
Submitted:
22 January 2026
Posted:
22 January 2026
You are already at the latest version
Abstract
Background: Collagen XIα1, encoded by the COL11A1 gene, is a minor fibrillar collagen that is overexpressed in various human cancers, in which its presence correlates with tumor aggressiveness and progression. Methods: In this study, we developed two novel mouse monoclonal antibodies (mAbs), Anti-colXIα1 clone 3 and Anti-colXIα1 clone 9, that target the putative C-telopeptide of human collagen XIα1. The antibodies were raised to the RRHTEGMQA sequence, a unique nine-amino acid stretch within the putative C-telopeptide of human collagen XIα1. Results: Corresponding to nearly identical V(D)J gene segments and complementarity-determining regions (CDRs), the antibodies specifically bound the RRHTEGMQA epitope in ELISAs but did not react with the C-propeptide. This specificity was further confirmed with the purified Anti-colXIα1 clone 9 mAb, which demonstrated strong reactivity to recombinant proteins containing the RRHTEGMQA sequence in both ELISAs and Western blot assays. This sequence seems to behave as a linear B-cell neoepitope, in which the RRHT motif is crucial for epitope recognition. Otherwise, no immunodetections were observed either in cultures and lysates from the COL11A1-highly expressing A204 human cell line or on tissue sections from specimens of human pancreatic ductal adenocarcinoma (PDAC), with strong desmoplastic reactions, Conclusions: Lacking a precise knowledge of the characteristics of the putative C-telopeptide of human collagen XIα1, these antibodies could enhance our understanding of the processing of human procollagen XIα1 and contribute to a better characterization of the tumor microenvironment of COL11A1-expressing cancers.
Keywords:
mouse monoclonal antibodies
; COL11A1
; human collagen XIα1
; putative C-telopeptide
1. Introduction
The COL11A1 human gene codes for the α1 chain of human procollagen XI and mature collagen XI, an extracellular minor fibrillar collagen. (Pro)collagen XIα1 is highly synthesized in diverse human cancers, and this expression is correlated with tumor aggressiveness and progression [1,2,3,4,5,6]. Together with the INHBA and THBS2 genes, the COL11A1 gene was initially correlated with a multicancer invasion- and metastasis-associated gene expression signature [1]. Later on, this gene was also identified as a member of a pancancer signature present in late-stage aggressive cancers [4]. As components of the extracellular matrix, collagens remodel the tumor microenvironment. Matrix stiffening and collagen fiber alignment, in which collagen XIα1 is thought to be involved, have been shown to promote cancer cell migration [5]. Conversely, knockdown of the COL11A1 gene significantly diminishes the invasive potential of cancer cells [6].
COL11A1/(pro)collagen XIα1 is mainly expressed by a subset of myofibroblastic cancer-associated fibroblasts (CAFs), which are also specifically characterized by their surface expression of Leucine-Rich Repeat-Containing 15 (LRRC15) protein and integrin alpha-11 (ITGA11) as well as their secretion of Matrix Metalloproteinase-11 (MMP-11, stromelysin 3). These COL11A1+ myofibroblasts are prominent in the desmoplastic reaction of human invasive carcinomas [7,8,9,10,11].
Some well-established human cancer cell lines also express high levels of COL11A1 mRNA/(pro)collagen XIα1 [12] and are useful as models to track the expression and biological significance of this extracellular matrix component.
The canonical sequence of the P12107-1 Name A isoform of human procollagen XIα1 consists of 1806 amino acid residues and has an estimated molecular weight of 181 kDa [13]. Based on comparison with other better-characterized collagens, this procollagen, upon secretion, is expected to be processed by extracellular proteinases excising the N- and C-terminal propeptides [14]. The 21 C-terminal residues of the processed mature extracellular collagen XIα1 -(1543) IQPLPILSSKKTRRHTEGMQA (1563)- would then comprise the putative nonhelical C-telopeptide [13].
Monoclonal antibodies to the C-telopeptide of human collagen XIα1 could help shed light on the actual processing of human procollagen XIα1 and lead to a better characterization of COL11A1-expressing tumors and the extracellular matrix microenvironment. In addition, the tuned derivatives of these antibodies, naked, as antibody–drug conjugates (ADCs) or as components of targeted immunovesicles (liposomes, exosomes)/immunonanoparticles, could be employed as therapeutic weapons to combat the progression of COL11A1-expressing tumors. However, to date, no antibodies have been generated to this region.
In this study, we developed two novel mouse monoclonal antibodies (mAbs)—Anti-colXIα1 clone 3 and Anti-colXIα1 clone 9—to the putative C-telopeptide of human collagen XIα1 and assessed their reactivity and genetic features.
2. Materials and Methods
2.1. Human Collagen XIα1 and C-Propeptide Recombinant Forms
The putative extracellular processed form of human collagen XIα1 (Figure 1), with an added 6xHis head, recombinantly expressed in CHO cells, was provided by GenScript (Piscataway, NJ 08854, USA). It has a theoretical molecular weight of 98.37 kDa.
Similarly, the putative C-propeptide, of 243 amino acid residues (Figure 2), with a theoretical molecular weight of 27.45 kDa, was also recombinantly expressed in CHO cells using GenScript.
The COL11A1 Fusion Protein, with an N-terminal GST tag, from ChromoTek GmbH & Proteintech, Germany (Cat. No. Ag16509), was recombinantly expressed in Escherichia coli. After removal of the GST tag (Ag37791), the first 19 N-terminal amino acid residues PLPILSSKKTRRHTEGMQA corresponded to the putative C-telopeptide (Figure 3). This product has a theoretical molecular weight of 29.6 kDa.
2.2. Mouse Immunization and Hybridoma Generation
Peptide conjugates KLH-Cys-GG-RRHTEGMQA and KLH-Cys-GG-EGMQADADD, corresponding to different amino acid stretches of the human procollagen XIα1 around the C-telopeptide, were provided by Abyntek Biopharma S.L. (Zamudio, Bizkaia, Spain). Cysteine and glycine residues were added to ensure correct linking to the carrier protein, as previously conducted [15]. Upon reconstitution in sterile water B Braun and in saline for injections, an emulsion was made with the Sigma Adjuvant System (Sigma-Aldrich, Cat. No. S6322) for immunization purposes, following the manufacturer’s instructions.
Female 6-week-old DBA/1J mice were obtained from The Jackson Laboratory through Charles River Laboratories S.A., Spain, and female 4-week-old BALB/c mice were obtained from Charles River Laboratories S.A., Spain. The mice were housed in the facilities of the Bioterio of the University of Oviedo under conventional conditions. They were handled following the Guidelines of the Experimental Animal Ethics Committee of the University of Oviedo and after the approval of animal experimental procedures by the Government of the Principality of Asturias, Spain, Resolutions PROAE 62/2019 and PROAE 4/2022.
Mice were i.p. injected with 200 µg of conjugate/500 µl of emulsion, at least 15 days apart. Once hyperimmunized, for hybridoma generation, they were euthanized via cervical dislocation. Spleens were aseptically removed and gently homogenized for fusion with Sp2/0 mouse myeloma cells using standard methods [16].
Hybridoma supernatants, in RPMI Medium/Hypoxanthine–Aminopterin–Thymidine (HAT)/20% Fetal Bovine Serum (FBS), were primarily screened using an indirect ELISA against KLH, mechanical lysates from A204 and A549 cell cultures, and the human collagen XIα1 recombinant form (Figure 1).
A 1:1000 dilution of anti-mouse γ-chain-HRPO (Sigma-Aldrich, Cat, No. A3673) in 10 mM Phosphate-Buffered Saline (PBS)–1% Bovine Serum Albumin (BSA)–0.1% Tween 20 was applied to identify mouse IgG-containing supernatants.
The selected antibody-secreting hybridomas were subcloned twice to ensure antibody monoclonality. Antibody subisotyping was conducted by means of the IsoStrip™ Mouse Monoclonal Antibody Isotyping Kit (11493027001 Roche).
For purified antibody preparations, subclones were progressively adapted to grow in RPMI Medium–20% FBS, subsequently in protein-free and serum-free CD Hybridoma Gibco™ Medium (ThermoFisher Scientific, Cat. No. 11279023)–20% FBS, and finally weaned from FBS.
Cultures were assessed to be free of Mycoplasma by means of the MycoStrip™ Mycoplasma Detection Kit (InvivoGen, Cat. No. rep-mys-10), following the manufacturer´s instructions.
Monoclonal antibodies were purified using an Amersham Biosciences ÄKTA Purifier 10 FPLC System w/ UPC-900 and 1 mL MabCaptureC™ MiniChrom Columns (ThermoFisher Scientific, Cat. No. 5943662001), following the manufacturer´s instructions; dialyzed against 10 mM of PBS, pH 7.4; concentrated via centrifugation in 10K concentrators; and finally filtered through sterile 0.25 μm filters.
2.3. Isotype-Matched Negative Control mAb
Anti-pneumolysin IgG1, kappa PLY-7 mAb [17] served as the isotype-matched negative control.
