Submitted:
15 January 2026
Posted:
21 January 2026
You are already at the latest version
Abstract
Native to the lakes and wetlands of southern Mexico, the axolotl (Ambystoma mexicanum) is a neotenic salamander that is renowned for its remarkable capacity for regeneration. Beyond regeneration, increasing evidence suggests that axolotls may also display resistance to cancer, making them a valuable model for exploring mechanisms of tumor suppression. This review provides an updated overview of molecular and physiological traits potentially underlying this resistance. Central aspects include their permanent larval state, high cellular plasticity, and tightly regulated cell proliferation during regeneration, all of which may limit malignant transformation. Additional mechanisms considered are their low metabolic rate, reduced thyroid hormone levels, decreased insulin sensitivity, presence of natural antitumor compounds, distinctive genomic and epigenetic features, and a robust innate immune system. Particular emphasis is placed on how these mechanisms may counteract established cancer hallmarks. Studying these features not only deepens understanding of axolotl biology but may also inspire novel strategies for cancer prevention and therapy in humans.
Keywords:
regeneration
; metamorphosis
; resistance to cancer development
; molecular mechanisms
1. Introduction
The axolotl, a species of salamander belonging to the genus Ambystoma (commonly known as the mole salamander) and specifically the species Ambystoma mexicanum, is native to the lakes and wetlands of southern Mexico [1]. Recognizable by their feathery external gills and perpetual “smile”, axolotls have become popular around the world. Their extraordinary ability to regenerate complex body parts, such as limbs, eyes, heart, spinal cord and even parts of the brain, has made them the subject of intense scientific and genetic research [2]. Notably, this dual status has created a conservation paradox: while axolotls abound in laboratories and aquariums around the world, their wild populations have plummeted due to severe habitat degradation, leaving the species critically endangered in the wild [3].
On the other hand, cancer represents a major global challenge, with profound societal, public health, and economic impacts. It accounts for nearly one in six deaths worldwide (16.8%) and approximately one in four deaths (22.8%) caused by noncommunicable diseases (NCDs) [4]. Cancer is characterized by the abnormal growth of cells that expand beyond their normal boundaries. These uncontrolled cells can invade nearby tissues and have the potential to spread to distant organs through a process known as metastasis [5,6].
Carcinogenesis is a gradual, multistep process driven by the accumulation of genetic alterations that disrupt normal cellular regulation and endow malignant cells with specific traits collectively known as the hallmarks of cancer [7]. Core hallmarks include the ability to sustain proliferative signaling by generating continuous growth cues, evade growth suppressors that normally act as brakes on cell division, and resist programmed cell death, ensuring survival despite damage or stress. Cancer cells also achieve replicative immortality by maintaining telomeres and bypassing senescence, induce or co-opt angiogenesis to secure oxygen and nutrients, and activate invasion and metastasis, spreading from their tissue of origin to distant organs. At the metabolic level, cancer cells reprogram energy pathways to fuel rapid growth even under adverse conditions, while simultaneously evading immune destruction, allowing them to persist despite immune surveillance. Beyond these established features, additional mechanisms are increasingly recognized. Cancer cell plasticity is increasingly appreciated as a key factor, supported by mechanisms such as non-mutational epigenetic reprogramming and variations in organ- or tissue-specific microbiomes, which may serve as novel enablers of hallmark capabilities. Moreover, tumor-promoting inflammation emerges as a critical process that cooperates with genetic mutations and other oncogenic alterations to sustain malignant progression. Finally, senescent cells of diverse origins, including both tumor cells and stromal cells, have been identified as functional contributors to cancer development and progression [7].
Several animal species in nature have demonstrated remarkable resistance to cancer, a disease often considered an inevitable consequence of complex life [8]. Notable examples include the naked mole rat [9], the blind mole rat [10], bats [11], whales [12], and elephants [13], all of which exhibit unique biological traits that seem to protect them against tumor development.
However, what about axolotls? The idea that these amphibians might also be resistant to cancer has circulated for many years (since 1971, it was suggested) [14]. However, scientific evidence to support this claim is limited. In fact, as more researchers begin studying salamanders—including axolotls—reports of cancer cases in these animals are increasing. Furthermore, axolotls are rarely subjected to systematic cancer monitoring, and most individuals kept in laboratories are only observed until around 3 to 4 years of age. In the wild, axolotls generally live 5 to 6 years but can live up to 21 years in captivity [15]. As a result, many potential cancer cases may go undetected, especially those that develop later in life.
Particularly intriguing are the biological factors that may contribute to this resilience (Table 1). Among these, the species’ ability to retain a larval state throughout life —a form of “perpetual youth”—without showing the typical decline associated with aging could play a key role [16]. This is accompanied by a high degree of cellular plasticity, allowing its cells to adapt and reprogram in response to external stimuli [17]. Another remarkable characteristic of the axolotl is its extraordinary capacity to regenerate complex tissues and organs without developing tumors. This regenerative capacity requires precise control over cell proliferation and mechanisms that prevent uncontrolled growth, as well as early suppression of potentially cancerous cells [18].
With this in mind, the aim of this review was to compile current knowledge and provide an overview of the molecular mechanisms that may underlie resistance of the axolotl to cancer (Figure 1). Key factors under analysis include the axolotl’s neoteny (retention of juvenile characteristics in a sexually mature adult animal), its remarkable cellular plasticity, and the tightly regulated cell proliferation observed during regeneration. Other factors not previously considered are their low metabolic rate, their reduced thyroid hormone levels, their lower insulin sensitivity, the presence of natural antitumor compounds, their unique genomic and epigenetic traits, and the characteristics of their innate immune system.
2. Tumor Cases in Axolotls
Few cases of spontaneous tumors have been described in the axolotl, and a relatively small proportion of them have been identified as malignant. Early and most commonly described cases include cutaneous and subcutaneous melanomas [20,21]. This could be explained by the fact that axolotls, particularly wild-type strains, possess a large number of pigment-producing cells called melanophores [22]. These cells are especially sensitive to mutagenic changes due to their high metabolic activity and the generation of reactive oxygen species (ROS) during melanin production [23]. As a result, melanophores are more susceptible to malignant transformation. Interestingly, in one group of animals, melanoma growth was observed to occur in parallel with tissue regeneration. However, the infiltration of melanotic tissue into the regeneration zone was limited and temporary, so it did not interfere with the process and resulted in the formation of normal tissue [24]. A case of chromatophoroma (a tumor also arising from pigment-producing cells) was reported in the tail of a 3-year-old female axolotl [25]. Another report documented a teratoma arising in the dorsal muscles of the proximal tail in a 2.5-year-old animal [26]. In addition, spontaneous mastocytomas have been observed in older members (11–17 years old) of an inbred laboratory colony [27]. Similarly, a case series reported ten instances of spontaneous myeloid leukemia in genetically related adult axolotls that were housed together in a zoo [28]. Another reported case described the development of a malignant abdominal tumor, identified as either lymphangiosarcoma or lymphosarcoma [29]. Two cases of olfactory neuroblastoma have been described in the axolotl, presenting as masses in the oral and nasal cavities [30,31]. Over a 27-year period, 16 cases of testicular tumors were reported within a laboratory axolotl colony in which all individuals were genetically related [32].
An important consideration is that human influence may have played a role in increasing cancer susceptibility in some of the reported cases. Observations from some laboratory colonies suggest a familial link (inbred) in cases where tumors such as mastocytomas [27], testicular tumors [32], melanoma [24], and acute myeloid leukemia [28] have occurred. It is possible that a small founder population in these colonies caused inbreeding and reduced genetic diversity, thus increasing the susceptibility of these individuals to develop tumors and cancer.
On the other hand, experimental induction of tumors in axolotls has also shown signs of resistance to cancer development. When exposed to polycyclic hydrocarbons such as dibenzanthracene and methylcholanthrene, the animals showed a low incidence of papilloma tumors (28.83%). The occurrence of malignant tumors was even lower (1.8%) and required a long latency period of 2 to 3 years to develop [14]. A case of lymphosarcoma was observed in an axolotl during experiments with allogeneic skin transplants. The cancer emerged after the third round of transplants and could only be transmitted through viable tumor cells to individuals that were both immunodeficient and genetically compatible. Interestingly, very young individuals of allogeneic strains acted as “regressors”, allowing transient tumor growth followed by regression, whereas adults of these same strains were “rejecters” and did not develop tumors at all [33].
3. Possible Molecular Mechanisms Involved in the Reduction of Cancer Incidence in Axolotls
3.1. Habitat and Low Metabolic Rate
Tumor growth has been reported to slow under low-temperature conditions [34]. Axolotls, which naturally inhabit cool environments (15–20 °C), could benefit from this factor as part of their arsenal against cancer development. Low temperatures reduce the metabolic rate in axolotls, and a lower basal metabolic rate may contribute to a decreased risk of cancer [35]. Axolotls exhibit a lower metabolic rate compared to mammals (Table 1) [36,37], which may create an unfavorable environment for cancer cells whose reprogrammed metabolism must sustain rapid proliferation—a well-recognized hallmark of cancer (Table 2). Similarly, exposure to cold can reduce inflammation [38], including tumor-promoting inflammation, representing inhibition of another established hallmark of cancer (Table 2).
