evaluation of the newly synthesized nucleobase-containing statin analogues are currently in progress, with the aim of assessing their pharmacological potential and identifying promising candidates for further development.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: DNA sequence of Pectobacterium atrosepticum DERA. Figure S2: Amino acid sequences of PaDERA and PaDERA C49M Figure S3: 1H NMR analysis of 2a: 2,4,6-trideoxy-D-erythro-hexapyranose obtained with DERA as biocatalyst. Figure S4: 13C NMR analysis of 2a: 2,4,6-trideoxy-D-erythro-hexapyranose obtained with DERA as biocatalyst. Figure S5: 1H NMR analysis of 2b: 6-chloro-2,4,6-trideoxy-D-erythro-hexapyranose obtained with DERA as biocatalyst. Figure S6: 13C NMR analysis of 2b: 6-chloro-2,4,6-trideoxy-D-erythro-hexapyranose obtained with DERA as biocatalyst. Figure S7: 1H NMR analysis of 6i: 6-thyminyl-2,4,6-trideoxy-D-erythro-hexopyranose obtained with DERA as biocatalyst. Figure S8: 13C NMR analysis of 6i: 6-thyminyl-2,4,6-trideoxy-D-erythro-hexopyranose obtained with DERA as biocatalyst. Figure S9: 1H NMR analysis of 4i: N-1-(2,2-dimethoxyethyl) thymine obtained by chemical synthesis. Figure S10: 1H NMR analysis of 4ii: N-1-(2,2-dimethoxyethyl) cytosine obtained by chemical synthesis. Figure S11: 13C NMR analysis of 4ii: N-1-(2,2-dimethoxyethyl) cytosine obtained by chemical synthesis. Figure S12: 1H NMR analysis of 4iii: N-9-(2,2-dimethoxyethyl) adenine obtained by chemical synthesis. Figure S13: 13C NMR analysis of 4iii: N-9-(2,2-dimethoxyethyl) adenine obtained by chemical synthesis. Figure S14: GC analysis of 2a: 2,4,6-trideoxy-D-erythro-hexapyranose (TDHP). Rt: 13.9 minutes. Figure S15: GC analysis of 2c: 6-methyl-2,4,6-trideoxy-D-erythro-hexapyranose. Rt: 15.3 minutes. Figure S16: GC analysis of 2d: 6-phenyl-2,4,6-trideoxy-D-erythro-hexapyranose. Rt: 18.7 minutes. Figure S17: LC-ESI-MS analysis for 6i: 6-thyminyl-2,4,6-trideoxy-D-erythro-hexapyranose (256 m/z). A) total ion chromatogram in the positive mode for compound 6i at Rt 7.49 mins. B) mass spectra found as [M+H+] corresponding to the product [C11H17N2O5]+: 257.1124). Figure S18: LC-ESI-MS analysis for 6ii: 6-cytosyl-2,4,6-trideoxy-D-erythro-hexapyranose (243 m/z). A) total ion chromatogram in the positive mode for compound 6ii at Rt 3.19 mins. B) mass spectra found as [M-H+] corresponding to the product [C10H16N3O4]-: 242.1127). Figure S19: LC-ESI-MS analysis for 6iii: 6-adenyl-2,4,6-trideoxy-d-erythro-hexapyranose (265 m/z). A) total ion chromatogram in the positive mode for compound 6iii at Rt 8.27 mins. B) mass spectra found as [M+H+] corresponding to the product [C11H16N5O3]+: 266.1240). Table S1: The qualitative analysis of whole-cell biotransformations (2a-d) were performed by TLC using acetonitrile:ethylether 1:1 (v/v) as a solvent and the spots were visualized by heating after spraying with 5 % v/v H2SO4 in ethanol. Table S2: The qualitative analysis of free-enzyme biotransformations were performed by TLC using dichloromethane:methanol 9:1 (v/v) as solvent and the spots were visualized by UV light. Table S3: The qualitative analysis of substrate synthesis was performed by TLC using dichloromethane:methanol 9:1 (v/v) as solvent and the spots were visualized by UV light.