2.4. Cell Cultures and Lysates
Human rhabdomyosarcoma A204 (no. HTB-82), large cell lung carcinoma NCI-H661 (no. HTB-183), and alveolar lung carcinoma A549 (no. CCL-185) cell lines were obtained from the American Type Culture Collection (ATCC) and cultured in DMEM supplemented with sodium pyruvate, L-glutamine, non-essential amino acids, 10% FBS, ascorbate 2-phosphate (37.5 µg/mL) (Sigma-Aldrich, Cat. No. A8960), and 10 ng/mL of recombinant TGF-β1 (PeproTech, Cat. No. 100-21C), in a humidified atmosphere of 5% CO2 in air at 37 °C.
Cells were cultured for at least 15 days, in T-25 and T-75 flasks (Sarstedt, Cat. No. 83.3910.302 and 83.3911.302, respectively). Passages and cell collections were conducted using trypsinization. Different harvests from each cell culture condition were pooled.
For immunocytochemistry (ICC), cells were cultured in 4-well culture slides (BD Falcon™, ref. 354114 or Nunc™ Lab-Tek™ II CC2™ (ThermoFisher Scientific, Cat. No. 154917PK or Sarstedt, Cat. No. 94.6170.402) for three–five days.
To generate cell lysates, after the removal of spent media, the flasks were washed with 10 mM of cold PBS–0.1% sodium azide (PBS-SA). Cell cultures were then vigorously collected in cold PBS-SA or a hot SDS-PAGE sample buffer with the help of cell scrapers. Cell lysates were frozen and thawed several times.
2.5. ELISA for Antibody Reactivity Screening and Assessment
Flat-bottom 96-well MaxiSorp Nunc-Immuno plates (ThermoFisher Scientific, Cat. No. 439454) were coated with 1 µg/100 µl well of the human collagen XIα1 recombinant form, the human collagen XIα1 C-propeptide recombinant form, the COL11A1 Fusion Protein with no GST tag (Ag37791), the peptide conjugates used as immunogens, or with 10 µg/100 µl well of mechanical cell lysates, in PBS-SA, pH 7.3, for 6 h at 37 ºC, and they were then blocked with 200 µl/well of PBS-1% BSA-SA for 1 h at 37 ºC and left overnight at 4 ºC. After being washed, the wells were successively incubated with 100 µl/well of crude hybridoma supernatants or with 1 µg/100 µl well of purified monoclonal antibody in PBS-1% BSA-SA (2 h at 37º C); 100 µl well of a 1:1000 dilution of anti-mouse γ-chain-HRPO (Sigma-Aldrich, Cat, No. A3673) in PBS-1% BSA-0.1% Tween 20 (1 h at 37 ºC); and 100 µl well of ready-to-use supersensitive TMB solution for the ELISA (Sigma-Aldrich, Cat, No. T4444) (5 min at 37 ºC). Color development was stopped by adding 100 µl well of 2 M H2SO4. Plates were read at 450 nm on a Synergy LX Microplate Reader (BioTek Instruments, Inc., Winooski, VT, USA). All determinations were made in triplicate and averaged. Data were analyzed using the BioTek Gen5 software. Optical density data were blanked by subtracting blanks (culture media or PBS-1% BSA-SA) from the readings.
Plates were washed three times with 200 µl/well of PBS–0.1% Tween-20 between incubations.
2.6. ELISA Competition Tests with Soluble Peptides
ELISA competition tests with soluble peptides were performed essentially as already described [18]. Immunograde, ≥ 95% pure, synthetic N-acetylated RRHTEGMQA, EGMQADADD, and (450)KRTISIWGT(458) from pneumolysin -as a scrambled control- peptides were supplied by Abyntek Biopharma S.L. (Zamudio, Bizkaia, Spain).
ELISA plates were coated with the human collagen XIα1 recombinant form, as above. The reactivity of each supernatant was titered in ELISAs; an assessed dilution of each supernatant, which rendered a reliable absorbance signal, was chosen as a positive control.
For competition tests, the supernatant dilutions were mixed, at equal volumes, with 1:10 serial dilutions of the synthetic peptide in PBS-1% BSA-SA, and preincubated for 2 h at 37 ºC and overnight at 4 ºC. The next day, the mixtures were then added to the ELISA coated wells and incubated for 2 h at 37 ºC. The rest of the ELISA continued as described above. No peptide was added in the blanks. The inhibitory activity of any blocking peptide sample was estimated from its color reading in relation to the absorbance of the positive controls.
2.7. SDS-PAGE and Western Blot Assays
Human Immunization-Grade Type XI Collagen was purchased from Chondrex (Catalog # 1085).
Purified protein concentrations were estimated using absorbance at 280 nm with a UV-1280 UV-VIS Spectrophotometer (Shimadzu).
A total of 15 μg/lane of recombinant protein or 50 μg/lane or 40 μl/lane of whole cell lysates and 10 µl/lane of PageRuler™ Plus Prestained Protein Ladder, 10 to 250 kDa (ThermoFisher Scientific, Cat. No. 26619), was subjected to 12, 10, or 6% polyacrylamide SDS-PAGE under reducing conditions and, subsequently, electrotransferred onto an Immobilon®-NC Transfer Membrane, with a 0.45 µm pore size (Sigma-Aldrich, Cat, No. HATF00010), using the Mini-PROTEAN® Tetra Cell (2-gel) and Tetra Blotting Module (BIO-RAD).
As already stated, some cell lysates were also generated by scraping cell cultures in the presence of hot sample buffer.
After blocking overnight at 4 ˚C in PBS-3% BSA-SA, the membranes were then probed with 10 mL of crude hybridoma supernatants or with 10 µg of purified mAb in 10 mL of PBS-1% BSA-SA with gentle rocking for 2 h at room temperature. After several washing steps with PBS–0.1% Tween-20 and PBS, the blots were incubated with 10 mL of a 1:4000 dilution of anti-mouse γ chain-specific HRPO conjugated (Sigma-Aldrich, Cat, No. A3673) in PBS–1% BSA–0.1% Tween-20, for 2 h as above, and finally developed with 1-Step™ Ultra TMB Blotting Solution (ThermoFisher Scientific, Cat. No. 37574).
2.8. PEP-FOLD4 Peptide Structure Predictions
Peptide sequences were submitted to the PEP-FOLD4 on-line server under the current default conditions [19].
2.9. V Gene Sequencing of Hybridomas
Sequencing of the whole heavy and light chains of mAbs was performed using whole transcriptome shotgun sequencing (RNA-Seq), Absolute Antibody Ltd., United Kingdom.
Nucleotide sequences were analyzed using IMGT/V-QUEST (http://imgt.cines.fr) [20].
2.10. Immunocytochemistry (ICC)
ICC procedures were performed by personnel of the Molecular Histopathology Unit in Animal Models of Cancer (IUOPA).
Cells were fixed in 4 % formaldehyde for 15 min in the chamber slide. Primary antibodies were applied, as described in Table 1
The primary antibody was omitted in the negative controls. Subsequently, slides were incubated with the EnVision FLEX HRP (DAKO K8000) system for 30 min at room temperature. The samples were then visualized with diaminobenzidine (DAB) for 30 sec. Finally, the immunostained slides were dehydrated, mounted, studied under an Olympus BX61 Automatic Microscope, and photographed using the DP Controller/Manager software, with the assistance of personnel of the Photonic Microscopy and Image Processing Unit, of the Scientific and Technical Services (SCTs) of the Universidad de Oviedo.
2.11. Immunohistochemistry (IHC)
For immunohistochemical techniques, specimens of pancreatic ductal adenocarcinoma (PDAC), with strong desmoplastic reactions, and normal pancreas counterparts from the same patients were provided by the Principality of Asturias BioBank (PT23/077). The specimens were processed following standard operating procedures with the appropriate approval of the Ethical and Scientific Committees (Comité de Ética de la Investigación con Medicamentos del Principado de Asturias, reference CEImPA 2022.462).
IHC procedures were performed by personnel of the BioBank. The DAKO Autostainer system was used.
The samples were fixed with 10% formaldehyde for 24 h and embedded in paraffin.
Tissue sections (3 µm) were deparaffinized, rehydrated, and subjected to epitope retrieval using heat induction (HIER) at 95 ºC for 20 min, at pH 9 and pH 6 (Agilent-DAKO), in the Pre-Treatment Module, PT-LINK (DAKO). Endogenous peroxidase activity was blocked with the EnVision™ FLEX Peroxidase-Blocking Reagent (DM821) for 5 min. Subsequently, sections were first incubated with Anti-colXIα1 clone 9 and PLY-7 (dilution 1:100 from 1 mg/mL, both) for 30 min. Subsequently, the Dako EnVision® + Dual Link System-HRP (Agilent-DAKO) was applied. The samples were then stained using DAB chromogen as a substrate in Dako EnVision™ FLEX/HRP (Agilent-DAKO), counterstained with hematoxylin, dehydrated, and then mounted with permanent medium (Agilent-DAKO Mounting Medium, CS703). The sections were studied under a light microscope (Nikon - ECLIPSE Ci).
The negative controls were processed by omitting the primary antibody
3. Results
3.1. mAb Reactivity
Upon immunization with the two designed immunogens, KLH-Cys-GG-RRHTEGMQA and KLH-Cys-GG- EGMQADADD, we sought to generate mAbs with the following abilities: besides interfering with proteinases, mAbs targeting the RRHTEGMQA sequence that could recognize the extracellular C-telopeptide and those targeting the EGMQADADD sequence that could even recognize procollagen before its processing. However, we were only successful by rescuing hybridomas from BALB/c mice after immunization with the KLH-Cys-GG-RRHTEGMQA immunogen.
