
Submitted:
20 January 2026
Posted:
21 January 2026
You are already at the latest version
Abstract
T-cell malignancies represent a complex spectrum of clinically and biologically heteroge-neous diseases. Effective translational research and drug development are critically de-pendent on preclinical models that faithfully recapitulate this diversity. This review ana-lyzes the current preclinical landscape, identifying a profound disparity between the clin-ical spectrum of T-cell neoplasms and the available in vitro tools. We demonstrate that the existing armamentarium of cell lines is heavily skewed, with an abundance of models for T-cell lymphoblastic leukemia/lymphoma (T-ALL), cutaneous T-cell lymphoma (CTCL), and anaplastic large cell lymphoma (ALCL). This skew is a direct result of a biological se-lection bias, as these entities are often driven by potent, TME-independent oncogenes (e.g., NOTCH1 mutations, NPM1-ALK fusions) conducive to immortalization. Conversely, the majority of peripheral T-cell lymphoma (PTCL) subtypes, which are frequently TME-dependent and clinically aggressive, remain "preclinical orphans" with few or no authenticated models. This "preclinical void" constitutes a major bottleneck, impeding mechanistic studies and therapeutic progress. We discuss the limitations of 2D cultures and highlight the necessity of adopting advanced platforms, such as patient-derived xen-ografts (PDX) and 3D organoid systems. These "avatar" models preserve vital tumor het-erogeneity and microenvironmental context, offering superior predictive value. The sys-tematic development and integration of these next-generation models are essential to bridge the translational gap and advance precision medicine for all patients with T-cell malignancies.
Keywords:
T-cell lymphoma
; preclinical models
; cell lines
; patient-derived xenograft (PDX)
; translational research
; peripheral T-cell lymphoma (PTCL)
1. Introduction
Human leukemia-lymphoma cell lines represent a cornerstone of modern cancer research. First established in the 1960s, these immortalized cell populations provide an unlimited, genetically stable, and globally accessible resource for investigating the fundamental pathophysiology of hematopoietic tumors. [1,2,3] Their utility in dissecting molecular pathways, identifying novel therapeutic targets, and conducting high-throughput drug screening is undisputed. However, the value of this armamentarium is contingent upon its ability to faithfully represent the vast clinical and biological diversity of the diseases under study. For T-cell malignancies, in particular, such an endeavor presents a formidable challenge. [4]
T-cell neoplasms are not a monolithic entity but rather a complex collection of dozens of distinct diseases, as defined by the World Health Organization (WHO) and International Consensus Classification (ICC) systems. [5] These malignancies arise from T-lymphocytes at various stages of differentiation, ranging from immature thymic precursors, which give rise to T-cell lymphoblastic leukemia/lymphoma (T-ALL/LBL), to a wide spectrum of mature, post-thymic T-cells that form the basis of peripheral T-cell lymphomas (PTCLs). [6,7,8] Our group and others have dedicated significant effort to elucidating the profound heterogeneity within the PTCL category, which is characterized by immense morphological, immunophenotypic, and molecular diversity [9]. This complexity makes PTCLs exceptionally difficult to classify, diagnose, and treat, earning them the unfortunate designation of "orphan diseases". [10,11]
This review sought to show that a critical disparity exists between the clinical diversity of T-cell malignancies and the preclinical models available to study them. The landscape of available cell lines is heavily skewed, with an abundance of well-characterized models for a few specific subtypes—namely, T-ALL/LBL, cutaneous T-cell lymphoma (CTCL), and anaplastic large cell lymphoma (ALCL). In stark contrast, for the majority of PTCL entities, which are often highly aggressive and associated with dismal prognoses, reliable and authenticated cell line models are either exceedingly rare or entirely non-existent. [11,12] This imbalance is not a mere artifact of historical collection but a direct consequence of the underlying biology of these tumors. Malignancies such as ALK-positive ALCL and many T-ALLs are driven by potent oncogenes, like the NPM1-ALK fusion protein or activating NOTCH1 mutations, which confer robust, cytokine-independent proliferative signals that are highly conducive to establishment and maintenance in artificial in vitro culture conditions. [13] Conversely, many rare PTCLs, such as angioimmunoblastic T-cell lymphoma (AITL), are intrinsically dependent on a complex tumor microenvironment (TME) for survival and proliferation—a condition that is lost in standard two-dimensional culture systems. This biological selection bias has created a vicious cycle: the availability of models drives research and drug discovery, leading to disproportionate progress in ALCL and T-ALL [14], while the entities lacking models remain neglected, perpetuating their status as preclinical and clinical orphans (Figure 1).
2. The Established Landscape: Available T-Cell Malignancy Cell Lines
The current collection of T-cell malignancy cell lines, curated by repositories such as the American Type Culture Collection (ATCC) and the German Collection of Microorganisms and Cell Cultures (DSMZ), reflects this skewed distribution. A systematic review reveals a clear pattern of abundance for some entities and a profound void for others (Table 1, Table 2 and Table 3).
2.1. Models for Immature T-Cell Neoplasms: T-Lymphoblastic Leukemia/Lymphoma (T-ALL/LBL)
T-ALL/LBL are aggressive neoplasms of T-cell precursors, accounting for approximately 15% of pediatric and 25% of adult ALL cases. [7,15,16] The molecular pathogenesis is characterized by a multi-step accumulation of genetic lesions. Key events include the aberrant expression of oncogenic transcription factors (e.g., TAL1, TLX1, HOXA), which often arise from chromosomal translocations involving T-cell receptor (TCR) loci and define distinct molecular subgroups that correspond to specific stages of T-cell developmental arrest. [15,16] These events are frequently coupled with hyperactive NOTCH1 signaling, due to activating mutations in over 60% of cases, and loss of the CDKN2A/B tumor suppressor locus in up to 70% of cases. [15,17] The strong proliferative signals conferred by these mutations have facilitated the establishment of numerous cell lines (Table 1).