Interestingly, the axolotl tumor protein p53 (TP53) harbors several amino acid substitutions, including changes at residues T155 and V157, which, in humans, are associated with reduced tumor suppressor activity. Notably, some of these changes may have been selected to maintain TP53 activity at the low physiological temperatures of this amphibian [39]. TP53 is known as the “guardian of the genome” because it preserves DNA integrity by monitoring cellular stress and genetic damage. When such damage is detected, TP53 acts as a master regulator of cell fate by halting cell-cycle progression to allow DNA repair or, if the damage is irreparable, by inducing programmed cell death (apoptosis). Through these mechanisms, TP53 prevents the propagation of genetically unstable cells and plays a central role in cancer suppression [40].
3.2. Low Levels of Thyroid Hormones
Although axolotls typically exhibit lifelong neoteny both in the wild and in captivity, metamorphosis can be experimentally triggered by the administration of thyroid hormones [41]. It is well established that neoteny in axolotls is a consequence of the low activity of the hypothalamo-pituitary-thyroid (HPT) axis [42]. In this context, thyroid hormones play a central role in regulating metabolism, growth, and numerous other physiological processes [43]. Some evidence indicates that both subclinical and clinical hyperthyroidism are associated with an increased risk of various solid tumors. Conversely, hypothyroidism has been linked to reduced tumor aggressiveness and, in some cases, a delayed onset of cancer [44]. In particular, thyroid hormones regulate key processes associated with cancer hallmarks, including metabolism, angiogenesis, cell proliferation, and survival (Table 2) [45]. In axolotls, low thyroid hormone levels may dampen growth signaling, restrict angiogenesis, prevent metabolic reprogramming, and increase susceptibility to apoptosis [46]. In this context, the naturally low levels of thyroid hormones in axolotls may play a role in their apparent resistance to cancer. However, it remains unclear whether cancer incidence is significantly higher in terrestrial salamanders that have undergone metamorphosis triggered by thyroid hormones, as comparative data are limited. The naked mole rat, another organism with exceptional resistance to cancer, also exhibits low levels of thyroid hormones [9,47].
3.3. Low Insulin Sensitivity
Hyperinsulinemia is associated with an increased risk of various types of cancer and increased mortality from cancer [48]. Unlike mammals, axolotls display lower blood glucose levels and are also less sensitive to insulin, resulting in slow blood glucose regulation [36]. Since most cancer cells rely heavily on glycolysis to support their rapid growth, invasion, and metastasis [49], the combination of low body temperature, reduced metabolic activity, and impaired glucose-insulin responsiveness in axolotls may create a physiological environment that is less conducive (or even hostile) to cancer cell proliferation and metastasis (Table 2). In addition, the insulin and insulin-like growth factor (IGF-1) pathways act as potent mitogenic and anti-apoptotic signals (Figure 2) [48]. Therefore, reduced activation of these pathways can impair uncontrolled cell proliferation and resistance to cell death, both of which represent central hallmarks of cancer (Table 2).
3.4. Intrinsic Natural Compounds with Antitumor Activity
Antimicrobial peptides isolated from axolotl skin mucus have demonstrated antitumor activity against breast cancer cells at concentrations as low as 1 µg/mL, primarily by upregulating tumor suppressor genes and downregulating oncogenes [50]. The antitumor activity of crude axolotl extract has also been observed in vitro at concentrations of 2 mg/mL, where it induced apoptosis and promoted differentiation toward the granulocytic lineage in a human acute myeloid leukemia cell line [51]. Breast cancer cells were reprogrammed using axolotl prophase oocyte extracts (5000 cells/μL extract). This reprogramming induced DNA demethylation and removed repressive histone marks at the promoters of tumor suppressor genes. As a result, cancer cell proliferation was reduced in vitro, along with decreased tumor growth in mouse xenograft models [52].
3.5. Genome and Epigenetic Characteristics
The axolotl genome is extraordinarily large, with approximately 32 billion base pairs (bp), about ten times larger than the human genome (Table 1) [19]. Although replication of such a large genome could increase the risk of mutations, axolotls appear to possess highly efficient and responsive DNA repair mechanisms that help maintain genomic stability [53,54,55]. In this regard, axolotl TP53 was phosphorylated and activated following exposure to UV irradiation (100 J/m2) or treatment with an alkylating agent, indicating a conserved DNA damage response pathway [39].
In addition, despite this vast difference in genome size, the number of protein-coding genes in axolotls is comparable to that of other vertebrates with much smaller genomes. Axolotl genome expansion is largely attributed to a prolonged period of transposon activity, followed by a more recent and ongoing burst of repeat element proliferation. It is especially enriched in repetitive sequences, dominated by diverse classes of long terminal repeat (LTR) retroelements, some of which exceed 10,000 bp in length. This high repeat content has significantly impacted genome architecture, leading to a median intron length of 22,759 bp (approximately 13, 16, and 25 times longer than the median intron sizes in human [1750 bp], mouse [1469 bp], and frog [906 bp], respectively). Intergenic regions have also expanded substantially, ranging from 12 to 17 times larger than those in these other species [19]. This suggests that mutations in the axolotl genome are more likely to occur within LTR retroelements and other non-coding regions rather than in coding sequences. It remains unknown whether these genomic sequences contain unique features specific to the axolotl that might contribute to its ability to avoid cancer development, such as distinctive non-coding RNAs or genomic regions that act as “mutation buffers,” preventing potentially harmful mutations from affecting functionally or phenotypically important sites in cells. In this sense, it has been suggested that some redundancies in DNA, such as certain introns and repetitive sequences, may dampen sequence-dependent conformational effects, preventing these effects from interfering with the accurate reading of genetic information [56].
The exact number of tumor suppressor genes in the axolotl is not yet known or fully cataloged, but it is presumed to be comparable to that in humans. The most recent genome assembly report lists 39,753 transcripts based on homologous sequences and 1398 transcripts that have no matches in BLAST (basic local alignment search tool) but possess an open reading frame of at least 200 amino acids, suggesting that they could represent new protein-coding genes [19]. By examining the list of axolotl genes with sequences homologous to those of humans (using www.axolotl-omics.org [57]), approximately 150 well-characterized tumor suppressor genes were identified (e.g., TP53, RB1, PTEN, BRCA1, BRCA2, APC, and CDKN2A). However, further validation is required to confirm both their identity and functional activity. Additionally, some of the axolotl’s unique genes might include species-specific tumor suppressors that contribute to its enhanced protection, though this possibility remains to be explored.
Another noteworthy aspect is that axolotls exhibit minimal changes in global DNA methylation levels throughout their life span, suggesting a remarkably stable epigenome compared to other species [58]. This epigenetic stability may contribute to their resistance to age-related diseases, including cancer. DNA methylation plays a key role in tumorigenesis, as hypermethylation of promoter regions can silence tumor suppressor genes, while hypomethylation of regulatory sequences can activate proto-oncogenes and retrotransposons [59].
Taken together, these findings suggest that axolotls possess genomic and epigenetic features that help protect them against key cancer hallmarks, including sustaining proliferative signaling, evading growth suppressors, enabling replicative immortality, activating invasion and metastasis, and reprogramming cellular metabolism (Table 2). Efficient DNA repair systems likely minimize genome instability and mutations. This stability may prevent proto-oncogene activation, maintain tumor suppressor gene expression, and ensure proper telomere regulation. The complementary action of tumor suppressor genes may further inhibit the evasion of growth suppression. Finally, robust epigenetic regulation appears to shield axolotls from cancer-associated epigenetic reprogramming.
3.6. Innate Immune System
Like higher vertebrates, the axolotl possesses both innate and adaptive branches of the immune system comprising the major classes of myeloid (neutrophils, eosinophils, and basophils) and lymphoid (T and B cells) lineages observed in mammals [60,61,62]. However, the overall diversity and functional specialization of these immune cell subtypes remain insufficiently characterized.
Axolotls possess a robust innate immune system [63], which may contribute to their apparent resistance to cancer, given that the ability to evade immune-mediated destruction is a fundamental hallmark of cancer (Table 2 and Figure 3) [64]. In this sense, non-specific cytotoxic cell receptor protein 1 (NCCRP1) expression has been detected in several axolotl tissues, including peripheral blood mononuclear cells, skin, lung, spleen, and limb blastema [64]. NCCRP1 is involved in antigen recognition and is a key receptor on nonspecific cytotoxic cells (NCCs), which mediate lysis of target cells (tumor cells and protozoan parasites) by binding to their natural ligand, the natural killer target antigen (NK Tag) [65]. NCCs are considered evolutionary precursors of mammalian natural killer (NK) cells [66] and may represent a component of the axolotl’s antitumoral immune system. Furthermore, axolotl major histocompatibility complex (MHC) class I molecules exhibit strong conservation of nonpolymorphic peptide-binding regions on the α chain, along with a high degree of variability at other amino acid sites [62]. This structural arrangement suggests that axolotl class I MHC molecules are capable of presenting a wide range of antigenic epitopes, including potentially tumor-associated antigens.