The two mAbs described herein were generated from the same BALB/c mouse; both are IgG1 kappa subtype.
Table 2 shows the immunoreactivity of initial crude supernatants, in RPMI-HAT-20% FBS, from the two hybridomas against the components of the conjugates which were used as immunogens.
According to these results, the Anti-colXIα1 clone 3 mAb seems to exclusively recognize the KLH-C-GG-RRHTEGMQA conjugate, while the Anti-colXIα1 clone 9 mAb seems to also recognize the KLH-C-GG-EGMQADADD sequence, although with lower intensity.
Table 3 shows some other reactivity characteristics against different antigen preparations.
In the ELISA, the Anti-colXIα1 clone 3 and Anti-colXIα1 clone 9 supernatants showed strong reactivity against the recombinant human collagen XIα1 but not against the recombinant C-propeptide or with cell lysates from the A204 and A549 cell lines. The human rhabdomyosarcoma A204 cell line, which expresses the highest recorded levels of COL11A1 mRNA [12], was used as a positive control, while alveolar lung carcinoma A549 cells expressing very low levels of COL11A1 mRNA served as a quasi-negative control. All the Western blots indicated in Table 3 were negative.
To confirm the fine specificity of the two mAbs, blocking assays with soluble free N-acetylated peptides were performed. As shown in Figure 4, there was a clear concentration-dependent inhibition of the recognition of collagen XIα1 by the RRHTEGMQA peptide but not by the EGMQADADD peptide or by the KRTISIWGT scrambled control peptide.
These results show that both the Anti-colXIα1 clone 3 and Anti-colXIα1 clone 9 supernatants specifically recognize the RRHTEGMQA sequence of the human collagen XIα1 putative C-telopeptide. The lack of inhibition of this recognition by the EGMQADADD peptide points to a prominent role of the RRHT amino acid residues in the recognition of the epitope by these two mAbs.
As the genetic characterization of these two clones showed that they were almost identical (see below), the following observations were made only with the Anti-colXIα1 clone 9 mAb.
The recombinant COL11A1 Fusion Protein from Proteintech was also assayed in an ELISA and Western blot with already finally purified preparations of the Anti-colXIα1 clone 9 and PLY-7 mAbs. Table 4 shows their ELISA immunoreactivity characteristics. The Anti-colXIα1 clone 9 mAb was reactive with collagen XIα1 and, to a lesser extent, with the COL11A1 Fusion Protein but not with the C-propeptide, which lacks the RRHTEGMQA sequence and served as a negative control.
In Western blots, from 12 % SDS-PAGE gels and 15 µg/lane of purified proteins, as shown in Figure 5, the anti-ColXIa1 clone 9 mAb recognized recombinant collagen XIa1 protein (containing degradation products), as well as the COL11A1 Fusion Protein, but not the C-propeptide (see also Supplementary Material S1).
These results confirm that the Anti-colXIα1 clone 9 mAb specifically recognizes, by both ELISA and Western blot, the RRHTEGMQA amino acid stretch which is contained in the sequence of the putative C-telopeptide of human collagen XIα1.
3.2. Peptide Structure Predictions
To gain more insight into the nature of the epitope recognized by the Anti-colXIα1 clone 9 mAb, the structures of peptide sequences corresponding to some of the assayed antigens were predicted using the PEP-FOLD4 server.
Figure 6A shows the structural prediction of the 50 N-terminal amino acid sequence PLPILSSKKTRRHTEGMQADADDNILDYSDGMEEIFGSLNSLKQDIEHMK of the COL11A1 Fusion Protein from Proteintech, with the first 19 N-terminal PLPILSSKKTRRHTEGMQA amino acid residues of the putative C-telopeptide. In this model, the RRH amino acid residues are part of a disordered stretch preceding an α-helix. The prediction for the free RRHTEGMQA peptide is a frank α-helix (Panel B). The structural prediction (Panel C) for the C-terminal RRHTEGMQA stretch is very similar to the one in Panel A above for the last 50 amino acid sequence NKGSTGPAGQKGDSGLPGPPGSPGPPGEVIQPLPILSSKKTRRHTEGMQA of the GenScript´s recombinant collagen XIα1 form, whose last 21 C-terminal IQPLPILSSKKTRRHTEGMQA amino acid residues correspond to the putative C-telopeptide. Thus, according to these predictions and the abovementioned ELISA and Western blot analyses, the epitope recognized by the Anti-colXIα1 clone 9 mAb behaves like a linear epitope, with essential recognition of the RRHT amino acid residues.
3.3. Genetic Characterization of the Two mAbs
Sequencing of the whole heavy and light chains of the two mAbs confirmed that both were IgG1, kappa.
Table 5 shows the most probable usage of germinal V(D)J gene segments by the two mAbs according to the IMGT/V-QUEST (https://www.imgt.org/IMGTindex/V-QUEST.php) analysis. The VH and VL domains of the two mAbs were most likely made from the same germinal gene segments.
Table 6 shows the amino acid sequences of the CDRs of the two mAbs, as identified by IMGT/V-QUEST. With very limited nucleotide differences, the CDRs of the VH domains of the two mAbs are almost identical, as well as the CDRs of their VL domains, which explains their very similar monoclonal reactivity.
When the sequences of these CDRs were compared to sequence databases, some individual CDRs were found to be present in different immunoglobulins, but neither the combination of the three CDRs of each V domain nor the combination of the six CDRs of each of the two mAbs were identified in any immunoglobulin reported so far. Thus, this may affirm that they are two novel mAbs.
For patent purposes, in accordance with The Budapest Treaty of 1977, the Anti-colXIα1 clone 3 and Anti-colXIα1 clone 9 mouse hybridomas were deposited in the European Collection of Authenticated Cell Cultures (ECACC) repository with accession numbers 23112901 and 23112902, respectively, given by the International Depositary Authority. They were determined to be free of mycoplasma contamination.
The VH and VL nucleotide sequences of the Anti-colXIα1 clone 3 and Anti-colXIα1 clone 9 mAbs were deposited in the GenBank Nucleotide Sequence Database [21] with accession numbers PP150425 and PP150426 and PP150423 and PP150424, respectively.
3.4. Immunocytochemistry (ICC) and Immunohistochemistry (IHC) Analyses
Immunocytochemistry and immunohistochemistry analyses were performed with the finally purified preparations of the Anti-colXIα1 clone 9 and PLY-7 mAbs.
As shown in Figure 7, the Anti-colXIα1 clone 9 mAb developed a rather faint immunostaining in the well-known COL11A1-positive A204 and NCI-H661 cell lines but not in the COL11A1-quasi-negative A549 cell line. These three cell lines are known to express the cytoskeletal intermediate filament vimentin; their anti-vimentin staining served as an ICC positive control.
No clear immunostaining patterns were observed in any of the IHC pancreatic ductal adenocarcinoma (PDAC) specimens studied.
4. Discussion
The molecular biology underlying the expression of human procollagen XIα1 is far from being understood. In particular, the function and structural characteristics of (pro)collagen XIα1 have been studied in tendons, vitreous humor, and cartilages from mice, rats, cows, and chickens, among other origins and body locations [22,23], but not in humans. No detailed molecular studies have been conducted on human cancer either. A purified recombinant form of human procollagen XIα1 is not available, and no study has been carried out on its C-terminal enzymatic processing by furin or BMP-1, as in the case of human procollagen Vα1 [24].
Based on similarity with other collagens, it is assumed that the terminal C-telopeptide of the mature human collagen XIα1 corresponds to the (1543) IQPLPILSSKKTRRHTEGMQA (1563) amino acid sequence of procollagen XIα1. According to diverse studies, the C-telopeptide would be involved in intrafibrillar cross-linking with triple-helical regions of adjacent molecules. Minor collagen XIα1 would act as a nucleator in the assembling of extracellular collagen fibrils [14,22,23], and being massively surrounded by major I, II, and III fibrillar collagens [14], this C-telopeptide could remain buried and not exposed on the surface of fibrils, thus hindering its recognition by antibodies. Nevertheless, it could be exposed at the time of enzymatic cleavage by extracellular proteinases and of further processing steps.
In the case of human collagens Iα1, IIα1, and IIIα1, soluble C-terminal cross-linked telopeptides (CTXs) are released and may be detected in biological fluids for diagnostic purposes [25,26,27,28] and serve as biomarkers of bone resorption or osteoarthritis. Different antibody preparations, used in immunoassays, are available for detecting and measuring these CTXs and for the immunostaining of extracellular collagens [29,30].
The aim of this study was to rescue and partially characterize two mAbs—Anti-colXIα1 clone 3 and Anti-colXIα1 clone 9— raised to the putative C-telopeptide amino acid stretch of the mature human collagen XIα1. They recognize the RRHTEGMQA amino acid sequence but not the EGMQADADD sequence, which includes the DADD amino acids of the N-terminus of the C-propeptide, or the recombinant C-propeptide form. In humans, this RRHTEGMQA nine-amino acid sequence is only present as such in (pro)collagen XIα1 and partially found in some other unrelated proteins.
These two mAbs used the same set of V, D, and J gene segments for the confection of their V domains, and their CDRs are almost identical. Their reactivity is also very similar.