A comprehensive analysis of 16 widely used T-ALL cell lines demonstrated that, despite their common classification, each line possesses a unique immunophenotype and TCR gene rearrangement status. This study proposed a categorization into pro-T, pre-T, cortical T, and mature T differentiation stages, underscoring the heterogeneity that these models collectively represent. [18]
2.2. Models for Cutaneous T-Cell Lymphomas (CTCL)
CTCLs are a group of extranodal non-Hodgkin lymphomas defined by the infiltration of malignant T-cells into the skin, with Mycosis Fungoides (MF) and the related leukemic form, Sézary Syndrome (SS), being the most prevalent subtypes. [19] Several key cell lines have been established, primarily from patients with advanced-stage, aggressive disease (Table 2). [20,21] These models have been instrumental in preclinical studies, particularly for evaluating novel therapeutic agents. For instance, the HUT 78 cell line was used to demonstrate the sensitivity of T-cell lymphoma to the histone deacetylase inhibitor (HDACi) depsipeptide, [22] providing a key preclinical rationale for the development of this drug class for T-cell malignancies. [23]
2.3. Models for Anaplastic Large Cell Lymphoma (ALCL)
ALCL is a mature T-cell lymphoma characterized by strong expression of the CD30 antigen. [24] It is divided into two main categories based on the expression of Anaplastic Lymphoma Kinase (ALK). ALK-positive ALCL, which is more common in younger patients, is defined by chromosomal rearrangements involving the ALK gene on chromosome 2p23, most frequently the t(2;5)(p23;q35) translocation that generates the potent NPM1-ALK fusion oncogene. [25] The constitutive kinase activity of NPM1-ALK drives robust downstream signaling through pathways including JAK/STAT and PI3K/AKT, promoting cell survival and proliferation. [26] This powerful oncogenic driver has made ALK+ ALCL one of the most successfully modeled PTCL subtypes, with a wealth of available cell lines (Table 3). [27] This oncogenic potency has made ALK-positive ALCL one of the most successful PTCL subtypes for modeling, with numerous cell lines available (Table 3). For example, the characterization of the L-82 cell line not only confirmed the presence of the NPM1-ALK fusion but also established its tumorigenicity in SCID mice, providing a crucial preclinical in vivo model. These models have proven essential not only for in vitro studies but also for confirming tumorigenicity in vivo, as demonstrated by the characterization of novel lines in SCID mice. [28] In contrast, ALK-negative ALCL, which lacks ALK rearrangements and generally has a poorer prognosis, is less well-represented by cell line models, though some exist (Table 3; Figure 2). [5,29,30]
2.4. The Void: Scarcity of Models for Other Peripheral T-Cell Lymphomas (PTCL)
Beyond ALCL and CTCL, the landscape of preclinical models for mature T-cell malignancies is remarkably sparse. This gap is particularly acute for the diverse and aggressive subtypes that fall under the PTCL umbrella. As our group has demonstrated through extensive gene expression profiling studies, many of these entities arise from distinct functional T-cell subsets, such as the derivation of AITL from T-follicular helper (TFH) cells. [9,31] This cellular origin implies a profound dependence on signals from a complex TME, a feature that is lost in conventional cell culture and likely explains the extreme difficulty in establishing stable cell lines for these diseases.
Consequently, there are no widely available, authenticated cell lines for many of the most challenging PTCLs, including among others angioimmunoblastic T-cell lymphoma (AITL), hepatosplenic T-cell lymphoma (HSTCL) [32], enteropathy-associated T-cell lymphoma (EATL) and extranodal NK/T-cell lymphoma, nasal type (ENKTL) [33].
A notable and recent exception is the T8ML-1 cell line, the first authenticated model for PTCL, not otherwise specified (PTCL-NOS) harboring the recurrent t(14;19)(q11.2;q13.3) translocation. [30,34] This translocation results in the juxtaposition of the TCR alpha (TRA@) locus and the PVRL2 gene, leading to its overexpression. [34] The successful characterization of T8ML-1 provides a crucial and unique tool for studying this specific genetic subtype of PTCL-NOS, but it also highlights the profound scarcity of models for this "wastebasket" category of PTCL. [34] The complexity of this category is further exemplified by cell lines like FE-PD, a CD30-positive line originally established from a patient diagnosed with Hodgkin’s disease, later reclassified as an aggressive anaplastic large cell lymphoma, and now considered a model for ALK-negative ALCL or CD30+ PTCL-NOS. [35,36]
This striking imbalance between diseases and models has tangible consequences. The entities with robust in vitro models, such as ALCL and CTCL, are precisely those that have seen the most significant advances in targeted therapy, including the clinical approval of ALK inhibitors and the early indication of HDAC inhibitors, respectively. [13] In contrast, diseases like PTCL/NOS, AITL and HSTCL, for which cell lines are so scant, continue to be treated with conventional, non-specific chemotherapy regimens with poor outcomes. [8,11,12,37,38,39] This correlation underscores a direct link: the availability of a preclinical model system is a critical prerequisite for successful translational research and the development of targeted therapies.

Table 1.
Characteristics of Selected T-Lymphoblastic Leukemia/Lymphoma (T-ALL/LBL) Cell Lines.
| Cell Line | ATCC/DSMZ No. | Patient Origin | Key Genetic Alterations | Immunophenotypic Stage | Key Reference(s) (PMID) |
|---|---|---|---|---|---|
| Jurkat | TIB-152 / ACC 282 | 14Y, M, Relapsed T-ALL | PTEN null, CDKN2A del, TP53 R196* | Cortical/Mature T | [5] |
| CCRF-CEM | CCL-119 / ACC 240 | 3Y, F, Relapsed T-ALL | FBXW7 R465C, TP53 R248Q | Cortical T | [5] |
| MOLT-3 | CRL-1552 / ACC 316 | 19Y, M, Relapsed T-ALL | FBXW7 R505C, TP53 R248Q | Cortical T | [5] |
| SUP-T1 | CRL-1942 / ACC 140 | 8Y, M, Relapsed T-LBL | FBXW7 R465H, TP53 null | Mature T (CD4+/CD8+) | [7] |
Abbreviations: Y, year; M, male; F, female; del, deletion.
Table 2.
Characteristics of Selected Cutaneous T-Cell Lymphoma (CTCL) Cell Lines.
| Cell Line | ATCC/DSMZ No. | Histotype | HTLV-1 Status | Key Genetic Alterations | Key Reference(s) (PMID) |
|---|---|---|---|---|---|
| HUT 78 | TIB-161 / ACC 398 | Sézary Syndrome | Negative | TP53 C176F | [8] |
| H9 | HTB-176 | CTCL (HUT 78 clone) | Negative | CDKN2A del, RB1 del, SOCS1 del, TP53 R196* | [4] |
| HH | CRL-2105 / ACC 707 | Aggressive CTCL | Negative | FOXK2::TP63 fusion, TP53 splice site mut | [10,31] |
| MJ [G11] | CRL-8294 | CTCL | Positive | - | [10] |
Abbreviations: del, deletion; mut, mutation.
Table 3.
Characteristics of Selected Anaplastic Large Cell Lymphoma (ALCL) and Other Peripheral T-Cell Lymphoma (PTCL) Cell Lines.
Table 3.
Characteristics of Selected Anaplastic Large Cell Lymphoma (ALCL) and Other Peripheral T-Cell Lymphoma (PTCL) Cell Lines.
| Cell Line | ATCC/DSMZ No. | Histotype | Key Genetic Alterations | Key Reference(s) (PMID) |
|---|---|---|---|---|
| SU-DHL-1 | CRL-2955 / ACC 356 | ALCL, ALK+ | NPM1-ALK t(2;5), TP53 R273H | [9] |
| Karpas 299 | - / ACC 31 | ALCL, ALK+ | NPM1-ALK t(2;5) | [41] |
| SUP-M2 | - / ACC 509 | ALCL, ALK+ | NPM1-ALK t(2;5) | [44] |
| KI-JK | - / ACC 695 | ALCL, ALK+ | NPM1-ALK t(2;5) | [25] |
| L-82 | - / ACC 597 | ALCL, ALK+ | NPM1-ALK t(2;5) | [28] |
| FE-PD | - | PTCL-NOS / ALK- ALCL (CD30+) | JAK1 G1097V, STAT3 G618R, DUSP22 rearrangement | [35] |
| TLBR-2 | - / ACC 905 | BIA-ALCL, ALK- | STAT3 D661Y | [29] |
| T8ML-1 | - | PTCL-NOS | TRA@::PVRL2 t(14;19) | [34] |
Abbreviations: BIA-ALCL, Breast Implant-Associated ALCL; PTCL-NOS, Peripheral T-cell lymphoma, not otherwise specified.