Another remarkable feature of the axolotl is its ability to regenerate the thymus de novo after its complete removal [67]. The thymus is the main organ for T lymphocyte development and a key site for establishing self-tolerance and adaptive immune function [68]. This regenerative process of the axolotl completely restores thymus morphology, cellular diversity, and immune function, an exceptional ability among vertebrates [67]. In contrast, mammals undergo age-related thymic involution, leading to a progressive decline in T cell production and overall immune competence [69]. The absence of thymic involution in the axolotl may therefore allow it to maintain robust immune surveillance and effective antitumor activity for a longer period of its life.
On the other hand, axolotls exhibit a markedly attenuated adaptive immune response compared to other vertebrates [70]. Only two classes of immunoglobulins (IgM and IgY) have been identified in these animals, and both lack anamnestic (memory) responses [71]. As a consequence, axolotls generate antibodies more slowly and show minimal humoral or cytotoxic responses to viral infections [71,72]. This limited adaptive immunity is thought to be related to their neotenic state, as induction of metamorphosis has been shown to enhance humoral responses [73]. Despite this, the attenuated adaptive immune response of axolotls could offer some advantages in the context of cancer. For example, inflammatory responses, often associated with multiple stages of tumorigenesis [74,75], would be more tightly regulated and less likely to become chronic, thus counteracting, at least to some extent, another important hallmark of cancer: tumor-promoting inflammation (Table 2).
3.7. Regeneration Capacity
The axolotl’s regenerative capacity is extraordinary, enabling it to restore complex structures such as limbs, the tail, parts of the eye and brain, and several internal organs [2]. In axolotls, the regenerative-permissive environment, comprising both cellular and acellular components, shares several tumor-like features. In fact, axolotl blastema cells can transiently express antigens similar to oncofetal antigens found in tumors, but their strict temporal regulation and immunological integration prevent malignant transformation and immune escape [76]. Notably, the initiation of proliferative signaling and blastema formation in axolotls involves a transient dedifferentiation process, during which mature cells revert to a progenitor-like state. These dedifferentiated cells display features commonly associated with cancer, including increased proliferation, proto-oncogene expression, and chromatin remodeling [16]. However, unlike the uncontrolled growth seen in tumors, axolotl regeneration is tightly regulated. The process is guided by precise control of cell cycle regulators to ensure balanced and orderly tissue restoration [77]. Similarly, Ty3 retrotransposon expression is suppressed during regeneration [78]. Additionally, these cells retain a memory of their original tissue identity and positional information, reducing the risk of malignant transformation [2]. Cancer cells often lose their normal identity and structure [79]. The remarkably stable epigenetic landscape of the axolotl probably also plays a crucial role in protecting against transformation [58].
Another key difference is that tumors are marked by extensive extracellular matrix (ECM) deposition, remodeling, and cross-linking, which contribute to fibrosis, stiffen the surrounding stroma, and promote malignant progression. This stiffened stroma supports tumor cell growth, survival, and migration, while also driving mesenchymal transition [80]. In contrast, axolotl wounds display only a temporary fibrotic response, ultimately leading to the regeneration of normal tissue architecture [81]. This regenerative response is marked by reduced levels of fibronectin and elevated levels of tenascin-C, creating a distinct tissue environment compared to mammalian wound repair [82]. Following limb amputation in axolotls, the number of macrophages and neutrophils increases at the wound site. Neutrophils play a key role in clearing cellular and molecular debris and promote collagen degradation by stimulating the production of matrix metalloproteinases. They also help activate anti-inflammatory macrophages by suppressing NF-κB signaling [83]. Macrophages, in turn, are essential for dampening the inflammatory response. Their presence is particularly critical during the early stages of heart regeneration, where they help regulate fibroblast activity and prevent excessive fibrosis [84]. On the other hand, ROS may also play a key role in extracellular matrix remodeling during blastema formation. In particular, hydrogen peroxide (H2O2) functions as an essential early signal that promotes both blastema initiation and progenitor cell reentry into the cell cycle during axolotl limb regeneration [85]. In addition, it has been proposed that ROS facilitate the recruitment of leukocytes to the site of injury to prevent fibrosis [86]. Specifically, inhibition of ROS activity has been shown to cause fibrotic tissue formation between the epithelium and the underlying remnant tissue, an effect similar to that observed with macrophage clearance [87]. Although transient ROS could cause a wide range of cellular damage, axolotl regeneration is supported by an antioxidant response that counteracts these harmful effects. At controlled or physiological levels—below the threshold of antioxidant capacity—ROS instead serve important roles in cell signaling, proliferation, and the maintenance of cellular homeostasis [88,89]. Notably, axolotl heart regeneration demonstrates remarkable resilience to the detrimental impact of hyperoxia [37].
Remarkably, axolotls retain their ability to heal without scarring even after undergoing metamorphosis, although the regeneration process becomes slower [82]. Likewise, the animals show a reduction in both the speed and fidelity of tissue regeneration, likely due to a slower cell cycle progression and decreased proliferative capacity [90].
4. Conclusions and Future Perspectives
Many questions remain unanswered regarding whether axolotls are truly resistant to cancer, as well as the underlying mechanisms that might contribute to a reduced risk of cancer development in this species (Table 3). Nevertheless, this manuscript reviews several well-established traits of the axolotl, such as neoteny, regenerative capacity, and high cellular plasticity, which may contribute to the development of cancer resistance. In addition, it brings attention to lesser-studied features, including their low metabolic rate, reduced thyroid hormone levels, decreased insulin sensitivity, presence of natural antitumor compounds, unique genomic and epigenetic profiles, and characteristics of axolotl innate immune system. These findings reinforce the potential of the axolotl as a valuable, yet underutilized, model in cancer research. Exploration of these traits would not only deepen our understanding of axolotl biology but also offers promising prospects for cancer prevention and treatment in humans.
This review highlights that axolotls possess certain protective mechanisms that may enable them to evade several distinctive features of cancer (Table 2). Their low insulin sensitivity helps limit sustained proliferative signaling, while efficient DNA repair systems minimize genome instability and mutation. A well-developed repertoire of tumor suppressor genes further supports growth inhibition, and low levels of thyroid hormones interfere with the metabolic and proliferative demands of tumor cells, restricting cancer-associated metabolic reprogramming. In addition, robust epigenetic regulation protects against tumor-related epigenetic alterations, and a strong innate immune system enhances the recognition and elimination of abnormal cells. Finally, favorable environmental and immune conditions help prevent the chronic inflammation and fibrosis that often promote tumor development.
An important avenue for research is the examination of cancer incidence in aging axolotls, especially given that most human cancers arise in older individuals. Studying older axolotls could provide valuable insights into their purported resistance to cancer and how it may change with age. Future research could also consider determining the mutation rate of the axolotl as a means of assessing the efficacy of its DNA replication and repair mechanisms. Although this is challenging due to the large size and highly repetitive nature of the axolotl genome, such investigations could provide valuable data. Identification of the genetic and metabolic mechanisms activated by the axolotl to maintain homeostasis and respond to various forms of cellular stress, such as replicative, oxidative, metabolic, and endoplasmic stress, could also provide valuable information.
The identification, characterization, and purification of compounds with antitumor activity in axolotl extracts would be highly valuable. Equally important is the molecular elucidation of the mechanisms underlying their effects. These compounds could hold significant potential for the pharmaceutical industry, either as direct-acting anticancer agents or as adjuvants to enhance existing cancer therapies.
Investigating the unique tumor suppressor mechanisms present in the axolotl has the potential to significantly advance our understanding of cancer biology and could have a profound impact on the scientific community. Equally crucial is uncovering the evolutionary origins of these mechanisms—understanding how and why axolotls developed such effective cancer resistance could reveal fundamental principles applicable to other species, including humans.
Future research should also extend beyond traditional studies of DNA methylation to explore other epigenetic modifications that may contribute to tumor suppression. Additionally, a comprehensive characterization of all immune cell subtypes in the axolotl is needed. Particular attention should be paid to elucidating the specific antitumor roles played by components of the innate immune system, as well as investigating the potential involvement of adaptive immunity in cancer resistance. Such insights could open new avenues for the development of innovative cancer therapies inspired by the axolotl’s biology.