Based on structural predictions and on the results obtained from the ELISA and Western blot assay, the Anti-colXIα1 clone 9 mAb recognizes a linear epitope within the C-telopeptide, in which the RRHT amino acid residues are central to mAb recognition .
Altogether, bearing in mind the low-intensity immunostaining of cultured cells by the Anti-colXIα1 clone 9 mAb, in comparison with its reactivity with the free collagen XIα1 recombinant form and in agreement with the putative C-terminal processing of procollagen XIα1, the RRHTEGMQA epitope sequence should be mostly envisaged as a linear B-cell neoepitope as it is not identified as such in procollagen.
A plethora of antibodies reactive with diverse collagens has been reported and described. A few recognize B-cell neoepitopes, which become exposed after enzymatic processing [31,32,33,34,35,36]. Thus, the epitope recognized by the Anti-colXIα1 clone 9 mAb may be added to the list of collagen B-cell neoepitopes.
On tissue sections from human PDAC samples, the purified Anti-colXIα1 clone 9 mAb did not stain fibroblast-like cells associated with carcinoma cell niches, as the 1E8.33 mAb does [37]; additionally, no staining of extracellular matrix components was observed in A204 cell cultures. Thus, these observations seem to corroborate the suggested buried status of the C-telopeptide of human collagen XIα1 in the extracellular matrix of the tumor microenvironment; alternatively, the conformation assumed by the RRHTEGMQA stretch in vivo is not recognized by the mAbs.
A human collagen Iα2 C-telopeptide binding antibody has been shown to reduce the rate of cleavage of the C-terminal propeptide by BMP-1 and to limit extracellular fibril formation both in vitro and in organo-typic keloid-like constructs [31,38]. Blocking antibodies to secretory components of myofibroblastic CAFs have been shown to affect carcinoma progression. For instance, an antibody to the microfibrillar-associated protein 5 (MFAP5) suppressed tumor growth in vivo, with a concurrent reduction in the expression of the COL11A1 gene [39]. Similarly, an antibody to MMP-11 showed significant anti-tumoral effects in animal models [40], as did a humanized antibody to secretory Collagen triple helix repeat containing-1 (CTHRC1) protein [41]. Therefore, antibodies to the C-telopeptide of the mature human collagen XIα1, by interfering with (pro)collagen processing steps, could show a progression restraining capacity of tumors expressing the COL11A1 gene, thus pointing to therapeutic potential as well.
One main limitation of this study is that there was not available a purified recombinant form of human procollagen XIα1 and that it was not assayed its experimental enzymatic processing. Therefore, it remains to be precisely determined the actual nature and characteristics of the putative C-telopeptide of human collagen XIα1 and if our mAbs do actually recognize the product(s) resulting from in vitro and in vivo processing.
5. Conclusions
To our knowledge, this is the first time that highly specific mAbs to a unique B-cell neoepitope of the putative C-telopeptide of human collagen XIα1 have been generated. These mAbs may be of help to researchers in analyzing the cell biology of human (pro)collagen XIα1 and for a better characterization of COL11A1-positive tumors, and, if able to interfere with COL11A1-expressing stromal and cancer cells, they could have in vivo therapeutic potential as well
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization; JRT, LJG-F, Data curation: JRT, Formal analysis: MG-O, LL-O, SR-R, JRT, IF-V, Funding acquisition: ML-M, LJG-F, Investigation: MG-O, LL-O, SR-R, JRT, PR-F, Methodology; MG-O, LL-O, SR-R, JRT, IF-V, Project administration: LJG-F, Resources: MG-O, IF-V, LJG-F, Software; JRT, Supervision: JRT, ML-M, LJG-F, Validation: MG-O, LL-O, SR-R, JRT, IF-V, Visualization: MG-O, LL-O, SR-R, PR-F, IF-V, ML-M, JRT, LJG-F, Writing—original draft preparation: JRT, Writing—review and editing: JRT. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partially financed by Startquake, S.L. (ETERNA Diagnostics), Spain and Proyecto IDI/2021/000282, FICYT, Principality of Asturias, Spain, provided to Luis J. García-Flórez.
Institutional Review Board Statement
This study was conducted in accordance with the Declaration of Helsinki and approved by the Ethical and Scientific Committees of the Principality of Asturias, Project no 42/12 and reference CEImPA 2022.462 (February 6, 2023). Human specimens, currently obtained after informed consent to participate, were from the Principality of Asturias BioBank (PT23/077). Mice were handled following the Guidelines of the Experimental Animal Ethics Committee of the University of Oviedo and after the approval of animal experimental procedures by the Government of the Principality of Asturias, Spain, Resolutions PROAE 62/2019 (December 19, 2019) and PROAE 4/2022 (March 21, 2022).
Data Availability Statement
The VH and VL nucleotide sequences of the Anti-colXIα1 clone 3 and Anti-colXIα1 clone 9 mAbs were deposited in the GenBank Nucleotide Sequence Database [21] with accession numbers PP150425 and PP150426 and PP150423 and PP150424, respectively. Anti-colXIα1 clone 9 and PLY-7 mAbs may be sent upon request, depending on the available stocks.
Acknowledgments
The authors greatly appreciate the excellent technical assistance of María Álvarez-García, Ana M. López-Fernández and Lucía Ronderos-López (Molecular Histopathology Unit in Animal Models of Cancer, IUOPA); Marta M. Alonso-Guervós, . (Photonic Microscopy and Image Processing Unit, of the Scientific and Technical Services [SCTs]) of the Universidad de Oviedo); and M. Daniela Corte-Torres (Biobank of the Principality of Asturias, ISPA-FINBA). We want to particularly acknowledge the patients and the Biobank of the Principality of Asturias (BioPA]) (National Registry of Biobanks B.0000827) (PT23/00077 funded by ISCIII and co-funded by the European Union) integrated into the Platform ISCIII Biobanks and Biomodels for their collaboration.
Conflicts of Interest
Marcos Ladreda-Mochales is a shareholder of Startquake, S.L.
Abbreviations
The following abbreviations are used in this manuscript:
| ADCs | Antibody–Drug Conjugates |
| ATCC | American Type Culture Collection |
| BMP-1 | Bone Morphogenetic Protein-1 |
| BSA | Bovine Serum Albumin |
| CAFs | Cancer-Associated Fibroblasts |
| CDRs | Complentarity-Determining Regions |
| CHO cells | Chinese Hamster Ovary cells |
| COL11A1 | Collagen Type XI Alpha 1 Chain gene |
| ColXIα1 | Collagen XIα1 |
| CTHRC1 | Collagen Triple Helix Repeat Containing-1 |
| CTXs | Cross-linked Telopeptides |
| DAB | DiAminoBenzidine |
| ELISA | Enzyme-Linked ImmunoSorbent Assay |
| ECACC | European Collection of Authenticated Cell Cultures |
| FBS | Fetal Bovine Serum |
| GST | Glutathione S-Transferase |
| HAT | Hypoxanthine–Aminopterin–Thymidine |
| HRPO | HorseRadish PerOxidase |
| ICC | ImmunoCytoChemistry |
| IHC | ImmunoHistoChemistry |
| INHBA | Inhibin Subunit Beta A gene |
| ITGA11 | Jntegrin Alpha-11 |
| KLH | Keyhole Limpet Hemocyanin |
| LRRC15 | Leucine-Rich Repeat-Containing 15 |
| mAbs | Mouse Monoclonal Antibodies |
| MFAP5 | MicroFibrillar-Associated Protein 5 |
| MMP-11 | Matrix MetalloProteinase-11 |
| PDAC | Pancreatic Ductal AdenoCarcinoma |
| PBS | Phosphate-Buffered Saline |
| SA | Sodium Azide |
| SDS-PAGE | Sodium Dodecyl Sulfate-PolyAcrylamide Gel Electrophoresis |
| TGF-β1 | Transforming Growth Factor Beta1 |
| THBS2 | Thrombospondin 2 gene |
| TMB | TetraMethylBenzidine |
References
- Kim, H.; Watkinson, J.; Varadan, V.; Anastassiou, D. Multi-cancer computational analysis reveals invasion-associated variant of desmoplastic reaction involving INHBA, THBS2 and COL11A1. BMC Med. Genomics 2010, 3, 51. [Google Scholar] [CrossRef] [PubMed]
- Raglow, Z.; Thomas, S.M. Tumor matrix protein collagen XIα1 in cancer. Cancer Lett. 2015, 357, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Villa, F.; García-Ocaña, M.; Galván, J.A.; García-Martínez, J.; García-Pravia, C.; Menéndez-Rodríguez, P.; González-del Rey, C.; Barneo-Serra, L.; de Los Toyos, J.R. COL11A1/(pro)collagen 11A1 expression is a remarkable biomarker of human invasive carcinoma-associated stromal cells and carcinoma progression. Tumour Biol. 2015, 36, 2213–2222. [Google Scholar] [CrossRef] [PubMed]
- Nallanthighal, S.; Heiserman, J.P.; Cheon, D.J. Collagen Type XI Alpha 1 (COL11A1): A Novel Biomarker and a Key Player in Cancer. Cancers (Basel) 2021, 13, 935. [Google Scholar] [CrossRef] [PubMed]
- Popova, N.V.; Jücker, M. The Functional Role of Extracellular Matrix Proteins in Cancer. Cancers (Basel) 2022, 14, 238. [Google Scholar] [CrossRef]
- Freire, J.; García-Berbel, P.; Caramelo, B.; García-Berbel, L.; Ovejero, V.J.; Cadenas, N.; Azueta, A.; Gómez-Román, J. Usefulness of COL11A1 as a Prognostic Marker of Tumor Infiltration. Biomedicines 2023, 11, 2496. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, C.X.; Müller, S.; Keerthivasan, S.; Koeppen, H.; Hung, J.; Gierke, S.; Breart, B.; Foreman, O.; Bainbridge, T.W.; Castiglioni, A.; et al. Single-Cell RNA Sequencing Reveals Stromal Evolution into LRRC15+ Myofibroblasts as a Determinant of Patient Response to Cancer Immunotherapy. Cancer Discov. 2020, 10, 232–253. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Cai, L.; Cui, C.; de Los Toyos, J.R.; Anastassiou, D. Single-cell analysis reveals the pan-cancer invasiveness-associated transition of adipose-derived stromal cells into COL11A1-expressing cancer-associated fibroblasts. PLOS Comput. Biol. 2021, 17, 34283835. [Google Scholar] [CrossRef]
- Zhang, J.; Lu, S.; Lu, T.; Han, D.; Zhang, K.; Gan, L.; Wu, X.; Li, Y.; Zhao, X.; Li, Z.; et al. Single-cell analysis reveals the COL11A1+ fibroblasts are cancer-specific fibroblasts that promote tumor progression. Front. Pharmacol. 2023, 14, 1121586. [Google Scholar] [CrossRef] [PubMed]
- Thorlacius-Ussing, J.; Jensen, C.; Nissen, N.I.; Cox, T.R.; Kalluri, R.; Karsdal, M.; Willumsen, N. The collagen landscape in cancer: Profiling collagens in tumors and in circulation reveals novel markers of cancer-associated fibroblast subtypes. J. Pathol. 2024, 262, 22–36. [Google Scholar] [CrossRef] [PubMed]
- Nissen, N.I.; Karsdal, M.A.; Cox, T.R.; Willumsen, N. Chapter 38 - The roles of collagens and fibroblasts in cancer, In Biochemistry of Collagens, Laminins and Elastin, 3rd ed., Morten A. Karsdal, Ed. Academic Press, Elsevier B.V.; Amsterdam, The Netherlands, 2024, pp. 419-434, ISBN 9780443156175. [CrossRef]
- The Human Protein Atlas. https://www.proteinatlas.org/ENSG00000060718-COL11A1/cell+line. Accessed 21 January 2026.