3. Utility and Limitations: The Role of Cell Lines in Translational Research and Drug Screening
The available T-cell lymphoma cell lines, though limited in scope, have been indispensable for major advances in both basic and translational research. The ALCL cell lines, for example, were fundamental in validating ALK as a bona fide therapeutic target, elucidating its downstream signaling pathways, and serving as the primary preclinical platform for testing the first generation of ALK inhibitors like crizotinib. [13,26] Similarly, CTCL cell lines have been crucial for demonstrating the potent anti-lymphoma activity of HDACis, providing the preclinical proof-of-concept that paved the way for their successful clinical development and approval. [22] In T-ALL, cell lines have been workhorses for dissecting the complex transcriptional networks downstream of NOTCH1 activation and for evaluating the therapeutic potential of gamma-secretase inhibitors. [15,40,41]
4. Key Scientific Questions Driving the Need for Better Models
While invaluable, the current armamentarium of cell lines primarily allows researchers to address questions pertinent to the biology of self-propagating, TME-independent tumors. Key scientific questions that have been successfully explored using these models include several issues. First, the dissection of of oncogenic pathways. Cell lines like Karpas 299 and SU-DHL-1 have been instrumental in mapping the signaling cascades downstream of the NPM1-ALK fusion protein, identifying critical dependencies on pathways like JAK/STAT and PI3K/AKT. [26,42] Similarly, T-ALL lines have clarified the central role of NOTCH1 signaling in driving leukemogenesis. [40,41,43]. Second, regarding target validation and preclinical drug screening, the availability of these models provides a high-throughput platform to test the efficacy of targeted drugs. The validation of ALK inhibitors in ALCL and HDAC inhibitors in CTCL are prime examples of successful translational research pipelines built upon cell line models. Third, cell lines are powerful tools for elucidating how novel drugs exert their anti-tumor effects and for investigating the molecular mechanisms that lead to acquired drug resistance.
Unfortunately, the most pressing challenges in the field of T-cell lymphomas lie in the entities for which these models are lacking. The scarcity of representative cell lines for the majority of PTCL subtypes means that numerous fundamental scientific questions remain largely unanswered, underscoring the urgent need for new and more sophisticated preclinical tools [44]. One of the most challenging issues in PTCLs is the definition of the role of the tumor microenvironment (TME) [45]. Many PTCLs, particularly AITL, are defined by a complex and pathologically integral TME. Standard cell lines, which are selected for growth in isolation, cannot be used to study the critical survival signals provided by the TME or to test therapies that target these tumor-stroma interactions. In addition, the cell of origin and even the normal counterparts of PTCLS are still largely unknown. While gene expression profiling has linked entities like AITL to T-follicular helper cells [46,47], the exact developmental stage and functional state of the cell of origin for most PTCL-NOS cases remain obscure. [9,31,48,49] Functional models derived from these specific subtypes are required to validate these hypotheses and understand the initial steps of transformation.
Another valuable point is understanding how rare, subtype-defining genetic lesions do drive lymphomagenesis. The discovery of recurrent alterations like the TRA@::PVRL2 translocation in a subset of PTCL-NOS was a major step forward, and the T8ML-1 cell line provides a unique model to study its consequences. [34] However, for dozens of other recurrent but rare mutations, no models exist, preventing any functional investigation into their pathogenic role. [50]
Of note, the lack of models for entities like AITL and HSTCL [12] creates a vicious cycle: without a platform for preclinical testing, rational drug development is stalled, and these patients continue to be treated with non-specific chemotherapy that yields poor outcomes. [11,37,39,50] Similarly, the mechanisms of intrinsic and acquired chemoresistance in PTCL are not well defined. Most PTCLs are notoriously resistant to standard chemotherapy regimens. Understanding the underlying biological basis for this resistance requires models that faithfully recapitulate the genetic and phenotypic heterogeneity of patient tumors, a feature that clonally-selected, long-passaged cell lines often lose. Finally, the development of treatments like CAR-T cells requires preclinical models that allow for the assessment of immune-mediated killing. [51] This is a significant challenge in systems lacking a competent immune system or the relevant tumor antigens, highlighting the need for syngeneic models or humanized PDX models. [52,53]
Addressing these fundamental questions is not possible with the current, skewed collection of cell lines. It necessitates a concerted effort to develop a new generation of preclinical models, including authenticated cell lines from rare entities and, critically, PDX models that better preserve the biological complexity of the primary disease. [54]
Despite these successes, the inherent limitations of cell line models are significant and well-documented. Having been selected for robust growth in artificial, two-dimensional culture, they represent a clonally homogeneous population that fails to capture the extensive intra-tumoral heterogeneity that is a hallmark of cancer in patients and a major driver of therapeutic resistance. [55,56] Furthermore, these models are completely divorced from the native TME, which provides critical survival signals and contributes to drug resistance. [57] This is a particularly glaring deficiency for lymphomas like AITL, where the TME is not merely supportive but is a defining pathological feature of the disease. [58] The cumulative result of these shortcomings is a poor predictive value for clinical success; promising results from cell line studies frequently fail to translate into patient benefit, with over 90% of drugs that show preclinical activity ultimately failing in clinical trials. [51,57]
The translational research efforts of our own group, which have focused on identifying molecular signatures and prognostic markers in PTCL, are directly impacted by this lack of representative models.11 While our work and that of others have identified promising therapeutic avenues, such as targeting aberrantly activated tyrosine kinases or epigenetic regulators, the ability to rigorously test these hypotheses in a disease-specific context is severely hampered by the absence of appropriate preclinical tools. [26,59]
5. Future Perspectives: Expanding the Preclinical Toolkit with Patient-Derived Xenografts (PDX)
Addressing the profound deficit in preclinical models for T-cell lymphomas requires a multi-pronged approach. First, a renewed, concerted international effort to establish novel cell lines from underrepresented PTCL subtypes is warranted, perhaps employing advanced co-culture or three-dimensional organoid systems that better recapitulate the TME. [44] The rigorous authentication and characterization of the T8ML-1 cell line serve as a valuable blueprint for such endeavors. [34]