Author Contributions
Performed the experiments, R.F.V., E.A., L.G., L.C., L.L.; writing—review and editing, R.F.V., E.A., P.-L. H., U. H., L.G., L.L., A.V. and E.L.; funding acquisition and supervision, U.H., A.I., and E.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Universidad Nacional de Quilmes, Agencia Nacional de Promoción Científica y Tecnológica (ANPCYT), National Council of Science and Technology, CONICET, Argentina, and ERACoBiotech (grant 053.80.737; BioDiMet); L.L., A.I. and E.S.L. are research members of CONICET; L.A.M.G. and R.N.F.V are CONICET fellows.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author(s).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Laranjo, L.; Lanas, F.; Sun, M.C.; Chen, D.A.; Hynes, L.; Imran, T.F.; Kazi, D.S.; Kengne, A.P.; Komiyama, M.; Kuwabara, M.; et al. World Heart Federation Roadmap for Secondary Prevention of Cardiovascular Disease: 2023 Update. Global Heart 2024, 19. [Google Scholar] [CrossRef] [PubMed]
- Di Cesare, M.; McGhie, D.V.; Perel, P.; Mwangi, J.; Taylor, S.; Pervan, B.; Kabudula, C.; Narula, J.; Bixby, H.; Pineiro, D.; et al. The Heart of the World. Global Heart 2024, 19. [Google Scholar] [CrossRef] [PubMed]
- Münzel, T.; Sørensen, M.; Lelieveld, J.; Landrigan, P.J.; Kuntic, M.; Nieuwenhuijsen, M.; Miller, M.R.; Schneider, A.; Daiber, A. A Comprehensive Review/Expert Statement on Environmental Risk Factors of Cardiovascular Disease. Cardiovascular Research 2025, 121, 1653–1678. [Google Scholar] [CrossRef] [PubMed]
- Virani, S.S.; Ramsey, D.J.; Westerman, D.; Kuebeler, M.K.; Chen, L.; Akeroyd, J.M.; Gobbel, G.T.; Ballantyne, C.M.; Petersen, L.A.; Turchin, A.; et al. Cluster Randomized Trial of a Personalized Clinical Decision Support Intervention to Improve Statin Prescribing in Patients with Atherosclerotic Cardiovascular Disease. Circulation 2023, 147, 1411–1413. [Google Scholar] [CrossRef]
- Tang, X. L.; Yu, J. W.; Geng, Y. H.; Wang, J. R.; Zheng, R. C.; Zheng, Y.G. From Discovery to Mass Production: A Perspective on Bio-Manufacturing Exemplified by the Development of Statins. Engineering 2023, 24, 138–150. [Google Scholar] [CrossRef]
- Zeng, W.; Deng, H.; Luo, Y.; Zhong, S.; Huang, M.; Tomlinson, B. Advances in Statin Adverse Reactions and the Potential Mechanisms: A Systematic Review. Journal of Advanced Research 2025, 76, 781–797. [Google Scholar] [CrossRef]
- Ward, N.C.; Watts, G.F.; Eckel, R.H. Statin Toxicity: Mechanistic Insights and Clinical Implications. Circulation Research 2019, 124, 328–350. [Google Scholar] [CrossRef]
- Kim, T.; Stogios, P.J.; Khusnutdinova, A.N.; Nemr, K.; Skarina, T.; Flick, R.; Joo, J.C.; Mahadevan, R.; Savchenko, A.; Yakunin, A.F. Rational Engineering of 2-Deoxyribose-5-Phosphate Aldolases for the Biosynthesis of (R)-1,3-Butanediol. Journal of Biological Chemistry 2020, 295, 597–609. [Google Scholar] [CrossRef]
- Roldán, R.; Sanchez-Moreno, I.; Scheidt, T.; Hélaine, V.; Lemaire, M.; Parella, T.; Clapés, P.; Fessner, W.D.; Guérard-Hélaine, C. Breaking the Dogma of Aldolase Specificity: Simple Aliphatic Ketones and Aldehydes Are Nucleophiles for Fructose-6-Phosphate Aldolase. Chemistry - A European Journal 2017, 23, 5005–5009. [Google Scholar] [CrossRef]