- UniProt. https://www.uniprot.org/uniprotkb/P12107/entry Accessed 21 January 2026.
- Bella, J.; Hulmes, D.J.S. Fibrillar Collagens. Subcell. Biochem. 2017, 82, 457–490. [Google Scholar] [CrossRef]
- Nissen, N.I.; Kehlet, S.; Johansen, A.Z.; Chen, I.M.; Karsdal, M.; Johansen, J.S.; Diab, H.M.H.; Jørgensen, L.N.; Sun, S.; Manon-Jensen, T.; et al. Noninvasive prognostic biomarker potential of quantifying the propeptides of Type XI collagen alpha-1 chain (PRO-C11) in patients with pancreatic ductal adenocarcinoma. Int. J. Cancer 2021, 149, 228–238. [Google Scholar] [CrossRef] [PubMed]
- Liddell JR, Cryer A. A practical guide to monoclonal antibodies, John Wiley and Sons Ltd. Chichester, United Kingdom; 1991. https://www.wiley.com/en-us/A+Practical+Guide+to+Monoclonal+Antibodies-p-9780471929055.
- de los Toyos, J, R.; Méndez, F.J.; Aparicio, J.F.; Vázquez, F.; Del Mar García Suárez, M.; Fleites, A.; Hardisson, C.; Morgan, P.J.; Andrew, P.W.; Mitchell, T.J. Functional analysis of pneumolysin by use of monoclonal antibodies. Infect. Immun. 1996, 64, 480–484. [Google Scholar] [CrossRef] [PubMed]
- Suárez-Alvarez, B.; García-Suárez, M del M.; Méndez, F.J.; de los Toyos, J.R. Characterisation of mouse monoclonal antibodies for pneumolysin: Fine epitope mapping and V gene usage. Immunol. Lett. 2003, 88, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Rey, J.; Murail, S.; de Vries, S.; Derreumaux, P.; Tufféry, P. PEP-FOLD4: a pH-dependent force field for peptide structure prediction in aqueous solution. Nucleic Acids Res., 2023, Web server issue.
- Manso, T.; Folch, G.; Giudicelli, V.; Jabado-Michaloud, J.; Kushwaha, A.; Nguefack Ngoune, V.; Georga, M.; Papadaki, A.; Debbagh, C.; Pégorier, P.; et al. IMGT® databases, related tools and web resources through three main axes of research and development. Nucleic Acids Res. 2022, 50(D1), D1262–D1272. [Google Scholar] [CrossRef] [PubMed]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2013, 41(D1), D36–D42. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Luo, E.Y.; Adams, S.M.; Adams, T.; Ye, Y.; Shetye, S.S.; Soslowsky, L. J.; Bir, D.E. Collagen XI regulates the acquisition of collagen fibril structure, organization and functional properties in tendon. Matrix Biol. 2020, 94, 77–94. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.Y.; Szlarski, P.M.; Kehlet, S.N.; Karsdal, M.A. Chapter 11 - Type XI collagen. In Biochemistry of Collagens, Laminins and Elastin, 2nd ed. Morten A. Karsdal, Ed. Academic Press, Elsevier B.V.; Amsterdam, The Netherlands, 2019, pp: 99-106, ISBN 978-0-12-817068-7. [CrossRef]
- Kessler, E.; Fichard, A.; Chanut-Delalande, H.; Brusel, M.; Ruggiero, F. Bone morphogenetic protein-1 (BMP-1) mediates C-terminal processing of procollagen V homotrimer. J. Biol. Chem. 2001, 276, 27051–27057. [Google Scholar] [CrossRef] [PubMed]
- Fledelius, C.; Johnsen, A.H.; Cloos, P.A.; Bonde, M.; Qvist, P. Characterization of urinary degradation products derived from type I collagen. Identification of a beta-isomerized Asp-Gly sequence within the C-terminal telopeptide (alpha1) region. J. Biol. Chem. 1997, 272, 9755–9763. [Google Scholar] [CrossRef] [PubMed]
- Chubb, S.A. Measurement of C-terminal telopeptide of type I collagen (CTX) in serum. Clin. Biochem. 2012, 45, 928–935. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Zhao, J.; Huang, Y.; Dai, L.; Zhang, X. Meta-analysis of urinary C-terminal telopeptide of type II collagen as a biomarker in osteoarthritis diagnosis. J. Orthop. Translat. 2017, 13, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Necula, L.; Matei, L.; Dragu, D.; Pitica, I.; Neagu, A.; Bleotu, C.; Diaconu, C.C.; Chivu-Economescu, M. Collagen Family as Promising Biomarkers and Therapeutic Targets in Cancer. Int. J. Mol. Sci. 2022, 23, 12415. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.K.; MacFarlane, G.; Srivastava, V.P.; Mohan, S.; Baylink, D.J. A new monoclonal antibody ELISA for detection and characterization of C-telopeptide fragments of type I collagen in urine. Calcif, Tissue Int 2001, 69, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Anti-Collagen 1, alpha 1 telopeptide Antibody SKU: 322-COLT. https://www.aveslabs.com/products/anti-collagen-1-alpha-1-telopeptide-antibody-322-colt. Accessed 20 January 2026.