However, the most promising and immediately impactful strategy is the development and widespread adoption of patient-derived xenograft (PDX) models. [53,60,61] Created by the direct implantation of fresh patient tumor tissue into severely immunodeficient mice (e.g., NOD-scid IL2Rgamma null or NSG mice), PDX models circumvent many of the critical limitations of traditional cell lines. [54,55,61] These models largely preserve the original tumor architecture, maintain the cellular and genetic heterogeneity of the primary disease, and retain key components of the human TME, including stromal and vascular elements. [52,55] Consequently, PDX models have demonstrated a significantly higher predictive value for clinical drug responses, making them a superior platform for translational research (Figure 3). [52,53]
The establishment of public repositories has been a transformative step, democratizing access to these powerful but resource-intensive models. The Public Repository of Xenografts (PRoXe), founded at the Dana-Farber Cancer Institute, is a landmark initiative that has generated and made available a large collection of well-characterized hematologic malignancy PDXs. [60,62,63] Crucially, this and other efforts have succeeded in creating stable, transplantable PDX models for T-cell lymphomas that have historically been impossible to culture in vitro. [62]
An AITL PDX has been successfully established, maintaining the characteristic CD4+/PD1+ immunophenotype of TFH cells and harboring hallmark mutations in genes such as TET2. [62] The ability to propagate this TME-dependent disease in vivo is a breakthrough, finally providing a renewable model to study AITL biology and test therapies targeted against its unique genetic drivers, such as mutations in RHOA, TET2, and IDH2. [5,39]. In addition, a PTCL-NOS PDX was shown to retain the clonal TCR gene rearrangement of the primary tumor and recapitulate a complex microenvironment containing non-malignant human T- and B-cells. [62]
PDX models of both systemic and breast implant-associated ALK-negative ALCL have been developed, providing invaluable tools for a disease with a poorer prognosis than its ALK-positive counterpart. [29,30]
Finally, a PDX model derived from an SS patient demonstrated appropriate disease dissemination, with malignant cells trafficking to the skin, spleen, and bone marrow, thus faithfully recapitulating the systemic nature of the disease. [62]
The availability of these models enables a paradigm shift in preclinical drug development toward "co-clinical trials" or "mouse avatar" studies. In this approach, cohorts of mice bearing PDXs from different patients can be used to conduct randomized, phase II-like trials, allowing for the robust evaluation of novel agents and the identification of biomarkers that predict response or resistance. [55,64]
6. Conclusions
The landscape of preclinical models for T-cell malignancies is one of stark contrasts. While a rich and well-characterized collection of cell lines exists for T-ALL, CTCL, and ALK-positive ALCL, this abundance is a direct reflection of a biological selection bias favoring tumors with potent, culture-permissive oncogenic drivers. For the majority of PTCL subtypes—many of which are rare, aggressive, and lack effective therapies—the preclinical toolkit is profoundly deficient. This gap has created a significant bottleneck in research, hindering our understanding of disease biology and impeding the development of novel targeted therapies. To break this cycle of neglect, the field must move beyond its reliance on a small, biased panel of historical cell lines. The future of translational research in T-cell lymphoma lies in the development and utilization of more clinically relevant models. Patient-derived xenografts, which preserve the heterogeneity and microenvironment of the original tumor, represent the most powerful tool currently available. The expansion and broad accessibility of PDX models through public repositories like PRoXe offer the most promising path forward to bridge the translational gap, enabling biologically-informed drug development and ultimately improving outcomes for patients with the full spectrum of T-cell lymphomas and leukemias. [60,63]
Author Contributions
Conceptualization, P.P.P.; methodology, L.C. and P.P.P..; investigation, L.C., V.T., P. C., C.I., S.S., G.C., F.D.P..; resources, P.P.P.; data curation, L.C., V.T., P. C.,; writing—original draft preparation, P.P.P., and L.C..; writing—review and editing, P.P.P. and G.T. supervision, P.P.P..; project administration, P.P.P.; funding acquisition, P.P.P. All authors have read and agreed to the published version of the manuscript.”.
Funding
The work reported in this publication was funded by the Italian Ministry of Health, RC-2025-2797263.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Prof. Pier Paolo Piccaluga is currently affiliated to the University of Nairobi (Nairobi, Kenya), and the University of Botswana (Gaborone, Botswana). VT and LC were supported in part by PRIN 2022NXK38S.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Drexler HG, Matsuo Y. Malignant hematopoietic cell lines: in vitro models for the study of natural killer cell leukemia-lymphoma. Leukemia. 2000 May;14(5):777–82. [CrossRef]
- Drexler HG, MacLeod RAF. Leukemia-lymphoma cell lines as model systems for hematopoietic research. Ann Med. 2003;35(6):404–12. [CrossRef]
- Drexler HG. Establishment and culture of leukemia-lymphoma cell lines. Methods Mol Biol. 2011;731:181–200.
- Sehn LH, Soulier J. Introduction to the review series on T-cell malignancies. Blood. 2017 Mar 2;129(9):1059–60. [CrossRef]
- Savage KJ, De Leval L. Erratum to: Introduction to the peripheral T-cell lymphoma review series: advances in molecular characterization, classification refinement and treatment optimization. Haematologica. 2024 Feb 1;109(2):693–4.
- Gootenberg JE, Ruscetti FW, Mier JW, Gazdar A, Gallo RC. Human cutaneous T cell lymphoma and leukemia cell lines produce and respond to T cell growth factor. J Exp Med. 1981 Nov 1;154(5):1403–18. [CrossRef]
- Kroeze E, Loeffen JLC, Poort VM, Meijerink JPP. T-cell lymphoblastic lymphoma and leukemia: different diseases from a common premalignant progenitor? Blood Adv. 2020 Jul 28;4(14):3466–73. [CrossRef]
- Lage LA de PC, Cabral TCDS, Costa R de O, Gonçalves M de C, Levy D, Zerbini MCN, et al. Primary nodal peripheral T-cell lymphomas: diagnosis and therapeutic considerations. Rev Bras Hematol Hemoter. 2015;37(4):277–84.
- Piccaluga PP, Fuligni F, De Leo A, Bertuzzi C, Rossi M, Bacci F, et al. Molecular profiling improves classification and prognostication of nodal peripheral T-cell lymphomas: results of a phase III diagnostic accuracy study. J Clin Oncol. 2013 Aug 20;31(24):3019–25. [CrossRef]
- Agostinelli C, Piccaluga PP, Went P, Rossi M, Gazzola A, Righi S, et al. Peripheral T cell lymphoma, not otherwise specified: the stuff of genes, dreams and therapies. J Clin Pathol. 2008 Nov;61(11):1160–7. [CrossRef]
- Piccaluga PP, Agostinelli C, Gazzola A, Mannu C, Bacci F, Sabattini E, et al. Prognostic markers in peripheral T-cell lymphoma. Curr Hematol Malig Rep. 2010 Oct;5(4):222–8.
- Bron D, De Leval L, Michiels S, Wittnebel S, EuroBloodNet for rare diseases. Hepatosplenic T-cell lymphoma: treatment challenges. Curr Opin Oncol. 2021 Sep 1;33(5):406–11. [CrossRef]
- Subbiah V, Kuravi S, Ganguly S, Welch DR, Vivian CJ, Mushtaq MU, et al. Precision therapy with anaplastic lymphoma kinase inhibitor ceritinib in ALK-rearranged anaplastic large cell lymphoma. ESMO Open. 2021 Aug;6(4):100172. [CrossRef]
- Gazdar AF, Carney DN, Bunn PA, Russell EK, Jaffe ES, Schechter GP, et al. Mitogen requirements for the in vitro propagation of cutaneous T-cell lymphomas. Blood. 1980 Mar;55(3):409–17.