- Chambre, D.; Guérard-Hélaine, C.; Darii, E.; Mariage, A.; Petit, J.L.; Salanoubat, M.; De Berardinis, V.; Lemaire, M.; Hélaine, V. 2-Deoxyribose-5-Phosphate Aldolase, a Remarkably Tolerant Aldolase towards Nucleophile Substrates. Chemical Communications 2019, 55, 7498–7501. [Google Scholar] [CrossRef]
- Haridas, M.; Abdelraheem, E.M.M.; Hanefeld, U. 2-Deoxy-d-Ribose-5-Phosphate Aldolase (DERA): Applications and Modifications. Applied Microbiology and Biotechnology 2018, 102, 9959–9971. [Google Scholar] [CrossRef] [PubMed]
- Laurent, V.; Hélaine, V.; Vergne-Vaxelaire, C.; Nauton, L.; Traikia, M.; Petit, J.L.; Salanoubat, M.; De Berardinis, V.; Lemaire, M.; Guérard-Hélaine, C. Achiral Hydroxypyruvaldehyde Phosphate as a Platform for Multi-Aldolases Cascade Synthesis of Diuloses and for a Quadruple Acetaldehyde Addition Catalyzed by 2-Deoxyribose-5-Phosphate Aldolases. ACS Catalysis 2019, 9, 9508–9512. [Google Scholar] [CrossRef]
- Gijsen, H.J.M.; Wong, C.H. Sequential One-Pot Aldol Reactions Catalyzed by 2-Deoxyribose-5-Phosphate Aldolase and Fructose-1,6-Diphosphate Aldolase. Journal of the American Chemical Society 1995, 117, 2947–2948. [Google Scholar] [CrossRef]
- Patel, R.N. Biocatalysis for Synthesis of Pharmaceuticals. Bioorganic and Medicinal Chemistry 2018, 26, 1252–1274. [Google Scholar] [CrossRef]
- Wong, C.H.; Gijsen, H.J.M. Unprecedented Asymmetric Aldol Reactions with Three Aldehyde Substrates Catalyzed by 2-Deoxyribose-5-Phosphate Aldolase. Journal of the American Chemical Society 1994, 8422–8423. [Google Scholar] [CrossRef]
- Dick, M.; Hartmann, R.; Weiergräber, O.H.; Bisterfeld, C.; Classen, T.; Schwarten, M.; Neudecker, P.; Willbold, D.; Pietruszka, J. Mechanism-Based Inhibition of an Aldolase at High Concentrations of Its Natural Substrate Acetaldehyde: Structural Insights and Protective Strategies. Chemical Science 2016, 7, 4492–4502. [Google Scholar] [CrossRef]
- Fei, H.; Zheng, C. cai; Liu, X. ye; Li, Q. An Industrially Applied Biocatalyst: 2-Deoxy-D-Ribose-5- Phosphate Aldolase. Process Biochemistry 2017, 63, 55–59. [Google Scholar] [CrossRef]
- Voutilainen, S.; Heinonen, M.; Andberg, M.; Jokinen, E.; Maaheimo, H.; Pääkkönen, J.; Hakulinen, N.; Rouvinen, J.; Lähdesmäki, H.; Kaski, S.; et al. Substrate Specificity of 2-Deoxy-D-Ribose 5-Phosphate Aldolase (DERA) Assessed by Different Protein Engineering and Machine Learning Methods. Applied Microbiology and Biotechnology 2020, 104, 10515–10529. [Google Scholar] [CrossRef]
- He, F. fan; Xin, Y. yao; Ma, Y. xin; Yang, S.; Fei, H. Rational Design to Enhance the Catalytic Activity of 2-Deoxy-D-Ribose-5-Phosphate Aldolase from Pseudomonas Syringae Pv. Syringae B728a. Protein Expression and Purification 2021, 183, 105863. [Google Scholar] [CrossRef]
- Yin, X.; Wang, Q.; Zhao, S. juan; Du, P.F.; Xie, K.L.; Jin, P.; Xie, T. Cloning and Characterization of a Thermostable 2- Deoxy-D-Ribose-5-Phosphate Aldolase from Aciduliprofundum Boonei. African Journal of Biotechnology 2011, 10, 16260–16266. [Google Scholar] [CrossRef]
- Bramski, J.; Dick, M.; Pietruszka, J.; Classen, T. Probing the Acetaldehyde-Sensitivity of 2-Deoxy-Ribose-5-Phosphate Aldolase (DERA) Leads to Resistant Variants. Journal of Biotechnology 2017, 258, 56–58. [Google Scholar] [CrossRef]