This review has several limitations, primarily due to the limited availability of data, which at times meant that certain observations were based on a single original source. In general, the article aims to present interpretations based on the available evidence; however, in some cases, reasoned speculation is offered when data are limited or suggestive. Further research in this field is necessary to draw definitive conclusions, and this manuscript aims to encourage and inspire continued investigation.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable to this article as no new data were created or analyzed in this study.
Conflicts of Interest
The author declares no conflicts of interest.
Abbreviations
| NCDs | Noncommunicable diseases |
| ROS | Reactive oxygen species |
| TP53 | Tumor protein p53 |
| HPT | Hypothalamo-pituitary-thyroid (axis) |
| NCCRP1 | Non-specific cytotoxic cell receptor protein 1 |
| NCCs | Nonspecific cytotoxic cells |
| NK | Natural killer (Cells) |
| NK Tag | Natural killer target antigen |
| MHC | Major histocompatibility complex |
| ECM | Extracellular matrix |
| H2O2 | Hydrogen peroxide |
| SASP | Senescence-associated secretory phenotype |
| IGF-1 | Insulin-like growth factor |
| BLAST | basic local alignment search tool |
References
- T. Sandoval-Guzmán, The axolotl, Nat Methods 20 (2023) 1117–1119. [CrossRef]
- C. McCusker, S. V. Bryant, D.M. Gardiner, The axolotl limb blastema: cellular and molecular mechanisms driving blastema formation and limb regeneration in tetrapods, Regeneration 2 (2015) 54–71. [CrossRef]
- R. Stone, In Mexico, the world’s last wild axolotls face extinction, Science (1979) 380 (2023) 570–570. [CrossRef]
- D.M. Parkin, F. Bray, J. Ferlay, P. Pisani, Global cancer statistics, 2002., CA Cancer J Clin 55 (2005) 74–108. [CrossRef]
- F.A. Lagunas-Rangel, DNA damage accumulation and repair defects in FLT3-ITD acute myeloid leukemia: Implications for clonal evolution and disease progression, Hematol Oncol 41 (2023) 26–38. [CrossRef]
- F.A. Lagunas-Rangel, Role of circular RNAs in DNA repair, RNA Biol 21 (2024) 149–161. [CrossRef]
- D. Hanahan, Hallmarks of Cancer: New Dimensions, Cancer Discov 12 (2022) 31–46. [CrossRef]
- L. Nunney, The real war on cancer: the evolutionary dynamics of cancer suppression, Evol Appl 6 (2013) 11–19. [CrossRef]
- F.A. Lagunas-Rangel, V. Chávez-Valencia, Learning of nature: The curious case of the naked mole rat, Mech Ageing Dev 164 (2017) 76–81. [CrossRef]
- F.A. Lagunas-Rangel, Cancer-free aging: Insights from Spalax ehrenbergi superspecies, Ageing Res Rev 47 (2018) 18–23. [CrossRef]
- F.A. Lagunas-Rangel, Why do bats live so long?—Possible molecular mechanisms, Biogerontology 21 (2020) 1–11. [CrossRef]
- F.A. Lagunas-Rangel, Deciphering the whale’s secrets to have a long life, Exp Gerontol 151 (2021) 111425. [CrossRef]
- A. Seluanov, V.N. Gladyshev, J. Vijg, V. Gorbunova, Mechanisms of cancer resistance in long-lived mammals, Nat Rev Cancer (2018) 1. [CrossRef]
- A.J. Ingram, The reactions to carcinogens in the axolotl ( Ambystoma mexicanum ) in relation to the ‘regeneration field control’ hypothesis, Development 26 (1971) 425–441. [CrossRef]
- R. Tacutu, D. Thornton, E. Johnson, A. Budovsky, Di. Barardo, T. Craig, E. Diana, G. Lehmann, D. Toren, J. Wang, V.E. Fraifeld, J.P. de Magalhães, Human Ageing Genomic Resources: new and updated databases, Nucleic Acids Res 46 (2018) D1083–D1090. [CrossRef]
- W.A. Vieira, K.M. Wells, C.D. McCusker, Advancements to the Axolotl Model for Regeneration and Aging, Gerontology 66 (2020) 212–222. [CrossRef]
- K. Sousounis, A.T. Athippozhy, S.R. Voss, P.A. Tsonis, Plasticity for axolotl lens regeneration is associated with age-related changes in gene expression, Regeneration 1 (2014) 47–57. [CrossRef]
- A. Bölük, M. Yavuz, T. Demircan, Axolotl: A resourceful vertebrate model for regeneration and beyond, Developmental Dynamics 251 (2022) 1914–1933. [CrossRef]
- S. Nowoshilow, S. Schloissnig, J.-F. Fei, A. Dahl, A.W.C.W.C. Pang, M. Pippel, S. Winkler, A.R.R. Hastie, G. Young, J.G.G. Roscito, F. Falcon, D. Knapp, S. Powell, A. Cruz, H. Cao, B. Habermann, M. Hiller, E.M.M. Tanaka, E.W.W. Myers, The axolotl genome and the evolution of key tissue formation regulators, Nature 554 (2018) 50–55. [CrossRef]
- Multiple Melanomas in the Axolot Ambystoma mexicanum, JNCI: Journal of the National Cancer Institute (1979). [CrossRef]
- H.G. Schlumberger, B.H.E.W. Lucke, Tumors of fishes, amphibians, and reptiles., Cancer Res 8 (1948) 657–753. http://www.ncbi.nlm.nih.gov/pubmed/18119893.
- S.K. Frost, L.G. Epp, S.J. Robinson, The pigmentary system of developing axolotls, Development 81 (1984) 105–125. [CrossRef]
- L. Denat, A.L. Kadekaro, L. Marrot, S.A. Leachman, Z.A. Abdel-Malek, Melanocytes as Instigators and Victims of Oxidative Stress, Journal of Investigative Dermatology 134 (2014) 1512–1518. [CrossRef]
- E.A.A. Sheremetieva, Spontaneous melanoma in regenerating tails of axolotls, Journal of Experimental Zoology 158 (1965) 101–121. [CrossRef]
- K.S. Vuong, C. Metral, W. Sheley, M. Liu, A.K. Carlson, M.P. Jones, Chromatophoromas on the tail of a pet axolotl (Ambystoma mexicanum) with presumptive metastasis, J Exot Pet Med 45 (2023) 8–13. [CrossRef]
- V.V. V Brunst, A.L.L. Roque, A spontaneous teratoma in an axolotl (Siredon mexicanum)., Cancer Res 29 (1969) 223–9.