- Spinella JF, Cassart P, Richer C, Saillour V, Ouimet M, Langlois S, et al. Genomic characterization of pediatric T-cell acute lymphoblastic leukemia reveals novel recurrent driver mutations. Oncotarget. 2016 Oct 4;7(40):65485–503. [CrossRef]
- Bardelli V, Arniani S, Pierini V, Di Giacomo D, Pierini T, Gorello P, et al. T-Cell Acute Lymphoblastic Leukemia: Biomarkers and Their Clinical Usefulness. Genes (Basel). 2021 Jul 23;12(8). [CrossRef]
- Moharram SA, Shah K, Kazi JU. T-cell Acute Lymphoblastic Leukemia Cells Display Activation of Different Survival Pathways. J Cancer. 2017;8(19):4124. [CrossRef]
- Burger R, Hansen-Hagge TE, Drexler HG, Gramatzki M. Heterogeneity of T-acute lymphoblastic leukemia (T-ALL) cell lines: suggestion for classification by immunophenotype and T-cell receptor studies. Leuk Res. 1999 Jan;23(1):19–27. [CrossRef]
- PDQ Adult Treatment Editorial Board. Peripheral T-Cell Non-Hodgkin Lymphoma Treatment (PDQ®): Health Professional Version. 2002.
- Starkebaum G, Loughran TP, Waters CA, Ruscetti FW. Establishment of an IL-2 independent, human T-cell line possessing only the p70 IL-2 receptor. Int J Cancer. 1991 Sep 9;49(2):246–53. [CrossRef]
- Popovic M, Sarin PS, Robert-Gurroff M, Kalyanaraman VS, Mann D, Minowada J, et al. Isolation and transmission of human retrovirus (human t-cell leukemia virus). Science. 1983 Feb 18;219(4586):856–9. [CrossRef]
- Irimia R, Piccaluga PP. Histone Deacetylase Inhibitors for Peripheral T-Cell Lymphomas. Cancers (Basel). 2024 Sep 30;16(19). [CrossRef]
- Piekarz RL, Robey RW, Zhan Z, Kayastha G, Sayah A, Abdeldaim AH, et al. T-cell lymphoma as a model for the use of histone deacetylase inhibitors in cancer therapy: impact of depsipeptide on molecular markers, therapeutic targets, and mechanisms of resistance. Blood. 2004 Jun 15;103(12):4636–43. [CrossRef]
- Epstein AL, Levy R, Kim H, Henle W, Henle G, Kaplan HS. Biology of the human malignant lymphomas. IV. Functional characterization of ten diffuse histiocytic lymphoma cell lines. Cancer. 1978 Nov;42(5):2379–91.
- Shimakage M, Dezawa T, Tamura S, Tabata T, Aoyagi N, Koike M, et al. A Ki-1-positive cell line expressing Epstein-Barr virus antigens, established from a child with Ki-1-positive lymphoma. Intervirology. 1993;36(4):215–24. [CrossRef]
- Piccaluga PP, Cascianelli C, Inghirami G. Tyrosine kinases in nodal peripheral T-cell lymphomas. Front Oncol. 2023;13:1099943. [CrossRef]
- Drexler HG, MacLeod RAF. Malignant hematopoietic cell lines: in vitro models for the study of anaplastic large-cell lymphoma. Leukemia. 2004 Oct;18(10):1569–71.
- Merz H, Lange K, Gaiser T, Müller A, Kapp U, Bittner C, et al. Characterization of a novel human anaplastic large cell lymphoma cell line tumorigenic in SCID mice. Leuk Lymphoma. 2002 Jan;43(1):165–72. [CrossRef]
- Marra A, Viale G, Pileri SA, Pravettoni G, Viale G, De Lorenzi F, et al. Breast implant-associated anaplastic large cell lymphoma: A comprehensive review. Cancer Treat Rev. 2020 Mar;84:101963.
- Chai KXY, Lee ECY, Li Z, Muhammad NAB, Lim JQ, Huang D, et al. Establishment and characterization of a novel patient-derived TP63-rearranged anaplastic large-cell lymphoma model PTCL-S1. Hum Cell. 2025 Jul 26;38(5):135. [CrossRef]
- Piccaluga PP, Agostinelli C, Tripodo C, Gazzola A, Bacci F, Sabattini E, et al. Peripheral T-cell lymphoma classification: the matter of cellular derivation. Expert Rev Hematol. 2011 Aug;4(4):415–25.
- Schwartz LF, Devine KJ, Xavier AC. Hepatosplenic T-cell lymphoma in children and adolescents. Blood Adv. 2025 Apr 22;9(8):1847–58. [CrossRef]
- Feng Y, Ma Y, Li T, Liu M, Hong Z, Yin Q, et al. Angioimmunoblastic T-cell lymphoma: a concise overview encompassing the pathogenetic, pathological, clinical, therapeutical characteristics, and recent advances. Clin Exp Med. 2025 Jun 25;25(1):218. [CrossRef]
- Ehrentraut S, Nagel S, Pommerenke C, Dirks WG, Quentmeier H, Kaufmann M, et al. Peripheral T-cell lymphoma cell line T8ML-1 highlights conspicuous targeting of PVRL2 by t(14;19)(q11.2;q13.3). Haematologica. 2017 Sep;102(9):e356–9. [CrossRef]
- del Mistro A, Leszl A, Bertorelle R, Calabro ML, Panozzo M, Menin C, et al. A CD30-positive T cell line established from an aggressive anaplastic large cell lymphoma, originally diagnosed as Hodgkin’s disease. Leukemia. 1994 Jul;8(7):1214–9.
- Piccaluga PP, Rossi M, Agostinelli C, Ricci F, Gazzola A, Righi S, et al. Platelet-derived growth factor alpha mediates the proliferation of peripheral T-cell lymphoma cells via an autocrine regulatory pathway. Leukemia. 2014 Aug;28(8):1687–97. [CrossRef]
- PDQ Adult Treatment Editorial Board. Mycosis Fungoides and Other Cutaneous T-Cell Lymphomas Treatment (PDQ®): Health Professional Version. 2002.
- Devine KJ, Schwartz L, El-Mallawany NK. Peripheral T-cell lymphoma-NOS in children and adolescents: a review from the Children’s Oncology Group NHL Committee. Blood Adv. 2025 Mar 25;9(6):1420–31. [CrossRef]
- Mohammed Saleh MF, Kotb A, Abdallah GEM, Muhsen IN, El Fakih R, Aljurf M. Recent Advances in Diagnosis and Therapy of Angioimmunoblastic T Cell Lymphoma. Curr Oncol. 2021 Dec 20;28(6):5480–98.
- Smith SD, Morgan R, Link MP, McFall P, Hecht F. Cytogenetic and immunophenotypic analysis of cell lines established from patients with T cell leukemia/lymphoma. Blood. 1986 Mar;67(3):650–6.