- Chung, H.J.; Steplewski, A.; Chung, K.Y.; Uitto, J.; Fertala, A. Collagen fibril formation. A new target to limit fibrosis. J. Biol. Chem. 2008, 283, 25879–25886. [Google Scholar] [CrossRef] [PubMed]
- Barascuk, N.; Veidal, S.S.; Larsen, L.; Larsen, D.V.; Larsen, M.R.; Wang, J.; Zheng, Q.; Xing, R.; Cao, Y.; Rasmussen, L.M.; et al. A novel assay for extracellular matrix remodeling associated with liver fibrosis: An enzyme-linked immunosorbent assay (ELISA) for a MMP-9 proteolytically revealed neo-epitope of type III collagen. Clin. Biochem. 2010, 43, 899–904. [Google Scholar] [CrossRef] [PubMed]
- Leeming; D, J.; He, Y.; Veidal, S.; Nguyen, Q.; Larsen, D.; Koizumi, M.; Segovia-Silvestre, T.; Zhang, C.; Zheng, Q.; Sun, S.; et al. A novel marker for assessment of liver matrix remodeling: An enzyme-linked immunosorbent assay (ELISA) detecting a MMP generated type I collagen neo-epitope (C1M). Biomarkers 2011, 16, 616–628. [Google Scholar] [CrossRef] [PubMed]
- Veidal, S.S.; Karsdal, M.A.; Nawrocki, A.; Larsen, M.R.; Dai, Y.; Zheng, Q.; Hägglund, P.; Vainer, B.; Skjøt-Arkil, H.; Leeming, D.J. Assessment of proteolytic degradation of the basement membrane: A fragment of type IV collagen as a biochemical marker for liver fibrosis. Fibrogenesis Tissue Repair 2011, 4, 22. [Google Scholar] [CrossRef] [PubMed]
- Willumsen, N.; Bager, C.L.; Leeming, D.J.; Smith, V.; Karsdal, M.A.; Dornan, D.; Bay-Jensen, A-C. Extracellular matrix specific protein fingerprints measured in serum can separate pancreatic cancer patients from healthy controls. BMC Cancer 2013, 13, 554. [Google Scholar] [CrossRef] [PubMed]
- Brachmann, C.B.; Zhang, Y.; Zavodovskaya, M.; Hu, J.; Maltzman, J.D.; Smith, V.; Xiao, Y.; Patterson, S.D. Evaluating collagen neoepitopes as pharmacodynamic biomarkers of GS-5745, an MMP9 inhibitor, in advanced gastric cancer. J. Clin. Oncol. 2017, 55 (suppl.): 58. [CrossRef]
- García-Ocaña, M.; Vázquez, F.; García-Pravia, C.; Fuentes-Martínez, N.; Menéndez-Rodríguez, P.; Fresno-Forcelledo, F.; Barneo-Serra, L.; Del Amo-Iribarren, J.; Simón-Buela, L.; De Los Toyos, J.R. Characterization of a novel mouse monoclonal antibody, clone 1E8.33, highly specific for human procollagen 11A1, a tumor-associated stromal component. Int. J. Oncol. 2012, 40, 1447–1454. [Google Scholar] [CrossRef]
- Steplewski, A.; Fertala, A. Inhibition of collagen fibril formation. Fibrogenesis Tissue Repair 2012, 5 (Suppl 1): S29, https://fibrogenesis.biomedcentral.com/articles/10.1186/1755-1536-5-S1-S29. [CrossRef] [PubMed]
- Yeung, T.L.; Leung, C.S.; Yip, K.; Sheng, J.; Vien, L.; Bover, L.C.; Birrer, M.J.; Wong, S.T.C.; Mok, S.C. Anticancer Immunotherapy by MFAP5 Blockade Inhibits Fibrosis and Enhances Chemosensitivity in Ovarian and Pancreatic Cancer. Clin. Cancer Res. 2019, 25, 6417–6428. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Jiang, P.; Liu, D.; Wang, H.Q.; Deng, Q.; Niu, X.; Lu, L.; Dai, H.; Wang, H.; Yang, W. Matrix Metalloproteinase 11 Is a Potential Therapeutic Target in Lung Adenocarcinoma. Mol. Ther. Oncolytics 2019, 14, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.K.; Jiang, F.; Kim, Y.J.; Ryu, K.; Masamune, A.; Hamada, S.; Park, Y.Y.; Koh, S.S. CTHRC1 Induces Pancreatic Stellate Cells (PSCs) into Myofibroblast-like Cancer-Associated Fibroblasts (myCAFs). Cancers (Basel) 2023, 15. 3370. [CrossRef]
Figure 1.
The amino acid sequence of the putative extracellular processed form of human collagen XIα1 (residues 512 to 1563 of the P12107-1 A isoform), recombinantly expressed in CHO cells, provided by GenScript. It encompasses the C-terminal telopeptide -(1543) IQPLPILSSKKTRRHTEGMQA (1563)- sequence. It is preceded by the MGWSCIILFLVATATGVHS signal peptide and by a 6xHis head.
Figure 1.
The amino acid sequence of the putative extracellular processed form of human collagen XIα1 (residues 512 to 1563 of the P12107-1 A isoform), recombinantly expressed in CHO cells, provided by GenScript. It encompasses the C-terminal telopeptide -(1543) IQPLPILSSKKTRRHTEGMQA (1563)- sequence. It is preceded by the MGWSCIILFLVATATGVHS signal peptide and by a 6xHis head.

Figure 2.
The amino acid sequence of the putative C-propeptide, from human procollagen XIα1 (residues 1564 to 1806 of the P12107-1 A isoform), recombinantly expressed in CHO cells, provided by GenScript. It is preceded by the MGWSCIILFLVATATGVHS signal peptide.
Figure 2.
The amino acid sequence of the putative C-propeptide, from human procollagen XIα1 (residues 1564 to 1806 of the P12107-1 A isoform), recombinantly expressed in CHO cells, provided by GenScript. It is preceded by the MGWSCIILFLVATATGVHS signal peptide.

Figure 3.
The amino acid sequence of the COL11A1 Fusion Protein (residues 1545 to 1806 of the P12107-1 A isoform), recombinantly expressed in Escherichia coli, after the removal of an N-terminal GST tag, provided by Proteintech. The first 19 N-terminal (1545) PLPILSSKKTRRHTEGMQA (1563) amino acid residues are part of the putative C-telopeptide. The next 243 amino acid residues (1564 to 1806) constitute the C-propeptide.
Figure 3.
The amino acid sequence of the COL11A1 Fusion Protein (residues 1545 to 1806 of the P12107-1 A isoform), recombinantly expressed in Escherichia coli, after the removal of an N-terminal GST tag, provided by Proteintech. The first 19 N-terminal (1545) PLPILSSKKTRRHTEGMQA (1563) amino acid residues are part of the putative C-telopeptide. The next 243 amino acid residues (1564 to 1806) constitute the C-propeptide.

Figure 4.
ELISA blocking of recognition of recombinant collagen XIα1, with soluble free N-acetylated peptides. Figure drawn using GraphPad Prism Version 10.0.3 (275) for Windows (GraphPad Software, Boston, Massachusetts USA, www.graphpad.com).
Figure 4.
ELISA blocking of recognition of recombinant collagen XIα1, with soluble free N-acetylated peptides. Figure drawn using GraphPad Prism Version 10.0.3 (275) for Windows (GraphPad Software, Boston, Massachusetts USA, www.graphpad.com).
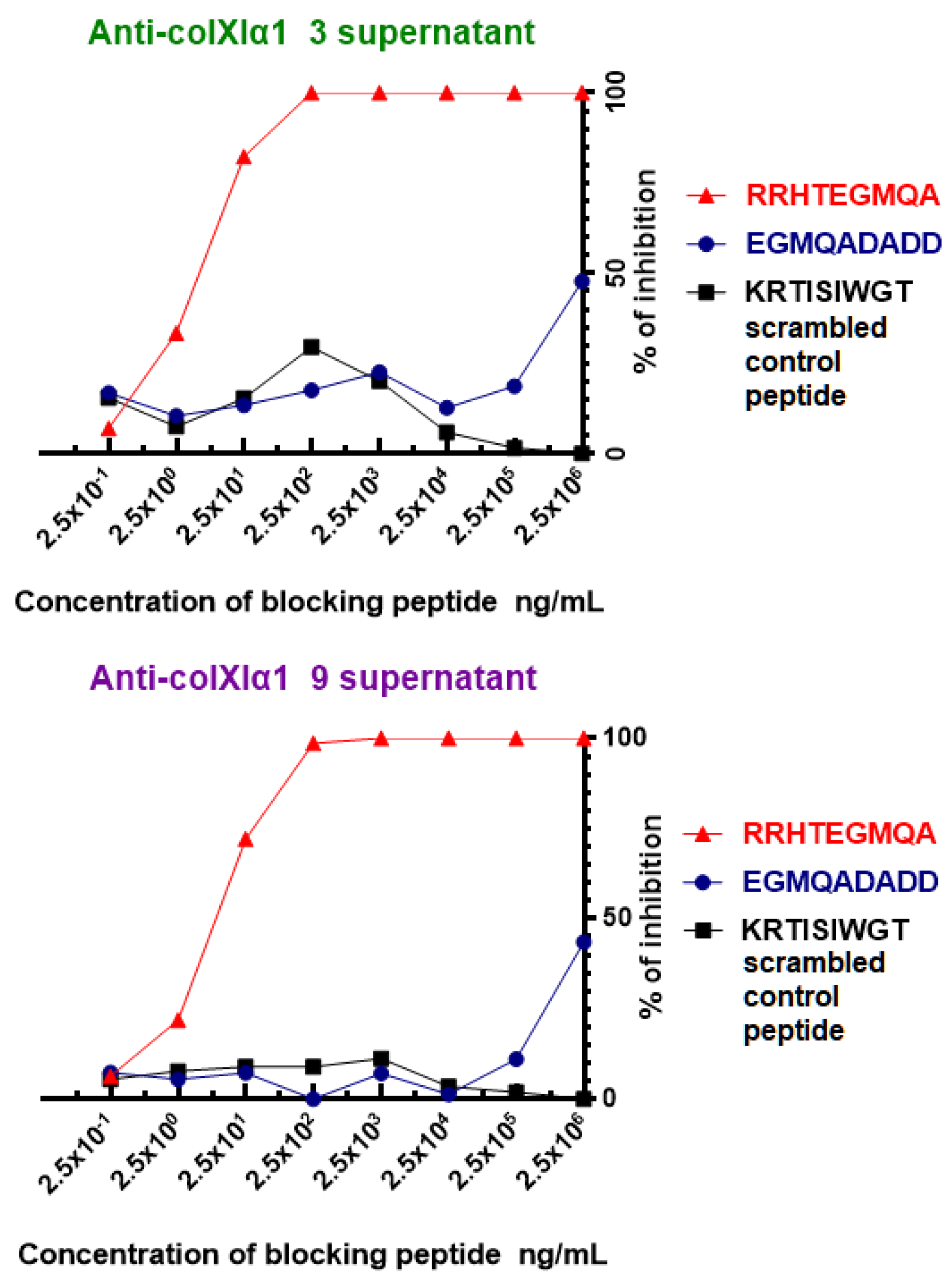
Figure 5.