- J.C.C. Harshbarger, S.C.C. Chang, L.E.E. DeLanney, F.L.L. Rose, D.E.E. Green, Cutaneous mastocytomas in the neotenic caudate amphibians Ambystoma mexicanum (axolotl) and Ambystoma tigrinum (tiger salamander), J Cancer Res Clin Oncol 125 (1999) 187–192. [CrossRef]
- M. Lo, P.R. Badial, P.M. DiGeronimo, E.M. Littman, N. Aquilino, T. Stokol, N.I. Stacy, R.J. Ossiboff, Acute myeloid leukemia in a genetically related captive population of axolotls ( Ambystoma mexicanum ), Vet Pathol (2025). [CrossRef]
- B. Menger, P.M. Vogt, I.D. Jacobsen, C. Allmeling, J.W. Kuhbier, F. Mutschmann, K. Reimers, Resection of a large intra-abdominal tumor in the Mexican axolotl: a case report., Vet Surg 39 (2010) 232–3. [CrossRef]
- F. Modesto, A. Nicolier, C. Hurtrel, J. Benoît, Excisional biopsy and radiotherapy for management of an olfactory neuroblastoma in an axolotl (Ambystoma mexicanum), J Am Vet Med Assoc 260 (2022) 436–441. [CrossRef]
- C. Shioda, K. Uchida, H. Nakayama, Pathological features of olfactory neuroblastoma in an axolotl (Ambystoma mexicanum)., J Vet Med Sci 73 (2011) 1109–11. [CrossRef]
- R.R. Humphrey, Tumors of the Testis in the Mexican Axolotl (Ambystoma, or Siredon, mexicanum), in: 1969: pp. 220–228. [CrossRef]
- L.E.E. DeLanney, Lymphosarcoma in the Mexican Axolotl, Ambystoma mexicanum, in: Bibl Haematol, 1970: pp. 642–642. [CrossRef]
- T. Seki, Y. Yang, X. Sun, S. Lim, S. Xie, Z. Guo, W. Xiong, M. Kuroda, H. Sakaue, K. Hosaka, X. Jing, M. Yoshihara, L. Qu, X. Li, Y. Chen, Y. Cao, Brown-fat-mediated tumour suppression by cold-altered global metabolism, Nature 608 (2022) 421–428. [CrossRef]
- J.C.M. Ng, C.M. Schooling, Effect of Basal Metabolic Rate on Cancer: A Mendelian Randomization Study, Front Genet 12 (2021). [CrossRef]
- H. Ma, G. Peng, Y. Hu, B. Lu, Y. Zheng, Y. Wu, W. Feng, Y. Shi, X. Pan, L. Song, I. Stützer, Y. Liu, J. Fei, Revealing the biological features of the axolotl pancreas as a new research model, Front Cell Dev Biol 13 (2025). [CrossRef]
- A. Dittrich, S.A. Andersson, M. Busk, K. Hansen, C.B. Foldager, J. Palmfeldt, A. Andersen, M. Pedersen, M. Vendelbo, K.L. Nielsen, H. Lauridsen, Metabolic changes during cardiac regeneration in the axolotl, Developmental Dynamics (2025). [CrossRef]
- T.P. Grazioso, N. Djouder, The forgotten art of cold therapeutic properties in cancer: A comprehensive historical guide, IScience 26 (2023) 107010. [CrossRef]
- É. Villiard, H. Brinkmann, O. Moiseeva, F.A. Mallette, G. Ferbeyre, S. Roy, Urodele p53 tolerates amino acid changes found in p53 variants linked to human cancer, BMC Evol Biol 7 (2007) 180. [CrossRef]
- Y. Liu, Z. Su, O. Tavana, W. Gu, Understanding the complexity of p53 in a new era of tumor suppression, Cancer Cell 42 (2024) 946–967. [CrossRef]
- K. V. Prahlad, L.E. DeLanney, A study of induced metamorphosis in the Axolotl, Journal of Experimental Zoology 160 (1965) 137–145. [CrossRef]
- B. De Groef, S.V.H. Grommen, V.M. Darras, Forever young: Endocrinology of paedomorphosis in the Mexican axolotl (Ambystoma mexicanum), Gen Comp Endocrinol 266 (2018) 194–201. [CrossRef]
- R. Mullur, Y.-Y. Liu, G.A. Brent, Thyroid Hormone Regulation of Metabolism, Physiol Rev 94 (2014) 355–382. [CrossRef]
- E. Krashin, A. Piekiełko-Witkowska, M. Ellis, O. Ashur-Fabian, Thyroid Hormones and Cancer: A Comprehensive Review of Preclinical and Clinical Studies, Front Endocrinol (Lausanne) 10 (2019). [CrossRef]
- H. Lin, Y. Chin, Y.S.H. Yang, H. Lai, J. Whang-Peng, L.F. Liu, H. Tang, P.J. Davis, Thyroid Hormone, Cancer, and Apoptosis, in: Compr Physiol, Wiley, 2016: pp. 1221–1237. [CrossRef]
- B.R. Gauthier, A. Sola-García, M.Á. Cáliz-Molina, P.I. Lorenzo, N. Cobo-Vuilleumier, V. Capilla-González, A. Martin-Montalvo, Thyroid hormones in diabetes, cancer, and aging, Aging Cell 19 (2020). [CrossRef]
- F.A. Lagunas-Rangel, Naked mole-rat hyaluronan, Biochimie 220 (2024) 58–66. [CrossRef]
- B.P. Leitner, S. Siebel, N.D. Akingbesote, X. Zhang, R.J. Perry, Insulin and cancer: a tangled web, Biochemical Journal 479 (2022) 583–607. [CrossRef]
- D. Guo, Y. Meng, G. Zhao, Q. Wu, Z. Lu, Moonlighting functions of glucose metabolic enzymes and metabolites in cancer, Nat Rev Cancer 25 (2025) 426–446. [CrossRef]
- N. Dastagir, C. Liebsch, J. Kutz, S. Wronski, A. Pich, D. Obed, P.M. Vogt, V. Bucan, S. Strauß, Identification of antimicrobial peptides from the Ambystoma mexicanum displaying antibacterial and antitumor activity, PLoS One 20 (2025) e0316257. [CrossRef]
- S. Suleiman, R. Di Fiore, A. Cassar, M.M. Formosa, P. Schembri-Wismayer, J. Calleja-Agius, Axolotl Ambystoma mexicanum extract induces cell cycle arrest and differentiation in human acute myeloid leukemia HL-60 cells., Tumour Biol 42 (2020) 1010428320954735. [CrossRef]
- C. Allegrucci, M.D. Rushton, J.E. Dixon, V. Sottile, M. Shah, R. Kumari, S. Watson, R. Alberio, A.D. Johnson, Epigenetic reprogramming of breast cancer cells with oocyte extracts, Mol Cancer 10 (2011) 7. [CrossRef]
- R. Stewart, C.A. Rascón, S. Tian, J. Nie, C. Barry, L.-F. Chu, H. Ardalani, R.J. Wagner, M.D. Probasco, J.M. Bolin, N. Leng, S. Sengupta, M. Volkmer, B. Habermann, E.M. Tanaka, J.A. Thomson, C.N. Dewey, Comparative RNA-seq Analysis in the Unsequenced Axolotl: The Oncogene Burst Highlights Early Gene Expression in the Blastema, PLoS Comput Biol 9 (2013) e1002936. [CrossRef]
- S.R. Voss, A. Palumbo, R. Nagarajan, D.M. Gardiner, K. Muneoka, A.J. Stromberg, A.T. Athippozhy, Gene expression during the first 28 days of axolotl limb regeneration I: Experimental design and global analysis of gene expression, Regeneration 2 (2015) 120–136. [CrossRef]
- B. Corradetti, P. Dogra, S. Pisano, Z. Wang, M. Ferrari, S. Chen, R.L. Sidman, R. Pasqualini, W. Arap, V. Cristini, Amphibian regeneration and mammalian cancer: Similarities and contrasts from an evolutionary biology perspective, BioEssays 43 (2021). [CrossRef]
- M. Conrad, The mutation buffering concept of biomolecular structure, J Biosci 8 (1985) 669–679. [CrossRef]
- S. Nowoshilow, E.M. Tanaka, Introducing www.axolotl-omics.org – an integrated -omics data portal for the axolotl research community, Exp Cell Res 394 (2020) 112143. [CrossRef]
- Y. Haluza, J.A. Zoller, A.T. Lu, H.E. Walters, M. Lachnit, R. Lowe, A. Haghani, R.T. Brooke, N. Park, M.H. Yun, S. Horvath, Axolotl epigenetic clocks offer insights into the nature of negligible senescence, BioRxiv (2024). [CrossRef]
- S. Sonar, S. Nyahatkar, K. Kalele, M.D. Adhikari, Role of DNA methylation in cancer development and its clinical applications, Clinical and Translational Discovery 4 (2024). [CrossRef]
- D. Lopez, L. Lin, J.R. Monaghan, C.R. Cogle, F.J. Bova, M. Maden, E.W. Scott, Mapping hematopoiesis in a fully regenerative vertebrate: the axolotl, Blood 124 (2014) 1232–1241. [CrossRef]
- J.S. Fellah, F. Kerfourn, A.-M. Dumay, G. Aubet, J. Charlemagne, Structure and diversity of the T-cell receptor α chain in the Mexican axolotl, Immunogenetics 45 (1997) 235–241. [CrossRef]
- A. Tournefier, V. Laurens, C. Chapusot, P. Ducoroy, M.R. Padros, F. Salvudori, B. Sammut, Structure of MHC class I and class II cDNAs and possible immunodeficiency linked to class II expression in the Mexican axolotl, Immunol Rev 166 (1998) 259–277. [CrossRef]
- J.D. Cotter, A. Storfer, R.B. Page, C.K. Beachy, S.R. Voss, Transcriptional response of Mexican axolotls to Ambystoma tigrinum virus (ATV) infection, BMC Genomics 9 (2008) 493. [CrossRef]
- K. Reimers, M. Abu Qarn, C. Allmeling, V. Bucan, P.M. Vogt, Identification of the non-specific cytotoxic cell receptor protein 1 (NCCRP1) in regenerating axolotl limbs, Journal of Comparative Physiology B 176 (2006) 599–605. [CrossRef]
- M. Huang, Z. Zhang, X. Li, J. Feng, Y.Y. Huang, A. Kwaku, Y.Y. Huang, J. Jian, Molecular characteristics and functional analysis of non-specific cytotoxic cell receptor (NCCRP1) in golden pompano (Trachinotus ovatus), Fish Shellfish Immunol 145 (2024) 109344. [CrossRef]
- D.T. Harris, L. Jaso-Friedmann, R.B. Devlin, H.S. Koren, D.L. Evans, Identification of an evolutionarily conserved, function-associated molecule on human natural killer cells., Proceedings of the National Academy of Sciences 88 (1991) 3009–3013. [CrossRef]
- A. Czarkwiani, M. Lobo, L.A.B. Castro, A. Petzold, F. Rost, R. Maehr, M.H. Yun, Molecular basis for de novo thymus regeneration in a vertebrate, the axolotl, Sci Immunol 10 (2025). [CrossRef]
- F.A. Lagunas-Rangel, Neutrophil-to-lymphocyte ratio in aging: Trends and clinical implications, Exp Gerontol 211 (2025) 112908. [CrossRef]
- A. Mier-Briseño, M.A. Benavides-Huerto, I. Padilla-Ponce, F.A. Lagunas-Rangel, Atypical Carcinoid of the Thymus: Early Diagnosis in a Case Report, Medical Sciences 13 (2025) 96. [CrossRef]
- Y.-C. Ching, R.J. Wedgwood, Immunologic Responses in the Axolotl, Siredon Mexicanum, The Journal of Immunology 99 (1967) 191–200. [CrossRef]
- A. Tournefier, S. Fellah, J. Charlemagne, Monoclonal antibodies to axolotl immunoglobulins specific for different heavy chains isotypes expressed by independent lymphocyte subpopulations, Immunol Lett 18 (1988) 145–148. [CrossRef]
- J. Charlemagne, Thymus independent anti-horse erythrocyte antibody response and suppressor T cells in the Mexican axolotl (Amphibia, Urodela, ambystoma mexicanum)., Immunology 36 (1979) 643–8.