- Girardi T, Vicente C, Cools J, De Keersmaecker K. The genetics and molecular biology of T-ALL. Blood. 2017 Mar 2;129(9):1113–23.
- Havre PA, Dang LH, Ohnuma K, Iwata S, Morimoto C, Dang NH. CD26 expression on T-anaplastic large cell lymphoma (ALCL) line Karpas 299 is associated with increased expression of versican and MT1-MMP and enhanced adhesion. BMC Cancer. 2013 Nov 1;13:517.
- Tan SH, Bertulfo FC, Sanda T. Leukemia-Initiating Cells in T-Cell Acute Lymphoblastic Leukemia. Front Oncol. 2017;7:218.
- Mulloy JC. Peripheral T cell lymphoma: new model + new insight. J Exp Med. 2010 May 10;207(5):911–3.
- Gaulard P, de Leval L. The microenvironment in T-cell lymphomas: emerging themes. Semin Cancer Biol. 2014 Feb;24:49–60.
- de Leval L, Rickman DS, Thielen C, Reynies A de, Huang YL, Delsol G, et al. The gene expression profile of nodal peripheral T-cell lymphoma demonstrates a molecular link between angioimmunoblastic T-cell lymphoma (AITL) and follicular helper T (TFH) cells. Blood. 2007 Jun 1;109(11):4952–63. [CrossRef]
- Piccaluga PP, Agostinelli C, Califano A, Carbone A, Fantoni L, Ferrari S, et al. Gene expression analysis of angioimmunoblastic lymphoma indicates derivation from T follicular helper cells and vascular endothelial growth factor deregulation. Cancer Res. 2007 Nov 15;67(22):10703–10.
- Piccaluga PP, Agostinelli C, Califano A, Rossi M, Basso K, Zupo S, et al. Gene expression analysis of peripheral T cell lymphoma, unspecified, reveals distinct profiles and new potential therapeutic targets. J Clin Invest. 2007 Mar;117(3):823–34.
- Iqbal J, Wright G, Wang C, Rosenwald A, Gascoyne RD, Weisenburger DD, et al. Gene expression signatures delineate biological and prognostic subgroups in peripheral T-cell lymphoma. Blood. 2014 May 8;123(19):2915–23.
- Kuczynski EA, Morlino G, Peter A, Coenen-Stass AML, Moss JI, Wali N, et al. A preclinical model of peripheral T-cell lymphoma GATA3 reveals DNA damage response pathway vulnerability. EMBO Mol Med. 2022 Jun 8;14(6):e15816. [CrossRef]
- Andreu-Sanz D, Gregor L, Carlini E, Scarcella D, Marr C, Kobold S. Predictive value of preclinical models for CAR-T cell therapy clinical trials: a systematic review and meta-analysis. J Immunother Cancer. 2025 Jun 12;13(6).
- Goto T. Patient-Derived Tumor Xenograft Models: Toward the Establishment of Precision Cancer Medicine. J Pers Med. 2020 Jul 18;10(3). [CrossRef]
- Hidalgo M, Amant F, Biankin A V, Budinská E, Byrne AT, Caldas C, et al. Patient-derived xenograft models: an emerging platform for translational cancer research. Cancer Discov. 2014 Sep;4(9):998–1013.
- Idrisova KF, Simon HU, Gomzikova MO. Role of Patient-Derived Models of Cancer in Translational Oncology. Cancers (Basel). 2022 Dec 26;15(1). [CrossRef]
- Jung J, Seol HS, Chang S. The Generation and Application of Patient-Derived Xenograft Model for Cancer Research. Cancer Res Treat. 2018 Jan;50(1):1–10.
- Yu K, Chen B, Aran D, Charalel J, Yau C, Wolf DM, et al. Comprehensive transcriptomic analysis of cell lines as models of primary tumors across 22 tumor types. Nat Commun. 2019 Aug 8;10(1):3574. [CrossRef]
- Xing Y, Huang J, Zhang Y, Wang Y, Qi S. Advancing the understanding and management of angioimmunoblastic T-cell lymphoma: insights into its pathogenesis, clinical features, and emerging therapeutic strategies. Front Oncol. 2025;15:1479179. [CrossRef]
- Mhaidly R, Krug A, Gaulard P, Lemonnier F, Ricci JE, Verhoeyen E. New preclinical models for angioimmunoblastic T-cell lymphoma: filling the GAP. Oncogenesis. 2020 Aug 14;9(8):73. [CrossRef]
- Pileri SA, Piccaluga PP. New molecular insights into peripheral T cell lymphomas. J Clin Invest. 2012 Oct;122(10):3448–55.
- Huo KG, D’Arcangelo E, Tsao MS. Patient-derived cell line, xenograft and organoid models in lung cancer therapy. Transl Lung Cancer Res. 2020 Oct;9(5):2214–32. [CrossRef]
- Steel C, James ER, Matthews JD, Turner SD. Establishing Patient-Derived Xenograft (PDX) Models of Lymphomas. Methods Mol Biol. 2025;2865:429–48.
- Christie AL, Ng SY, Koch R, Christodoulou AN, DeSouza T, Murakami MA, et al. T-Cell Lymphoma Patient-Derived Xenografts and Newly Developed Cell Lines Recapitulate Aspects of Disease Biology and Represent Novel Tools for Preclinical Drug Development. Blood. 2016 Dec 2;128(22):3015–3015. [CrossRef]
- Murakami MA, Christodoulou AN, Christie AL, DeSouza T, Louissaint A, Vojinovic U, et al. Proxe: A Public Repository of Xenografts to Facilitate Studies of Biology and Expedite Preclinical Drug Development in Leukemia and Lymphoma. Blood. 2015 Dec 3;126(23):3252–3252.
- Townsend EC, Murakami MA, Christodoulou A, Christie AL, Köster J, DeSouza TA, et al. The Public Repository of Xenografts Enables Discovery and Randomized Phase II-like Trials in Mice. Cancer Cell. 2016 Jul 11;30(1):183.
Figure 1.
Disparity in Preclinical Models: Clinical Incidence vs. In Vitro Representation of T-Cell Malignancies. Panel A illustrates the approximate clinical incidence of major T-cell malignancy subtypes. Panel B shows the number of currently available, authenticated cell line models for these subtypes. The marked imbalance highlights a major research gap, particularly for PTCL-NOS, AITL, and HSTCL, which lack suitable in vitro models for mechanistic and preclinical studies.
Figure 1.
Disparity in Preclinical Models: Clinical Incidence vs. In Vitro Representation of T-Cell Malignancies. Panel A illustrates the approximate clinical incidence of major T-cell malignancy subtypes. Panel B shows the number of currently available, authenticated cell line models for these subtypes. The marked imbalance highlights a major research gap, particularly for PTCL-NOS, AITL, and HSTCL, which lack suitable in vitro models for mechanistic and preclinical studies.