- Jiao, X.C.; Pan, J.; Kong, X.D.; Xu, J.H. Protein Engineering of Aldolase LbDERA for Enhanced Activity toward Real Substrates with a High-Throughput Screening Method Coupled with an Aldehyde Dehydrogenase. Biochemical and Biophysical Research Communications 2017, 482, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Kullartz, I.; Pietruszka, J. Cloning and Characterisation of a New 2-Deoxy-d-Ribose-5-Phosphate Aldolase from Rhodococcus Erythropolis. Journal of Biotechnology 2012, 161, 174–180. [Google Scholar] [CrossRef]
- Sakuraba, H.; Yoneda, K.; Yoshihara, K.; Satoh, K.; Kawakami, R.; Uto, Y.; Tsuge, H.; Takahashi, K.; Hori, H.; Ohshima, T. Sequential Aldol Condensation Catalyzed by Hyperthermophilic 2-Deoxy-D-Ribose-5-Phosphate Aldolase. Applied and Environmental Microbiology 2007, 73, 7427–7434. [Google Scholar] [CrossRef] [PubMed]
- Sakuraba, H.; Tsuge, H.; Shimoya, I.; Kawakami, R.; Goda, S.; Kawarabayasi, Y.; Katunuma, N.; Ago, H.; Miyano, M.; Ohshima, T. The First Crystal Structure of Archaeal Aldolase. Unique Tetrameric Structure of 2-Deoxy-D-Ribose-5-Phosphate Aldolase from the Hyperthermophilic Archaea Aeropyrum Pernix. Journal of Biological Chemistry 2003, 278, 10799–10806. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Chen, X.; Feng, J. Cloning, Purification and Characterization of Promising 2-Deoxy-D-Ribose 5-Phosphate Aldolase from Staphylococcus Aureus N315. Chin. J. Bioprocess Eng 2013, 11, 47–53. [Google Scholar]
- Valino, A.L.; Palazzolo, M.A.; Iribarren, A.M.; Lewkowicz, E. Selection of a New Whole Cell Biocatalyst for the Synthesis of 2-Deoxyribose 5-Phosphate. Applied Biochemistry and Biotechnology 2012, 166, 300–308. [Google Scholar] [CrossRef]
- Haridas, M.; Bisterfeld, C.; Chen, L.M.; Marsden, S.R.; Tonin, F.; Médici, R.; Iribarren, A.; Lewkowicz, E.; Hagedoorn, P.-L.; Hanefeld, U.; et al. Discovery and Engineering of an Aldehyde Tolerant 2-Deoxy-D-Ribose 5-Phosphate Aldolase (DERA) from Pectobacterium Atrosepticum. Catalysts 2020, 10, 883. [Google Scholar] [CrossRef]
- Rouvinen, J.; Andberg, M.; Pääkkönen, J.; Hakulinen, N.; Koivula, A. Current State of and Need for Enzyme Engineering of 2-Deoxy-D-Ribose 5-Phosphate Aldolases and Its Impact. Applied Microbiology and Biotechnology 2021, 105, 6215–6228. [Google Scholar] [CrossRef]
- Greenberg, W.A.; Varvak, A.; Hanson, S.R.; Wong, K.; Huang, H.; Chen, P.; Burk, M.J. Development of an Efficient, Scalable, Aldolase-Catalyzed Process for Enantioselective Synthesis of Statin Intermediates. Proceedings of the National Academy of Sciences of the United States of America 2004, 101, 5788–5793. [Google Scholar] [CrossRef]
- Jennewein, S.; Schürmann, M.; Wolberg, M.; Hilker, I.; Luiten, R.; Wubbolts, M.; Mink, D. Directed Evolution of an Industrial Biocatalyst 2-Deoxy-d-Ribose 5-Phosphate Aldolase. Biotechnology Journal 2006, 1, 537–548. [Google Scholar] [CrossRef] [PubMed]
- Jiao, X.C.; Pan, J.; Xu, G.C.; Kong, X.D.; Chen, Q.; Zhang, Z.J.; Xu, J.H. Efficient Synthesis of a Statin Precursor in High Space-Time Yield by a New Aldehyde-Tolerant Aldolase Identified from Lactobacillus Brevis. Catalysis Science and Technology 2015, 5, 4048–4054. [Google Scholar] [CrossRef]
- Xuan, K.; Yang, G.; Wu, Z.; Xu, Y.; Zhang, R. Efficient Synthesis of (3R,5S)-6-Chloro-2,4,6-Trideoxyhexapyranose by Using New 2-Deoxy-D-Ribose-5-Phosphate Aldolase from Streptococcus Suis with Moderate Activity and Aldehyde Tolerance. Process Biochemistry 2020, 92, 113–119. [Google Scholar] [CrossRef]