SDS-PAGE gel staining and Western blot of recombinant antigens with finally purified preparations of Anti-colXIα1 clone 9 and PLY-7 mAbs. Lane 1: PageRuler™ Plus Prestained Protein Ladder. Lane 2: COL11A1 Fusion Protein (Proteintech). Lane 3: Collagen XIα1 (GenScript). Lane 4: C-propeptide (GenScript). Full-length blots/gels are presented in Supplementary Figure S5.
Figure 5.
SDS-PAGE gel staining and Western blot of recombinant antigens with finally purified preparations of Anti-colXIα1 clone 9 and PLY-7 mAbs. Lane 1: PageRuler™ Plus Prestained Protein Ladder. Lane 2: COL11A1 Fusion Protein (Proteintech). Lane 3: Collagen XIα1 (GenScript). Lane 4: C-propeptide (GenScript). Full-length blots/gels are presented in Supplementary Figure S5.
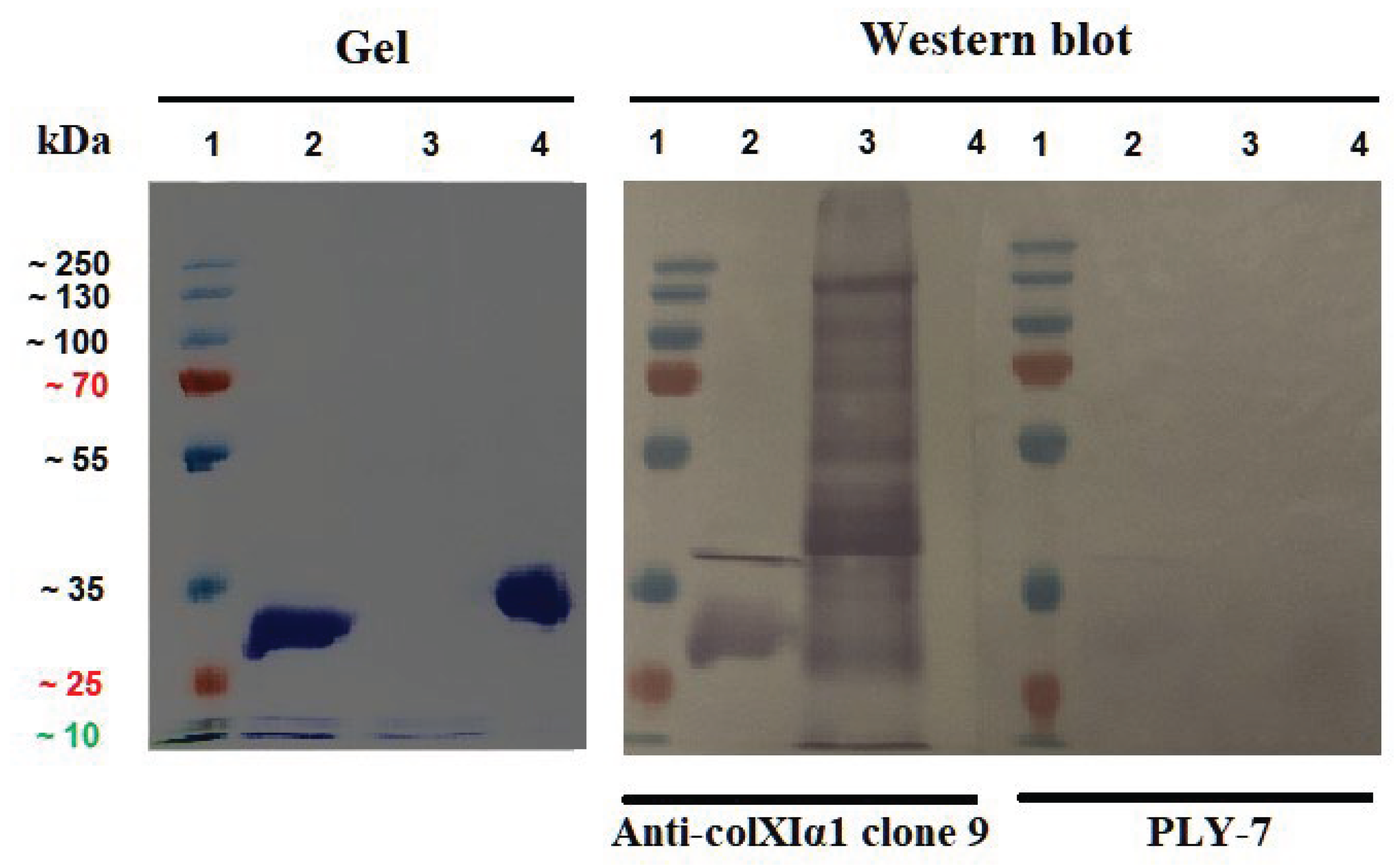
Figure 6.
The PEP-FOLD4-derived structural predictions of peptides related to the putative C-telopeptide. Panel A: The 50 N-terminal amino acid sequence of the COL11A1 Fusion Protein from Proteintech, with the first 19 N-terminal PLPILSSKKTRRHTEGMQA amino acid residues of the putative C-telopeptide. Panel B: A free RRHTEGMQA peptide. Panel C: The 50 C-terminal amino acid sequence of the GenScript´s recombinant collagen XIα1 form, whose last 21 C-terminal IQPLPILSSKKTRRHTEGMQA amino acid residues correspond to the putative C-telopeptide. In Panels A and B, the N-terminus of the peptides is on the left, but in Panel C, it is on the right.
Figure 6.
The PEP-FOLD4-derived structural predictions of peptides related to the putative C-telopeptide. Panel A: The 50 N-terminal amino acid sequence of the COL11A1 Fusion Protein from Proteintech, with the first 19 N-terminal PLPILSSKKTRRHTEGMQA amino acid residues of the putative C-telopeptide. Panel B: A free RRHTEGMQA peptide. Panel C: The 50 C-terminal amino acid sequence of the GenScript´s recombinant collagen XIα1 form, whose last 21 C-terminal IQPLPILSSKKTRRHTEGMQA amino acid residues correspond to the putative C-telopeptide. In Panels A and B, the N-terminus of the peptides is on the left, but in Panel C, it is on the right.
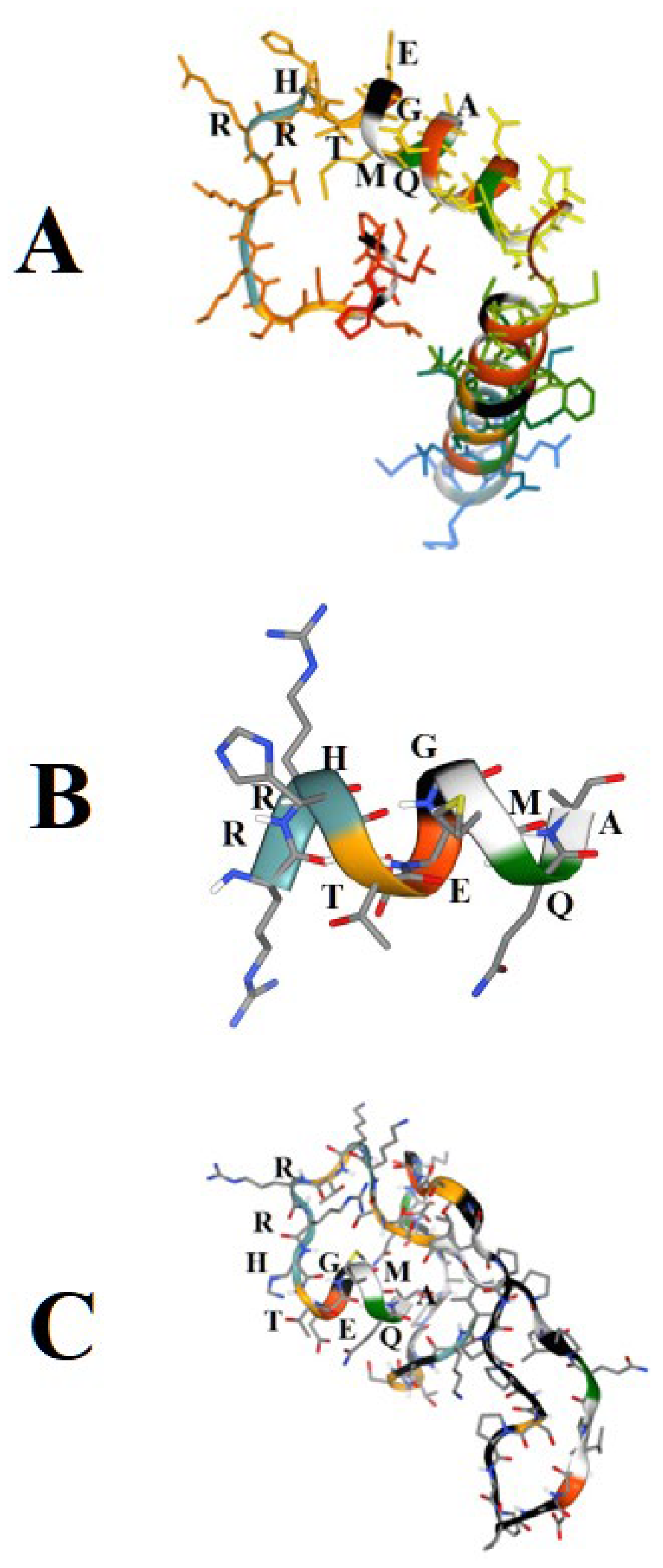
Figure 7.