- A.P. Ussing, P. Rosenkilde, Effect of Induced Metamorphosis on the Immune System of the Axolotl, Ambystoma mexicanum, Gen Comp Endocrinol 97 (1995) 308–319. [CrossRef]
- Y. Wen, Y. Zhu, C. Zhang, X. Yang, Y. Gao, M. Li, H. Yang, T. Liu, H. Tang, Chronic inflammation, cancer development and immunotherapy, Front Pharmacol 13 (2022). [CrossRef]
- F.A. Lagunas-Rangel, Neutrophil-to-lymphocyte ratio in aging: Trends and clinical implications, Exp Gerontol 211 (2025) 112908. [CrossRef]
- E.M. Tanaka, P.W. Reddien, The Cellular Basis for Animal Regeneration, Dev Cell 21 (2011) 172–185. [CrossRef]
- A. Espinal-Centeno, M. Dipp-Álvarez, C. Saldaña, L. Bako, A. Cruz-Ramírez, Conservation analysis of core cell cycle regulators and their transcriptional behavior during limb regeneration in Ambystoma mexicanum, Mech Dev 164 (2020) 103651. [CrossRef]
- S. Ruiz-Pérez, N. Alcaraz, K. Torres-Arciga, J.A. Ocampo-Cervantes, A. Cervera, C. Castro-Hernández, C.G. Sámano-Salazar, E. Soto-Reyes, R. González-Barrios, Retrotransposon Expression Is Upregulated in Adulthood and Suppressed during Regeneration of the Limb in the Axolotl ( Ambystoma mexicanum ), Adv Biol (2025). [CrossRef]
- A. Zippo, S. Beyes, Molecular mechanisms altering cell identity in cancer, Oncogene (2025). [CrossRef]
- B. Piersma, M.-K. Hayward, V.M. Weaver, Fibrosis and cancer: A strained relationship, Biochimica et Biophysica Acta (BBA) - Reviews on Cancer 1873 (2020) 188356. [CrossRef]
- T. Endo, S. V Bryant, D.M. Gardiner, A stepwise model system for limb regeneration, Dev Biol 270 (2004) 135–145. [CrossRef]
- A.W. Seifert, J.R. Monaghan, S.R. Voss, M. Maden, Skin Regeneration in Adult Axolotls: A Blueprint for Scar-Free Healing in Vertebrates, PLoS One 7 (2012) e32875. [CrossRef]
- A.K. Rodgers, J.J. Smith, S.R. Voss, Identification of immune and non-immune cells in regenerating axolotl limbs by single-cell sequencing, Exp Cell Res 394 (2020) 112149. [CrossRef]
- J.W. Godwin, R. Debuque, E. Salimova, N.A. Rosenthal, Heart regeneration in the salamander relies on macrophage-mediated control of fibroblast activation and the extracellular landscape, NPJ Regen Med 2 (2017) 22. [CrossRef]
- B. Carbonell-M, J. Zapata Cardona, J.P. Delgado, Post-amputation reactive oxygen species production is necessary for axolotls limb regeneration, Front Cell Dev Biol 10 (2022). [CrossRef]
- B. Carbonell M, J. Zapata Cardona, J.P. Delgado, Hydrogen peroxide is necessary during tail regeneration in juvenile axolotl, Developmental Dynamics 251 (2022) 1054–1076. [CrossRef]
- J.W. Godwin, A.R. Pinto, N.A. Rosenthal, Macrophages are required for adult salamander limb regeneration, Proceedings of the National Academy of Sciences 110 (2013) 9415–9420. [CrossRef]
- F.A. Lagunas-Rangel, J.V. Linnea-Niemi, B. Kudłak, M.J. Williams, J. Jönsson, H.B. Schiöth, Role of the Synergistic Interactions of Environmental Pollutants in the Development of Cancer., Geohealth 6 (2022) e2021GH000552. [CrossRef]
- P.D. Ray, B.-W. Huang, Y. Tsuji, Reactive oxygen species (ROS) homeostasis and redox regulation in cellular signaling, Cell Signal 24 (2012) 981–990. [CrossRef]
- J.R. Monaghan, A.C. Stier, F. Michonneau, M.D. Smith, B. Pasch, M. Maden, A.W. Seifert, Experimentally induced metamorphosis in axolotls reduces regenerative rate and fidelity, Regeneration 1 (2014) 2–14. [CrossRef]
Figure 1.
Key mechanisms identified in the axolotl that could contribute to cancer resistance.
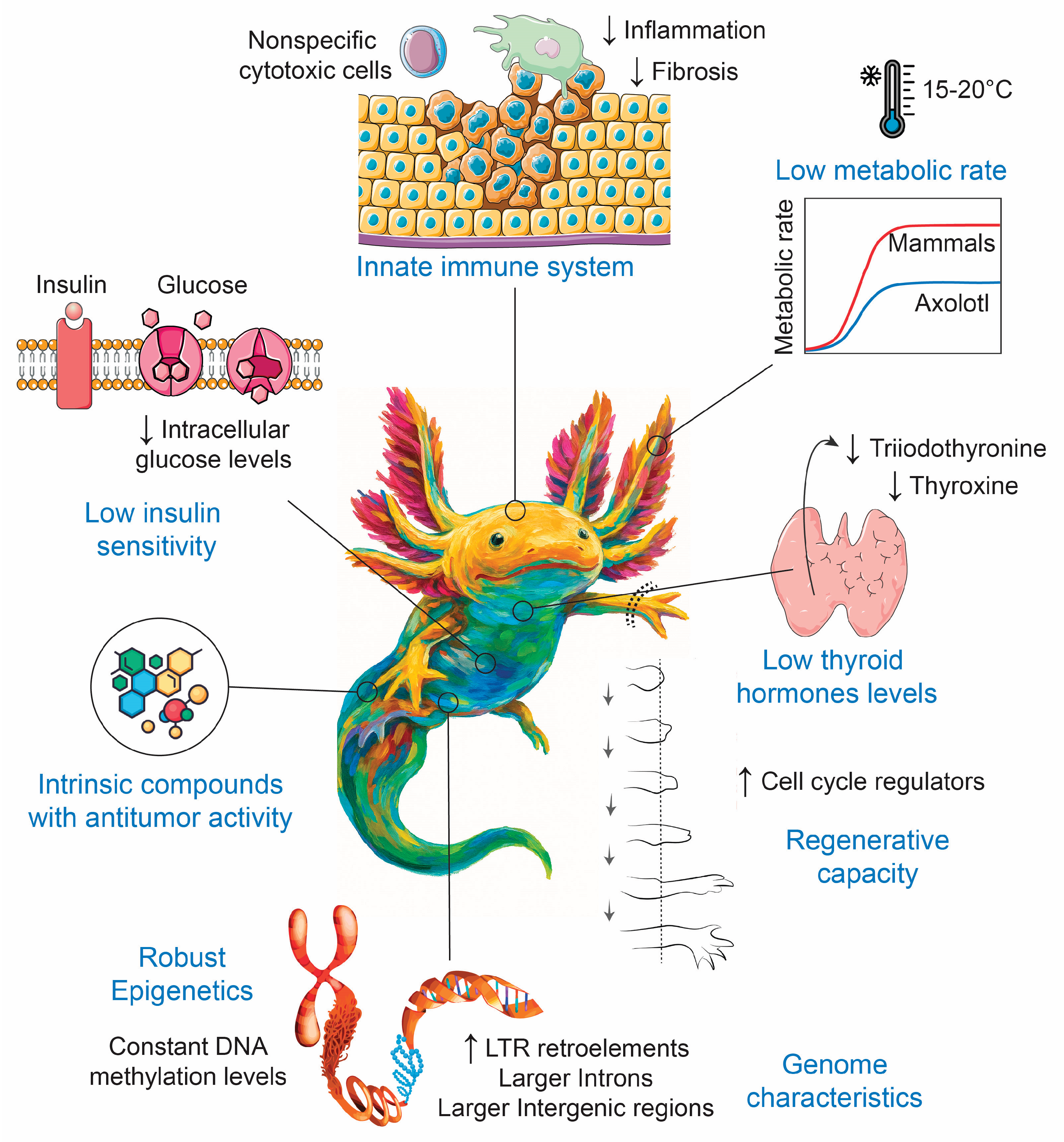
Figure 2.