- Švarc, A.; Fekete, M.; Hernandez, K.; Clapés, P.; Findrik Blažević, Z.; Szekrenyi, A.; Skendrović, D.; Vasić-Rački, Đ.; Charnock, S.J.; Presečki, A.V. An Innovative Route for the Production of Atorvastatin Side-Chain Precursor by DERA-Catalysed Double Aldol Addition. Chemical Engineering Science 2021, 231, 116312. [Google Scholar] [CrossRef]
- Ošlaj, M.; Cluzeau, J.; Orkić, D.; Kopitar, G.; Mrak, P.; Časar, Z. A Highly Productive, Whole-Cell DERA Chemoenzymatic Process for Production of Key Lactonized Side-Chain Intermediates in Statin Synthesis. PLoS ONE 2013, 8, e62250. [Google Scholar] [CrossRef]
- Liu, J.; Hsu, C.C.; Wong, C.H. Sequential Aldol Condensation Catalyzed by DERA Mutant Ser238Asp and a Formal Total Synthesis of Atorvastatin. Tetrahedron Letters 2004, 45, 2439–2441. [Google Scholar] [CrossRef]
- Bauer, D.W.; O’Nell, P.M.; Watson, T.J.; Hu, S. United States Patent 8,642,783 B2, 2011.
- Ručigaj, A.; Krajnc, M. Optimization of a Crude Deoxyribose-5-Phosphate Aldolase Lyzate-Catalyzed Process in Synthesis of Statin Intermediates. Organic Process Research and Development 2013, 17, 854–862. [Google Scholar] [CrossRef]
- García-Bofill, M.; Sutton, P.W.; Guillén, M.; Álvaro, G. Enzymatic Synthesis of a Statin Precursor by Immobilised Alcohol Dehydrogenase with NADPH Oxidase as Cofactor Regeneration System. Applied Catalysis A: General 2021, 609. [Google Scholar] [CrossRef]
- Istvan, E.S.; Deisenhofer, J. Structural Mechanism for Statin Inhibition of HMG-CoA Reductase. Science 2001, 292, 1160–1164. [Google Scholar] [CrossRef]
- Sadowska, A.; Osiński, P.; Roztocka, A.; Kaczmarz-Chojnacka, K.; Zapora, E.; Sawicka, D.; Car, H. Statins — From Fungi to Pharmacy. International Journal of Molecular Sciences 2023, 25, 466–491. [Google Scholar] [CrossRef]
- Li, X.Z.; Jiang, S.Y.; Li, G.Q.; Jiang, Q.R.; Li, J.W.; Li, C.C.; Han, Y.Q.; Song, B.L.; Ma, X.R.; Qi, W.; et al. Synthesis of Heterocyclic Ring-Fused Analogs of HMG499 as Novel Degraders of HMG-CoA Reductase That Lower Cholesterol. European Journal of Medicinal Chemistry 2022, 236, 114323. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Komoto, J.; Lou, K.; Ueki, A.; Hua, D.H.; Sugiyama, K.; Takata, Y.; Ogawa, H.; Takusagawa, F. Structure and Function of Eritadenine and Its 3-Deaza Analogues: Potent Inhibitors of S-Adenosylhomocysteine Hydrolase and Hypocholesterolemic Agents. Biochemical Pharmacology 2007, 73, 981–989. [Google Scholar] [CrossRef] [PubMed]
- Ishara, J.; Buzera, A.; Mushagalusa, G. N.; Hammam, A. R.; Munga, J.; Karanja, P.; Kinyuru, J. Nutraceutical Potential of Mushroom Bioactive Metabolites and Their Food Functionality. Journal of Food Biochemistry 2022, 46, e14025. [Google Scholar] [CrossRef] [PubMed]
- Fernández Varela, R.; Valino, A.L.; Abdelraheem, E.; Médici, R.; Sayé, M.; Pereira, C.A.; Hagedoorn, P.; Hanefeld, U.; Iribarren, A.; Lewkowicz, E. Synthetic Activity of Recombinant Whole Cell Biocatalysts Containing 2-Deoxy-D-ribose-5-phosphate Aldolase from Pectobacterium Atrosepticum. ChemBioChem 2022, 202200147, 1–7. [Google Scholar] [CrossRef]