Representative images of immunostaining of cultured cancer cell lines with the purified Anti-colXIα1 clone 9 and irrelevant PLY-7 mAbs. Original magnification: 400x; scale bar: 20.0 µm.
Figure 7.
Representative images of immunostaining of cultured cancer cell lines with the purified Anti-colXIα1 clone 9 and irrelevant PLY-7 mAbs. Original magnification: 400x; scale bar: 20.0 µm.
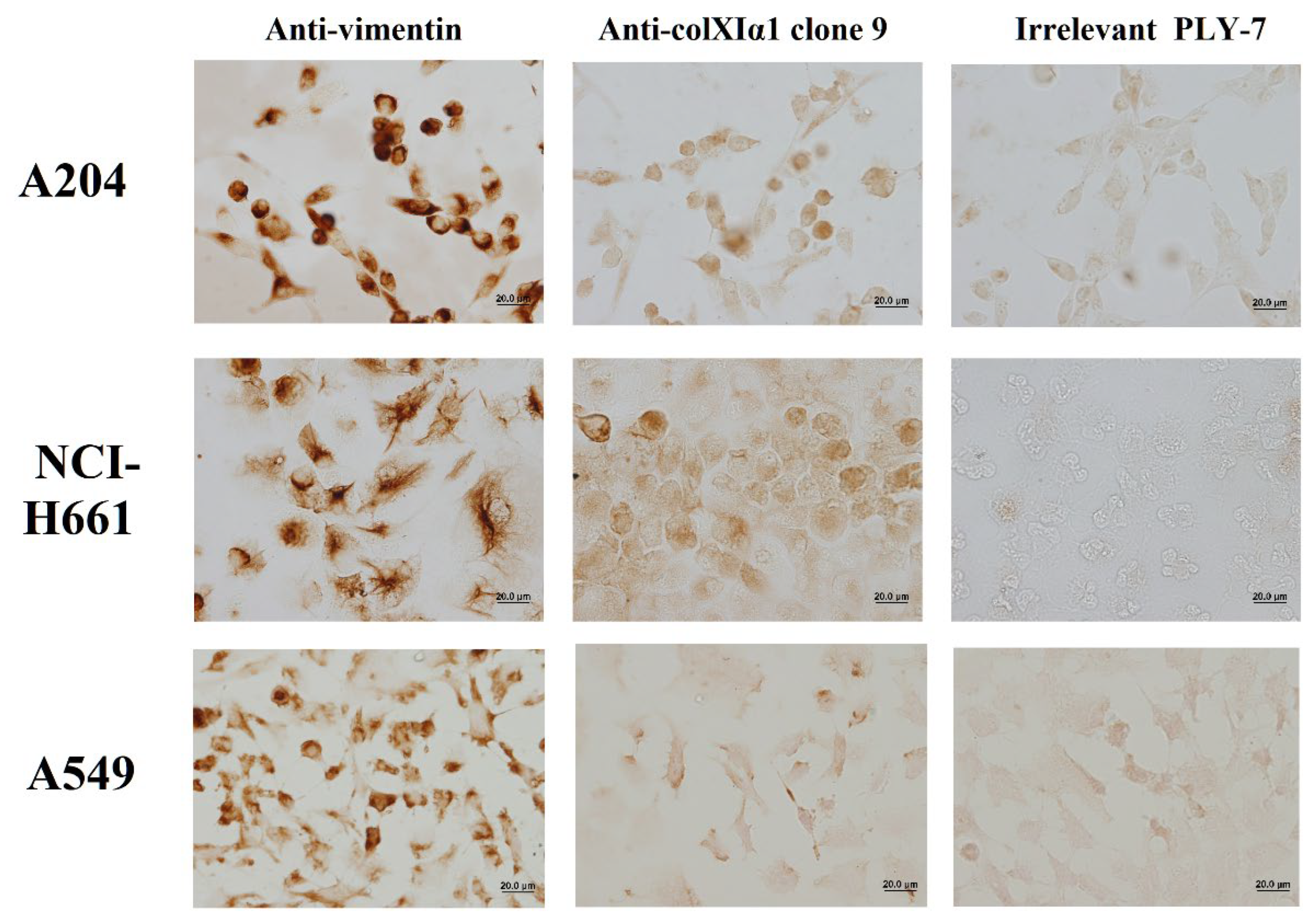

Table 1.
Antibodies used in ICC analyses.
| Primary antibody (species) |
Protein recognized |
Source | Dilution | Incubation conditions |
|---|---|---|---|---|
| Anti-colXIα1 clone 9 (mAb) |
Human collagen XIα1 |
In-house | 1:300 from 1 mg/mL |
48 h at 4 ºC |
| PLY-7 (mAb) |
Pneumococcal pneumolysin |
In-house | 1:300 from 1 mg/mL |
48 h at 4 ºC |
| Vimentin Antibody (V9) (mAb) |
Human vimentin |
Santa Cruz Biotechnology, Inc. SC-6260 |
1:500 | 48 h at 4 ºC |
Table 2.
Immunoreactivity against components of the immunogens.
| ELISAa | |||
|---|---|---|---|
| Antigen | |||
| Supernatant/mAb | Immunogen 1 KLH- C - GG- EGMQADADD |
Immunogen 2 KLH- C - GG- RRHTEGMQA |
KLH |
| Anti-colXIα1 clone 3 #14/08/2023 |
- | 3+ | - |
| Anti-colXIα1 clone 9 #14/08/2023 |
2+ | 3+ | - |
| Medium RPMI-HAT-20% FBS | - | - | - |
a Color development was estimated by eye. # From supernatants in RPMI-HAT-20% FBS, with indication of collection date.
Table 3.
Some other immunoreactivity characteristics.
|
ELISAa |
Conventional denaturing and reducing Western blot |
|||||||
|---|---|---|---|---|---|---|---|---|
| Supernatant/ mAb |
Recombinant collagen XIα1 |
Recombinant C-propeptide |
A204 cell lysate |
A549 cell lysate |
6% gel | 12% gel | 10% gel | |
| Purified human collagen XI |
Recombinant C-propeptide |
A204 cell lysate |
A549 cell lysate |
|||||
| Anti-colXIα1 clone 3 |
3.259 | 0.000 | 0.105 | 0.029 | - | - | - | - |
| Anti-colXIα1 clone 9 |
3.519 | 0.000 | 0.055 | 0.064 | - | - | - | - |
a ELISA optical density scale of blanked data from 0 to 4.
Table 4.
The ELISA immunoreactivity of the finally purified preparations of the Anti-colXIα1 clone 9 and PLY-7 mAbs.
Table 4.
The ELISA immunoreactivity of the finally purified preparations of the Anti-colXIα1 clone 9 and PLY-7 mAbs.
| ELISAa Antigen (1 µg/ 100 µl well) |
|||
|---|---|---|---|
| Purified mAb (1 µg/ 100 µl well) |
Recombinant COL11A1 Fusion Protein (Proteintech) |
Recombinant collagen XIα1 (GenScript) |
Recombinant C-propeptide (GenScript) |
| Anti-colXIα1 clone 9 | 0.699 | 2.228 | 0.012 |
| Irrelevant PLY-7 | 0.250 | 0.116 | 0.109 |
a ELISA optical density scale of blanked data from 0 to 4.
Table 5.
The most probable germinal V(D)J gene segment usage by the two mAbs.
| mAb | VH | DH | JH | Vkappa | J kappa |
|---|---|---|---|---|---|
| Anti-colXIα1 clone 3 |
IGHV9-2-1*01 F | IGHD2-10*01 F | IGHJ3*01 F | IGKV8-30*01 F | IGKJ4*01 F |
| Anti-colXIα1 clone 9 |
IGHV9-2-1*01 F | IGHD2-10*01 F | IGHJ3*01 F | IGKV8-30*01 F | IGKJ4*01 F |
Table 6.
Deduced IMGT CDRs.
| mAb | CDRH1 | CDRH2 | CDRH3 | CDRL1 | CDRL2 | CDRL3 |
|---|---|---|---|---|---|---|
| Anti-colXIα1 clone 3 |
GYTFTDYS | INTETGEP | VRRANYGNAWFVY | QSLLYRSNQKNY | WAS | QQYYDYPFT |
| Anti-colXIα1 clone 9 |
GYTFTDYS | INTETGQP | IRRANYGNAWFAY | QNLLYRSNHKNY | WAS | QQYYDYPFT |
Amino acids that are not identical between the mAbs are in bold.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