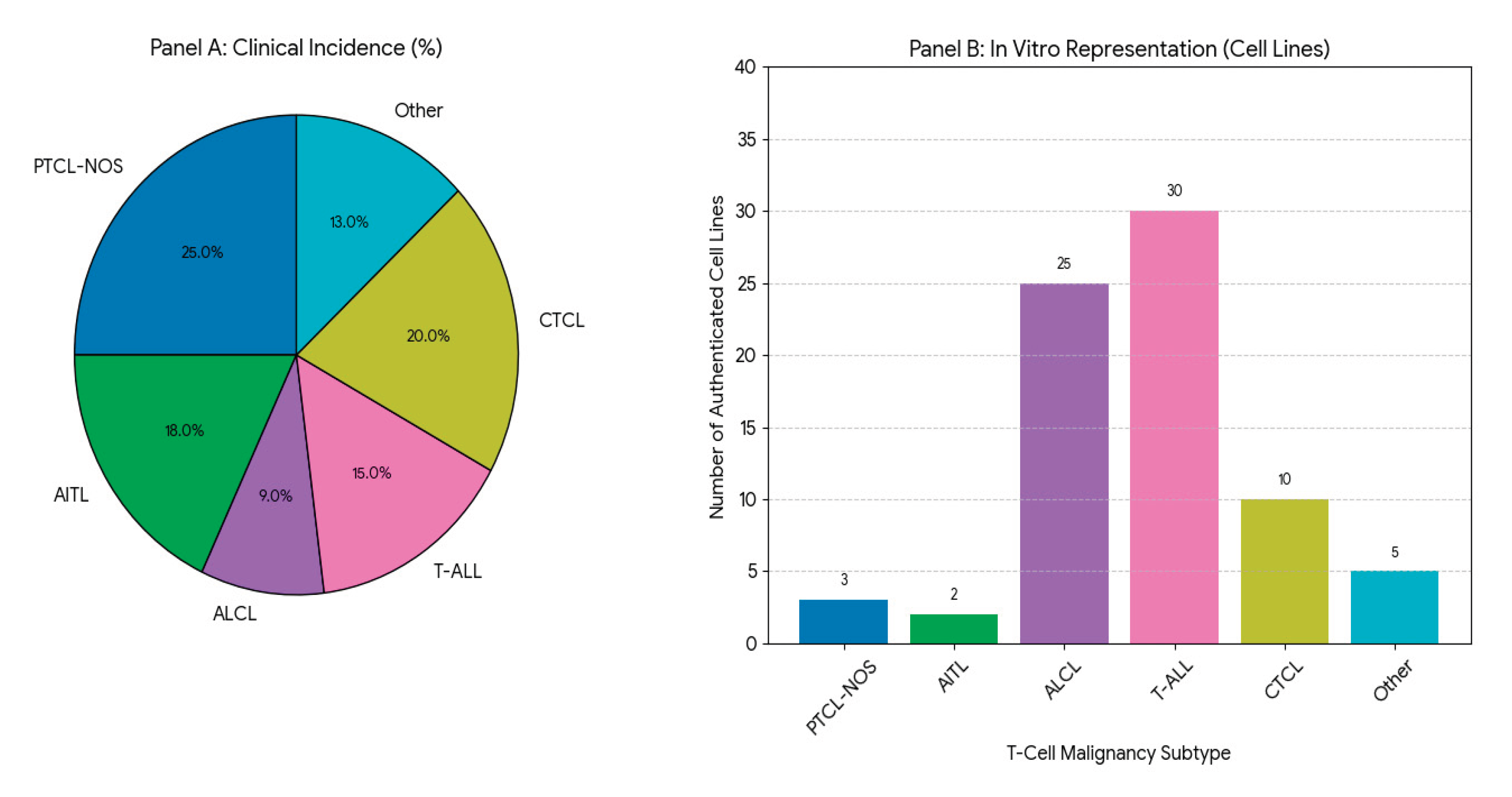
Figure 2.
Constitutively active signaling pathways in amenable T-cell lymphoma subtypes. This diagram illustrates potent, self-sustaining oncogenic signaling mechanisms in ALCL (NPM1–ALK fusion) and T-ALL (NOTCH1 mutation) that drive robust cell line establishment and survival in vitro. In ALCL, the NPM1–ALK fusion results in constitutive activation of the JAK/STAT3 and PI3K/AKT/mTOR pathways, promoting MYC-driven proliferation and survival. In T-ALL, activating NOTCH1 mutations lead to ligand-independent cleavage and release of the NOTCH1 intracellular domain (NICD), which interacts with CSL/MAML to upregulate target genes such as MYC and CYCLIN D1, sustaining uncontrolled cell growth. This highlights a biological selection bias favoring cell line establishment from subtypes with strong autonomous signaling activity, such as ALCL and T-ALL.
Figure 2.
Constitutively active signaling pathways in amenable T-cell lymphoma subtypes. This diagram illustrates potent, self-sustaining oncogenic signaling mechanisms in ALCL (NPM1–ALK fusion) and T-ALL (NOTCH1 mutation) that drive robust cell line establishment and survival in vitro. In ALCL, the NPM1–ALK fusion results in constitutive activation of the JAK/STAT3 and PI3K/AKT/mTOR pathways, promoting MYC-driven proliferation and survival. In T-ALL, activating NOTCH1 mutations lead to ligand-independent cleavage and release of the NOTCH1 intracellular domain (NICD), which interacts with CSL/MAML to upregulate target genes such as MYC and CYCLIN D1, sustaining uncontrolled cell growth. This highlights a biological selection bias favoring cell line establishment from subtypes with strong autonomous signaling activity, such as ALCL and T-ALL.