- Švarc, A.; Findrik Blažević, Z.; Vasić-Rački, Đ.; Szekrenyi, A.; Fessner, W.D.; Charnock, S.J.; Vrsalović Presečki, A. 2-Deoxyribose-5-Phosphate Aldolase from Thermotoga Maritima in the Synthesis of a Statin Side-Chain Precursor: Characterization, Modeling and Optimization. Journal of Chemical Technology and Biotechnology 2019, 94, 1832–1842. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantification of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Analytical Biochemistry 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Palazzolo, M.A.; Nigro, M.J.; Iribarren, A.M.; Lewkowicz, E.S. A Chemoenzymatic Route to Prepare Acyclic Nucleoside Analogues. European Journal of Organic Chemistry 2016, 2016, 921–924. [Google Scholar] [CrossRef]
- Nigro, M.J.; Brardinelli, J.I.; Lewkowicz, E.S.; Iribarren, A.M.; Laurella, S.L. Aldehyde-Hydrate Equilibrium in Nucleobase 2-Oxoethyl Derivatives: An NMR, ESI-MS and Theoretical Study. Journal of Molecular Structure 2017, 1144, 49–57. [Google Scholar] [CrossRef]
- Butler, D.E.; Le, T.V.; Millar, A.; Nanninga, T.N. United States Patent 5155251, 1992.
- Wolberg, M.; Filho, M.V.; Bode, S.; Geilenkirchen, P.; Feldmann, R.; Liese, A.; Hummel, W.; Müller, M. Chemoenzymatic Synthesis of the Chiral Side-Chain of Statins: Application of an Alcohol Dehydrogenase Catalysed Ketone Reduction on a Large Scale. Bioprocess and Biosystems Engineering 2008, 31, 183–191. [Google Scholar] [CrossRef]
- Hoyos, P.; Pace, V.; Alcántara, A.R. Biocatalyzed Synthesis of Statins: A Sustainable Strategy for the Preparation of Valuable Drugs. Catalysts 2019, 9, 1–32. [Google Scholar] [CrossRef]
- Rizzo, A.; Aranda, C.; Galman, J.; Alcasabas, A.; Pandya, A.; Bornadel, A.; Costa, B.; Hailes, H.C.; Ward, J.M.; Jeffries, J.W.E.; et al. Broadening The Substrate Scope of Aldolases Through Metagenomic Enzyme Discovery. ChemBioChem 2024, 25. [Google Scholar] [CrossRef]
- Hélaine, V.; Gastaldi, C.; Lemaire, M.; Clapés, P.; Guérard-Hélaine, C. Recent Advances in the Substrate Selectivity of Aldolases. ACS Catalysis 2022, 12, 733–761. [Google Scholar] [CrossRef]
- Schulte, M.; Petrović, D.; Neudecker, P.; Hartmann, R.; Pietruszka, J.; Willbold, S.; Willbold, D.; Panwalkar, V. Conformational Sampling of the Intrinsically Disordered C-Terminal Tail of DERA Is Important for Enzyme Catalysis. ACS Catalysis 2018, 8, 3971–3984. [Google Scholar] [CrossRef] [PubMed]
- Abdelraheem, E.; Kuijpers, R.; Hagedoorn, P.L.; Hollmann, F.; Hanefeld, U. Enzymatic Cascade of DERA and ADH for Lactone Synthesis. Catalysis Science and Technology 2024, 14, 2739–2751. [Google Scholar] [CrossRef]
- Müller, M. Chemoenzymatic Synthesis of Building Blocks for Statin Side Chains. Angewandte Chemie - International Edition 2005, 44, 362–365. [Google Scholar] [CrossRef]
- Nigro, M.J.; Palazzolo, M.A.; Colasurdo, D.; Iribarren, A.M.; Lewkowicz, E.S. N-Acetylneuraminic Acid Aldolase-Catalyzed Synthesis of Acyclic Nucleoside Analogues Carrying a 4-Hydroxy-2-Oxoacid Moiety. Catalysis Communications 2019, 121, 73–77. [Google Scholar] [CrossRef]
- Nigro, M.; Sánchez-Moreno, I.; Benito-Arenas, R.; Valino, A.L.; Iribarren, A.M.; Veiga, N.; García-Junceda, E.; Lewkowicz, E.S. Synthesis of Chiral Acyclic Pyrimidine Nucleoside Analogues from DHAP-Dependent Aldolases. Biomolecules 2024, 14, 1–24. [Google Scholar] [CrossRef]
- Rosen, T.; Taschner, M.J. Synthetic and Biological Studies of Compactin and Related Compounds. 2. Synthesis of the Lactone Moiety of Compactin. J. Org. Chem. 1984, 21, 3994–4003. [Google Scholar] [CrossRef]
Figure 1.