Axolotls show low insulin signaling. Axolotls exhibit lower blood glucose levels and reduced sensitivity to insulin, factors that may contribute to their lower incidence of cancer. Insulin binds to the extracellular α-subunit of the insulin receptor (INSR), leading to autophosphorylation and activation of its intracellular β-subunit. Once activated, the INSR phosphorylates several intracellular substrates, including members of the insulin receptor substrate family (IRS1–IRS4) and SHC. Phosphorylated tyrosine residues on IRS proteins provide docking sites for the p85 regulatory subunit of phosphatidylinositol 3-kinase (PI3K). Activated PI3K initiates downstream signaling cascades, most prominently the activation of AKT (also known as protein kinase B), which promotes cell survival, metabolism, and growth. In parallel, phosphorylation of SHC by the INSR activates the mitogen-activated protein kinase (MAPK) pathway. SHC recruits growth factor receptor-binding protein 2 (GRB2), which, in turn, activates the guanine nucleotide exchange factor son of sevenless (SOS), leading to activation of the small GTP-binding protein RAS. Activated RAS triggers a kinase cascade involving RAF–MEK–MAPK, ultimately resulting in full activation of the MAPK pathway.
Figure 2.
Axolotls show low insulin signaling. Axolotls exhibit lower blood glucose levels and reduced sensitivity to insulin, factors that may contribute to their lower incidence of cancer. Insulin binds to the extracellular α-subunit of the insulin receptor (INSR), leading to autophosphorylation and activation of its intracellular β-subunit. Once activated, the INSR phosphorylates several intracellular substrates, including members of the insulin receptor substrate family (IRS1–IRS4) and SHC. Phosphorylated tyrosine residues on IRS proteins provide docking sites for the p85 regulatory subunit of phosphatidylinositol 3-kinase (PI3K). Activated PI3K initiates downstream signaling cascades, most prominently the activation of AKT (also known as protein kinase B), which promotes cell survival, metabolism, and growth. In parallel, phosphorylation of SHC by the INSR activates the mitogen-activated protein kinase (MAPK) pathway. SHC recruits growth factor receptor-binding protein 2 (GRB2), which, in turn, activates the guanine nucleotide exchange factor son of sevenless (SOS), leading to activation of the small GTP-binding protein RAS. Activated RAS triggers a kinase cascade involving RAF–MEK–MAPK, ultimately resulting in full activation of the MAPK pathway.
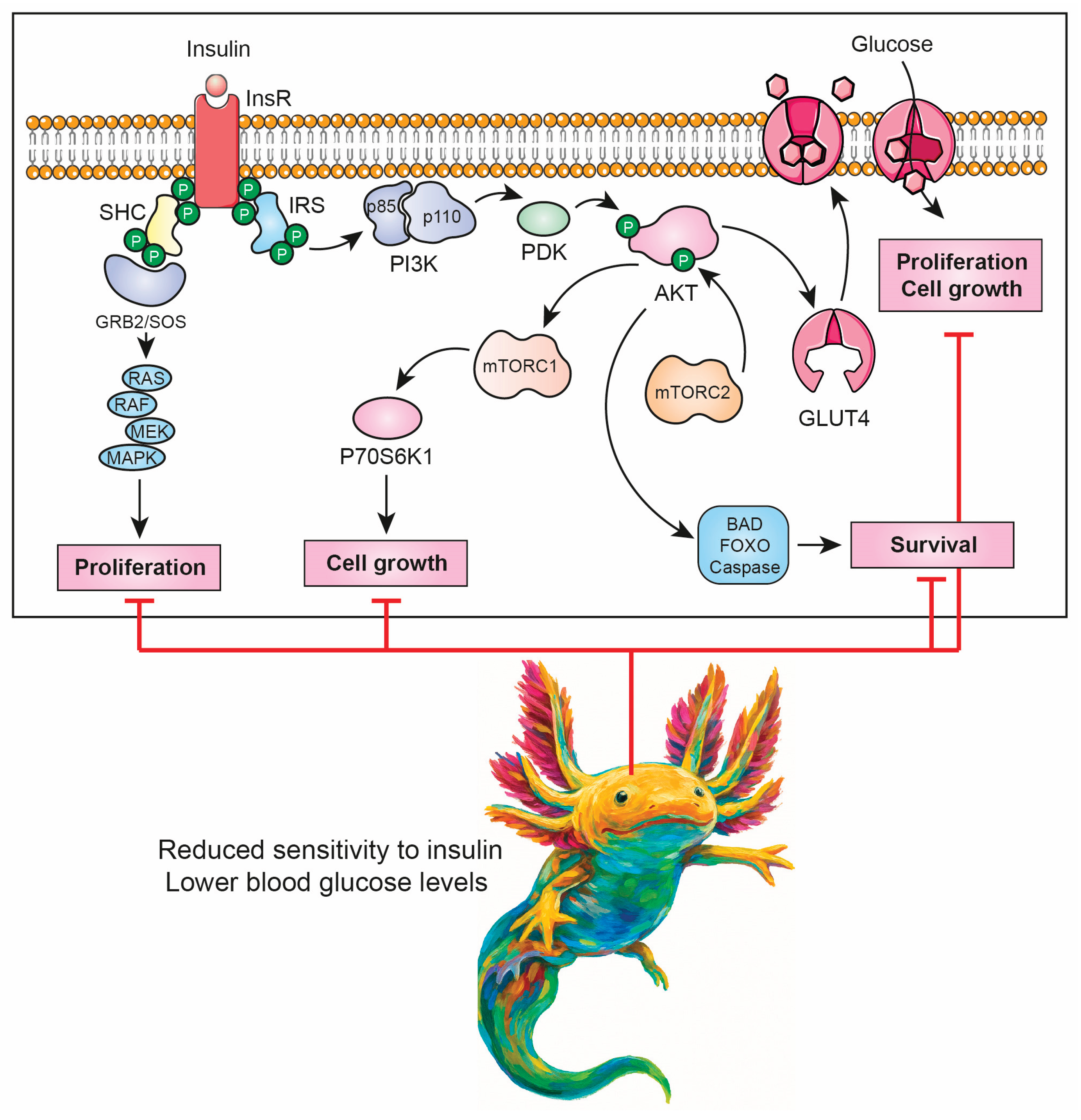
Figure 3.
Innate immune system in axolotls. Non-specific cytotoxic cells (NCCs) may play a key role in the axolotl’s innate immune defense. They can mediate the lysis of target cells, such as tumor cells, by binding to their natural ligand—the natural killer target antigen (NK Tag)—and releasing lytic granules. Expression of the nonspecific cytotoxic cell receptor protein 1 (NCCRP1), which is involved in antigen recognition, has been detected in multiple axolotl tissues, including peripheral blood mononuclear cells, skin, lung, spleen, and limb blastema. NCCs are considered evolutionary precursors of mammalian natural killer (NK) cells and may contribute to the antitumoral immune response in axolotls.
Figure 3.
Innate immune system in axolotls. Non-specific cytotoxic cells (NCCs) may play a key role in the axolotl’s innate immune defense. They can mediate the lysis of target cells, such as tumor cells, by binding to their natural ligand—the natural killer target antigen (NK Tag)—and releasing lytic granules. Expression of the nonspecific cytotoxic cell receptor protein 1 (NCCRP1), which is involved in antigen recognition, has been detected in multiple axolotl tissues, including peripheral blood mononuclear cells, skin, lung, spleen, and limb blastema. NCCs are considered evolutionary precursors of mammalian natural killer (NK) cells and may contribute to the antitumoral immune response in axolotls.