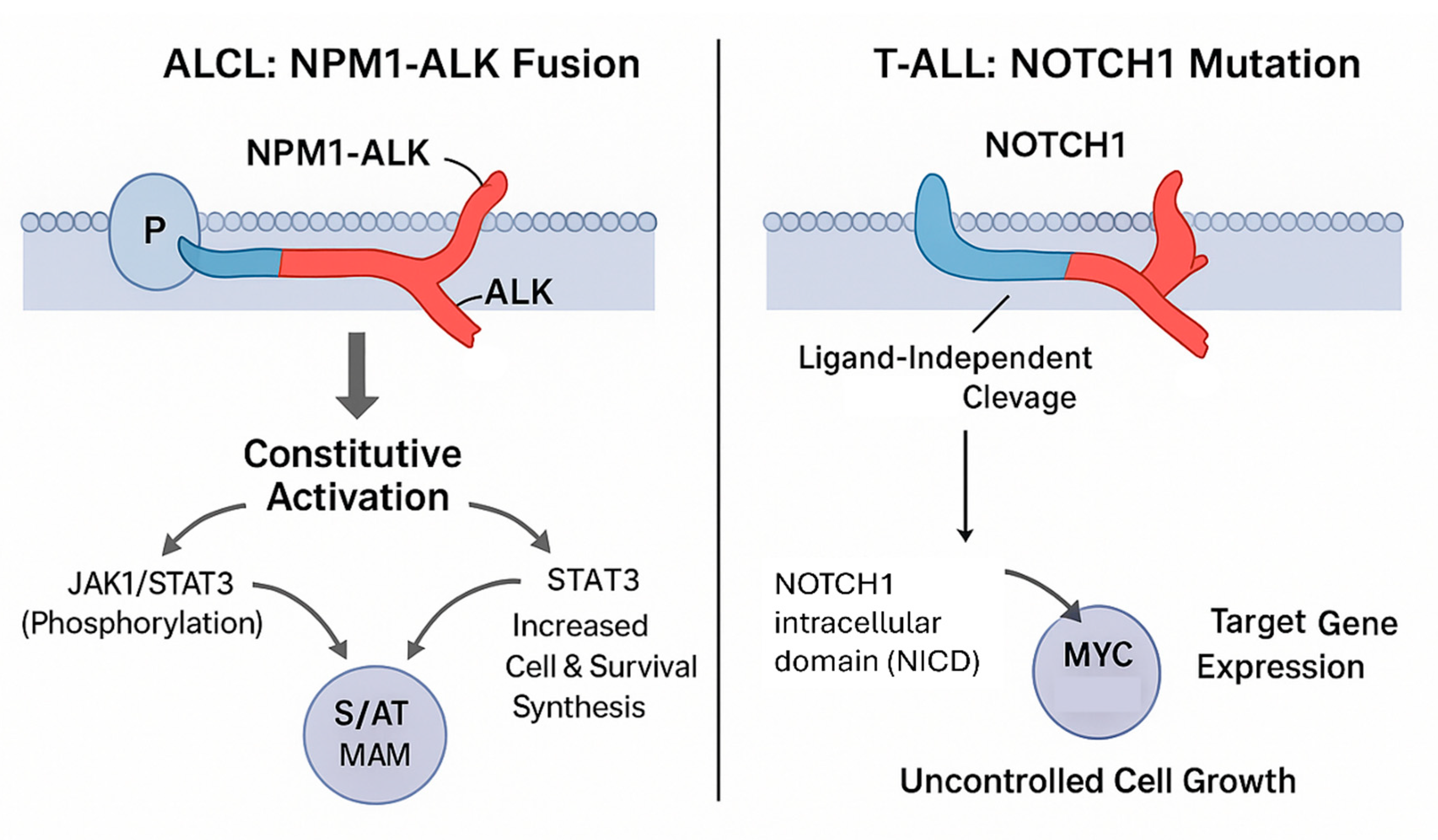
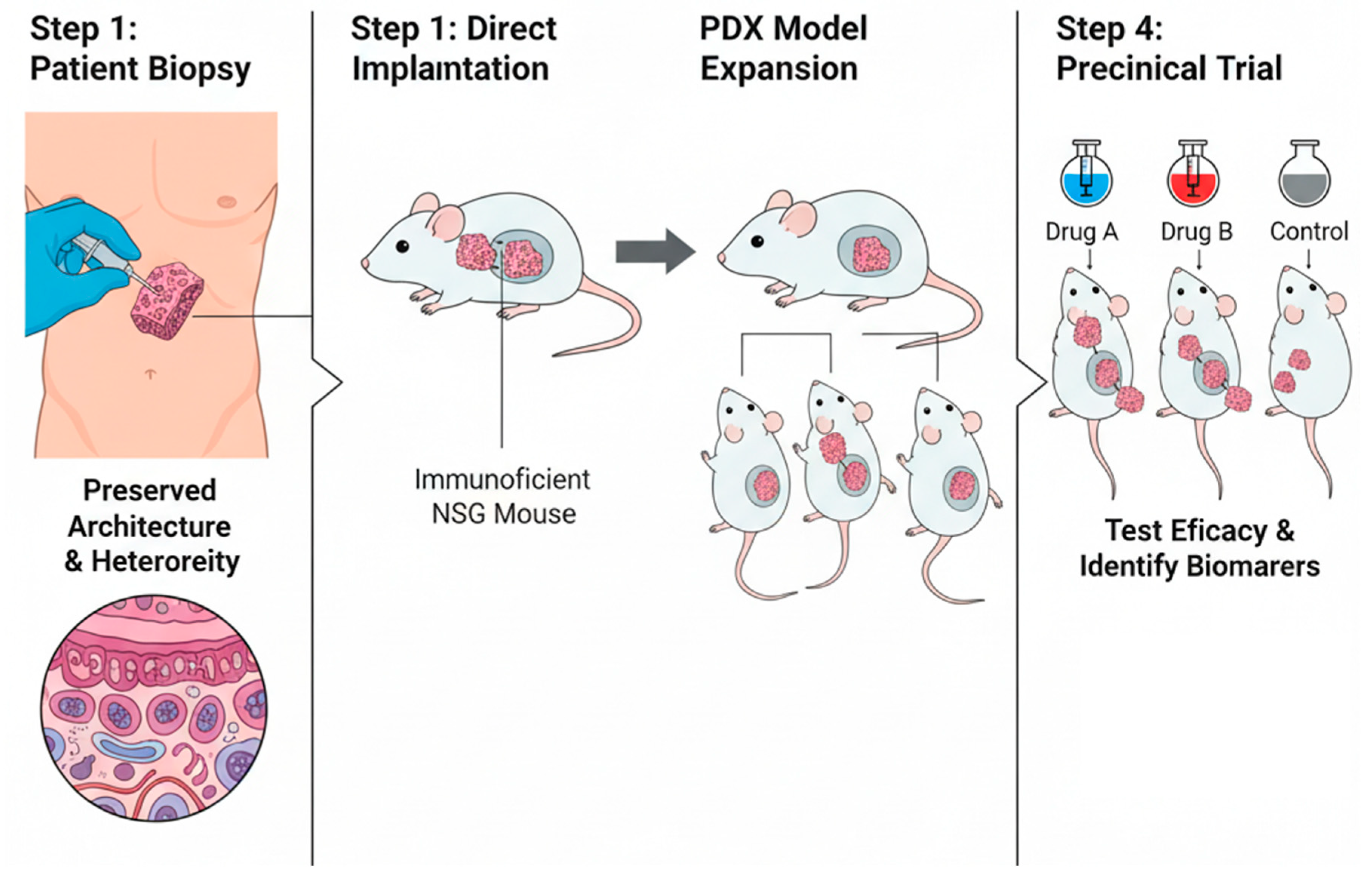
Figure 3.
Schematic representation of Patient-Derived Xenograft (PDX) generation and application in translational research. Patient tumor biopsies are directly implanted into immunodeficient NSG mice, preserving the original tumor architecture and heterogeneity. After successful engraftment, PDX models are expanded for subsequent preclinical testing. These “mouse avatar” models allow evaluation of drug efficacy (e.g., Drug A, Drug B, Control) and identification of predictive biomarkers of response or resistance.
Figure 3.
Schematic representation of Patient-Derived Xenograft (PDX) generation and application in translational research. Patient tumor biopsies are directly implanted into immunodeficient NSG mice, preserving the original tumor architecture and heterogeneity. After successful engraftment, PDX models are expanded for subsequent preclinical testing. These “mouse avatar” models allow evaluation of drug efficacy (e.g., Drug A, Drug B, Control) and identification of predictive biomarkers of response or resistance.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