Some U.S. Food & Drug Administration (FDA) approved statins.
Scheme 1.
Sequential aldol addition by DERA.
Scheme 2.
Chemoenzymatic approach to nucleobase containing lactol precursor of potential statins.
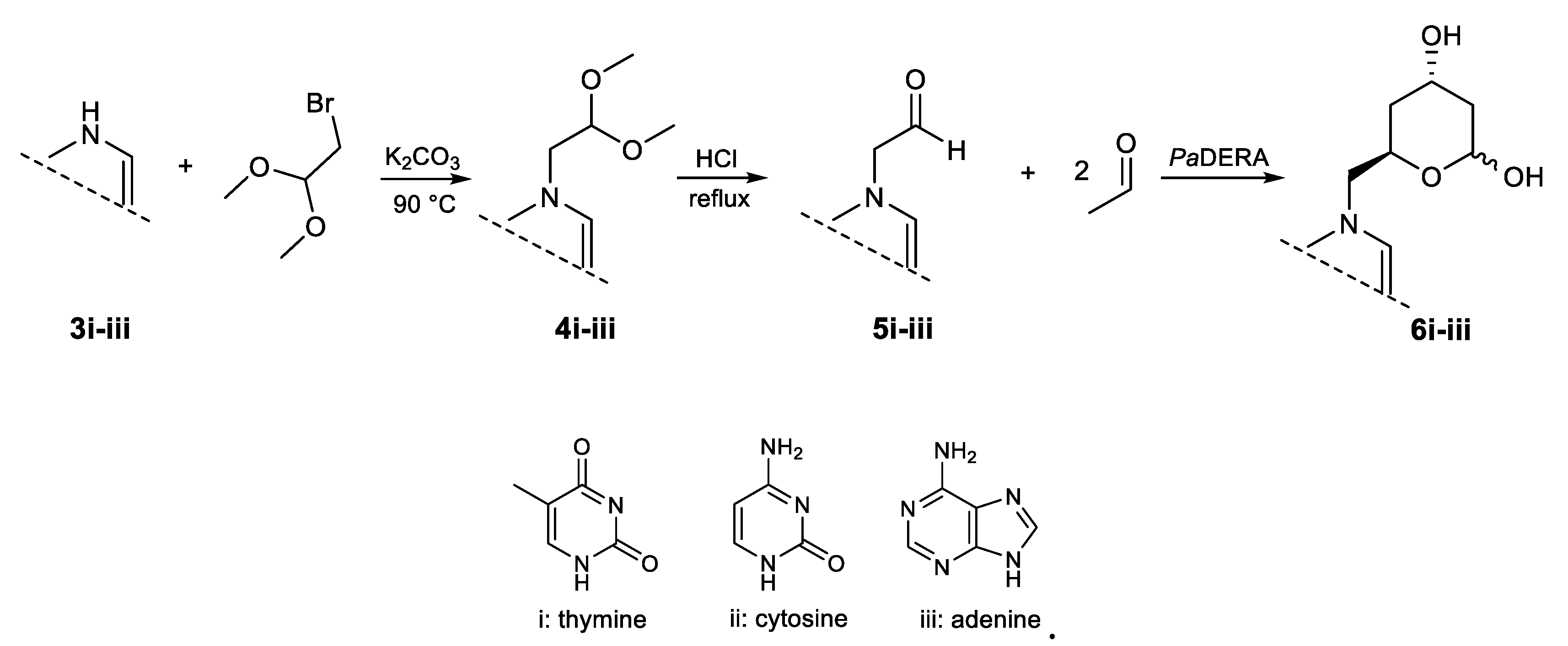
Figure 2.
Partial view of the 1H NMR spectrum of 6i obtained by PaDERA C49M biocatalyzed aldol addition of 5i and acetaldehyde as substrates. The inset shows C1 hydrogen signals of the anomers.