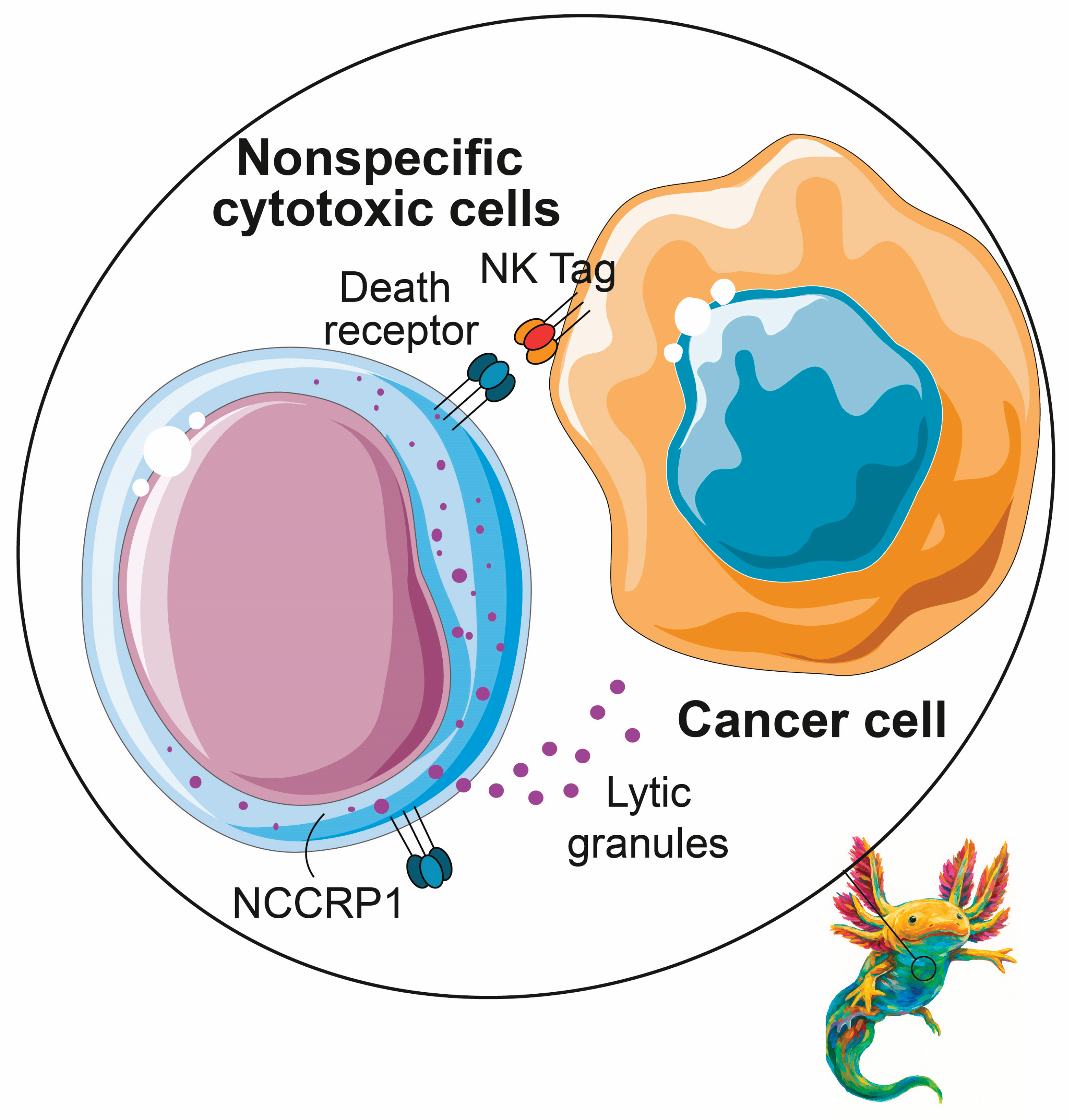

Table 1.
Comparative analysis of key aspects relevant to cancer across axolotls, zebrafish, mice, and humans.
Table 1.
Comparative analysis of key aspects relevant to cancer across axolotls, zebrafish, mice, and humans.
| Organism | ||||
|---|---|---|---|---|
| Feature | Axolotl | Zebrafish | Mice | Human |
| Maximum longevity [15] | 21 years (captivity) |
5.5 years (captivity) |
4 years (captivity) |
122.5 years |
| Basal metabolic rate [15] | ~0.025 W | ~0.00056 W | 0.2710 W | 82.7800 W |
| Body mass [15] | ~60–150 g (lab-bred individuals) |
~0.3–0.9 g (lab-reared adults) |
18.0 g | 70,000.0 g |
| Typical body temperature [15] | ~16–20 °C (laboratory) |
~28 °C (laboratory) |
36.9 °C | 37.0 °C |
| Cancer incidence | Rare (suggested) |
Moderate | High | Moderate |
| Regenerative capacity | Very high | High (embryos) Moderate (adults) |
Limited | Very limited |
| Genome size [19] | ∼32 Gb | ∼1.5 Gb | ∼2.7 Gb | ∼3.2 Gb |
| Chromosomes [19] | 56 | 50 | 40 | 46 |
| Protein-coding genes [19] | 23,251 | 26,206 | 21,529 | 19,435 |
| Median intron length [19] | 22,759 bp | ~1350 bp | 1469 bp | 1750 bp |
| LTR retroelements [19] | High | Low | Moderate | Moderate |
| Innate immune system | Robust and active | Functional and efficient | Functional and balanced | Functional and balanced |
| Adaptive immune system | Limited | Limited | Diversified and efficient | Highly diversified and efficient |
Table 2.
Hallmarks of cancer and potential mechanisms by which axolotls counteract it. The hallmarks of cancer framework serves as a powerful heuristic tool to distill the extraordinary complexity of cancer phenotypes and genotypes into a defined set of underlying biological principles [7]. In this context, axolotls display traits that may counteract cancer hallmarks, suggesting mechanisms that could underlie their resistance to cancer. Section 3 provides a more detailed explanation of the axolotl’s mechanisms.
Table 2.
Hallmarks of cancer and potential mechanisms by which axolotls counteract it. The hallmarks of cancer framework serves as a powerful heuristic tool to distill the extraordinary complexity of cancer phenotypes and genotypes into a defined set of underlying biological principles [7]. In this context, axolotls display traits that may counteract cancer hallmarks, suggesting mechanisms that could underlie their resistance to cancer. Section 3 provides a more detailed explanation of the axolotl’s mechanisms.
| Hallmark of Cancer | Hallmark Description | Axolotl Mechanisms That May Counteract the Hallmark Trait |
|---|---|---|
| Sustaining proliferative signaling | Cancer cells maintain chronic growth-promoting signals that drive uncontrolled proliferation. | - Low levels of thyroid hormones - Low insulin sensitivity - Genomic stability |
| Evading growth suppressors | Cancer cells inactivate tumor suppressor pathways that normally restrict cell division. | - Genomic stability - Action of tumor suppressor genes |
| Resisting cell death | Malignant cells circumvent apoptosis and other programmed cell death mechanisms. | - Low levels of thyroid hormones - Low insulin sensitivity |
| Enabling replicative immortality | Telomere maintenance and bypassing senescence mechanisms permit unlimited cellular replication. | - Genomic stability |
| Inducing or accessing vasculature | Tumors promote angiogenesis or exploit existing blood vessels to secure oxygen and nutrients. | - Low levels of thyroid hormones - Low insulin sensitivity |
| Activating invasion and metastasis | Cancer cells acquire abilities to invade tissues and colonize distant organs. | - Low levels of thyroid hormones - Low insulin sensitivity - Genomic stability |
| Reprogramming cellular metabolism | Tumor cells adapt metabolic pathways to support growth, survival, and biosynthesis. | - Cool environment - Low levels of thyroid hormones - Low insulin sensitivity - Genomic stability |
| Avoiding immune destruction | Cancer cells evade recognition and elimination by the host immune system. | - Nonspecific cytotoxic cells - Thymus regeneration |
| Non-mutational epigenetic reprogramming | Epigenetic changes, such as DNA methylation or histone modification, alter gene expression without changing the DNA sequence, allowing cancer cells to adapt and progress more rapidly. | - Robust epigenetic characteristics |
| Polymorphic microbiomes | Variations in host-associated microbiota influence tumor initiation, progression, and therapeutic response. | --- |
| Tumor-promoting inflammation | Inflammatory mediators and cells create a microenvironment that supports tumor progression. | - Cool environment - Attenuated adaptive immune response |
| Senescent cells | Senescent cells have been identified as functional contributors to cancer development and progression | --- |
Table 3.
Outstanding questions and research opportunities in axolotl cancer research. Axolotl model could transform both basic biology and cancer research. Understanding whether axolotls are truly resistant to cancer and how their oncogenic pathways, immune system, and epigenetic regulation differ from mammals could reveal protective mechanisms that suppress tumor development. Likewise, dissecting the fine line between regeneration and tumorigenesis may provide insights into how proliferative programs can be directed toward healing rather than cancer. Comparative studies across urodeles and mammals could highlight evolutionary strategies of tumor suppression, opening new avenues for therapies.
Table 3.
Outstanding questions and research opportunities in axolotl cancer research. Axolotl model could transform both basic biology and cancer research. Understanding whether axolotls are truly resistant to cancer and how their oncogenic pathways, immune system, and epigenetic regulation differ from mammals could reveal protective mechanisms that suppress tumor development. Likewise, dissecting the fine line between regeneration and tumorigenesis may provide insights into how proliferative programs can be directed toward healing rather than cancer. Comparative studies across urodeles and mammals could highlight evolutionary strategies of tumor suppression, opening new avenues for therapies.
UNANSWERED QUESTIONS
|
UNANSWERED QUESTIONS
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