Figure 2.
Partial view of the 1H NMR spectrum of 6i obtained by PaDERA C49M biocatalyzed aldol addition of 5i and acetaldehyde as substrates. The inset shows C1 hydrogen signals of the anomers.
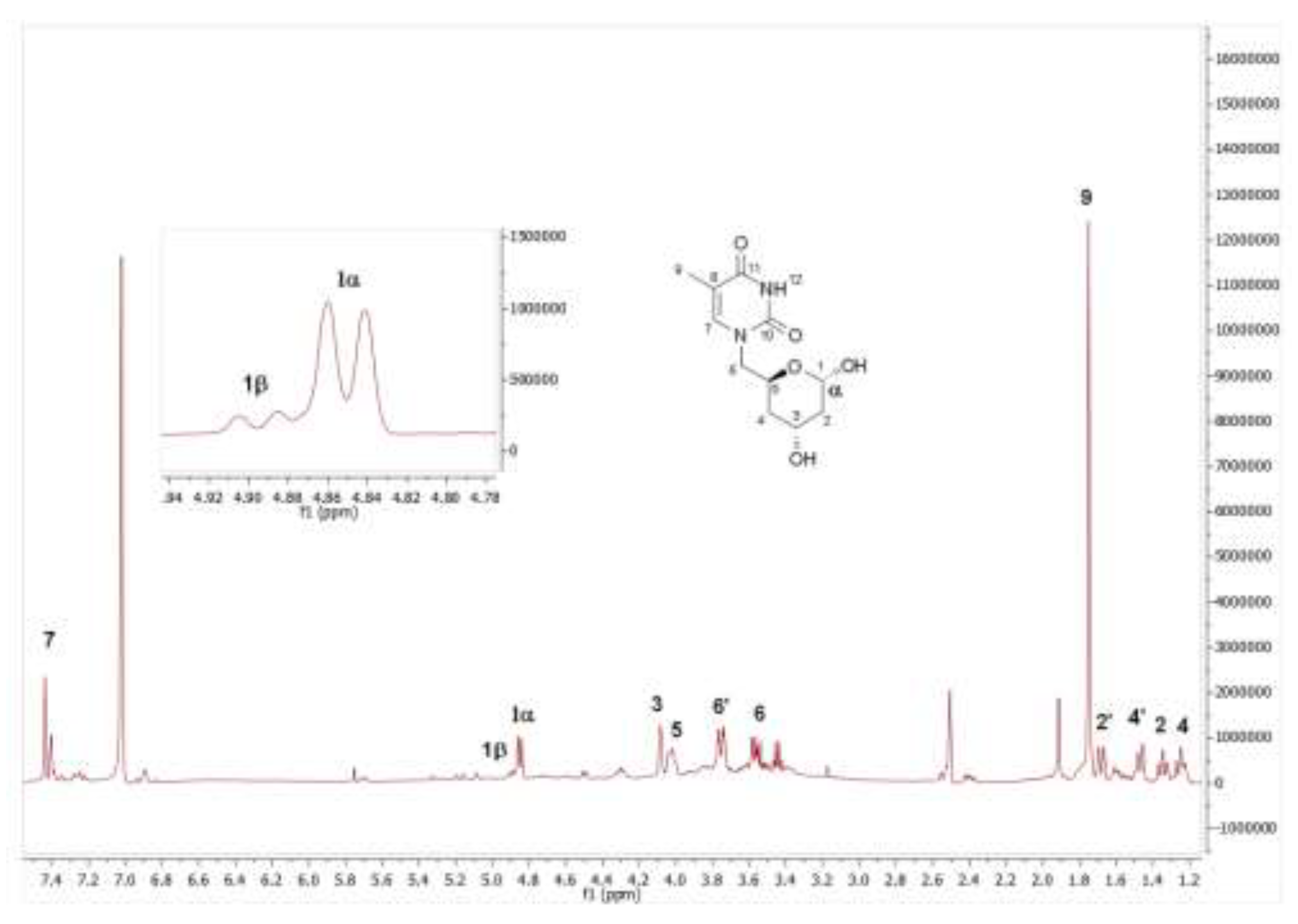

Table 1.
6-substituted lactols synthesized by E. coli (PaDERA C49M)a.
 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



